
Submitted:
28 October 2024
Posted:
28 October 2024
You are already at the latest version
Abstract
Fungi can remarkably sense and adapt to various extracellular stimuli and stress conditions. Oxidative stress, which results from an imbalance between reactive oxygen species production and antioxidant defenses, leads to cellular damage and death. In Trichophyton rubrum, oxidative stress is managed by a complex antioxidant system including thioredoxins, glutathione, catalases, peroxidases, and superoxide dismutase, with glutathione playing a crucial role. The fungus also responds to oxidative stress through critical pathways such as the glycerol high osmolarity pathway, activator protein 1 transcription factor, and responsive regulator SKN7. To better understand the role of the transcription factor StuA in regulating oxidative stress-related genes within these pathways, we conducted gene expression studies in ΔstuA mutant and wild-type strains of T. rubrum cultivated in keratin and under oxidative stress induced by hydrogen peroxide. Our results revealed significant downregulation of essential antioxidant genes, including glutathione transferases and catalases, in the ΔstuA mutant. Moreover, catalase and glutathione S-transferase activities were impaired in the mutants under stress conditions, highlighting the impact of this mutation. These findings underscore the critical role of StuA in the oxidative stress response and fungal pathogenesis and provide new insights into T. rubrum's adaptive mechanisms.
Keywords:
oxidative stress
; transcription factors
; glutathione
; catalase
; Trichophyton rubrum
; StuA
; HOG pathway
; AP1-SKN7
; hydrogen peroxide
1. Introduction
Trichophyton rubrum is the most common agent of dermatophytosis worldwide [1], primarily causing cutaneous infections in keratin-rich structures, such as nails and skin. This fungus secretes proteolytic enzymes that degrade keratinized structures, which are metabolized and used as a nutritional source during infection [2,3]. T. rubrum senses and adapts to the host environment to survive and thrive during infection, responding appropriately to environmental cues and the cellular stresses they induce. To this end, the fungus has evolved diverse adaptation mechanisms, including a complex interplay between signaling molecules and stress response pathways [4].
Oxidative stress occurs when there is an imbalance between the production of reactive oxygen species (ROS) and antioxidant defenses, leading to ROS levels that exceed the antioxidant capacity required to maintain the intracellular redox environment in a reduced state, ultimately resulting in cellular damage. ROS are generated as byproducts of normal cellular metabolism in response to environmental factors. Therefore, the ability of fungi to detect environmental signals, perform robust signal transduction, and trigger accurate responses is fundamental for their survival [5,6].
The systems responsible for removing or detoxifying ROS include superoxide dismutase, catalase, glutathione, and thioredoxins, with glutathione being the primary antioxidant system [6]. Additionally, glutathione is involved in iron metabolism owing to its requirement in Fe-S cluster assembly, sensing and regulation of iron levels, iron trafficking, and biosynthesis of iron cofactors [7,8].
However, literature on how fungi respond to oxidative stress is scarce. The prominent roles of three significant modulators have been highlighted: the glycerol high osmolarity (HOG) pathway, the yeast activation protein 1-like basic-leucine zipper (bZIP) transcription factor, and the response regulator and transcription factor SKN7 [5,6]. The HOG pathway, which is analogous to the p38 and Jun N-terminal kinase pathways in mammals, is highly conserved across the fungal kingdom, regardless of the species niche. This pathway plays a prominent role in adaptation to various stresses in fungal environments, including antifungals, antimicrobial peptides, and osmotic and oxidative stresses, and contributes to virulence [5,9]. Transcription factors can also orchestrate a robust signaling pathway by either activating or suppressing molecular responses based on the specific biological context to which the fungus is exposed [10,11]. The bZIP family of transcription factors plays a role in stress adaptation in fungi. Among these, the transcription factor activator protein 1 (Ap1) is a major oxidative stress response regulator and is homologous to the mammalian Jun/Fos bZIP transcription factors. Ap1 is activated via oxidation by redox-sensitive glutathione peroxidase 3 in Saccharomyces cerevisiae or thioredoxin peroxidase 3 in S. pombe [5,6]. Oxidized Ap1 accumulates in the nucleus and triggers expression of oxidative stress-related genes. Additionally, Ap1 interacts with the transcription factor SKN7 to enhance the transcription of oxidative stress-related genes [12].
Regarding transcription factors involved in regulating oxidative stress, previous reports have demonstrated that the APSES transcription factor StuA may play a role in this response [13,14]. However, the mechanism by which StuA regulates the expression of oxidative stress-related genes remains unclear. Additionally, this transcription factor is multifaceted as a critical regulator of metabolism-, adhesion-, and pathogenicity-related genes [13,15,16,17].
Mining RNA sequencing data from the mutant strain ΔstuA during growth in keratin [13], we identified that some genes involved in oxidative stress were downregulated in the mutant strain compared with the wild-type (WT). Based on these findings, we hypothesized that StuA plays a role in the regulation of oxidative stress-related genes. Therefore, this study aimed to evaluate the transcriptional regulation of oxidative stress pathways, specific genes, and metabolic aspects, focusing on catalase and glutathione detoxification systems in WT and ΔstuA strains during growth in keratin and under oxidative stress induced by hydrogen peroxide. Our results demonstrated, for the first time, the importance of StuA in a specific class of glutathiones, enhancing our understanding of oxidative stress response mechanisms in T. rubrum and underscoring the pivotal role of transcription factors, such as StuA, in fungal pathogenesis.
2. Results
2.1. Search for Genes in T. rubrum Homologous to Genes Related to Oxidative Stress in Other Trichophyton Species
We searched for oxidative stress-related gene homologs in T. rubrum, considering that the most involved in oxidative stress pathways are hypothetical proteins. Table 1 shows the homologous oxidative stress-related genes, considering genes with >85% identity.
2.2. The Transcription Factor StuA Regulates Genes Related to Oxidative Stress Pathways in T. rubrum
We investigated the influence of StuA on the regulation of the transcriptional response of the central genes involved in oxidative stress pathways during fungal growth in keratin (Figure 1) and oxidative stress induction with fungal exposure to hydrogen peroxide (Figure 2). Our results showed that the gene encoding the response regulator Ssk1 (TERG_07855) of the HOG pathway was downregulated under both conditions compared with that in the WT strain. In contrast, the gene ypd1 (TERG_07131), also involved in the HOG pathway, only reduced transcript levels when ΔstuA strain was grown in keratin for 24 h.
Regarding the AP1-SKN7 pathway, we observed that the transcript levels of the gene encoding the transcription factor Ap1 (TERG_02940) were lower in the ΔstuA strain than in the WT strain only when fungi were exposed to hydrogen peroxide. The transcriptional response of the gene encoding transcription factor Skn7 (TERG_01117) varied in the mutant strain depending on the duration of fungal culture in keratin or exposure to hydrogen peroxide. The absence of StuA consistently reduced the transcript levels of the gene encoding the Seb1 transcription factor (TERG_06759) under all the conditions evaluated in this study.
2.3. Transcript Levels of Glutathione S-Transferase and a Catalase-like Gene Are Reduced in the Mutant Strain
Given that some genes involved in oxidative stress pathways were downregulated in the mutant strain compared with those in the WT strain, we analyzed the transcript levels of genes encoding members of the glutathione family and catalases during fungal growth in keratin (Figure 3) and during hydrogen peroxide exposure (Figure 4).
Our results showed that genes encoding glutathione S-transferases (TERG_07326 and TERG_01685) and a catalase-like gene (TERG_02005) were downregulated in the mutant strain compared with those in the WT strain under all conditions evaluated in this study. The glutathione peroxidase-encoding gene (TERG_01349) was downregulated only during hydrogen peroxide exposure. The transcript levels of a glutathione synthetase gene (TERG_04073) were overexpressed in the ΔstuA strain compared with the WT strain during fungal growth in keratin and after 3 h of hydrogen peroxide exposure, suggesting that StuA may act as a regulator of the gene responsible for glutathione production. Regarding the glutathione reductase-coding gene (TERG_08069) and the catalase A-coding gene (TERG_01252), the mutant strain maintained expression levels similar to the WT strain after 48 h of fungal growth in keratin or showed induced expression after 3 h of hydrogen peroxide exposure.
2.4. The Transcription Factor StuA as a Consensus-Binding Site for a Gene Encoding a Glutathione S-Transferase in T. rubrum
We conducted a bioinformatics analysis to determine the existence of a consensus-binding site for the transcription factor StuA in all genes evaluated in this study. Our analysis revealed a consensus-binding site for StuA in TERG_07326, which encodes a glutathione S-transferase (Figure 5). No consensus-binding site for StuA was found in the other evaluated genes.
2.5. The Mutant ΔstuA Exhibited Lower Catalase Activity
To investigate the regulatory role of the transcription factor StuA in the oxidative stress response, we measured the intracellular catalase activity for 10 and 15 min in protein extracts from fungal growth in keratin for 24 h (Figure 6A) and 48 h (Figure 6B), and from fungal exposure to hydrogen peroxide for 3 h (Figure 6C). Our results showed reduced catalase activity in the ΔstuA mutant strain under all conditions evaluated in this study. Catalase activity was not observed in protein extracts from fungi exposure to hydrogen peroxide for 1 h.
2.6. The ΔstuA Mutant Exhibited Reduced Glutathione S-Transferase Activity During Cultivation in Keratin and Under Hydrogen Peroxide Exposure
We investigated whether glutathione S-transferase activity was affected by the stuA mutation. Our results showed that the activity of glutathione S-transferase was notably impaired in the ΔstuA mutant compared with the WT strain during cultivation in keratin (Figure 7A) and under hydrogen peroxide exposure (Figure 7B).
2.7. Genes Involved in Iron Metabolism Are Downregulated When the ΔstuA Strain is Exposed to Hydrogen Peroxide
Iron is an essential metal that acts as a cofactor for antioxidant enzymes. However, in excess, it can participate in Fenton reactions, producing highly reactive hydroxyl radicals and exacerbating oxidative stress. In this study, we analyzed two genes involved in the siderochrome iron transport (TERG_03174 and TERG_04949) and a nonribosomal siderophore peptide synthase (TERG_00697) during fungal growth in keratin and hydrogen peroxide exposure.
Our results reveal that these genes were upregulated in the ΔstuA mutant strain, particularly after 48 h of growth in keratin. However, when the mutant strain was exposed to hydrogen peroxide, the transcript levels of these genes decreased compared with those in the WT strain, except for TERG_03174, which remained upregulated after 3 h of exposure. (Figure 8).
2.8. The Intracellular Iron Content Is Higher in the ΔstuA Strain After Three Hours of Exposure to Hydrogen Peroxide
We also investigated the intracellular iron content in both the WT and mutant strains during cultivation in keratin and under oxidative stress. Our results revealed higher intracellular iron content in the WT strain after 48 h of growth in keratin. However, when exposed to oxidative stress with hydrogen peroxide, the ΔstuA mutant strain exhibited increased intracellular iron levels after 3 h (Figure 9).
3. Discussion
Regulation of the oxidative stress response is crucial for pathogen survival and thrive in the host. Transcription factors play an essential role in modulating the expression of genes involved in detoxifying ROS and managing oxidative stress pathways, thereby maintaining cellular redox homeostasis.
In this study, we investigated the role of the transcription factor StuA in the regulation of oxidative stress response pathways in T. rubrum. Our findings revealed, for the first time, that StuA significantly influences the expression of genes involved in oxidative stress and other related transcription factors. Several genes, including those involved in oxidative stress pathways, are identified in T. rubrum as encoding hypothetical proteins. Therefore, we conducted an orthological analysis to identify potential orthologs of these genes in T. rubrum based on other dermatophytes.
The absence of StuA substantially impaired the expression of TERG_07855, a gene that encodes the response regulator (ssk1), which is crucial for activating Hog1. The Hog1 pathway, one of the most evolutionarily conserved pathways in eukaryotes, forms the backbone of stress signaling via a two-component system-dependent mechanism. Although not present in mammals, the eukaryotic two-component system involves a multistep histidine-aspartate phosphorylation mechanism consisting of a hybrid histidine kinase and a histidine-containing phosphotransfer protein, such as TERG_07131, which encodes the Ypd1 gene. Histidine-containing phosphotransfer mediates histidine kinase and the response regulator ssk1, leading to a cascade of mitogen-activated protein kinase phosphorylation and subsequent Hog1 activation [5,18]. Our results suggest that even if Ypd1 expression is not significantly affected by StuA deletion during fungal growth in keratin or hydrogen peroxide exposure, the cascade of mitogen-activated protein kinase phosphorylation leading to Hog1 activation may be compromised because of the lower transcript levels of ssk1 in the mutant strain than in the WT strain under all conditions evaluated. Additionally, reduced levels of hog1 transcripts in the T. rubrum ΔstuA mutant during fungal cultivation in keratin [16] suggest that StuA may play a critical role in modulating the oxidative stress response by influencing key components of this pathway.
Our research reveals a complex interplay between transcription factors in response to environmental stress. We found that transcript levels of the ap1 coding gene (TERG_02940) were downregulated in the ΔstuA strain when exposed to oxidative stress compared with the WT strain. The opposite effect was observed during cultivation in keratin-containing medium. Ap1 can also interact with other transcription factors, such as Skn7, which is involved in the antioxidant response. Approximately half of the genes regulated by Ap1 require interaction with Skn7, whereas the other half are independent of Skn7 [19]. The exact mechanism of Skn7 activation via phosphorylation remains unclear. However, once activated, Skn7 cooperates with oxidized Ap1 to regulate oxidative stress-related genes [5]. In the present study, we showed that the skn7 coding gene (TERG_01117) did not show a consistent pattern of regulation by StuA, fluctuating between upregulation and downregulation across different conditions and time points.
Analysis of a Δap1 strain of T. rubrum indicated that Ap1 acts as a negative regulator of specific virulence attributes. However, deletion of ap1 did not affect susceptibility to oxidative stress induced by hydrogen peroxide [20]. To the best of our knowledge, there is no evidence of an interaction between Ap1 and StuA. Based on our findings, we propose that StuA acts as a positive regulator of Ap1 during oxidative stress exposure to hydrogen peroxide and a negative regulator of Ap1 during keratin decomposition.
Additionally, the gene encoding the C2H2 transcription factor Seb-1 (TERG_06759) was consistently downregulated in the ΔstuA strain compared with the WT strain, suggesting that StuA positively regulates this transcription factor. Although not specifically a part of the oxidative stress response pathway, Seb-1 binds to the Stress Response Element under heat stress. In Neurospora crassa, a Δseb1 mutant strain was more sensitive to oxidative stress during hydrogen peroxide exposure [21], and Seb-1 is also involved in oxidative stress sensitivity in Valsa mali [22]. However, there is no evidence that StuA directly regulates or cooperates with Seb1 or other transcription factors. Additionally, no consensus-binding site for StuA was identified in any of the genes involved in the oxidative stress pathways evaluated in this study, suggesting that this regulation might be indirect or mediated through other transcription factors not addressed here.
Considering the obtained results regarding genes involved in oxidative stress pathways, we hypothesize that the ΔstuA mutant strain may be more susceptible to oxidative stress effects during hydrogen peroxide exposure owing to the reduced transcript levels of essential genes in the main AP1-SKN7 and HOG1 oxidative stress response pathways.
Regarding antioxidant enzyme-coding genes, we focused on evaluating the transcript levels of the glutathione family and catalases. Our results suggest that StuA can indirectly regulate the transcriptional response of TERG_02005, a catalase-like gene, as the transcript levels were lower in the mutant strain than in the WT under all conditions evaluated. In contrast, the catalase A-coding gene (TERG_01252) did not show a consistent regulation pattern, alternating between down and upregulation depending on time and culture conditions. The regulatory mechanisms of specific gene transcription factors are complex and involve several factors. Similarly, previous RNA sequencing data of the ΔstuA strain during growth in keratin showed that catalase genes in T. rubrum were downregulated [13].
Additionally, our analysis of intracellular catalase enzymatic activity showed reduced activity in the mutant strain in protein extracts during fungal growth in keratin for 24 and 48 h and in the protein extract after fungal exposure to hydrogen peroxide for 3 h. These findings further confirm the reduced levels of transcription and enzymatic activity of catalases in mutants lacking the transcription factor StuA, which has significant implications for our understanding of the critical role of this transcription factor in regulating catalase activity and managing oxidative stress responses [14,16,23].
Our findings also indicate that the absence of StuA affects the transcriptional regulation of multiple glutathione-related genes. Specifically, StuA appears to act as both a positive and negative regulator of transcript levels for genes encoding glutathione synthetase (TERG_04073), which is responsible for glutathione synthesis, and glutathione reductase, which maintains reduced glutathione levels in cells for cellular homeostasis, depending on time and culture conditions. The absence of StuA diminished the transcription of the glutathione peroxidase gene (TERG_01349) in the presence of hydrogen peroxide, suggesting that the ability to reduce peroxide in cells could be compromised in the mutant strain.
Our study showed that the absence of StuA significantly affected a specific class within the glutathione family. This led to a downregulation of both glutathione S-transferase coding genes (TERG_07326 and TERG_01685) in the ΔstuA strain compared with the WT strain across all conditions evaluated in this study. Furthermore, we identified a consensus StuA-binding site in TERG_07326, suggesting direct regulation by this transcription factor. Analysis of the enzymatic activity of this class of enzymes also revealed diminished activity in the mutant strain.
Glutathione S-transferases (GSTs) are multifunctional enzymes primarily involved in detoxification. They catalyze the conjugation of glutathione to various toxic compounds, facilitating their excretion from the cells. Additionally, GSTs are involved in cell signaling, regulation of apoptosis, and resistance to chemotherapeutic drugs [24]. Accordingly, mutants with deletions in the GST gene of various fungi show increased sensitivity to xenobiotics and different stressors [8]. For instance, in S. pombe, gst1Δ, gst2Δ, and gst3Δ mutants were sensitive to the antifungal fluconazole, indicating the role of GSTs in mediating drug resistance [8]. In S. cerevisiae, gtt1Δ gtt2Δ, and gtt1Δgtt2Δ mutants displayed increased sensitivity to heat shock or growth defects at high temperatures. In Aspergillus nidulans, the gstAΔ mutant was sensitive to heavy metals [8,25]. To the best of our knowledge, no previous studies have investigated the role of StuA in regulating glutathione family genes. However, it is plausible to hypothesize that a deficiency in the conjugation and excretion of toxic compounds in the glutathione-mediated detoxification system may occur in the mutant strain, which could increase its sensitivity to toxic compounds and oxidative stress.
Deficiency of the glutathione-mediated detoxification system leads to an imbalance in iron metabolism. Abnormal iron metabolism causes iron overload, leading to the Fenton reaction and increasing the generation of ROS [7,26,27]. Iron acquisition is essential during the infectious process of dermatophytosis because this metal is vital for many cellular functions, including respiration and energy production. Iron plays crucial roles in the survival and virulence of pathogens during dermatophyte infections. Dermatophytes such as T. rubrum need to acquire iron from the environment to sustain their growth and pathogenicity, mainly when infecting keratinized tissues such as the skin and nails [28,29].
Our results showed that genes encoding siderochrome iron transporters and a nonribosomal siderophore peptide synthase, which are essential for iron acquisition, transport, and metabolism, were upregulated in the mutant strain during fungal growth in keratin compared with the WT strain. The WT strain presented higher intracellular iron content under the same growth conditions. It is possible to hypothesize that during growth in keratin, the absence of StuA may lead to the upregulation of genes involved in iron metabolism as a compensatory strategy to enhance the uptake of this essential metabolite and mitigate the harmful effects of the mutation. However, despite this upregulation, there was no accumulation of intracellular iron, suggesting that iron transport and distribution within mutant cells may also be compromised.
During exposure to hydrogen peroxide, we observed that most genes involved in iron metabolism were downregulated in the mutant strain compared with those in the WT strain. However, the intracellular iron levels of the mutant strain increased from 3 h onwards, which may indicate that despite the lower transcript levels of iron metabolism genes in the mutant strain, there was an increase in intracellular iron as a strategy to enhance the efficiency of its antioxidants, as iron can act as a cofactor for catalases [30] which was shown to be impaired in our results. Alternatively, it could also indicate increased cellular intoxication due to a deficiency in GSTs or other components of the glutathione system, potentially leading to significant cellular toxicity.
4. Materials and Methods
4.1. Search for Oxidative Stress-Related Gene Homologs in T. rubrum CBS118892
We searched for the stress response transcription factor SKN7, C2H2 transcription factor SEB1, phosphotransfer protein Ypd1, response regulator SSK1, glutathione S-transferase, and catalase homolog proteins in T. rubrum CBS 118892 using T. benhamiae, T. equinum, T. verrucosum, and T. tonsurans, respectively, as queries (ARB_07479, ARB_00366, TEQG_03843, TRV_02413, ARB_02229, and TESG_07337, available at Ensembl Fungi https://fungi.ensembl.org/). We performed multiple sequence alignments using the Standard Protein BLAST (Basic Local Alignment Search Tool, National Center for Biotechnology Information, USA (https://www.ncbi.nlm.nih.gov/)), using non-redundant protein sequences as a database.
4.2. Fungal Strains and Culture Conditions
We used the T. rubrum strain CBS118892 (Westerdijk Biodiversity Institute, Utrecht, The Netherlands) as a reference (WT) and the previously constructed null mutant strain ΔstuA [15]. The strains were grown on solid malt extract agar medium (2% glucose, 2% malt extract, 0.1% peptone, and 2% agar, pH 5.7) at 28°C for 20 days. A conidial suspension was then prepared by flooding the plates with a 0.9% sterile NaCl solution and filtering the suspension through the fiberglass to remove hyphal fragments. Conidial concentration was estimated using a Neubauer chamber. Approximately 1 × 106 conidia of each strain were inoculated into 100 mL of Sabouraud dextrose broth and incubated at 28°C for 96 h in an orbital shaker with agitation (120 rpm).
For experiments using keratin-containing medium, the resulting mycelia were transferred to 100 mL of minimal medium [31] containing 70 mM sodium nitrate (Sigma-Aldrich, St. Louis, MO, USA) and 0.5% bovine keratin (m/v). The cultures were incubated for 24 and 48 h at 28°C with constant agitation (120 rpm). The biological material from three independent replicates was filtered at each time point and stored at −80°C until RNA extraction.
4.3. Oxidative Stress Induction with Hydrogen Peroxide
For experiments involving fungal growth in a medium containing hydrogen peroxide, we first performed a minimum inhibitory concentration (MIC) assay according to the M38-A reference method recommended by the Clinical and Laboratory Standards Institute, with the following modification: 100 μL of the conidial suspension, amounting to 6 × 10⁴ conidia/mL of WT and mutant strains, was added to each well in a 96-well plate. The final conidial concentration was adjusted to approximately 3 × 10⁴ conidia per well in the Sabouraud medium. Serial dilutions of hydrogen peroxide ranged between 1.2 and 0.018 mM. MIC was established at 0.3 mM for both strains.
Next, conidial suspensions of both strains were prepared as described in Section 4.2. The resulting mycelia were transferred to 100 mL of Sabouraud medium containing 70% of the MIC (0.21 mM). The cultures were incubated for 1 and 3 h at 28°C with constant agitation (120 rpm). Sabouraud medium without hydrogen peroxide was used as the control.
Subsequently, biological material from three independent replicates of Sabouraud cultures with hydrogen peroxide was filtered at each time point and stored at −80°C until RNA extraction.
4.4. RNA Extraction and cDNA Synthesis
According to the manufacturer's instructions, total RNA was extracted using the Illustra RNAspin Mini Isolation Kit (GE Healthcare, Chicago, IL, USA). RNA concentration and purity were assessed using a NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Total RNA was treated with DNase I (Thermo Fisher Scientific, Waltham, MA, USA) to prevent genomic DNA contamination. Subsequently, cDNA synthesis was performed using the Platus Transcriber RNase H-cDNA First Strand Kit (Sinapse Inc., Miami, FL, USA) following the manufacturer’s instructions.
To assess the quality of the cDNA, we conducted a polymerase chain reaction using oligonucleotides to amplify a region of the constitutive β-tubulin gene, followed by analysis using agarose gel electrophoresis. cDNA was diluted to 70 ng/μL for reverse transcription-quantitative polymerase chain reaction.
4.5. Reverse Transcription-Quantitative Polymerase Chain Reaction Analyses
We used a QuantStudio 3 Real-Time PCR System (Applied Biosystems, Waltham, MA, USA) with the primers listed in Supplementary Table S1 for transcript quantification. The concentration of each primer was standardized to achieve reaction efficiencies between 90% and 110%. The reactions were prepared using Power SYBR™ Green PCR Master Mix (Applied Biosystems, Waltham, MA, USA) with ROX dye as a fluorescent normalizer [32]. The 2−ΔΔCt method was used for relative expression analysis [33], with the T. rubrum genes gapdh and rpb2 serving as robust endogenous controls [34]. The results are presented as the mean relative expression values from three independent replicates with standard deviations.
4.6. Metabolic Assays
We used the macerated mycelium of WT and ΔstuA strains during fungal growth in keratin and hydrogen peroxide exposure, as described previously [17].
4.6.1. Catalase Activity Assay
For the catalase activity assay, a hydrogen peroxide solution (2 mL at a final concentration of 10 mM) prepared in 50 mM phosphate buffer (pH 7.0) was used as a substrate for the reaction, followed by the addition of 100 μg of protein extract diluted in 50 μL of the appropriate buffer, as described previously [16]. The decomposition of hydrogen peroxide was monitored over time by measuring the absorbance at 240 nm for 10 and 15 min at room temperature. To infer the enzymatic activity of catalase in each protein extract, the amount of hydrogen peroxide consumed per minute per milligram of protein was measured. Values are presented as means followed by the standard deviation of three independent biological replicates.
4.6.2. Intracellular Iron Assay
For the intracellular iron assay, we used a Serico Iron Kit (Labtest Diagnóstica, Lagoa Santa, MG, Brazil) according to the manufacturer’s instructions. Absorbance readings were taken at 550 nm using a Multiskan™ Microplate Photometer (ThermoFisher Scientific, Waltham, MA, USA), and the results are presented as means followed by the standard deviation of three independent biological replicates.
4.6.3. GST Enzymatic Activity Assay
GST-specific enzymatic activity was quantified using a GST Activity Assay Kit (Elabscience, Houston, TX, USA) according to the manufacturer’s instructions. First, 100 μg of protein extract was mixed with the appropriate buffer and substrate solution. The GST activity was calculated as U/mg by measuring the rate of increase in absorbance at 340 nm. The results are presented as the mean ± standard deviation of three independent biological replicates. One unit of GST activity was defined as the amount of enzyme in 1 mL of the sample that catalyzed the conjugation of 1 μmol of 1-chloro-2,4-dinitrobenzene with reduced glutathione at 37°C per minute.
4.7. Statistical Analysis
An unpaired t-test was used to statistically analyze transcript quantification and enzymatic assay results. Statistical significance was determined using the Holm-Sidak method, with p < 0.05. Significance is represented in the graphs as * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001. GraphPad Prism software v.6 (GraphPad Software, San Diego, CA, USA) [35] was used for statistical analysis and graph design.
5. Conclusions
Our results showed that the transcription factor StuA is crucial for regulating genes involved in oxidative stress response pathways and iron homeostasis in T. rubrum. The absence of stuA impairs the activation of the Hog1 pathway genes and negatively affects the expression of oxidative stress-related genes, such as ap1 and seb-1 while altering intracellular iron levels, catalase, and glutathione S-transferase activity. These findings highlight the role of StuA in the fungus's adaptation and survival under stress, thereby enhancing our understanding of the mechanisms underlying the pathogenicity of T. rubrum during infection.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: List of primers used.
Author Contributions
M.F.P. designed and conducted the experiments and wrote the manuscript. L.M.-S. supported the catalase enzymatic activity assay and contributed to the final version of the manuscript. V.M.O. performed fungal cultures and assisted with RNA and protein extraction. P.R.S. conducted the bioinformatic analysis. N.M.M.-R. and A.R. supervised the experimental design and execution and edited the manuscript. All authors reviewed and approved the submitted manuscript and agreed to the published version.
Funding
This work was supported by grants from the Brazilian Agencies: São Paulo Research Foundation—FAPESP (proc. No. 2019/22596-9, postdoctoral scholarship Nos. 2021/10359-2 to MP and 2021/10255-2 to LM-S); the National Council for Scientific and Technological Development—CNPq (grant Nos. 307871/2021-5 and 307876/2021-7); the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)—Finance Code 001; and the Fundação de Apoio ao Ensino, Pesquisa e Assistência—FAEPA.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article and the accompanying Supplementary Data.
Acknowledgments
The authors thank Mendelson Mazzucato and Marcos D. Martins for their technical support.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflict of interest.
References
- Petrucelli, M.F.; Abreu, M.H.; Cantelli, B.A.M.; Segura, G.G.; Nishimura, F.G.; Bitencourt, T.A.; Fachin, A. Epidemiology and Diagnostic Perspectives of Dermatophytoses. Journal of Fungi 2020, 6, 310. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Kawa, A.; Kanarek, K.; Sobon, A.; Szewczyk, R. Metabolomic Analysis of Trichophyton rubrum and Microsporum canis During Keratin Degradation. Scientific Reports 2021, 11, 3959. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rossi, N.M.; Peres, N.T.A.; Bitencourt, T.A.; Martins, M.P.; Rossi, A. State-of-the-Art Dermatophyte Infections: Epidemiology Aspects, Pathophysiology, and Resistance Mechanisms. Journal of Fungi (Basel) 2021, 7, 629. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rossi, N.M.; Bitencourt, T.A.; Peres, N.T.A.; Lang, E.A.S.; Gomes, E.V.; Quaresemin, N.R.; Martins, M.P.; Lopes, L.; Rossi, A. Dermatophyte Resistance to Antifungal Drugs: Mechanisms and Prospectus. Frontiers in Microbiology 2018, 9, 1108. [Google Scholar] [CrossRef]
- Yaakoub, H.; Mina, S.; Calenda, A.; Bouchara, J.-P.; Papon, N. Oxidative Stress Response Pathways in Fungi. Cellular and Molecular Life Sciences 2022, 79, 333. [Google Scholar] [CrossRef]
- Zadrąg-Tęcza, R.; Maślanka, R.; Bednarska, S.; Kwolek-Mirek, M. Response Mechanisms to Oxidative Stress in Yeast and Filamentous Fungi, Stress Response Mechanisms in Fungi: Theoretical and Practical Aspects; Springer International Publishing: Cham, 2018; pp. 1–34. [Google Scholar]
- Berndt, C.; Lillig, C.H. Glutathione, Glutaredoxins, and Iron. Antioxidants & Redox Signaling 2017, 27, 1235–1251. [Google Scholar]
- Wangsanut, T.; Pongpom, M. The Role of the Glutathione System in Stress Adaptation, Morphogenesis and Virulence of Pathogenic Fungi. International Journal of Molecular Science 2022, 23, 10645. [Google Scholar] [CrossRef]
- Yaakoub, H.; Sanchez, N.S.; Ongay-Larios, L.; Courdavault, V.; Calenda, A.; Bouchara, J.; Coria, R.; Papon, N. The High Osmolarity Glycerol (HOG) Pathway in Fungi. Critical Reviews in Microbiology 2022, 48, 657–695. [Google Scholar] [CrossRef]
- Weidemller, P.; Kholmatov, M.; Petsalaki, E.; Zaugg, J.B. Transcription Factors: Bridge Between Cell Signaling and Gene Regulation. Proteomics 2021, 21, 2000034. [Google Scholar] [CrossRef]
- Shelest, E. Transcription Factors in Fungi. FEMS microbiology letters 2008, 286, 145–151. [Google Scholar] [CrossRef]
- Mulford, K.E.; Fassler, J.S. Association of the Skn7 and Yap1 Transcription Factors in the Saccharomyces cerevisiae Oxidative Stress Response. Eukaryotic Cell 2011, 10, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Bitencourt, T.A.; Neves-Da-Rocha, J.; Martins, M.P.; Sanches, P.R.; Lang, E.A.S.; Bortolossi, J.C.; Rossi, A.; Martinez-Rossi, N.M. StuA-Regulated Processes in the Dermatophyte Trichophyton rubrum: Transcription Profile, Cell-Cell Adhesion, and Immunomodulation. Frontiers in Cellular and Infection Microbiology 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D. C.; Doedt, T.; Chiang, L.Y.; Kim, H.S.; Chen, D.; Nierman, W.C.; Filler, S.G. The Aspergillus fumigatus StuA Protein Governs the Up-Regulation of a Discrete Transcriptional Program During the Acquisition of Developmental Competence. Molecular biology of the cell 2005, 16, 5866–5879. [Google Scholar] [CrossRef] [PubMed]
- Elza, A.S.; Bitencourt, T.A.; Peres, N.T.A.; Lopes, L.; Silva, L.G.; Cazzaniga, R.A.; Rossi, A.; Martinez-Rossi, N.M. The stuA Gene Controls Development, Adaptation, Stress Tolerance, and Virulence of the dermatophyte Trichophyton rubrum. Microbiological Research 2020, 241, 126592. [Google Scholar]
- Martins-Santana, L.; Petrucelli, M.F.; Sanches, P.R.; Almeida, F.; Martinez-Rossi, N.M.; Rossi, A. The StuA Transcription Factor and Alternative Splicing Mechanisms Drive the Levels of MAPK Hog1 Transcripts in the Dermatophyte Trichophyton rubrum. Mycopathologia 2024, 189, 37. [Google Scholar] [CrossRef]
- Petrucelli, M.F.; Martins-Santana, L.; Sanches, P.R.; Oliveira, V.M.; Rossi, A.; Martinez-Rossi, N.M. The Transcription Factor StuA Regulates the Glyoxylate Cycle in the Dermatophyte Trichophyton rubrum under Carbon Starvation. International Journal of Molecular Sciences 2024, 25, 405. [Google Scholar] [CrossRef]
- Liao, B.; Ye, X.; Chen, X.; Zhou, Y.; Cheng, L.; Zhou, X.; Ren, B. The Two-Component Signal Transduction System and its Regulation in Candida albicans. Virulence 2021, 12, 1884–1899. [Google Scholar]
- Mendoza-Martínez, A.E.; Cano-Domínguez, N.; Aguirre, J. Yap1 Homologs Mediate More than the Redox Regulation of the Antioxidant Response in Filamentous Fungi. Fungal Biology 2020, 124, 253–262. [Google Scholar] [CrossRef]
- Peres, N.T.A.; Lang, E.A.; Bitencourt, T.A.; Oliveira, V.M.; Fachin, A.L.; Rossi, A.; Martinez-Rossi, N.M. The bZIP Ap1 Transcription Factor is a Negative Regulator of Virulence Attributes of the Anthropophilic Dermatophyte Trichophyton rubrum. Current Research in Microbial Sciences 2022, 3, 100132. [Google Scholar] [CrossRef]
- Freitas, F.Z.; Virgilio, S.; Cupertino, F.B.; Kowbel, D.J.; Fioramonte, M.; Gozzo, F.C.; Glass, L.; Bertoloni, M.C. The SEB-1 Transcription Factor Binds to the STRE Motif in Neurospora crassa and Regulates a Variety of Cellular Processes Including the Stress Response and Reserve Carbohydrate Metabolism. G3 (Genes, Genomes, Genetics 2016, 6, 1327–1343. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, L.; Yin, Z.; Feng, H.; Huang, L. Transcription Factor VmSeb1 is Rrequired for the Growth, Development, and Virulence in Valsa mali. Microbial Pathogenesis 2018, 123, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Scherer, M.; Wei, H.; Liese, R.; Fischer, R. Aspergillus nidulans Catalase-Peroxidase Gene (cpeA) is Transcriptionally Induced During Sexual Development Through the Transcription Factor StuA. Eukaryotic Cell 2002, 1, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.R.; Reindl, K.M. Glutathione S-Transferases in Cancer. Antioxidants 2021, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.A.; Davis, M.A.; Hynes, M.J. A Gene from Aspergillus nidulans With Similarity to URE2 of Saccharomyces cerevisiae Encodes a Glutathione S-transferase Which Contributes to Heavy Metal and Xenobiotic Resistance. Applied and Environmental Microbiology 2002, 68, 2802–2808. [Google Scholar] [CrossRef]
- Lok, H.C.; Richardson, V.; Kalinowski, D. S.; Kovacevic, Z.; Lane, D.J.R.; Richardson, D.R. Glutathione S-transferase and MRP1 Form an Integrated System Involved in the Storage and Transport of Dinitrosyl–dithiolato Iron Complexes in Cells. Free Radical Biology and Medicine 2014, 75, 14–29. [Google Scholar] [CrossRef]
- Gao, X.; Hu, W.; Qian, D.; Bai, X.; He, H.; Li, L.; Sun, S. The Mechanisms of Ferroptosis Under Hypoxia. Cellular and Molecular Neurobiology 2023, 43, 3329–3341. [Google Scholar] [CrossRef]
- Stelitano, G.; Cocorullo, M.; Mori, M.; Villa, S.; Meneghetti, F.; Chiarelli, L.R. Iron Acquisition and Metabolism as a Promising Target for Antimicrobials (Bottlenecks and Opportunities): Where Do We Stand? International Journal of Molecular Sciences 2023, 24, 6181. [Google Scholar] [CrossRef]
- Kröber, A.; Scherlach, K.; Hortschansky, P.; Shelest, E.; Staib, P.; Kniemeyer, O.; Brakhage, A.A. HapX Mediates Iron Homeostasis in the Pathogenic Dermatophyte Arthroderma benhamiae but Is Dispensable for Virulence. Plos One 2016, 11, e0150701. [Google Scholar] [CrossRef]
- Pradhan, A.; Herrero-de-Dios, C.; Belmonte, R.; Budge, S.; Garcia, A.L.; Kolmogorova, A.; Lee, K.K.; Martin, B.D.; Ribeiro, A.; Bebes, A.; Yuecel, R.; Gow, N.A.R.; Munro, C.A.; MacCallum, D.M.; Quinn, J.; Brown, A.J. Elevated Catalase Expression in a Fungal Pathogen is a Double-Edged Sword of Iron. 2017, 13, e1006405. [Google Scholar] [CrossRef]
- Cove, D.J. The Induction and Repression of Nitrate Reductase in the Fungus Aspergillus nidulans. Biochimica et Biophysica Acta (BBA) - Enzymology and Biological Oxidation 1966, 113, 51-56.
- Jacobson, L.S.; Mclntyre, L.; Mykusz, J. Assessment of Real-Time PCR Cycle Threshold Values in Microsporum canis Culture-Positive and Culture-Negative Cats in an Animal Shelter: a Field Study. Journal of feline medicine and surgery 2018, 20, 108–113. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2- ΔΔCT method. methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jacob, T.R.; Peres, N.T.A.; Persinoti, G.F.; Silva, L.G.; Mazucato, M.; Rossi, A.; Martinez-Rossi, N.M. rpb2 is a Reliable Reference Gene for Quantitative Gene Expression Analysis in the Dermatophyte Trichophyton rubrum. Medical Mycology 2012, 50, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Swift, M.L. GraphPad Prism, Data Analysis, and Scientific Graphing. Journal of chemical information and computer sciences 1997, 37, 411–412. [Google Scholar] [CrossRef]
Figure 1.
Relative expression analysis of genes involved in oxidative stress pathways during fungal growth in keratin (minimal medium (MM) containing 0.5% of bovine keratin) for 24 and 48 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
Figure 1.
Relative expression analysis of genes involved in oxidative stress pathways during fungal growth in keratin (minimal medium (MM) containing 0.5% of bovine keratin) for 24 and 48 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
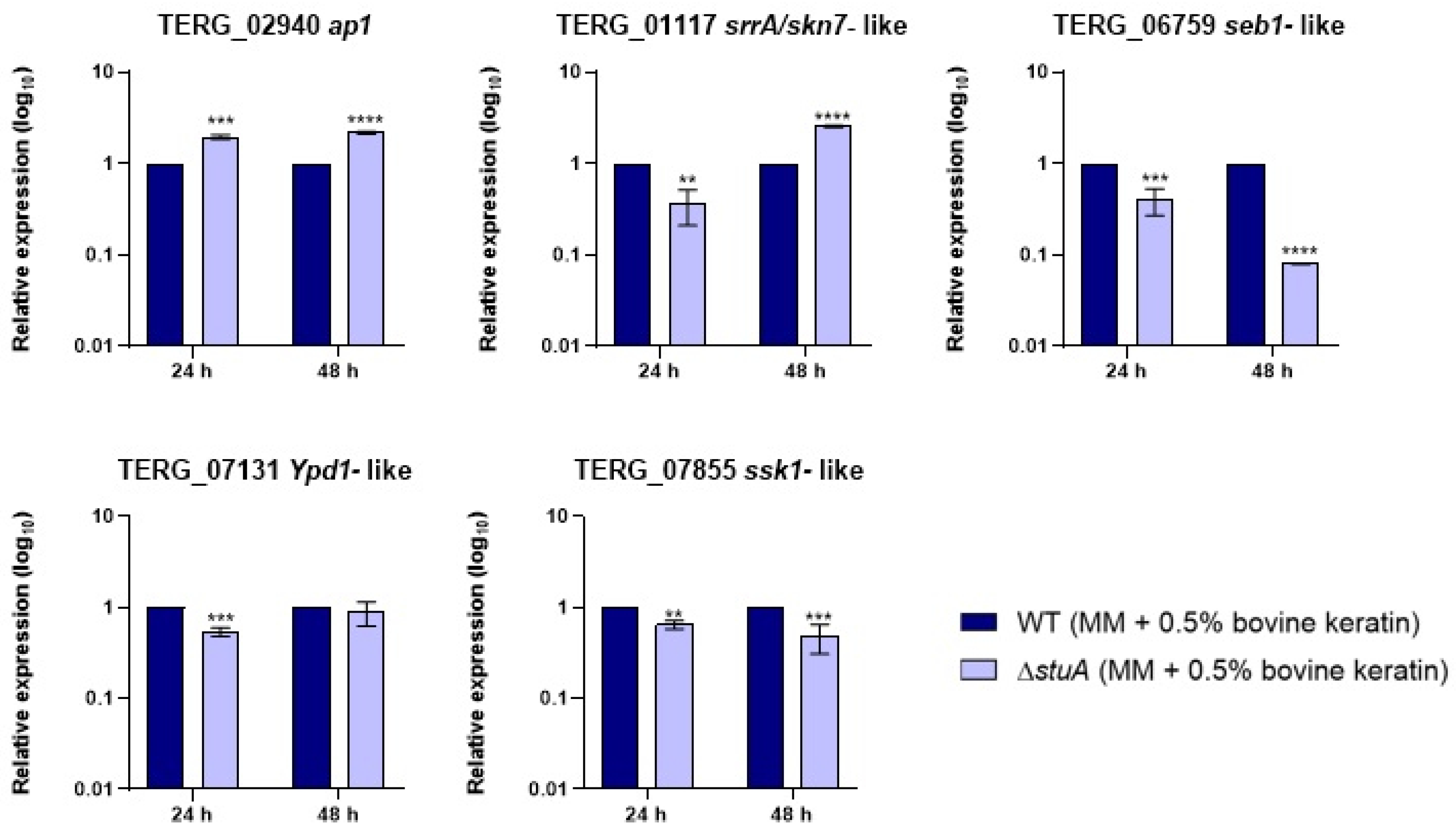
Figure 2.
Relative expression analysis of genes involved in oxidative stress pathways during fungal exposure to oxidative stress induction with hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM of hydrogen peroxide (H2O2)) for 1 and 3 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. * p < 0.05, ** p < 0.01, and *** p < 0.001.
Figure 2.
Relative expression analysis of genes involved in oxidative stress pathways during fungal exposure to oxidative stress induction with hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM of hydrogen peroxide (H2O2)) for 1 and 3 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. * p < 0.05, ** p < 0.01, and *** p < 0.001.
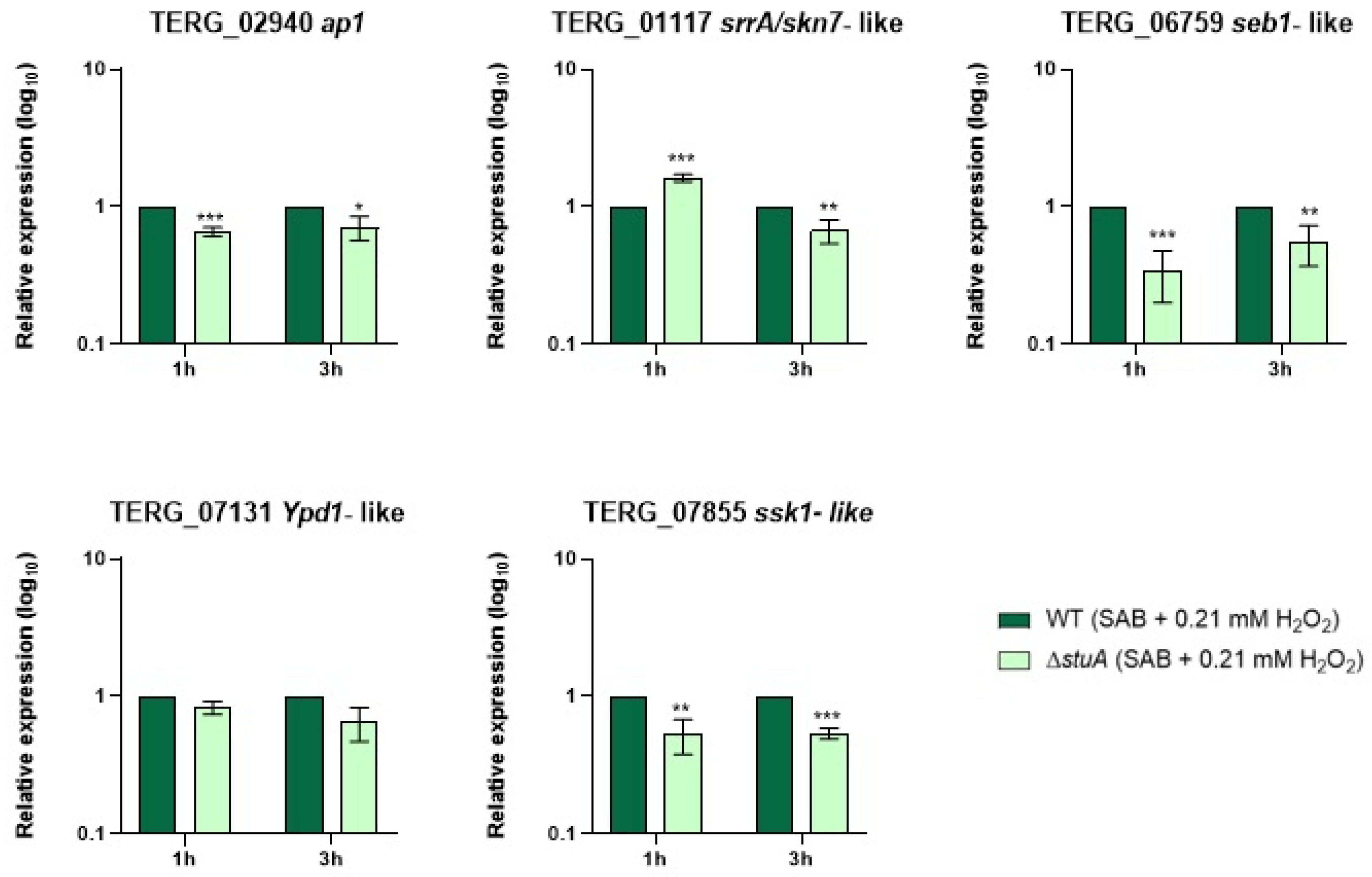
Figure 3.
Relative expression analysis of genes encoding the glutathione family and catalases during fungal growth in keratin (minimal medium (MM) containing 0.5% bovine keratin) for 24 and 48 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. *** p < 0.001, and **** p < 0.0001.
Figure 3.
Relative expression analysis of genes encoding the glutathione family and catalases during fungal growth in keratin (minimal medium (MM) containing 0.5% bovine keratin) for 24 and 48 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. *** p < 0.001, and **** p < 0.0001.
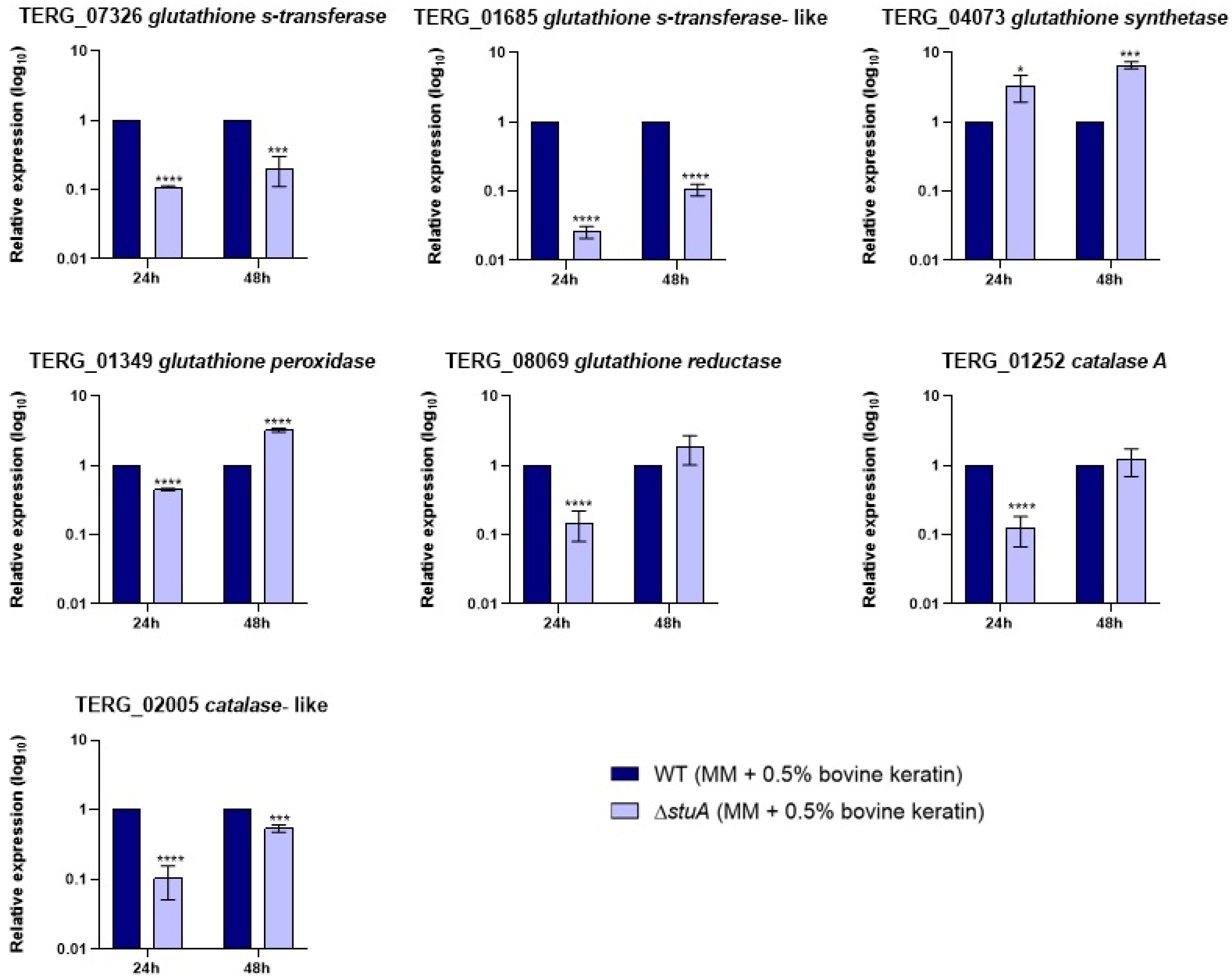
Figure 4.
Relative expression analysis of genes encoding the glutathione family and catalases during fungal exposure to oxidative stress induction with hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) for 1 and 3 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
Figure 4.
Relative expression analysis of genes encoding the glutathione family and catalases during fungal exposure to oxidative stress induction with hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) for 1 and 3 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
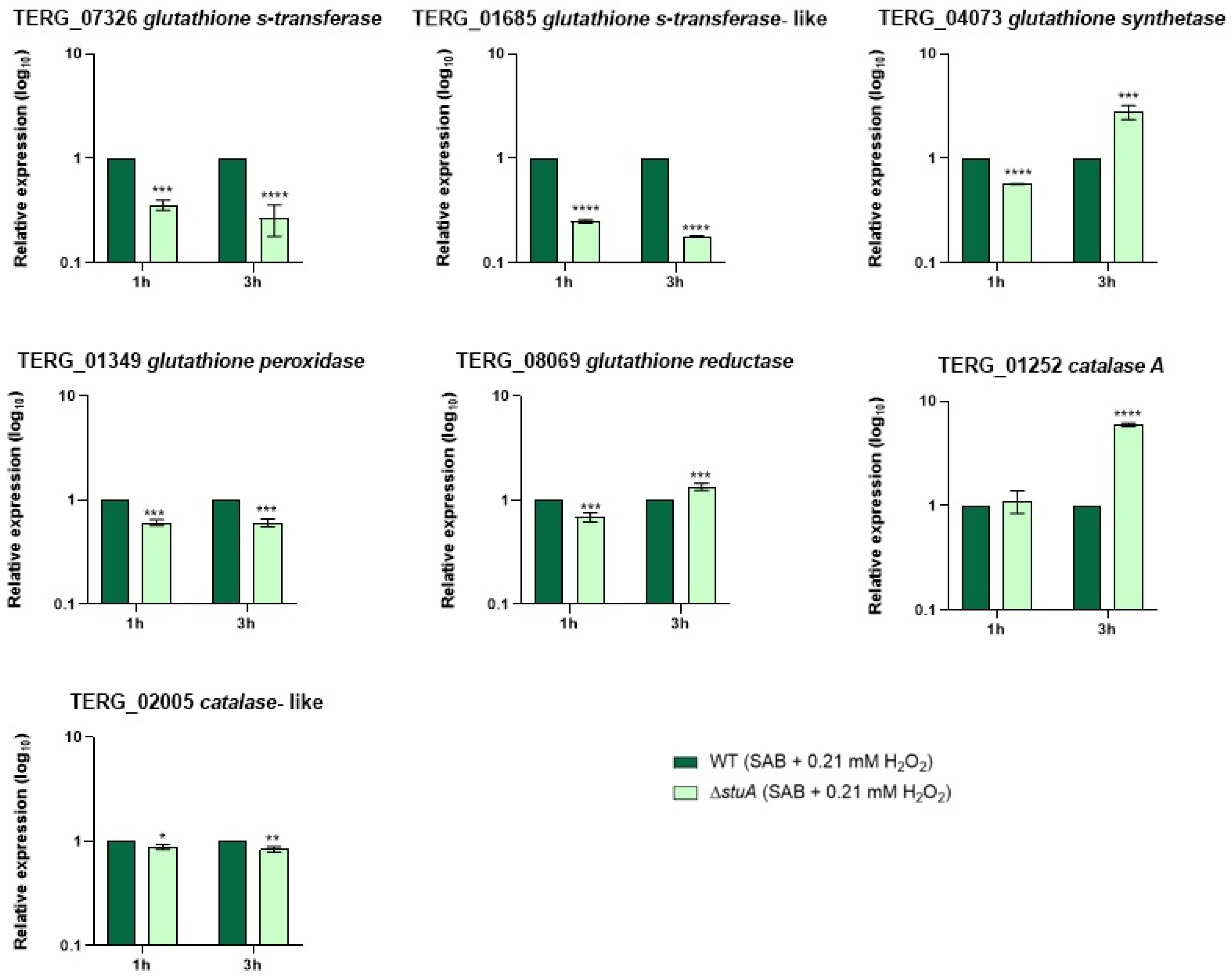
Figure 5.
Consensus sequence position for StuA in the promoter region of the gene TERG_07326. The consensus-binding sites were determined by identifying DNA sequences upstream (1,000 bp) that matched the StuA 5’-[A/T]CGCG[T/A]N[A/C]-3’ DNA-binding consensus. (A) Representation of the disposition of the cis-regulatory elements found in the sense strand (blue box). (B) StuA-interaction consensus sequence flanked by 35 adjacent base pairs. Yellow boxes depict the StuA consensus sequence found in the gene promoter.
Figure 5.
Consensus sequence position for StuA in the promoter region of the gene TERG_07326. The consensus-binding sites were determined by identifying DNA sequences upstream (1,000 bp) that matched the StuA 5’-[A/T]CGCG[T/A]N[A/C]-3’ DNA-binding consensus. (A) Representation of the disposition of the cis-regulatory elements found in the sense strand (blue box). (B) StuA-interaction consensus sequence flanked by 35 adjacent base pairs. Yellow boxes depict the StuA consensus sequence found in the gene promoter.

Figure 6.
Intracellular catalase enzymatic activity in the ΔstuA mutant and WT strains. Catalase activity was measured in protein extracts from fungal growth in keratin (minimal medium (MM) containing 0.5% bovine keratin) for 24 h (A) and 48 h (B), and from fungal exposure to hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) for 3 h (C). The activity was assessed at 10- and 15-minute intervals. The statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. *** p < 0.001, and **** p < 0.0001.
Figure 6.
Intracellular catalase enzymatic activity in the ΔstuA mutant and WT strains. Catalase activity was measured in protein extracts from fungal growth in keratin (minimal medium (MM) containing 0.5% bovine keratin) for 24 h (A) and 48 h (B), and from fungal exposure to hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) for 3 h (C). The activity was assessed at 10- and 15-minute intervals. The statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. *** p < 0.001, and **** p < 0.0001.
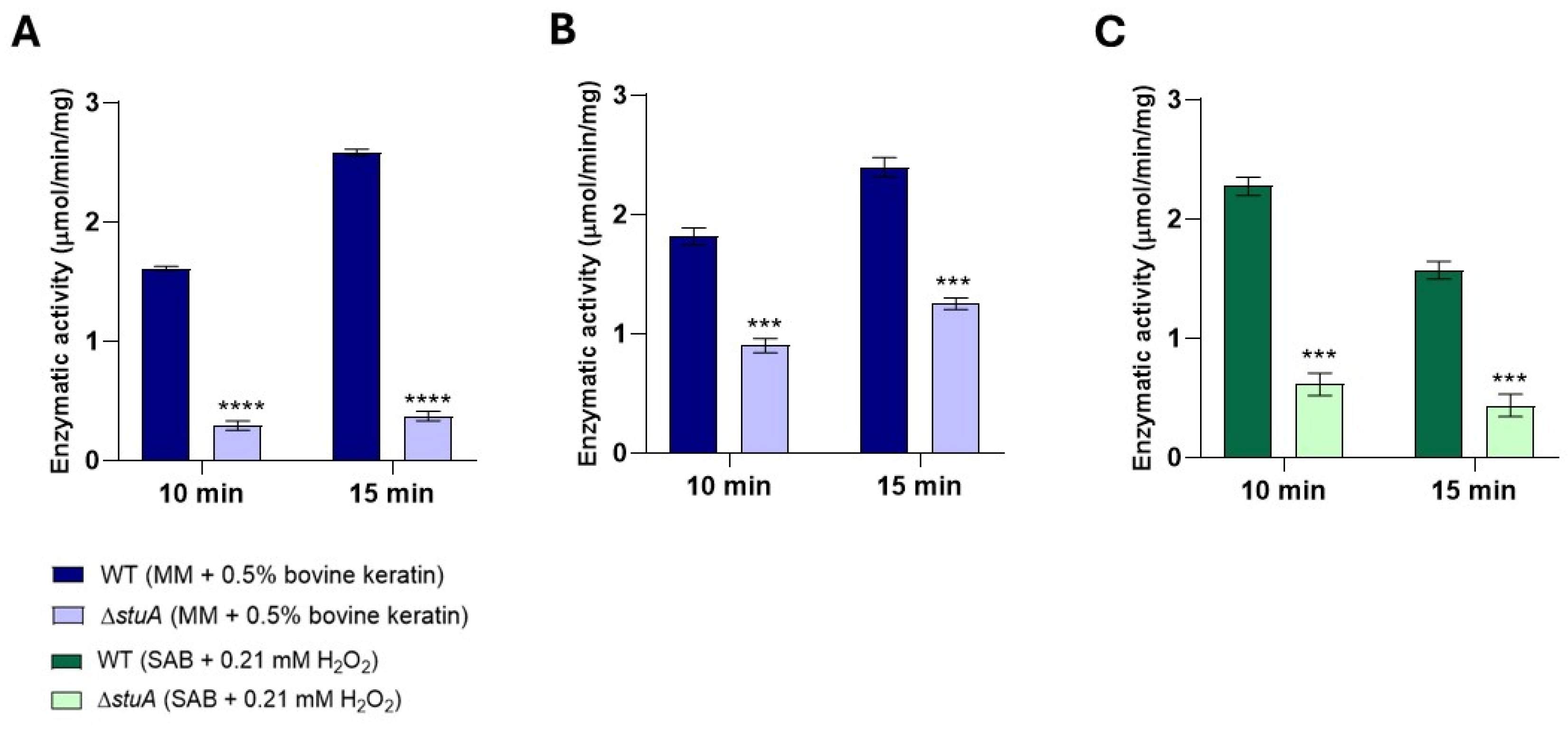
Figure 7.
Intracellular glutathione S-transferase activity was measured in the ΔstuA mutant and WT strains. The activity was assessed in protein extracts from fungal cultivation in keratin (minimal medium (MM) containing 0.5% bovine keratin) at 24 and 48 h (A) and from fungal exposure to hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) at 1 and 3 h (B). The enzyme activity was measured at 5-minute intervals. The statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. ** p < 0.01.
Figure 7.
Intracellular glutathione S-transferase activity was measured in the ΔstuA mutant and WT strains. The activity was assessed in protein extracts from fungal cultivation in keratin (minimal medium (MM) containing 0.5% bovine keratin) at 24 and 48 h (A) and from fungal exposure to hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) at 1 and 3 h (B). The enzyme activity was measured at 5-minute intervals. The statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. ** p < 0.01.
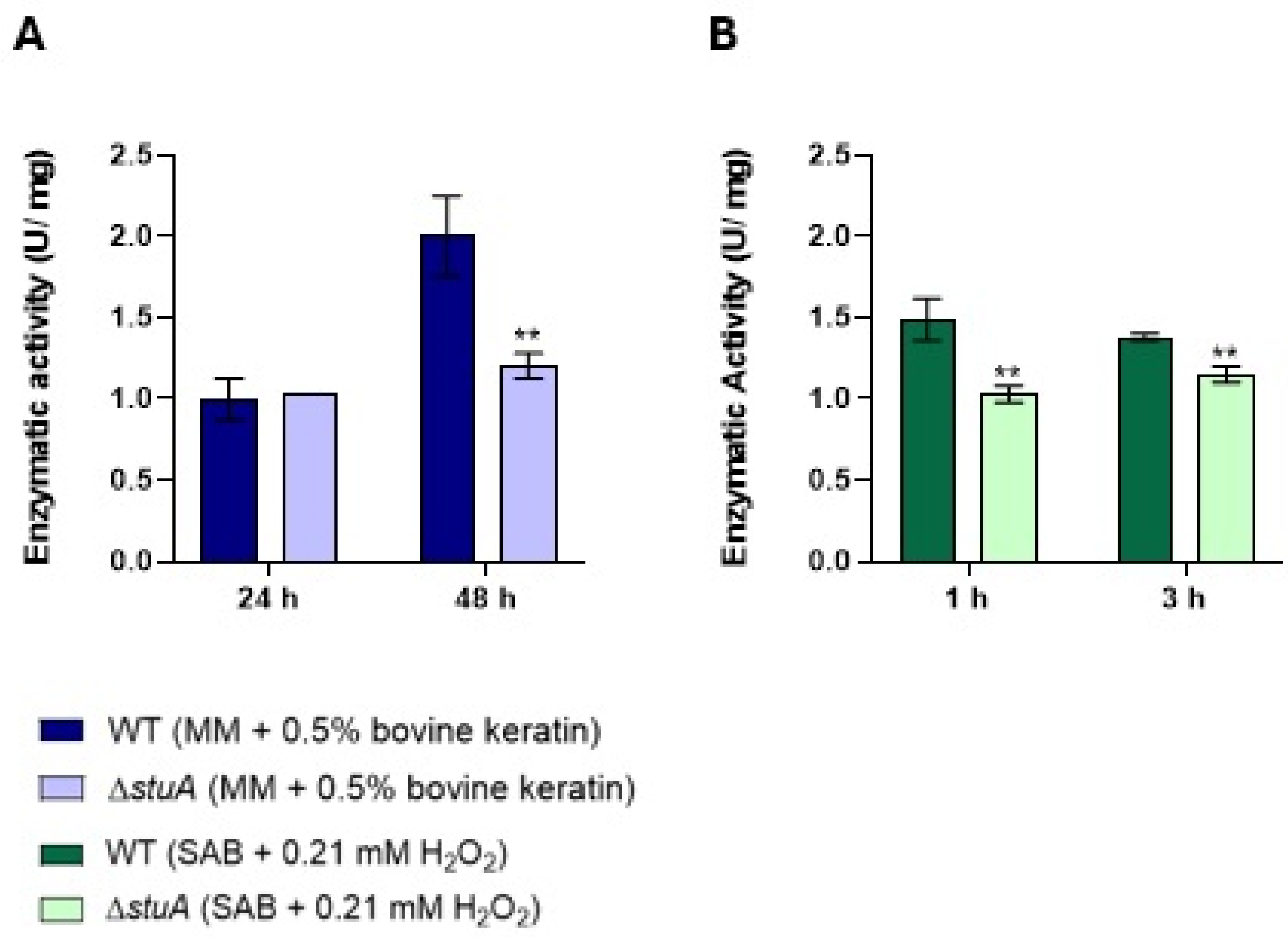
Figure 8.
Relative expression analysis of genes encoding siderochrome iron transporters (TERG_03174 and TERG_04949) and nonribosomal siderophore peptide synthase (TERG_00697) during fungal growth in keratin (minimal medium (MM) with 0.5% bovine keratin) for 24 and 48 h and during exposure to oxidative stress induced by hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) for 1 and 3 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
Figure 8.
Relative expression analysis of genes encoding siderochrome iron transporters (TERG_03174 and TERG_04949) and nonribosomal siderophore peptide synthase (TERG_00697) during fungal growth in keratin (minimal medium (MM) with 0.5% bovine keratin) for 24 and 48 h and during exposure to oxidative stress induced by hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) for 1 and 3 h. The WT strain was used as the normalizing condition. Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
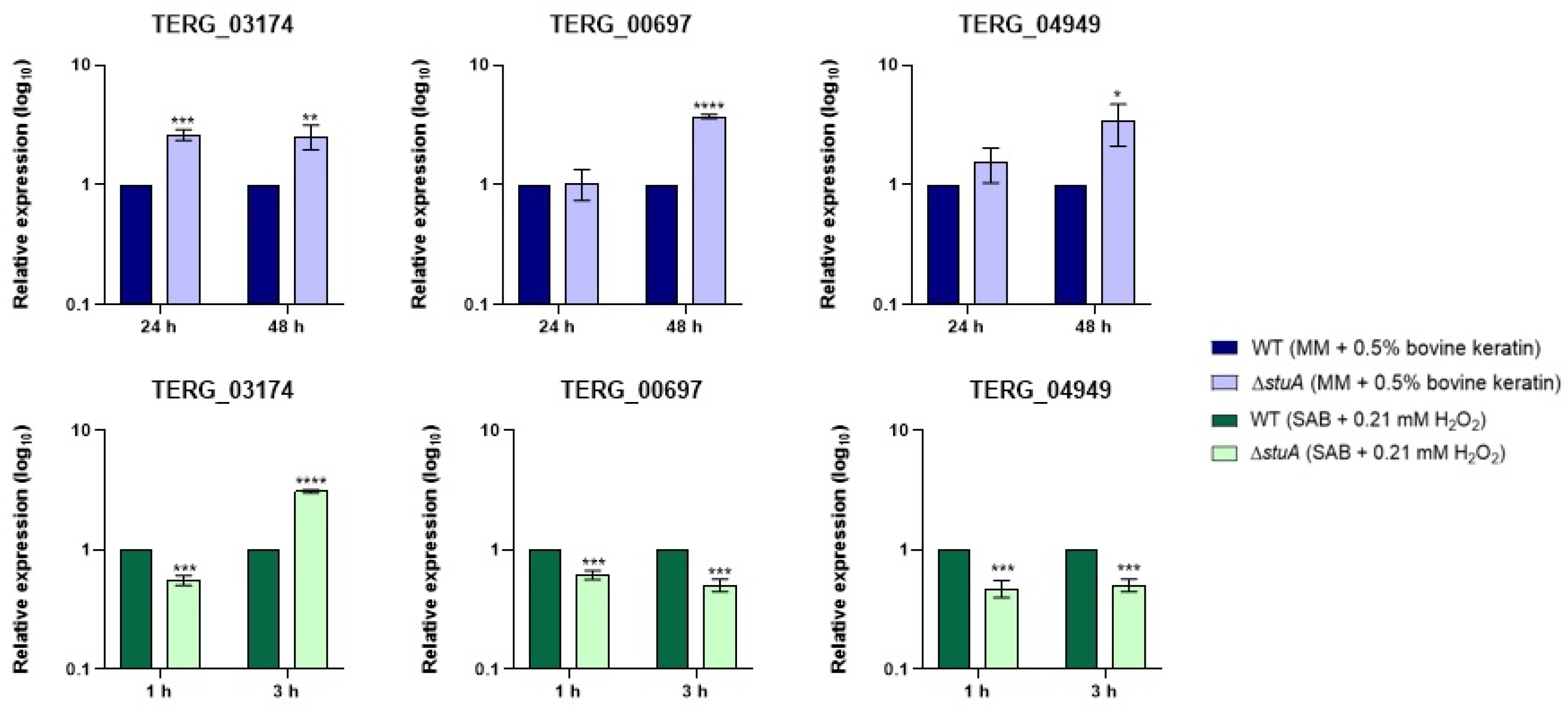
Figure 9.
Intracellular iron levels in the ΔstuA mutant and WT strains. Intracellular iron levels were measured in protein extracts from fungal growth in keratin (minimal medium (MM) containing 0.5% bovine keratin) for 24 and 48 h (A) and from fungal exposure to hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) for 1 and 3 h (B). Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. * p < 0.05, ** p < 0.01, and *** p < 0.001.
Figure 9.
Intracellular iron levels in the ΔstuA mutant and WT strains. Intracellular iron levels were measured in protein extracts from fungal growth in keratin (minimal medium (MM) containing 0.5% bovine keratin) for 24 and 48 h (A) and from fungal exposure to hydrogen peroxide (Sabouraud medium (SB) with 0.21 mM hydrogen peroxide (H2O2)) for 1 and 3 h (B). Statistical significance was determined using an unpaired Student’s t-test with Holm-Sidak correction for multiple comparisons. * p < 0.05, ** p < 0.01, and *** p < 0.001.
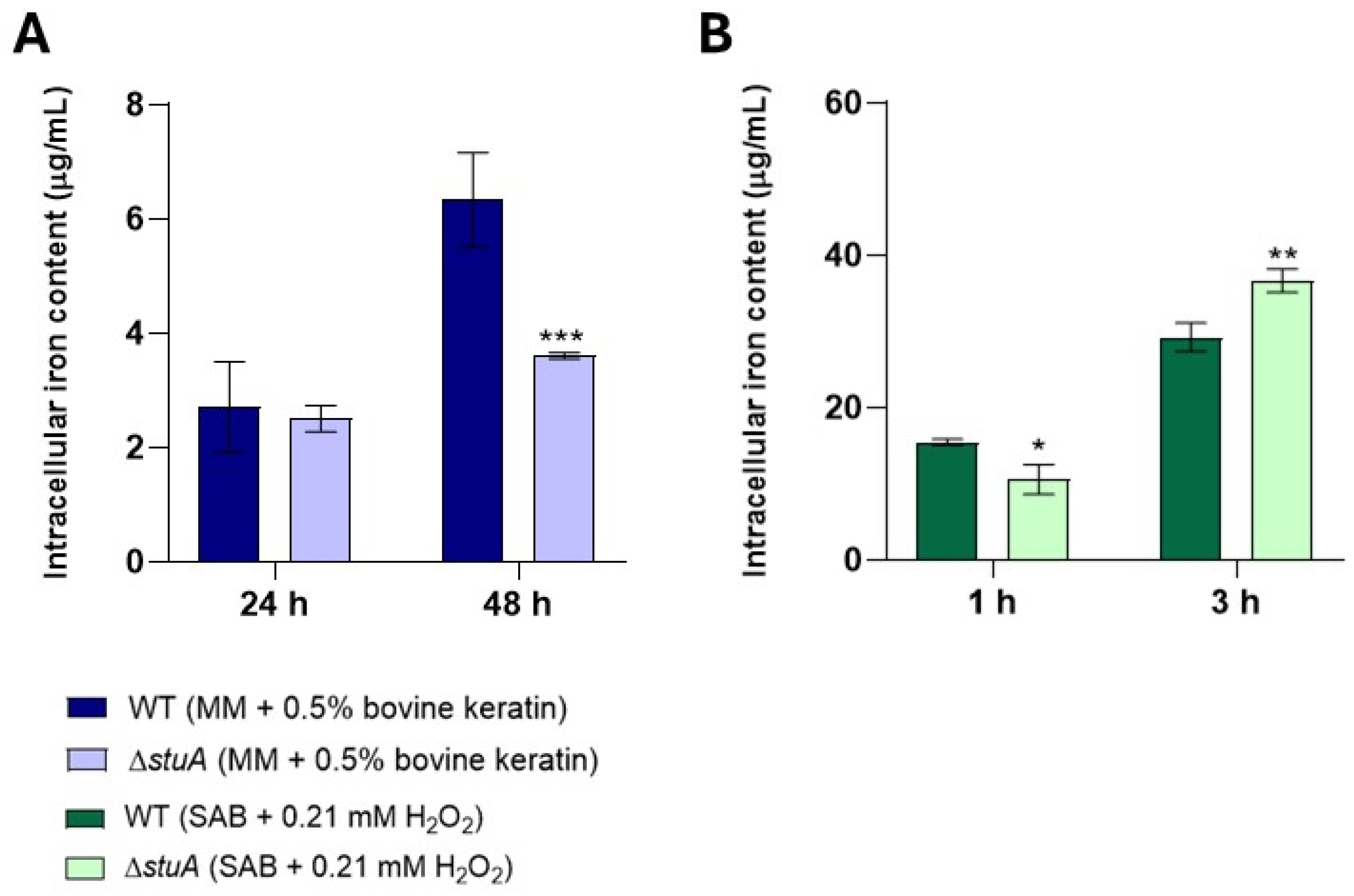

Table 1.
Oxidative stress-related genes with homologs in various Trichophyton species.
| Gene ID | Gene Product Name | Homologous |
|---|---|---|
| TERG_01117 | Stress response transcription factor SrrA/Skn7 | ARB_07479 (Trichophyton benhamiae) |
| TERG_06759 | C2H2 transcription factor (Seb1) | ARB_00366 (Trichophyton benhamiae) |
| TERG_07131 | Phosphotransmitter protein Ypd1 | TEQG_03843 (Trichophyton equinum) |
| TERG_07855 | Response regulator ssk1 | TRV_02413 (Trichophyton verrucosum) |
| TERG_01685 | Glutathione S-transferase | ARB_02229 (Trichophyton benhamiae) |
| TERG_02005 | Catalase | TESG_07337 (Trichophyton tonsurans) |
TERG (T. rubrum gene identification), ARB (T. benhamiae gene identification), TEQG (T. equinum gene identification), TRV (T. verrucosum gene identification), TESG (T. tonsurans gene identification).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



