
Submitted:
28 October 2024
Posted:
30 October 2024
You are already at the latest version
Abstract
The present study aimed to evaluate the potential synergy between pharmaceutical formulations containing Bixa orellana L. (granulated – CHR OR and injectable nanodispersion – CHR IN) in conjunction with a cannabidiol (CBD)-rich extract of Cannabis sativa L. (CSE) on experimental pain models in Wistar rats, including acetic acid-induced writhing, formalin, hot plate, and cold-water tail withdrawal tests. Chemical analyses by chromatography confirmed that the oil from B. orellana is rich in δ-tocotrienol (72.0 ± 1.0%), while the oil from Cannabis sativa highlighted the presence of cannabidiol (CBD). The results from the experimental pain tests indicated that the combined administration of formulations containing Bixa orellana and C. sativa, such as the granulated CHR OR (400 mg/kg, orally) with CSE (40 mg/kg, orally) or the nanodispersion CHR In (10 mg/kg, intramuscularly) with CSE (40 mg/kg, orally), demonstrated significant results (p < 0.001) in pain reduction. Although the formulations containing Bixa orellana extract showed statistical significance in the tests when used in isolation, their effects were inferior compared to the combined use with CSE or the isolated use of CSE. These findings suggest that combining formulations containing extracts of these plant species may represent a viable therapeutic option, considering the synergistic action in reducing pain under the experimental conditions employed. These results imply that combining the phytocomplexes present in B. orellana and C. sativa may be a promising approach for pain treatment.
Keywords:
Cannabis sativa
; Bixa orellana
; Nanodispersion
; Granules
; Analgesia
1. Introduction
Pain is a distressing experience associated with actual or potential tissue damage, with sensory, emotional, cognitive, and social components (Williams; Craig, 2016). Although pain generally plays an adaptive role, it can have adverse effects on function, social well-being, and psychological health, especially in the context of chronic pain, which affects approximately 20% of the population [1] (Raja et al., 2020; Urits et al., 2020).
Pain treatments can take diverse pharmacological approaches depending on the type of pain experienced by the patient. For nociceptive pain, nonsteroidal anti-inflammatory drugs (NSAIDs), non-opioid analgesics, and opioids can be used. For neuropathic pain or central sensitization, tricyclic antidepressants and serotonin-norepinephrine reuptake inhibitors (SNRIs) are recommended options (Urits et al., 2020). In response to the opioid epidemic and the risks associated with prescribing opioids for chronic pain management, alternative therapies have been introduced as viable treatment options, and the study of bioactive plant-derived compounds has garnered increasing interest (Hill et al., 2017).
Bixa orellana L., commonly known as "annatto" or "achiote," is a small perennial tree native to the tropical rainforests of Central and South America. This species holds significant cultural importance in India, China, the Philippines, Brazil, and Guyana (Batista et al., 2022). Additionally, the plant exhibits a wide range of medicinal properties. It is widely used to treat renal discomfort, diabetes mellitus, constipation, and hypertension, as well as serving as an antibiotic, laxative, cardiotonic, anti-inflammatory, antipyretic, and analgesic (Aktary et al., 2019; Shilpi et al., 2006). These benefits are partly attributed to the presence of tocotrienols and geranylgeraniol in its composition (Batista et al., 2022).
The literature provides preclinical and clinical evidence supporting the analgesic effects attributed to tocotrienols, likely due to their anti-inflammatory properties (Tejpal Singh et al., 2023). Specifically, δ-tocotrienol, identified as the predominant isoform in Bixa orellana oil, has demonstrated significant efficacy in reducing inflammation and protecting cells from oxidative damage, as evidenced by studies (Aggarwal et al., 2019; Ranasinghe et al., 2022).
Cannabis sativa L. is an herbaceous plant belonging to the Cannabaceae family, characterized by its growth in tropical and subtropical regions (de Oliveira Carvalho et al., 2021). The extensive medicinal history of C. sativa dates back to ancient Chinese texts from 2900 BC, highlighting its ancestral use for pain relief (Hill et al., 2017). Although Cannabis sativa contains a variety of cannabinoids, ∆9-tetrahydrocannabinol (∆9-THC) and cannabidiol (CBD) are the predominant components in terms of concentration (Mlost et al., 2020).
CBD, a prominent non-psychoactive component of Cannabis sativa, has been the subject of extensive research due to its analgesic and anti-inflammatory properties. CBD demonstrates a limited affinity for the CB1 and CB2 receptors of the endocannabinoid system. Studies indicate that CBD may influence non-cannabinoid pathways, including serotonin 5-HT1A receptors, TRPV1, and GPR55, which play crucial roles in pain regulation (Neelakantan et al., 2015; Urits et al., 2020)
The present study aimed to evaluate the potential synergy between pharmaceutical formulations containing Bixa orellana L. (granules - Chronic® and injectable nanosuspension - Chronic In®) combined with CBD-rich Cannabis sativa L. extract in experimental pain models using Wistar rats.
2. Results and Discussion
2.1. Chemical Analysis of Bixa orellana and Cannabis sativa Oils
As analyzed by gas chromatography, the chromatogram (Figure 1) allowed for the integration of the peak corresponding to δ-tocotrienol (C27H40O2), with a retention time of 28.6 minutes. The average concentration of δ-tocotrienol was found to be 0.725 ± 0.062 mg/mL (72.0 ± 1.0%), calculated based on the equation of the line obtained from δ-tocotrienol at various concentrations. The coefficient of linearity (R²) was equal to 0.9973, with the regression equation given by y = 8.30 × 106x - 2.818 × 106.
The chemical results obtained for Bixa orellana oil are consistent with the literature, which reports δ-tocotrienol values ranging from 53% to 93% as the major component (Sales et al., 2023; Yong et al., 2014)
In the chemical analysis of Cannabis sativa oil via gas chromatography (Figure 1), the chromatographic peak of cannabidiol was identified at 30.54 minutes. The fragmentation profile was confirmed using the chromatograph's NIST library. Other compounds were identified, including methyl esters of medium-chain fatty acids used to solubilize the Extract, with the methyl ester of capric acid showing a retention time of 13.75 minutes.
Cannabidiol (CBD) has emerged as a promising substance due to its documented analgesic potential in various studies. This non-psychoactive compound, found in the Cannabis sativa plant, has been extensively investigated for its therapeutic properties, particularly in pain management (Hill et al., 2017).
2.2. Characterization of Injectable Nanodispersion (Chronic in®) of B. orellana
The nanoprecipitation method, adapted from Oliveira Carvalho et al. (2022), was employed to prepare the nanodispersion containing Bixa orellana oil. No visual alterations indicating loss of stability were observed throughout the evaluation period. The formulation did not exhibit phase separation or precipitation of components, demonstrating a visibly stable appearance with a slightly yellow color and a pH of 5.8 ± 0.01.
In this study, the nanoparticle size remained stable, with an average size of 53.15 ± 0.64 nm and a PDI of 0.574 ± 0.032 on the first day. On day 30, the particle size was 59.90 ± 3.63 nm with a PDI of 0.574 ± 0.032. The nanoformulation exhibited a Zeta potential of 19.86 ± 0.60 mV on day zero and 19.66 ± 1.45 mV on day 30 (Table 1).
The studies by Oliveira Carvalho et al. (2021) and Mitchell et al. (2021) have indicated that nanoformulations with particle sizes ranging from 50 to 200 nm significantly maintain stability, aligning with our findings. According to Maia Filho et al. (2023), the reduction of nanoparticle size is crucial as it directly impacts the properties and stability of nanoemulsions. Smaller particles possess a larger surface area, leading to better dispersion and reduced influence of repulsive forces, thereby preventing adjacent particles' aggregation. Additionally, Zeta potential, as highlighted by Zielinska et al. (2020), is a crucial parameter for evaluating the physicochemical stability of nanoemulsions, with formulations above ±15 mV considered ideal.
2.3. Evaluation of Treatments in Pain Models
This study investigated the efficacy of subchronic treatment using a combination of Cannabis sativa extract with a high cannabidiol content and the nutraceuticals Chronic® and Chronic In®, developed from the phytocomplex oil extract of Bixa orellana L., to alleviate pain in animals. We assessed the analgesic activity using four distinct methods: the acetic acid-induced writhing test, the formalin test, the hot plate test, and the cold-water tail withdrawal test.
The acetic acid-induced writhing technique is commonly employed to assess the efficacy of peripheral analgesic agents or visceral inflammatory pain management (Bunman et al., 2022). Acetic acid triggers an inflammatory response in the abdominal cavity, activating the nociceptor (Foss et al., 2021a). This model of visceral pain, induced by acetic acid nociception, is non-specific and peripheral, susceptible to modulation by various therapeutic agents, including anti-inflammatories, opioid analgesics, and other centrally acting compounds (Foss et al., 2021a; Silva et al., 2021).
The pain-inducing action of acetic acid arises from its chemical stimulation in the peritoneal cavity, leading to the release of inflammatory mediators such as interleukins IL-1, IL-6, IL-8, tumor necrosis factor α (TNF-α), as well as chemokines, histamine, serotonin, and bradykinin. These substances promote the increased synthesis of lipoxygenase (LOX) and cyclooxygenase (COX) enzymes, resulting in the production of leukotrienes and prostaglandins, notably prostaglandin E2 (PGE2) and F2α (PGF2α) (Menezes et al., 2021; Silva et al., 2021). Additionally, pro-inflammatory cytokines such as IL-8, IL-1β, and TNF-α, released by macrophages and basophils, amplify the release of aspartate, glutamate, and other endogenous mediators that activate visceral nociceptive neurons (Batista et al., 2016).
Several recent studies have investigated the therapeutic potential of cannabidiol (CBD) in managing acetic acid-induced abdominal pain, consistently demonstrating its ability to significantly reduce abdominal writhing compared to placebo groups (Bunman et al., 2022; Foss et al., 2021a; Neelakantan et al., 2015; Silva et al., 2021). Neelakantan et al. (2015) and Foss et al. (2021a) report that CBD decreases the production of various inflammatory markers, such as TNF-α, IL-6, and COX-2, through mechanisms that do not involve CB1 or CB2 cannabinoid receptors. Thus, the antinociceptive effects of CBD observed in the acetic acid-induced writhing test may be attributed to its potent anti-inflammatory properties.
Furthermore, earlier studies have corroborated the therapeutic efficacy of Bixa orellana in this same test. Aktary et al. (2019) and Shilpi et al. (2006) suggest that the antinociceptive activity observed in Bixa orellana extract may result from its ability to interfere with the synthesis and release of endogenous substances induced by acetic acid, such as prostacyclin (PGI2) and other prostanoids. Additionally, the desensitization of sensory C fibers may be involved in pain transmission.
Regarding the number of abdominal writhing induced by acetic acid (Figure 2), a statistically significant reduction was observed in all experimental groups, with a p-value < 0.001. Notably, the groups subjected to the combined administration of CSE (40 mg/kg, oral) + CHR OR (400 mg/kg, oral), as well as those receiving this combination together with naloxone (2 mg/kg, intraperitoneal), showed results comparable to the standard drug (morphine).
Considering that the acetic acid-induced abdominal writhing test is a non-specific model and does not allow for determining the exact nociceptive pathways through which a drug may act, other procedures were employed to elucidate possible mechanisms of antinociception (Silva et al., 2021). The formalin test stands out from conventional pain models as it evaluates an animal's response to continuous and moderate pain from tissue injury. It is considered more representative of clinical pain than tests that use short-duration mechanical or thermal stimuli (Tj et al., 1992).
Two distinct phases characterize the nociceptive response to formalin. The first phase, known as the neurogenic phase, occurs immediately after formalin injection and lasts approximately 10 minutes. It is marked by releasing substances P, glutamate, bradykinin, and nitric oxide, which stimulate C-type nerve fibers and some Aδ fibers. The second phase, known as the inflammatory phase, begins 15-20 minutes after injection and can last for over 60 minutes, with the release of inflammatory mediators such as histamine, serotonin, bradykinin, and prostaglandins (Bannon & Malmberg, 2007; Bunman et al., 2022; Menezes et al., 2021). Central-acting analgesics, such as opioids, inhibit both phases of the response, while peripherally-acting anti-inflammatory drugs, such as nonsteroidal anti-inflammatory drugs (NSAIDs) and steroids (SAIDs), inhibit only the second phase (Silva et al., 2021).
Figure 3 graphically presents the results obtained in this study for both test phases regarding the treatments with the orally administered nutraceutical Chronic®, either alone or in combination with Cannabis sativa extract rich in cannabidiol (CSE). In both the early and late phases, all treated groups showed significant differences from the control group, which received distilled water (4 ml/kg), with p-values < 0.001.
Regarding the results of the formalin test in the groups that received Chronic In®, administered intramuscularly, all groups showed significant differences compared to the control group (p < 0.001) in both the early and late phases (Figure 4). However, significant differences were observed between the experimental groups CSE + CHR IN and CHR IN alone in the early phase (p < 0.05) and the late phase (p < 0.01), with the Group receiving the combination of treatments yielding the best results. This suggests that the combination of Cannabis sativa extract rich in cannabidiol may enhance the analgesic effects of Bixa orellana.
Additionally, in the late (inflammatory) phase, the combination of naloxone with CSE and CHR IN had a significantly negative impact (p < 0.05), increasing the nociceptive behavior of the animals. This effect was not reported when naloxone was combined with CSE and CHR OR, suggesting that the relevance of this interaction in the opioid pathway is still unclear. Therefore, further studies are necessary to elucidate this issue.
The findings of this study align with previous research highlighting the efficacy of CBD (Bunman et al., 2022; Razavi et al., 2021; Silva et al., 2021) and Bixa orellana (Aktary et al., 2019) in this pain model. It is worth noting that CBD demonstrated a more pronounced influence in the inflammatory phase than in the neurogenic phase in all these studies.
Several studies corroborate the anti-inflammatory effects of CBD. Costa et al. (2004) reported that in a carrageenan-induced paw edema experiment, CBD significantly reduced tissue cyclooxygenase activity, plasma levels of prostaglandin E2, and the production of oxygen free radicals and nitric oxide. Atalay et al. (2020) affirmed that CBD acts as an agonist of the PPARγ receptor, modulating inflammation by suppressing the expression of pro-inflammatory genes, including cyclooxygenase (COX2), and various pro-inflammatory mediators such as TNF-α, IL-1β, and IL-6. Additionally, Urits et al. (2020) documented that, in combination with THC, even low concentrations of CBD can interact with CB2 receptors, inhibiting the inflammatory response, especially by suppressing mast cell degranulation and neutrophil migration to areas near pain centers. Therefore, the enhanced efficacy of CBD during the late phase of the formalin test could be attributed to its anti-inflammatory effects.
As for the action of Bixa orellana extract in the formalin test, its analgesic properties can also be associated with its anti-inflammatory characteristics (Aktary et al., 2019). As mentioned earlier, tocotrienols exhibit significant anti-inflammatory and antioxidant activity, reducing the secretion of critical pro-inflammatory mediators involved in pain sensitization and providing protection against tissue injury (Aggarwal et al., 2019; Ranasinghe et al., 2022; Tejpal Singh et al., 2023). Geranylgeraniol is another compound found in B. orellana, which demonstrates anti-inflammatory properties. Batista et al. (2022) indicate that geranylgeraniol inhibits the expression of interleukin-1 receptor-associated kinase-1 (IRAK1) and tumor necrosis factor receptor-associated factor 6 (TRAF6), preventing excessive NF-κB activation in response to lipopolysaccharide-induced inflammation in THP-1 cells.
The hot plate and tail withdrawal tests are widely recognized as effective for evaluating central antinociceptive activity (Aktary et al., 2019). In this assay, the nociceptive experience is mediated by the activation of vanilloid receptors and the stimulation of C-type and Aδ-type nociceptive fibers (Menezes et al., 2021). Preliminary studies suggest that centrally acting drugs, such as NMDA receptor antagonists, opioids, and tricyclic antidepressants, show significant efficacy in this model. At the same time, anti-inflammatory agents like indomethacin, dexamethasone, and nimesulide do not exhibit relevant efficacy (Bannon & Malmberg, 2007).
In the literature, there is some disagreement regarding the expected outcomes when assessing CBD in the hot plate test. While some studies report modest or no significant analgesic activity in this assay (Foss et al., 2021b; Neelakantan et al., 2015; Sofia et al., 1975), casting doubt on the possibility of central analgesic action by CBD, other studies have found positive results for both CBD (Crivelaro do Nascimento et al., 2020) and Cannabis sativa roots (Menezes et al., 2021).
Regarding the analgesic effects of CBD in this test, Nascimento et al. (2020) demonstrated that the selective CB1 antagonist AM251 can block CBD's effect in the hot plate test. In contrast, the CB2 inverse agonist SCH 336 does not influence the antinociceptive effects of CBD, suggesting that the analgesic action of CBD in this test is mediated through the CB1 receptor rather than the CB2 receptor. Moreover, the antagonism of TRPV1 receptors amplified the antinociceptive effect of CBD. These findings indicate that CBD reduces nociception through FAAH inhibition, leading to increased levels of anandamide (AEA). Consequently, AEA may bind to both CB1 and TRPV1 receptors, which have opposing effects on pain modulation.
As for the antinociceptive effects of Bixa orellana, there is a notable gap in the literature, with few studies available. Aktary et al. (2019) demonstrated that the methanolic Extract of B. orellana showed a latency time comparable to the control group in the hot plate test, suggesting a potential central analgesic action by this plant.
In our study, as illustrated in Figure 5, both the Group treated with CSE alone and the groups that received CSE in combination with Chronic® or Chronic In® exhibited statistically significant results (p < 0.001). The isolated use of CHR OR and CHR IN, although significant (p < 0.05), revealed inferior results compared to the other experimental groups, supporting the theory that Bixa orellana has a central analgesic effect, albeit less potent than that of CBD. It is important to note that the groups treated with naloxone, an opioid antagonist, displayed altered results, suggesting a potential interaction of CSE within the opioid pathway in this pain model. Some studies have already suggested a possible action of CBD on μ and δ opioid receptors for pain regulation (Mlost et al., 2020b; Urits et al., 2020), emphasizing further research to clarify this interaction.
As previously mentioned, the tail withdrawal test is widely recognized for assessing central antinociceptive activity. Unlike the hot plate test, where the endpoint involves a complex behavior (licking of the hind paw), nociceptive stimulation by temperature on the tail produces a simple nociceptive reflex response mediated by the spinal cord (tail withdrawal) (Bannon & Malmberg, 2007). According to studies (Adams et al., 1994; Pizziketti et al., 1985), the cold-water tail withdrawal test is particularly suited for evaluating opioid-mediated antinociception. In this method, central nervous system depressants, such as antipsychotics or anxiolytics, as well as non-narcotic analgesics like aspirin and acetaminophen, are ineffective (Pizziketti et al., 1985)
In current literature, the results of the tail withdrawal test involving Cannabis sativa, particularly the phytocannabinoids CBD and THC, are conflicting. While specific studies fail to demonstrate therapeutic efficacy for CBD in this method, indicating that analgesic action is attributable solely to THC (Moore & Weerts, 2022; Sadaka et al., 2023), other studies suggest analgesic effects of CBD, as documented by Nascimento et al. (2020), who observed a significant reduction in thermal hyperalgesia in both acute and chronic CBD treatments. Regarding the action of Bixa orellana in this context, Aktary et al. (2019) identified significant neuropharmacological activity in a dose-dependent manner comparable to standard drugs.
Figure 6 presents the results of the cold-water tail immersion test, which measures the time required for tail withdrawal. The groups treated with CSE (40 mg/kg V.O.) + CHR OR (400 mg/kg V.O.) and CSE (40 mg/kg V.O.) + CHR IN (10 mg/kg V.O.), as well as their combinations with naloxone (2 mg/kg intraperitoneally), demonstrated a significant increase in response time. This increase is evidenced by p-values of < 0.001 compared to the control group and is comparable to both the positive control group (MOR) and the Group treated with CSE alone, with p < 0.001.
The Group that received CHR IN alone showed statistical significance (p < 0.05), although inferior to the groups that received the combinations. However, the Group that received CHR OR did not show significant differences compared to the control group. These results suggest that combining CSE with CHR OR or CHR IN enhances analgesic effects, while naloxone does not appear to interfere with these effects significantly.
3. Materials and Methods
3.1. Acquisition of Cannabis sativa Extract and Chemical Analysis by G.C.
The Cannabis sativa extract (Batch, 2007) was obtained from Zion Medpharma (São Paulo, SP, Brazil), with an extract concentration of 200 mg/mL, containing 50 mg/mL of CBD and < 2 mg/mL of THC. The gas chromatography (G.C.) analysis conditions were capillary column RTX-5 (30 m × 0.25 mm ID × 0.25 µm) and helium as the carrier gas, with a 14 mL/min flow rate. The oven temperature was programmed to remain at 60°C for 1 minute, then increase by 5°C/min up to 300°C. The injector temperature was set to 280°C, and the detector temperature to 250°C. A 1 µL injection was performed in split mode at a 10:1 ratio. The fragmentation spectra of the compounds were evaluated and identified using the NIST library of the equipment.
3.2. Acquisition of B. orellana Oil and Chemical Analysis
The B. orellana oil (Chronic®) was provided by Ages Bioactive Compounds, batch 0012/2022. For the chemical analysis, δ-tocotrienol quantification was performed. A stock solution of δ-tocotrienol in hexane (Sigma-Aldrich, MKCF5755, G.C. grade, Germany) was prepared at 10 mg/mL concentration. From this stock solution, additional solutions were prepared to construct a calibration curve with the following concentrations: 2.5 mg/mL, 1.0 mg/mL, 0.5 mg/mL, 0.25 mg/mL, and 0.1 mg/mL. The analyses were conducted using gas chromatography-mass spectrometry (GC-MS), following the previously described methodology (Rodrigues et al., 2022).
3.3. Obtention of the Granules of Bixa orellana (Chronic®)
The granules of Bixa orellana (Chronic®) were sourced from Ages Bioactive Compounds Co. (São Paulo, SP, Brazil). The extraction technique follows a standardized process protected by a patent.
3.4. Obtention of Polymeric Nanoparticles Containing Bixa orellana oil (Chronic In®)
The nanoprecipitation method, as adopted by de Oliveira Carvalho et al. (2022), was used to create a polymeric nanoparticle system containing Bixa orellana oil. A formulation of 1% purified B. orellana oil, 2% polyethylene glycol 4000 (PEG 4000), 2% Tween 80, 25% absolute ethanol, and 70% injectable-grade water was used to produce a final volume of 20 mL of nanosuspension (Chronic In®).
First, the organic phase was prepared by solubilizing B. orellana oil and PEG 4000 in absolute ethanol and stirring the mixture with a magnetic stirrer at 800 rpm for 20 minutes. Simultaneously, the aqueous phase was prepared by solubilizing Tween 80 in water and stirring for 20 minutes. The organic phase was poured into the aqueous phase under agitation using a mechanical stirrer (Fisatom, SP, Brazil) for 20 minutes. After preparation, the resulting nanosuspension was stored in injectable ampoules and subjected to characterization.
3.5. Characterization of B. orellana Nanosuspension (Chronic In®)
For stability purposes, the nanosuspension was characterized on the day of preparation and after 7 and 30 days. Visual appearance, phase separation, and sedimentation were macroscopically evaluated. Zeta potential, polydispersity index (PDI), and particle size were assessed in triplicate using a Zeta-Sizer (Malvern Pan-Analytics, UK), following the methodology described by (Borges et al., 2018)
3.6. Study Design and Ethical Considerations
This study was a randomized, controlled, non-clinical trial utilizing quantitative methods. The research was conducted at the Drug Research Laboratory located in the Health Sciences building of the Federal University of Amapá (UNIFAP), situated at Rodovia Juscelino Kubitscheck, Km-02, s/n, Zerão, Macapá, Amapá, Brazil.
As the study involved the use of animals, all procedures were conducted following the recommendations of the Universal Declaration of Animal Rights, the Brazilian College of Animal Experimentation (COBEA), and the resolutions of the Federal Council of Veterinary Medicine, as well as other applicable laws governing ethical practices for experimental research involving animals.
The study protocol was submitted to the Animal Research Ethics Committee (CEUA) of the Federal University of Amapá (UNIFAP) for review and was approved under protocol number 02/2022.
3.7. Animals Used
A total of 45 female Wistar rats were used in this study. The rats were obtained from the Multidisciplinary Center for Biological Research in Laboratory Animal Science (CEMIB) – University of Campinas (UNICAMP) at 21 days of age. The animals were acclimatized in groups of five per polyethylene cage, maintained under controlled temperature conditions (21 ± 1°C), with free access to food and potable water, a 12-hour light/dark cycle, and sawdust bedding changes, at the Drug Research Laboratory of the Federal University of Amapá.
3.8. Experimental Design
The present study utilized nine experimental groups comprising five Wistar strain Rattus norvegicus. For 28 days, seven of these groups were subjected to subchronic treatment administered daily.
The Negative Control Group (CON) received distilled water orally at 4 mL/kg. The Chronic® Group (CHR OR) was treated with Chronic® orally at 400 mg/kg. The Chronic In® Group (CHR IN) received Chronic In® via intramuscular injection (I.M.) at 10 mg/kg.
Other groups received combinations of treatments: the Cannabis sativa extract Group + Chronic® (CSE + CHR OR) was treated with CSE orally at a dosage of 40 mg/kg and Chronic® orally at a dosage of 400 mg/kg; the Cannabis sativa extract + Chronic In® Group (CSE + CHR IN) was treated with CSE orally at a dosage of 40 mg/kg and Chronic In® I.M. at a dosage of 10 mg/kg. The Cannabis sativa extract + Chronic® + Naloxone Group (CSE + CHR OR + NAL) was treated with CSE orally at a dosage of 40 mg/kg, Chronic® at a dosage of 400 mg/kg, and intraperitoneal Naloxone (I.P.) at a dosage of 2 mg/kg. Lastly, the Cannabis sativa extract + Chronic In® + Naloxone Group (CSE + CHR IN + NAL) received CSE orally at a dosage of 40 mg/kg, Chronic In® I.M. at a dosage of 10 mg/kg, and intraperitoneal Naloxone at a dosage of 2 mg/kg.
Two of the nine experimental groups received treatment only one hour before each experiment: the Positive Control Group (MOR), treated with morphine orally at a dosage of 10 mg/kg, and the Cannabis sativa extract Group (CSE), treated with CSE orally at a dosage of 40 mg/kg.
3.9. Formalin Test
This test was conducted according to the methodology proposed by Dubuisson and Dennis (1977). A total of 0.02 mL of a 1% formalin solution was administered intraplantarly in the right hind paws of the animals. The time each animal spent licking its paw was recorded in two phases: the first from 0 to 5 minutes and the second from 20 to 30 minutes. The groups were subjected to daily treatments for 28 days. The test was performed on the 28th day, 60 minutes after administering treatments. The MOR and CSE groups were exceptions, receiving treatment only on the day of the experiment and a 60-minute interval before the test.
3.10. Hot Plate Test
The animals were placed on a hot plate at a fixed temperature (55 ± 0.5 ºC). The latency to heat was assessed by measuring the time the animal took to withdraw its hind paw from the hot plate and lick it. The cut-off time was thirty seconds (İnaltekin; Kivrak, 2021). The groups were subjected to daily treatments for 28 days. The test was conducted on the 28th day, with measurements initiated 60 minutes after administering treatments. The MOR and CSE groups were exceptions, receiving treatment only on the day of the experiment and a 60-minute interval before the measurements.
3.11. Cold-Water Tail Withdrawal Test
Tail withdrawal tests in cold water were performed according to the method described by Sharma et al., 2008. The distal portion of each rat's tail was immersed in cold water (10 ± 1 ºC). Baseline latency for tail movement (tail withdrawal response) or signs of struggling were observed. The cut-off time was set at 50 seconds. The groups were subjected to daily treatments for 28 days. The test was conducted on the 28th day, 60 minutes after administering treatments. The MOR and CSE groups were exceptions, receiving treatment only on the day of the experiment and a 60-minute interval before the test.
3.12. Acetic Acid-Induced Abdominal Writhing
An intraperitoneal injection of 0.9% acetic acid (volume 0.1 mL/10 g) diluted in saline (0.9%) was administered. After the administration of acetic acid, the rats were placed in an acrylic box and observed for counting abdominal writhing from the fifth to the twentieth minute (adapted by Collier et al., 1968). The groups were subjected to daily treatments for 28 days. The test was conducted on the 28th day, 60 minutes after administering treatments. The MOR and CSE groups were exceptions, receiving treatment only on the day of the experiment and a 60-minute interval before the test.
3.13. Statistical Analysis
All experimental quantitative results were expressed as mean ± standard deviation for statistical analysis. Tukey's multiple comparisons test followed a one-way analysis of variance (ANOVA). Results with significance levels of p < 0.05 were considered statistically significant. The statistical software used for analysis was GraphPad Instat and Prism (version 5.03).
4. Conclusions
The simultaneous administration of formulations containing Bixa orellana and C. sativa, such as the granules CHR OR (400 mg/kg, orally) with CSE (40 mg/kg, orally) or the nanodispersion CHR IN (10 mg/kg, intramuscularly) with CSE (40 mg/kg, orally), yielded significant and promising results in reducing pain in Wistar rat pain models, suggesting a potential synergy between the formulations. While the Bixa orellana extract showed statistical significance when used alone, its effects were less pronounced than the combined use with CSE or the isolated use of CSE. These findings imply that combining formulations containing extracts of these plant species may offer a viable therapeutic option, as it enhances pain reduction under the experimental conditions.
5. Patents
The work is part of patent BR 10 2023 006029 3 A2 "Composition, the process for obtaining polymeric nanoparticles containing Bixa orellana oil standardized in total tocotrienols, the process of the polar microparticulate system or natural microcluster and its uses," deposited at the National Institute of Industrial Property - Brazil.
Author Contributions
A.M.S., H.O.C., D.E.S.G., L.P.G., N.N.D.C., A.V.T.L.T.S., and A.Y.S.S., realized the analytics experiments and pharmacological assays. H.O.C. supervised the pharmacologic tests. T.A.T. supervised the analytics experiments. J.C.T.C. reviewed the results and final text and was the Study General Coordinator. All authors read and approved the final manuscript.
Funding
CAPES (1723/2018-00 PROCAD-AMAZONIA), CNPq - MAI-DAI Program (Proc. 421808/2022-5), and INCT North-Northeast Network of Phytoproducts – CNPq.
Institutional Review Board Statement
The authors followed the guidelines of the legislation for animal experimentation established by the National Council for Animal Experimentation of Brazil (CONCEA). The Ethics Committee on the Use of Animals (CEUA) of the Federal University of Amapá-UNIFAP approved the project under protocol number 02/2022.
Acknowledgments
The CAPES funded this study - PROCAD AMAZONIA, CNPq - MAI-DAI Program, and INCT North-Northeast Network of Phytoproducts - CNPq.
Conflicts of Interest
The authors declare that they have no known competing for financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Adams, J. U., Geller, E. B., & Adler, M. W. (1994). Receptor selectivity of icv morphine in the rat cold water tail-flick test.
- Aggarwal, V., Kashyap, D., Sak, K., Tuli, H. S., Jain, A., Chaudhary, A., Garg, V. K., Sethi, G., & Yerer, M. B. (2019). Molecular mechanisms of action of tocotrienols in cancer: Recent trends and advancements. In International Journal of Molecular Sciences (Vol. 20, Número 3). MDPI AG. [CrossRef]
- Aktary, N., Sultana, S., & Hossain, M. L. (2019). Assessment of analgesic and neuropharmacological activity of leaves of Bixa orellana (Family: Bixaceae). International Journal of Scientific Reports, 6(1), 13. [CrossRef]
- Atalay, S., Jarocka-karpowicz, I., & Skrzydlewskas, E. (2020). Antioxidative and anti-inflammatory properties of cannabidiol. Em Antioxidants (Vol. 9, Número 1). MDPI. [CrossRef]
- Bannon, A. W., & Malmberg, A. B. (2007). Models of nociception: hot-plate, tail-flick, and formalin tests in rodents. Current protocols in neuroscience / editorial board, Jacqueline N. Crawley ... [et al.], Chapter 8. [CrossRef]
- Batista, E. K. F., Trindade, H. I., Lira, S. R. S., Muller, J. B. B. S., Silva, L. L. B., & Batista, M. C. S. (2016). Atividades antinociceptiva e antiinflamatória do extrato etanólico de Luehea divaricata. Revista Brasileira de Plantas Medicinais, 18(2), 433–441. [CrossRef]
- Batista, M. A., Dos Santos, A. V. T. de L. T., Nascimento, A. L. Do, Moreira, L. F., Souza, I. R. S., Silva, H. R. da, Pereira, A. C. M., Hage-Melim, L. I. da S., & Carvalho, J. C. T. (2022). Potential of the Compounds from Bixa orellana Purified Annatto Oil and Its Granules (Chronic®) against Dyslipidemia and Inflammatory Diseases: In Silico Studies with Geranylgeraniol and Tocotrienols. Molecules, 27(5). [CrossRef]
- Borges, R. S., Lima, E. S., Keita, H., Ferreira, I. M., Fernandes, C. P., Cruz, R. A. S., Duarte, J. L., Velázquez-Moyado, J., Ortiz, B. L. S., Castro, A. N., Ferreira, J. V., da Silva Hage-Melim, L. I., & Carvalho, J. C. T. (2018). Anti-inflammatory and antialgic actions of a nanoemulsion of Rosmarinus officinalis L. essential oil and a molecular docking study of its major chemical constituents. Inflammopharmacology, 26(1), 183–195. [CrossRef]
- Bunman, S., Muengtaweepongsa, S., Piyayotai, D., Charlermroj, R., Phuengwas, S., & Makornwattana, M. (2022). Study the Effect of Cannabidiol Topical on Antinociceptive and Anti-inflammatory Activities in Animal Model. [CrossRef]
- Collier, J., Dinneen, L. C., Johnson, C. A., & Schneider, C. (1968). THE ABDOMINAL CONSTRICTION RESPONSE AND ITS SUPPRESSION BY ANALGESIC DRUGS IN THE MOUSE. Em Br. J. Pharmac. Chemother (Vol. 32).
- Costa, B., Colleoni, M., Conti, S., Parolaro, D., Franke, C., Trovato, A. E., & Giagnoni, G. (2004). Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn-Schmiedeberg’s Archives of Pharmacology, 369(3), 294–299. [CrossRef]
- de Oliveira Carvalho, H., Gonçalves, D. E. S., Picanço, K. R. T., de Lima Teixeira dos Santos, A. V. T., Lucia, M., Hu, X., Fernandes, C. P., Ferreira, I. M., & Carvalho, J. C. T. (2021). Actions of Cannabis sativa L. fixed oil and nanoemulsion on venom-induced inflammation of Bothrops moojeni snake in rats. Inflammopharmacology, 29(1), 123–135. [CrossRef]
- de Oliveira Carvalho, H., Laura, A., Sauma, R., Emanuelle, D., Gonçalves, S., Gomes, L., Ribeiro Da Silva, H., César, A., Pereira, M., Carlos, J., & Carvalho, T. (2022). Intramuscular compatibility of an injectable nanodispersion anti-inflammatory (Chronic®) from a standardized Bixa orellana oil: a toxicological study in Wistar rats. [CrossRef]
- Dubuisson, D., & Dennis, S. G. (1977). THE FORMALIN TEST: A QUANTITATIVE STUDY OF THE ANALGESIC EFFECTS OF MORPHINE, MEPERIDINE, AND BRAIN STEM STIMULATION IN RATS AND CATS—Em Pain (Vol. 4). ElsevirrlNorth-Holland Biomedical Press.
- Foss, J. D., Farkas, D. J., Huynh, L. M., Kinney, W. A., Brenneman, D. E., & Ward, S. J. (2021a). Behavioural and pharmacological effects of cannabidiol (CBD) and the cannabidiol analogue KLS-13019 in mouse models of pain and reinforcement. British Journal of Pharmacology, 178(15), 3067–3078. [CrossRef]
- Foss, J. D., Farkas, D. J., Huynh, L. M., Kinney, W. A., Brenneman, D. E., & Ward, S. J. (2021b). Behavioural and pharmacological effects of cannabidiol (CBD) and the cannabidiol analogue KLS-13019 in mouse models of pain and reinforcement. British Journal of Pharmacology, 178(15), 3067–3078. [CrossRef]
- Hill, K. P., Palastro, M. D., Johnson, B., & Ditre, J. W. (2017). Cannabis and Pain: A Clinical Review. Em Cannabis and cannabinoid research (Vol. 2, Número 1, p. 96–104). NLM (Medline). [CrossRef]
- İnaltekin, A., & Kivrak, Y. (2021). Evaluation of the effect of vortioxetine on pain threshold by hot-plate test in mice. Noropsikiyatri Arsivi, 58(4), 274–277. [CrossRef]
- Menezes, P. M. N., Araújo, T. C. de L., Pereira, E. C. V., Neto, J. A., Silva, D. S., Brito, M. C., Lima, K. S. B., Monte, A. P. O. do, Matos, M. H. T. de, Lima, R. S. de, Ribeiro, L. A. de A., Silva, F. S., & Rolim, L. A. (2021). Investigation of antinociceptive, antipyretic, antiasthmatic, and spasmolytic activities of Brazilian Cannabis sativa L. roots in rodents. Journal of Ethnopharmacology, 278, 114259. [CrossRef]
- Mitchell, M. J., Billingsley, M. M., Haley, R. M., Wechsler, M. E., Peppas, N. A., & Langer, R. (2021). Engineering precision nanoparticles for drug delivery. Em Nature Reviews Drug Discovery (Vol. 20, Número 2, p. 101–124). Nature Research. [CrossRef]
- Mlost, J., Bryk, M., & Starowicz, K. (2020a). Cannabidiol for pain treatment: Focus on pharmacology and mechanism of action. Em International Journal of Molecular Sciences (Vol. 21, Número 22, p. 1–22). MDPI AG. [CrossRef]
- Mlost, J., Bryk, M., & Starowicz, K. (2020b). Cannabidiol for pain treatment: Focus on pharmacology and mechanism of action. Em International Journal of Molecular Sciences (Vol. 21, Número 22, p. 1–22). MDPI AG. [CrossRef]
- Moore, C. F., & Weerts, E. M. (2022). Cannabinoid tetrad effects of oral Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD) in male and female rats: sex, dose-effects and time course evaluations. Psychopharmacology, 239(5), 1397–1408. [CrossRef]
- Nascimento, G., Ferrari, D. P., Guimaraes, F. S., Del Bel, E. A., Bortolanza, M., & Ferreira- Junior, N. C. (2020). Cannabidiol increases the nociceptive threshold in a preclinical model of Parkinson’s disease. Neuropharmacology, 163. [CrossRef]
- Neelakantan, H., Tallarida, R. J., Reichenbach, Z. W., Tuma, R. F., Ward, S. J., & Walker, E. A. (2015). Distinct interactions of cannabidiol and morphine in three nociceptive behavioral models in mice. Behavioural Pharmacology, 26(3), 304–314. [CrossRef]
- P1zziketti, R. J., Pressman, N. S., Geller, E. B., Cowan, A., Adler, M. W., & Pizziketti, R. J. (1985). 23 Elsevier RAT COLD WATER TAlL-FLICK: A NOVEL ANALGESIC TEST THAT DISTINGUISHES OPIOID AGONISTS FROM MIXED AGONIST-ANTAGONISTS. in European Journal of Pharmacology (Vol. 119).
- Raja, S. N., Carr, D. B., Cohen, M., Finnerup, N. B., Flor, H., Gibson, S., Keefe, F. J., Mogil, J. S., Ringkamp, M., Sluka, K. A., Song, X. J., Stevens, B., Sullivan, M. D., Tutelman, P. R., Ushida, T., & Vader, K. (2020). The revised International Association for the Study of Pain definition of pain: concepts, challenges, and compromises. Em Pain (Vol. 161, Número 9, p. 1976–1982). Lippincott Williams and Wilkins. [CrossRef]
- Ranasinghe, R., Mathai, M., & Zulli, A. (2022). Revisiting the therapeutic potential of tocotrienol. Em BioFactors (Vol. 48, Número 4, p. 813–856). John Wiley and Sons Inc. [CrossRef]
- Razavi, Y., Rashvand, M., Sharifi, A., Haghparast, A., Keyhanfar, F., & Haghparast, A. (2021). Cannabidiol microinjection into the nucleus accumbens attenuated nociceptive behaviors in an animal model of tonic pain. Neuroscience Letters, 762, 136141. [CrossRef]
- Rodrigues, A. P. S., da Silva Barbosa, R., Pereira, A. C. M., Batista, M. A., Sales, P. F., Ferreira, A. M., Colares, N. N. D., da Silva, H. R., Soares, M. O. dos S., da Silva Hage-Melim, L. I., & Carvalho, J. C. T. (2022). Ormona® SI and Ormona® RC—New Nutraceuticals with Geranylgeraniol, Tocotrienols, Anthocyanins, and Isoflavones—Decrease High-Fat Diet-Induced Dyslipidemia in Wistar Rats. Nutraceuticals, 2(4), 311–322. [CrossRef]
- Sadaka, A. H., Canuel, J., Febo, M., Johnson, C. T., Bradshaw, H. B., Ortiz, R., Ciumo, F., Kulkarni, P., Gitcho, M. A., & Ferris, C. F. (2023). Effects of inhaled cannabis high in Δ9-THC or CBD on the aging brain: A translational MRI and behavioral study. Frontiers in Aging Neuroscience, 15. [CrossRef]
- Sales, P. F., Nascimento, A. L. do, Pinheiro, F. C., Alberto, A. K. M., Teixeira dos Santos, A. V. T. de L., Carvalho, H. de O., de Souza, G. C., & Carvalho, J. C. T. (2023). Effect of the Association of Fixed Oils from Abelmoschus esculentus (L.) Moench, Euterpe oleracea Martius, Bixa orellana Linné and Chronic SM® on Atherogenic Dyslipidemia in Wistar Rats. Molecules, 28(18). [CrossRef]
- Sharma, S. S., Kumar, A., & Kaundal, R. K. (2008). Protective effects of 4-amino1,8-napthalimide, a poly (ADP-ribose) polymerase inhibitor in experimental diabetic neuropathy. Life Sciences, 82(11–12), 570–576. [CrossRef]
- Shilpi, J. A., Taufiq-Ur-Rahman, M., Uddin, S. J., Alam, M. S., Sadhu, S. K., & Seidel, V. (2006). Preliminary pharmacological screening of Bixa orellana L. leaves. Journal of Ethnopharmacology, 108(2), 264–271. [CrossRef]
- Silva, J. C., Oliveira-Júnior, R. G., Silva, M. G., Lavor, E. M., Soares, J. M. D., Lima-Saraiva, S. R. G., Diniz, T. C., Medeiros, M. A. M. B., Lima, L. M., Barreiro, E. J., & Almeida, J. R. G. S. (2021). In Mice Evaluation of Antinociceptive and Anti-inflammatory Activity of N-acylhydrazone Derivative LASSBio-1587. Revista Virtual de Quimica, 13(6), 1467–1472. [CrossRef]
- Sofia, R. D., Vassar, H. B., & Knobloch, L. C. (1975). Comparative Analgesic Activity of Various Naturally Occurring Cannabinoids in Mice and Rats. In Psychopharmacology (Berl.) (Vol. 40). Springer-Verlag.
- Tejpal Singh, H. S., Aminuddin, A. A., Pang, K. L., Ekeuku, S. O., & Chin, K. Y. (2023). The Role of Tocotrienol in Arthritis Management—A Scoping Review of Literature. Em Pharmaceuticals (Vol. 16, Número 3). MDPI. [CrossRef]
- Tj, A., Berge, O.-G., Hunskaar, S., Henrik Rosland, J., & Hole, K. (1992). The formalin test: an evaluation of the method. Em Puin (Vol. 51).
- Urits, I., Gress, K., Charipova, K., Habib, K., Lee, D., Lee, C., Jung, J. W., Kassem, H., Cornett, E., Paladini, A., Varrassi, G., Kaye, A. D., & Viswanath, O. (2020). Use of cannabidiol (CBD) for the treatment of chronic pain. In Best Practice and Research: Clinical Anaesthesiology (Vol. 34, Número 3, p. 463–477). Bailliere Tindall Ltd. [CrossRef]
- Williams, A. C. D. C., & Craig, K. D. (2016). Updating the definition of pain. Em Pain (Vol. 157, Número 11, p. 2420–2423). Lippincott Williams and Wilkins. [CrossRef]
- Yong, S. T., Wong, H., & Mohammad, M. (2014). Tocotrienol and tocopherol contents of annatto seed accessions. https://www.researchgate.net/publication/318601954.
- Zielinska, A., Carreiró, F., Oliveira, A. M., Neves, A., Pires, B., Nagasamy Venkatesh, D., Durazzo, A., Lucarini, M., Eder, P., Silva, A. M., Santini, A., & Souto, E. B. (2020). Polymeric Nanoparticles: Production, Characterization, Toxicology and Ecotoxicology. Em Molecules (Vol. 25, Número 16). MDPI AG. [CrossRef]
Figure 1.
(A) Chromatographic profile of Bixa orellana oil with a major peak corresponding to δ-tocotrienol (C27H40O2) at a retention time of 28.6 minutes. (B) Chromatographic profile of Cannabis sativa extract showing the retention time of cannabidiol at 30.54 minutes.
Figure 1.
(A) Chromatographic profile of Bixa orellana oil with a major peak corresponding to δ-tocotrienol (C27H40O2) at a retention time of 28.6 minutes. (B) Chromatographic profile of Cannabis sativa extract showing the retention time of cannabidiol at 30.54 minutes.
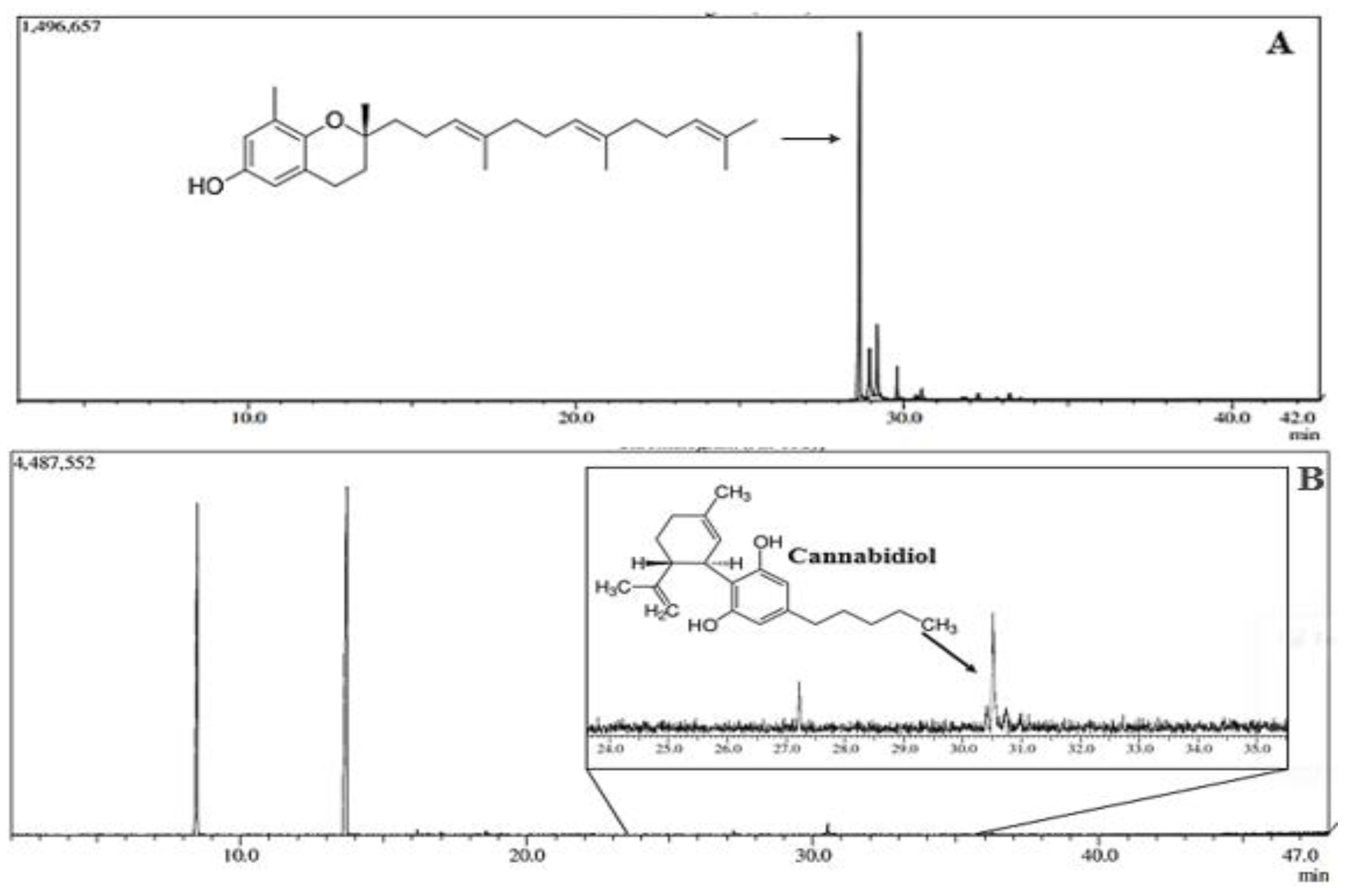
Figure 2.
Effect of treatment with CSE (40 mg/kg) associated with Chronic® (400 mg/kg) and CSE associated with Chronic In® (10 mg/kg) in the acetic acid-induced abdominal writhing test. A. Represents the number of writhes in the CSE + CHR OR, CHR OR, and CSE + CHR OR + NAL groups. B. Represents the number of writhes in the CSE + CHR IN, CHR IN, and CSE + CHR IN + NAL groups. Results are presented as mean ± SD (n=5). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. *** indicates p < 0.001.
Figure 2.
Effect of treatment with CSE (40 mg/kg) associated with Chronic® (400 mg/kg) and CSE associated with Chronic In® (10 mg/kg) in the acetic acid-induced abdominal writhing test. A. Represents the number of writhes in the CSE + CHR OR, CHR OR, and CSE + CHR OR + NAL groups. B. Represents the number of writhes in the CSE + CHR IN, CHR IN, and CSE + CHR IN + NAL groups. Results are presented as mean ± SD (n=5). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. *** indicates p < 0.001.
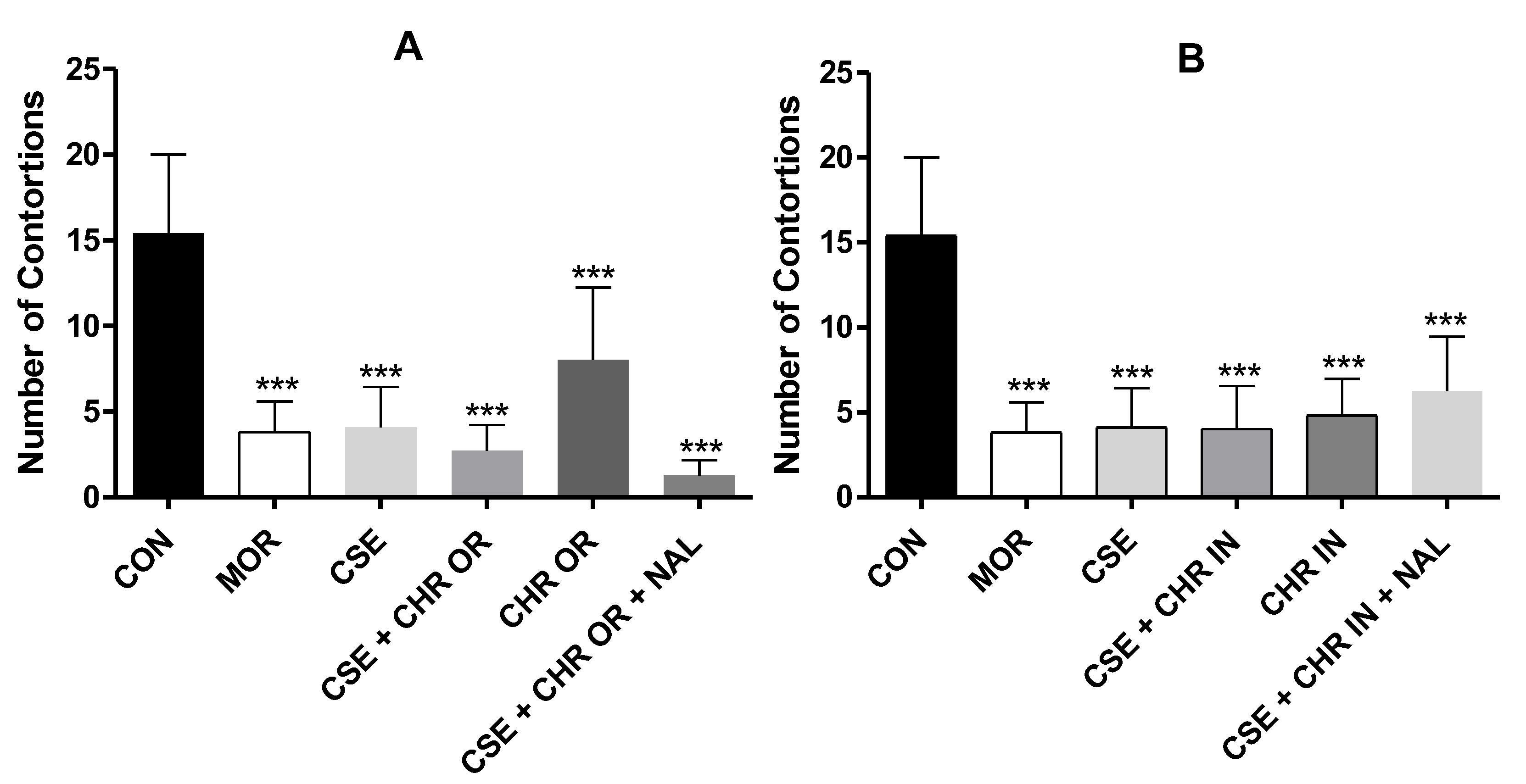
Figure 3.
Formalin test. The rats (n=5) were treated with placebo, morphine (10 mg/kg), CSE (40 mg/kg), CSE + CHR OR (400 mg/kg), and CSE + CHR OR + NAL (2 mg/kg). Nociceptive behaviors were recorded over time (seconds). (A) Early phase (neurogenic phase: 0-5 minutes) and (B) Late phase (inflammatory phase: 15-30 minutes). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. *** indicates p < 0.001.
Figure 3.
Formalin test. The rats (n=5) were treated with placebo, morphine (10 mg/kg), CSE (40 mg/kg), CSE + CHR OR (400 mg/kg), and CSE + CHR OR + NAL (2 mg/kg). Nociceptive behaviors were recorded over time (seconds). (A) Early phase (neurogenic phase: 0-5 minutes) and (B) Late phase (inflammatory phase: 15-30 minutes). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. *** indicates p < 0.001.
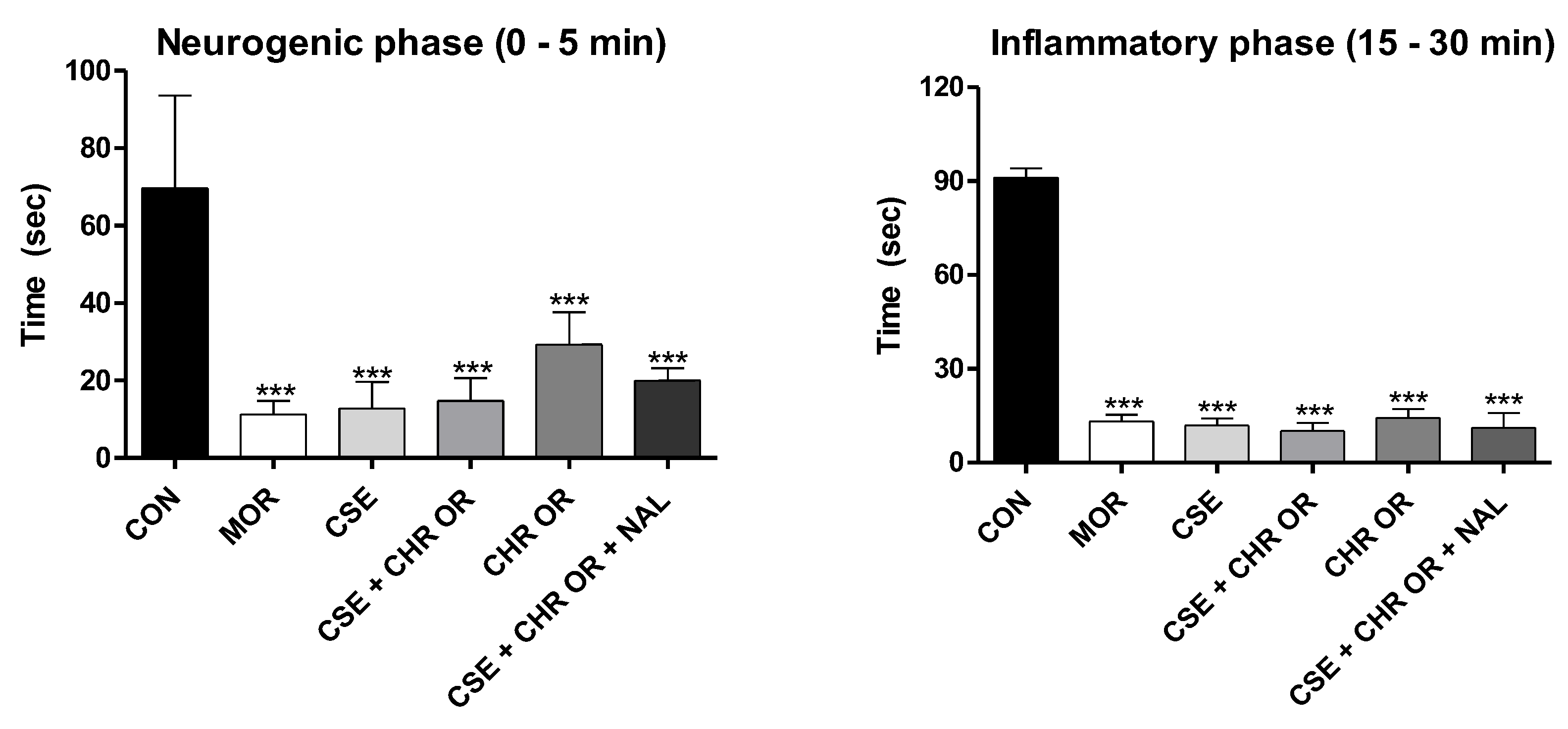
Figure 4.
Formalin test. The rats (n=5) were treated with placebo, morphine (10 mg/kg), CSE (40 mg/kg), CSE + CHR IN (10 mg/kg), and CSE + CHR IN + NAL (2 mg/kg). Nociceptive behaviors were recorded over time (seconds). (A) Early phase (neurogenic phase: 0-5 minutes) and (B) Late phase (inflammatory phase: 15-30 minutes). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. *, **, and *** indicate p < 0.05, p < 0.01, and p < 0.001, respectively.
Figure 4.
Formalin test. The rats (n=5) were treated with placebo, morphine (10 mg/kg), CSE (40 mg/kg), CSE + CHR IN (10 mg/kg), and CSE + CHR IN + NAL (2 mg/kg). Nociceptive behaviors were recorded over time (seconds). (A) Early phase (neurogenic phase: 0-5 minutes) and (B) Late phase (inflammatory phase: 15-30 minutes). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. *, **, and *** indicate p < 0.05, p < 0.01, and p < 0.001, respectively.
Figure 5.
Effect of treatment with CSE (40 mg/kg) associated with Chronic® (400 mg/kg) and CSE associated with Chronic In® (10 mg/kg) in the hot plate test. A. Represents the latency time (seconds) in the CSE + CHR OR, CHR OR, and CSE + CHR OR + NAL groups. B. Represents the latency time (seconds) in the CSE + CHR IN, CHR IN, and CSE + CHR IN + NAL groups. Results are presented as mean ± SD (n=5). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. * and *** indicate p < 0.05 and p < 0.001, respectively.
Figure 5.
Effect of treatment with CSE (40 mg/kg) associated with Chronic® (400 mg/kg) and CSE associated with Chronic In® (10 mg/kg) in the hot plate test. A. Represents the latency time (seconds) in the CSE + CHR OR, CHR OR, and CSE + CHR OR + NAL groups. B. Represents the latency time (seconds) in the CSE + CHR IN, CHR IN, and CSE + CHR IN + NAL groups. Results are presented as mean ± SD (n=5). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. * and *** indicate p < 0.05 and p < 0.001, respectively.
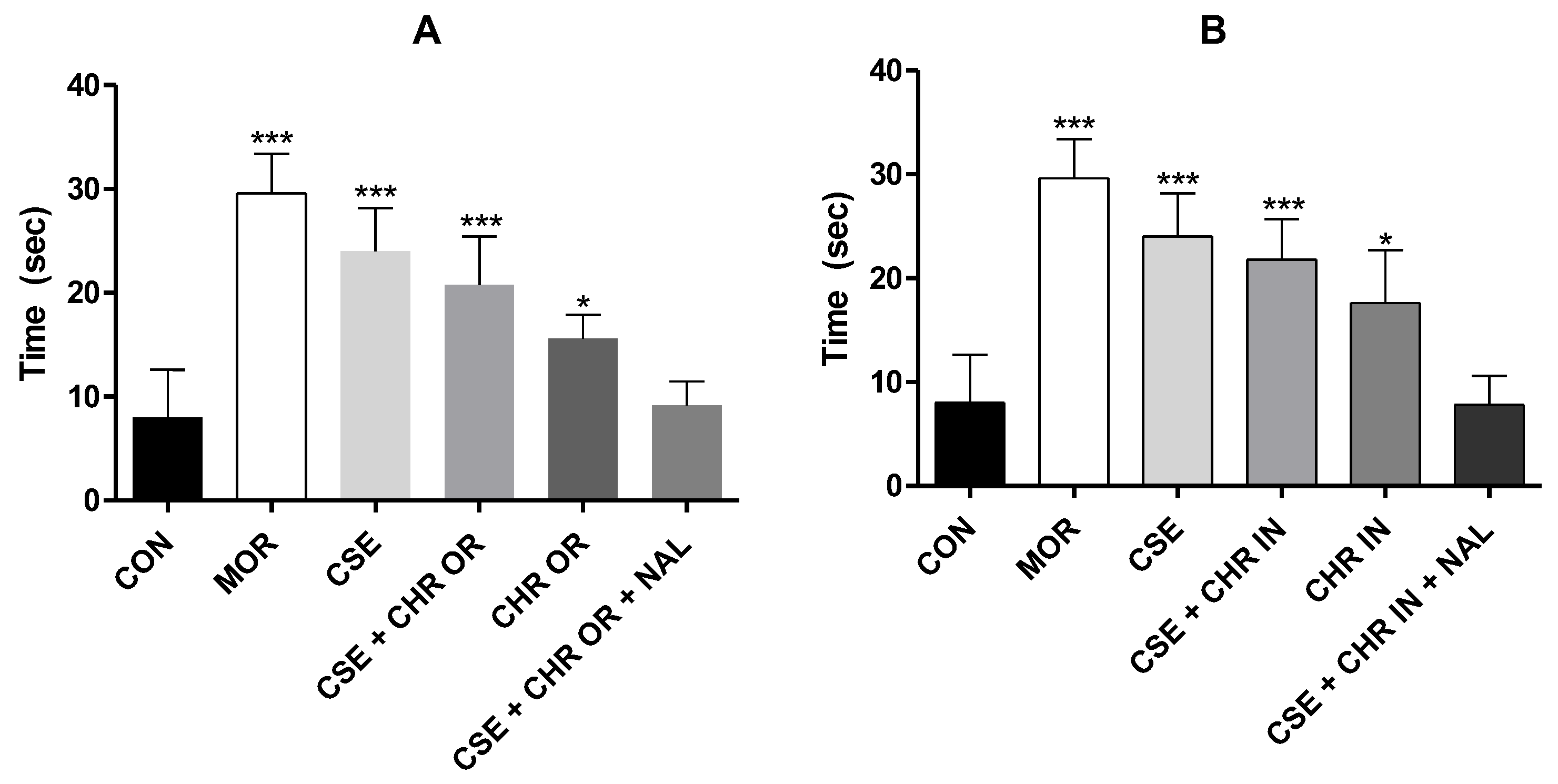
Figure 6.
Effect of treatment with CSE (40 mg/kg) associated with Chronic (400 mg/kg) and CSE associated with Chronic In (10 mg/kg) in the cold-water tail withdrawal test. A. Represents the tail withdrawal time (seconds) in the CSE + CHR OR, CHR OR, and CSE + CHR OR + NAL groups. B. Represents the tail withdrawal time (seconds) in the CSE + CHR IN, CHR IN, and CSE + CHR IN + NAL groups. Results are presented as mean ± SD (n=5). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. * and *** indicate p < 0.05 and p < 0.001, respectively.
Figure 6.
Effect of treatment with CSE (40 mg/kg) associated with Chronic (400 mg/kg) and CSE associated with Chronic In (10 mg/kg) in the cold-water tail withdrawal test. A. Represents the tail withdrawal time (seconds) in the CSE + CHR OR, CHR OR, and CSE + CHR OR + NAL groups. B. Represents the tail withdrawal time (seconds) in the CSE + CHR IN, CHR IN, and CSE + CHR IN + NAL groups. Results are presented as mean ± SD (n=5). Differences between groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s test. * and *** indicate p < 0.05 and p < 0.001, respectively.
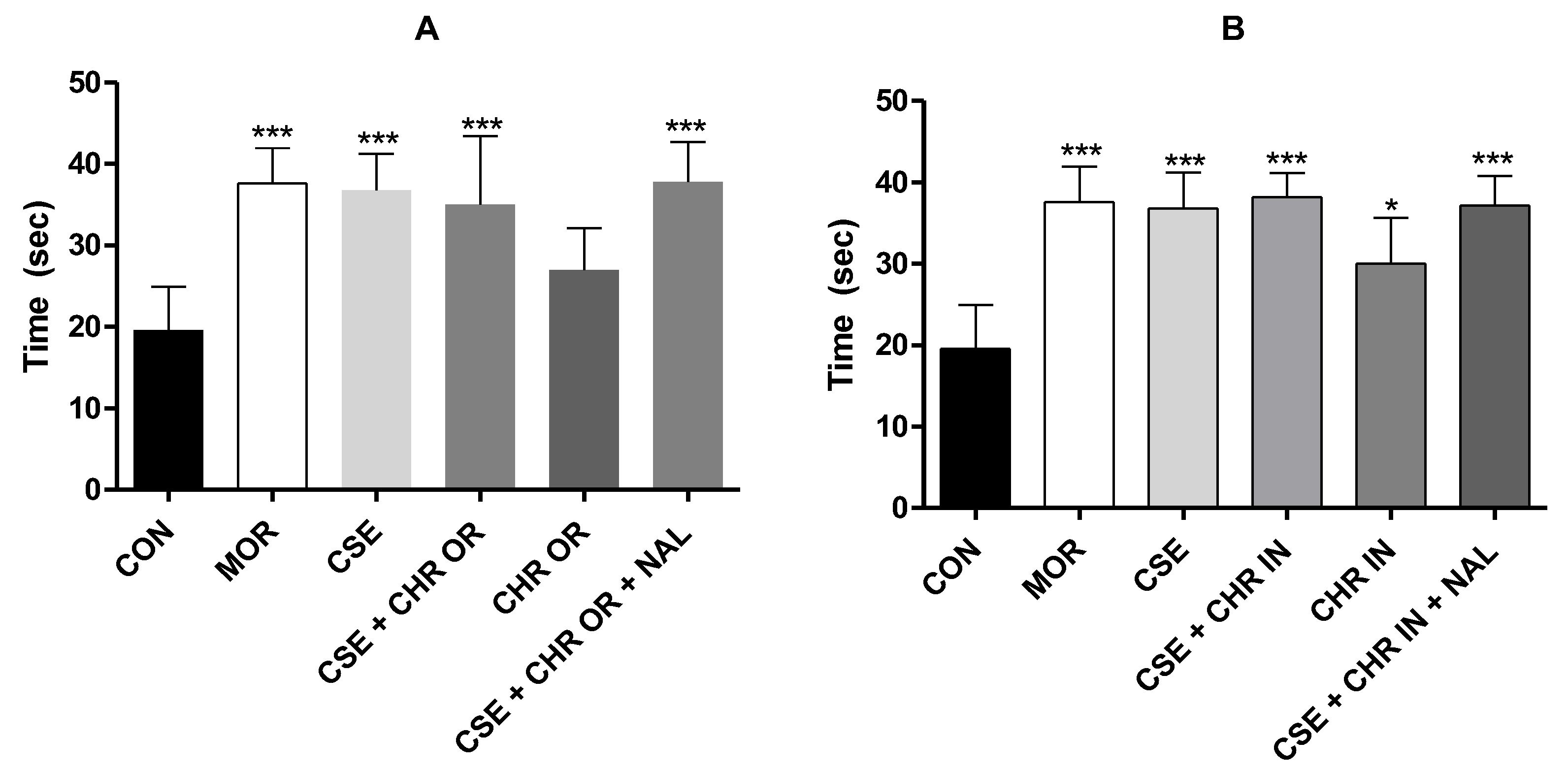

Table 1.
Analysis of Zeta potential (mV), average size (nm), and polydispersity index (PDI) of the nanodispersion (Chronic In®), analyzed at 1 and 30 days after the NBO was obtained, presenting mean and standard deviation (n=3).
Table 1.
Analysis of Zeta potential (mV), average size (nm), and polydispersity index (PDI) of the nanodispersion (Chronic In®), analyzed at 1 and 30 days after the NBO was obtained, presenting mean and standard deviation (n=3).
| P. Zeta (mV) | Size (nm) | PDI | pH | |
|---|---|---|---|---|
| Day 1 | 19,86±0,60 | 53,15±0,64 | 0,533±0,008 | 5,8±0,01 |
| Day 30 | 19,66±1,45 | 59,90±3,63 | 0,594±0,014 | 5,8±0,01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



