Submitted:
25 October 2024
Posted:
28 October 2024
You are already at the latest version
Abstract
Despite the increasing number of placenta accreta spectrum (PAS) cases in recent years, its im-pact on neonatal outcomes and respiratory morbidity has not yet been extensively studied. In this context, it is crucial to understand the underlying mechanisms of neonatal complications. Moreover, no study has yet demonstrated the effectiveness of antenatal corticosteroid therapy (CT) for prevention of the respiratory distress syndrome (RDS) in newborns of mothers with PAS at the molecular level. In this regard, miRNA profiling was performed on 160 blood plasma sam-ples from preterm infants (gestational age 33–36 weeks) and their mothers diagnosed with or without PAS. The samples with PAS were categorized into groups: without antenatal RDS prophylaxis, and with CT admin-istered either 2–7 days, 7–14 days, or more than 14 days before delivery. These groups were compared to a control group without PAS in the absence of antenatal CT. Deep sequencing was conducted using the NEB-Next® Multiplex Small RNA Library Prep Set for Illumina® on the NextSeq 500/550 platform, followed by validation by quantitative real-time PCR. A significant increase in hsa-miR-199a-3p and hsa-miR-382-5p levels was observed in the blood plasma of newborns with PAS (placenta accreta and increta) compared to the control group. In maternal blood, hsa-miR-199a-3p levels were markedly higher than hsa-miR-382-5p. The timing of antenatal CT significantly influenced the levels of hsa-miR-199a-3p and hsa-miR-382-5p in neonatal plasma from mothers with placenta accreta or increta. A clear trend toward normalization was observed when CT was administered within 14 days before delivery, particularly in the seven days before delivery, but not beyond 14 days. Among newborns with PAS, higher levels of hsa-miR-382-5p were significantly associated with increased Neomod scale severity scores of 2, 4, and 5 compared to a score of 0. Additional-ly, a direct correlation was found between hsa-miR-382-5p level in neonatal plasma and hsa-miR-199a-3p level in the same sample (r = 0.49; p = 0.0001), oxygen requirements in the NICU (r = 0.41; p = 0.0016), duration of NICU stay (r = 0.31; p = 0.019), and the severity of the newborn’s condition based on the NEOMOD scale (r = 0.36; p = 0.0051). Logistic regression mod-els based on maternal plasma levels of hsa-miR-199a-3p and hsa-miR-382-5p predicted the need for cardiotonic therapy, invasive mechanical ventilation, or high-frequency oscillatory ventila-tion in newborns during the early neonatal period, with a sensitivity of 95-100%. It was con-cluded that elevated circulating levels of miRNAs, hsa-miR-199a-3p and hsa-miR-382-5p, in ma-ternal and fetal blood are crucial in the development of respiratory and cardiac complications in newborns from pregnancies affected by PAS. These miRNAs regulate surfactant synthesis in al-veolar cells, fetal organogenesis via IGF-1, the formation of proper lung tissue architecture, and vascular tone.
Keywords:
placenta accreta spectrum
; RDS
; antenatal corticosteroid therapy
; miRNA
; deep sequencing
; PCR
; neonatal complication
; blood plasma
1. Introduction
Abnormal placental implantation occurs when trophoblasts invade the superficial uterine endometrium (placenta accreta), the myometrium (placenta increta), or beyond the uterine serosa (placenta percreta). Collectively, these conditions are referred to as placenta accreta spectrum (PAS). The primary cause of PAS is thought to be defective decidualization at the implantation site, leading to the absence of both the decidua basalis and Nitabuch’s layer. This results in the direct attachment of chorionic villi to the myometrium [1,2]. The incidence of PAS is estimated to be as high as 1.1% of all births [3], and this rate is rising globally due to an increase in cesarean deliveries and other uterine surgeries, such as surgical uterine evacuations, myomectomies, and infertility treatments [4,5]. Among the types of PAS, placenta accreta is the most common. In a pooled analysis of hysterectomy specimens with confirmed abnormal placentation, the distribution was as follows: placenta accreta (79%), placenta increta (14%), and placenta percreta (7%) [6].
Several clinical studies have shown that PAS is associated with an increased incidence of respiratory distress syndrome (RDS) and a greater need for neonatal respiratory support, including continuous positive airway pressure [7] [8]. RDS occurs due to surfactant deficiency and immature lung development. Although it is well-known that preterm infants (those born before 37 weeks of gestation) are at higher risk for RDS, especially those born before 32 weeks [9] [10], and that the risk decreases with increasing gestational age as organ systems mature [11,12], earlier analysis at the V.I. Kulakov National Medical Research Center of Obstetrics, Gynecology, and Perinatology revealed a more severe course of the early neonatal period and a higher incidence of RDS in preterm infants born to mothers with PAS compared to those born to mothers without PAS [13]. Despite the increasing number of PAS cases in recent years, its impact on neonatal outcomes and respiratory morbidity has not yet been extensively studied in large multicenter clinical trials. Therefore, it is crucial to understand the underlying mechanisms of neonatal complications in the context of PAS.
Andrew Parsons discussed the concept of a placental-pulmonary connection in his review article [14], hypothesizing that placental disorders during pregnancy may uniquely affect the developing fetal lungs due to similarities in their structure and function. Both the placenta and lungs undergo parallel branching morphogenesis during gestation, leading to the formation of functional subunits for gas exchange—placental villi in the placenta and the air-epithelial interface in the lung alveoli. Parsons also explored the relationship between bronchopulmonary dysplasia and hypertensive pregnancy disorders, such as preeclampsia, which is associated with the release of the anti-angiogenic soluble fms-like tyrosine kinase 1 (sFLT-1) from the placenta into the maternal circulation. Increased sFLT-1 levels have been detected in fetal cord blood and amniotic fluid. In an antenatal model, intraamniotic exposure to anti-angiogenic sFLT-1 [15] [16] led to postnatal lung changes, including simplified alveolar structure, altered vascularization, and flattening of bronchial airway epithelium.
Given that PAS is characterized by a proangiogenic placental phenotype and increased trophoblast invasiveness, which contrasts with the antiangiogenic profile seen in preeclamptic placentas, there may be an antenatal link between placental characteristics in PAS and structural changes in the newborn’s lung tissue. Investigating this potential relationship was the focus of the current study.
Corticosteroids have become the standard of care for women at risk of preterm birth before 32 to 34 weeks of gestation in many countries [17]. In the fetal lungs, corticosteroids stimulate the production of proteins, promote the biosynthesis of phospholipids, and increase the production of surfactant [18]. Despite the widespread use of antenatal corticosteroids to prevent RDS in preterm infants, there is still no consensus on the optimal corticosteroid type, dosage, frequency, timing, or administration route [19]. The reduction in the incidence of RDS with antenatal corticosteroid therapy is effective for up to seven days after treatment [20]. A Cochrane Review evaluated whether women who remain undelivered and at risk of preterm birth should receive a repeat course of corticosteroids seven days after the initial treatment [21].
The first large study demonstrating the effectiveness of antenatal corticosteroids for RDS prevention in late preterm infants (34/0–36/6 weeks) was conducted by Gyamfi-Bannerman et al. and published in 2016. It showed a reduction in the need for respiratory and surfactant therapy, as well as supplemental oxygen, in infants whose mothers received antenatal prophylaxis with betamethasone [22]. The study also found a significant decrease in transient tachypnea of the newborn (TTN) and bronchopulmonary dysplasia (BPD). However, there was a noted increase in neonatal hypoglycemia. A systematic review with meta-analysis, published in 2016 and including Gyamfi-Bannerman's study, showed that neonates whose mothers received corticosteroids after 34 weeks had a significantly lower risk of developing RDS, TTN, and required less surfactant and mechanical ventilation. These infants also had shorter NICU stays, higher Apgar scores, and required lower peak oxygen concentrations [23].
The effectiveness of antenatal corticosteroids for preventing RDS in late preterm infants (34/0 – 36/6 weeks) born to mothers with placenta accreta was specifically assessed at the Kulakov National Medical Research Center of Obstetrics, Gynecology, and Perinatology [24]. The study found that when corticosteroids were administered no later than seven days before delivery, there was a reduction in the severity of respiratory disorders and a decrease in the need for invasive respiratory therapy, including high-frequency oscillatory ventilation (HFOVL). RDS prevention earlier in pregnancy was not always determinative, and the positive effects did not depend on the frequency of corticosteroid administration.
Despite these findings, no study has yet demonstrated the effectiveness of antenatal corticosteroid therapy in newborns of mothers with PAS at the molecular level. In earlier research, we identified microRNA markers of PAS in the blood of women during the first trimester of pregnancy [25] and near the time of delivery [26]. MicroRNAs (miRNAs) are small non-coding RNAs that regulate protein-coding mRNAs post-transcriptionally [27]. They are expressed in various cell types and act as biological regulators. In reproductive biology, miRNAs are involved in processes such as spermatogenesis, folliculogenesis, endometrial functions, embryogenesis, maternal recognition of pregnancy, embryo implantation, and placental development [28] [29] [30] [31]. Aberrant miRNA expression has been linked to numerous pathological conditions, including pregnancy complications [32,33,34]. Their ability to be secreted into biological fluids, combined with their measurability, sensitivity, and stability (average half-life of 119 hours), makes them promising markers for identifying pathological conditions [35] [36].
In this study, we aimed to investigate whether there are changes in plasma miRNA levels in premature infants born to mothers with PAS compared to infants of similar gestational age born to mothers without PAS. Additionally, we explored whether these changes are associated with the morphological type of PAS, the severity of respiratory and cardiovascular disorders in the newborn, and the timing of antenatal RDS prophylaxis.
2. Materials and Methods
2.1. Patients
All patients included in the study were admitted to the National Medical Research Center for Obstetrics, Gynecology, and Perinatology, named after Academician V.I. Kulakov of the Ministry of Healthcare of the Russian Federation, for pregnancy and delivery management. They signed informed consent to participate, and the study was approved by the Ethics Committee of the Center.
In the main group (n=69), all women underwent operative delivery via cesarean section due to PAS. In 66 cases, delivery was planned, while 3 cases required emergency cesarean section due to bleeding.
In the control group (n=11), all women also underwent cesarean sections. In 2 cases, the procedure was planned, with indications being preeclampsia in one case and threatened preterm labor in the other. Nine women required emergency cesarean sections, for reasons including bleeding (1 case), onset of labor (2 cases), fetal condition deterioration (3 cases), preeclampsia (1 case), suspected uterine scar failure (1 case), and maternal somatic pathology (1 case).
Antenatal prophylaxis for RDS was conducted following current clinical guidelines for preterm labor management. The drug "Dexamethasone" (manufacturer "Ellara," Russia) was administered intramuscularly at a dose of 8 mg three times, with an 8-hour interval between doses (total dose: 24 mg).
2.2. Isolation of RNA from Peripheral Blood Plasma Samples
Peripheral blood samples were collected into VACUETTE® EDTA tubes, centrifuged for 20 minutes at 300g at 4°C, plasma was collected and centrifuged again for 10 minutes at 16,000g. RNA was isolated from 200 μl of plasma using the miRNeasy Serum/Plasma kit (Qiagen).
2.3. Deep Sequencing of miRNA
cDNA libraries were synthesized using 6 μl of total RNA eluate from neonatal plasma samples with the NEBNext® Multiplex Small RNA Library Prep Set for Illumina® (Set2, New England Biolab®, Germany), following the manufacturer’s protocol. The cDNA libraries were amplified and purified using 6% polyacrylamide gel, with the 140–160 base pair fraction extracted. The quantity and quality of the cDNA libraries were assessed with an Agilent 2100 Bioanalyzer (Agilent Technologies, USA) using the High Sensitivity DNA reagents kit (Agilent Technologies, USA). Sequencing of the cDNA libraries was performed on the NextSeq 500 platform (Illumina, USA), following the manufacturer’s instructions. For sequence annotation, the GRCh38.p15 and miRBase v21 databases were utilized, with the STAR RNAseq aligner program. The DESeq2 software package was used to normalize the cDNA read counts in each sample.
2.4. Reverse Transcription and Quantitative Real-Time PCR
Five microliters of the 14 µL eluate obtained from the miRNeasy Serum/Plasma Kit column (Qiagen, Hilden, Germany), which contained plasma RNA, were used for cDNA synthesis following the manufacturer's protocol with the miRCURY LNA RT Kit (Qiagen, Hilden, Germany). Quantitative real-time PCR was then carried out using the miRCURY LNA SYBR Green PCR Kit (Qiagen, Hilden, Germany) and miRNA-specific primers (miRCURY LNA miRNA PCR Assay, Qiagen, Hilden, Germany) according to the manufacturer's instructions, using a StepOnePlus™ thermal cycler (Applied Biosystems). Relative miRNA expression in plasma was calculated using the ∆Ct method, with UniSp6 serving as the reference RNA.
2.5. Statistical Data Processing
Scripts written in R [37] and the RStudio software [38] were used for statistical analysis. The Shapiro-Wilk test was applied to assess the normality of the data. For non-normally distributed data, paired comparisons were made using the Mann-Whitney test. Variables that did not follow a normal distribution were described as the median (Me) and quartiles Q1 and Q3 in the format Me (Q1; Q3). A significance threshold of p = 0.05 was set, and if the p-value was less than 0.001, it was indicated as p < 0.001.
Logistic regression models were developed in RStudio through stepwise inclusion and exclusion of miRNA marker molecules based on their contribution to the model. The predictive performance of the model was evaluated using ROC (Receiver Operating Characteristic) analysis, assessing the AUC (Area Under the Curve), statistical significance, specificity, and sensitivity.
3. Results
3.1. Deep Sequencing of Neonatal BLOOD plasma miRNA
In the initial phase of the study, a deep sequencing method was employed to analyze the miRNA profiles in the blood plasma of day-old newborns, aiming to identify differences based on the presence or absence of PAS.
Using the partial least squares regression (PLS-A) method, a distinct cluster of neonatal plasma samples from mothers with PAS was observed, separate from the cluster of samples from mothers without PAS (Figure 1). The most significant contribution to this separation came from the read counts of 42 miRNAs, each with a VIP parameter greater than 1. As shown in Figure 1, all peripheral blood plasma samples of neonates born to mothers with PAS were markedly different from those of neonates born to mothers without PAS. The separation was primarily driven by the following 42 miRNAs: hsa-miR-152-3p, miR-339-3p, miR-675-3p, miR-34c-5p, miR-199a-5p, miR-22-3p, miR-625-5p, miR-625-3p, miR-6511a, miR-101-3p, miR-324-3p, let-7d-5p, miR-339-5p, miR-199a-3p, miR-199b-3p, miR-382-5p, miR-1908-5p, miR-382-3p, miR-30c-5p, miR-485-5p, let-7g-5p, let-7f-5p, miR-493-5p, let-7d-3p, miR-136-3p, miR-330-3p, miR-98-5p, miR-335-3p, miR-127-3p, miR-432-5p, miR-205-5p, miR-1180-3p, miR-1306-3p, miR-326, miR-379-5p, miR-3131, miR-26b-5p, miR-320d, miR-421, miR-3180-3p, and miR-6842-3p, miR-195-3p.
Since antenatal corticosteroid therapy to reduce the incidence of RDS is effective up to seven days after treatment [39], this study aimed to compare the miRNA profiles in the blood plasma of newborns from mothers without PAS who did not receive RDS prophylaxis with those from newborns of mothers with PAS who did receive RDS prophylaxis within the 7-14 days preceding delivery. The goal was to identify marker miRNAs associated with PAS rather than those influenced by antenatal RDS prophylaxis itself. Statistically significant differences were observed between the compared groups in the levels of 38 miRNAs (see Table 1).
Notably, this list included seven miRNAs (hsa-miR-199a-3p, hsa-miR-199b-3p, hsa-miR-382-5p, hsa-let-7g-5p, hsa-let-7f-5p, hsa-let-7d-3p, hsa-miR-26b-5p) that contributed to the separation of two clusters based on the presence or absence of PAS, as determined by PLS-A analysis (Figure 1). Two miRNAs, hsa-miR-199a-3p and hsa-miR-382-5p, were selected for validation of the quantitative real-time RT-PCR sequencing data across all 160 maternal and neonatal plasma samples collected.
3.2. Validation of miRNAs Sequencing Data by Quantitative Real-Time PCR
The levels of hsa-miR-382-5p and hsa-miR-199a-3p were analyzed through quantitative real-time RT-PCR in 160 plasma samples from preterm infants and their mothers across several groups: those without PAS and without RDS prophylaxis (Control without CT), those with PAS (accreta, increta, percreta) without RDS prophylaxis, and those who received antenatal corticosteroid therapy (CT) more than 14 days before delivery, 7-14 days before delivery, and within 7 days before delivery (Figure 2).
Table 2 summarizes the clinical parameters of the newborns from mothers, categorized by the presence or absence of PAS and the timing of antenatal corticosteroid therapy. A statistically significant decrease in RDW-SD and MCV was noted, alongside an increase in weight across all groups of newborns from mothers with PAS, regardless of whether they received corticosteroid therapy. Notably, newborns from mothers with PAS who received CT within 7 days or 7-14 days before delivery exhibited higher weights than those from mothers with PAS who received CT more than 14 days before delivery or did not receive CT at all (Table 2).
Additionally, the PAS group without antenatal CT showed significantly higher intrapartum blood loss in pregnant women (p = 0.033, Table 3) and a lower MCH value in their newborns (p = 0.01, Table 3) compared to the PAS group without CT.
In the analysis of hsa-miR-382-5p levels in neonatal blood plasma (Figure 2A, Table 3), a significant increase was observed in the “accreta without CT” and “increta without CT” groups compared to the “Control without CT” group. Additionally, the timing of antenatal corticosteroid therapy (CT) influenced hsa-miR-382-5p levels in neonates with placenta accreta or increta. Specifically, the groups “without CT” and “CT more than 14 days before delivery” exhibited significant differences from the “CT during 7-14 days before delivery” and “CT during 7 days before delivery” groups. Notably, the hsa-miR-382-5p level in the “CT during 7 days before delivery” group was closest to that in the “Control without CT” group, suggesting that corticosteroid therapy within this timeframe may be more effective. No significant differences were detected among the compared groups of newborns from mothers with placenta percreta, nor were any changes in hsa-miR-382-5p levels found in the blood plasma of mothers depending on the timing of antenatal CT (Figure 2B, Table 4).
Regarding the analysis of hsa-miR-199a-3p levels in neonatal plasma (Figure 2C, Table 3), a significant increase was noted in the “accreta without CT” and “increta without CT” groups compared to the “Control without CT” group. The timing of antenatal CT also impacted hsa-miR-199a-3p levels in neonatal plasma for mothers with placenta accreta or increta. The “CT more than 14 days before delivery” group showed significant differences from the “Control without CT” group, while no significant differences were observed between the “CT during 7-14 days before delivery” or “CT during 7 days before delivery” groups and the “Control without CT” group. Similar to hsa-miR-382-5p, no significant differences were found among the groups of neonates from mothers with placenta percreta. Interestingly, the analysis of hsa-miR-199a-3p in the blood plasma of mothers revealed a significant increase in miRNA levels across all PAS groups (accreta, increta, percreta) compared to the “Control without CT” group, regardless of the timing of CT (Figure 2D, Table 4).
A notable 3.5-4.6-fold increase in hsa-miR-199a-3p levels was detected in neonatal plasma compared to maternal plasma (Figure 3, Table 5). Although significant increases in hsa-miR-199a-3p levels were observed in both neonatal and maternal plasma in cases with PAS (Figure 2C, Table 3, and Figure 2D, Table 4), this increase was more pronounced in maternal plasma. This is evidenced by the relative decrease in hsa-miR-199a-3p levels in neonatal plasma from mothers with PAS compared to neonates from mothers without PAS (Figure 3).
As indicated in Table 5, antenatal CT is most effective when administered 2-7 days or 7-14 days prior to delivery, as there were no significant differences between these groups and the “control without CT” group. In contrast, the “PAS without CT” and “PAS with CT more than 14 days before delivery” groups showed significant differences from the “control without CT” group.
When analyzing newborns with PAS according to the severity score on the Neomod scale, a significant increase in hsa-miR-382-5p levels was observed in the blood plasma of newborns with scores of 2, 4, and 5, compared to those with a score of 0 (Figure 4, Table 6). A similar trend was noted in the quantitative analysis of hsa-miR-199a-3p levels in the blood plasma of newborns, although this did not reach statistical significance (Table 6).
It is important to highlight that the group with a score of 1 on the Neomod scale included newborns with only moderate respiratory dysfunction. In contrast, the group with a score of 2 included newborns either with severe respiratory dysfunction or with a combination of moderate respiratory dysfunction and moderate dysfunction of the cardiovascular or urinary systems. The groups with scores of 4-5 included newborns experiencing severe respiratory dysfunction combined with moderate dysfunction of the cardiovascular and/or urinary systems and/or acid-base balance.
The significant changes in hsa-miR-382-5p levels in the blood plasma of newborns from mothers with PAS, based on the severity of the condition according to the Neomod scale, indicate a relationship between this miRNA and dysfunctions in the respiratory, cardiovascular, and urinary systems.
Using the nonparametric Spearman rank correlation method, the study found several significant relationships regarding the levels of specific miRNAs and clinical parameters in newborns and their mothers:
- a direct correlation between the levels of hsa-miR-382-5p and hsa-miR-199a-3p in the blood plasma of newborns (r = 0.49; p = 0.0001);
- an inverse correlation between the level of hsa-miR-199a-3p in the blood plasma of mothers and their newborns with the depth of trophoblast invasion (r = -0.46; p = 0.0003 for mothers and r = -0.29; p = 0.0285 for newborns);
- an inverse correlation between the level of hsa-miR-199a-3p in the blood plasma of newborns with the volume of maternal blood loss (r = -0.28; p = 0.0321);
- an inverse relationship between hsa-miR-382-5p levels in newborns and their weight (r = -0.39; p = 0.0027) and platelet levels (r = -0.27; p = 0.0426);
- direct relationship between the level of hsa-miR-382-5p in the blood plasma of the newborn and the required fraction of oxygen in the NICU (r = 0.41; p = 0.0016), duration of stay in the NICU (r = 0.31; p = 0.019), and the severity of the newborn's condition according to the NEOMOD scale (r = 0.36; p = 0.0051).
In turn, significant correlations were noted between the required oxygen fraction in the NICU for newborns of mothers with PAS and various hematological parameters, including fetal red blood cell count (r = -0.47; p = 0.0002), hemoglobin (HGB) (r = -0.37; p = 0.0038), hematocrit (r = -0.36; p = 0.0051), and the coefficient of variation of red blood cell distribution width (r = -0.36; p = 0.0052). Additionally, there were strong correlations with the duration of NICU stay (r = 0.71; p = 0), total hospitalization duration (r = 0.49; p = 0.0001), and the severity of the newborn's condition according to the NEOMOD scale (r = 0.68; p = 0).
Based on these correlations, the study aimed to evaluate the potential of using the levels of hsa-miR-199a-3p and hsa-miR-382-5p in maternal blood plasma to predict neonatal complications. Previous meta-analysis results [23] and our own observations [24] indicated that newborns whose mothers received antenatal corticosteroids after 34 weeks of gestation had a significantly lower risk of developing RDS and transient tachypnea of the newborn (TTN), along with reduced surfactant and mechanical ventilation use, shorter durations of oxygen supplementation, lower peak inspired oxygen concentrations, shorter NICU stays, and higher Apgar scores than controls. In this regard, the overall dynamics of changes in the level of hsa-miR-199a-3p and hsa-miR-382-5p in the blood plasma of pregnant women without PAS and in the case of PAS with different timing of antenatal corticosteroid therapy without subdividing into morphological types of PAS was assessed (Figure 5). As illustrated in Figure 5, a significant increase in hsa-miR-199a-3p and hsa-miR-382-5p levels was observed across different PAS groups compared to the control group without PAS (Figure 5, Table 7). It was decided to use hsa-miR-181a-5p as a reference endogenous miRNA instead of the exogenous UniSp6 for the quantitative assessment of hsa-miR-199a-3p and hsa-miR-382-5p in pregnant women's blood when constructing logistic regression models for predicting neonatal complications, since no significant differences in hsa-miR-181a-5p levels were found among the compared groups of maternal blood plasma samples (Figure 5, Table 7) as well as it didn’t contribute to the separation of clusters of neonatal plasma samples from mothers with and without PAS while using PLS-A method (Figure 1).
The probabilities of neonatal complications—specifically respiratory disorders (including RDS, congenital pneumonia, and transient tachypnea) and cardiovascular disorders—were calculated by constructing logistic regression models (see Figure 6A and Figure 6B) based on quantitative real-time PCR data (-ΔCt values). This analysis assessed the levels of miR-199a-3p and/or miR-382-5p in the blood plasma of pregnant women with PAS, using endogenous RNA miR-181a-5p as a reference. In this context, the dependent variable (response variable) was the presence of neonatal complications, coded as follows: 0 for absence of complications and 1 for presence of complications.
The characteristics of these models are detailed in Table 8. Among the constructed models for predicting respiratory disorders in newborns, Model 2 (shown in Figure 6A) demonstrated the best diagnostic value. It can predict, with 100% sensitivity, the need for invasive mechanical ventilation (IMV) or high-frequency oscillatory ventilation (HFOV) in newborns during the early neonatal period, based on the levels of miR-199a-3p and miR-382-5p in the maternal blood plasma shortly before delivery.
For predicting cardiovascular disorders in newborns, Model 1 (illustrated in Figure 6B) also exhibited strong diagnostic value. This model can predict, with 95% sensitivity (as shown in Table 9), the need for cardiotonic therapy for the newborn in the early neonatal period, based solely on the level of miR-199a-3p in the maternal blood plasma prior to delivery.
To understand the role of hsa-miR-382-5p and hsa-miR-199a-3p in the pathogenesis of neonatal complications in newborns of mothers with PAS, we identified their potential and experimentally validated target genes using the miRTargetLink 2.0 program. This was followed by an analysis of the identified gene sets in the FunRich software tool (Version 3.1.3) for functional enrichment, considering a significance threshold of p < 0.05 (Figure 7).
The expression sites of 35-77% of the gene targets for hsa-miR-382-5p and hsa-miR-199a-3p were found across various organs and systems, including the placenta, kidney, lung, heart, uterine corpus, serum, and plasma (Figure 7). In terms of cellular components, 46.50% (p < 0.001) of the gene targets of hsa-miR-382-5p and 50.55% (p < 0.001) of those for hsa-miR-199a-3p were located in the nucleus. Additionally, 45.16% (p < 0.001) and 45.47% (p = 0.001) of the targets were found in the cytoplasm, while 9.23% (p = 0.008) of the gene targets for hsa-miR-199a-3p were located in the Golgi apparatus (Figure 7).
The significantly enriched pathways associated with the gene targets of these miRNAs included the glypican pathway, which is known to regulate cell growth, motility, and differentiation through fibroblast growth factors (FGF), vascular endothelial growth factor-A (VEGF-A), transforming growth factor-β (TGF-β), and Wnt signaling [40]; the mTOR (mammalian target of rapamycin) signaling pathway, which controls cell proliferation, migration, cytoskeleton remodeling, ion transport, and glucose metabolism [41]; pathways involved in inflammatory processes, such as sphingosine 1-phosphate (S1P), thrombin/protease-activated receptor (PAR), endothelin, TGF-beta receptor, and IL-1- and IL-3-mediated signaling pathways; Arf6 signaling events, which play a crucial role in innate immunity and host-pathogen interactions [42]; cell death signaling involving TRAIL and TNF receptors; and LKB1 and IGF1 pathways that regulate lipid, cholesterol, and glucose metabolism [43,44]. Additionally, pathways associated with epithelial-to-mesenchymal transition were identified (Figure 7).
4. Discussion
While maternal outcomes following pregnancies complicated by PAS are well documented, reports on neonatal outcomes in these cases are limited. Previous retrospective studies consistently indicated high rates of admissions to neonatal intensive care units (NICUs) and a significant need for mechanical ventilation in pregnancies affected by PAS [45]. The primary perinatal complications observed in premature infants born to mothers with PAS in this study included transient tachypnea of the newborn (44%), RDS (12%), congenital pneumonia (41%), congenital anemia (20%), and intraventricular hemorrhage (8%). RDS, which results from a primary deficiency of surfactant and the immaturity of lung tissue due to prematurity, along with congenital pneumonia, can lead to the development of acute respiratory distress syndrome (ARDS) [46]. The mortality rate associated with ARDS remains high, accounting for 30% of all fatalities in intensive care units. [47] [48] [49]. Morphologically, RDS and ARDS exhibit similar characteristics, including immaturity and antenatal damage to the structures of the air-blood barrier, as well as pneumonia and pulmonary ischemia with the formation of hyaline membranes [46].
Recently, numerous studies have been published on the molecular mechanisms involved in the pathogenesis and pathophysiology of ARDS, many of which are detailed in a review article by Huang Q. et al.[50]. The author summarizes that lung barrier dysfunction during ARDS results from the death of alveolar epithelial and pulmonary endothelial cells, which can be triggered by apoptosis pathways such as FasL, TNF-α/TNFR1, and TNF-related apoptosis-inducing ligand (TRAIL) signaling events. Additionally, the article discusses various signals that regulate inflammatory processes during ARDS, particularly those known to activate the RhoA/ROCK pathway, including IL-1, TGF-β, thrombin, sphingosine-1 phosphate (S1P), and endothelin-1. It also highlights factors that alter the activity of the PI3K/AKT pathway through the mammalian target of rapamycin (mTOR) or NF-κB, leading to NLRP3 inflammasome activation or increased levels of inflammatory cytokines. Furthermore, the epithelial–mesenchymal transition (EMT) is identified as a major factor contributing to epithelial barrier dysfunction and worsening pulmonary edema through the modulation of Wnt signaling in the alveolar epithelium. This process results in the loss of epithelial morphology and the acquisition of mesenchymal characteristics, along with the expression of profibrotic proteins that contribute to pulmonary fibrosis. In this study, we found that these signaling pathways are potentially regulated by two microRNAs, miR-382-5p and miR-199a-3p, which were significantly elevated in the blood plasma of day-old neonates and/or their mothers with PAS.
In recent times, there has been an increasing emphasis on the role of miRNAs in RDS, particularly through their ability to target specific genes to regulate signaling pathways [51] [52]. Certain miRNAs play significant roles in the inflammatory response associated with ARDS. For instance, miR-199a-3p has been linked to inflammatory lung diseases, including sepsis-induced ARDS [53]. Notably, this miRNA regulates the synthesis and release of various inflammatory mediators by macrophages [54], which account for nearly half of the immune cells in the lungs[55] [56]. Emerging evidence highlights the critical role of extracellular vesicles from alveolar macrophages in the inflammatory processes of ARDS, particularly secretory autophagosomes (SAPs) [57]. One of the regulators of SAP secretion is miR-199a-3p, which influences the expression of the target gene PAK4 [54], a serine/threonine kinase identified as a key regulator of TNF-induced microparticle release [58]. Studies have shown that SAPs derived from alveolar macrophages contribute to ARDS through excessive secretion of IL-1β, which exacerbates inflammation and pathological injury in lung tissue [57]. Overexpression of miR-199a-3p has been observed in the lungs of mice with ARDS, where the miR-199a-3p antagomir significantly inhibited SAP release, while the miR-199a-3p mimetic promoted SAP release in bronchoalveolar lavage fluid (BALF), resulting in alleviation or intensification of LPS-stimulated ARDS, respectively [54]. These results are consistent with findings from this study that noted an increase in hsa-miR-199a-3p levels in the blood plasma of newborns from mothers with PAS. This increase manifests as severe respiratory distress in the early neonatal period, necessitating invasive ventilation or high-frequency ventilation (HFVL).
Another possible pathogenetic mechanism for respiratory disorders in premature infants born to mothers with PAS, particularly concerning the elevated levels of hsa-miR-199a-3p circulating in maternal and fetal blood, is its negative impact on the differentiation of alveolar type II cells, consequently affecting surfactant protein production [59]. The major protein component of pulmonary surfactant, SP-A (a product of the SFTPA gene), is developmentally regulated in fetal lung. It serves as a marker of alveolar type II cell differentiation Additionally, SP-A plays a vital role in innate immunity by enhancing the uptake and destruction of various pathogens by alveolar macrophages [60] [61]. Moreover, it is secreted into the amniotic fluid from the fetal lung, acting as a signaling molecule for the initiation of labor [62] [63] [64].
During a normal pregnancy, there is a developmental decline in the expression of the miR-199a/-214 cluster in the fetal lung, which leads to increased expression of key gene targets responsible for alveolar type II cell differentiation and enhanced SP-A expression by term [59]. This dependence of miR-199a/-214 cluster expression on gestational age can be explained by increased TGF-β signaling during early to mid-gestation, when the fetal lung is relatively hypoxic. This signaling enhances the expression of ZEB1, a transcription factor that stimulates miR-199a/miR-214 cluster expression. As vascularization of the fetal lung increases during the third trimester and near term, heightened oxygen tension leads to decreased TGF-β signaling and repression of ZEB1, resulting in reduced expression of miR-199a/miR-214.Overexpression of miR-199a-3p, -5p, and miR-214 in human fetal lung epithelial cells has been shown to inhibit SP-A expression as well as the expression of transcription factors CREB1 and C/EBPβ, which are crucial for fetal lung development [65] [66]. Interestingly, ZEB1 is an EMT (epithelial-mesenchymal transition) factor that downregulates epithelial genes while activating mesenchymal genes, promoting a highly invasive cell phenotype [67] [68]. This is typical for extravillous trophoblast cells of the placenta in the case of PAS, which exhibit abnormally aggressive EMT that does not cease at the end of the first trimester but continues throughout pregnancy [69] [70].
Thus, the following mechanism of pathogenesis of respiratory disorders in neonates from mothers with PAS. The elevated level of miR-199a-3p in maternal blood plasma in cases of PAS may reflect excessive EMT of extracellular trophoblasts under chronic inflammatory conditions in the uterine decidua due to endometritis, antecedent curettage, or incompetent uterine scars following cesarean sections. According to Kalluri R. [71], macrophages and activated resident fibroblasts secrete growth factors such as TGF-β, chemokines, and matrix metalloproteinases (MMP-2, -3, -9) in these circumstances. The presence of chorionic villi in the layers of the myometrium results in abnormal gas exchange in the maternal-fetal system, creating hypoxic conditions for the fetus, including the lung tissue. Under these conditions, TGF-β signaling in lung tissue increases, raising the expression of ZEB1 and, consequently, hsa-miR-199a-3p, leading to immature lung structures and reduced surfactant synthesis.Additionally, we observe elevated levels of hsa-miR-199a-3p in the blood plasma of neonates. As indicated in Figure 3, in cases of PAS, the level of hsa-miR-199a-3p in maternal blood plasma is higher than that in neonatal blood plasma compared to pregnancies without PAS. This represents an additional negative factor influencing the damage to fetal lung tissue due to circulating maternal hsa-miR-199a-3p. Moreover, this study revealed significant negative correlations between the levels of hsa-miR-199a-3p in maternal and fetal blood plasma and the severity of PAS; specifically, lower levels of hsa-miR-199a-3p in the maternal and fetal bloodstream are associated with deeper placental invasion into the myometrial layers. The elevated level of hsa-miR-382-5p detected in the blood plasma of newborns from mothers with placenta accreta may represent an additional pathogenetic link in the occurrence of neonatal complications. Furthermore, levels of hsa-miR-199a-3p and hsa-miR-382-5p in the blood plasma of newborns were found to correlate significantly and positively with each other. This correlation may be explained by the presence of a common experimentally validated target gene, PTEN (according to miRTargetLink), which is involved in cell functions including proliferation, migration, and metabolism [72]. Dysregulated PTEN expression was found in blastocyst implantation [73], preeclampsia [74] [75], pulmonary diseases [76], and PAS [77]. Localized primarily in the syncytiotrophoblast (STB), endothelial cells surrounding fetal blood vessels, and to a lesser extent in the stroma of normal placenta [77], increased expression of PTEN impairs human trophoblast cell invasion and is associated with the development of preeclampsia [78]. In contrast, PTEN mRNA and protein levels are reduced in placenta tissue affected by PAS compared to normal placenta [77], suggesting its critical role during pregnancy.
It is known that miR-382-5p is a member of the chromosome 14 miRNA cluster (C14MC), which is one of the largest clusters of pregnancy-related miRNAs, comprising 52 miRNAs [28]. This cluster is involved in embryonic development, endothelial cell migration, and angiogenesis during placental development [79]. miR-382-5p, as an ortholog of the C14MC found in equines, has been shown to be enriched in the blood serum of pregnant mares compared to non-pregnant mares [80]. Additionally, aberrant expression of miR-382-5p in rat lung tissues has been reported as a potential cause of bronchopulmonary dysplasia (BRD) through the suppression of M1 macrophage polarization[81,82].
Regarding the regulation of macrophage function, miR-382-5p may play a significant role in the pathogenesis of ARDS, as macrophages are a crucial component of pulmonary innate immunity, comprising nearly half of the immune cells in the lungs, and the balance between M1 and M2 macrophage phenotypes influences the various stages of ARDS [83] [84] [85] [86] [87]. In the acute exudative phase of ARDS, macrophages are predominantly M1-polarized, releasing pro-inflammatory factors that induce a severe inflammatory response. In the later stages of ARDS, macrophages mainly adopt an M2-polarized phenotype, which can lead to pathological fibroplasia and pulmonary fibrosis.
Mechanisms regulating macrophage function involving miR-382-5p have been demonstrated using microglial cells, which are resident macrophages in the central nervous system and perform immune surveillance in the brain and spinal cord [88]. Through the upregulation of Circ_0006640, which can directly sequester miR-382-5p, and the elevation of IGF1, a target of miR-382-5p, microglial cells showed protection from LPS-induced apoptotic, inflammatory, and oxidative injuries. IGF-1 is a major growth hormone critical for prenatal lung growth and organogenesis [89]. Local synthesis of IGF-1 in lung tissue occurs in type II pneumocytes, alveolar macrophages, and mesenchymal cells. In animal models, mutations in the IGF-1 gene disrupt the architecture of lung tissue, leading to atelectatic lungs, respiratory failure, and high postnatal mortality.
In our study, the level of hsa-miR-382-5p in the blood plasma of premature infants born to mothers with PAS was significantly higher in cases where antenatal prophylaxis for RDS was absent or implemented more than 14 days before delivery, compared to premature infants born to mothers without PAS and without antenatal prophylaxis for RDS. The level of hsa-miR-382-5p in the blood plasma of newborns from mothers with PAS tended to normalize after antenatal prophylaxis for RDS 2-14 days before delivery and did not significantly differ from levels in the blood plasma of newborns from mothers without PAS. A markedly increased level of hsa-miR-382-5p in the blood plasma of premature infants from mothers with PAS, particularly in the absence of antenatal prophylaxis for RDS or when implemented more than 14 days before delivery, likely causes a decrease in IGF-1 across various organs and tissues of the newborn, including the lungs. This decrease helps explain the presence of respiratory disorders in this group of patients, as well as the statistically significant correlations between the level of hsa-miR-382-5p in the blood plasma of the newborn and factors such as weight (r = -0.39; p = 0.0027), required oxygen fraction in the NICU (r = 0.41; p = 0.0016), length of stay in the NICU (r = 0.31; p = 0.019), and severity of the newborn's condition according to the NEOMOD scale (r = 0.36; p = 0.0051).
In addition to respiratory support, newborns from mothers with PAS require cardiotonic therapy due to cardiovascular dysfunction. It was found that miRNAs derived from the precursor miR-199a play a key role in maintaining cardiac homeostasis, particularly through the regulation of endothelial nitric oxide synthase (eNOS) in the endothelium [90] [91] [92]. A common mechanism underlying many cardiovascular diseases is endothelial dysfunction, which is characterized by reduced availability of nitric oxide (NO) [93]. It has been demonstrated that inhibition of miR-199a-3p enhances eNOS activity and decreases the degradation of NO, thereby increasing its bioavailability and modulating vascular contractility [92].
Given the relationships identified in this study between the levels of hsa-miR-199a-3p and hsa-miR-382-5p in the blood plasma of pregnant women and their newborns, as well as the severity of respiratory and cardiac disorders during the neonatal period, we constructed logistic regression models to predict these disorders. These models take into account the established roles of these miRNAs in surfactant synthesis by alveolar cells, fetal organogenesis, the formation of proper lung tissue architecture, and the regulation of the cardiovascular system as reported in the literature. The models developed in this study allow for the prediction of the need for cardiotonic therapy and invasive mechanical ventilation (IMV) or high-frequency oscillatory ventilation (HFOV) for newborns in the early neonatal period, with a sensitivity of 95-100%. However, the implementation of these models in clinical practice will require large-scale studies using independent test samples.
Author Contributions
Conceptualization, A.V.T., A.D.N. and E.N.B.; methodology, A.V.T. and I.S.F.; software, I.S.F.; validation, A.V.T., A.D.N. and E.N.B.; formal analysis, D.N.D.; investigation, A.V.T., I.S.F. and A.M.T.; resources, A.V.T. and A.D.N.; data curation, A.V.T., E.N.B., and D.N.D.; writing—original draft preparation, A.V.T.; writing—review and editing, A.V.T.; visualization, A.V.T. and I.S.F.; supervision, G.T.S.; project administration, A.V.T.; funding acquisition, A.V.T. and G.T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Russian Science Foundation under grant no. 22-15-00363 “Epigenetic and biochemical aspects of the pathology of pregnancy in violations of the invasive properties of the trophoblast: from early diagnosis to the prevention of maternal and perinatal morbidity”.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and was approved by the Institutional Ethics Committee of National Medical Research Center for Obstetrics, Gynecology, and Perinatology, named after the Academician V.I. Kulakov of Ministry of Healthcare of the Russian Federation (protocol No. 11, approval date: 11 November 2021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tantbirojn, P.; Crum, C.P.; Parast, M.M. Pathophysiology of placenta creta: the role of decidua and extravillous trophoblast. Placenta 2008, 29, 639–645. [Google Scholar] [CrossRef]
- Obstetric Care Consensus, No. 7: Placenta Accreta Spectrum. Obstet. Gynecol. 2018, 132, e259–e275. [Google Scholar] [CrossRef]
- Jauniaux, E.; Bunce, C.; Grønbeck, L.; Langhoff-Roos, J. Prevalence and main outcomes of placenta accreta spectrum: a systematic review and meta-analysis. Am. J. Obstet. Gynecol. 2019, 221, 208–218. [Google Scholar] [CrossRef]
- Hull, A.D.; Moore, T.R. Multiple repeat cesareans and the threat of placenta accreta: incidence, diagnosis, management. Clin. Perinatol. 2011, 38, 285–296. [Google Scholar] [CrossRef]
- Baldwin, H.J.; Patterson, J.A.; Nippita, T.A.; Torvaldsen, S.; Ibiebele, I.; Simpson, J.M.; Ford, J.B. Antecedents of Abnormally Invasive Placenta in Primiparous Women: Risk Associated With Gynecologic Procedures. Obstet. Gynecol. 2018, 131, 227–233. [Google Scholar] [CrossRef]
- Wu, S.; Kocherginsky, M.; Hibbard, J.U. Abnormal placentation: twenty-year analysis. Am. J. Obstet. Gynecol. 2005, 192, 1458–1461. [Google Scholar] [CrossRef] [PubMed]
- Spillane, N.T.; Zamudio, S.; Alvarez-Perez, J.; Andrews, T.; Nyirenda, T.; Alvarez, M.; Al-Khan, A. Increased incidence of respiratory distress syndrome in neonates of mothers with abnormally invasive placentation. PLoS One 2018, 13, e0201266. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J.L.; Kimura, A.M.; Julia, J.; Tunnell, C.; Hernandez, B.; Curbelo, J.; Ramsey, P.S.; Ireland, K.E. Impact of placenta accreta spectrum (PAS) pathology on neonatal respiratory outcomes in cesarean hysterectomies. J. Matern. neonatal Med. Off. J. Eur. Assoc. Perinat. Med. Fed. Asia Ocean. Perinat. Soc. Int. Soc. Perinat. Obstet. 2022, 35, 10692–10697. [Google Scholar] [CrossRef]
- Haram, K.; Mortensen, J.H.S.; Wollen, A.-L. Preterm delivery: an overview. Acta Obstet. Gynecol. Scand. 2003, 82, 687–704. [Google Scholar] [CrossRef]
- Patel, R.M. Short- and Long-Term Outcomes for Extremely Preterm Infants. Am. J. Perinatol. 2016, 33, 318–328. [Google Scholar] [CrossRef]
- Doyle, L.W. Outcome at 5 years of age of children 23 to 27 weeks’ gestation: refining the prognosis. Pediatrics 2001, 108, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Saigal, S.; Doyle, L.W. An overview of mortality and sequelae of preterm birth from infancy to adulthood. Lancet (London, England) 2008, 371, 261–269. [Google Scholar] [CrossRef]
- Balashova E.N., Ionov O.Balashova E.N., Ionov O.V. The features of respiratory and cardiovascular disorders in preterm infants born to mothers with abnormally invasive placenta. Obstet. Gynegology 2021, 5. [Google Scholar] [CrossRef]
- Parsons, A.; Netsanet, A.; Seedorf, G.; Abman, S.H.; Taglauer, E.S. Understanding the role of placental pathophysiology in the development of bronchopulmonary dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 323, L651–L658. [Google Scholar] [CrossRef]
- Wallace, B.; Peisl, A.; Seedorf, G.; Nowlin, T.; Kim, C.; Bosco, J.; Kenniston, J.; Keefe, D.; Abman, S.H. Anti-sFlt-1 Therapy Preserves Lung Alveolar and Vascular Growth in Antenatal Models of Bronchopulmonary Dysplasia. Am. J. Respir. Crit. Care Med. 2018, 197, 776–787. [Google Scholar] [CrossRef]
- Tang, J.-R.; Karumanchi, S.A.; Seedorf, G.; Markham, N.; Abman, S.H. Excess soluble vascular endothelial growth factor receptor-1 in amniotic fluid impairs lung growth in rats: linking preeclampsia with bronchopulmonary dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L36–46. [Google Scholar] [CrossRef]
- Jobe, A.H.; Soll, R.F. Choice and dose of corticosteroid for antenatal treatments. Am. J. Obstet. Gynecol. 2004, 190, 878–881. [Google Scholar] [CrossRef]
- Ballard, P.L.; Ballard, R.A. Scientific basis and therapeutic regimens for use of antenatal glucocorticoids. Am. J. Obstet. Gynecol. 1995, 173, 254–262. [Google Scholar] [CrossRef]
- Williams, M.J.; Ramson, J.A.; Brownfoot, F.C. Different corticosteroids and regimens for accelerating fetal lung maturation for babies at risk of preterm birth. Cochrane database Syst. Rev. 2022, 8, CD006764. [Google Scholar] [CrossRef]
- Roberts, D.; Brown, J.; Medley, N.; Dalziel, S.R. Antenatal corticosteroids for accelerating fetal lung maturation for women at risk of preterm birth. Cochrane database Syst. Rev. 2017, 3, CD004454. [Google Scholar] [CrossRef]
- Walters, A.; McKinlay, C.; Middleton, P.; Harding, J.E.; Crowther, C.A. Repeat doses of prenatal corticosteroids for women at risk of preterm birth for improving neonatal health outcomes. Cochrane database Syst. Rev. 2022, 4, CD003935. [Google Scholar] [CrossRef] [PubMed]
- Gyamfi-Bannerman, C.; Thom, E.A.; Blackwell, S.C.; Tita, A.T.N.; Reddy, U.M.; Saade, G.R.; Rouse, D.J.; McKenna, D.S.; Clark, E.A.S.; Thorp, J.M.J.; et al. Antenatal Betamethasone for Women at Risk for Late Preterm Delivery. N. Engl. J. Med. 2016, 374, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Saccone, G.; Berghella, V. Antenatal corticosteroids for maturity of term or near term fetuses: systematic review and meta-analysis of randomized controlled trials. BMJ 2016, 355, i5044. [Google Scholar] [CrossRef] [PubMed]
- Nikonets, A.D. , Balashova E.N., Ionov O.V., Kirtbaya A.R., Zubkov V.V., Shmakov R.G., D.D.N. Prevention of respiratory disorders in late preterm neonates born to mothers with abnormally invasive placenta. Obstet. Gynegology 2024, 1. [Google Scholar] [CrossRef]
- Timofeeva, A. V; Fedorov, I.S.; Suhova, Y. V; Tarasova, A.M.; Ezhova, L.S.; Zabelina, T.M.; Vasilchenko, O.N.; Ivanets, T.Y.; Sukhikh, G.T. Diagnostic Role of Cell-Free miRNAs in Identifying Placenta Accreta Spectrum during First-Trimester Screening. Int. J. Mol. Sci. 2024, 25. [Google Scholar] [CrossRef]
- Timofeeva, A. V; Fedorov, I.S.; Pirogova, M.M.; Vasilchenko, O.N.; Chagovets, V. V; Ezhova, L.S.; Zabelina, T.M.; Shmakov, R.G.; Sukhikh, G.T. Clusterin and Its Potential Regulatory microRNAs as a Part of Secretome for the Diagnosis of Abnormally Invasive Placenta: Accreta, Increta, and Percreta Cases. Life (Basel, Switzerland) 2021, 11. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Morales-Prieto, D.M.; Ospina-Prieto, S.; Chaiwangyen, W.; Schoenleben, M.; Markert, U.R. Pregnancy-associated miRNA-clusters. J. Reprod. Immunol. 2013, 97, 51–61. [Google Scholar] [CrossRef]
- Reza, A.M.M.T.; Choi, Y.-J.; Han, S.G.; Song, H.; Park, C.; Hong, K.; Kim, J.-H. Roles of microRNAs in mammalian reproduction: from the commitment of germ cells to peri-implantation embryos. Biol. Rev. Camb. Philos. Soc. 2019, 94, 415–438. [Google Scholar] [CrossRef]
- Hayder, H.; O’Brien, J.; Nadeem, U.; Peng, C. MicroRNAs: crucial regulators of placental development. Reproduction 2018, 155, R259–R271. [Google Scholar] [CrossRef]
- Timofeeva, A. V; Fedorov, I.S.; Shamina, M.A.; Chagovets, V. V; Makarova, N.P.; Kalinina, E.A.; Nazarenko, T.A.; Sukhikh, G.T. Clinical Relevance of Secreted Small Noncoding RNAs in an Embryo Implantation Potential Prediction at Morula and Blastocyst Development Stages. Life (Basel, Switzerland) 2021, 11. [Google Scholar] [CrossRef]
- Zhao, Z.; Moley, K.H.; Gronowski, A.M. Diagnostic potential for miRNAs as biomarkers for pregnancy-specific diseases. Clin. Biochem. 2013, 46, 953–960. [Google Scholar] [CrossRef]
- Timofeeva, A. V; Fedorov, I.S.; Sukhova, Y. V; Ivanets, T.Y.; Sukhikh, G.T. Prediction of Early- and Late-Onset Pre-Eclampsia in the Preclinical Stage via Placenta-Specific Extracellular miRNA Profiling. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef]
- Timofeeva, A. V; Fedorov, I.S.; Brzhozovskiy, A.G.; Bugrova, A.E.; Chagovets, V. V; Volochaeva, M. V; Starodubtseva, N.L.; Frankevich, V.E.; Nikolaev, E.N.; Shmakov, R.G.; et al. miRNAs and Their Gene Targets-A Clue to Differentiate Pregnancies with Small for Gestational Age Newborns, Intrauterine Growth Restriction, and Preeclampsia. Diagnostics (Basel, Switzerland) 2021, 11. [Google Scholar] [CrossRef]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. MicroRNA profiling: approaches and considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef]
- Gantier, M.P.; McCoy, C.E.; Rusinova, I.; Saulep, D.; Wang, D.; Xu, D.; Irving, A.T.; Behlke, M.A.; Hertzog, P.J.; Mackay, F.; et al. Analysis of microRNA turnover in mammalian cells following Dicer1 ablation. Nucleic Acids Res. 2011, 39, 5692–5703. [Google Scholar] [CrossRef]
- Team, R.C. A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria Available online:. Available online: https://www.r-project.org (accessed on Mar 10, 2021).
- Team, Rs. RStudio: Integrated Development for R. Available online: http://www.rstudio.com/. (accessed on Mar 23, 2021).
- McGoldrick, E.; Stewart, F.; Parker, R.; Dalziel, S.R. Antenatal corticosteroids for accelerating fetal lung maturation for women at risk of preterm birth. Cochrane database Syst. Rev. 2020, 12, CD004454. [Google Scholar] [CrossRef]
- Pan, J.; Ho, M. Role of glypican-1 in regulating multiple cellular signaling pathways. Am. J. Physiol. Cell Physiol. 2021, 321, C846–C858. [Google Scholar] [CrossRef]
- Panwar, V.; Singh, A.; Bhatt, M.; Tonk, R.K.; Azizov, S.; Raza, A.S.; Sengupta, S.; Kumar, D.; Garg, M. Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal Transduct. Target. Ther. 2023, 8, 375. [Google Scholar] [CrossRef]
- Van Acker, T.; Tavernier, J.; Peelman, F. The Small GTPase Arf6: An Overview of Its Mechanisms of Action and of Its Role in Host−Pathogen Interactions and Innate Immunity. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Shackelford, D.B.; Shaw, R.J. The LKB1-AMPK pathway: metabolism and growth control in tumour suppression. Nat. Rev. Cancer 2009, 9, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, A. Insulin-Like Growth Factor 1 (IGF-1) Signaling in Glucose Metabolism in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Toussia-Cohen, S.; Castel, E.; Friedrich, L.; Mor, N.; Ohayon, A.; Levin, G.; Meyer, R. Neonatal outcomes in pregnancies complicated by placenta accreta- a matched cohort study. Arch. Gynecol. Obstet. 2024, 310, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Perepelitsa, S.A. Acute Respiratory Distress Syndrome in Preterm Newborns (Morphological Study). Gen. Reanimatol. 2020, 16. [Google Scholar] [CrossRef]
- Heidemann, S.M.; Nair, A.; Bulut, Y.; Sapru, A. Pathophysiology and Management of Acute Respiratory Distress Syndrome in Children. Pediatr. Clin. North Am. 2017, 64, 1017–1037. [Google Scholar] [CrossRef]
- Johnson, E.R.; Matthay, M.A. Acute lung injury: epidemiology, pathogenesis, and treatment. J. Aerosol Med. Pulm. Drug Deliv. 2010, 23, 243–252. [Google Scholar] [CrossRef]
- Ragaller, M.; Richter, T. Acute lung injury and acute respiratory distress syndrome. J. Emerg. Trauma. Shock 2010, 3, 43–51. [Google Scholar] [CrossRef]
- Huang, Q.; Le, Y.; Li, S.; Bian, Y. Signaling pathways and potential therapeutic targets in acute respiratory distress syndrome (ARDS). Respir. Res. 2024, 25, 30. [Google Scholar] [CrossRef]
- Lu, Q.; Yu, S.; Meng, X.; Shi, M.; Huang, S.; Li, J.; Zhang, J.; Liang, Y.; Ji, M.; Zhao, Y.; et al. MicroRNAs: Important Regulatory Molecules in Acute Lung Injury/Acute Respiratory Distress Syndrome. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Rajasekaran, S.; Pattarayan, D.; Rajaguru, P.; Sudhakar Gandhi, P.S.; Thimmulappa, R.K. MicroRNA Regulation of Acute Lung Injury and Acute Respiratory Distress Syndrome. J. Cell. Physiol. 2016, 231, 2097–2106. [Google Scholar] [CrossRef]
- Liu, Y.; Guan, H.; Zhang, J.-L.; Zheng, Z.; Wang, H.-T.; Tao, K.; Han, S.-C.; Su, L.-L.; Hu, D. Acute downregulation of miR-199a attenuates sepsis-induced acute lung injury by targeting SIRT1. Am. J. Physiol. Cell Physiol. 2018, 314, C449–C455. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, X.; Dong, X.; Yang, Y.; Liu, L. MiR-199a-3p-regulated alveolar macrophage-derived secretory autophagosomes exacerbate lipopolysaccharide-induced acute respiratory distress syndrome. Front. Cell. Infect. Microbiol. 2022, 12, 1061790. [Google Scholar] [CrossRef]
- Deprez, M.; Zaragosi, L.-E.; Truchi, M.; Becavin, C.; Ruiz García, S.; Arguel, M.-J.; Plaisant, M.; Magnone, V.; Lebrigand, K.; Abelanet, S.; et al. A Single-Cell Atlas of the Human Healthy Airways. Am. J. Respir. Crit. Care Med. 2020, 202, 1636–1645. [Google Scholar] [CrossRef]
- Jardine, L.; Haniffa, M. Human lung macrophages: roll up for the MISTRG tour. Immunity 2021, 54, 194–196. [Google Scholar] [CrossRef]
- Xu, X.; Liu, X.; Dong, X.; Qiu, H.; Yang, Y.; Liu, L. Secretory Autophagosomes from Alveolar Macrophages Exacerbate Acute Respiratory Distress Syndrome by Releasing IL-1β. J. Inflamm. Res. 2022, 15, 127–140. [Google Scholar] [CrossRef]
- Zhang, S.; Yin, Y.; Li, C.; Zhao, Y.; Wang, Q.; Zhang, X. PAK4 suppresses TNF-induced release of endothelial microparticles in HUVECs cells. Aging (Albany. NY). 2020, 12, 12740–12749. [Google Scholar] [CrossRef]
- Mishra, R.; Benlhabib, H.; Guo, W.; Lerma Cervantes, C.B.; Mendelson, C.R. Developmental Decline in the MicroRNA 199a (miR-199a)/miR-214 Cluster in Human Fetal Lung Promotes Type II Cell Differentiation by Upregulating Key Transcription Factors. Mol. Cell. Biol. 2018, 38. [Google Scholar] [CrossRef]
- Wright, J.R. Immunoregulatory functions of surfactant proteins. Nat. Rev. Immunol. 2005, 5, 58–68. [Google Scholar] [CrossRef]
- Lawson, P.R.; Reid, K.B. The roles of surfactant proteins A and D in innate immunity. Immunol. Rev. 2000, 173, 66–78. [Google Scholar] [CrossRef]
- Condon, J.C.; Jeyasuria, P.; Faust, J.M.; Mendelson, C.R. Surfactant protein secreted by the maturing mouse fetal lung acts as a hormone that signals the initiation of parturition. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 4978–4983. [Google Scholar] [CrossRef]
- Montalbano, A.P.; Hawgood, S.; Mendelson, C.R. Mice deficient in surfactant protein A (SP-A) and SP-D or in TLR2 manifest delayed parturition and decreased expression of inflammatory and contractile genes. Endocrinology 2013, 154, 483–498. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Rabbitt, E.H.; Condon, J.C.; Renthal, N.E.; Johnston, J.M.; Mitsche, M.A.; Chambon, P.; Xu, J.; O’Malley, B.W.; Mendelson, C.R. Steroid receptor coactivators 1 and 2 mediate fetal-to-maternal signaling that initiates parturition. J. Clin. Invest. 2015, 125, 2808–2824. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.D.; Flecknoe, S.J.; Tan, K.H.; Olsson, P.F.; Antony, N.; Mantamadiotis, T.; Mollard, R.; Hooper, S.B.; Cole, T.J. cAMP response element binding protein is required for differentiation of respiratory epithelium during murine development. PLoS One 2011, 6, e17843. [Google Scholar] [CrossRef] [PubMed]
- Roos, A.B.; Berg, T.; Barton, J.L.; Didon, L.; Nord, M. Airway epithelial cell differentiation during lung organogenesis requires C/EBPα and C/EBPβ. Dev. Dyn. an Off. Publ. Am. Assoc. Anat. 2012, 241, 911–923. [Google Scholar] [CrossRef]
- Garg, M. Targeting microRNAs in epithelial-to-mesenchymal transition-induced cancer stem cells: Therapeutic approaches in cancer. Expert Opin. Ther. Targets 2015, 19, 285–297. [Google Scholar] [CrossRef]
- Greening, D.W.; Gopal, S.K.; Mathias, R.A.; Liu, L.; Sheng, J.; Zhu, H.-J.; Simpson, R.J. Emerging roles of exosomes during epithelial-mesenchymal transition and cancer progression. Semin. Cell Dev. Biol. 2015, 40, 60–71. [Google Scholar] [CrossRef]
- Bartels, H.C.; Postle, J.D.; Downey, P.; Brennan, D.J. Placenta accreta spectrum: A review of pathology, molecular biology, and biomarkers. Dis. Markers 2018, 2018. [Google Scholar] [CrossRef]
- Das, V.; Bhattacharya, S.; Chikkaputtaiah, C.; Hazra, S.; Pal, M. The basics of epithelial–mesenchymal transition (EMT): A study from a structure, dynamics, and functional perspective. J. Cell. Physiol. 2019, 234, 14535–14555. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Invest. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Chen, J.; He, L.; Stiles, B.L. PTEN: Tumor Suppressor and Metabolic Regulator. Front. Endocrinol. (Lausanne). 2018, 9, 338. [Google Scholar] [CrossRef]
- Makker, A.; Goel, M.M.; Nigam, D.; Mahdi, A.A.; Das, V.; Agarwal, A.; Pandey, A.; Gautam, A. Aberrant Akt Activation During Implantation Window in Infertile Women With Intramural Uterine Fibroids. Reprod. Sci. 2018, 25, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Tao, T.; Yin, Y.; Zhao, L.; Yang, L.; Hu, L. miR-144 may regulate the proliferation, migration and invasion of trophoblastic cells through targeting PTEN in preeclampsia. Biomed. Pharmacother. 2017, 94, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Lou, C.-X.; Zhou, X.-T.; Tian, Q.-C.; Xie, H.-Q.; Zhang, J.-Y. Low expression of microRNA-21 inhibits trophoblast cell infiltration through targeting PTEN. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6181–6189. [Google Scholar] [CrossRef]
- Tian, Y.; Li, H.; Qiu, T.; Dai, J.; Zhang, Y.; Chen, J.; Cai, H. Loss of PTEN induces lung fibrosis via alveolar epithelial cell senescence depending on NF-κB activation. Aging Cell 2019, 18, e12858. [Google Scholar] [CrossRef]
- Murrieta-Coxca, J.M.; Barth, E.; Fuentes-Zacarias, P.; Gutiérrez-Samudio, R.N.; Groten, T.; Gellhaus, A.; Köninger, A.; Marz, M.; Markert, U.R.; Morales-Prieto, D.M. Identification of altered miRNAs and their targets in placenta accreta. Front. Endocrinol. (Lausanne). 2023, 14, 1021640. [Google Scholar] [CrossRef]
- Xue, P.; Zheng, M.; Diao, Z.; Shen, L.; Liu, M.; Gong, P.; Sun, H.; Hu, Y. miR-155* mediates suppressive effect of PTEN 3’-untranslated region on AP-1/NF-κB pathway in HTR-8/SVneo cells. Placenta 2013, 34, 650–656. [Google Scholar] [CrossRef]
- Dini, P.; Daels, P.; Loux, S.C.; Esteller-Vico, A.; Carossino, M.; Scoggin, K.E.; Ball, B.A. Kinetics of the chromosome 14 microRNA cluster ortholog and its potential role during placental development in the pregnant mare. BMC Genomics 2018, 19, 954. [Google Scholar] [CrossRef]
- Dini, P.; El-Sheikh Ali, H.; Carossino, M.; C Loux, S.; Esteller-Vico, A.; E Scoggin, K.; Daels, P.; A Ball, B. Expression Profile of the Chromosome 14 MicroRNA Cluster (C14MC) Ortholog in Equine Maternal Circulation throughout Pregnancy and Its Potential Implications. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Xing, Y.; Fu, J.; Yang, H.; Yao, L.; Qiao, L.; Du, Y.; Xue, X. MicroRNA expression profiles and target prediction in neonatal Wistar rat lungs during the development of bronchopulmonary dysplasia. Int. J. Mol. Med. 2015, 36, 1253–1263. [Google Scholar] [CrossRef]
- Lv, Y.; Li, Y.; Wang, J.; Li, M.; Zhang, W.; Zhang, H.; Shen, Y.; Li, C.; Du, Y.; Jiang, L. MiR-382-5p suppresses M1 macrophage polarization and inflammatory response in response to bronchopulmonary dysplasia through targeting CDK8: Involving inhibition of STAT1 pathway. Genes Cells 2021, 26, 772–781. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, H.; Chen, B. Identification of macrophage-related genes in sepsis-induced ARDS using bioinformatics and machine learning. Sci. Rep. 2023, 13, 9876. [Google Scholar] [CrossRef] [PubMed]
- Dang, W.; Tao, Y.; Xu, X.; Zhao, H.; Zou, L.; Li, Y. The role of lung macrophages in acute respiratory distress syndrome. Inflamm. Res. Off. J. Eur. Histamine Res. Soc.... [et al.] 2022, 71, 1417–1432. [Google Scholar] [CrossRef]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef]
- Chen, X.; Tang, J.; Shuai, W.; Meng, J.; Feng, J.; Han, Z. Macrophage polarization and its role in the pathogenesis of acute lung injury/acute respiratory distress syndrome. Inflamm. Res. Off. J. Eur. Histamine Res. Soc.... [et al.] 2020, 69, 883–895. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.-A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Yang, D.; Wei, H.; Sheng, Y.; Peng, T.; Zhao, Q.; Xie, L.; Yang, J. Circ_0006640 transferred by bone marrow-mesenchymal stem cell-exosomes suppresses lipopolysaccharide-induced apoptotic, inflammatory and oxidative injury in spinal cord injury. J. Orthop. Surg. Res. 2024, 19, 50. [Google Scholar] [CrossRef]
- Hellström, A.; Ley, D.; Hansen-Pupp, I.; Hallberg, B.; Löfqvist, C.; van Marter, L.; van Weissenbruch, M.; Ramenghi, L.A.; Beardsall, K.; Dunger, D.; et al. Insulin-like growth factor 1 has multisystem effects on foetal and preterm infant development. Acta Paediatr. 2016, 105, 576–586. [Google Scholar] [CrossRef]
- Catalucci, D.; Latronico, M.V.G.; Condorelli, G. MicroRNAs control gene expression: importance for cardiac development and pathophysiology. Ann. N. Y. Acad. Sci. 2008, 1123, 20–29. [Google Scholar] [CrossRef]
- Neppl, R.L.; Wang, D.-Z. The myriad essential roles of microRNAs in cardiovascular homeostasis and disease. Genes Dis. 2014, 1, 18–39. [Google Scholar] [CrossRef]
- Joris, V.; Gomez, E.L.; Menchi, L.; Lobysheva, I.; Di Mauro, V.; Esfahani, H.; Condorelli, G.; Balligand, J.-L.; Catalucci, D.; Dessy, C. MicroRNA-199a-3p and MicroRNA-199a-5p Take Part to a Redundant Network of Regulation of the NOS (NO Synthase)/NO Pathway in the Endothelium. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2345–2357. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Gladwin, M.T.; Weitzberg, E. Strategies to increase nitric oxide signalling in cardiovascular disease. Nat. Rev. Drug Discov. 2015, 14, 623–641. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PLS-A analysis of deep sequencing data of miRNA in the peripheral blood plasma of day-old newborns from mothers with PAS and without PAS (control).
Figure 1.
PLS-A analysis of deep sequencing data of miRNA in the peripheral blood plasma of day-old newborns from mothers with PAS and without PAS (control).
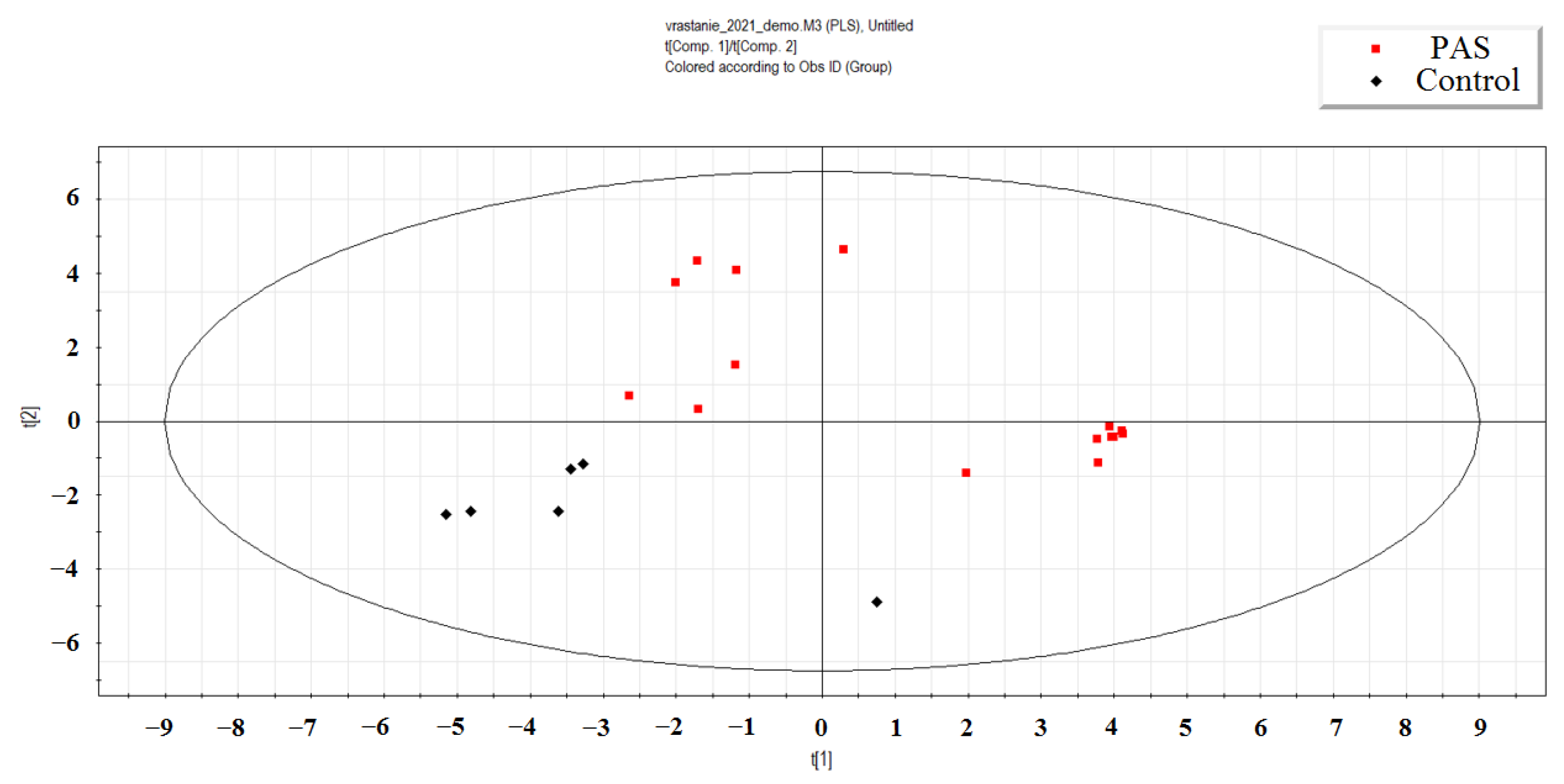
Figure 2.
The dependence of hsa-miR-382-5p and hsa-miR-199a-3p content in the blood plasma of newborns and their mothers on the severity of placenta accreta spectrum (PAS) and the timing of corticosteroid therapy (CT).
Figure 2.
The dependence of hsa-miR-382-5p and hsa-miR-199a-3p content in the blood plasma of newborns and their mothers on the severity of placenta accreta spectrum (PAS) and the timing of corticosteroid therapy (CT).

Figure 3.
Dynamics of changes in hsa-miR-199a-3p levels in the blood plasma of newborns compared to their mothers' blood plasma, with and without PAS, depending on the timing of antenatal corticosteroid therapy (CT).
Figure 3.
Dynamics of changes in hsa-miR-199a-3p levels in the blood plasma of newborns compared to their mothers' blood plasma, with and without PAS, depending on the timing of antenatal corticosteroid therapy (CT).
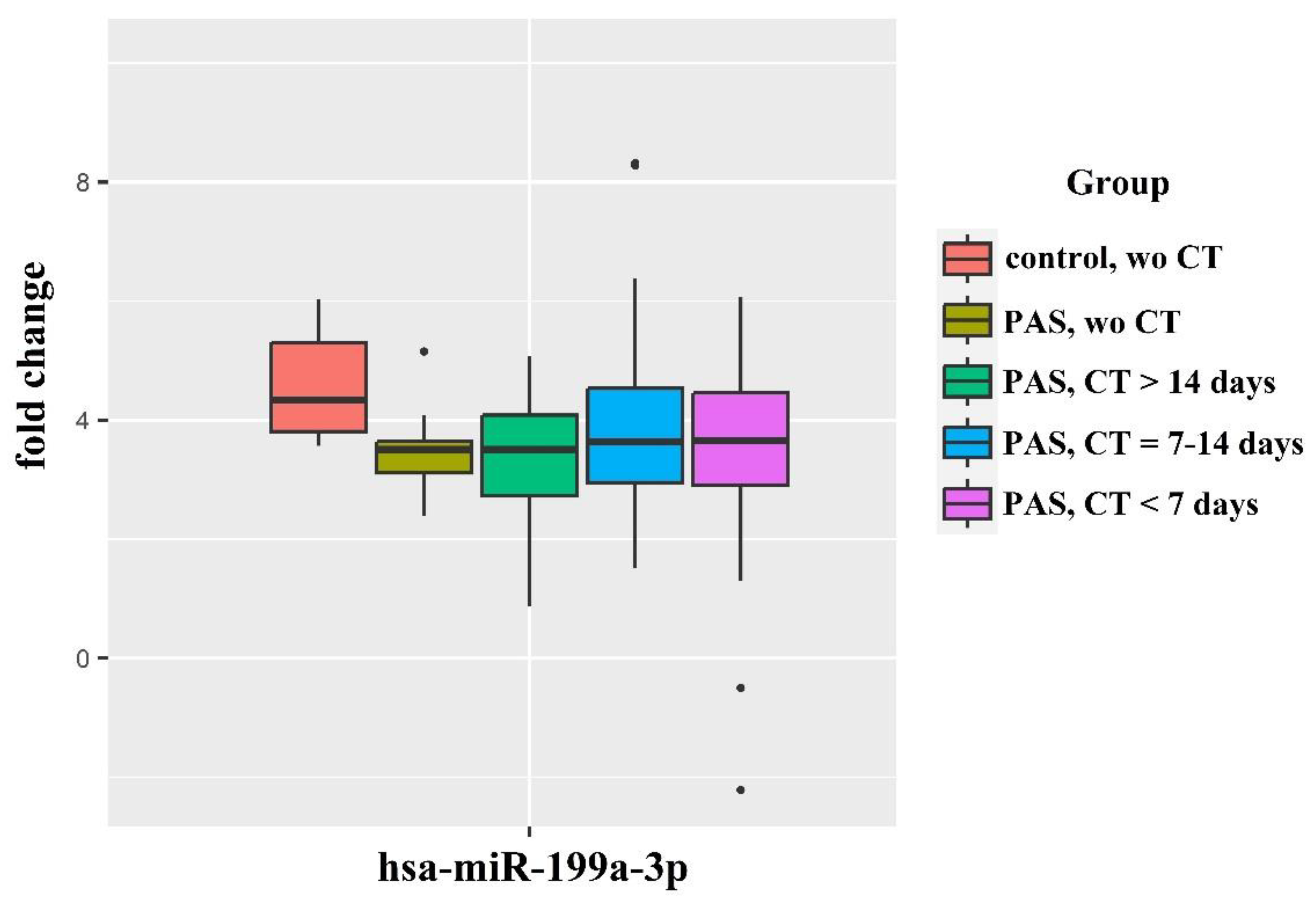
Figure 4.
Levels of hsa-miR-199a-3p and hsa-miR-382-5p in the blood plasma of newborns with PAS, categorized by their severity score according to the Neomod scale.
Figure 4.
Levels of hsa-miR-199a-3p and hsa-miR-382-5p in the blood plasma of newborns with PAS, categorized by their severity score according to the Neomod scale.

Figure 5.
Levels of miR-181a-5p, miR-199a-3p and miR-382-5p in blood plasma of pregnant women with/without PAS and with/without antenatal corticosteroid therapy.
Figure 5.
Levels of miR-181a-5p, miR-199a-3p and miR-382-5p in blood plasma of pregnant women with/without PAS and with/without antenatal corticosteroid therapy.

Figure 6.
Logistic regression models for predicting neonatal complications by plasma miR-199a-3p and/or miR-382-5p levels in pregnant women with PAS using miR-181a-5p as a reference endogenous RNA. (A) Respiratory complications probability models. (B) Cardiovascular complications probability models.
Figure 6.
Logistic regression models for predicting neonatal complications by plasma miR-199a-3p and/or miR-382-5p levels in pregnant women with PAS using miR-181a-5p as a reference endogenous RNA. (A) Respiratory complications probability models. (B) Cardiovascular complications probability models.
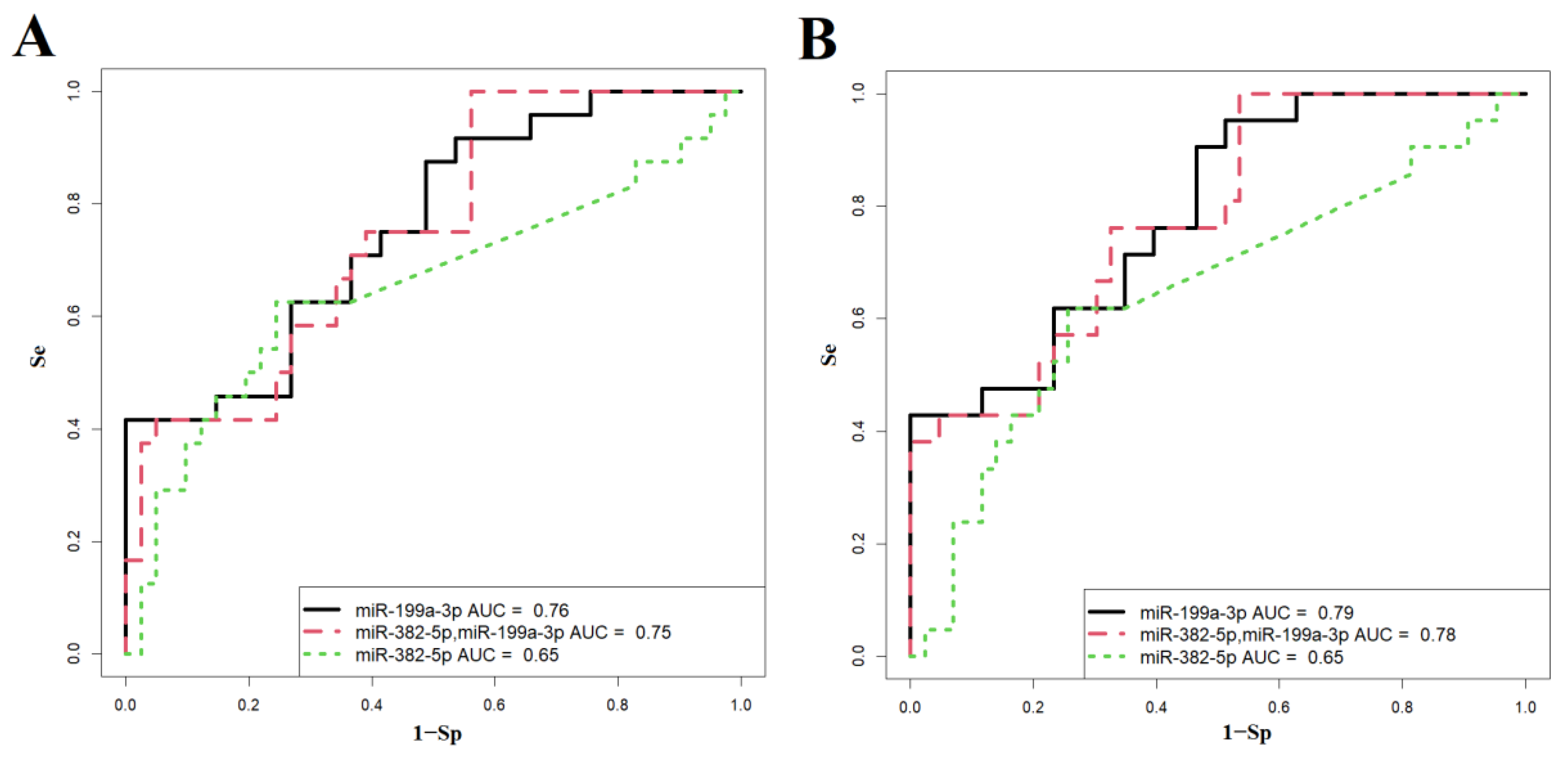
Figure 7.
Enrichment analysis of gene-targets of hsa-miR-382-5p and hsa-miR-199a-3p using FunRich software tool.
Figure 7.
Enrichment analysis of gene-targets of hsa-miR-382-5p and hsa-miR-199a-3p using FunRich software tool.


Table 1.
Statistically significant changes in the level of miRNA in the blood plasma of newborns from mothers with PAS and RDS prophylaxis 7-14 days before delivery compared to newborns from mothers without PAS and without RDS prophylaxis.
Table 1.
Statistically significant changes in the level of miRNA in the blood plasma of newborns from mothers with PAS and RDS prophylaxis 7-14 days before delivery compared to newborns from mothers without PAS and without RDS prophylaxis.
| miRNA | baseMean | log2FoldChange | lfcSE | p-value | |
| 1 | hsa-miR-215-5p | 98.71519 | 5.819687 | 1.265354 | 4.24E-06 |
| 2 | hsa-miR-516b-5p | 215.1577 | 5.248942 | 1.166537 | 6.81E-06 |
| 3 | hsa-miR-182-5p | 55.26162 | 4.71655 | 1.105862 | 2.00E-05 |
| 4 | hsa-miR-183-5p | 143.454 | 4.126976 | 1.034286 | 6.60E-05 |
| 5 | hsa-miR-192-5p | 503.9136 | 1.635789 | 0.46376 | 0.00042 |
| 6 | hsa-miR-1323 | 30.67836 | 3.847168 | 1.192338 | 0.001253 |
| 7 | hsa-miR-760 | 15.02699 | -3.21625 | 1.009398 | 0.001441 |
| 8 | hsa-let-7f-5p | 992.7216 | 2.217035 | 0.745282 | 0.002932 |
| 9 | hsa-miR-26a-5p | 1195.92 | 1.756481 | 0.610171 | 0.003994 |
| 10 | hsa-miR-199a-3p | 320.7235 | -1.81477 | 0.635226 | 0.004278 |
| 11 | hsa-miR-200c-3p | 121.4978 | -4.12153 | 1.450851 | 0.004501 |
| 12 | hsa-miR-199b-3p | 160.3617 | -1.7853 | 0.631627 | 0.004706 |
| 13 | hsa-let-7g-5p | 1207.634 | 1.872991 | 0.679862 | 0.00587 |
| 14 | hsa-miR-10a-5p | 1121.615 | 2.724361 | 1.00493 | 0.006708 |
| 15 | hsa-miR-146b-5p | 130.2105 | 1.470428 | 0.550239 | 0.007532 |
| 16 | hsa-miR-99b-3p | 8.964436 | -3.41312 | 1.28876 | 0.008088 |
| 17 | hsa-miR-218-5p | 9.343058 | -4.06315 | 1.600639 | 0.011134 |
| 18 | hsa-miR-150-5p | 24.77196 | 1.48045 | 0.631319 | 0.019026 |
| 19 | hsa-miR-29a-3p | 35.61049 | 1.99514 | 0.86954 | 0.021763 |
| 20 | hsa-miR-181b-5p | 124.9393 | -2.39485 | 1.093218 | 0.028478 |
| 21 | hsa-miR-378c | 8.895485 | 1.825745 | 0.838232 | 0.029399 |
| 22 | hsa-miR-26b-5p | 102.9558 | 1.269355 | 0.58317 | 0.029507 |
| 23 | hsa-miR-30e-3p | 45.69012 | 1.591149 | 0.740817 | 0.031727 |
| 24 | hsa-miR-483-3p | 37.44106 | 2.100013 | 0.979484 | 0.032033 |
| 25 | hsa-miR-194-5p | 209.4698 | 1.657005 | 0.780456 | 0.033743 |
| 26 | hsa-miR-99a-5p | 1362.079 | -1.48541 | 0.714392 | 0.037592 |
| 27 | hsa-miR-2110 | 38.68886 | -1.90428 | 0.917955 | 0.038034 |
| 28 | hsa-let-7d-3p | 244.3945 | 1.280001 | 0.628354 | 0.041643 |
| 29 | hsa-miR-382-5p | 125.1091 | -2.21816 | 1.205711 | 0.04581 |
Table 2.
Clinical characteristics of premature infants in mothers with and without antenatal corticosteroid therapy, along with comparisons of the corresponding groups using the Mann-Whitney U test.
Table 2.
Clinical characteristics of premature infants in mothers with and without antenatal corticosteroid therapy, along with comparisons of the corresponding groups using the Mann-Whitney U test.
| Clinical parameters | Control, without CT (n=11) | PAS, Without CT (n=10) | “Control without CT” vs “PAS without CT” | PAS, CT more than 14 days before delivery (n=13) | “Control, without CT” vs “PAS, CT more than 14 days before delivery” | PAS, CT during 7-14 days before delivery (n=25) | “Control, without CT” vs PAS from 7 to 14 | PAS, CT during 7 days before delivery (n=21) | “Control, without CT” vs “PAS, CT during 7 days before delivery” |
| Me(Q1;Q3) | Me(Q1;Q3) | p-value | Me(Q1;Q3) | p-value | Me(Q1;Q3) | p-value | Me(Q1;Q3) | p-value | |
| Mother’sbloodless during delivery | 750(750;825) | 1350(812.5;2675) | 0.033 | 800(700;1200) | 0.841 | 800(750;1000) | 0.279 | 800(750;1000) | 0.419 |
| Weight of newborn, g | 2250(1965;2437.5) | 2795.5(2542;3042.25) | 0.001 | 2520(2390;2652) | 0.089 | 2863(2780;3030) | <0.001 | 2850(2730;2960) | 0.001 |
| Apgar score, 1 min | 8(7.5;8) | 7(7;8) | 0.205 | 8(7;8) | 0.702 | 8(7;8) | 0.606 | 8(7;8) | 0.973 |
| Apgar score, 5 min | 8(8;9) | 8(8;8) | 0.084 | 8(8;8) | 0.067 | 8(8;9) | 0.425 | 8(8;9) | 0.447 |
| WBC | 11.42(9.75;12.68) | 12.25(9.94;18.02) | 0.417 | 10.46(9.39;13.35) | 0.757 | 14.11(9.5;16.98) | 0.207 | 13.28(10.64;16.5) | 0.189 |
| ACHN | 4225(3806.5;4561) | 4776.5(3236.25;8941) | 0.475 | 3872(3448;5440) | 0.937 | 5664(4323;7874) | 0.148 | 6190(4131;7722) | 0.155 |
| Ni | 0.07(0.04;0.08) | 0.05(0.02;0.11) | 0.659 | 0.06(0.03;0.09) | 0.781 | 0.07(0.03;0.11) | 0.714 | 0.06(0.05;0.09) | 0.979 |
| RBС | 4.51(4.36;4.84) | 4.78(4.11;4.9) | 1 | 4.76(4.42;4.89) | 0.938 | 4.46(4.06;4.83) | 0.48 | 4.66(4.45;4.84) | 0.75 |
| RDW-CV | 16(15.35;17.2) | 15.75(15.27;16.28) | 0.769 | 15.8(15.4;16.6) | 0.721 | 15.8(15.4;16.1) | 0.437 | 15.8(15.3;16.5) | 0.652 |
| RDW-SD | 63.1(61.9;67.95) | 57.45(51.85;59.35) | 0.007 | 58.8(55.9;60.4) | 0.047 | 58.9(56.7;59.7) | 0.009 | 60.1(57.7;62.9) | 0.08 |
| MCV | 105.8(105;108.3) | 98(95.38;102.12) | 0.001 | 101.4(99.4;103.2) | 0.008 | 102.2(98.5;103.3) | 0.002 | 101.9(100.4;105.6) | 0.027 |
| HGB, g/L | 163(155.5;180.5) | 161(145.5;167.75) | 0.806 | 168(158;179) | 0.936 | 158(146;173) | 0.583 | 168(161;171) | 0.121 |
| MCH | 36.6(35.8;38.2) | 35.05(34;35.4) | 0.01 | 36.2(35.2;36.7) | 0.427 | 35.5(35.1;36.5) | 0.068 | 35.9(35.1;36.6) | 0.185 |
| MCHC | 34.6(34.55;34.95) | 35.45(35.05;36.22) | 0.05 | 35.7(35.2;36.1) | 0.039 | 35.4(35;35.7) | 0.079 | 35.1(34.6;35.6) | 0.287 |
| HTC | 47.3(45.1;52.15) | 42.75(40.07;49.5) | 0.13 | 47.2(45.1;49.8) | 0.606 | 44.8(41.2;50.6) | 0.171 | 47.7(46.4;48.9) | 0.958 |
| Platelets | 324(288;356) | 323(280.25;399) | 0.696 | 281(224;335) | 0.428 | 354(317;402) | 0.092 | 339(296;413) | 0.533 |
| MPV | 9.7(9.05;9.95) | 9.45(9.2;9.67) | 0.302 | 9.8(9.4;10) | 0.72 | 9.5(8.9;10) | 1 | 9.6(9;10.1) | 0.811 |
| PTC | 0.31(0.26;0.38) | 0.3(0.27;0.37) | 0.883 | 0.28(0.22;0.32) | 0.341 | 0.35(0.3;0.38) | 0.283 | 0.34(0.28;0.37) | 0.594 |
| PDW | 10.4(9.55;10.55) | 9.7(8.98;10.88) | 0.807 | 10.2(9.5;10.7) | 0.873 | 9.1(8.6;10) | 0.273 | 9.8(9;10.1) | 0.381 |
| PLCR | 22.3(17.6;24) | 19.95(18.5;23.18) | 0.66 | 22.8(19.2;24.5) | 0.751 | 19.7(15.9;24.2) | 0.789 | 21(17.8;25.1) | 1 |
| DHR | 2(1;4) | 4.5(3;6) | 0.115 | 5(2;6) | 0.118 | 2(2;4) | 0.591 | 2(2;3) | 0.978 |
| HD | 13(9;14.5) | 10(8;14) | 0.305 | 11(11;13) | 0.937 | 10(7;15) | 0.315 | 9(7;11) | 0.77 |
*PAS, placenta accreta spectrum; CT, corticosteroid therapy; WBC, white blood cells; ACHN, absolute neutrophil count; Ni, neutrophil index; RBС, red blood cells; RDW-CV, RBC distribution width, the coefficient of variation; RDW-SD, RBC distribution width, standard deviation; MCV, mean corpuscular volume; MCH, mean concentration hemoglobin; MCHC, mean corpuscular hemoglobin concentration; HTC, hematocrit; MPV, mean platelets volume; PTC, thrombocrit; PDW, platelet distribution width; PLCR, percentage of giant (˃12µm) platelets (%); DHR, length of stay in the NICU, days; HD, duration of hospitalization, days. The clinical blood test data and the weight of the day-old newborn are presented in the Table.
Table 3.
Quantitative RT-PCR data assessing hsa-miR-382-5p and hsa-miR-199a-3p levels in the blood plasma of newborns from mothers without PAS in the absence of antenatal corticosteroid therapy, as well as from mothers with PAS without CT or with CT administered during different time periods.
Table 3.
Quantitative RT-PCR data assessing hsa-miR-382-5p and hsa-miR-199a-3p levels in the blood plasma of newborns from mothers without PAS in the absence of antenatal corticosteroid therapy, as well as from mothers with PAS without CT or with CT administered during different time periods.
| RT-PCR data, -ΔCt | p-value, Mann-Whitney U test | |||||||
| Group | Me | Q1 | Q3 | Without CT | Without CT | CT more than 14 days before delivery | CT during 7-14 days before delivery | CT during 7 days before delivery |
| miR-382-5p | ||||||||
| control | placenta accreta | |||||||
| Control, without CT | -13.28 | -13.34 | -12.81 | 1 | 0.0471 | 0.2 | 0.486 | 0.415 |
| placenta accreta, without CT | -10.28 | -10.74 | -10.05 | 0.0471 | 1 | 0.1 | 0.049 | 0.05 |
| placenta accreta, CT more than 14 days before delivery | -11.66 | -11.67 | -11.47 | 0.2 | 0.1 | 1 | 0.8 | 0.25 |
| placenta accreta, CT during 7-14 days before delivery | -11.68 | -12.36 | -11.32 | 0.486 | 0.049 | 0.8 | 1 | 0.9 |
| placenta accreta, CT during 7 days before delivery | -12.02 | -12.89 | -11.72 | 0.415 | 0.05 | 0.25 | 0.9 | 1 |
| control | placenta increta | |||||||
| Control, without CT | -13.28 | -13.34 | -12.81 | 1 | 0.0491 | 0.016 | 0.079 | 0.19 |
| placenta increta, without CT | -10.38 | -11.06 | -10.07 | 0.0491 | 1 | 0.7 | 0.1 | 0.1 |
| placenta increta, CT more than 14 days before delivery | -10.51 | -10.98 | -9.78 | 0.016 | 0.7 | 1 | 0.008 | 0.03 |
| placenta increta, CT during 7-14 days before delivery | -11.31 | -12.01 | -11.11 | 0.079 | 0.1 | 0.008 | 1 | 0.5 |
| placenta increta, CT during 7 days before delivery | -12.13 | -12.15 | -11.65 | 0.19 | 0.1 | 0.03 | 0.5 | 1 |
| control | Placenta percreta | |||||||
| Control, without CT | -13.28 | -13.34 | -12.81 | 1 | 0.2 | 0.171 | 0.28 | 0.226 |
| placenta percreta, without CT | -11.89 | -12.48 | -11.35 | 0.2 | 1 | 0.7 | 0.55 | 0.785 |
| placenta percreta, CT more than 14 days before delivery | -11.33 | -12.11 | -10.89 | 0.171 | 0.7 | 1 | 0.122 | 0.462 |
| placenta percreta, CT during 7-14 days before delivery | -12.5 | -13 | -11.98 | 0.28 | 0.55 | 0.122 | 1 | 0.101 |
| placenta percreta, CT during 7 days before delivery | -11.89 | -11.99 | -11.72 | 0.226 | 0.785 | 0.462 | 0.101 | 1 |
| miR-199a-3p | ||||||||
| control | placenta accreta | |||||||
| Control, without CT | -11.44 | -11.87 | -10.91 | 1 | 0.05 | 0.05 | 0.1 | 0.28 |
| placenta accreta, without CT | -9.72 | -9.86 | -9.72 | 0.05 | 1 | 0.7 | 0.2 | 0.9 |
| placenta accreta, CT more than 14 days before delivery | -9.91 | -9.96 | -9.87 | 0.05 | 0.7 | 1 | 0.6 | 0.3 |
| placenta accreta, CT during 7-14 days before delivery | -10.13 | -10.38 | -9.72 | 0.1 | 0.2 | 0.6 | 1 | 0.9 |
| placenta accreta, CT during 7 days before delivery | -10.07 | -10.41 | -9.99 | 0.28 | 0.9 | 0.3 | 0.9 | 1 |
| control | placenta increta | |||||||
| Control, without CT | -11.44 | -11.87 | -10.91 | 1 | 0.05 | 0.05 | 0.1 | 0.06 |
| placenta accreta, without CT | -9.72 | -10.05 | -9.38 | 0.05 | 1 | 0.7 | 0.07 | 0.45 |
| placenta accreta, CT more than 14 days before delivery | -9.51 | -10.08 | -8.39 | 0.05 | 0.7 | 1 | 0.04 | 0.3 |
| placenta accreta, CT during 7-14 days before delivery | -10.43 | -10.54 | -9.87 | 0.1 | 0.07 | 0.04 | 1 | 0.1 |
| placenta accreta, CT during 7 days before delivery | -9.82 | -10.14 | -9.63 | 0.06 | 0.45 | 0.3 | 0.1 | 1 |
| control | placenta percreta | |||||||
| Control, without CT | -11.44 | -11.87 | -10.91 | 1 | 0.6 | 0.35 | 0.4 | 0.6 |
| placenta accreta, without CT | -10.51 | -10.96 | -10.44 | 0.6 | 1 | 0.9 | 0.6 | 0.9 |
| placenta accreta, CT more than 14 days before delivery | -11.09 | -11.49 | -10.12 | 0.35 | 0.9 | 1 | 0.8 | 0.8 |
| placenta accreta, CT during 7-14 days before delivery | -10.32 | -11.46 | -9.89 | 0.4 | 0.6 | 0.8 | 1 | 0.5 |
| placenta accreta, CT during 7 days before delivery | -10.59 | -11.94 | -10.24 | 0.6 | 0.9 | 0.8 | 0.5 | 1 |
Table 4.
The quantitative RT-PCR data for evaluating hsa-miR-382-5p and hsa-miR-199a-3p levels in the blood plasma of pregnant women without PAS in the absence of antenatal CT, in mothers with PAS without CT, or with CT at various time points.
Table 4.
The quantitative RT-PCR data for evaluating hsa-miR-382-5p and hsa-miR-199a-3p levels in the blood plasma of pregnant women without PAS in the absence of antenatal CT, in mothers with PAS without CT, or with CT at various time points.
| RT-PCR data, -ΔCt | p-value, Mann-Whitney U test | |||||||
| Group | Me | Q1 | Q3 | Without CT | Without CT | CT more than 14 days before delivery | CT during 7-14 days before delivery | CT during 7 days before delivery |
| hsa-miR-382-5p | ||||||||
| control | placenta accreta | |||||||
| Control, without CT | -19.23 | -19.35 | -19.06 | 1 | 0.0167 | 0.9 | 0.23 | 0.106 |
| placenta accreta, without CT | -17.05 | -17.06 | -15.68 | 0.0167 | 1 | 0.1 | 0.161 | 0.39 |
| placenta accreta, CT more than 14 days before delivery | -19.16 | -19.27 | -19.11 | 0.9 | 0.1 | 1 | 0.22 | 0.25 |
| placenta accreta, CT during 7-14 days before delivery | -18.67 | -19.1 | -18.24 | 0.23 | 0.161 | 0.22 | 1 | 0.9 |
| placenta accreta. CT during 7 days before delivery | -18.92 | -19.13 | -18.2 | 0.106 | 0.39 | 0.25 | 0.9 | 1 |
| control | placenta increta | |||||||
| Control, without CT | -19.23 | -19.35 | -19.06 | 1 | 0.41 | 0.1 | 0.07 | 0.0177 |
| placenta accreta, without CT | -19.13 | -19.22 | -19.01 | 0.41 | 1 | 0.413 | 0.304 | 0.111 |
| placenta accreta, CT more than 14 days before delivery | -18.81 | -19.2 | -18.11 | 0.1 | 0.413 | 1 | 0.9 | 0.548 |
| placenta accreta, CT during 7-14 days before delivery | -18.9 | -19.08 | -16.86 | 0.07 | 0.304 | 0.9 | 1 | 0.513 |
| placenta accreta, CT during 7 days before delivery | -18.64 | -18.99 | -16.07 | 0.0177 | 0.111 | 0.548 | 0.513 | 1 |
| control | placenta percreta | |||||||
| Control, without CT | -19.23 | -19.35 | -19.06 | 1 | 0.383 | 0.731 | 0.0853 | 0.0268 |
| placenta accreta, without CT | -19.14 | -19.17 | -19.06 | 0.383 | 1 | 0.905 | 0.368 | 0.291 |
| placenta accreta, CT more than 14 days before delivery | -19.18 | -19.32 | -16.11 | 0.731 | 0.905 | 1 | 0.808 | 0.216 |
| placenta accreta, CT during 7-14 days before delivery | -18.98 | -19.1 | -18.36 | 0.0853 | 0.368 | 0.808 | 1 | 0.606 |
| placenta accreta, CT during 7 days before delivery | -18.9 | -19.07 | -15.96 | 0.0268 | 0.291 | 0.216 | 0.606 | 1 |
| hsa-miR-199a-3p | ||||||||
| control | placenta accreta | |||||||
| Control, without CT | -15.52 | -15.88 | -15.3 | 1 | 0.015 | 0.0167 | 0.006 | 0.002 |
| placenta accreta, without CT | -12.88 | -13.53 | -12.67 | 0.015 | 1 | 0.7 | 0.161 | 0.786 |
| placenta accreta, CT more than 14 days before delivery | -13.69 | -13.77 | -13.33 | 0.0167 | 0.7 | 1 | 0.629 | 0.786 |
| placenta accreta, CT during 7-14 days before delivery | -12.96 | -13.33 | -12.67 | 0.006 | 0.161 | 0.629 | 1 | 0.19 |
| placenta accreta, CT during 7 days before delivery | -13.41 | -13.85 | -13.39 | 0.002 | 0.786 | 0.786 | 0.19 | 1 |
| control | placenta increta | |||||||
| Control, without CT | -15.52 | -15.88 | -15.3 | 1 | 0.00606 | 0.04 | 0.0185 | 0.00253 |
| placenta accreta, without CT | -13.54 | -14.11 | -12.84 | 0.00606 | 1 | 0.9 | 0.188 | 0.905 |
| placenta accreta, CT more than 14 days before delivery | -13.09 | -13.46 | -12.39 | 0.04 | 0.9 | 1 | 0.206 | 0.841 |
| placenta accreta, CT during 7-14 days before delivery | -14.37 | -15.01 | -13.32 | 0.0185 | 0.188 | 0.206 | 1 | 0.31 |
| placenta accreta, CT during 7 days before delivery | -13.84 | -14.09 | -13.04 | 0.00253 | 0.905 | 0.841 | 0.31 | 1 |
| control | placenta percreta | |||||||
| Control, without CT | -15.52 | -15.88 | -15.3 | 1 | 0.0167 | 0.05 | 0.0346 | 0.0154 |
| placenta accreta, without CT | -14.72 | -14.78 | -14.59 | 0.0167 | 1 | 0.7 | 0.885 | 0.291 |
| placenta accreta, CT more than 14 days before delivery | -14.87 | -15.17 | -13.85 | 0.05 | 0.7 | 1 | 0.961 | 0.462 |
| placenta accreta, CT during 7-14 days before delivery | -14.56 | -15.27 | -14.08 | 0.0346 | 0.885 | 0.961 | 1 | 0.3 |
| placenta accreta, CT during 7 days before delivery | -14.13 | -14.78 | -13.59 | 0.0154 | 0.291 | 0.462 | 0.3 | 1 |
Table 5.
Relative content of hsa-miR-199a-3p in the blood plasma of newborns from pregnant women without PAS in the absence of corticosteroid therapy, as well as in the blood plasma of newborns from pregnant women with PAS with or without CT administered during different time periods.
Table 5.
Relative content of hsa-miR-199a-3p in the blood plasma of newborns from pregnant women without PAS in the absence of corticosteroid therapy, as well as in the blood plasma of newborns from pregnant women with PAS with or without CT administered during different time periods.
| RT-PCR data, -ΔCt | p-value, Mann-Whitney U test | |||||||
| group | Ме | Q1 | Q3 | Control, without CT | PAS, without CT | PAS, CT more than 14 days before delivery | PAS, CT during 7-14 days before delivery | PAS, CT during 7 days before delivery |
| Control, without CT | 4.34 | 3.81 | 5.3 | 1 | 0.005 | 0.037 | 0.2 | 0.09 |
| PAS, without CT | 3.51 | 3.12 | 3.64 | 0.005 | 1 | 0.93 | 0.68 | 0.66 |
| PAS, CT more than 14 days before delivery | 3.51 | 2.73 | 4.09 | 0.037 | 0.93 | 1 | 0.55 | 0.8 |
| PAS, CT during 7-14 days before delivery | 3.64 | 2.95 | 4.53 | 0.2 | 0.68 | 0.55 | 1 | 0.58 |
| PAS, CT during 7 days before delivery | 3.66 | 2.91 | 4.46 | 0.09 | 0.66 | 0.8 | 0.58 | 1 |
Table 6.
Comparison of newborns groups from mothers with PAS based on the levels of hsa-miR-199a-3p and hsa-miR-382-5p relative to their scores on the Neomod scale.
Table 6.
Comparison of newborns groups from mothers with PAS based on the levels of hsa-miR-199a-3p and hsa-miR-382-5p relative to their scores on the Neomod scale.
| miR-382-5p | miR-199a-3p | |||||||
| RT-PCR data, -ΔCt | p-value, Mann-Whitney U test | ОТ-ПЦР данные, -ΔCt | p-value, Mann-Whitney U test | |||||
| Groups according to the Neomod scale | Me | Q1 | Q3 | Neomod, 0 | Me | Q1 | Q3 | Neomod, 0 |
| Neomod, 0 | -12.15 | -12.81 | -11.89 | 1 | -10.36 | -11.08 | -10.13 | 1 |
| Neomod, 1 | -11.75 | -12.81 | -11.08 | 0.251 | -10.3 | -11.12 | -9.68 | 0.672 |
| Neomod, 2 | -11.25 | -11.59 | -10.13 | 0.073 | -9.72 | -10.17 | -9.38 | 0.1807 |
| Neomod, 4 | -11.21 | -11.64 | -10.82 | 0.0134 | -10.25 | -10.58 | -9.82 | 0.8868 |
| Neomod, 5 | -11.43 | -11.68 | -10.88 | 0.0503 | -10.15 | -10.98 | -9.45 | 0.927 |
| Neomod, > 4 | -11.23 | -11.65 | -10.82 | 0.0096 | -10.24 | -10.58 | -9.81 | 0.855 |
Table 7.
Comparison of groups of pregnant women by the level of hsa-miR-382-5p, hsa-miR-199a-3p and hsa-miR-181a-5p depending on the presence of PAS and the time of corticosteroid therapy.
Table 7.
Comparison of groups of pregnant women by the level of hsa-miR-382-5p, hsa-miR-199a-3p and hsa-miR-181a-5p depending on the presence of PAS and the time of corticosteroid therapy.
| Group | miR-382-5p | miR-199a-3p | miR-181a-5p | |||||||||
| RT-PCR data, -ΔCt | p-value, Mann-Whitney U test | RT-PCR data, -ΔCt | p-value, Mann-Whitney U test | RT-PCR data, -ΔCt | p-value, Mann-Whitney U test | |||||||
| Me | Q1 | Q3 | Control, without CT | Me | Q1 | Q3 | Control, without CT | Me | Q1 | Q3 | Control, without CT | |
| Control, without CT | -19.23 | -19.35 | -19.06 | 1 | -15.52 | -15.88 | -15.3 | 1 | -16.12 | -17.05 | -15.99 | 1 |
| PAS, without CT | -18.93 | -19.12 | -17.35 | 0.02 | -14.13 | -14.39 | -12.91 | <0.001 | -15.72 | -19.03 | -14.48 | 0.66 |
| PAS, CT more than 14 days before delivery | -19.13 | -19.3 | -18.29 | 0.3 | -13.61 | -14.89 | -13.18 | 0.004 | -15.63 | -17.87 | -14.45 | 0.255 |
| PAS, CT during 7-14 days before delivery | -18.98 | -19.1 | -18.31 | 0.03 | -14.25 | -15.01 | -13.22 | 0.003 | -17.03 | -18.79 | -15.03 | 0.89 |
| PAS, CT during 7 days before delivery | -18.9 | -19.09 | -16.07 | 0.007 | -13.9 | -14.43 | -13.39 | <0.001 | -18.49 | -19.13 | -15.2 | 0.53 |
Table 8.
Parameters of the logistic regression models presented in Figure 6A and Figure 6B.
| Figure 6A | Wald | p_value | coefficients | threshold | sensitivity | specificity |
| 1 model | 0.642 | 0.4167 | 1 | |||
| (Intercept) | 1.879 | 0.060 | 0.974 | |||
| miR-199a-3p | -3.281 | 0.001 | -0.548 | |||
| 2 model | 0.2028 | 1 | 0.439 | |||
| (Intercept) | 1.706 | 0.088 | 1.540 | |||
| miR-382-5p | 0.796 | 0.426 | 0.119 | |||
| miR-199a-3p | -2.662 | 0.008 | -0.699 | |||
| 3 model | 0.4223 | 0.625 | 0.7561 | |||
| (Intercept) | -2.616 | 0.009 | -0.804 | |||
| miR-382-5p | -2.049 | 0.040 | -0.206 | |||
| Рисунoк 6В | Wald | p_value | coefficients | threshold | sensitivity | specificity |
| 1 model | 0.16 | 0.95 | 0.49 | |||
| (Intercept) | 1.887 | 0.050 | 1.046 | |||
| miR-199a-3p | -3.473 | 0.001 | -0.635 | |||
| 2 model | 0.15 | 1 | 0.47 | |||
| (Intercept) | 2.005 | 0.045 | 2.127 | |||
| miR-382-5p | 1.282 | 0.200 | 0.217 | |||
| miR-199a-3p | -2.940 | 0.003 | -0.924 | |||
| 3 model | 0.38 | 0.62 | 0.74 | |||
| (Intercept) | -3.092 | 0.002 | -1.002 | |||
| miR-382-5p | -2.031 | 0.042 | -0.217 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



