
Submitted:
27 October 2024
Posted:
28 October 2024
You are already at the latest version
Abstract
Phytophthora-induced crop diseases, commonly known as “plant plagues,” pose a significant threat to global food security. In this study, strain ASG80 was isolated from sisal roots and demonstrated broad-spectrum antagonistic activity against several Phytophthora species and fungal pathogens. Strain ASG80 was identified as Streptomyces luteireticuli via phylogenetic analysis, digital DNA-DNA hybridization (dDDH), and average nucleotide identity (ANI). Whole-genome sequencing identified 40 biosynthetic gene clusters (BGCs) related to secondary metabolite production, including antimicrobial compounds. Strain ASG80 extract exhibited broad-spectrum inhibitory activity against Phytophthora nicotianae, P. vignae, P. cinnamomi, and P. sojae. Pot experiments showed that strain ASG80 extract significantly reduced sisal zebra disease incidence, with efficacy comparable to the fungicide metalaxyl. These findings suggest that strain ASG80 is a promising biocontrol agent with substantial potential for managing Phytophthora-related diseases in agriculture.
Keywords:
Phytophthora
; Streptomyces
; antimicrobial activity
; biocontrol agent
; whole genome sequencing
1. Introduction
Phytophthora, a genus of oomycetes, is widely recognized for causing severe plant diseases that have a substantial impact on global agriculture [1]. This genus, often referred to as the "Plant Destroyer," currently encompasses over 400 described species and subspecies (www.mycobank.org, accessed August 2024). In recent years, the incidence of diseases caused by Phytophthora has been increasing, a trend attributed to climate change and intensified agricultural activities [2]. The pathogens within this genus exhibit varying degrees of virulence and host specificity, resulting in a diverse range of effects on crops. For example, Phytophthora infestans, the primary pathogen responsible for potato late blight, instigated the Irish potato famine in the 19th century and continues to cause an estimated annual economic loss of over $6 billion globally [3]. P. ramorum, the causal agent of sudden oak death, has significantly disrupted forest ecosystems throughout North America [4]. P. cinnamomi, a major pathogen of root rot in avocados, poses a substantial threat to the global avocado industry [5]. Additionally, P. capsici infects a range of economically important vegetables, causing root, stem, leaf, and fruit rot. This pathogen has a broad host range exceeding 50 cultivated plant species, leading to considerable economic losses [6,7]. P. sojae is responsible for root and stem rot in soybeans, causing estimated global losses ranging from $1 to $2 billion annually [8]. P. nicotianae has adversely affected numerous economically significant crops, with tobacco production losses in North Carolina, USA, alone exceeding $30 million annually [9]. Lastly, P. palmivora is a major pathogen affecting tropical crops, most notably causing bud rot in African oil palms, a disease ranked among the most destructive in this category [10]. In efforts to manage Phytophthora-induced diseases, researchers have investigated a range of strategies. Chemical control remains among the most widely used methods; however, prolonged application of single-target pesticides may result in resistance and residue accumulation, raising concerns regarding food safety and human health. Consequently, biological control has become an essential alternative to chemical fungicides in agricultural practices [11].
Biological control, as an environmentally sustainable approach to disease management, has gained substantial attention in recent years. Biological control involves the use of biological agents, such as beneficial microorganisms or their metabolites, to suppress or control plant pathogens, offering multiple advantages. Firstly, biocontrol is environmentally friendly. In contrast to chemical pesticides, biological control agents generally exert a lower environmental impact, thereby reducing the risks of soil and water pollution [12,13]. Secondly, biocontrol effectively addresses challenges associated with pathogen resistance. Long-term use of chemical pesticides often results in the development of pathogen resistance. In contrast, biocontrol strategies employ multiple mechanisms—such as competition, antagonism, and induced resistance—which collectively reduce the likelihood of resistance evolution [14,15,16,17]. Additionally, biocontrol is safe and sustainable. Many biocontrol agents are safe for humans and animals, and they can be integrated with other agricultural practices as part of an integrated disease management strategy [18,19]. Lastly, biocontrol agents often offer long-term disease control by persisting in the soil and continuing to combat pathogens over extended periods [19,20]. Currently, agricultural biocontrol methods include microbial control, natural enemies, insect pathogens, plant extracts, and insect pheromones. Microbial control has attracted significant attention in plant disease management. Microbial antagonists, which include beneficial bacteria, fungi, and viruses, are utilized to control plant diseases [21]. Certain secondary metabolites produced by these microorganisms have been proven effective in managing plant pathogens. For instance, secondary metabolites from Trichoderma spp. demonstrate strong antimicrobial activity against Lasiodiplodia theobromae, Xanthomonas campestris, and Meloidogyne incognita [22,23]. Additionally, metabolites produced by Bacillus subtilis JF-4, B. amylum JF-5 and Pseudomonas aeruginosa have shown efficacy in controlling banana anthracnose [24,25].
Research has shown that secondary metabolites from Streptomyces species exhibit broad-spectrum antibacterial, antiviral, and anthelmintic properties, highlighting the potential of Streptomyces as biocontrol agents [26,27,28]. For instance, Park solated Streptomyces roseoflavus LS-A24, which produces staurosporine in its fermentation broth, exhibiting inhibitory activity against Phytophthora capsici, the pathogen responsible for pepper blight [29]. Sun reported an exopolysaccharide, EPS66A, from Streptomyces sp. HL-66, which induces resistance in tobacco against Tobacco Mosaic Virus [30]. Moreover, Wang isolated novel macrocyclic lactones from S. avermitilis NEAU1069 that show strong acaricidal and anthelmintic activities [31]. Arasu isolated a novel polyketide compound from Streptomyces sp. AP-123, which displays potent broad-spectrum antibacterial, antifungal, and cytotoxic activities, comparable to erythromycin [32]. Therefore, the utilization of Streptomyces species for plant disease control emerges as a promising strategy.
In this study, we isolated and screened an antimicrobial strain, ASG80, from the roots of sisal. Whole-genome sequencing was performed on strain ASG80 to predict potential biocontrol mechanisms, likely linked to secondary metabolites with antifungal properties. Subsequently, the strain ASG80 extract was evaluated against plant pathogens, demonstrating a broad inhibitory spectrum against Phytophthora species and indicating its potential as a biocontrol agent. Additionally, pot trials were conducted to confirm its efficacy against sisal zebra disease, providing strong evidence for its application in the management of Phytophthora-related diseases. These results suggest that strain ASG80 represents a valuable microbial resource for the future control of Phytophthora plant diseases.
2. Materials and Methods
2.1. Isolation of Streptomyces
Soil samples were collected from the root zone of sisal in Haikou, Hainan Province, China. The samples were stored in sealed bags at 4°C until further processing. A 10 g portion of the root sample was ground using a sterile mortar and pestle, transferred to an Erlenmeyer flask containing 90 mL of sterile water, and agitated for 20 minutes. A 1 mL aliquot of the suspension was serially diluted up to 10⁴-fold, and the 10⁻², 10⁻³, and 10⁻⁴ dilutions were spread onto Gauze’s No. 1 solid medium plates (soluble starch, 20.0 g; KNO₃, 1.0 g; K₂HPO₄, 0.5 g; MgSO₄·7H₂O, 0.5 g; NaCl, 0.5 g; FeSO₄·7H₂O, 0.01 g; agar, 20.0 g; pH 7.4–7.6), containing 1 µg/mL nystatin, nalidixic acid, and potassium dichromate as selective agents. Plates were incubated at 28°C, with each dilution plated in triplicate. Colonies were subsequently selected from the plates, and strain was isolated and purified on fresh ISP2 solid medium (yeast extract, 4.0 g; malt extract, 10.0 g; glucose, 4.0 g; trace salt solution, 1.0 mL; agar, 20.0 g; distilled water, 1000 mL; pH 7.2). The trace salt solution consisted of FeSO₄·7H₂O, 0.1 g; MnCl₂·4H₂O, 0.1 g; ZnSO₄·7H₂O, 0.1 g; distilled water, 1000 mL. Plates were autoclaved at 121°C for 20 minutes. The purified strain was preserved in 20% glycerol at -80°C for long-term storage.
2.2. Screening of Streptomyces Strains with Anti-Phytophthora Activity and Antifungal Spectrum Assays
The antagonistic activity of Streptomyces strains against Phytophthora nicotianae was assessed using the dual-culture plate assay. A mycelial plug of P. nicotianae was placed at the center of PDA plate, and each Streptomyces strain was inoculated at four equidistant points approximately 2 cm from the center. Plates inoculated with P. nicotianae alone served as the control [33]. The plates were incubated at 28°C until the fungal growth in the control group completely covered the plate. Each treatment was conducted in triplicate. The fungal colony diameter was measured using the cross-measurement method, and the inhibition rate of each Streptomyces strain was calculated. The inhibition rate (%) was determined using the formula: Inhibition rate (%) = [(colony diameter of control - colony diameter of treatment) / (colony diameter of control - diameter of fungal plug)] × 100%.
The strain exhibiting the highest antimicrobial activity was selected for further antifungal spectrum assays. The evaluated pathogens included plant pathogenic oomycetes: Phytophthora nicotianae, P. cinnamomi, P. palmivora, P. capsici, P. vignae, P. melonis, P. colocasiae, P. sojae, and P. melonis. Additionally, several plant pathogenic fungi were tested, including Colletotrichum gloeosporioides, Fusarium graminearum, Magnaporthe grisea, Fusarium oxysporum, and Pestalotiopsis palmarum. For each pathogen, a 5 mm mycelial plug was placed at the center of a PDA plate (90 mm diameter), and strain ASG80 was inoculated at two positions, 2 cm above and below the plug. Plates inoculated only with the pathogen served as controls. The plates were incubated at 28°C until fungal growth in the control group fully covered the plate. Each treatment was conducted in triplicate. The colony diameter was measured using the cross-measurement method, and the inhibition rate of strain ASG80 against each pathogen was calculated as follows: Inhibition rate (%) = [(control colony diameter - treatment colony diameter ) / (control colony diameter - pathogen plug diameter)]× 100%.
2.3. Genome Assembly and Annotation
Sample preparation and genome sequencing were performed as previously described by Wang. The ASG80 sample was sequenced at Biomarker Technologies (Beijing, China) using single-molecule real-time sequencing on the PacBio platform. Genome assembly and functional annotation followed the methods outlined by Wang [34]. The genome of Streptomyces luteireticuli ASG80 was annotated using Prodigal (v2.6.3) [35]. rRNA and tRNA genes were identified with Infernal (v1.1.3) [36]and tRNAscan-SE (v2.0) [37], respectively. Functional annotation was conducted using BLASTx against multiple databases, including the NCBI non-redundant protein database (Nr), gene ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG), Clusters of Orthologous Groups (COG), and Swiss-Prot. Gene clusters involved in secondary metabolite synthesis were identified using antiSMASH 7.1.0 [38].
The 16S rRNA gene sequence of strain ASG80 was extracted using Barrnap (version 0.9). This sequence, along with 16S rRNA sequences from closely related Streptomyces strains downloaded from the EZBioCloud database, was used to construct a preliminary 16S rRNA-based phylogenetic tree to clarify the taxonomic position of strain ASG80 and its relationships with related strains. A phylogenetic tree was generated using Molecular Evolutionary Genetic Analysis software (version 7.0, MEGA, Auckland, New Zealand). Based on the results of the 16S rRNA phylogenetic tree, genomes of closely related species were retrieved from the GenBank database. A whole-genome phylogenetic tree was then constructed online using the Type Strain Genome Server (https://tygs.dsmz.de/), and digital DNA-DNA hybridization (dDDH) values were calculated [38]. Additionally, whole-genome comparisons between strain ASG80 and its closely related species were conducted using JSpeciesWS (http://jspecies.ribohost.com/jspeciesws/#analyse) to obtain average nucleotide identity (ANI) values. The taxonomic classification of the isolated Streptomyces strain was confirmed based on the dDDH and ANI values.
2.4. Comparative Genomic Analysis
To analyze the phylogenetic relationships and genomic similarities, three strains most closely related to strain ASG80 were identified based on phylogenetic analysis, ANI values, and dDDH values comparisons. The selected strains were Streptomyces luteireticuli JCM 4788 (GenBank Accession No.: GCA_039521205.1), S. thioluteus JCM 4087 (GCA_039535865.1), and S. caatingaensis CMAA 1322 (GCA_001187435.1). A comparative genomic analysis was performed using OrthoMCL to classify protein families from strain ASG80 and the reference genomes [39]. A Venn or petal diagram was constructed to illustrate the gene family statistics. A BLAST analysis was executed to compare the protein sequences of strain ASG80 with those of the reference genomes, allowing identification of homologous genes. Syntenic relationships at the nucleotide level were established based on their respective genomic loci. Pairwise synteny plots between the ASG80 genome and each reference genome were generated using MCScanX software [40].
2.5. Preparation of Ethyl Acetate Extract
Strain ASG80 was first activated on ISP2 solid medium plates and subsequently inoculated into ISP2 liquid medium. The culture was incubated at 28°C with shaking at 180 rpm for 3 days. A 1% (v/v) inoculum was then transferred to a soybean meal infusion medium (containing sucrose, 10.0 g; peptone, 2.0 g; soluble starch, 5.0 g; yeast extract, 2.0 g; NaCl, 2.0 g; K₂HPO₄, 0.5 g; MgSO₄·7H₂O, 0.5 g; CaCO₃, 1.0 g; soybean meal, 20.0 g; distilled water, 1000 mL; pH 7.2). The medium was sterilized by autoclaving at 121°C for 20 minutes. The culture was then incubated at 28°C and 180 rpm for 8 days to obtain the fermentation broth. The fermentation broth was extracted three times with an equal volume of ethyl acetate. The combined ethyl acetate extracts were concentrated to dryness under reduced pressure at 45°C to yield the crude extract, representing the fermentation product of strain ASG80.
2.6. Inhibition Rate of Mycelial Growth by ASG80 Extract
The inhibitory activity of the ASG80 extract against mycelial growth of plant pathogens was evaluated using the growth rate method. The extract was dissolved in DMSO at a concentration of 10.0 mg/mL and added to PDA or V8 medium at 50-60°C. A series of PDA or V8 plates with varying concentrations of the active compound were prepared using two-fold serial dilutions, resulting in final concentrations of 0.125 μg/mL, 0.25 μg/mL, 0.5 μg/mL, 1 μg/mL, 2 μg/mL, 4 μg/mL, and 8 μg/mL. Plates with an equal volume of DMSO served as controls. Each plant pathogen was inoculated at the center of the plates, which were then incubated at 28°C until the mycelia in the control group reached the plate edge. The average vertical diameter of the colonies was measured. Each treatment was performed in triplicate. The median effective concentration (EC₅₀) values for mycelial growth inhibition were calculated to determine the extract's efficacy against plant pathogens [41].
2.7. Pot Experiment for Disease Control with ASG80 Extract
The inoculum of Phytophthora nicotianae was prepared following established methods. Millet grains were boiled until approximately two-thirds of the husks cracked, then filtered through gauze and air-dried to approximately 40% moisture content. The grains were placed in Erlenmeyer flasks and sterilized at 121°C for 20 minutes. P. nicotianae was cultured on V8 medium for 4 days, after which five 5-mm mycelial plugs were inoculated onto the sterilized millet medium. The cultures were then incubated at 28°C for 14 days [42].
The test soil, obtained from Huizhou Bida Landscape Materials Co., Ltd., was mixed with P. nicotianae-infested millet to create infected soil. Each pot was filled with 1 kg of soil, containing 4 g of inoculated millet. Uniform 0.5-leaf stage sisal seedlings were transplanted into the treated soil and immediately received the designated treatments. Five treatment groups were established: CK (no P. nicotianae, sterile water), Pn (P. nicotianae, sterile water), T-1 (P. nicotianae, ASG80 extract at a 1:1000 dilution), T-2 (P. nicotianae, ASG80 extract at a 1:2000 dilution), and Me-2 (P. nicotianae, metalaxyl at a 1:2000 dilution). For each treatment, 20 mL of solution was applied to each pot for root drenching. Disease incidence and severity were assessed on individual plants 30 days after transplanting, with disease index and control efficacy subsequently calculated.
3. Results
3.1. Screening of Streptomyces
A total of 28 strains of Streptomyces with antagonistic activity against Phytophthora nicotianae were isolated from the roots of sisal. Notably, strain ASG80 exhibited the strongest antibacterial activity, achieving a growth inhibition rate of 80.51 ± 0.16% (Figure 1A). On ISP2 solid medium, strain ASG80 formed light yellow substrate mycelium, while the aerial mycelium appeared white (Figure 1B). Scanning electron microscopy (SEM) revealed that the aerial mycelium exhibited a filamentous structure (Figure 1C).
3.2. Antifungal Spectrum of Strain ASG80
The antifungal spectrum of strain ASG80 was evaluated using a standard spot inoculation method. Strain ASG80 exhibited broad-spectrum antifungal activity, with varying degrees of inhibition observed against all 12 tested pathogens. Notably, it showed strong inhibitory effects against oomycetes such as Phytophthora cinnamomi, P. palmivora, P. capsici, P. vignae, P. melonis, P. colocasiae, P. sojae, and P. melonis, achieving a maximum inhibition rate of 88.43 ± 0.31. The inhibition rate against Magnaporthe oryzae was 85.87 ± 0. In contrast, the inhibitory effects on Colletotrichum gloeosporioides, Fusarium graminearum, M. oryzae, F. oxysporum, and Pestalotiopsis palmarum were relatively lower (Table 1; Supplementary Figure S1).
3.3. Complete Genome Sequence and Biosynthesis-Related Gene Clusters of Strain ASG80
The complete genome of strain ASG80 was sequenced using single-molecule real-time (SMRT) sequencing on the PacBio platform and assembled de novo into four contigs, consisting of one chromosomal DNA (accession number: CP167927 ) and three plasmid DNAs (accession numbers: CP167926-CP167930). The total genome length of ASG80 is 8,724,805 bp, with a GC content of 71.77%, and it encodes 7,867 genes(Figure 2). Coding sequences constitute 87.01% of the genome, with an average gene length of 964 bp. Structural analysis revealed that the ASG80 genome contains 21 rRNAs, 80 tRNAs, four CRISPR arrays, four genomic islands, three prophages, and five operons. Functional annotation of the 7,867 genes was performed using eggNOG, GO, KEGG, Nr, Swiss-Prot, TrEMBL, and Pfam databases, resulting in the annotation of 7,549 genes (97.50%) across all databases (Table 2).
The antiSMASH v7.1.0 analysis predicted 40 biosynthetic gene clusters for secondary metabolites within the ASG80 genome (Table 3). Most bioactive compounds produced by Streptomyces, such as insecticides, antibiotics, and anticancer agents, are synthesized through polyketide synthase (PKS) and non-ribosomal peptide synthase (NRPS) pathways. The strain ASG80 genome contains 25 gene clusters related to PKS and NRPS pathways, accounting for 62.5% of all gene clusters. These include 13 T1PKS, one T2PKS, 14 NRPS, and six NRPS-like clusters. Additional gene clusters identified include nine Terpene, two Lassopeptide, one NAPAA, three Phenazine, one Thioamide-NRP, one Butyrolactone, one CDPS, one Furan, three RiPP-like, one Thiopeptide, two NI-siderophore, and one Indole cluster.
Among the 40 gene clusters encoded in strain ASG80 genome, four clusters (Clusters 1, 8, 11, and 12) showed 100% similarity to known gene clusters. An additional three clusters (Clusters 16, 38, and 39) exhibited over 80% similarity, indicating that strain ASG80 likely has the potential to produce these secondary metabolites. Meanwhile, 27 clusters displayed similarities ranging from 4% to 76%, suggesting the potential for strain ASG80 to produce known metabolites or their structural analogs. The remaining six clusters showed no significant matches, suggesting they may represent novel gene clusters.
Phylogenetic and homology analysis using the 16S rDNA sequence revealed that strain ASG80 shares the highest similarity (99.86%) with Streptomyces luteireticuli NBRC 13422T, clustering closely with this species (Figure 3). A phylogenetic tree was constructed based on the whole-genome sequences of ASG80, S. luteireticuli JCM 4788T, S. thioluteus JCM 4087T, S. caatingaensis CMAA 1322T, S. huiliensis SCA2-4T, S. mobaraensis DSM 40847T, S. cinnamoneus JCM 4633T, S. sichuanensis SCA3-4T, S. griseocarneus JCM 4580T, S. olivoverticillatus CECT 3266T, S. hiroshimensis JCM 4586T, S. telluris AA8T, S. abikoensis JCM 4002T, and S. luteosporeus JCM 4542T (Figure 4A). The results indicate that strain ASG80 is most closely related to S. luteireticuli JCM 4788, while it is more distantly related to S. telluris AA8 and S. hiroshimensis JCM 4586.
Species delineation among Streptomyces spp. was determined using digital DNA-DNA hybridization (dDDH) and average nucleotide identity (ANI) values, with thresholds set at ≥70% for dDDH and ≥95% for ANI. Comparative analysis of dDDH and ANI values for 13 Streptomyces strains based on whole-genome alignments revealed that strain ASG80 shares the highest dDDH (86.10%) and ANI (98.22%) values with S. luteireticuli JCM 4788, clustering together in the phylogenetic tree(Figure 4B). These findings confirm that strain ASG80 belongs to the same species as S. luteireticuli JCM 4788, and we propose naming this strain Streptomyces luteireticuli ASG80.
3.4. Comparative Genomic Analysis
Based on the phylogenetic tree and supported by dDDH and ANI values, the genomes of the most closely related strains, Streptomyces luteireticuli JCM 4788, S. thioluteus JCM 4087, and S. caatingaensis CMAA 1322, were selected for comparative genomic analysis with strain ASG80. This analysis was conducted to examine the differences between strain ASG80 and related strains at both intra-species and inter-species levels. Phylogenetic analysis using PhyML software generated an evolutionary tree illustrating the relationships among these species (Figure 5A). The results positioned strain ASG80 and JCM 4788 on the same branch, indicating a close evolutionary relationship. In terms of gene family classification, ASG80 contains 6,340 gene families, while strain JCM 4788, JCM 4087, and CMAA 1322 contain 6,219, 2,100, and 4,967 gene families, respectively. Strain ASG80 shares 6,093, 2,012, and 4,752 genes with strain JCM 4788, JCM 4087, and CMAA 1322, respectively. A total of 1,702 gene families are conserved across all four strains, with strain ASG80, JCM 4788, JCM 4087, and CMAA 1322 containing 121, 14, 5, and 58 unique genes, respectively (Figure 5B). Further genomic comparisons using Mauve were conducted to align the genomes of strain ASG80, JCM 4788, JCM 4087, and CMAA 1322, revealing multiple rearrangements in syntenic regions among these genomes (Figure 5C).
3.5. Broad-Spectrum Antifungal Activity Assay
Increasing evidence indicates that secondary metabolites from Streptomyces species can effectively inhibit the growth of Phytophthora and other fungal pathogens. To assess the antagonistic activity of strain ASG80 against Phytophthora and fungal pathogens, ethyl acetate extracts of strain ASG80 were tested for antifungal activity against 13 different Phytophthora and fungal pathogens in vitro(Table 4). Strain ASG80 extract exhibited strong inhibitory effects on the growth of all tested fungal pathogens, with EC₅₀ values ranging from 0.8688 to 51.2052 µg/mL. The lowest EC₅₀ value was observed against P. cinnamomi (0.8688 µg/mL), followed by P. vignae (1.3427 µg/mL), P. nicotianae (1.6120 µg/mL), and P. sojae (1.6806 µg/mL). In contrast, the highest EC₅₀ was observed against Fusarium oxysporum (51.2052 µg/mL).
3.6. Effect of Strain ASG80 Extract on Sisal Zebra Disease under Greenhouse Conditions
To assess the in vivo inhibitory effect of strain ASG80 extract on Phytophthora nicotianae, a pot experiment was conducted under controlled greenhouse conditions. After 30 days of treatment, the disease severity index in the treated groups (ranging from 8.33 to 18.43) was significantly lower than that of the control group (52.45). The disease indices in the T-1 and T-2 treatment groups were reduced by 64.86% and 84.12%, respectively, indicating a positive correlation with the extract concentration(Table 5; Supplementary Figure S2).
4. Discussion
In numerous studies, antagonistic Streptomyces species have been utilized for plant disease control. For instance, S. olivoreticuli ZZ-21 has been effectively utilized against tobacco target spot, caused by Rhizoctonia solani [43], while Streptomyces sp. Y1-14 has proven effective against banana Fusarium wilt [44]. Additionally, Streptomyces species produce diverse secondary metabolites that have been formulated as biopesticides for agricultural applications, including Jinggangmycin [45], Zhongshengmycin [45], and Avermectin [46]. In this study, antagonistic Streptomyces strains were screened, leading to the identification of a highly effective strain ASG80. In vivo and in vitro experiments both confirmed the exceptional biocontrol activity exhibited by strain ASG80. Accordingly, the genome of strain ASG80 was sequenced for detailed analysis.
Phylogenetic analysis, informed by whole-genome sequencing, average nucleotide identity (ANI) of 98.22%, and a digital DNA-DNA hybridization (dDDH) value of 86.10%, identified ASG80 as Streptomyces luteireticuli. Comparative genomic analysis with Streptomyces strains JCM4788, JCM4087, and CMAA1322 revealed that strain ASG80 and JCM4788 belong to the same evolutionary lineage. Gene family clustering indicated the sharing of 1,702 genes, with strain ASG80 containing 121 unique genes. Syntenic analysis revealed poor genomic alignment between strain ASG80 and strains JCM4788, JCM4087, and CMAA1322, reflecting numerous insertions, deletions, inversions, and translocations among genes. These findings imply that strain ASG80 possesses distinctive characteristics, and its biocontrol mechanisms may significantly differ from those observed in other Streptomyces strains.
Streptomyces species produce a diverse range of secondary metabolites, primarily through the biosynthesis of polyketides, nucleosides, peptides, and hydrolytic enzymes. These metabolites are capable of inhibiting or even eliminating pathogens. In addition, Streptomyces generates various bioactive compounds with antimicrobial properties, including enzymes, organic acids, amino acids, immunomodulators, and vitamins [47,48,49]. Whole-genome sequencing enables comprehensive genomic analysis, identification, and classification of strain ASG80, and facilitates the exploration of potential biosynthetic gene clusters (BGCs) responsible for secondary metabolites. A total of 40 secondary metabolite biosynthesis gene clusters were identified in the genome of strain ASG80. Among them, 11 clusters exhibited significant similarity (greater than 50%) to known BGCs, with four clusters—antipain, geosmin, diisonitrile antibiotic SF2768, and aureothin—demonstrating 100% similarity. Antipain is a novel analogue that inhibits pain by suppressing PAR signaling via protease inhibition, subsequently reducing excitatory neuropeptide release [50]. Diisonitrile antibiotic SF2768 acts as a copper carrier by binding specifically to copper [51], while aureothin exhibits a range of biological activities, including antifungal [52,53], antiviral [54], antitumor [53], nematicidal [55], and larvicidal effects [56]. Of note, a significant number of BGCs exhibited minimal similarity, with similarity scores below 50%, and many others under 20%. Six BGCs displayed no detectable similarity to known clusters, suggesting that strain ASG80 harbors numerous genes with potentially novel functions, thereby highlighting its significant research potential.
In recent years, an increasing number of Streptomyces species have been identified to possess anti-oomycete activity, positioning them as potential biocontrol agents against Phytophthora diseases. For example, Streptomyces sp. FXP04 and Streptomyces sp. A2-16 have been demonstrated to control late blight in potatoes [57,58], while S. rochei IT20 and S. vinaceusdrappus SS14 effectively inhibit Phytophthora in chili peppers [59]. However, compared to bacterial and fungal antagonists, relatively few studies report on the use of Streptomyces for Phytophthora biocontrol. In this study, strain ASG80 and its extracts showed potent inhibitory effects against both oomycete and fungal pathogens. The extracts exhibited EC50 values between 0.8 and 3.0 µg/mL against oomycete pathogens, whereas EC50 values exceeded 8.0 µg/mL for fungal pathogens, indicating a stronger inhibitory effect on oomycetes.
Sisal zebra disease, induced by Phytophthora nicotianae, represents a significant threat to sisal cultivation. Chemical fungicides are commonly utilized to manage this pathogen. In an effort to minimize reliance on synthetic fungicides, recent research has increasingly centered on identifying effective biological control agents, such as Bacillus [60], Pseudomonas [61], Trichoderma [62], and yeast strains [63], that demonstrate efficacy in inhibiting P. nicotianae. In this study, strain ASG80 extracts exhibited broad-spectrum in vitro anti-Phytophthora activity and substantially decreased zebra stripe disease incidence in sisal in vivo, demonstrating a control effect comparable to that of metalaxyl. The observed reduction in disease incidence was concentration-dependent, suggesting that the extracts function as the principal inhibitory component. Additionally, strain ASG80 displayed broad-spectrum activity against Phytophthora and fungal pathogens, exhibiting significant inhibitory effects on 13 different pathogens. This finding aligns with previous reports indicating that promising biocontrol strains frequently exhibit a broad antimicrobial spectrum [64]. To the best of our knowledge, this study represents the first investigation into the biocontrol potential of Streptomyces luteireticuli against Phytophthora species. Strain ASG80 demonstrates significant potential as a biocontrol agent for the management of Phytophthora diseases.
5. Conclusion
In this study, the complete genomic profile of strain ASG80 was comprehensively analyzed, and the functional roles of its associated genes were investigated. The findings provide insights that could support further research into the regulatory mechanisms of active secondary metabolite biosynthesis in strain ASG80. Furthermore, strain ASG80 extracts exhibited broad-spectrum in vitro anti-Phytophthora activity and significantly reduced the incidence of sisal zebra disease in vivo. These results indicate that strain ASG80 possesses considerable potential as a biocontrol agent against Phytophthora diseases in plants. The development and application of strain ASG80 could contribute significantly to green control strategies for plant diseases, providing a valuable resource for utilizing antagonistic Streptomyces in biocontrol. Additionally, this strain offers a foundation for the experimental exploration and application of novel antibacterial compounds.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
We express our gratitude to Zhuoyu Sun at the Modern Agricultural Development Service Center of Changjiang Li Autonomous County, for his invaluable assistance in guiding our team through the collection of sisal seedlings and soil samples.
References
- Judelson, H.S.; Blanco, F.A. The spores of Phytophthora: weapons of the plant destroyer. Nature reviews. Microbiology 2005, 3, 47-58, doi:10.1038/nrmicro1064. [CrossRef]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nature Reviews Microbiology 2023, 21, 640-656, doi:10.1038/s41579-023-00900-7. [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. NATURE ECOLOGY & EVOLUTION 2019, 3, 430-+, doi:10.1038/s41559-018-0793-y. [CrossRef]
- Grunwald, N.J.; Goss, E.M.; Press, C.M. Phytophthora ramorum: a pathogen with a remarkably wide host range causing sudden oak death on oaks and ramorum blight on woody ornamentals. Molecular plant pathology 2008, 9, 729-740, doi:10.1111/j.1364-3703.2008.00500.x. [CrossRef]
- Farooq, Q.U.A.; McComb, J.; Hardy, G.E.S.J.; Burgess, T.I. Soil amendments for management of Phytophthora root rot in avocado and their impact on the soil microbiome. JOURNAL OF PLANT PATHOLOGY 2024, 106, 439-455, doi:10.1007/s42161-024-01604-4. [CrossRef]
- Lu, X.H.; Zhu, S.S.; Bi, Y.; Liu, X.L.; Hao, J.J. Baseline Sensitivity and Resistance-Risk Assessment of Phytophthora capsici to Iprovalicarb. PHYTOPATHOLOGY 2010, 100, 1162-1168, doi:10.1094/PHYTO-12-09-0351. [CrossRef]
- Fang, Y.; Wang, Z.; Zhang, S.; Peng, Q.; Liu, X. Characterization and proteome analysis of the extracellular vesicles of Phytophthora capsici. JOURNAL OF PROTEOMICS 2021, 238, doi:10.1016/j.jprot.2021.104137. [CrossRef]
- Cheng, W.; Lin, M.; Qiu, M.; Kong, L.; Xu, Y.; Li, Y.; Wang, Y.; Ye, W.; Dong, S.; He, S.; et al. Chitin synthase is involved in vegetative growth, asexual reproduction and pathogenesis of Phytophthora capsici and Phytophthora sojae. ENVIRONMENTAL MICROBIOLOGY 2019, 21, 4537-4547, doi:10.1111/1462-2920.14744. [CrossRef]
- Gallup, C.A.; McCorkle, K.L.; Ivors, K.L.; Shew, D. Characterization of the Black Shank Pathogen, Phytophthora nicotianae, Across North Carolina Tobacco Production Areas. PLANT DISEASE 2018, 102, 1108-1114, doi:10.1094/PDIS-02-17-0295-RE. [CrossRef]
- Garcia-Gaona, M.; Botero-Rozo, D.; Araque, L.; Romero, H.M. The Dynamic Interaction between Oil Palm and Phytophthora palmivora in Bud Rot Disease: Insights from Transcriptomic Analysis and Network Modelling. JOURNAL OF FUNGI 2024, 10, doi:10.3390/jof10030164. [CrossRef]
- Riseh, R.S.; Hassanisaadi, M.; Vatankhah, M.; Babaki, S.A.; Barka, E.A. Chitosan as a potential natural compound to manage plant diseases. INTERNATIONAL JOURNAL OF BIOLOGICAL MACROMOLECULES 2022, 220, 998-1009, doi:10.1016/j.ijbiomac.2022.08.109. [CrossRef]
- Liu, K.; McInroy, J.A.; Hu, C.-H.; Kloepper, J.W. Mixtures of Plant-Growth-Promoting Rhizobacteria Enhance Biological Control of Multiple Plant Diseases and Plant-Growth Promotion in the Presence of Pathogens. PLANT DISEASE 2018, 102, 67-72, doi:10.1094/PDIS-04-17-0478-RE. [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. MICROORGANISMS 2022, 10, doi:10.3390/microorganisms10030596. [CrossRef]
- Zhou, X.; Wang, J.; Liu, F.; Liang, J.; Zhao, P.; Tsui, C.K.M.; Cai, L. Cross-kingdom synthetic microbiota supports tomato suppression of Fusarium wilt disease. NATURE COMMUNICATIONS 2022, 13, doi:10.1038/s41467-022-35452-6. [CrossRef]
- Salazar, B.; Ortiz, A.; Keswani, C.; Minkina, T.; Mandzhieva, S.; Pratap Singh, S.; Rekadwad, B.; Borriss, R.; Jain, A.; Singh, H.B.; et al. Bacillus spp. as Bio-factories for Antifungal Secondary Metabolites: Innovation Beyond Whole Organism Formulations. MICROBIAL ECOLOGY 2023, 86, 1-24, doi:10.1007/s00248-022-02044-2. [CrossRef]
- Kulkova, I.; Dobrzynski, J.; Kowalczyk, P.; Belzecki, G.; Kramkowski, K. Plant Growth Promotion Using Bacillus cereus. INTERNATIONAL JOURNAL OF MOLECULAR SCIENCES 2023, 24, doi:10.3390/ijms24119759. [CrossRef]
- Hou, Q.; Kolodkin-Gal, I. Harvesting the complex pathways of antibiotic production and resistance of soil bacilli for optimizing plant microbiome. FEMS MICROBIOLOGY ECOLOGY 2020, 96, doi:10.1093/femsec/fiaa142. [CrossRef]
- Liu, L.; Zhao, K.; Cai, L.; Zhang, Y.; Fu, Q.; Huang, S. Combination effects of tebuconazole with Bacillus subtilis to control rice false smut and the related synergistic mechanism. PEST MANAGEMENT SCIENCE 2023, 79, 234-243, doi:10.1002/ps.7193. [CrossRef]
- Xiong, W.; Guo, S.; Jousset, A.; Zhao, Q.; Wu, H.; Li, R.; Kowalchuk, G.A.; Shen, Q. Bio-fertilizer application induces soil suppressiveness against Fusarium wilt disease by reshaping the soil microbiome. SOIL BIOLOGY & BIOCHEMISTRY 2017, 114, 238-247, doi:10.1016/j.soilbio.2017.07.016. [CrossRef]
- Santos, M.; Dianez, F.; Sanchez-Montesinos, B.; Huertas, V.; Moreno-Gavira, A.; Esteban Garcia, B.; Garrido-Cardenas, J.A.; Gea, F.J.J. Biocontrol of Diseases Caused by Phytophthora capsici and P. parasitica in Pepper Plants. JOURNAL OF FUNGI 2023, 9, doi:10.3390/jof9030360. [CrossRef]
- Pandit, M.A.; Kumar, J.; Gulati, S.; Bhandari, N.; Mehta, P.; Katyal, R.; Rawat, C.D.; Mishra, V.; Kaur, J. Major Biological Control Strategies for Plant Pathogens. PATHOGENS 2022, 11, doi:10.3390/pathogens11020273. [CrossRef]
- Khan, R.A.A.; Najeeb, S.; Mao, Z.; Ling, J.; Yang, Y.; Li, Y.; Xie, B. Bioactive Secondary Metabolites from Trichoderma spp. against Phytopathogenic Bacteria and Root-Knot Nematode. MICROORGANISMS 2020, 8, doi:10.3390/microorganisms8030401. [CrossRef]
- Zhan, X.; Khan, R.A.A.; Zhang, J.; Chen, J.; Yin, Y.; Tang, Z.; Wang, R.; Lu, B.; Liu, T. Control of postharvest stem-end rot on mango by antifungal metabolites of Trichoderma pinnatum LS029-3. SCIENTIA HORTICULTURAE 2023, 310, doi:10.1016/j.scienta.2022.111696. [CrossRef]
- Duan, Y.; Pang, Z.; Yin, S.; Xiao, W.; Hu, H.; Xie, J.; Moussa, E.J. Screening and Analysis of Antifungal Strains Bacillus subtilis JF-4 and B. amylum JF-5 for the Biological Control of Fusarium Wilt of Banana. JOURNAL OF FUNGI 2023, 9, doi:10.3390/jof9090886. [CrossRef]
- Xie, J.; Singh, P.; Qi, Y.; Singh, R.K.; Qin, Q.; Jin, C.; Wang, B.; Fang, W. Pseudomonas aeruginosa Strain 91: A Multifaceted Biocontrol Agent against Banana Fusarium Wilt. JOURNAL OF FUNGI 2023, 9, doi:10.3390/jof9111047. [CrossRef]
- Chaudhary, A.K.; Dhakal, D.; Sohng, J.K. An Insight into the "-Omics" Based Engineering of Streptomycetes for Secondary Metabolite Overproduction. BIOMED RESEARCH INTERNATIONAL 2013, 2013, doi:10.1155/2013/968518. [CrossRef]
- Nguyen, C.T.; Dhakal, D.; Pham, V.T.T.; Nguyen, H.T.; Sohng, J.-K. Recent Advances in Strategies for Activation and Discovery/Characterization of Cryptic Biosynthetic Gene Clusters in Streptomyces. MICROORGANISMS 2020, 8, doi:10.3390/microorganisms8040616. [CrossRef]
- Rey, T.; Dumas, B. Plenty Is No Plague: Streptomyces Symbiosis with Crops. TRENDS IN PLANT SCIENCE 2017, 22, 30-37, doi:10.1016/j.tplants.2016.10.008. [CrossRef]
- Park, H.J.; Lee, J.Y.; Hwang, I.S.; Yun, B.S.; Kim, B.S.; Hwang, B.K. Isolation and Antifungal and Antioomycete Activities of Staurosporine from Streptomyces roseoflavus Strain LS-A24. Journal of Agricultural and Food Chemistry 2006, 54, 3041-3046, doi:10.1021/jf0532617. [CrossRef]
- Sun, Y.; Wu, H.; Xu, S.; Tang, S.; Hao, J.; Liu, X.; Zhang, H.; Han, L. Roles of the EPS66A polysaccharide from Streptomyces sp. in inducing tobacco resistance to tobacco mosaic virus. INTERNATIONAL JOURNAL OF BIOLOGICAL MACROMOLECULES 2022, 209, 885-894, doi:10.1016/j.ijbiomac.2022.04.081. [CrossRef]
- Wang, X.-J.; Wang, M.; Wang, J.-D.; Jiang, L.; Wang, J.-J.; Xiang, W.-S. Isolation and Identification of Novel Macrocyclic Lactones from Streptomyces avermitilis NEAU1069 with Acaricidal and Nematocidal Activity. JOURNAL OF AGRICULTURAL AND FOOD CHEMISTRY 2010, 58, 2710-2714, doi:10.1021/jf902496d. [CrossRef]
- Arasu, M.V.; Duraipandiyan, V.; Ignacimuthu, S. Antibacterial and antifungal activities of polyketide metabolite from marine Streptomyces sp AP-123 and its cytotoxic effect. CHEMOSPHERE 2013, 90, 479-487, doi:10.1016/j.chemosphere.2012.08.006. [CrossRef]
- Li, X.; Jing, T.; Zhou, D.; Zhang, M.; Qi, D.; Zang, X.; Zhao, Y.; Li, K.; Tang, W.; Chen, Y.; et al. Biocontrol efficacy and possible mechanism of Streptomyces sp. H4 against postharvest anthracnose caused by Colletotrichum fragariae on strawberry fruit. POSTHARVEST BIOLOGY AND TECHNOLOGY 2021, 175, doi:10.1016/j.postharvbio.2020.111401. [CrossRef]
- Wang, Y.; Zhao, Q.; Sun, Z.; Li, Y.; He, H.; Zhang, Y.; Yang, X.; Wang, D.; Dong, B.; Zhou, H.; et al. Whole-genome analysis revealed the growth-promoting mechanism of endophytic bacterial strain Q2H1 in potato plants. FRONTIERS IN MICROBIOLOGY 2022, 13, doi:10.3389/fmicb.2022.1035901. [CrossRef]
- Hyatt, D.; Chen, G.-L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC BIOINFORMATICS 2010, 11, doi:10.1186/1471-2105-11-119. [CrossRef]
- Nawrocki, E.P.; Eddy, S.R. Infernal 1.1: 100-fold faster RNA homology searches. BIOINFORMATICS 2013, 29, 2933-2935, doi:10.1093/bioinformatics/btt509. [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. Methods in molecular biology (Clifton, N.J.) 2019, 1962, 1-14, doi:10.1007/978-1-4939-9173-0_1. [CrossRef]
- Naughton, L.M.; Romano, S.; O'Gara, F.; Dobson, A.D.W. Identification of Secondary Metabolite Gene Clusters in the Pseudovibrio Genus Reveals Encouraging Biosynthetic Potential toward the Production of Novel Bioactive Compounds. FRONTIERS IN MICROBIOLOGY 2017, 8, doi:10.3389/fmicb.2017.01494. [CrossRef]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: identification of ortholog groups for eukaryotic genomes. Genome research 2003, 13, 2178-2189, doi:10.1101/gr.1224503. [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. NUCLEIC ACIDS RESEARCH 2012, 40, doi:10.1093/nar/gkr1293. [CrossRef]
- Van Ewijk, P.H.; Hoekstra, J.A. Calculation of the EC50 and its confidence interval when subtoxic stimulus is present. Ecotoxicology and environmental safety 1993, 25, 25-32, doi:10.1006/eesa.1993.1003. [CrossRef]
- Zhang, X.; Wang, Y.; Han, X.; Gou, J.; Li, W.; Zhang, C. A Novel Bio-Fertilizer Produced by Prickly Ash Seeds with Biochar Addition Induces Soil Suppressiveness against Black Shank Disease on Tobacco. APPLIED SCIENCES-BASEL 2021, 11, doi:10.3390/app11167261. [CrossRef]
- Zhong, J.; Sui, W.W.; Bai, X.Y.; Qiu, Z.L.; Li, X.G.; Zhu, J.Z. Characterization and biocontrol mechanism of Streptomyces olivoreticuli as a potential biocontrol agent against Rhizoctonia solani. Pesticide Biochemistry and Physiology 2023, 197, 105681, doi:doi.org/10.1016/j.pestbp.2023.105681. [CrossRef]
- Cao, M.; Cheng, Q.; Cai, B.; Chen, Y.; Wei, Y.; Qi, D.; Li, Y.; Yan, L.; Li, X.; Long, W.; et al. Antifungal Mechanism of Metabolites from Newly Isolated Streptomyces sp. Y1-14 against Banana Fusarium Wilt Disease Using Metabolomics. JOURNAL OF FUNGI 2022, 8, doi:10.3390/jof8121291. [CrossRef]
- Dong, W.; Wu, W.-J.; Song, C.-Y.; Li, T.; Zhang, J.-Z. Jinggangmycin stimulates reproduction and increases CHCs-dependent desiccation tolerance in Drosophila melanogaster. PESTICIDE BIOCHEMISTRY AND PHYSIOLOGY 2023, 194, doi:10.1016/j.pestbp.2023.105484. [CrossRef]
- Li, M.; Chen, Z.; Zhang, X.; Song, Y.; Wen, Y.; Li, J. Enhancement of avermectin and ivermectin production by overexpression of the maltose ATP-binding cassette transporter in Streptomyces avermitilis. BIORESOURCE TECHNOLOGY 2010, 101, 9228-9235, doi:10.1016/j.biortech.2010.06.132. [CrossRef]
- Sharma, V.; Kaur, R.; Salwan, R. Streptomyces: host for refactoring of diverse bioactive secondary metabolites. 3 BIOTECH 2021, 11, doi:10.1007/s13205-021-02872-y. [CrossRef]
- Ferraiuolo, S.B.; Cammarota, M.; Schiraldi, C.; Restaino, O.F. Streptomycetes as platform for biotechnological production processes of drugs. APPLIED MICROBIOLOGY AND BIOTECHNOLOGY 2021, 105, 551-568, doi:10.1007/s00253-020-11064-2. [CrossRef]
- Lacey, H.J.; Rutledge, P.J. Recently Discovered Secondary Metabolites from Streptomyces Species. MOLECULES 2022, 27, doi:10.3390/molecules27030887. [CrossRef]
- Nakae, K.; Kojima, F.; Sawa, R.; Kubota, Y.; Igarashi, M.; Kinoshita, N.; Adachi, H.; Nishimura, Y.; Akamatsu, Y. Antipain Y, a new antipain analog that inhibits neurotransmitter release from rat dorsal root ganglion neurons. JOURNAL OF ANTIBIOTICS 2010, 63, 41-44, doi:10.1038/ja.2009.109. [CrossRef]
- Wang, L.; Zhu, M.; Zhang, Q.; Zhang, X.; Yang, P.; Liu, Z.; Deng, Y.; Zhu, Y.; Huang, X.; Han, L.; et al. Diisonitrile Natural Product SF2768 Functions As a Chalkophore That Mediates Copper Acquisition in Streptomyces thioluteus. ACS CHEMICAL BIOLOGY 2017, 12, 3067-3075, doi:10.1021/acschembio.7b00897. [CrossRef]
- Werneburg, M.; Busch, B.; He, J.; Richter, M.E.A.; Xiang, L.; Moore, B.S.; Roth, M.; Dahse, H.-M.; Hertweck, C. Exploiting Enzymatic Promiscuity to Engineer a Focused Library of Highly Selective Antifungal and Antiproliferative Aureothin Analogues. JOURNAL OF THE AMERICAN CHEMICAL SOCIETY 2010, 132, 10407-10413, doi:10.1021/ja102751h. [CrossRef]
- Busch, B.; Hertweck, C. Evolution of metabolic diversity in polyketide-derived pyrones: Using the non-colinear aureothin assembly line as a model system. PHYTOCHEMISTRY 2009, 70, 1833-1840, doi:10.1016/j.phytochem.2009.05.022. [CrossRef]
- Herrmann, A.; Roesner, M.; Werner, T.; Hauck, S.M.; Koch, A.; Bauer, A.; Schneider, M.; Brack-Werner, R. Potent inhibition of HIV replication in primary human cells by novel synthetic polyketides inspired by Aureothin. SCIENTIFIC REPORTS 2020, 10, doi:10.1038/s41598-020-57843-9. [CrossRef]
- Kang, M.-K.; Kim, J.-H.; Liu, M.-J.; Jin, C.-Z.; Park, D.-J.; Kim, J.; Sung, B.-H.; Kim, C.-J.; Son, K.-H. New discovery on the nematode activity of aureothin and alloaureothin isolated from endophytic bacteria Streptomyces sp. AE170020. SCIENTIFIC REPORTS 2022, 12, doi:10.1038/s41598-022-07879-w. [CrossRef]
- Kim, S.J.; Cantrell, C.L.; Avula, B.; Chen, J.; Schrader, K.K.; Santo, S.N.; Ali, A.; Khan, I.A. Streptomyces distallicus, a Potential Microbial Biolarvicide. JOURNAL OF AGRICULTURAL AND FOOD CHEMISTRY 2022, doi:10.1021/acs.jafc.2c03537. [CrossRef]
- Fu, X.; Liu, S.; Ru, J.; Tang, B.; Zhai, Y.; Wang, Z.; Wang, L. Biological control of potato late blight by Streptomyces sp. FXP04 and potential role of secondary metabolites. BIOLOGICAL CONTROL 2022, 169, doi:10.1016/j.biocontrol.2022.104891. [CrossRef]
- Feng, S.; Tang, S.; Jian, Y.; Huang, X.; Jin, L.; Zhu, Z.; Dong, P.; Li, Z. Complete Genome Sequence Data of a Novel Streptomyces sp. Strain A2-16, a Potential Biological Control Agent for Potato Late Blight. PLANT DISEASE 2022, 106, 723-726, doi:10.1094/PDIS-04-21-0858-A. [CrossRef]
- Abbasi, S.; Safaie, N.; Sadeghi, A.; Shamsbakhsh, M. Tissue-specific synergistic bio-priming of pepper by two Streptomyces species against Phytophthora capsici</i>. PLOS ONE 2020, 15, doi:10.1371/journal.pone.0230531. [CrossRef]
- Ji, S.; Tian, Y.; Li, J.; Xu, G.; Zhang, Y.; Chen, S.; Chen, Y.; Tang, X. Complete genome sequence of Bacillus cereus Z4, a biocontrol agent against tobacco black shank, isolated from the Western Pacific Ocean. MARINE GENOMICS 2023, 72, doi:10.1016/j.margen.2023.101071. [CrossRef]
- Rocio Suarez-Moreno, Z.; Marcela Vinchira-Villarraga, D.; Isabel Vergara-Morales, D.; Castellanos, L.; Ramos, F.A.; Guarnaccia, C.; Degrassi, G.; Venturi, V.; Moreno-Sarmiento, N. Plant-Growth Promotion and Biocontrol Properties of Three Streptomyces spp. Isolates to Control Bacterial Rice Pathogens. FRONTIERS IN MICROBIOLOGY 2019, 10, doi:10.3389/fmicb.2019.00290. [CrossRef]
- Bae, S.-J.; Mohanta, T.K.; Chung, J.Y.; Ryu, M.; Park, G.; Shim, S.; Hong, S.-B.; Seo, H.; Bae, D.-W.; Bae, I.; et al. Trichoderma metabolites as biological control agents against Phytophthora pathogens. BIOLOGICAL CONTROL 2016, 92, 128-138, doi:10.1016/j.biocontrol.2015.10.005. [CrossRef]
- Liu, Z.; Tian, J.; Yan, H.; Li, D.; Wang, X.; Liang, W.; Wang, G. Ethyl acetate produced by Hanseniaspora uvarum is a potential biocontrol agent against tomato fruit rot caused by Phytophthora nicotianae. FRONTIERS IN MICROBIOLOGY 2022, 13, doi:10.3389/fmicb.2022.978920. [CrossRef]
- Zheng, T.-w.; Liu, L.; Nie, Q.-w.; Hsiang, T.; Sun, Z.-x.; Zhou, Y. Isolation, identification and biocontrol mechanisms of endophytic bacterium D61-A from Fraxinus hupehensis against Rhizoctonia solani. BIOLOGICAL CONTROL 2021, 158, doi:10.1016/j.biocontrol.2021.104621. [CrossRef]
Figure 1.
Isolation of strain ASG80 with strong antifungal activity against Phytophthora nicotiana. (A) The colony morphology of strain ASG80. (B) Morphological characteristics of aerial mycelia of strain ASG80 using SEM . (C) Strain Y1-14 extracts inhibiting mycelial growth of Phytophthora nicotiana.
Figure 1.
Isolation of strain ASG80 with strong antifungal activity against Phytophthora nicotiana. (A) The colony morphology of strain ASG80. (B) Morphological characteristics of aerial mycelia of strain ASG80 using SEM . (C) Strain Y1-14 extracts inhibiting mycelial growth of Phytophthora nicotiana.
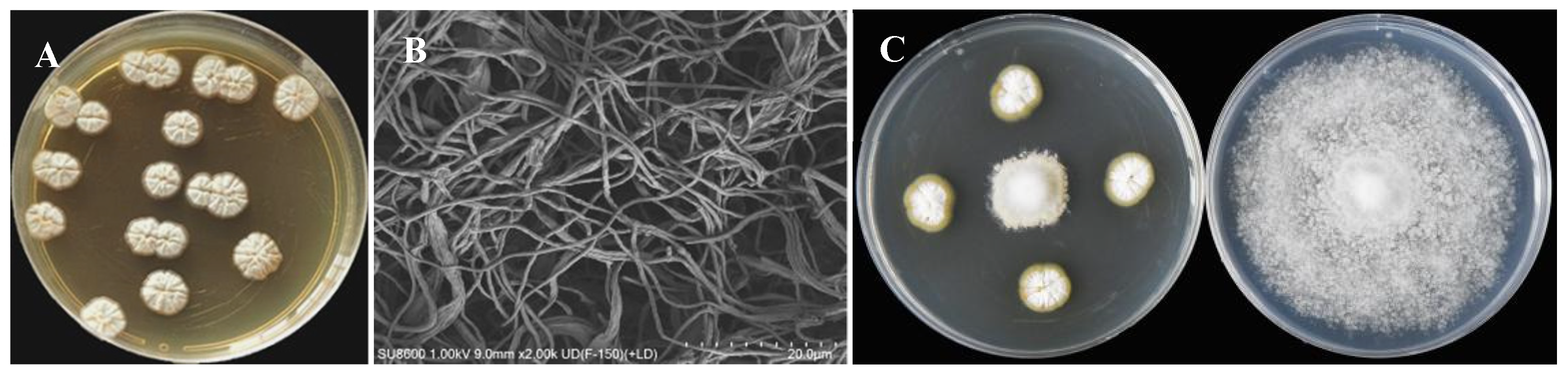
Figure 2.
Genome map of strain ASG80: The outermost circle represents the genome size. The second and third circles depict genes on the positive and negative strands of the genome, with different colors indicating various COG functional classifications. The fourth circle displays repeat sequences, while the fifth indicates tRNA (blue) and rRNA (purple). The sixth circle represents GC content, and the innermost circle shows the GC skew. The letters A–Z correspond to the functional classification of CDS genes in the chromosome.
Figure 2.
Genome map of strain ASG80: The outermost circle represents the genome size. The second and third circles depict genes on the positive and negative strands of the genome, with different colors indicating various COG functional classifications. The fourth circle displays repeat sequences, while the fifth indicates tRNA (blue) and rRNA (purple). The sixth circle represents GC content, and the innermost circle shows the GC skew. The letters A–Z correspond to the functional classification of CDS genes in the chromosome.
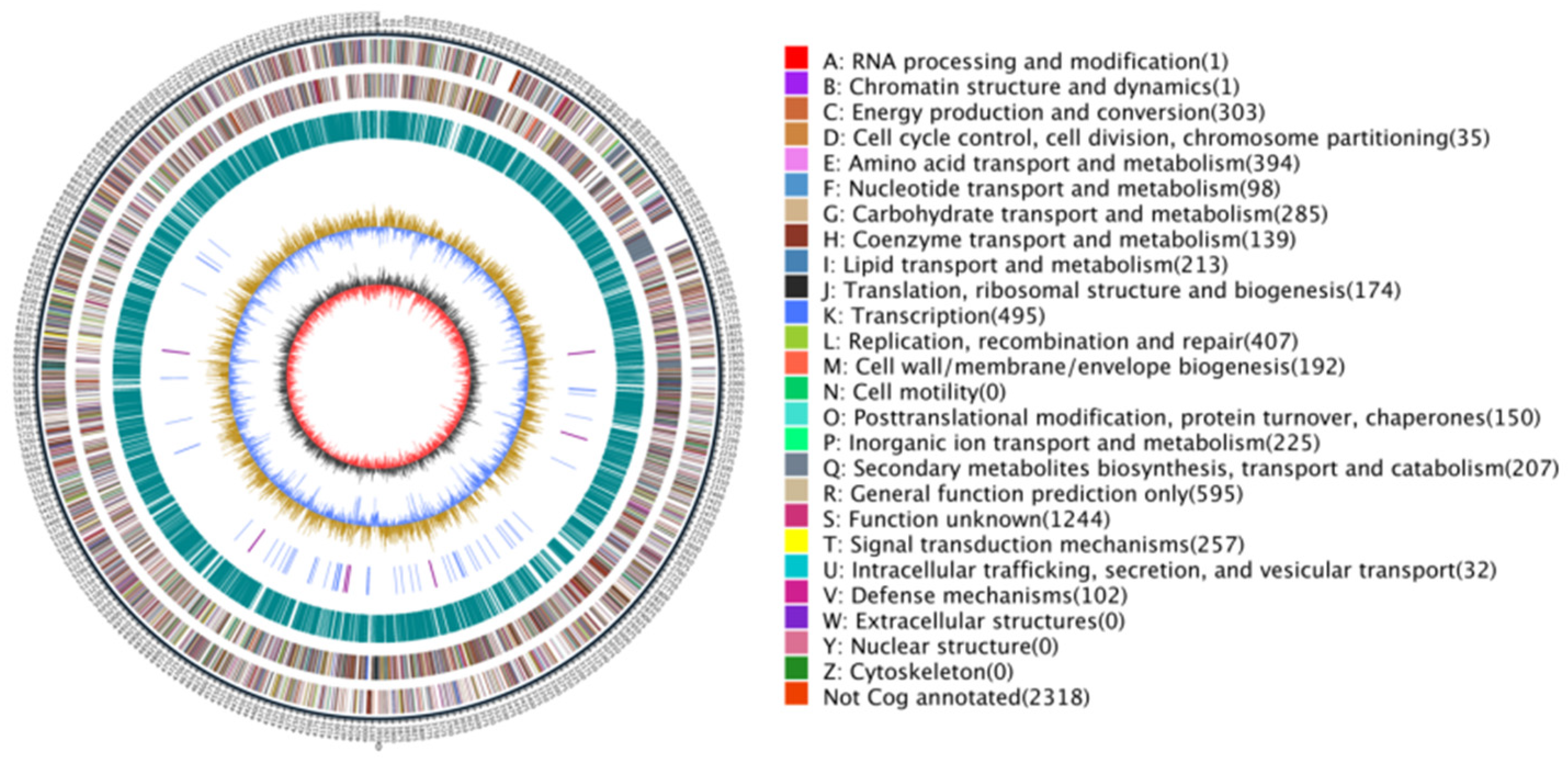
Figure 3.
Phylogenetic tree of strain ASG80 based on 16S rRNA gene sequence analysis. The bootstrap values (%) at the branches were calculated from 1000 replications.
Figure 3.
Phylogenetic tree of strain ASG80 based on 16S rRNA gene sequence analysis. The bootstrap values (%) at the branches were calculated from 1000 replications.
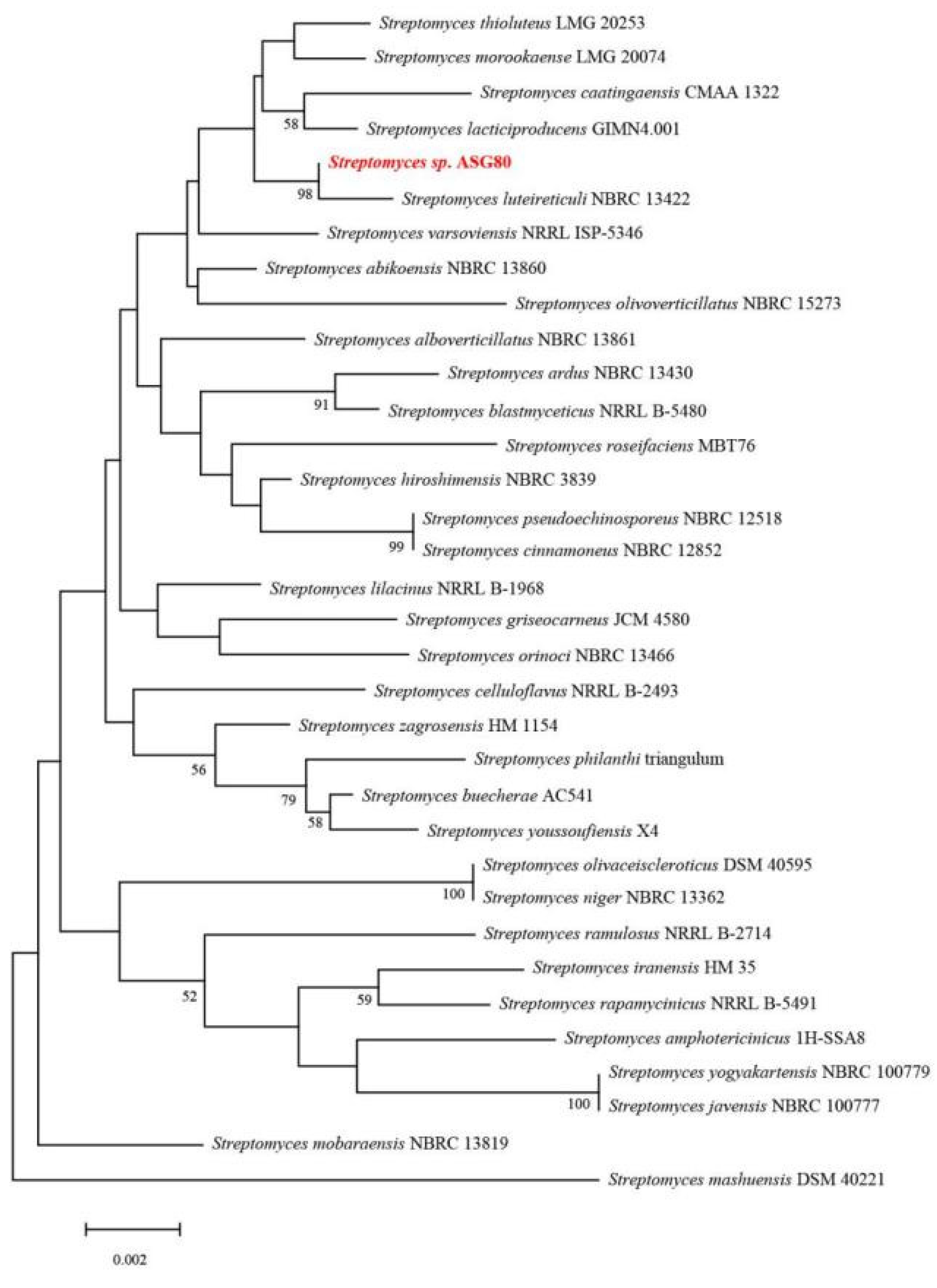
Figure 4.
Analysis of strain ASG80 and related Streptomyces species. (A) A phylogenetic tree was constructed using the Type Strain Genome Server to analyze the whole genome sequences. (B) Heatmaps were generated to visualize the calculated average nucleotide identity (ANI) values and digital DNA-DNA Hybridization (dDDH) values.
Figure 4.
Analysis of strain ASG80 and related Streptomyces species. (A) A phylogenetic tree was constructed using the Type Strain Genome Server to analyze the whole genome sequences. (B) Heatmaps were generated to visualize the calculated average nucleotide identity (ANI) values and digital DNA-DNA Hybridization (dDDH) values.

Figure 5.
Comparative genomics analysis of strain ASG80. (A)Evolutionary relationship between ASG80 and three species. (B) Venn diagram of gene family statistics. (C) Genome collinearity of ASG80/JCM4788、ASG80/CMAA1322 and ASG80/JCM4087.
Figure 5.
Comparative genomics analysis of strain ASG80. (A)Evolutionary relationship between ASG80 and three species. (B) Venn diagram of gene family statistics. (C) Genome collinearity of ASG80/JCM4788、ASG80/CMAA1322 and ASG80/JCM4087.
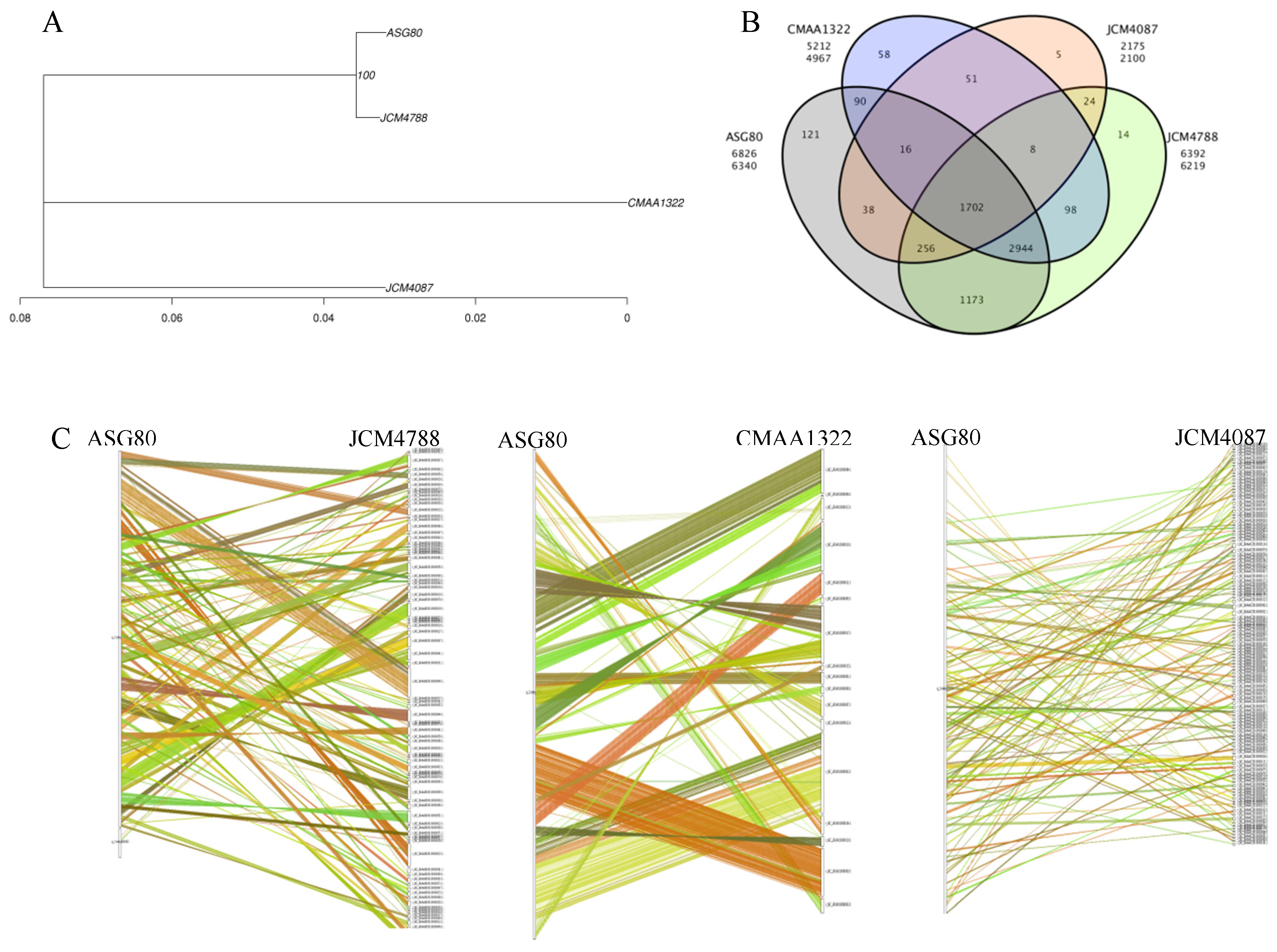

Table 1.
The inhibition rate (%) of the pathogen by strain ASG80.
| Pathogens | Mycelial inhibition(%) |
|---|---|
| Phytophthora colocasiae | 83.74±0 |
| Phytophthora cinnamomi | 72.87±0.46 |
| Phytophthora palmivora | 75.63±0.27 |
| Phytophthora capsici | 61.27±0.18 |
| Phytophthora vignae | 88.43±0.31 |
| Phytophthora sojae | 56.70±0.26 |
| Phytophthora melonis | 70.53±0.15 |
| Colletotrichum gloeosporioides | 62.78±0.60 |
| Magnaporthe grisea | 85.87±0 |
| Fusarium oxysporum | 43.21±0.16 |
| Fusarium graminearum | 44.96±0.31 |
| Pestalotiopsis palmarum | 58.95±0.16 |
Table 2.
Genome characteristics of strain ASG80.
| Database | Number of annotated functional proteins | Proportion/% |
|---|---|---|
| eggNOG | 5,549 | 70.54 |
| GO | 4,598 | 58.45 |
| KEGG | 2,385 | 30.32 |
| Nr | 7,531 | 95.73 |
| Pfam | 5,793 | 73.64 |
| Swiss-prot | 3,440 | 43.73 |
| TrEMB | 7,510 | 95.46 |
| All database | 7,549 | 95.96 |
Table 3.
Secondary metabolite clusters in strain ASG80 predicted by antiSMASH.
| Region | Type | From | To | Most similar known cluster |
Similarity |
|---|---|---|---|---|---|
| Clusters 1 | Terpene,NRPS,NRPS-like | 60,185 | 117,028 | antipain | 100% |
| Clusters 2 | T1PKS | 169,015 | 216,487 | himastatin | 8% |
| Clusters 3 | T1PKS | 242,821 | 288,100 | petrichorin | 5% |
| Clusters 4 | NRPS,T1PKS | 473,674 | 553,694 | tallysomycin | 55% |
| Clusters 5 | Terpene | 578,046 | 600,082 | tylactone | 6% |
| Clusters 6 | Terpene | 610,891 | 631,859 | - | |
| Clusters 7 | Lassopeptide,T1PKS | 638,187 | 696,947 | ulleungdin | 75% |
| Clusters 8 | Thiopeptide | 748,305 | 808,571 | geosmin | 100% |
| Clusters 9 | T1PKS,NRPS | 816,029 | 911,078 | peucechelin | 20% |
| Clusters 10 | T1PKS,NRPS-like | 930,265 | 1,105,264 | conglobatin | 26% |
| Clusters 11 | NRPS | 1,122,162 | 1,166,496 | diisonitrile antibiotic SF2768 | 100% |
| Clusters 12 | T1PKS,phenazine | 1,194,939 | 1,257,441 | aureothin | 100% |
| Clusters 13 | NRPS,T1PKS | 1,295,327 | 1,467,303 | sceliphrolactam | 48% |
| Clusters 14 | NRPS-like,arylpolyene | 1,547,967 | 1,590,881 | lankacidin C | 20% |
| Clusters 15 | RiPP-like | 1,632,931 | 1,644,841 | - | |
| Clusters 16 | Lassopeptide | 1,746,532 | 1,769,080 | lagmysin | 80% |
| Clusters 17 | Terpene | 1,779,551 | 1,800,171 | - | |
| Clusters 18 | T1PKS | 1,890,303 | 1,957,177 | cremimycin | 10% |
| Clusters 19 | NI-siderophore | 2,265,382 | 2,295,133 | - | |
| Clusters 20 | Terpene,NRPS | 2,725,486 | 2,770,746 | pyrroloformamide | 37% |
| Clusters 21 | Thioamide-NRP,T1PKS | 3,661,290 | 3,719,169 | 5-isoprenylindole-3-carboxylate β-D-glycosyl ester | 33% |
| Clusters 22 | Butyrolactone,T3PKS | 4,656,145 | 4,697,411 | neocarzinostatin | 8% |
| Clusters 23 | Terpene | 4,924,188 | 4,945,345 | ebelactone | 5% |
| Clusters 24 | T3PKS | 5,734,254 | 5,775,303 | violapyrone B | 28% |
| Clusters 25 | NI-siderophore | 6,121,786 | 6,151,618 | kinamycin | 19% |
| Clusters 26 | RiPP-like | 6,210,115 | 6,220,963 | - | |
| Clusters 27 | T3PKS,phenazine | 6,247,692 | 6,295,310 | endophenazine | 47% |
| Clusters 28 | T2PKS | 6,367,291 | 6,439,815 | allocyclinone | 43% |
| Clusters 29 | Phenazine,NRPS | 6,617,890 | 6,676,804 | streptophenazine | 24% |
| Clusters 30 | Indole | 6,688,763 | 6,712,068 | AT2433-A1 | 14% |
| Clusters 31 | NAPAA,terpene | 6,721,859 | 6,774,236 | hopene | 76% |
| Clusters 32 | NRPS-like | 6,831,419 | 6,873,260 | kitacinnamycin | 7% |
| Clusters 33 | NRPS-like | 6,898,154 | 6,961,149 | cyphomycin | 5% |
| Clusters 34 | Other,terpene | 6,985,496 | 7,026,419 | ECO-0501 | 4% |
| Clusters 35 | Other | 7,182,203 | 7,223,099 | griseusin | 15% |
| Clusters 36 | RiPP-like | 7,298,812 | 7,310,096 | - | |
| Clusters 37 | NRPS,T1PKS,terpene | 7,364,647 | 7,465,253 | hexacosalactone A | 18% |
| Clusters 38 | NRPS-like,CDPS | 7,500,628 | 7,543,702 | guanipiperazine | 80% |
| Clusters 39 | T1PKS,NRPS | 7,647,760 | 7,783,943 | qinichelins | 83% |
| Clusters 40 | T3PKS,T1PKS,furan | 7,816,344 | 7,894,681 | paerucumarin | 60% |
Table 4.
Effects of strain ASG80 extracts on mycelial growth of phytopathogenic.
| Pathogens | Regression equation | EC50 (μg/mL) | Correlation coefficient | |
|---|---|---|---|---|
| Phytophthora vignae | y = 1.3297x + 4.8299 | 1.3427 | 0.9864 | |
| Phytophthora sojae | y = 1.6772x + 4.6219 | 1.6806 | 0.9985 | |
| Phytophthora palmivora | y = 1.7603x + 4.1724 | 2.9521 | 0.9920 | |
| Phytophthora nicotiana | y = 0.8279x + 4.8283 | 1.6120 | 0.9952 | |
| Phytophthora melonis | y = 1.1621x + 4.5614 | 2.3846 | 0.9994 | |
| Phytophthora colocasiae | y = 1.7260x + 4.4184 | 2.1725 | 0.9687 | |
| Phytophthora cinnamomi | y = 1.6358x + 5.0999 | 0.8688 | 0.9998 | |
| Phytophthora capsici | y = 1.7841x + 4.1978 | 2.8159 | 0.9933 | |
| Pestalotiopsis palmarum | y = 1.3937x + 2.8532 | 34.6959 | 0.9895 | |
| Magnaporthe grisea | y = 1.1531x + 3.9087 | 8.8389 | 0.9807 | |
| Fusarium oxysporum | y = 1.1694x + 3.0012 | 51.2052 | 0.9956 | |
| Fusarium graminearum | y = 1.5994x + 3.1874 | 13.5908 | 0.9958 | |
| Colletotrichum gloeosporioides | y = 1.7642x + 3.1930 | 10.5738 | 0.9938 | |
Table 5.
The inhibitory effect of strain ASG80 extract on sisal zebra disease.
| Treatment | Incidence rate(%) | Disease index | Control effect (%) |
|---|---|---|---|
| CK | 0 | / | / |
| Pn | 100.00 | 52.45 | / |
| T-1 | 30.00 | 8.33 | 84.12 |
| T-2 | 46.67 | 18.43 | 64.86 |
| Me-2 | 36.67 | 11.99 | 77.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



