
Submitted:
31 October 2024
Posted:
04 November 2024
You are already at the latest version
Abstract
Vaccine adjuvants are crucial for reinforcing the immunogenicity of vaccines. Therefore, they are widely used in the aquaculture sector as vaccine components facilitating the efficient prevention of infectious diseases and promoting sustainable teleost fish growth. Despite their benefits, there has been a growing concern about the potential adverse effects of vaccine adjuvants in teleost fish, connoting a valid impact on their overall health and welfare. Among adjuvants used in aquaculture vaccinology, nanoparticle-based formulations have given rise to a promising new alternative to traditional options, such as oil-based emulsions and aluminum compounds, offering the benefit of minimizing relevant side-effects. The aim of this paper was to review the current status of the adjuvants used in aquaculture, provide a description and an evaluation of their mode of action and side-effects, and explore the potential use of nanoparticle formulations as adjuvants to improve the efficacy of aquaculture vaccines. By demonstrating and assessing the equilibrium between teleost fish welfare and immunological efficacy, this review presents a collective perspective that will assist in establishing a framework for the utilization of effective, species-specific practices around adjuvant use in aquaculture, while also addressing the challenges of welfare-friendly immunization.
Keywords:
aquaculture
; vaccines
; adjuvants
; nanoparticles
; immunization
; inflammation
; granulomas
; lesions
; sustainability
; welfare
; health
; side effects
; teleost fish
1. Introduction
In the dynamic landscape of aquaculture, the use of vaccines has emerged as a cornerstone for disease prevention, marking an essential checkpoint in sustainable industry growth [1]. Vaccines constitute the most efficient solution to combat impeding pathogen threats, especially in the context of modern aquaculture, where increased rearing densities have caused a spike in microbial agent transmission [2,3]. The ecological pressure exerted by aquaculture practices in the global transmission of pathogens in wild fish populations has added an additional layer to the issue, proving that the efficient and timely development of vaccines is paramount in safeguarding the health of teleost fish species [4,5,6].
To further boost the immunogenicity and the elicitation of robust immune responses, auxiliary substances called adjuvants have long been used in aquaculture vaccine formulations to maximize the effectiveness of the administered vaccine preparations. Traditionally, these substances were abstractly described as agents that either improved vaccine potency or efficiency, two terms that were used to signify boosts to adaptive immune responses and the prevention of infection respectively [7]. Considerable advancements in the field of immunology, however, led to a novel, clearer, and more refined term for adjuvants, defining them as structurally heterogenous compounds that regulate the inherent immunogenicity of vaccine antigens [7]. This new definition acknowledges the role of adjuvants in boosting immunizations through specific and intricate mechanisms of action, thereby consolidating a promising field for scientific exploration in the field of immunology.
The use of adjuvants in aquaculture is compatible with every method of vaccine administration, including injection, immersion, and oral vaccination. The vast majority of vaccines utilized in aquaculture, however, are administered via intraperitoneal (IP) injection, since this method of vaccine delivery provides the most adequate and long-lasting immunization in teleost fish [7,8,9]. Nevertheless, the use of specific adjuvants has enabled other vaccine delivery methods, such as immersion vaccination, to be efficiently utilized in aquaculture, thus opening the gateway for alternative methods of vaccine administration. By enhancing the effectiveness of alternative vaccination methods, the industry can potentially shift from the exclusive use of injection vaccination to other forms of vaccine delivery that will offer benefits not only in logistical and cost-related aspects, but also in animal welfare, by reducing the stress induced during the immunization process.
In this quest for innovative and sustainable immunization strategies, nanoparticles have paved the way for a promising alternative to traditional adjuvant formulations in the aquaculture sector. Thanks to their small size and their ability to provide tailored platforms for controlled release, these formulations offer enhanced antigen delivery and improved immune response, while minimizing the required dosages and mitigating any relevant side effects associated with traditional adjuvants [10,11,12]. Recent data suggest that nanoparticles not only boost the efficacy of conventional injectable vaccines, but also improve immunization methods that involve mucosal delivery such as immersion and oral vaccination [13]. By promoting enhanced antigen uptake and by stimulating both innate and adaptive immunity, nanoparticle-based adjuvants can revolutionize vaccine protocols in aquaculture, potentially reducing the reliance on invasive methods in favor of more cost-efficient and welfare-friendly routes. Furthermore, their capacity to encapsulate and protect antigens improves vaccine stability, prospectively broadening their use across various aquatic species and environmental conditions [14].
This review article aims to synthesize a comprehensive perspective on the most popular adjuvants currently used in aquaculture vaccinology, drawing information from literature not only about their exact modes of action, but also about advances made in understanding and mitigating their side-effects. Additionally, the potential of utilizing nanoparticle formulations as an alternative to traditional vaccine adjuvants in the industry of aquaculture is discussed, offering a complete overview of the future directions for optimizing vaccine safety and enhancing immune responses. Spreading research findings regarding adjuvant side effects across different and diverse combinations of adjuvants, vaccines, fish species, and rearing conditions is paramount in broadening the knowledge on the assessment of these adverse effects, thereby safeguarding aquatic organism health and sustainability in practice. Delving deeper into the matter, understanding exactly how adjuvants work is essential for unraveling their importance and integration in vaccination strategies and their implications for teleost fish welfare.
2. Mode of Action: The Two-Signal Model
The precise mechanisms by which adjuvants enhance immune responses have long been, and continue to be, a subject of significant debate and investigation. While adjuvants are known to enhance the body’s immune response to antigens, the detailed processes by which they achieve this remain elusive [15,16]. Researchers have proposed many assumptions, including the stimulation of innate immune receptors, the creation of a “depot effect” that prolongs antigen exposure, and the modulation of the immune environment; yet, complete understanding is still lacking, especially in the context of aquatic organisms [7]. In this quest for understanding how adjuvants work, Schijns’ model, also known as the two-signal model [17], stands out as the most comprehensive model for explaining and categorizing their mechanisms of action, providing a structured approach to these complex interactions, while highlighting the need for continued research to fully unravel the intricacies of adjuvant function to improve vaccine development. According to this proposed model, vaccine adjuvants can be categorized based on their capacity to facilitate two types of signals: type 1 and type 2 signals.
2.1. Type 1 Signals
Type 1 signals typically refer to the process of antigen presentation by specific types of immune cells, called professional antigen presenting cells (pAPCs) [7]. These cells can bind to antigens through highly polymorphic MHC class II proteins expressed on their surface and subsequently display them to T-cells to facilitate their differentiation into CD4+ T-cells, known as helper T-cells or Th cells. This sequence of events is essential not only to the production of antibodies, but also to the induction of innate immune responses, since Th cells can aid by providing necessary co-stimulatory signals that are unique to the type of antigen that is recognized [18,19,20,21]. Several immune cells are known to possess antigen-presenting functions, including B-cells, macrophages, and dendritic cells (DCs), and this is a pattern that is conserved in teleost fish, albeit with some differences [22].
Adjuvants that influence type 1 signals and antigen presentation thereof can be categorized in a distinct group that encompasses type 1 signal facilitators that can essentially catalyze the antigen presentation process. The exact way in which type 1 adjuvants influence can vary, however it all boils down to improving the immunoavailability of the vaccine’s antigens to antigen presenting cells [16]. Based on that, and by taking the geographical concept of immune reactivity into consideration [23], adjuvants that have a significant effect on an antigen’s presentability in the context of space, time, and availability can collectively be categorized as type 1 adjuvants according to the two-signal model.
Although type 1 adjuvants are widely implemented in vaccinology and are considered crucial for eliciting robust immune responses, the precise mechanisms behind their effects remain largely elusive until today [16]. Since type 1 adjuvants are used extensively in injectable vaccines, the hypothesis for the most important mechanism of action thus far proposes that these adjuvants facilitate a “depot effect”, in which a localized reservoir of the antigen is formed at the site of injection [7,17]. This allows for a controlled, gradual, and sustained release of the antigen over an extended period of time, therefore affecting its overall immunoavailability to the host’s immune system cells [16]. By maintaining the antigen’s presence in the body for a longer period, the depot effect maximizes the immune system’s exposure to the vaccine’s antigen, thereby promoting a more robust and prolonged immune response.
The evidence on whether the “depot effect” is solely responsible for the immunostimulating effects of type 1 adjuvants or whether it simply affects it remains debatable, keeping many researchers conflicted on the topic [24]. Many studies imply that the tissue microenvironment of the injection site plays an important role in immunostimulation, as many antigen presenting cells are attracted to chemotactic factors produced locally at this site [16,24]. Many key players of the innate wing of the immune system enter the injured tissue and influence the function of APCs. Based on that, it appears that type 1 adjuvants can potentially promote inflammatory signals, adding an additional layer of complexity to the mode of actions supporting their immune-enhancing effects.
2.2. Type 2 Signals
On the other hand, type 2 signals according to the two-signal model are regarded as secondary, ancillary signals that facilitate the activation of adaptive immune cells like B-cells and T-cells [7,16,17]. These signals provide necessary co-stimulation during the process of antigen presentation, which is fulfilled through the equilibrated and combinatory effects of co-stimulatory or co-inhibitory signals [16]. These signals can thus tailor the scale and the effect of adaptive immune responses, resulting in varying outcomes of immunostimulation that aid in enhancing the process of immunization. As such, adjuvants that facilitate type 2 signals can collectively be grouped into type 2 adjuvants.
The activity of adjuvants that facilitate type 2 signals has been well described, even down to the molecular level, as opposed to those that facilitate type 1 signals [16]. The cells of the innate immune system express pattern recognition receptors (PRRs) that are used to recognize pathogens by binding to specific pathogen-associated molecular patterns (PAMPs) [16]. These molecular patterns are typically structural elements that are unique to different classes of pathogens and are recognized by different kinds of PRRs [25,26]. The recognition of pathogen-associated molecular patterns by the receptors of the innate immune cells orchestrates signaling cascades that facilitate the induction of both humoral and cellular immune responses. Some of the most important examples of pattern recognition receptors subfamilies and their associated ligands identified so far in teleost fish are summarized in Table 1.
Type 2 adjuvants facilitate the stimulation of the immune system by simulating the recognition of PAMPs by specific pattern recognition receptors. However, another way by which type 2 adjuvants can stimulate the innate immune system lies in the induction of the so-called danger signals that are associated with tissue injury and damage. According to the model proposed by Matzinger, the immune system does not necessarily focus on the recognition of microbial agents by themselves, but rather on the co-recognition of antigens associated with danger signals [43]. Upon entry and infection, microbial agents cause damage and stress to surrounding tissues, enabling the induction of signals that are materialized through the release of damage-associated molecular patterns, or DAMPs. These patterns can include both segments of dead cells and molecules like heat-shock proteins, uric acid, and cytokines that promote the activation of stimulatory signals for the pAPCs [16]. According to Ribeiro & Schijns, this can elucidate the mechanism of action behind several type 1 adjuvants as well, adding an interesting element of overlapping between the immunostimulating machineries involved in both type 1 and type 2 adjuvants.
Overall, it is accepted that the categorization of adjuvants to type 1 or type 2 facilitators is not necessarily mutually exclusive [16]. The intricate mechanisms behind the activation of signaling cascades that modulate immune responses can be affected in numerous ways, highlighting that the study of different adjuvants is important in the elucidation of the interplay between immune-enhancing compounds and the elements of the immune system. Adjuvants that facilitate type 2 signals can influence a plethora of immunostimulating pathways, and that is evidenced by the fact that one of the most well-studied PRR receptors, the toll-like receptors (TLRs), can aid in the activation of both innate and adaptive immunity [16], bridging the gap between the two immune subsystems. Upon binding to pattern recognition receptors, different molecules can have different outcomes in terms of immune-enhancing capabilities, thus facilitating the induction of different stimulatory signals that are materialized through the secretion of cytokines [26,27,28,29,44].
2.3. Further Elaborations on the Two-Signal Model
Since the binding of PRR ligands precedes the secretion of cytokines and co-stimulatory molecules, it has been suggested that the binding of PAMPs to the PRRs constitutes a separate type of signal, often referred to as a type 0 signal [16]. According to this proposal, this type of signal facilitates the initiation of all down-stream pathways involved in the secretion of co-stimulatory signals that make up the type 2 signal category, ultimately making a distinction between the effects of PRR ligand binding and the stimulatory effects of type 2 signals on the activation of naive B-cells and T-cells. This distinction can potentially aid in advancing the field of adjuvant immunology by allowing research to focus on adjuvants that have profound stimulatory effects on each type of signal. This can include studies on PRR agonists co-delivered with antigens, inhibitory molecules that block immune-attenuating signals, and recombinant co-stimulatory molecules that effectively emulate the immune-amplifying capabilities of endogenous cytokines directly [16].
In a recent update of events, further elaborations on Schijns’ proposed model appear to have been made, making the case for a fourth type of signal (type 3 signal) that regulates the proliferation of T-cells into different antigen-specific subpopulations, as with regulatory T-cells (Tregs) and different subsets of Th cells [45]. Additionally, a fifth type of signal called type 4 signal can be used to describe adjuvants that affect the localization of vaccine antigens by directing them to specific anatomical regions within the host’s body [45]. This aspect adds another source of complexity to the activity of adjuvants, as vaccine formulations need to not only activate the immune system, but also ensure the correct direction of immune effector cells and the induced immune response. In aquaculture vaccinology this can be especially important for vaccines targeting mucosal routes, like the skin, the gills, and the intestines.
The mucosal surfaces are primary entry points for many aquatic pathogens [46,47,48]. By potentially utilizing adjuvants that enhance mucosal imprinting of type 4 homing signals, vaccines can be designed to induce robust and localized immune responses that will reduce disease incidence and improve overall teleosts health and aquaculture sustainability. This lies in accordance with the current direction of aquaculture vaccinology that aims to develop vaccines that will not only be able to elicit mucosal immunity, but also be administered through routes that are more preferrable to the industry, such as immersion and oral vaccination [1]. Effectively, this can also reduce the stress induced on fish during the vaccination process, therefore promoting their overall welfare. A schematic overview synthesizing the most recent insights incorporated into signal modelling of vaccine adjuvant activity can be seen in Figure 1.
3. Traditional Adjuvants Used in Aquaculture Vaccinology
In the realm of aquaculture vaccinology, adjuvants are key components that enhance the immunogenicity of vaccines, which makes them indispensable in the development of efficient vaccines for aquatic species. This section will delve into major adjuvants currently employed in aquaculture vaccines, exploring their modes of action, efficacy, and their influence on immune cell activation and homing. By gaining a deeper grasp on the unique properties and applications of these adjuvants, a better understanding of their potential exploitation can be established, leading to the improved assessment of their side-effects and their mitigation thereof in different fish species. An illustration summarizing the main adjuvants discussed below can be seen in Figure 2.
3.1. Oil-Based Emulsions
Oil-based emulsions are widely used in aquaculture vaccine formulations, especially due to their proven efficacy in enhancing immunization [49]. Emulsions consist of a dispersed liquid distributed inside a second liquid, forming two phases, the continuous and the dispersed, where typically the two liquids would not be able to mix naturally. In vaccine formulations, these phases typically consist of an aqueous continuous phase, loaded with the vaccine antigen, and an oily phase that is dispersed [7]. The formation of the emulsion is facilitated by the use of emulsifiers that act as surfactants, stabilizing the emulsion by reducing the surface tension between the two liquids thanks to their polar groups which are hydrophilic and their non-polar groups which are hydrophobic [7,49]. Surfactants can be categorized by taking into consideration the value of the analogy of the hydrophilic to the hydrophobic groups, known as hydrophilic-lipophilic balance (HLB), a term first proposed by Griffin in 1949 [50].
The HLB value of surfactants is a determining factor for the production of different types of emulsions [50,51,52], that have different efficiencies as vaccine adjuvants [7]. Emulsifiers with low HLB values lead to the creation of water-in-oil emulsions (W/O), whereas those with high HLB values tend to form oil-in-water emulsions (O/W) that are typically better tolerated but exhibit weaker immune responses [7]. Certain surfactant system with an intermediate HLB value allow for the creation of W/O/W emulsions, where the continuous phase is aqueous and the dispersed phase is oil, within which an entrapped aqueous phase containing water-soluble or suspended antigens is found. This third type of emulsion has been shown to elicit robust immune responses with a variety of antigens as summarized by Dalmo et al. [7].
The majority of aquaculture vaccines rely on adjuvants that are based on mineral oil emulsions [7,53]. The most common example of such adjuvants consists of Freund’s Complete Adjuvant (FCA), which is a mineral oil-based formulation used to create a W/O emulsion with the addition of a surfactant and heat-killed Mycobacteria [7,53]. Despite being effective however, this adjuvant has raised concerns due to its association with severe side effects during administration, making its implementation restricted to mainly experimental settings, while gradually being replaced by Freund’s Incomplete Adjuvant (FIA), which lacks the mycobacterial elements [7]. Montanide adjuvants are a series of commercially available adjuvants based on various oil emulsions formulated by the company SEPPIC and licensed under the common trade name of MontanideTM. These adjuvants are not exclusively mineral oil-based, but their use has been well-established in the realm of veterinary vaccines, constituting the adjuvant of choice for many commercially available vaccines alongside other mineral oil-based adjuvant brand series in the market. A table listing the currently commercially available MontanideTM series formulations intended for use in teleost fish can be read below in Table 2.
Some of the most commonly used MontanideTM adjuvants in aquaculture vaccines, included the Montanide™ ISA 763A and 763B VG series, whose efficiency has been highlighted in numerous publications dealing with different species of fish and pathogens [54,55,56,57]. However, in accordance with evidence provided by the recent elaborations on the two-signal model, recent studies appear to suggest that the use of different adjuvants, including even commercialized ones, with different vaccine antigens, can result in a totally different type and scale of immunization [58,59,60,61]. This further strengthens the notion that the appropriate selection of the adjuvant-antigen combination is paramount in ensuring a robust and long-lasting immune response upon vaccination in the context of aquatic organisms, alluding to the fact that more research should be focused on such combinations to shed light into the optimization of vaccine formulations.
3.2. Aluminum-Based Compounds
Another category of popular vaccine adjuvants for aquaculture vaccines are aluminum compounds that contain aluminum salts, such as aluminum hydroxide or aluminum phosphate, collectively referred to as alum [7]. The mechanism of action of these adjuvants remains elusive, though quite a number of mechanisms have been proposed to explain their immune-enhancing capabilities [15,62]. Similarly to oil-based emulsions, it has been suggested that alum provides a “depot” effect by adsorbing the vaccine antigen onto its particles and creating an antigen reservoir at the site of administration [62,63,64,65]. Studies, however, have shown that an antigen depot is not necessary for the adjuvanticity of alum [64], alluding to the fact that there might be other mechanisms at play.
Today, it is strongly suggested that alum has a profound effect on phagocytosis, inflammation, dendritic cell maturation, and the secretion of cytokines [63,64]. Several studies have highlighted its role in the activation of the NLRP3 inflammasome, yet the definite mode of action is still under debate, as there is no universally established standard as far as immunization protocol, type of aluminum adjuvant, and animal model are concerned [63]. Alum does not appear to have the ability to influence dendritic cells directly through TLR binding pathways, though some level of interaction has been reported via TLR-independent binding to lipid cues in the dendritic cell membrane [64]. Additionally, alum has been shown to promote the induction of endogenous danger signals and facilitate the activation of the complement system, with the primary characteristic of its adjuvant activity lying in the generation of Th2-mediated humoral immune responses as summarized by He et al.’s review [64].
The validation of these hypotheses and the elucidation of the intrinsic machinery involved in alum’s adjuvant action remains an interesting and elusive topic for researchers so far. In the context of aquatic organism immunology, the scarcity of information makes understanding the action of aluminum-based adjuvants additionally challenging; nevertheless, recent studies have shown that alum is a potent adjuvant in teleost fish, able to enhance immune responses upon vaccination. In Angosto et al.’s recent study, alum was shown to trigger the expression of the gene encoding IL-1β in both the European seabass (Dicentrarchus labrax) and gilthead seabream (Sparus aurata), matching the hypothesis that aluminum-based adjuvants can promote the secretion of inflammatory cytokines by activating the NLRP3 inflammasome [66]. Surprisingly though, no IL-1β secretion was observed in seabream leukocytes and macrophages, alluding to the fact that alum’s adjuvant action might be independent of the inflammasome in teleost fish, as further strengthened by the lack of conservation of caspase-1 processing sites in non-mammalian IL-1β, the inability of caspase-1 to modify IL-1β, and the failure of caspase-1 activation through NLRP3 inflammasome facilitators previously reported in gilthead seabream [67,68,69].
Despite the forementioned findings, alum remains a strong adjuvant in aquaculture vaccinology, able to facilitate the production of reactive oxygen species through a NADPH oxidase-mediated route [66]. Adding to the fact that alum crystals can potentially induce endogenous signals like uric acid and released DNA from dead cells [70,71,72], these results tilt towards the validation of the notion that alum’s adjuvant capacity is achieved through the induction of danger signals in the host’s body. However, the existence of side-effects can also be attributed to alum’s immunostimulating capacity, as inflammatory reactions, granulomas, melanizations, and toxicity in the splenic melanomacrophage centers have been documented with alum use in teleost fish [73].
3.3. Synthetic Adjuvants and Cytokines
Synthetically-derived adjuvants represent a unique category of adjuvants that have been facilitated by the relatively recent advent of scientific and technological innovation. In the realm of aquaculture, one of the most popular adjuvants of this type explored consists of synthetic oligodeoxynucleotides (ODNs) that express unmethylated motifs known as CpG. Such motifs are abundant in microbial DNA and can be recognized by immune cells expressing TLR9 [7], though recent reviews also portray TLR21 as a receptor [27,28]. Synthetic double-stranded polyribonucleotides called Poly I:C (Polyinosinic:polycytidylic acid) have also been used as adjuvants in aquaculture vaccinology, particularly to combat pathogens of viral origins. Their mode of action relies on simulating a viral infection, leading to the production of antiviral cytokines like type-1 interferons (IFNs), through binding to the TLR3 receptor [7]. However, as mentioned previously, recent data has shown that Poly I:C can also act as a ligand to the TLR22, TLR26, and TLR28 receptors in teleost fish, opening the possibilities for additional signaling pathways to be involved in the adjuvanticity of Poly I:C.
The exploration of fish cytokines as potential vaccine adjuvants in aquaculture was recently summarized in our previous paper [1], incorporating insights from Guo and Li’s review on the topic [74], along with findings from several recent studies highlighting their effects. Overall, recombinant cytokines and cytokine genes incorporated into vectors have shown promising results when incorporated as adjuvants into vaccine formulations. Although in its infancy, this approach has recently garnered attention, as cytokines can modulate immune responses and provide certain immunopotentiation effects during fish vaccination, without the danger of a strong generalized response and potential side effects [75]. Th0-derived cytokines like IL-1β, IL-8, IL-12, IL-15, IL-17, G-CSF, and TNF-α have been shown to have potential adjuvant activity in various teleost fish species and in combination with various pathogens and vaccine technologies, as is the case with Th1-derived IFN-α, IFN-γ, IFN-c, IL-2, and Th-2 derived IL-6 and β-chemokines [1,74,75].
Lastly, for oral vaccination, synthetic and biodegradable poly(lactic-co-glycolic) acid polymers, known simply as PLGA, have long been prevalent as vaccine carriers and potential adjuvants in teleost fish vaccination. Even though the vast majority of studies has explored PLGA microparticles as a vaccine delivery carrier through the method of antigen encapsulation, there have been a few reported publications that exhibit PLGA’s adjuvant activity as per Dalmo et al.’s review [7]. Since then, recent reports have successfully implemented PLGA-mediated delivery of DNA [76] and inactivated vaccines [77,78,79] in teleost fish immunization protocols, showcasing PLGA’s ability in conferring increased levels of protection against both viral and bacterial pathogens. In an exciting and promising new series of events, recent studies combining PLGA microparticles with cytokines [80] and CpG-ODNs [81] as adjuvants have also surfaced, demonstrating that synthetic adjuvants remain a fertile area of research in the future of aquaculture vaccinology, since these combinations show better immunization efficacy when compared to using each component individually.
3.4. Structural Microbial Components and Natural Compounds
Naturally occurring substances have similarly been explored for their adjuvant and immunostimulating capabilities in teleost fish immunization. Typically, this category encompasses structural components of microorganisms that exhibit promising adjuvant effects, since they constitute distinctive pathogen-associated molecular patterns and can act as PRR ligands for the host’s immune cells. Prime examples include lipopolysaccharides (LPS), flagellins, β-glucans, and peptidoglycans (PGN), which have been implemented as both feed immunostimulants and as adjuvants in aquaculture. These macromolecules induce the secretion of signaling cytokines and potentiate immune responses, opening the gateway for their application as both functional feed additives and vaccine adjuvants in a wide array of fish species, as evidenced by numerous publications on the topic [7,82,83,84,85,86,87]. Similarly, lipopeptides have been successfully used as vaccine adjuvants in fish, since they are abundant in many microbial species, including Mycobacteria and Mycoplasma. Polar glycopeptidolipids (pGPL) of mycobacterial origin have shown promising immune-enhancing capabilities in fish, without the presence of any adverse side-effects [88].
Besides PAMPs, however, other naturally occurring substances have exhibited promising results when used as potential aquaculture vaccine adjuvants. Saponins are a naturally occurring group of steroid or terpenoid glycosides that are typically extracted from plants. They have been shown to enhance innate immune responses in many animal species, including teleost fish and aquatic invertebrates [89]. In aquaculture vaccinology, saponins extracted from the soapbark tree (Quillaja Saponaria) have given rise to a promising vaccine adjuvant, known as Quil-A. The implementation of saponins as aquaculture vaccine adjuvants, however, has been met with adversity, as they can be toxic in high doses, and their stability is compromised in aqueous solutions [7,90]. Nevertheless, recent studies show that saponins can effectively augment immune responses, even when administered together with other adjuvants or through mucosal routes [91,92,93]. Additionally, several other plant-derived saponins, like ginseng stem leaf saponins, have successfully been reported to enhance immune responses in fish upon administration, as seen in recent studies [94,95].
The adjuvant capacity of several phytogenic substances for aquaculture vaccinology is well-described in a review recently published by Soltani et al. [96]. Furthermore, probiotics appear to gain traction as adjuvants in aquaculture vaccines, especially in oral vaccination applications dealing with encapsulation platforms. Coupled with modern genetic bioengineering techniques, probiotic bacteria like Bacilli, Lactobacilli, and Lactococci can have profound effects on fish immunization, opening the gateway for interesting vaccine delivery applications by combining the immune-stimulating effects with improved antigen delivery through the oral route [97,98,99,100,101,102,103,104,105,106]. Finally, the use of chitosan, the deacetylation product of the naturally occurring polysaccharide chitin derived from insects, fungi, and aquatic invertebrates, has been met with promise in the realm of aquaculture vaccinology, as it can be similarly used in oral routes as an immune-enhancing vehicle for vaccine delivery [107,108,109]. The adjuvant capacity of chitosan microparticles can be traced in a plethora of recent publications, both by itself, and in combination with other vaccine adjuvants or polymers [110,111,112,113,114,115,116,117,118].
4. Assessing the Side Effects of Traditional Adjuvants
Despite vaccine adjuvants being crucial components in enhancing the efficacy of aquaculture vaccines, it is well-recognized that their use can be accompanied by several side effects and drawbacks. Oil-based adjuvants are currently dominating formulations used in aquaculture as they are predominantly implemented in vaccines administered by way of intraperitoneal injection [8,119,120,121]. However, these vaccines are often associated with adverse side effects, stemming primarily from inflammatory reactions that are triggered during immunization. These reactions can exhibit varying degrees of intensity and severity, and the most typical symptoms include adhesions of tissues and organs to each other or to the peritoneal cavity, granuloma formation, and discoloration at the site of injection [120,121,122,123,124,125,126,127,128]. A schematic illustration summarizing the adverse effects of currently employed vaccine formulations in teleost fish can be seen below in Figure 3.
The Spielberg scale, developed in 1996 by Midtlyng et al., was designed to assess the severity of intraperitoneal symptoms in Atlantic salmon (Salmo salar) following immunization against furunculosis [129]. Remarkably, this scale and its underlying methodology remain valid and therefore prevalent in international literature, and are particularly referenced in studies examining the side effects of vaccines in teleost fish of similar species. In this framework, individual fish are evaluated based on the severity of their symptoms and the associated pathological changes post-vaccination, by implementing a scale that typically ranges from 0 to 6; a higher score indicates more pronounced post-vaccination pathological findings, offering a tool for establishing a standardized approach in evaluating the impact of adjuvanted vaccines in teleost fish health. Recently, in 2021, Tziouvas & Varvarigos adapted a Spielberg-type scale to the European sea bass (Dicentrarchus labrax) [128]; a side-by-side adaptation comparison of the two scales can be seen in Table 3.
This study involved the collection of data from fish growing in Greek aquaculture sites that were vaccinated through intraperitoneal injection with licensed oil-adjuvanted vaccines over a three-year period (2016-2018) and the meticulous evaluation of the pathological findings found within the peritoneal cavity. As expected, many of the sampled fish exhibited the side effects described in literature, such as chronic peritonitis, granulomatous lesions, discolorations, and tissue and organ adhesions. Notably, smaller-sized fish appeared to experience more severe side effects following vaccination, potentially due to the challenges associated with the administration method of IP injection, which requires careful handling and execution. Furthermore, vaccines formulated with mineral oil adjuvants that are not easily metabolized by fish, such as paraffin oils, resulted in more pronounced symptoms associated with intense inflammatory responses. The feed conversion rate (FCR) remained largely unaffected in the aquaculture sites, except in cases where there were severe granulomatous lesions and strong tissue adhesions hindering the physiological functioning of the digestive system.
The findings from the previous study, combined with simple macroscopic and histological analyses conducted during the autopsies, led to the development of a scale similar to that of Midtlyng et al. to estimate the severity of symptoms post-vaccination. The newly proposed six-point scale can potentially serve as a valuable tool for aquaculture industry professionals and scientist alike to assess the welfare of teleost fish and evaluate relevant side effects of oil-adjuvanted vaccines in aquaculture. This advancement holds particular significance for marine finfish and European aquaculture where species like the European sea bass (Dicentrarchus labrax) and the gilthead seabream (Sparus aurata), two of the most economically important species within the Mediterranean region, are routinely vaccinated with commercial vaccines. Simultaneously, this work also lays the groundwork for further research to be conducted into the implications of contemporary vaccines for farmed fish, expanding the scope to include various factors such as teleost fish species, type of vaccine and adjuvant used, growth rates, final body weight, and the quality of the final products. In conclusion, this showcases the necessity for similar studies to be carried out in order to fully understand the impact of current aquaculture vaccines on aquatic welfare and health. As such, the development of species-specific and tailored scales is essential for enhancing sustainability and promoting humane practices in the sector.
5. The Promise of Nanoparticle-Based Formulations for the Future
The association between traditional vaccine adjuvants and the adverse post-vaccination side effects on the welfare and health of teleost fish has prompted researchers to explore novel and alternative options, with nanoparticle-based formulations gaining significant traction as the future of this field. As technological expertise progresses, these innovative formulations pave the way for increased vaccination efficacy as novel adjuvants, with the capacity to minimize relevant side effects and adverse reactions. In this section, recent advances in the field of nanoparticle-based adjuvants for aquaculture vaccines will be reviewed, highlighting their position as a potential avenue for ongoing research, where the crossroads of aquatic immunology, aquaculture health management, and technological innovation are met.
5.1. Polymeric Nanoparticle Formulations
As already mentioned, polymers represent a significant portion of current nanoparticle-based research attempts focused on vaccine delivery in aquaculture. The shift in scale from microparticles to nanoparticles can offer advantages in immunization enhancement, as smaller-sized particles can activate the immune system more efficiently, triggering both cellular and humoral immune responses in teleost fish. In addition, nanoformulations seem to improve crucial vaccine factors, such as solubility, stability, targeting, permeability, and biocompatibility [130].
PLGA-based nanoparticle applications in aquaculture vaccinology have maintained a prominent presence in international literature over the past decade, as seen in some of earlier publications [131,132,133,134]. Focusing on more recent studies, PLGA nanoparticles have been utilized for immunization against a diverse range of aquatic pathogens, employing various vaccination technologies and administration methods, while targeting numerous teleost fish species. Dubey et al. have demonstrated that the oral administration of recombinant outer membrane protein W (OmpW) of Aeromonas hydrophila encapsulated in PLGA nanoparticles shows a dose-dependent protective effect in rohu fish (Labeo rohita) [135]. Since then, Harshita et al. have also recently shown that the recombinant OmpA can similarly be encapsulated in PLGA nanoparticles and be orally delivered to confer protection in Aeromonas hydrophila challenged zebrafish (Danio rerio) [136]. The compatibility of PLGA nanoparticle-based vaccine formulations has also been tested with DNA vaccines against Aeromonas hydrophila, both in oral and injectable applications, as seen in the recent works by Alishahi et al. [137,138]. In the case of viral pathogens, a very early first attempt of implementing a PLGA nanoparticle-based oral DNA vaccine against infectious haematopoietic necrosis was made in Adomako’s study [133], however recent endeavors have expanded towards more viruses, such as the spring viremia of carp virus (SVCV) and the hemorrhagic septicemia virus (HSV). In a study published in 2019, the efficacy of an inactivated viral vaccine against HSV encapsulated in PLGA nanoparticles was demonstrated by Kole et al. [139], whereas in 2022, Zhang et al. successfully developed a dual-targeting polymer containing PLGA and chitosan nanoparticles to encapsulate a DNA vaccine that was able to confer robust protective immunity in common carp (Cyprinus carpio) [140].
The combination of polymers with chitosan nanoparticles as vaccine adjuvants is not coincidental, as chitosan is non-toxic, biodegradable, and has potent mucoadhesive properties that can aid in eliciting strong protective effects upon mucosal immunization in fish [141,142,143]. Several studies combining chitosan nanoparticles with other polymers like alginate and PLGA for various vaccination applications in teleost fish have been published in recent years [144,145,146,147,148,149]. Apart from those, chitosan-complexed nano-vaccines have been implemented to counter both bacterial aquatic diseases [150,151,152,153,154,155,156,157,158,159,160,161,162] and viral ones [163,164,165,166,167,168] in a plethora of new studies. Additionally, novel technologies like chitosan nanoparticles complexed with oxygen nanobubbles have emerged, facilitating the enhanced uptake of vaccines and resulting in superior mucosal immune responses [169]. The synergy of chitosan nanoparticles with traditional adjuvants has also been studied, highlighting that the combination of aluminum adjuvants with chitosan-based nanovaccines can trigger both local and systemic immune responses [170], while on the contrary, Poly I:C’s adjuvanticity appears to deteriorate, presumably due to the delayed, gradual, and stable antigen release that the chitosan particles provide on the microscale [112].
Nevertheless, Collado-González & Esteban highlighted that the inconsistencies in the characterization of the physicochemical properties of chitosan particles, like the degree of acetylation (DA), the molecular weight (MW), and the different preparation methods used have made it difficult to determine which specific characteristics are most effective in stimulating the teleost fish immune system [171]. This underscores the need for standardizing chitosan nanoparticle-based treatments and improving reporting practices to enhance data comparability and reproducibility. Overall, studies that properly characterized chitosan suggest that low DA (10-15%) and a MW ranging from 50 to 800 kDa may enhance immune gene expression in teleost fish, though the differing amounts of chitosan used, and the different immunization regimens followed in different species make direct comparisons difficult at this stage. Collectively, the implementation of polymeric nanoparticle-based vaccine formulations in teleost fish appears promising so far and there is a pressing need for more studies to explore and correctly assess the advantages of the shift from micro to nano-scale polymeric materials in aquaculture vaccinology.
5.2. Lipid-Based Nanoparticle Formulations
Another type of nanoparticle-based formulation that has gained traction for vaccine delivery and the enhancement of immune responses encompasses lipid nanoparticles, either through nanoliposomes or by using cationic lipid surfactants. These substances typically aid in the stabilization of antigens and can serve as interesting tools for the development of nano-formulations in a wide array of vaccine technologies. Liposomes consist of a hydrophilic core surrounded by hydrophobic bilayers and can be used to encapsulate both hydrophilic and lipophilic compounds [172]. This offers advantages in enhanced solubility, bioavailability, and antigen stability, offering a solid delivery platform that could potentially reinforce immune responses. Alternatively, surfactants like cetrimonium bromide, known as CTAB, are currently being explored for the construction of cationic lipid-based vaccines due to their ability to modify nanoparticle size and surfaces. Additionally, they can alter vaccine charges, therefore opening the gateways for improved delivery, especially through mucosal routes [173,174].
The use of such technologies at the nano scale are not extensively studied in the field of aquaculture vaccinology, however there are a few studies that have delved into the use of lipid-based nanoparticle vaccine formulations. In 2019, Ji et al. published a study where nanoliposomes loaded with bacterial LPS and Poly I:C were successfully used to enhance innate immune responses in zebrafish larvae, leading to the upregulation of several immune-related genes and an increase in survival post Aeromonas hydrophila challenge following bath immersion vaccination [172]. These results have been succeeded by similar studies, where immunostimulant-loaded nanoliposomes were tested on adult zebrafish, resulting in protection against Pseudomonas aeruginosa and SVCV infection both in injection and in immersion applications [175,176]. These results also highlight the recent shift in size from liposome to nanoliposome formulations in teleost fish and pave the way for fish fry vaccination applications by lifting size limitations and improving antigen delivery [172]. A recent development was also made in 2024, where an mRNA-lipid nanoparticle vaccine formulation was successfully applied in the Atlantic salmon (Salmo salar), demonstrating that the relatively novel mRNA vaccine technology encapsulated in lipid nanoparticles can indeed be used to express antigens in teleost fish, both in vitro and in vivo [177].
Cationic lipid-based nanovaccines represent the other major part of this category as far as nanoparticle-based formulations in aquaculture vaccinology are concerned. Despite the lack of reports on teleost fish applications, recent years have witnessed the emergence of a few studies implementing this technology to develop mucoadhesive immersion nano- vaccines against bacterial pathogens. In Thangsunnan et al.’s published study, a mucoadhesive nanovaccine against Francisella noatunensis subsp. orientalis was successfully constructed using CTAB as a surfactant [174]. This formulation conferred protection comparable to that of a whole-cell inactivated vaccine, inducing strong immune responses after boost immunization. Gene expression analysis revealed significant upregulation of both IgM and IgT in vaccinated fish, confirming enhanced mucosal and systemic responses in red tilapia (Oreochromis sp.). Similarly, Bunnoy et al. developed a vaccine of similar technology to combat columnaris disease [175]. The nanovaccine effectively targeted the mucosal-associated lymphoid tissues (MALT) in the skin and gills of the Asian seabass (Lates calcarifer), resulting in a strong immune response, where vaccinated fish showed higher antibody levels and survival rates (65.83-72.50%) compared to the controls. This formulation stimulated both innate and adaptive immunity, resulting in increased expression of immune-related genes like IgM and MHC-IIα. However, while effective, the vaccine was linked with early mortality, indicating that further optimization is needed in vaccination protocols.
5.3. Carbon Nanotubes and Inorganic Nanoparticle Formulations
Lastly, carbon nanotubes (CNTs) represent a cutting-edge paradigm with regard to nanoparticle-based vaccine formulations for teleost fish, offering an alternative tool to enhance the delivery and immunogenicity of aquaculture vaccines, particularly through mucosal routes. Carbon nanotubes are comprised of hollow, cylindrical structures made from rolled up graphene sheets. These nanotubes can either include a single, or many graphene layers, making the distinction between single-walled carbon nanotubes (SWCNTs) and multi-walled carbon nanotubes (MWCNTs) [178].
Carbon nanotubes are considered inert and not immunogenic by themselves, having relatively low toxicity and high biocompatibility [179]. A recent study conducted by Cimbaluk et al. demonstrated that this nanomaterial does not induce any genotoxic effects like single or double DNA strand breaks in teleost fish erythrocytes. However, CNT-DNA crosslinks were observed, alluding to CNTs preventing the separation of DNA strands and potentially compromising the process of replication [180]. Chronic exposure to this material has indeed been linked to the induction of oxidative stress, pollutant accumulation, neurotoxicity, inflammation, apoptotic signaling, and histological pathologies in fish [180,181,182,183,184,185,186,187,188]. Nonetheless, the relevance of these studies to vaccination regimens remains to be elucidated and warrants further research to be conducted, especially since the physicochemical properties of the CNTs appear to make a difference in their toxicity according to Jiang et al. [189]. The modification of CNTs with other polymers also appears to lower toxicity claims, as is the case with chitosan grafting in Wisdom et al.’s study [190], which underpins the possibility of an ideal formulation to be discovered soon through the combination of nanopolymeric materials.
The functionalization of CNTs for immunization applications in teleost fish typically shows reduced toxicity and can have profound effects on the stimulation of immune cells [191]. During the past decade, several studies have accumulated in international literature dealing with CNT-based vaccination in a wide range of species, yet the focus seems to be particularly set in immunization applications against viral aquatic diseases. Nevertheless, there have been a few studies showcasing that CNTs can be used to combat bacterial diseases as well, with Aeromonas hydrophila centered as the main pathogen of interest. In 2015, Gong et al. developed a novel formulation using SWCNTs to deliver a recombinant vaccine against the pathogen in juvenile grass carp (Ctenopharyngodon idellus) [192]. The vaccine, administered via both injection and immersion, led to a higher antibody production and significant upregulation of immune markers, achieving similar survival rates to traditional injection methods. A year later, the same group published another study, highlighting the upregulation of immune-related genes which was induced by their SWCNT-based vaccine compared to a SWCNT-free control [193]. More recent efforts were reported in 2020, with Zhang et al. successfully demonstrating the immunoprotective effect of a whole-cell lysed inactivated vaccine utilizing SWCNTs as a carrier [194]. In 2024, Cao et al. shifted the focus by publishing an interesting study in which they constructed a CNT nanocarrier immersion vaccine encoding the immunogenic recombinant protein Sip [195]. This vaccine formulation promoted the expression of immune genes, induced relatively high serum antibody production, and enhanced related enzyme activities, ultimately conferring significant cross-protection against Streptococcus agalactiae and Streptococcus iniae in red tilapia.
As previously mentioned, however, the main body of work revolving around CNT-based vaccination in teleost fish stems primarily from applications dealing with viral diseases. The grass carp reovirus (GCRV) constitutes a prime example, since there are several publications documenting the efficacy of CNT-based nanovaccines in a plethora of different vaccine technologies, such as recombinant subunit vaccines [196,197,198] and DNA vaccines [199,200,201]. Another grass carp virus, the spring viraemia carp virus (SVCV) has similarly garnered the attention of CNT-based vaccination applications, as evidenced by recent studies [202,203,204,205,206]. Zhao et al. have successfully constructed and demonstrated the effect of a SWCNT-based subunit vaccine against the infectious spleen and kidney necrosis virus (ISKNV) in mandarin fish (Siniperca chuatsi) [207,208] and the same strategy was implemented for a DNA vaccine [209]. The most recent publications from this group are currently showcasing the efficacy of a mannose-modified CNT subunit vaccine designed to target antigen presenting cells both in injection and in immersion formats [210,211]. Apart from ISKNV, CNT-based vaccination has been reported to be efficient against another iridovirus, named the grouper iridovirus of Taiwan (TGIV) [212,213], as well as other viral families, such as herpesviruses [214,215], rhabdoviruses [216], betanodaviruses [217], and the recently classified largemouth bass ulcerative syndrome ranavirus [218]. An indicative table (Table 4) summarizing the recent studies of CNT-based vaccination studies in teleost fish mentioned can be read below:
Staying within the context of nanotube formulations, it is worth noting that an inorganic alternative of CNTs has recently emerged in the realm of aquaculture vaccines, offering an alternative to its organic relative and addressing the gap in literature regarding the use of inorganic nanoparticle-based formulations in teleost fish [219]. Halloysite nanotubes (HNTs) are naturally occurring tubular clay structures similar to CNTs, featuring a unique disparity in charge between their inner and outer surfaces [220]. They provide an environmentally safe and cost-efficient platform for vaccine delivery throughout the gastrointestinal tract, allowing for the encapsulation of susceptible antigens and their subsequent release in a pH-dependent manner, as well as for their modification with other materials to enhance their properties [220,221].
Pumchan et al. have recently harnessed the power of HNTs to test out and develop a novel oral nanovaccine delivery system to combat streptococcosis in tilapia (Oreochromis sp.) [221]; remarkably, they found that chitosan-modified HNTs demonstrated strong resilience to stomach acidity, bypassing gastric digestion and delivering a whole-cell inactivated vaccine to the intestines that promoted an immune response via the gut-associated lymphoid tissues (GALT). This formulation effectively induced immunity against Streptococcus agalactiae serotype Ia, exhibiting similar disease protection and survival rates when compared to traditional injectable vaccines. The chitosan coating of HNTs enhanced the immune response and adherence to intestinal cells, positioning this nanoparticle-based formulation as a promising candidate for efficacious oral vaccination. Taken together with Zhang et al.’s recent publication on the use of a mesoporous silica nanoparticle-based delivery system for oral vaccination against Vibrio alginolyticus [222], these few studies highlight the use of inorganic nanoparticles in teleost fish immunization, ultimately completing the puzzle of a much promising arsenal for the future of adjuvant research in aquaculture vaccinology.
6. Conclusions
Adjuvants are quintessential elements of modern aquaculture vaccines designed for teleost fish, as they aid in inducing potent and long-lasting immune responses that can protect reared species against aquatic diseases. Progresses made in understanding the exact mechanisms of action behind various adjuvants have facilitated the development of many innovative and novel vaccine formulations, designed specifically to induce desired immunization outcomes. Combined with recent insights into aquatic and teleost fish immunology, these advancements have enabled the introduction of progressively more sophisticated vaccine and adjuvant technologies to be implemented in aquaculture settings, indicating that the future of vaccinology research and vaccine development in one of the fastest growing sectors is an excellent field for investment and exploration.
Despite vaccine adjuvants forming an integral part of aquaculture vaccines, there has been growing concerns about their potential adverse effects in teleost fish health, making the question for product quality assurance and associated welfare issues in modern aquaculture. The use of contemporary vaccine adjuvants, such as oil-based emulsions, has been linked with side effects like tissue and organ adhesions, inflammatory reactions, granulomas, and spatial discolorations at the sites of injections, since intraperitoneal injection is currently the most widely used method of vaccine administration in the field. These findings have led researchers and aquaculture professionals alike to explore alternative ways of teleost fish immunization, in an effort to minimize cost, labor, and potential side effects, while maximizing the immunization outcomes. Apart from the shift in the administration route, as current trends dictate, the exploration of novel adjuvants offers another backroad for circumnavigating the limitations of current aquaculture vaccines, providing cutting-edge solutions to persisting issues, such as impotent immunization and compromised fish welfare. Taken together, these two elements can significantly improve the sustainability of the aquaculture industry and facilitate the establishment of more welfare-friendly practices that will ensure the quality of aquatic food products in accordance with the consumer demands.
Currently, the spearhead of such attempts, as far as modern innovation is concerned, lies in utilization of nanoparticle-based formulations as a means of replacing or improving the traditional vaccine adjuvants in aquaculture vaccinology. By harnessing the power of technological advances and biological knowledge, the utilization of nanoparticles can enhance vaccine delivery, potentiate immune responses, and ultimately confer superior protective effects in reared species, improving vaccine formulations and mitigating relative side effects linked to conventional vaccine adjuvants. Polymeric nanoparticles, lipid-based nanoformulations, and nanomaterials of organic or inorganic origin are at the forefront of current research endeavors employing nanoparticle-based formulations for immunization purposes in teleost fish. However, even though they appear promising, these applications are still considered to be in their relative infancy, as most of these technologies have only been tested in fish for no more than a decade. Ultimately, this warrants for further research efforts to be put into assessing the application of these materials, so that concrete and relative evidence will be accumulated on their efficacy, optimization, and safety in aquatic species.
Overall, the assessment of animal safety and welfare should be placed as a top priority in trials exploring novel formulations, as this can potentially ensure faster application approval and the establishment of welfare-friendly vaccination practices in the sector. The need for tools and attention to effectively monitor and evaluate teleost fish welfare is pressing, especially in the realm of aquaculture immunization, as many different technologies, employing various administration routes, and in several species are currently being utilized globally, in a plethora of aquaculture sites. A series of properly standardized technology- and species-specific guidelines can potentially help in establishing a framework where vaccine adjuvants and their effects can be tracked, compared, and evaluated on an evidential basis, in an effort to accelerate vaccine development and sort out potential candidates for novel vaccine formulations. By striking a balance between vaccination efficacy and teleost fish welfare, the next generation of aquaculture vaccines will be probably soon available in the mass market, ushering in a new era for the future of the aquaculture industry.
Author Contributions
Conceptualization, I.T., K.B., and A.G.; methodology, I.T., K.B., and A.G.; software, I.T.; validation, I.T., K.B., and A.G.; formal analysis, I.T.; investigation, I.T., K.B. and A.G.; resources, I.T. and A.G.; data curation, I.T., K.B., and A.G.; writing—original draft preparation, I.T.; writing—review and editing, K.B. and A.G.; visualization, I.T., K.B., and A.G.; supervision, K.B. and A.G.; project administration, K.B. and A.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
All of the figures in this manuscript were created using BioRender.com: (Figure 1: Created in BioRender. Tammas, I. (2024) https://BioRender.com/s47j443, Figure 2: Created in BioRender. Tammas, I. (2024) https://BioRender.com/n42u649, Figure 3: Created in BioRender. Tammas, I. (2024) https://BioRender.com/l57y747).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tammas, I.; Bitchava, K.; Gelasakis, A.I. Transforming Aquaculture through Vaccination: A Review on Recent Developments and Milestones. Vaccines 2024, 12, 732. [Google Scholar] [CrossRef] [PubMed]
- Schar, D.; Klein, E.Y.; Laxminarayan, R.; Gilbert, M.; Van Boeckel, T.P. Global Trends in Antimicrobial Use in Aquaculture. Sci Rep 2020, 10, 21878. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Gauthier, J.; Derome, N.; Charette, S.J. The Rise and Fall of Antibiotics in Aquaculture. In Microbial Communities in Aquaculture Ecosystems: Improving Productivity and Sustainability; Derome, N., Ed.; Springer International Publishing: Cham, 2019; pp. 1–19. ISBN 978-3-030-16190-3. [Google Scholar]
- Arechavala-Lopez, P.; Sanchez-Jerez, P.; Bayle-Sempere, J.T.; Uglem, I.; Mladineo, I. Reared Fish, Farmed Escapees and Wild Fish Stocks—a Triangle of Pathogen Transmission of Concern to Mediterranean Aquaculture Management. Aquaculture Environment Interactions 2013, 3, 153–161. [Google Scholar] [CrossRef]
- Mordecai, G.J.; Miller, K.M.; Bass, A.L.; Bateman, A.W.; Teffer, A.K.; Caleta, J.M.; Di Cicco, E.; Schulze, A.D.; Kaukinen, K.H.; Li, S.; et al. Aquaculture Mediates Global Transmission of a Viral Pathogen to Wild Salmon. Science Advances 2021, 7, eabe2592. [Google Scholar] [CrossRef] [PubMed]
- Johansen, L.-H.; Jensen, I.; Mikkelsen, H.; Bjørn, P.-A.; Jansen, P.A.; Bergh, Ø. Disease Interaction and Pathogens Exchange between Wild and Farmed Fish Populations with Special Reference to Norway. Aquaculture 2011, 315, 167–186. [Google Scholar] [CrossRef]
- Dalmo, R.; Bøgwald, J.; Tafalla, C. Adjuvants and Delivery Methods: Current and Novel. In Fish Vaccines; Adams, A., Ed.; Springer: Basel, 2016; pp. 75–103. ISBN 978-3-0348-0980-1. [Google Scholar]
- Mondal, H.; Thomas, J. A Review on the Recent Advances and Application of Vaccines against Fish Pathogens in Aquaculture. Aquacult Int 2022, 30, 1971–2000. [Google Scholar] [CrossRef]
- Subramani, P.A.; Michael, R.D. Chapter 4 - Prophylactic and Prevention Methods Against Diseases in Aquaculture. In Fish Diseases; Jeney, G., Ed.; Academic Press, 2017; pp. 81–117. ISBN 978-0-12-804564-0. [Google Scholar]
- Nasr-Eldahan, S.; Nabil-Adam, A.; Shreadah, M.A.; Maher, A.M.; El-Sayed Ali, T. A Review Article on Nanotechnology in Aquaculture Sustainability as a Novel Tool in Fish Disease Control. Aquacult Int 2021, 29, 1459–1480. [Google Scholar] [CrossRef]
- Shah, B.R.; Mraz, J. Advances in Nanotechnology for Sustainable Aquaculture and Fisheries. Reviews in Aquaculture 2020, 12, 925–942. [Google Scholar] [CrossRef]
- Sarkar, B.; Mahanty, A.; Gupta, S.K.; Choudhury, A.R.; Daware, A.; Bhattacharjee, S. Nanotechnology: A next-Generation Tool for Sustainable Aquaculture. Aquaculture 2022, 546, 737330. [Google Scholar] [CrossRef]
- Thompson, K.D.; Rodkhum, C.; Bunnoy, A.; Thangsunan, P.; Kitiyodom, S.; Sukkarun, P.; Yostawornkul, J.; Yata, T.; Pirarat, N. Addressing Nanovaccine Strategies for Tilapia. Vaccines 2023, 11, 1356. [Google Scholar] [CrossRef]
- Radhakrishnan, A.; Vaseeharan, B.; Ramasamy, P.; Jeyachandran, S. Oral Vaccination for Sustainable Disease Prevention in Aquaculture—an Encapsulation Approach. Aquacult Int 2023, 31, 867–891. [Google Scholar] [CrossRef] [PubMed]
- Turley, J.L.; Lavelle, E.C. Resolving Adjuvant Mode of Action to Enhance Vaccine Efficacy. Current Opinion in Immunology 2022, 77, 102229. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.M.S.; Schijns, V.E.J.C. Immunology of Vaccine Adjuvants. In Vaccine Adjuvants: Methods and Protocols; Davies, G., Ed.; Humana Press: Totowa, NJ, USA, 2010; pp. 1–14. ISBN 978-1-60761-585-9. [Google Scholar]
- Schijns, V.E. Immunological Concepts of Vaccine Adjuvant Activity: Commentary. Current Opinion in Immunology 2000, 12, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Qin, Z.; Liu, H.; Lin, L.; Ye, J.; Li, J. Recent Advances on Phagocytic B Cells in Teleost Fish. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C.J.; Belmonte, R. Overview of the Fish Adaptive Immune System. In Fish Vaccines; Adams, A., Ed.; Springer Basel: Basel, 2016; pp. 35–52. ISBN 978-3-0348-0978-8. [Google Scholar]
- Kordon, A.O.; Pinchuk, L.; Karsi, A. Adaptive Immune System in Fish. Turkish Journal of Fisheries and Aquatic Sciences 2021, 22. [Google Scholar] [CrossRef]
- Díaz-Rosales, P.; Muñoz-Atienza, E.; Tafalla, C. Role of Teleost B Cells in Viral Immunity. Fish & Shellfish Immunology 2019, 86, 135–142. [Google Scholar] [CrossRef]
- Johnstone, C.; Chaves-Pozo, E. Antigen Presentation and Autophagy in Teleost Adaptive Immunity. International Journal of Molecular Sciences 2022, 23, 4899. [Google Scholar] [CrossRef]
- Zinkernagel, R.M.; Ehl, S.; Aichele, P.; Oehen, S.; Kündig, T.; Hengartner, H. Antigen Localisation Regulates Immune Responses in a Dose- and Time-Dependent Fashion: A Geographical View of Immune Reactivity. Immunological Reviews 1997, 156, 199–209. [Google Scholar] [CrossRef]
- Awate, S.; Babiuk, L.A.; Mutwiri, G. Mechanisms of Action of Adjuvants. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef]
- Kordon, A.O.; Karsi, A.; Pinchuk, L. Innate Immune Responses in Fish: Antigen Presenting Cells and Professional Phagocytes. Turkish Journal of Fisheries and Aquatic Sciences 2018, 18, 1123–1139. [Google Scholar] [CrossRef]
- Sahoo, B.R. Structure of Fish Toll-like Receptors (TLR) and NOD-like Receptors (NLR). International Journal of Biological Macromolecules 2020, 161, 1602–1617. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Su, J. Progresses on Three Pattern Recognition Receptor Families (TLRs, RLRs and NLRs) in Teleost. Developmental & Comparative Immunology 2021, 122, 104131. [Google Scholar] [CrossRef]
- Zhang, J.; Kong, X.; Zhou, C.; Li, L.; Nie, G.; Li, X. Toll-like Receptor Recognition of Bacteria in Fish: Ligand Specificity and Signal Pathways. Fish & Shellfish Immunology 2014, 41, 380–388. [Google Scholar] [CrossRef]
- Sahoo, B.R. Structure of Fish Toll-like Receptors (TLR) and NOD-like Receptors (NLR). International Journal of Biological Macromolecules 2020, 161, 1602–1617. [Google Scholar] [CrossRef]
- Ferreira, I.A.; Peixoto, D.; Losada, A.P.; Quiroga, M.I.; do Vale, A.; Costas, B. Early Innate Immune Responses in European Sea Bass (Dicentrarchus Labrax L.) Following Tenacibaculum Maritimum Infection. Front. Immunol. 2023, 14. [Google Scholar] [CrossRef]
- Leiva-Rebollo, R.; Gémez-Mata, J.; Castro, D.; Borrego, J.J.; Labella, A.M. Immune Response of DNA Vaccinated-Gilthead Seabream (Sparus Aurata) against LCDV-Sa Infection: Relevance of the Inflammatory Process. Front. Immunol. 2023, 14. [Google Scholar] [CrossRef]
- Franch, R.; Cardazzo, B.; Antonello, J.; Castagnaro, M.; Patarnello, T.; Bargelloni, L. Full-Length Sequence and Expression Analysis of Toll-like Receptor 9 in the Gilthead Seabream (Sparus Aurata L.). Gene 2006, 378, 42–51. [Google Scholar] [CrossRef]
- Miccoli, A.; Buonocore, F.; Picchietti, S.; Scapigliati, G. The Sea Bass Dicentrarchus Labrax as a Marine Model Species in Immunology: Insights from Basic and Applied Research. Aquaculture and Fisheries 2024, 9, 136–143. [Google Scholar] [CrossRef]
- Muñoz, I.; Sepulcre, M.P.; Meseguer, J.; Mulero, V. Toll-like Receptor 22 of Gilthead Seabream, Sparus Aurata: Molecular Cloning, Expression Profiles and Post-Transcriptional Regulation. Developmental & Comparative Immunology 2014, 44, 173–179. [Google Scholar] [CrossRef]
- Chen, Z.; Ceballos-Francisco, D.; Guardiola, F.A.; Huang, D.; Esteban, M.Á. The Alleviation of Skin Wound-Induced Intestinal Barrier Dysfunction via Modulation of TLR Signalling Using Arginine in Gilthead Seabream (Sparus Aurata L). Fish & Shellfish Immunology 2020, 107, 519–528. [Google Scholar] [CrossRef]
- Wang, B.; Liu, X.; Zhao, J.; Cao, M.; Yu, Z.; Fu, Q.; Tan, F.; Yang, N.; Li, C. Characterization, Evolution and Expression Analysis of Toll-like Receptor 7 (TLR7) in Turbot (Scophthalmus Maximus L.). Fish & Shellfish Immunology 2022, 125, 9–16. [Google Scholar] [CrossRef]
- Li, S.; Wang, G.; Liu, D.; Liu, Q.; Hu, G. Cloning and Expression Analysis of a Toll-like Receptor 21 (TLR21) Gene from Turbot, Scophthalmus Maximus. Developmental & Comparative Immunology 2017, 73, 163–168. [Google Scholar] [CrossRef]
- Zhang, H.; Hu, G.; Liu, Q.; Zhang, S. Cloning and Expression Study of a Toll-like Receptor 2 (Tlr2) Gene from Turbot, Scophthalmus Maximus. Fish & Shellfish Immunology 2016, 59, 137–148. [Google Scholar] [CrossRef]
- Dong, X.; Su, B.; Zhou, S.; Shang, M.; Yan, H.; Liu, F.; Gao, C.; Tan, F.; Li, C. Identification and Expression Analysis of Toll-like Receptor Genes (TLR8 and TLR9) in Mucosal Tissues of Turbot (Scophthalmus Maximus L.) Following Bacterial Challenge. Fish & Shellfish Immunology 2016, 58, 309–317. [Google Scholar] [CrossRef]
- Yu, J.; Liu, X.; Yang, N.; Wang, B.; Su, B.; Fu, Q.; Zhang, M.; Tan, F.; Li, C. Characterization of Toll-like Receptor 1 (TLR1) in Turbot (Scophthalmus Maximus L.). Fish & Shellfish Immunology 2021, 115, 27–34. [Google Scholar] [CrossRef]
- Hu, G.-B.; Zhang, S.-F.; Yang, X.; Liu, D.-H.; Liu, Q.-M.; Zhang, S.-C. Cloning and Expression Analysis of a Toll-like Receptor 22 (Tlr22) Gene from Turbot, Scophthalmus Maximus. Fish & Shellfish Immunology 2015, 44, 399–409. [Google Scholar] [CrossRef]
- Liu, F.; Su, B.; Fu, Q.; Shang, M.; Gao, C.; Tan, F.; Li, C. Identification, Characterization and Expression Analysis of TLR5 in the Mucosal Tissues of Turbot (Scophthalmus Maximus L.) Following Bacterial Challenge. Fish & Shellfish Immunology 2017, 68, 272–279. [Google Scholar] [CrossRef]
- Matzinger, P. Tolerance, Danger, and the Extended Family. Annual Review of Immunology 1994, 12, 991–1045. [Google Scholar] [CrossRef]
- Tanekhy, M. The Role of Toll-like Receptors in Innate Immunity and Infectious Diseases of Teleost. Aquaculture Research 2016, 47, 1369–1391. [Google Scholar] [CrossRef]
- Savelkoul, H.F.J.; Ferro, V.A.; Strioga, M.M.; Schijns, V.E.J.C. Choice and Design of Adjuvants for Parenteral and Mucosal Vaccines. Vaccines 2015, 3, 148–171. [Google Scholar] [CrossRef]
- Salinas, I.; Ding, Y.; Fernández-Montero, Á.; Sunyer, J.O. Mucosal Immunity in Fish. In Principles of Fish Immunology: From Cells and Molecules to Host Protection; Buchmann, K., Secombes, C.J., Eds.; Springer International Publishing: Cham, 2022; pp. 387–443. ISBN 978-3-030-85420-1. [Google Scholar]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The Mucosal Immune System of Fish: The Evolution of Tolerating Commensals While Fighting Pathogens. Fish & Shellfish Immunology 2013, 35, 1729–1739. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, Q.; Huang, Z.; Ding, L.; Xu, Z. Immunoglobulins, Mucosal Immunity and Vaccination in Teleost Fish. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Villumsen, K.R.; Koppang, E.O.; Christensen, D.; Bojesen, A.M. Alternatives to Mineral Oil Adjuvants in Vaccines against Aeromonas Salmonicida Subsp. Salmonicida in Rainbow Trout Offer Reductions in Adverse Effects. Sci Rep 2017, 7, 5930. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, H.; Han, Y.; Cui, Y.; Han, X. Study on the Relationships between the Oil HLB Value and Emulsion Stabilization. RSC Advances 2023, 13, 24692–24698. [Google Scholar] [CrossRef] [PubMed]
- Hong, I.K.; Kim, S.I.; Lee, S.B. Effects of HLB Value on Oil-in-Water Emulsions: Droplet Size, Rheological Behavior, Zeta-Potential, and Creaming Index. Journal of Industrial and Engineering Chemistry 2018, 67, 123–131. [Google Scholar] [CrossRef]
- Schmidts, T.; Dobler, D.; Guldan, A.-C.; Paulus, N.; Runkel, F. Multiple W/O/W Emulsions—Using the Required HLB for Emulsifier Evaluation. Colloids and Surfaces A: Physicochemical and Engineering Aspects 2010, 372, 48–54. [Google Scholar] [CrossRef]
- Raman, R.P.; Kumar, S. Adjuvants for Fish Vaccines. In Fish immune system and vaccines; M., M., K.V., R., Eds.; Springer Nature: Singapore, 2022; pp. 231–244. ISBN 978-981-19126-8-9. [Google Scholar]
- Tepparin, S.; Unajak, S.; Hirono, I.; Kondo, H.; Areechon, N. Efficacy of Adjuvanted Streptococcus Agalactiae Vaccine by Montanide ISA 763 A VG in Nile Tilapia (Oreochromis Niloticus Linn.). Journal of Fisheries and Environment 2018, 42, 26–38. [Google Scholar]
- Soltani, M.; Mokhtari, A.; Mirzargar, S.S.; Taherimirghaed, A.; Zargar, A.; Shafiei, S.; Hosseini Shekarabi, S.P. Efficacy and Immune Response of Intraperitoneal Vaccination of Rainbow Trout (Oncorhynchus Mykiss) with a Yersinia Ruckeri Bacterin Formulated with Montanide (TM) ISA 763 AVG Adjuvant; 2016; p. 236.
- Xu, W.; Jiao, C.; Bao, P.; Liu, Q.; Wang, P.; Zhang, R.; Liu, X.; Zhang, Y. Efficacy of MontanideTM ISA 763 A VG as Aquatic Adjuvant Administrated with an Inactivated Vibrio Harveyi Vaccine in Turbot (Scophthalmus Maximus L.). Fish & Shellfish Immunology 2019, 84, 56–61. [Google Scholar] [CrossRef]
- Jaafar, R.M.; Chettri, J.K.; Dalsgaard, I.; Al-Jubury, A.; Kania, P.W.; Skov, J.; Buchmann, K. Effects of Adjuvant MontanideTM ISA 763 A VG in Rainbow Trout Injection Vaccinated against Yersinia Ruckeri. Fish & Shellfish Immunology 2015, 47, 797–806. [Google Scholar] [CrossRef]
- Wangkahart, E.; Thongsrisuk, A.; Vialle, R.; Pholchamat, S.; Sunthamala, P.; Phudkliang, J.; Srisapoome, P.; Wang, T.; Secombes, C.J. Comparative Study of the Effects of MontanideTM ISA 763A VG and ISA 763B VG Adjuvants on the Immune Response against Streptococcus Agalactiae in Nile Tilapia (Oreochromis Niloticus). Fish & Shellfish Immunology 2023, 134, 108563. [Google Scholar] [CrossRef]
- Wangkaghart, E.; Deville, S.; Wang, B.; Srisapoome, P.; Wang, T.; Secombes, C.J. Immune Response and Protective Efficacy of Two New Adjuvants, MontanideTM ISA 763B VG and MontanideTM GEL02, Administered with a Streptococcus Agalactiae Ghost Vaccine in Nile Tilapia (Oreochromis Niloticus). Fish & Shellfish Immunology 2021, 116, 19–29. [Google Scholar] [CrossRef]
- Veenstra, K.A.; Wang, T.; Russell, K.S.; Tubbs, L.; Ben Arous, J.; Secombes, C.J. MontanideTM ISA 763A VG and ISA 761 VG Induce Different Immune Pathway Responses in Rainbow Trout (Oncorhynchus Mykiss) When Used as Adjuvant for an Aeromonas Salmonicida Bacterin. Fish & Shellfish Immunology 2021, 114, 171–183. [Google Scholar] [CrossRef]
- Thim, H.L.; Villoing, S.; McLoughlin, M.; Christie, K.E.; Grove, S.; Frost, P.; Jørgensen, J.B. Vaccine Adjuvants in Fish Vaccines Make a Difference: Comparing Three Adjuvants (Montanide ISA763A Oil, CpG/Poly I:C Combo and VHSV Glycoprotein) Alone or in Combination Formulated with an Inactivated Whole Salmonid Alphavirus Antigen. Vaccines 2014, 2, 228–251. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wang, W. Factors Affecting Alum–Protein Interactions. International Journal of Pharmaceutics 2014, 466, 139–146. [Google Scholar] [CrossRef] [PubMed]
- HogenEsch, H. Mechanism of Immunopotentiation and Safety of Aluminum Adjuvants. Front. Immunol. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zou, Y.; Hu, Z. Advances in Aluminum Hydroxide-Based Adjuvant Research and Its Mechanism. Human Vaccines & Immunotherapeutics 2015, 11, 477–488. [Google Scholar] [CrossRef]
- HogenEsch, H.; O’Hagan, D.T.; Fox, C.B. Optimizing the Utilization of Aluminum Adjuvants in Vaccines: You Might Just Get What You Want. npj Vaccines 2018, 3, 1–11. [Google Scholar] [CrossRef]
- Angosto, D.; López-Muñoz, A.; García-Alcazar, A.; Meseguer, J.; Sepulcre, M.P.; Mulero, V. Aluminum Is a Powerful Adjuvant in Teleost Fish despite Failing to Induce Interleukin-1β Release. Developmental & Comparative Immunology 2018, 85, 18–24. [Google Scholar] [CrossRef]
- Compan, V.; Baroja-Mazo, A.; López-Castejón, G.; Gomez, A.I.; Martínez, C.M.; Angosto, D.; Montero, M.T.; Herranz, A.S.; Bazán, E.; Reimers, D.; et al. Cell Volume Regulation Modulates NLRP3 Inflammasome Activation. Immunity 2012, 37, 487–500. [Google Scholar] [CrossRef]
- Angosto, D.; López-Castejón, G.; López-Muñoz, A.; Sepulcre, M.P.; Arizcun, M.; Meseguer, J.; Mulero, V. Evolution of Inflammasome Functions in Vertebrates: Inflammasome and Caspase-1 Trigger Fish Macrophage Cell Death but Are Dispensable for the Processing of IL-1β. Innate Immun 2012, 18, 815–824. [Google Scholar] [CrossRef]
- Bird, S.; Zou, J.; Wang, T.; Munday, B.; Cunningham, C.; Secombes, C.J. Evolution of Interleukin-1β. Cytokine & Growth Factor Reviews 2002, 13, 483–502. [Google Scholar] [CrossRef]
- McKee, A.S.; Burchill, M.A.; Munks, M.W.; Jin, L.; Kappler, J.W.; Friedman, R.S.; Jacobelli, J.; Marrack, P. Host DNA Released in Response to Aluminum Adjuvant Enhances MHC Class II-Mediated Antigen Presentation and Prolongs CD4 T-Cell Interactions with Dendritic Cells. Proceedings of the National Academy of Sciences of the United States of America 2013, 110, E1122–E1131. [Google Scholar] [CrossRef] [PubMed]
- Marichal, T.; Ohata, K.; Bedoret, D.; Mesnil, C.; Sabatel, C.; Kobiyama, K.; Lekeux, P.; Coban, C.; Akira, S.; Ishii, K.J.; et al. DNA Released from Dying Host Cells Mediates Aluminum Adjuvant Activity. Nat Med 2011, 17, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Kool, M.; Pétrilli, V.; De Smedt, T.; Rolaz, A.; Hammad, H.; van Nimwegen, M.; Bergen, I.M.; Castillo, R.; Lambrecht, B.N.; Tschopp, J. Cutting Edge: Alum Adjuvant Stimulates Inflammatory Dendritic Cells through Activation of the NALP3 Inflammasome. The Journal of Immunology 2008, 181, 3755–3759. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Villegas, J.; García-Alcazar, A.; Meseguer, J.; Mulero, V. Aluminum Adjuvant Potentiates Gilthead Seabream Immune Responses but Induces Toxicity in Splenic Melanomacrophage Centers. Fish & Shellfish Immunology 2019, 85, 31–43. [Google Scholar] [CrossRef]
- Guo, M.; Li, C. An Overview of Cytokine Used as Adjuvants in Fish: Current State and Future Trends. Reviews in Aquaculture 2021, 13, 996–1014. [Google Scholar] [CrossRef]
- Wang, Q.; Ji, W.; Xu, Z. Current Use and Development of Fish Vaccines in China. Fish & Shellfish Immunology 2020, 96, 223–234. [Google Scholar] [CrossRef]
- Thirumalaikumar, E.; Vimal, S.; Sathishkumar, R.; Ravi, M.; Karthick, V.; Ramya, S.; Thomas, J.; Kumar, V.; Kamaraj, C.; Citarasu, T. DNA Vaccine Incorporated Poly (Lactic-Co-Glycolic) Acid (PLGA) Microspheres Offer Enhanced Protection against Aeromonas Hydrophila Infection. International Journal of Biological Macromolecules 2023, 253, 127182. [Google Scholar] [CrossRef]
- Kole, S.; Qadiri, S.S.N.; Shin, S.-M.; Kim, W.-S.; Lee, J.; Jung, S.-J. PLGA Encapsulated Inactivated-Viral Vaccine: Formulation and Evaluation of Its Protective Efficacy against Viral Haemorrhagic Septicaemia Virus (VHSV) Infection in Olive Flounder (Paralichthys Olivaceus) Vaccinated by Mucosal Delivery Routes. Vaccine 2019, 37, 973–983. [Google Scholar] [CrossRef]
- Kole, S.; Qadiri, S.S.N.; Shin, S.-M.; Kim, W.-S.; Lee, J.; Jung, S.-J. PLGA Encapsulated Inactivated-Viral Vaccine: Formulation and Evaluation of Its Protective Efficacy against Viral Haemorrhagic Septicaemia Virus (VHSV) Infection in Olive Flounder (Paralichthys Olivaceus) Vaccinated by Mucosal Delivery Routes. Vaccine 2019, 37, 973–983. [Google Scholar] [CrossRef]
- Kole, S.; Qadiri, S.S.N.; Shin, S.-M.; Kim, W.-S.; Lee, J.; Jung, S.-J. PLGA Encapsulated Inactivated-Viral Vaccine: Formulation and Evaluation of Its Protective Efficacy against Viral Haemorrhagic Septicaemia Virus (VHSV) Infection in Olive Flounder (Paralichthys Olivaceus) Vaccinated by Mucosal Delivery Routes. Vaccine 2019, 37, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Huo, X.; Tang, L.; Hu, M.; Yang, C.; Luo, D.; Su, J. Oral PLGA-Based DNA Vaccines Using Interferons as Adjuvants Remarkably Promote the Immune Protection of Grass Carp (Ctenopharyngodon Idella) against GCRV Infection. Water Biology and Security 2023, 2, 100143. [Google Scholar] [CrossRef]
- Yogeshwari, G. Poly D, L-Lactide-Co-Glycolic Acid (PLGA)-Encapsulated CpG-Oligonucleotide (ODN) on Immune Response in Cyprinus Carpio against Aeromonas Hydrophila. Journal of Aquaculture Research & Development 2015, 06. [Google Scholar] [CrossRef]
- Diao, J.; Ye, H.; Yu, X.; Fan, Y.; Xu, L.; Li, T.; Wang, Y. Adjuvant and Immunostimulatory Effects of LPS and β-Glucan on Immune Response in Japanese Flounder, Paralichthys Olivaceus. Veterinary Immunology and Immunopathology 2013, 156, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Wangkahart, E.; Secombes, C.J.; Wang, T. Studies on the Use of Flagellin as an Immunostimulant and Vaccine Adjuvant in Fish Aquaculture. Front. Immunol. 2019, 9. [Google Scholar] [CrossRef]
- Sahoo, L.; Parhi, J.; Debnath, C.; Prasad, K.P. Effect of Feeding Lipopolysaccharide as an Immunostimulant on Immune Response and Immune Gene Expression of Labeo Bata. Veterinary Immunology and Immunopathology 2017, 188, 48–58. [Google Scholar] [CrossRef]
- Munang’andu, H.M.; Salinas, I.; Tafalla, C.; Dalmo, R.A. Editorial: Vaccines and Immunostimulants for Finfish. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Vallejos-Vidal, E.; Reyes-López, F.; Teles, M.; MacKenzie, S. The Response of Fish to Immunostimulant Diets. Fish & Shellfish Immunology 2016, 56, 34–69. [Google Scholar] [CrossRef]
- Giri, S.S.; Chi, C.; Jun, J.W.; Park, S.C. Use of Bacterial Subcellular Components as Immunostimulants in Fish Aquaculture. Reviews in Aquaculture 2018, 10, 474–492. [Google Scholar] [CrossRef]
- Hoel, K.; Lillehaug, A. Adjuvant Activity of Polar Glycopeptidolipids fromMycobacterium Chelonaein Experimental Vaccines againstAeromonas Salmonicidain Salmonid Fish. Fish & Shellfish Immunology 1997, 7, 365–376. [Google Scholar] [CrossRef]
- Burakova, Y.; Madera, R.; McVey, S.; Schlup, J.R.; Shi, J. Adjuvants for Animal Vaccines. Viral Immunology 2018, 31, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Vinay, T.-N.; Park, C.-S.; Kim, H.-Y.; Jung, S.-J. Toxicity and Dose Determination of Quillaja Saponin, Aluminum Hydroxide and Squalene in Olive Flounder (Paralichthys Olivaceus). Veterinary Immunology and Immunopathology 2014, 158, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Huang, J.; Li, J. Adjuvant Effect of Quillaja Saponaria Saponin (QSS) on Protective Efficacy and IgM Generation in Turbot (Scophthalmus Maximus) upon Immersion Vaccination. International Journal of Molecular Sciences 2016, 17, 325. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.-H.; Jung, S.-J.; Kim, T. Saponin and Chitosan-Based Oral Vaccine against Viral Haemorrhagic Septicaemia Virus (VHSV) Provides Protective Immunity in Olive Flounder (Paralichthys Olivaceus). Fish & Shellfish Immunology 2022, 126, 336–346. [Google Scholar] [CrossRef]
- Jung, M.-H.; Kole, S.; Jung, S.-J. Efficacy of Saponin-Based Inactivated Rock Bream Iridovirus (RBIV) Vaccine in Rock Bream (Oplegnathus Fasciatus). Fish & Shellfish Immunology 2022, 121, 12–22. [Google Scholar] [CrossRef]
- Sun, F.; Zhou, J.; Zhang, Y.; Liu, Q.; Wang, Q.; Liu, X. A Compound Ginseng Stem Leaf Saponins and Aluminium Adjuvant Enhances the Potency of Inactivated Aeromonas Salmonicida Vaccine in Turbot. Fish & Shellfish Immunology 2022, 128, 60–66. [Google Scholar] [CrossRef]
- Sun, F.; Wu, Y.; Zhang, Y.; Liu, Q.; Wang, Q.; Liu, X. An Aluminium Adjuvant Compound with Ginseng Stem Leaf Saponins Enhances the Potency of Inactivated Pseudomonas Plecoglossicida Vaccine in Large Yellow Croaker (Larimichthys Crocea). Fish & Shellfish Immunology 2024, 144, 109243. [Google Scholar] [CrossRef]
- Soltani, M.; Lymbery, A.; Song, S.K.; Hosseini Shekarabi, P. Adjuvant Effects of Medicinal Herbs and Probiotics for Fish Vaccines. Reviews in Aquaculture 2019, 11, 1325–1341. [Google Scholar] [CrossRef]
- Beck, B.R.; Lee, S.H.; Kim, D.; Park, J.H.; Lee, H.K.; Kwon, S.-S.; Lee, K.H.; Lee, J.I.; Song, S.K. A Lactococcus Lactis BFE920 Feed Vaccine Expressing a Fusion Protein Composed of the OmpA and FlgD Antigens from Edwardsiella Tarda Was Significantly Better at Protecting Olive Flounder (Paralichthys Olivaceus) from Edwardsiellosis than Single Antigen Vaccines. Fish & Shellfish Immunology 2017, 68, 19–28. [Google Scholar] [CrossRef]
- Lee, S.H.; Beck, B.R.; Hwang, S.-H.; Song, S.K. Feeding Olive Flounder (Paralichthys Olivaceus) with Lactococcus Lactis BFE920 Expressing the Fusion Antigen of Vibrio OmpK and FlaB Provides Protection against Multiple Vibrio Pathogens: A Universal Vaccine Effect. Fish & Shellfish Immunology 2021, 114, 253–262. [Google Scholar] [CrossRef]
- Kim, D.; Beck, B.R.; Lee, S.M.; Jeon, J.; Lee, D.W.; Lee, J.I.; Song, S.K. Pellet Feed Adsorbed with the Recombinant Lactococcus Lactis BFE920 Expressing SiMA Antigen Induced Strong Recall Vaccine Effects against Streptococcus Iniae Infection in Olive Flounder (Paralichthys Olivaceus). Fish & Shellfish Immunology 2016, 55, 374–383. [Google Scholar] [CrossRef]
- Anuradha, K.; Foo, H.L.; Mariana, N.S.; Loh, T.C.; Yusoff, K.; Hassan, M.D.; Sasan, H.; Raha, A.R. Live Recombinant Lactococcus Lactis Vaccine Expressing Aerolysin Genes D1 and D4 for Protection against Aeromonas Hydrophila in Tilapia (Oreochromis Niloticus). Journal of Applied Microbiology 2010, 109, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Naderi-Samani, M.; Soltani, M.; Dadar, M.; Taheri-Mirghaed, A.; Zargar, A.; Ahmadivand, S.; Hassanzadeh, R.; Goudarzi, L.M. Oral Immunization of Trout Fry with Recombinant Lactococcus Lactis NZ3900 Expressing G Gene of Viral Hemorrhagic Septicaemia Virus (VHSV). Fish & Shellfish Immunology 2020, 105, 62–70. [Google Scholar] [CrossRef]
- Docando, F.; Nuñez-Ortiz, N.; Gonçalves, G.; Serra, C.R.; Gomez-Casado, E.; Martín, D.; Abós, B.; Oliva-Teles, A.; Tafalla, C.; Díaz-Rosales, P. Bacillus Subtilis Expressing the Infectious Pancreatic Necrosis Virus VP2 Protein Retains Its Immunostimulatory Properties and Induces a Specific Antibody Response. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-D.; Yao, Y.-Y.; Cui, Z.-W.; Zhang, X.-Y.; Guo, X.; Zhou, Y.-Y.; Zhang, Y.-A. Comparative Study of the Immunoprotective Effect of Two Grass Carp-Sourced Bacillus Subtilis Spore-Based Vaccines against Grass Carp Reovirus. Aquaculture 2019, 504, 88–95. [Google Scholar] [CrossRef]
- Gonçalves, G.; Santos, R.A.; Coutinho, F.; Pedrosa, N.; Curado, M.; Machado, M.; Costas, B.; Bonneville, L.; Serrano, M.; Carvalho, A.P.; et al. Oral Vaccination of Fish against Vibriosis Using Spore-Display Technology. Front. Immunol. 2022, 13. [Google Scholar] [CrossRef]
- Wang, Q.; Liang, X.; Ning, Y.; Liu, S.; Liang, Z.; Zhang, Z.; Chen, Y.; Cao, J.; Wang, F.; Lan, L.; et al. Surface Display of Major Capsid Protein on Bacillus Subtilis Spores against Largemouth Bass Virus (LMBV) for Oral Administration. Fish & Shellfish Immunology 2023, 135, 108627. [Google Scholar] [CrossRef]
- Huang, X.; Ma, Y.; Wang, Y.; Niu, C.; Liu, Z.; Yao, X.; Jiang, X.; Pan, R.; Jia, S.; Li, D.; et al. Oral Probiotic Vaccine Expressing Koi Herpesvirus (KHV) ORF81 Protein Delivered by Chitosan-Alginate Capsules Is a Promising Strategy for Mass Oral Vaccination of Carps against KHV Infection. Journal of Virology 2021, 95. [Google Scholar] [CrossRef]
- Wu, Y.; Rashidpour, A.; Almajano, M.P.; Metón, I. Chitosan-Based Drug Delivery System: Applications in Fish Biotechnology. Polymers 2020, 12, 1177. [Google Scholar] [CrossRef]
- Mohan, K.; Rajan, D.K.; Ganesan, A.R.; Divya, D.; Johansen, J.; Zhang, S. Chitin, Chitosan and Chitooligosaccharides as Potential Growth Promoters and Immunostimulants in Aquaculture: A Comprehensive Review. International Journal of Biological Macromolecules 2023, 251, 126285. [Google Scholar] [CrossRef]
- Collado-González, M.; Esteban, M.Á. Chitosan-Nanoparticles Effects on Mucosal Immunity: A Systematic Review. Fish & Shellfish Immunology 2022, 130, 1–8. [Google Scholar] [CrossRef]
- Tian, J.; Yu, J.; Sun, X. Chitosan Microspheres as Candidate Plasmid Vaccine Carrier for Oral Immunisation of Japanese Flounder (Paralichthys Olivaceus). Veterinary Immunology and Immunopathology 2008, 126, 220–229. [Google Scholar] [CrossRef] [PubMed]
- León-Rodríguez, L.; Luzardo-Álvarez, A.; Blanco-Méndez, J.; Lamas, J.; Leiro, J. A Vaccine Based on Biodegradable Microspheres Induces Protective Immunity against Scuticociliatosis without Producing Side Effects in Turbot. Fish & Shellfish Immunology 2012, 33, 21–27. [Google Scholar] [CrossRef]
- Jin, P.; Sun, F.; Liu, Q.; Wang, Q.; Zhang, Y.; Liu, X. An Oral Vaccine Based on Chitosan/Aluminum Adjuvant Induces Both Local and Systemic Immune Responses in Turbot (Scophthalmus Maximus). Vaccine 2021, 39, 7477–7484. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.-F.; Jiang, F.-Y.; Zhou, G.-Q.; Xia, J.-Y.; Yang, F.; Zhu, B. The Recombinant Subunit Vaccine Encapsulated by Alginate-Chitosan Microsphere Enhances the Immune Effect against Micropterus Salmoides Rhabdovirus. Journal of Fish Diseases 2022, 45, 1757–1765. [Google Scholar] [CrossRef]
- Wang, E.; Wang, X.; Wang, K.; He, J.; Zhu, L.; He, Y.; Chen, D.; Ouyang, P.; Geng, Y.; Huang, X.; et al. Preparation, Characterization and Evaluation of the Immune Effect of Alginate/Chitosan Composite Microspheres Encapsulating Recombinant Protein of Streptococcus Iniae Designed for Fish Oral Vaccination. Fish & Shellfish Immunology 2018, 73, 262–271. [Google Scholar] [CrossRef]
- Behera, T.; Swain, P. Antigen Encapsulated Alginate-Coated Chitosan Microspheres Stimulate Both Innate and Adaptive Immune Responses in Fish through Oral Immunization. Aquacult Int 2014, 22, 673–688. [Google Scholar] [CrossRef]
- Behera, T.; Swain, P. Alginate–Chitosan–PLGA Composite Microspheres Induce Both Innate and Adaptive Immune Response through Parenteral Immunization in Fish. Fish & Shellfish Immunology 2013, 35, 785–791. [Google Scholar] [CrossRef]
- Dezfuly, Z.T.; Alishahi, M.; Ghorbanpoor, M.; Tabandeh, M.R.; Mesbah, M. Immunogenicity and Protective Efficacy of Yersinia Ruckeri Lipopolysaccharide (LPS), Encapsulated by Alginate-Chitosan Micro/Nanoparticles in Rainbow Trout (Oncorhyncus Mykiss). Fish & Shellfish Immunology 2020, 104, 25–35. [Google Scholar] [CrossRef]
- González-Chavarría, I.; Roa, F.J.; Sandoval, F.; Muñoz-Flores, C.; Kappes, T.; Acosta, J.; Bertinat, R.; Altamirano, C.; Valenzuela, A.; Sánchez, O.; et al. Chitosan Microparticles Enhance the Intestinal Release and Immune Response of an Immune Stimulant Peptide in Oncorhynchus Mykiss. International Journal of Molecular Sciences 2023, 24, 14685. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Wiulsrød, R.; Fredriksen, B.N.; Lindmo, K.; Løkling, K.-E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and Future Perspectives of Vaccines for Industrialised Fin-Fish Farming. Fish & Shellfish Immunology 2013, 35, 1759–1768. [Google Scholar] [CrossRef]
- Hoare, R.; Jung, S.-J.; Ngo, T.P.H.; Bartie, K.; Bailey, J.; Thompson, K.D.; Adams, A. Efficacy and Safety of a Non-Mineral Oil Adjuvanted Injectable Vaccine for the Protection of Atlantic Salmon (Salmo Salar L.) against Flavobacterium Psychrophilum. Fish & Shellfish Immunology 2019, 85, 44–51. [Google Scholar] [CrossRef]
- Poppe, T.T.; Koppang, E.O. Side-Effects of Vaccination. In Fish Vaccination; John Wiley & Sons, Ltd., 2014; pp. 153–161. ISBN 978-1-118-80691-3. [Google Scholar]
- Mutoloki, S.; Cooper, G.A.; Marjara, I.S.; Koop, B.F.; Evensen, Ø. High Gene Expression of Inflammatory Markers and IL-17A Correlates with Severity of Injection Site Reactions of Atlantic Salmon Vaccinated with Oil-Adjuvanted Vaccines. BMC Genomics 2010, 11, 336. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Cheng, S.; Hu, Y.; Sun, L. Comparative Study of the Effects of Aluminum Adjuvants and Freund’s Incomplete Adjuvant on the Immune Response to an Edwardsiella Tarda Major Antigen. Vaccine 2010, 28, 1832–1837. [Google Scholar] [CrossRef]
- Gjessing, M.C.; Falk, K.; Weli, S.C.; Koppang, E.O.; Kvellestad, A. A Sequential Study of Incomplete Freund’s Adjuvant-Induced Peritonitis in Atlantic Cod. Fish & Shellfish Immunology 2012, 32, 141–150. [Google Scholar] [CrossRef]
- Spinos, E.; Kokkoris, G.D.; Bakopoulos, V. Prevention of Sea Bass (Dicentrarchus Labrax, L. 1758) Photobacteriosis and Vibriosis. Long Term Efficacy Study of Intraperitoneally Administered Bivalent Commercial Vaccines. Aquaculture 2017, 471, 172–184. [Google Scholar] [CrossRef]
- Veenstra, K.A.; Wang, T.; Alnabulsi, A.; Douglas, A.; Russell, K.S.; Tubbs, L.; Arous, J.B.; Secombes, C.J. Analysis of Adipose Tissue Immune Gene Expression after Vaccination of Rainbow Trout with Adjuvanted Bacterins Reveals an Association with Side Effects. Molecular Immunology 2017, 88, 89–98. [Google Scholar] [CrossRef]
- Li, J.; Tang, L.; Li, S.; Li, G.; Mo, Z. The Efficacy and Side-Effects of Oil-Based Adjuvants Emulsified Vibrio Anguillarum Bivalent Inactivated Vaccine in Turbot (Scophthalmus Maximus) under Production Mode. Aquaculture 2020, 524, 735259. [Google Scholar] [CrossRef]
- Tziouvas, H.; Varvarigos, P. Intensity Scale of Side Effects in European Sea Bass (Dicentrarchus Labrax) Post Intraperitoneal Injection with Commercial Oil-Adjuvanted Vaccines. Bulletin of the EAFP 2021, 41, 103–110. [Google Scholar] [CrossRef]
- Midtlyng, P.J.; Reitan, L.J.; Speilberg, L. Experimental Studies on the Efficacy and Side-Effects of Intraperitoneal Vaccination of Atlantic Salmon (Salmo salarL.) against Furunculosis. Fish & Shellfish Immunology 1996, 6, 335–350. [Google Scholar] [CrossRef]
- Harshitha, M.; Nayak, A.; Disha, S.; Akshath, U.S.; Dubey, S.; Munang’andu, H.M.; Chakraborty, A.; Karunasagar, I.; Maiti, B. Nanovaccines to Combat Aeromonas Hydrophila Infections in Warm-Water Aquaculture: Opportunities and Challenges. Vaccines 2023, 11, 1555. [Google Scholar] [CrossRef] [PubMed]
- Fredriksen, B.N.; Grip, J. PLGA/PLA Micro- and Nanoparticle Formulations Serve as Antigen Depots and Induce Elevated Humoral Responses after Immunization of Atlantic Salmon (Salmo Salar L.). Vaccine 2012, 30, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Fredriksen, B.N.; Sævareid, K.; McAuley, L.; Lane, M.E.; Bøgwald, J.; Dalmo, R.A. Early Immune Responses in Atlantic Salmon (Salmo Salar L.) after Immunization with PLGA Nanoparticles Loaded with a Model Antigen and β-Glucan. Vaccine 2011, 29, 8338–8349. [Google Scholar] [CrossRef]
- Adomako, M.; St-Hilaire, S.; Zheng, Y.; Eley, J.; Marcum, R.D.; Sealey, W.; Donahower, B.C.; LaPatra, S.; Sheridan, P.P. Oral DNA Vaccination of Rainbow Trout, Oncorhynchus Mykiss (Walbaum), against Infectious Haematopoietic Necrosis Virus Using PLGA [Poly(D,L-Lactic-Co-Glycolic Acid)] Nanoparticles. Journal of Fish Diseases 2012, 35, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Yu, J. Poly(Lactic-Co-Glycolic Acid) Nanoparticles as Candidate DNA Vaccine Carrier for Oral Immunization of Japanese Flounder (Paralichthys Olivaceus) against Lymphocystis Disease Virus. Fish & Shellfish Immunology 2011, 30, 109–117. [Google Scholar] [CrossRef]
- Dubey, S.; Avadhani, K.; Mutalik, S.; Sivadasan, S.M.; Maiti, B.; Paul, J.; Girisha, S.K.; Venugopal, M.N.; Mutoloki, S.; Evensen, Ø.; et al. Aeromonas Hydrophila OmpW PLGA Nanoparticle Oral Vaccine Shows a Dose-Dependent Protective Immunity in Rohu (Labeo Rohita). Vaccines 2016, 4, 21. [Google Scholar] [CrossRef]
- Harshitha, M.; D’souza, R.; Akshay, S.D.; Nayak, A.; Disha, S.; Aditya, V.; Akshath, U.S.; Dubey, S.; Munang’andu, H.M.; Chakraborty, A.; et al. Oral Administration of Recombinant Outer Membrane Protein A-Based Nanovaccine Affords Protection against Aeromonas Hydrophila in Zebrafish. World J Microbiol Biotechnol 2024, 40, 250. [Google Scholar] [CrossRef]
- Alishahi, M.; Vaseghi, M.; Tabandeh, M.R.; Khosravi, M. Immunogenic and Protective Effects of an Oral Polylactic-Co-Glycolic Acid Nano Encapsulated DNA Vaccine Encoding aopB Gene of Aeromonas Hydrophila in Common Carp. Aquacult Int 2024, 32, 1169–1190. [Google Scholar] [CrossRef]
- Alishahi, M.; Lababian, H.; Heidari, H.; Tabandeh, M.R.; Khosravi, M. Development of an Injectable DNA Vaccine Against Aeromonas Hydrophila Infection Nanoencapsulated With Poly(Lactic-Co-Glycolic) Acid (PLGA) in Common Carp. Aquaculture Research 2024, 2024, 7270489. [Google Scholar] [CrossRef]
- Kole, S.; Qadiri, S.S.N.; Shin, S.-M.; Kim, W.-S.; Lee, J.; Jung, S.-J. PLGA Encapsulated Inactivated-Viral Vaccine: Formulation and Evaluation of Its Protective Efficacy against Viral Haemorrhagic Septicaemia Virus (VHSV) Infection in Olive Flounder (Paralichthys Olivaceus) Vaccinated by Mucosal Delivery Routes. Vaccine 2019, 37, 973–983. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, P.-Q.; Guo, S.; Zhao, Z.; Wang, G.-X.; Zhu, B. Dual-Targeting Polymer Nanoparticles Efficiently Deliver DNA Vaccine and Induce Robust Prophylactic Immunity against Spring Viremia of Carp Virus Infection. Microbiology Spectrum 2022, 10, e03085-. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Soliman, F.M.; Adly, M.A.; Soliman, H.A.M.; El-Matbouli, M.; Saleh, M. Recent Progress in Biomedical Applications of Chitosan and Its Nanocomposites in Aquaculture: A Review. Research in Veterinary Science 2019, 126, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Rashidpour, A.; Almajano, M.P.; Metón, I. Chitosan-Based Drug Delivery System: Applications in Fish Biotechnology. Polymers 2020, 12, 1177. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, M.; et al. Applications of Chitosan and Chitosan Nanoparticles in Fish Aquaculture. Egyptian Journal of Aquatic Biology and Fisheries 2022, 26, 23–43. [Google Scholar] [CrossRef]
- Kole, S.; Dar, S.A.; Shin, S.-M.; Jeong, H.-J.; Jung, S.-J. Potential Efficacy of Chitosan-Poly (Lactide-Co-Glycolide)-Encapsulated Trivalent Immersion Vaccine in Olive Flounder (Paralichthys Olivaceus) Against Viral Hemorrhagic Septicemia Virus, Streptococcus Parauberis Serotype I, and Miamiensis Avidus (Scuticociliate). Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Ahmadivand, S.; Soltani, M.; Behdani, M.; Evensen, Ø.; Alirahimi, E.; Hassanzadeh, R.; Soltani, E. Oral DNA Vaccines Based on CS-TPP Nanoparticles and Alginate Microparticles Confer High Protection against Infectious Pancreatic Necrosis Virus (IPNV) Infection in Trout. Developmental & Comparative Immunology 2017, 74, 178–189. [Google Scholar] [CrossRef]
- Rao, B.M.; Kole, S.; Gireesh-Babu, P.; Sharma, R.; Tripathi, G.; Bedekar, M.K. Evaluation of Persistence, Bio-Distribution and Environmental Transmission of Chitosan/PLGA/pDNA Vaccine Complex against Edwardsiella Tarda in Labeo Rohita. Aquaculture 2019, 500, 385–392. [Google Scholar] [CrossRef]
- Halimi, M.; Alishahi, M.; Abbaspour, M.R.; Ghorbanpoor, M.; Tabandeh, M.R. Valuable Method for Production of Oral Vaccine by Using Alginate and Chitosan against Lactococcus Garvieae/Streptococcus Iniae in Rainbow Trout (Oncorhynchus Mykiss). Fish & Shellfish Immunology 2019, 90, 431–439. [Google Scholar] [CrossRef]
- Huo, X.; Tang, L.; Liu, Q.; Zhu, W.; Zhang, J.; Hu, M.; Zhao, F.; Wang, P.; Yuan, G.; Yang, C.; et al. Oral pcDNA3.1-VP4/VP56-FlaC DNA Vaccine Encapsulated by Chitosan/Sodium Alginate Nanoparticles Confers Remarkable Protection against GCRV Infection in Grass Carp. Aquaculture 2023, 577, 739996. [Google Scholar] [CrossRef]
- Leya, T.; Ahmad, I.; Sharma, R.; Tripathi, G.; Kurcheti, P.P.; Rajendran, K.V.; Bedekar, M.K. Bicistronic DNA Vaccine Macromolecule Complexed with Poly Lactic-Co-Glycolic Acid-Chitosan Nanoparticles Enhanced the Mucosal Immunity of Labeo Rohita against Edwardsiella Tarda Infection. International Journal of Biological Macromolecules 2020, 156, 928–937. [Google Scholar] [CrossRef]
- Li, L.; Lin, S.-L.; Deng, L.; Liu, Z.-G. Potential Use of Chitosan Nanoparticles for Oral Delivery of DNA Vaccine in Black Seabream Acanthopagrus Schlegelii Bleeker to Protect from Vibrio Parahaemolyticus. Journal of Fish Diseases 2013, 36, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Rajesh Kumar, S.; Ishaq Ahmed, V.P.; Parameswaran, V.; Sudhakaran, R.; Sarath Babu, V.; Sahul Hameed, A.S. Potential Use of Chitosan Nanoparticles for Oral Delivery of DNA Vaccine in Asian Sea Bass (Lates Calcarifer) to Protect from Vibrio (Listonella) Anguillarum. Fish & Shellfish Immunology 2008, 25, 47–56. [Google Scholar] [CrossRef]
- Sukkarun, P.; Kitiyodom, S.; Yostawornkul, J.; Chaiin, P.; Yata, T.; Rodkhum, C.; Boonrungsiman, S.; Pirarat, N. Chitosan-Polymer Based Nanovaccine as Promising Immersion Vaccine against Aeromonas Veronii Challenge in Red Tilapia (Oreochromis Sp.). Fish & Shellfish Immunology 2022, 129, 30–35. [Google Scholar] [CrossRef]
- Kitiyodom, S.; Yata, T.; Yostawornkul, J.; Kaewmalun, S.; Nittayasut, N.; Suktham, K.; Surassmo, S.; Namdee, K.; Rodkhum, C.; Pirarat, N. Enhanced Efficacy of Immersion Vaccination in Tilapia against Columnaris Disease by Chitosan-Coated “Pathogen-like” Mucoadhesive Nanovaccines. Fish & Shellfish Immunology 2019, 95, 213–219. [Google Scholar] [CrossRef]
- Dubey, S.; Avadhani, K.; Mutalik, S.; Sivadasan, S.M.; Maiti, B.; Girisha, S.K.; Venugopal, M.N.; Mutoloki, S.; Evensen, Ø.; Karunasagar, I.; et al. Edwardsiella Tarda OmpA Encapsulated in Chitosan Nanoparticles Shows Superior Protection over Inactivated Whole Cell Vaccine in Orally Vaccinated Fringed-Lipped Peninsula Carp (Labeo Fimbriatus). Vaccines 2016, 4, 40. [Google Scholar] [CrossRef]
- Thirumalaikumar, E.; Lelin, C.; Sathishkumar, R.; Vimal, S.; Anand, S.B.; Babu, M.M.; Citarasu, T. Oral Delivery of pVAX-OMP and pVAX-Hly DNA Vaccine Using Chitosan-Tripolyphosphate (Cs-TPP) Nanoparticles in Rohu, (Labeo Rohita) for Protection against Aeromonas Hydrophila Infection. Fish & Shellfish Immunology 2021, 115, 189–197. [Google Scholar] [CrossRef]
- Kole, S.; Kumari, R.; Anand, D.; Kumar, S.; Sharma, R.; Tripathi, G.; Makesh, M.; Rajendran, K.V.; Bedekar, M.K. Nanoconjugation of Bicistronic DNA Vaccine against Edwardsiella Tarda Using Chitosan Nanoparticles: Evaluation of Its Protective Efficacy and Immune Modulatory Effects in Labeo Rohita Vaccinated by Different Delivery Routes. Vaccine 2018, 36, 2155–2165. [Google Scholar] [CrossRef]
- Zhang, Z.; Jin, Z.; Wang, G.; Ling, F. Mannosylated Chitosan Nanoparticles Loaded with ABP Antigen Server as a Novel Nucleic Acid Vaccine against Nocardia Seriolae Infection in Micropterus Salmoides. Aquaculture 2023, 574, 739635. [Google Scholar] [CrossRef]
- Ponce, M.; Zuasti, E.; Anguís, V.; Fernández-Díaz, C. Anti-Bacterial and Immunostimulatory Properties of Ulvan-Loaded Chitosan Nanoparticles for Use in Aquaculture. Mar Biotechnol 2024, 26, 19–27. [Google Scholar] [CrossRef]
- Sukkarun, P.; Kitiyodom, S.; Kamble, M.T.; Bunnoy, A.; Boonanuntanasarn, S.; Yata, T.; Boonrungsiman, S.; Thompson, K.D.; Rodkhum, C.; Pirarat, N. Systemic and Mucosal Immune Responses in Red Tilapia (Oreochromis Sp.) Following Immersion Vaccination with a Chitosan Polymer-Based Nanovaccine against Aeromonas Veronii. Fish & Shellfish Immunology 2024, 146, 109383. [Google Scholar] [CrossRef]
- Kitiyodom, S.; Trullàs, C.; Rodkhum, C.; Thompson, K.D.; Katagiri, T.; Temisak, S.; Namdee, K.; Yata, T.; Pirarat, N. Modulation of the Mucosal Immune Response of Red Tilapia (Oreochromis Sp.) against Columnaris Disease Using a Biomimetic-Mucoadhesive Nanovaccine. Fish & Shellfish Immunology 2021, 112, 81–91. [Google Scholar] [CrossRef]
- Kitiyodom, S.; Yata, T.; Thompson, K.D.; Costa, J.; Elumalai, P.; Katagiri, T.; Temisak, S.; Namdee, K.; Rodkhum, C.; Pirarat, N. Immersion Vaccination by a Biomimetic-Mucoadhesive Nanovaccine Induces Humoral Immune Response of Red Tilapia (Oreochromis Sp.) against Flavobacterium Columnare Challenge. Vaccines 2021, 9, 1253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhao, Z.; Zhou, J.; Wang, W.; Su, J.; Yuan, G. Carboxymethyl Chitosan Nanoparticles Loaded with Ctenopharyngodon Idella Interferon-Γ2 (CiIFN-Γ2) Enhance Protective Efficacy against Bacterial Infection in Grass Carp. Aquaculture 2023, 572, 739554. [Google Scholar] [CrossRef]
- Tattiyapong, P.; Kitiyodom, S.; Yata, T.; Jantharadej, K.; Adamek, M.; Surachetpong, W. Chitosan Nanoparticle Immersion Vaccine Offers Protection against Tilapia Lake Virus in Laboratory and Field Studies. Fish & Shellfish Immunology 2022, 131, 972–979. [Google Scholar] [CrossRef]
- Zheng, F.; Liu, H.; Sun, X.; Zhang, Y.; Zhang, B.; Teng, Z.; Hou, Y.; Wang, B. Development of Oral DNA Vaccine Based on Chitosan Nanoparticles for the Immunization against Reddish Body Iridovirus in Turbots (Scophthalmus Maximus). Aquaculture 2016, 452, 263–271. [Google Scholar] [CrossRef]
- Kole, S.; Qadiri, S.S.N.; Shin, S.-M.; Kim, W.-S.; Lee, J.; Jung, S.-J. Nanoencapsulation of Inactivated-Viral Vaccine Using Chitosan Nanoparticles: Evaluation of Its Protective Efficacy and Immune Modulatory Effects in Olive Flounder (Paralichthys Olivaceus) against Viral Haemorrhagic Septicaemia Virus (VHSV) Infection. Fish & Shellfish Immunology 2019, 91, 136–147. [Google Scholar] [CrossRef]
- Vimal, S.; Abdul Majeed, S.; Nambi, K.S.N.; Madan, N.; Farook, M.A.; Venkatesan, C.; Taju, G.; Venu, S.; Subburaj, R.; Thirunavukkarasu, A.R.; et al. Delivery of DNA Vaccine Using Chitosan–Tripolyphosphate (CS/TPP) Nanoparticles in Asian Sea Bass, Lates Calcarifer (Bloch, 1790) for Protection against Nodavirus Infection. Aquaculture 2014, 420–421, 240–246. [Google Scholar] [CrossRef]
- Valero, Y.; Awad, E.; Buonocore, F.; Arizcun, M.; Esteban, M.Á.; Meseguer, J.; Chaves-Pozo, E.; Cuesta, A. An Oral Chitosan DNA Vaccine against Nodavirus Improves Transcription of Cell-Mediated Cytotoxicity and Interferon Genes in the European Sea Bass Juveniles Gut and Survival upon Infection. Developmental & Comparative Immunology 2016, 65, 64–72. [Google Scholar] [CrossRef]
- Valero, Y.; Awad, E.; Buonocore, F.; Arizcun, M.; Esteban, M.Á.; Meseguer, J.; Chaves-Pozo, E.; Cuesta, A. An Oral Chitosan DNA Vaccine against Nodavirus Improves Transcription of Cell-Mediated Cytotoxicity and Interferon Genes in the European Sea Bass Juveniles Gut and Survival upon Infection. Developmental & Comparative Immunology 2016, 65, 64–72. [Google Scholar] [CrossRef]
- Thu Lan, N.G.; Dong, H.T.; Vinh, N.T.; Salin, K.R.; Senapin, S.; Pimsannil, K.; St-Hilaire, S.; Shinn, A.P.; Rodkhum, C. A Novel Vaccination Strategy against Vibrio Harveyi Infection in Asian Seabass (Lates Calcarifer) with the Aid of Oxygen Nanobubbles and Chitosan. Fish & Shellfish Immunology 2024, 149, 109557. [Google Scholar] [CrossRef]
- Andresen, A.M.S.; Gjøen, T. Chitosan Nanoparticle Formulation Attenuates Poly (I:C) Induced Innate Immune Responses against Inactivated Virus Vaccine in Atlantic Salmon (Salmo Salar). Comparative Biochemistry and Physiology Part D: Genomics and Proteomics 2021, 40, 100915. [Google Scholar] [CrossRef] [PubMed]
- Collado-González, M.; Esteban, M.Á. Chitosan-Nanoparticles Effects on Mucosal Immunity: A Systematic Review. Fish & Shellfish Immunology 2022, 130, 1–8. [Google Scholar] [CrossRef]
- Ji, J.; Merino, S.; Tomás, J.M.; Roher, N. Nanoliposomes Encapsulating Immunostimulants Modulate the Innate Immune System and Elicit Protection in Zebrafish Larvae. Fish & Shellfish Immunology 2019, 92, 421–429. [Google Scholar] [CrossRef]
- Bunnoy, A.; Thangsunan, P.; Chokmangmeepisarn, P.; Yata, T.; Klongklaew, N.; Pirarat, N.; Kitiyodom, S.; Srisapoome, P.; Rodkhum, C. Mucoadhesive Cationic Lipid-Based Flavobacterium Oreochromis Nanoencapsulation Enhanced the Efficacy of Mucoadhesive Immersion Vaccination against Columnaris Disease and Strengthened Immunity in Asian Sea Bass (Lates Calcarifer). Fish & Shellfish Immunology 2022, 127, 633–646. [Google Scholar] [CrossRef]
- Thangsunan, P.; Kitiyodom, S.; Srisapoome, P.; Pirarat, N.; Yata, T.; Thangsunan, P.; Boonrungsiman, S.; Bunnoy, A.; Rodkhum, C. Novel Development of Cationic Surfactant-Based Mucoadhesive Nanovaccine for Direct Immersion Vaccination against Francisella Noatunensis Subsp. Orientalis in Red Tilapia (Oreochromis Sp.). Fish & Shellfish Immunology 2022, 127, 1051–1060. [Google Scholar] [CrossRef]
- Ruyra, A.; Cano-Sarabia, M.; MacKenzie, S.A.; Maspoch, D.; Roher, N. A Novel Liposome-Based Nanocarrier Loaded with an LPS-dsRNA Cocktail for Fish Innate Immune System Stimulation. PLOS ONE 2013, 8, e76338. [Google Scholar] [CrossRef]
- Ruyra, A.; Cano-Sarabia, M.; García-Valtanen, P.; Yero, D.; Gibert, I.; Mackenzie, S.A.; Estepa, A.; Maspoch, D.; Roher, N. Targeting and Stimulation of the Zebrafish (Danio Rerio) Innate Immune System with LPS/dsRNA-Loaded Nanoliposomes. Vaccine 2014, 32, 3955–3962. [Google Scholar] [CrossRef]
- Dahl, L.O.S.; Hak, S.; Braaen, S.; Molska, A.; Rodà, F.; Parot, J.; Wessel, Ø.; Fosse, J.H.; Bjørgen, H.; Borgos, S.E.; et al. Implementation of mRNA–Lipid Nanoparticle Technology in Atlantic Salmon (Salmo Salar). Vaccines 2024, 12, 788. [Google Scholar] [CrossRef]
- Giri, S.S.; Kim, S.G.; Kang, J.W.; Kim, S.W.; Kwon, J.; Lee, S.B.; Jung, W.J.; Park, S.C. Applications of Carbon Nanotubes and Polymeric Micro-/Nanoparticles in Fish Vaccine Delivery: Progress and Future Perspectives. Reviews in Aquaculture 2021, 13, 1844–1863. [Google Scholar] [CrossRef]
- Giri, S.S.; Park, S.C. Application of Carbon Nanotubes in the Advancement of Fish Vaccine. In Biotechnological Advances in Aquaculture Health Management; Gupta, S.K., Giri, S.S., Eds.; Springer Nature: Singapore, 2021; ISBN 9789811651953. [Google Scholar]
- Cimbaluk, G.V.; Ramsdorf, W.A.; Perussolo, M.C.; Santos, H.K.F.; Da Silva De Assis, H.C.; Schnitzler, M.C.; Schnitzler, D.C.; Carneiro, P.G.; Cestari, M.M. Evaluation of Multiwalled Carbon Nanotubes Toxicity in Two Fish Species. Ecotoxicology and Environmental Safety 2018, 150, 215–223. [Google Scholar] [CrossRef]
- Chowdhry, A.; Kaur, J.; Khatri, M.; Puri, V.; Tuli, R.; Puri, S. Characterization of Functionalized Multiwalled Carbon Nanotubes and Comparison of Their Cellular Toxicity between HEK 293 Cells and Zebra Fish in Vivo. Heliyon 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Yan, X.; Pu, Y.; Xiao, F.; Wang, D.; Yang, M. Risks of Single-Walled Carbon Nanotubes Acting as Contaminants-Carriers: Potential Release of Phenanthrene in Japanese Medaka (Oryzias Latipes). Environ. Sci. Technol. 2013, 47, 4704–4710. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zheng, X.; Gao, S.; Huang, Y.; Xiong, J.; Ren, H. Toxicity of Amine-Functionalized Single-Carbon Nanotube (NH2 f-SWCNT) to Channel Catfish (Ietalurus Punetaus): Organ Pathologies, Oxidative Stress, Inflammation, and Apoptosis. Chemosphere 2021, 282, 131133. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Shaw, B.J.; Handy, R.D. Toxicity of Single Walled Carbon Nanotubes to Rainbow Trout, (Oncorhynchus Mykiss): Respiratory Toxicity, Organ Pathologies, and Other Physiological Effects. Aquatic Toxicology 2007, 82, 94–109. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Liu, G.-L.; Ling, F.; Song, L.-S.; Wang, G.-X. Development Toxicity of Functionalized Single-Walled Carbon Nanotubes on Rare Minnow Embryos and Larvae. Nanotoxicology 2015, 9, 579–590. [Google Scholar] [CrossRef]
- Li, Y.; Men, B.; He, Y.; Xu, H.; Liu, M.; Wang, D. Effect of Single-Wall Carbon Nanotubes on Bioconcentration and Toxicity of Perfluorooctane Sulfonate in Zebrafish (Danio Rerio). Science of The Total Environment 2017, 607–608, 509–518. [Google Scholar] [CrossRef]
- Bisesi, J.H.; Ngo, T.; Ponnavolu, S.; Liu, K.; Lavelle, C.M.; Afrooz, A.R.M.N.; Saleh, N.B.; Ferguson, P.L.; Denslow, N.D.; Sabo-Attwood, T. Examination of Single-Walled Carbon Nanotubes Uptake and Toxicity from Dietary Exposure: Tracking Movement and Impacts in the Gastrointestinal System. Nanomaterials 2015, 5, 1066–1086. [Google Scholar] [CrossRef]
- Sohn, E.K.; Chung, Y.S.; Johari, S.A.; Kim, T.G.; Kim, J.K.; Lee, J.H.; Lee, Y.H.; Kang, S.W.; Yu, I.J. Acute Toxicity Comparison of Single-Walled Carbon Nanotubes in Various Freshwater Organisms. BioMed Research International 2015, 2015, 323090. [Google Scholar] [CrossRef]
- Jiang, T.; Alberto Amadei, C.; Gou, N.; Lin, Y.; Lan, J.; D. Vecitis, C.; Z. Gu, A. Toxicity of Single-Walled Carbon Nanotubes (SWCNTs): Effect of Lengths, Functional Groups and Electronic Structures Revealed by a Quantitative Toxicogenomics Assay. Environmental Science: Nano 2020, 7, 1348–1364. [Google Scholar] [CrossRef]
- Wisdom, K.S.; Bhat, I.A.; Chanu, T.I.; Kumar, P.; Pathakota, G.-B.; Nayak, S.K.; Walke, P.; Sharma, R. Chitosan Grafting onto Single-Walled Carbon Nanotubes Increased Their Stability and Reduced the Toxicity in Vivo (Catfish) Model. International Journal of Biological Macromolecules 2020, 155, 697–707. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; González-Ortega, O. Carbon Nanotubes-Based Mucosal Vaccines. In Nanovaccines: An Innovative Technology to Fight Human and Animal Diseases; Rosales-Mendoza, S., González-Ortega, O., Eds.; Springer International Publishing: Cham, 2019; pp. 159–179. ISBN 978-3-030-31668-6. [Google Scholar]
- Gong, Y.-X.; Zhu, B.; Liu, G.-L.; Liu, L.; Ling, F.; Wang, G.-X.; Xu, X.-G. Single-Walled Carbon Nanotubes as Delivery Vehicles Enhance the Immunoprotective Effects of a Recombinant Vaccine against Aeromonas Hydrophila. Fish & Shellfish Immunology 2015, 42, 213–220. [Google Scholar] [CrossRef]
- Liu, L.; Gong, Y.-X.; Liu, G.-L.; Zhu, B.; Wang, G.-X. Protective Immunity of Grass Carp Immunized with DNA Vaccine against Aeromonas Hydrophila by Using Carbon Nanotubes as a Carrier Molecule. Fish & Shellfish Immunology 2016, 55, 516–522. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, G.; Ma, R.; Qi, X.; Wang, G.; Zhu, B.; Ling, F. The Immunoprotective Effect of Whole-Cell Lysed Inactivated Vaccine with SWCNT as a Carrier against Aeromonas Hydrophila Infection in Grass Carp. Fish & Shellfish Immunology 2020, 97, 336–343. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, J.; Liu, G.; Du, H.; Liu, T.; Liu, T.; Li, P.; Yu, Q.; Wang, G.; Wang, E. A Nanocarrier Immersion Vaccine Encoding Surface Immunogenic Protein Confers Cross-Immunoprotection against Streptococcus Agalactiae and Streptococcus Iniae Infection in Tilapia. Fish & Shellfish Immunology 2024, 144, 109267. [Google Scholar] [CrossRef]
- Zhu, B.; Zhang, C.; Zhao, Z.; Wang, G.-X. Targeted Delivery of Mannosylated Nanoparticles Improve Prophylactic Efficacy of Immersion Vaccine against Fish Viral Disease. Vaccines 2020, 8, 87. [Google Scholar] [CrossRef]
- Zhu, B.; Liu, G.-L.; Gong, Y.-X.; Ling, F.; Song, L.-S.; Wang, G.-X. Single-Walled Carbon Nanotubes as Candidate Recombinant Subunit Vaccine Carrier for Immunization of Grass Carp against Grass Carp Reovirus. Fish & Shellfish Immunology 2014, 41, 279–293. [Google Scholar] [CrossRef]
- Qiu, D.-K.; Jia, Y.-J.; Gong, Y.-M.; Zheng, Y.-Y.; Wang, G.-X.; Zhu, B. Optimizing the Immunization Procedure of Single-Walled Carbon Nanotubes Based Vaccine against Grass Carp Reovirus for Grass Carp. Aquaculture 2021, 533, 736152. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, G.-L.; Li, D.-L.; Ling, F.; Zhu, B.; Wang, G.-X. The Protective Immunity against Grass Carp Reovirus in Grass Carp Induced by a DNA Vaccination Using Single-Walled Carbon Nanotubes as Delivery Vehicles. Fish & Shellfish Immunology 2015, 47, 732–742. [Google Scholar] [CrossRef]
- Zhu, B.; Liu, G.-L.; Gong, Y.-X.; Ling, F.; Wang, G.-X. Protective Immunity of Grass Carp Immunized with DNA Vaccine Encoding the Vp7 Gene of Grass Carp Reovirus Using Carbon Nanotubes as a Carrier Molecule. Fish & Shellfish Immunology 2015, 42, 325–334. [Google Scholar] [CrossRef]
- Zheng, Y.-Y.; Qiu, D.-K.; Guo, Z.-R.; Gong, Y.-M.; Wang, G.-X.; Zhu, B. Evaluation of SWCNTs-Loaded DNA Vaccine Encoding Predominant Antigen Epitope VP4-3 against Type II GCRV. Aquaculture 2021, 534, 736197. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, Z.; Liu, G.-Y.; Li, J.; Wang, G.-X.; Zhu, B. Immune Response and Protective Effect against Spring Viremia of Carp Virus Induced by Intramuscular Vaccination with a SWCNTs-DNA Vaccine Encoding Matrix Protein. Fish & Shellfish Immunology 2018, 79, 256–264. [Google Scholar] [CrossRef]
- Zhang, C.; Li, L.-H.; Wang, J.; Zhao, Z.; Li, J.; Tu, X.; Huang, A.-G.; Wang, G.-X.; Zhu, B. Enhanced Protective Immunity against Spring Viremia of Carp Virus Infection Can Be Induced by Recombinant Subunit Vaccine Conjugated to Single-Walled Carbon Nanotubes. Vaccine 2018, 36, 6334–6344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhao, Z.; Zha, J.-W.; Wang, G.-X.; Zhu, B. Single-Walled Carbon Nanotubes as Delivery Vehicles Enhance the Immunoprotective Effect of a DNA Vaccine against Spring Viremia of Carp Virus in Common Carp. Fish & Shellfish Immunology 2017, 71, 191–201. [Google Scholar] [CrossRef]
- Zhang, C.; Zheng, Y.-Y.; Gong, Y.-M.; Zhao, Z.; Guo, Z.-R.; Jia, Y.-J.; Wang, G.-X.; Zhu, B. Evaluation of Immune Response and Protection against Spring Viremia of Carp Virus Induced by a Single-Walled Carbon Nanotubes-Based Immersion DNA Vaccine. Virology 2019, 537, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.-M.; Zhang, C.; Li, Y.; Chen, G.; Wang, G.-X.; Zhu, B. Optimization of Immunization Procedure for SWCNTs-Based Subunit Vaccine with Mannose Modification against Spring Viraemia of Carp Virus in Common Carp. Journal of Fish Diseases 2021, 44, 1925–1936. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, C.; Jia, Y.-J.; Qiu, D.-K.; Lin, Q.; Li, N.-Q.; Huang, Z.-B.; Fu, X.-Z.; Wang, G.-X.; Zhu, B. Immersion Vaccination of Mandarin Fish Siniperca Chuatsi against Infectious Spleen and Kidney Necrosis Virus with a SWCNTs-Based Subunit Vaccine. Fish & Shellfish Immunology 2019, 92, 133–140. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiong, Y.; Zhang, C.; Jia, Y.-J.; Qiu, D.-K.; Wang, G.-X.; Zhu, B. Optimization of the Efficacy of a SWCNTs-Based Subunit Vaccine against Infectious Spleen and Kidney Necrosis Virus in Mandarin Fish. Fish & Shellfish Immunology 2020, 106, 190–196. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, C.; Lin, Q.; Li, N.; Huang, Z.-B.; Zhao, M.; Fu, X.-Z.; Wang, G.-X.; Zhu, B. Single-Walled Carbon Nanotubes as Delivery Vehicles Enhance the Immunoprotective Effect of an Immersion DNA Vaccine against Infectious Spleen and Kidney Necrosis Virus in Mandarin Fish. Fish & Shellfish Immunology 2020, 97, 432–439. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, Y.; Chen, G.; Zhang, C.; Wang, G.-X.; Zhu, B. Protective Immunity against Infectious Spleen and Kidney Necrosis Virus Induced by Mannose Modified Subunit Vaccine with Carbon Nanotubes in Mandarin Fish. Aquaculture Research 2022, 53, 2175–2184. [Google Scholar] [CrossRef]
- Zhao, Z.; Meng, Q.; Sun, T.-Z.; Zhu, B. Mannose Modified Targeted Immersion Vaccine Delivery System Improves Protective Immunity against Infectious Spleen and Kidney Necrosis Virus in Mandarin Fish (Siniperca Chuatsi). Vaccine 2024, 42, 2886–2894. [Google Scholar] [CrossRef]
- Liu, G.-Y.; Wang, E.-L.; Qu, X.-Y.; Yang, K.-C.; Zhang, Z.-Y.; Liu, J.-Y.; Zhang, C.; Zhu, B.; Wang, G.-X. Single-Walled Carbon Nanotubes Enhance the Immune Protective Effect of a Bath Subunit Vaccine for Pearl Gentian Grouper against Iridovirus of Taiwan. Fish & Shellfish Immunology 2020, 106, 510–517. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, G.; Liu, J.; Zhu, B.; Wang, G.; Ling, F. Epitope Screening of the Major Capsid Protein within Grouper Iridovirus of Taiwan and the Immunoprotective Effect with SWCNTs as the Vaccine Carrier. Fish & Shellfish Immunology 2021, 117, 17–23. [Google Scholar] [CrossRef]
- Hu, F.; Li, Y.; Wang, Q.; Wang, G.; Zhu, B.; Wang, Y.; Zeng, W.; Yin, J.; Liu, C.; Bergmann, S.M.; et al. Carbon Nanotube-Based DNA Vaccine against Koi Herpesvirus given by Intramuscular Injection. Fish & Shellfish Immunology 2020, 98, 810–818. [Google Scholar] [CrossRef]
- Hu, F.; Li, Y.; Wang, Q.; Zhu, B.; Wu, S.; Wang, Y.; Zeng, W.; Yin, J.; Liu, C.; Bergmann, S.M.; et al. Immersion Immunization of Koi (Cyprinus Carpio) against Cyprinid Herpesvirus 3 (CyHV-3) with Carbon Nanotube-Loaded DNA Vaccine. Aquaculture 2021, 539, 736644. [Google Scholar] [CrossRef]
- Guo, Z.-R.; Zhao, Z.; Zhang, C.; Jia, Y.-J.; Qiu, D.-K.; Zhu, B.; Wang, G.-X. Carbon Nanotubes-Loaded Subunit Vaccine Can Increase Protective Immunity against Rhabdovirus Infections of Largemouth Bass (Micropterus Salmoides). Fish & Shellfish Immunology 2020, 99, 548–554. [Google Scholar] [CrossRef]
- Liu, G.-Y.; Li, J.; Zhang, Z.-Y.; Liu, J.-Y.; Zhang, C.; Zhu, B.; Wang, G.-X. An Immersion Subunit Vaccine Loaded by Single-Walled Carbon Nanotube Protects Pearl Gentian Grouper from Viral Nervous Necrosis Virus. Aquaculture 2021, 541, 736813. [Google Scholar] [CrossRef]
- Jia, Y.-J.; Guo, Z.-R.; Ma, R.; Qiu, D.-K.; Zhao, Z.; Wang, G.-X.; Zhu, B. Immune Efficacy of Carbon Nanotubes Recombinant Subunit Vaccine against Largemouth Bass Ulcerative Syndrome Virus. Fish & Shellfish Immunology 2020, 100, 317–323. [Google Scholar] [CrossRef]
- Vijayaram, S.; Tsigkou, K.; Zuorro, A.; Sun, Y.-Z.; Rabetafika, H.; Razafindralambo, H. Inorganic Nanoparticles for Use in Aquaculture. Reviews in Aquaculture 2023, 15, 1600–1617. [Google Scholar] [CrossRef]
- Lisuzzo, L.; Cavallaro, G.; Milioto, S.; Lazzara, G. Layered Composite Based on Halloysite and Natural Polymers: A Carrier for the pH Controlled Release of Drugs. New J. Chem. 2019, 43, 10887–10893. [Google Scholar] [CrossRef]
- Pumchan, A.; Sae-Ueng, U.; Prasittichai, C.; Sirisuay, S.; Areechon, N.; Unajak, S. A Novel Efficient Piscine Oral Nano-Vaccine Delivery System: Modified Halloysite Nanotubes (HNTs) Preventing Streptococcosis Disease in Tilapia (Oreochromis Sp.). Vaccines 2022, 10, 1180. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, C.; Xiao, F.; Liu, X.; Xie, A.; Chen, F.; Dong, P.; Lin, P.; Zheng, C.; Zhang, H.; et al. pH-Controlled Release of Antigens Using Mesoporous Silica Nanoparticles Delivery System for Developing a Fish Oral Vaccine. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
An overview of all elaborations made in the two-signal model to effectively explain the mode of action behind several vaccine adjuvants. An effective immunization is usually the result of a well-orchestrated immune response and thus, the synergistic effect of antigens and vaccine adjuvants facilitating certain signals can aid in eliciting a robust and long-lasting protective effect.
Figure 1.
An overview of all elaborations made in the two-signal model to effectively explain the mode of action behind several vaccine adjuvants. An effective immunization is usually the result of a well-orchestrated immune response and thus, the synergistic effect of antigens and vaccine adjuvants facilitating certain signals can aid in eliciting a robust and long-lasting protective effect.
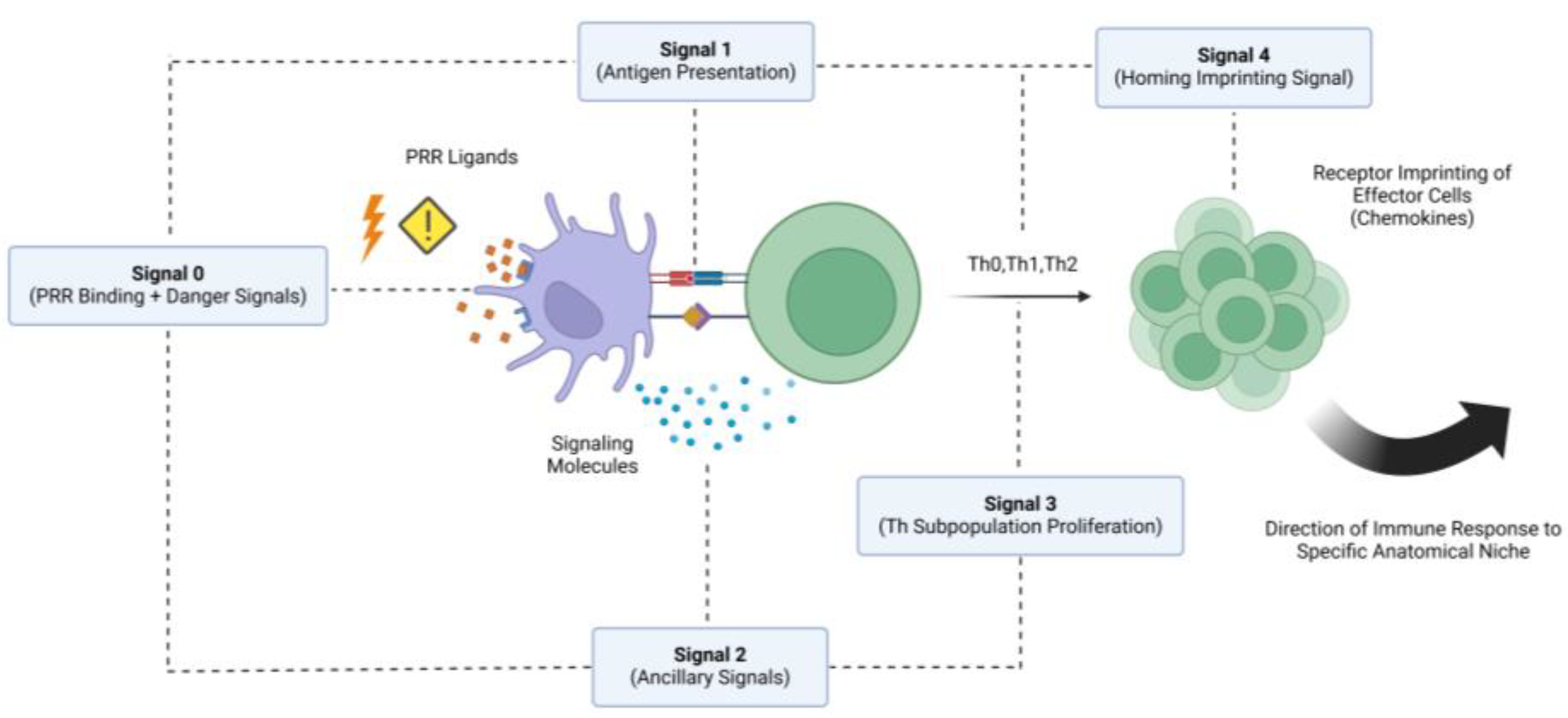
Figure 2.
Schematic illustration showcasing the main types of vaccine adjuvants used conventionally so far in teleost fish vaccinology.
Figure 2.
Schematic illustration showcasing the main types of vaccine adjuvants used conventionally so far in teleost fish vaccinology.

Figure 3.
Potential side effects of current adjuvanted injectable vaccines in teleost fish. In severe cases, the physiological functioning of the digestive system and thus the growth rate can be impaired, potentially undermining animal welfare and lowering the quality of aquatic food products.
Figure 3.
Potential side effects of current adjuvanted injectable vaccines in teleost fish. In severe cases, the physiological functioning of the digestive system and thus the growth rate can be impaired, potentially undermining animal welfare and lowering the quality of aquatic food products.


Table 1.
Cumulative summary of the three major pattern recognition receptor (PRR) types identified in teleost fish according to current and up-to-date literature. It is evident that the teleost fish species is an important parameter in the presence or absence of certain PRRs, as well as in their associated ligands. Homologues of mammalian PRRs have been characterized in a plethora of teleost fish species. This table includes some indicative ones for reference [26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42].
Table 1.
Cumulative summary of the three major pattern recognition receptor (PRR) types identified in teleost fish according to current and up-to-date literature. It is evident that the teleost fish species is an important parameter in the presence or absence of certain PRRs, as well as in their associated ligands. Homologues of mammalian PRRs have been characterized in a plethora of teleost fish species. This table includes some indicative ones for reference [26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42].
| Toll-like Receptors (TLRs) | ||
|---|---|---|
| Receptor | Associated Ligands | Indicative Teleost Fish Species |
| TLR1 | Lipopeptides | Rainbow Trout, Large Yellow Croaker, Carp, Pufferfish, Orange-Spotted Grouper, European Sea Bass, Turbot |
| TLR2 | Lipopeptides, PGN, LTA, Pam3CSK4 | Carp, Catfish, Orange-Spotted Grouper, European Sea Bass, Turbot, Gilthead Seabream |
| TLR3 | dsRNA, poly(I:C) | Carp, Pufferfish, Zebrafish |
| TLR4 | Unknown | Carp, Catfish, Rare Minnow, Zebrafish |
| TLR5 | Flagellin | Atlantic Salmon, Japanese Flounder, Channel Catfish, Gilthead Seabream, Rainbow Trout, Pufferfish, Zebrafish, Turbot |
| TLR7 | ssRNA | Channel Catfish, Grass Carp, Pufferfish, Rainbow Trout, Zebrafish, Turbot |
| TLR8 | ssRNA | Atlantic Salmon, Channel Catfish, Pufferfish, Rainbow Trout, Turbot |
| TLR9 | CpG motifs | Atlantic Salmon, Cobia, Japanese Flounder, Rainbow Trout, Zebrafish, European Seabass, Gilthead Seabream, Turbot |
| TLR13 | rRNA | Atlantic Salmon, Channel Catfish, Orange-Spotted Grouper |
| TLR14 | Unknown | Japanese Flounder, Orange-Spotted Grouper, Pufferfish |
| TLR18 | Unknown | Channel Catfish, Grass Carp, Zebrafish |
| TLR19 | dsRNA | Channel Catfish, Grass Carp, Zebrafish |
| TLR20 | Unknown | Carp, Channel Catfish, Zebrafish |
| TLR21 | CpG motifs | Channel Catfish, Grass Carp, Orange-Spotted Grouper, Zebrafish, Turbot |
| TLR22 | dsRNA, poly(I:C) | Atlantic Cod, Channel Catfish, Grass Carp, Pufferfish, Zebrafish, European Seabass, Turbot, Gilthead Seabream |
| TLR23 | Unknown | Atlantic Cod, Pufferfish |
| TLR25 | Unknown | Channel Catfish, Fathead Minnow, Nile Tilapia |
| TLR26 | LPS, poly(I:C) | Channel Catfish, Yellow Catfish |
| TLR28 | LPS, poly(I:C) | Brown Croaker |
| NOD-like Receptor (NLR) Superfamilies | ||
| NLR-A | LPS, PGN (iE-DAP, MDP), Poly (I:C) | Grass Carp, Nile Tilapia, Rainbow Trout, Channel Catfish, Zebrafish |
| NLR-B | Unknown | Grass Carp |
| NLR-C | Unknown | Grass Carp, Brown Croaker |
| RIG-1-like Receptors (RLRs) | ||
| RIG-1 | dsRNA | Zebrafish |
| MDA5 | dsRNA | Grass Carp |
| LGP2 | dsRNA | Grass Carp |
Table 2.
Commercially available MontanideTM adjuvants intended for use in teleost fish according to the manufacturer, at the time of writing. The vast majority are based on mineral oils and are intended for use via injection vaccination.
Table 2.
Commercially available MontanideTM adjuvants intended for use in teleost fish according to the manufacturer, at the time of writing. The vast majority are based on mineral oils and are intended for use via injection vaccination.
| Currently Available MontanideTM Adjuvants for Use in Aquaculture Vaccinology | ||
|---|---|---|
| Series Name | Technology | Route |
| MontanideTM ISA 61 VG | Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM ISA 50 V2 | Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM ISA 70 VG | Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM ISA 71 VG | Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM ISA 71 R VG | Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM ISA 761 VG | Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM ISA 78 VG | Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM ISA 763B VG | Non-Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM ISA 660 VG | Non-Mineral Oil base for W/O Emulsions | Injection |
| MontanideTM GR | Oil base for W/O Emulsions containing a Protective Matrix | Oral |
| MontanideTM IMS 1312 VG | Combination of Micro-Emulsions with an Immunostimulating Compound | Immersion |
Table 3.
A side-by-side adaptation comparison between Spielberg’s original scale for assessing the Atlantic salmon post-furunculosis vaccination and the newly proposed scale for assessing vaccination side effects in the European seabass by Tziouvas & Varvarigos.
Table 3.
A side-by-side adaptation comparison between Spielberg’s original scale for assessing the Atlantic salmon post-furunculosis vaccination and the newly proposed scale for assessing vaccination side effects in the European seabass by Tziouvas & Varvarigos.
| Score (0-6) | Spielberg Scale [129] (Atlantic Salmon) |
Tziouvas & Varvarigos Scale [128] (European Sea Bass) |
|---|---|---|
| 0 |
|
|
| 1 |
|
|
| 2 |
|
|
| 3 |
|
|
| 4 |
|
|
| 5 |
|
|
| 6 |
|
|
Table 4.
A summary of recent carbon nanotube-based applications in teleost fish vaccinology, based on recent literature. Findings suggest that CNT-based vaccination aligns predominantly with two underutilized vaccine technologies in commercial applications, recombinant subunit and DNA vaccine platforms, indicating a potential shift in the future of aquaculture vaccines thanks to nanoparticle formulations.
Table 4.
A summary of recent carbon nanotube-based applications in teleost fish vaccinology, based on recent literature. Findings suggest that CNT-based vaccination aligns predominantly with two underutilized vaccine technologies in commercial applications, recombinant subunit and DNA vaccine platforms, indicating a potential shift in the future of aquaculture vaccines thanks to nanoparticle formulations.
| CNT Type | Vaccine Technology | Delivery Route | Pathogen (Antigen) | Teleost Species | Source |
|---|---|---|---|---|---|
| Single-Walled | Recombinant Subunit |
Intramuscular Injection & Bath Immersion |
Aeromonas hydrophila (aerA) |
Grass Carp | [192] |
| Single-Walled | DNA Vaccine | Intramuscular Injection |
Aeromonas hydrophila (aerA) |
Grass Carp | [193] |
| Single-Walled | Whole-cell Inactivated Vaccine |
Intraperitoneal Injection & Bath Immersion |
Aeromonas hydrophila (Bacterial Lysate) |
Grass Carp | [194] |
| Single-Walled | Recombinant Subunit |
Bath Immersion |
Streptococcus sp. (rSip) |
Tilapia | [195] |
| Single-Walled (Mannose Modified) |
Recombinant Subunit |
Bath Immersion | GCRV (VP7) | Grass Carp | [196] |
| Single-Walled | Recombinant Subunit |
Bath Immersion | GCRV (VP7) | Grass Carp | [197] |
| Single-Walled | Recombinant Subunit |
Bath Immersion | GCRV (VP4-3) | Grass Carp | [198] |
| Single-Walled | DNA Vaccine | Intramuscular Injection & Bath Immersion |
GCRV (VP4-3, VP5, VP7) | Grass Carp | [199,200,201] |
| Single-Walled | DNA Vaccine | Intramuscular Injection | SVCV (M) | Common Carp | [202] |
| Single-Walled | Recombinant Subunit |
Intramuscular Injection & Bath Immersion |
SVCV (G) | Common Carp | [203] |
| Single-Walled | DNA Vaccine | Intramuscular Injection & Bath Immersion |
SVCV (G) | Common Carp | [204] |
| Single-Walled | DNA Vaccine | Bath Immersion | SVCV (M) | Common Carp | [205] |
| Single-Walled (Mannose Modified) |
Recombinant Subunit |
Bath Immersion | SVCV (G) | Common Carp | [206] |
| Single-Walled | Recombinant Subunit | Bath Immersion | ISKNV (MCP) | Mandarin Fish | [207,208] |
| Single-Walled | DNA Vaccine | Bath Immersion | ISKNV (MCP) | Mandarin Fish | [209] |
| Single-Walled (Mannose Modified) |
Recombinant Subunit | Intramuscular Injection & Bath Immersion |
ISKNV (MCP) | Mandarin Fish | [210,211] |
| Single-Walled | Recombinant Subunit | Bath Immersion | TGIV (MCP, P2) | Pearl Gentian Grouper | [212,213] |
| Single-Walled | DNA Vaccine | Intramuscular Injection & Bath Immersion |
KHV (ORF 149) | Koi Fish | [214,215] |
| Single-Walled | Recombinant Subunit | Bath Immersion | MSRV (G) | Largemouth Bass | [216] |
| Single-Walled | Recombinant Subunit | Bath Immersion | NNV (MCP) | Pearl Gentian Grouper | [217] |
| Single-Walled | Recombinant Subunit | Bath Immersion | LBUSV (MCP) | Largemouth Bass | [218] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



