
Submitted:
04 November 2024
Posted:
05 November 2024
You are already at the latest version
Abstract
An essential role in proper homeostasis of glucose is played by kidneys where glucose tranport is carried out across cell membranes by two families of glucose tansporters - facilitated diffusion glucose transporters (GLUTs) and Na (+) -dependent glucose co-transporters (SGLT family). Among transporters, sodium-dependent glucose co-transporters have a major role in the kidney ‘s ability to reabsorb glucose. Although localization of glucose transporters has been extensively studied in mammals there are still gaps in knowledge of localization of SGLTs in birds. The aim of the reasarch was to study comparatively immunolocalization of sodium-dependent glucose co-transporters SGLT1 and SGLT2 in kidneys of healthy and T-2 mycotoxicated chickens. Immunohistochemical staining was carried out using polyclonal primary antibodies SGLT1 and SGLT2 (Abcam, UK) on kidney tissue derived from three healthy and three T-2 mycotoxicated 7 days old female layer-type Ross chickens (Gallus gallus domesticus). Sections were stained using Immunohistochemistry kit (Abcam, UK). In kidneys of healthy birds strong staining of SGLT1 and SGLT2 was noted in cytoplasm of epithelial cells of the proximal straight and convoluted tubules. In T-2 toxin group birds' kidneys’ weak expression of SGLT1 and SGLT2 with morphological changes occurred, indicating to reduced glucose transport in urinary system during T-2 mycotoxicosis.
Keywords:
chicken
; immunohistochemistry
; sodium-dependent glucose co-transporter
; mycotoxicosis
1. Introduction
As glucose is the primary source of energy for all living organisms, the homeostasis of it is particulary important [1]. In the body’s glucose homeostasis, the kidneys play a major role in not only filtering but also in absorbing the glucose [2]. The primary filtration unit in the kidneys, the glomerulus, filters the glucose, which is then reabsorbed in the kidney’s proximal convoluted tubule by the sodium-dependent glucose co-transporters SGLT1 and SGLT2 as the glucose transport across the apical brush border of renal epithelial cells requires the presence of a sodium gradient [3,4,5,6]. The glucose transport across the cell membrane is mediated by two families of glucose transporters: the facilitated diffusion glucose transporters (GLUTs) and the Na (+) -dependent glucose co-transporters (SGLT family) [7]. GLUTs and SGLTs vary in their substrate specificity, regulatory mechanisms as well as their distribution [8]. Whilst the GLUTs transport glucose across the plasma membrane by the mechanism of a facilitated diffusion,the sodium-dependent glucose co-transporters belong to the family of active glucose transporters which simultaneously transport sodium and glucose using a concentration gradient [9]. A low intracellular sodium concentration is created and maintained by the basolaterally located sodium-potassium-adenosine triphosphatase pump forcing through the basolateral membrane intracellular sodium out of the cell after which the electrochemical gradient provides the driving force for the transport of sodium into the cell across the apical membrane and the glucose transport by specific sodium-dependent glucose transporters (SGLTs) [10]. When glucose is concentrated in epithelial cells to a level that exceeds the interstitial glucose level, it diffuses into the interstitium through specific facilitative glucose transporters located on the basolateral membrane. The facilitative glucose transporter GLUT2 mediates glucose transport across the basolateral membrane along its chemical gradient. Basolateral GLUT1 may help to reabsorb glucose or take up glucose from the peritubular space. Figure 1 represents the mechanism of transmembrane glucose transport by SGLT2, which is situated at the brush border of epithelial cells [11].
SGLT2 = Na (+) -dependent glucose co-transporter 2
GLUT2 = facilitative glucose transport
Since SGLT2 carries out 80–90% of glomerular filtered glucose reabsorption it is considered the main co-transporter involved in glucose reabsorption in the kidney [12,13]. The remainder of the glucose absorption, which is approximately 10%, is carried out by SGLT1 [4,14]. The localization of glucose transporters in the gastrointestinal tract of birds has previously been extensively studied and these studies have shown differences of glucose transporter immunohistochemical staining in different age groups of laying chickens and ostriches [15,16]. However, the immunolocalization of sodium-dependent glucose co-transporters in the kidney tissue of birds has only recently started [11,16,17]. Because the kidneys of avians possess different types of glomeruli: the mammalian type and the reptilian glomeruli type [3], they differ significantly from mammalian kidneys.
Mycotoxins, toxic secondary metabolites, are produced by filamentous fungi belonging to the phylum Ascomycota or molds, having a great importance in the health of humans and animals as being the cause of acute and chronic diseases [18]. Of the approximately 400 mycotoxins, 30 are considered as a threat to human or animal health, among which aflatoxins, ochratoxins, fumonisins and trichothecenes are the most important. Trichothecenes are groups of mycotoxins that are produced by diverse filamentous fungal species including Fusarium, Myrothecium, Stachybotrys, Trichoderma, Trichothecium, and Spicellum [19,20,21]. Among mycotoxins, T-2 toxin is the most toxic trichothecenes, and it is produced by different Fusarium species, including F. sporotrichioides, F. poae and F. acuminatum. The presence of these Fusarium species has been documented to occur in moderate to cold climates and wet storage conditions [22,23]. F. sporotrichioides, is the main T-2 producer and it is able to grow in a wide range of temperatures ranging from −2 to 35 °C [24]. Toxins produced by T-2 are a significant threat to human and animal health. It can increase susceptibility to infections by immunosuppression, are able to cause necrosis in the digestive tract and dystrophy in other organs like liver, kidneys, heart, brain or the peripheral ganglia of vegetative nervous system [25,26,27]. In poultry consumption of feed contaminated with T-2 reduces chicken’s weight gain, egg production and hatching ability [28]. The T-2 toxin is hypothesized to inhibit protein synthesis through binding and inactivation of peptidyl-transferase at the site of DNA transcription [29,30]. The main sources of Trichothecenes include contaminated wheat, barley, rye, oats, and maize, however they also occur in hay, straw, green feed as well as silage from contaminated cereals [26,31,32,33]. Trichothecenes can enter the human food chain by consumption of products of animal origin such as meat, milk and eggs that are derived from livestock fed with Trichothecene contaminated feed [31,34,35]. Although the localization of the sodium-dependent glucose co-transporters is relatively well established in mammal´s kidney tissue, the same comprehensive scientific studies on the immunolocalization of SGLT1 and SGLT2 in the kidney of healthy and diseased birds are lacking.
Since up to now there is a need for comparative studies about the effect of T-2 mycotoxin on the expression of glucose transporters, specifically sodium-dependent glucose co-transporters-1(SGLT1) and -2 (SGLT2) in avian kidneys, the aim of our experimental investigation was to immunolocalize the sodium-dependent glucose co-transporters-1 and -2 comparatively in healthy and T-2 intoxicated laying-type chicken.
2. Materials and Methods
Kidney material was collected from three 7-days-old healthy (control group) and three 7-days-old female layer-type broilers with T-2 toxicosis (T-2 toxin group). The Ross broilers (Gallus gallus domesticus), obtained from a commercial Macedonian hatchery, were placed in temperature-controlled brooders and raised in standard conditions with having free access to food and water. For the T-2 toxin group, starting from fourth day after hatching, T-2 toxin was applied in a dose of 0.250 mg/day/bird and was compulsive for three consecutive days. 24 h after the last dosage of the T-2 mycotoxin, the chickens were sacrificed by intracardiac overdose of 0.5 mL 20% sodium pentobarbital and the kidneys were removed. Specimens 0.5-1.0 cm in diameter were fixed in 10% buffered formalin solution for at least 72 hours, to ensure complete fixation. Thereafter the specimens were embedded into paraffin and slices 7 μm in thickness were cut. The prepared slides underwent standard Hematoxylin and eosin staining method [36] and immunohistochemical staining with polyclonal primary antibodies Rabbit anti-SGLT-1 and Rabbit anti-SGLT-2 (Abcam, UK). To immunolocalize SGLT1 and SGLT2 in chicken kidney tissue the sections were stained using an Immunohistochemistry kit (Abcam, UK) containing the secondary antibody, according to the manufacturer’s guidelines (IHC kit, Abcam, UK). The sections were pre-treated using heat-mediated antigen retrieval with sodium citrate buffer (pH 6) for 20 minutes, thereafter, incubated with the primary rabbit polyclonal antibodies Rabbit anti-SGLT1 and Rabbit anti-SGLT2 (Abcam, UK) at 1:1000 dilution for 30 min at 37 ºC. Biotinylated secondary antibody at 1:1000 dilution and streptavidin-conjugated peroxidase were used for detection using DAB as chromogen. Negative controls contained antibody diluent (Dako, S0809) instead of primary antibodies and ostriches’ chicken’s kidney tissue sections for identifying SGLT1 and SGLT2 were used as positive controls (Hussar et al. 2022).
The immunolocalization of SGLT1 and SGLT2 was studied by eye-visual control by two scientists in blind analysis and visualized photographically using an AxioCam HRc camera (Germany) connected to a Zeiss Axioplan-2 Imaging microscope (Germany).
The Ethical Committee of Ss. Cyril and Methodius University in Skopje, in conformity with the recommendation provided in the European Convention for the Protection of Vertebrate Animals used for Experimental and Other Scientific Purposes (ETS no.123, Approval No. 03-7534, 12.04.2013), approved the husbandry and experimental procedures of the study.
3. Results
3.1. Routine Histology
3.2. Immunohistochemistry
The results of the immunohistochemical study revealed the immunolocalization of the major glucose transporters in the kidney- SGLT1 and SGLT2 noticed in the proximal renal tubules of hen chickens in both study groups (Figure 3A; Figure 4A). In the group of healthy chicken’s strong expression of SGLT1 occurred in the apical part of the epithelial cells of renal proximal tubules (Figure 3A).
Compared to the healthy birds, in the T-2 toxin group the expression of the both studied antibodies was weaker, brush border membranes of proximal tubule’s epithelial cells were irregular and damaged (Figure 3B; Figure 4B).
In both studied groups no specific staining for SGLT1 and SGLT2 was observed in the distal tubules and collecting ducts of the birds’ kidneys (Figure 5).
Comparatively for both studied antibodies, the staining of the renal tissue of the control group was more intense than that of the T-2 toxin group. Furthermore, the expression of SGLT2 was noted to be comparatively much stronger than the staining of SGLT1. The results of the descriptive (eye visual) analysis on the expression through the intensity of staining of SGLT1 and SGLT2 in the proximal tubules are shown in Table 1
4. Discussion
In maintenance of the body’s homeostasis the role of kidneys is significant. Histologically kidneys consist of two zones – the cortex and the medulla. The morphofunctional unit of the kidney is the nephron. Funcionally nephrons consists of glomeruli and tubules. Nephrons are responsible for filtering the blood plasma to eliminate waste products as well as conserving glucose and water. In birds, the homeostasis of fluid and ions needs the proper functioning of several organ systems and is a more complex phenomenon than in other vertebrates [37]. Unlike mammals, the kidneys of chicken contain different types of glomeruli, all noticed in the cortical region: small reptilian type glomerulus near the surface of cortex; larger, mammalian type, glomerulus near medulla and intermediate type glomerulus in the cortex deeper regions. The kidney maintains the glucose homeostasis by reabsorption of glucose in the proximal tubules, release of glucose into the circulation via gluconeogenesis and uptake of glucose from the circulatory sytem to provide itself with energy [6]. Glomeruli are the primary filtration units, because they filter glucose from plasma, which is then reabsorbed through glucose transporter proteins that are found in the cell membranes within the proximal tubules of the kidney. The proximal renal tubule, a part of the nephron, can be further divided into two sections, the proximal convoluted tubule and the proximal straight tubule. These two sections are subdivided into ultrastructural divisions, so-called segments, that consist either of higher or lower cell complexity. While the S1 segment corresponds to the convoluted proximal tubule, the straight proximal tubule has the correspondence of the S2 and S3 [38]. The studies on mammalian renal tissue have shown that the kidney’s proximal tubules contain the necessary enzymes for gluconeogenesis (lactate, glutamine, glycerol, alanine) as well as the main transporters which are active in the process of glucose reabsorption - the Na (+) -dependent glucose co-transporters SGLT2 and SGLT1 on the apical membrane and the facilitated diffusion glucose transporter GLUT2 on the basolateral membrane. Glucose reabsorbed or produced in the proximal tubule (gluconeogenesis) is mainly absorbed in the peritubular capillaries and returned to the systemic circulation or used as an energy source in the distal tubule segments, which take up glucose via basolateral GLUT1. The SGLTs are considered active transporters and GLUTs belong to the family of passive transporters [39]. In our previous study the immunolocalization of SGLT1 and SGLT2 has been revealed in the kidneys of chickens of different ages [40]. For SGLT1 strong staining in the apical parts of the epithelial cells of the straight proximal tubules was noted and for SGLT2 strong staining of the renal proximal tubules and unstained distal tubules were observed in kidneys of laying chicken of different ages. The immunohistochemical localization of SGLT1 and SGLT2 in the kidneys of hen chickens of different ages was detected on the apical side of the epithelial cells of the proximal renal tubules. The findings on the localization of the sodium-dependent glucose co-transporters SGLT1 and SGLT2 in renal tissue of chickens of different age groups of the previous studies are in accordance with the findings of our current study on young laying chicken where the immunolocalization of SGLT1 and SGLT2 was detected on the apical membrane of the epithelial cells of the chicken kidney’s proximal tubules.
Mycotoxins are low-molecular-weight secondary metabolites of fungi [41]. Mycotoxins can be divided according to the organ affected as nephro-, hepato-, immunotoxic etc. or into general groups such as allergens, teratogens or carcinogens etc. [42]. The T-2 toxin studied in our current experiment, is a carcinogenic mycotoxin that belongs in the trichothecene family to the group of Fusarium mycotoxins that is widely encountered as natural contaminant known to elicit toxin responses in different organs and tissues [43]. Mycotoxins, which are secondary metabolites produced by several fungal species, can exert severe toxic effects on humans and animals [44]. In the trichothecene family, the T-2 mycotoxin is amongst the most toxic members as compared to other mycotoxins causing serous hemorrhagic inflammation, necrosis and ulceration in the digestive tract, dystrophy in kidney, liver, heart, brain and peripheral ganglia of the vegetative nervous system. Additionally, T-2 has been shown to reduce egg production and weight gain of chickens, as well as impeding the ability of egg hatching [45]. In poultry the T-2 toxin also has been observed as the causative agent for the impairment of immune responses, destruction of the hematopoietic system, declining egg production, the serous-haemorrhagic necrotic-ulcerative inflammation of the digestive tract, internal hemorrhaging, mouth and skin lesions [44]. Pathohistological studies usually reveal fatty changes and strong granular degeneration mainly in the kidneys and liver. In chronically T-2 intoxicated poultry, interstitial nephritis, glomerunephritis and kidney sclerosis are among the main pathologies that can be observed [46]. As the kidneys are among the main affected organs by T-2 mycotoxin, having an essential role in the glucose homeostasis, and as the immunolocalization of sodium-dependent glucose transporters in birds’ kidney tissue has not yet been fully elucidated, our current study was carried out on domestic chickens’ kidney material, investigating the effect of T-2 on SGLT1 and SGLT2 - members of the SLC5A gene family contributing to the renal glucose reabsorption [47]. Compared to the staining of SGLT1 in chicken kidney tissue our study revealed a strong staining of SGLT2 in the epithelial cells of the renal proximal tubules, presumambly because SGLT2 is considered as the main glucose transporter involved in glucose reabsorption in the kidneys carrying out 80–90% of glomerular filtered glucose reabsorption in renal tubular system [12,13]. In eye visual control it could be observed that compared to SGLT1, which localized mainly only in the apical region of the epithelial cells, SGLT2 was immunolocalized throughout the epithelial cells of the proximal tubules. In the T-2 toxin group pale staining of proximal renal tubules was noted- the weak expression of both studied antibodies and the morphological changes in the T-2 intoxicated bird group noted in our current study suggest the reduced glucose transport caused by the intoxication. Furthermore, irregular and damaged brush border membranes of proximal tubule’s epithelial cells in the T-2 toxicated group chicken has been observed in our present study which is also in accordance with previous studies showing that T-2 induces oxidative stress with detrimental effects like nuclear and mitochondrial DNA damage, disturbances in cell-signaling as the toxins can affect the cell cycle and lead to apoptosis and ultimatively death of the cells [48,49,50]. A few of the T-2 main toxic effects on the kidney specifically are apoptosis of proximal convoluted tubule’s epithelial cells, degeneration of the epithelial lining, swelling and vacuolar degeneration of the tubular epithelium and necrosis of renal tubules [25]. These pathological alterations indicate the nephrotoxicity of T-2 and are in accordance with the findings of our study. The pale staining of proximal renal tubules in the T-2 toxin group of both major glucose transporters in kidneys indicates to the reduced ability to adequately carry out glucose transport caused by T-2 mycotoxicosis supporting the hypothesis that there are morphological changes in the kidney tissue during intoxication. To specify the glucose transporters’ localization and clarify the patomorphological changes in laying chicken’s kidney tissue during mycotoxicosis in greater detail, more studies using different research methods such as Immunofluorescence or Western blot should be carried out in the future.
5. Conclusions
The study revealed the immunolocalization of SGLT1 and SGLT2 in kidney tissue of healthy and intoxicated young female chicks. The sodium-dependent glucose co-transporters SGLT1 and SGLT2 were localized in the epithelial cells of the renal proximal tubules. The expression of SGLT2 was noticed to be comparatively stronger than the expression of SGLT1 which localized mainly in the apical membranes of the epithelial cells of the proximal tubules. Comparing the expression of the studied antibodies in the kidneys of the chicken of the two study groups, the expression of both antibodies was relatively weaker in the kidneys of the chicks belonging to the T-2 toxin group. Together with the morphological changes- irregular and damaged brush border membranes of proximal tubule’s epithelial cells, the weak expression of both major glucose transporters SGLT1 and SGLT2 in the kidney tissue of the chicken in T-2 toxin group may indicate damage of the kidney tissue and subsequently a reduced functionality of glucose transport in the kidneys during T-2 mycotoxicosis.
Author Contributions
Conceptualization, Piret Hussar, Cristin Allmang, Florina Popovska-Percinic; Ilmārs Dūrītis.; methodology, Piret Hussar, Cristin Allmang, Florina Popovska-Percinic,; software, Cristin Allmang, Piret Hussar; validation, Piret Hussar, Cristin Allmang, Florina Popovska-Percinic; Ilmārs Dūrītis.; formal analysis, Cristin Allmang, Piret Hussar; investigation, Cristin Allmang, Piret Hussar; resources, Florina Popovska-Percinic, Cristin Allmang, Piret Hussar; data curation, Cristin Allmang, Piret Hussar; writing—original draft preparation, Cristin Allmang; writing—review and editing, Piret Hussar, Cristin Allmang, Florina Popovska-Percinic; Ilmārs Dūrītis; visualization, Cristin Allmang, Piret Hussar; supervision, Piret Hussar, Ilmārs Dūrītis; project administration, Piret Hussar, Ilmārs Dūrītis, Florina Popovska-Percinic. All authors have read and agreed to the published version of the manuscript.
Funding
Financing based on the EEC performance agreement.
Institutional Review Board Statement
The Ethical Committee of Ss. Cyril and Methodius University in Skopje, in conformity with the recommendation provided in the European Convention for the Protection of Vertebrate Animals used for Experimental and Other Scientific Purposes (ETS no.123, Approval No. 03-7534, 12.04.2013), approved the husbandry and experimental procedures of the study.
Data availability statement: The original contributions presented in this study are included in the article material. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors wish to thank Mare Tamm for her laboratory assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Hruby, V.J. In Molecular and Cellular Endocrinology, Bittar, E. ; Bittar, N., Eds.; JAI Press Inc., Greenwhich, USA, 1997, 10, 387–401. [Google Scholar]
- Mota, M.; Mota, E.; Dinu, I.R. In Treatment of Type 2 Diabetes; Croniger, C., Ed.; IntechOpen: Rijeka, Crotaia, 2015, Chapter 1, pp.13-17.
- König, H.E.; Korbel, R.; Liebich, H.G. Avian Anatomy textbook and colour atlas; 5m Publishing: Sheffield, UK, 2016; Volume 2, pp: 131-133. [Google Scholar]
- Vallon, V.; Thomson, S.C. Renal Function in Diabetic Disease Models: The Tubular System in the Pathophysiology of the Diabetic Kidney. Annu. Rev. Physiol. 2012, 74, 351–375. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.; Eckstein, N.; Pfeifer, V.; Mayer, P.; Hass, M.D.S. Efficacy, Safety and Regulatory Status of SGLT2 Inhibitors: Focus on Canagliflozin. Nutr. Diabetes. 2014, 4, e143. [Google Scholar] [CrossRef] [PubMed]
- Mather, A.; Pollock, C. Glucose Handling by the Kidney. Kidney Int. 2011, 79, S1–S6. [Google Scholar] [CrossRef]
- Takata, K. Glucose Transporters in the Transepithelial Transport of Glucose. J. Electron Microsc. 1996, 45, 275–284. [Google Scholar] [CrossRef]
- Sano, R.; Shinozaki, Y.; Ohta, T. Sodium–Glucose Cotransporters: Functional Properties and Pharmaceutical Potential. J. Diabetes Investig. 2020, 11, 770–782. [Google Scholar] [CrossRef]
- Navale, A.M.; Paranjape, A.N. Glucose Transporters: Physiological and Pathological Roles. Biophys. Rev. 2016, 8, 5–9. [Google Scholar] [CrossRef]
- Raja, M.; Puntheeranurak, T.; Hinterdorfer, P.; Kinne, R. SLC5 and SLC2 Transporters in Epithelia—Cellular Role and Molecular Mechanisms. In Current Topics in Membranes; Mark O. Bevensee Ed.; Academic Press, 2012; 70, pp.29-76.
- Allmang, C.; Hussar, P.; Järveots, T.; Duritis, I. Sodium-glucose co-transporter SGLT2 in kidneys of chicken in different ages. In ACB 2024, 57, 300: 21st Congress of the International Federation of Associations of Anatomists in conjunction with the 74th Annual Meeting of the Korean Association of Anatomists, Kimdaejung Convention Center, Gwangju, South-Korea, 5-8.09.2024.
- You, G.; Lee, W.-S.; Barros, E.J.G.; Kanai, Y.; Huo, T.-L.; Khawaja, S.; Wells, R.G.; Nigam, S.K.; Hediger, M.A. Molecular Characteristics of NA+-Coupled Glucose Transporters in Adult and Embryonic Rat Kidney. J. Biol. Chem. 1995, 270, 29365–29371. [Google Scholar] [CrossRef]
- Bonora, B.M.; Avogaro, A.; Fadini, G.P. Extraglycemic Effects of SGLT2 Inhibitors: A Review of the Evidence. Diabetes Metab. Syndr. Obes. 2020, 13, 161–174. [Google Scholar] [CrossRef]
- Horiba, N.; Masuda, S.; Takeuchi, A.; Takeuchi, D.; Okuda, M.; Inui, K.-I. Cloning and Characterization of a Novel NA+-Dependent Glucose Transporter (NAGLT1) in Rat Kidney. J. Biol. Chem. 2003, 278, 14669–14676. [Google Scholar] [CrossRef] [PubMed]
- Hussar, P.; Dūrītis, I.; Popovska-Percinic, F.; Järveots, T. Short communication : Immunohistochemical study of sodium-dependent glucose co-transporters in ostriches kidneys. Agraarteadus J. Agric. Sci. 2020, 31, 147–150. [Google Scholar] [CrossRef]
- Hussar, P.; Allmang, C.; Popovska-Percinic, F.; Järveots, T.; Dūrītis, I. Comparative Study of Sodium-Dependent Glucose Co-Transporters in Kidneys of Ostrich Chickens. Sci. Hori. 2022, 25, 30–35. [Google Scholar] [CrossRef]
- Hussar, P.; Kaerner, M.; Duritis, I.; Plivca, A.; Pendovski, L.; Jaerveots, T.; Popovska-Percinic, F. Temporospatial Study of Hexose Transporters and Mucin in the Epithelial Cells of Chicken (Gallus Gallus Domesticus) Small Intestine. Pol. J. Vet. Sci. 2017, 20, 627–633. [Google Scholar] [CrossRef]
- Varzakas, T.; Agriopoulou, S.; Stamatelopoulou, E.; Mycotoxin. Encyclopedia. 2024. Available online: https://encyclopedia.pub/entry/22939 (accessed on 26 September 2024).
- Nazari, L.; Pattori, E.; Terzi, V.; Morcia, C.; Rossi, V. Influence of Temperature on Infection, Growth, and Mycotoxin Production by Fusarium Langsethiae and F. Sporotrichioides in Durum Wheat. Food Microbiol. 2014, 39, 19–26. [Google Scholar] [CrossRef]
- Nathanail, A.V.; Varga, E.; Meng-Reiterer, J.; Bueschl, C.; Michlmayr, H.; Malachova, A.; Fruhmann, P.; Jestoi, M.; Peltonen, K.; Adam, G.; et al. Metabolism of the Fusarium Mycotoxins T-2 Toxin and HT-2 Toxin in Wheat. J. Agric. Food. Chem. 2015, 63, 7862–7872. [Google Scholar] [CrossRef]
- Nayakwadi, S.; Ramu, R.; Sharma, A.K.; Gupta, V.K.; Rajukumar, K.; Kumar, V.; Shirahatti, P.S.; L, R.; Basalingappa, K.M. Toxicopathological Studies on the Effects of T-2 Mycotoxin and Their Interaction in Juvenile Goats. PLoS One. 2020, 15, e0229463. [Google Scholar] [CrossRef]
- Edwards, S.G.; Imathiu, S.M.; Ray, R.V.; Back, M.; Hare, M.C. Molecular Studies to Identify the Fusarium Species Responsible for HT-2 and T-2 Mycotoxins in UK Oats. Int. J. Food Microbiol. 2012, 156, 168–175. [Google Scholar] [CrossRef]
- Lippolis, V.; Pascale, M.; Maragos, C.M.; Visconti, A. Improvement of Detection Sensitivity of T-2 and HT-2 Toxins Using Different Fluorescent Labeling Reagents by High-Performance Liquid Chromatography. Talanta 2008, 74, 1476–1483. [Google Scholar] [CrossRef]
- Kiš, M.; Vulić, A.; Kudumija, N.; Šarkanj, B.; Tkalec, V.J.; Aladić, K.; Škrivanko, M.; Furmeg, S.; Pleadin, J. A Two-Year Occurrence of Fusarium T-2 and HT-2 Toxin in Croatian Cereals Relative of the Regional Weather. Toxins. 2021, 13, 39. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Podogrocki, M.; Ceremuga, M.; Stela, M.; Bijak, M. T-2 Toxin—The Most Toxic Trichothecene Mycotoxin: Metabolism, Toxicity, and Decontamination Strategies. Molecules. 2021, 26, 6868. [Google Scholar] [CrossRef] [PubMed]
- Male, D.; Wu, W.; Mitchell, N.J.; Bursian, S.; Pestka, J.J.; Wu, F. Modeling the Emetic Potencies of Food-Borne Trichothecenes by Benchmark Dose Methodology. Food Chem. Toxicol. 2016, 94, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Oswald, I. Effect of Deoxynivalenol and Other Type B Trichothecenes on the Intestine: A Review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef] [PubMed]
- Chi, M.S.; Mirocha, C.J.; Kurtz, H.J.; Weaver, G.; Bates, F.; Shimoda, W. Effects of T-2 Toxin on Reproductive Performance and Health of Laying Hens, Poult. Sci. 1977, 56, 628–637. [Google Scholar] [CrossRef]
- Henghold, W.B. 2nd. Other biologic toxin bioweapons: ricin, staphylococcal enterotoxin B, and trichothecene mycotoxins. Dermatol Clin, 22. [CrossRef]
- Afsah-Hejri, L.; Jinap, S.; Hajeb, P.; Radu, S.; Shakibazadeh, S. A Review on Mycotoxins in Food and Feed: Malaysia Case Study. Compr Rev Food Sci Food Saf. 2013, 12, 629–651. [Google Scholar] [CrossRef]
- Cope, R.B. In Veterinary Toxicology; 3rd ed.; Gupta, R.C., Ed., Academic Press: Cambridge, MA, USA, 2018, 3, 1043–1053.
- Lancova, K.; Hajslova, J.; Poustka, J.; Krplova, A.; Zachariasova, M.; Dostalek, P.; Sachambula, L. Transfer ofFusariummycotoxins and ‘Masked’ Deoxynivalenol (Deoxynivalenol-3-Glucoside) from Field Barley through Malt to Beer. Ood Addit. Contam. - Chem. Anal. Control Expo. Risk Assess. 2008, 25, 732–744. [Google Scholar] [CrossRef]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in Cereal Grains – an Update. Toxins. 2019, 11, 634. [Google Scholar] [CrossRef]
- He, J.; Zhou, T.; Young, J.C.; Boland, G.J.; Scott, P.M. Chemical and Biological Transformations for Detoxification of Trichothecene Mycotoxins in Human and Animal Food Chains: A Review. Trends Food Sci. Technol. 2010, 21, 67–76. [Google Scholar] [CrossRef]
- Meneely, J.; Greer, B.; Kolawole, O.; Elliott, C. T-2 and HT-2 Toxins: Toxicity, Occurrence and Analysis: A Review. Toxins 2023, 15, 481. [Google Scholar] [CrossRef]
- Carson, F.L. Histotechnology: A Self-Instructional Text; 2nd ed. ASCP Press: Chicago, IL, USA, 1997. [Google Scholar]
- Deepa, K.P.; Sreeranjini, A.R.; Soumya, C.B.; Mayadany, S.; Sunilkumar, N.S.; Sumena, K.B. Comparative histological studies on the renal medulla in broiler chicken and broiler duck. Int J Vet Sci Anim Husbandry 2021, 6, 11–14. [Google Scholar] [CrossRef]
- Boron, W.F.; Boulpaep, E.L. Medical physiology. 3rd ed.; Elsevier, Philadelphia, USA, 2016; p. 727.
- Triplitt, C.L. Understanding the kidneys' role in blood glucose regulation. Am J Manag Care. 2012, 18, S11–S16. [Google Scholar] [PubMed]
- Allmang, C.; Hussar, P. ; Duriti,s I. ; Järveots, T. Immunolocalization of sodium-dependent glucose co-transporter 1 and sodium-dependent glucose co-transporter 2 in chicken’s (Gallus gallus domesticus) kidneys, Pol. J. Vet. Sci. 2024, 27, 323–328. [Google Scholar] [CrossRef]
- Bhatnagar, D.; Yu, J.; Ehrlich, K.C. Toxins of filamentous fungi. Chem Immunol. 2002, 81, 167–206. [Google Scholar] [CrossRef] [PubMed]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc., 2011, 2, 129–144. [Google Scholar] [CrossRef]
- Türker, L.; Gümüş, S. A Theoretical Study on Vomitoxin and Its Tautomers. J. Hazard. Mater. 2009, 163, 285–294. [Google Scholar] [CrossRef]
- Devreese, M.; De Backer, P.; Croubels, S. Verschillende Methoden Om Mycotoxineproductie En de Impact Op de Diergezondheid Tegen Te Gaan. Vlaams Diergeneeskundig Tijdschrift. 2013, 82. [Google Scholar] [CrossRef]
- Adhikari, M.; Negi, B.; Kaushik, N.; Adhikari, A.; Al-Khedhairy, A.A.; Kaushik, N.K.; Choi, E.H. T-2 mycotoxin: toxicological effects and decontamination strategies. Oncotarget. 2017, 8, 33933–33952. [Google Scholar] [CrossRef]
- Stoev, S.D.; Diakov, L.; Koynarski, V.; Angelov, A. In Special pathology and diagnostics of mycoses, mycotoxicoses, parasitoses, intoxications and avitaminoses; Publishing House CD Contrast: Stara Zagora; Bulgaria, 2010, pp. 1–239.
- Wright, E.M. Renal Na+-Glucose Cotransporters. Am. J. Physiol. 2001, 280, F10–F18. [Google Scholar] [CrossRef]
- Sudakin, D.L. Trichothecenes in the Environment: Relevance to Human Health. Toxicol. Lett. 2003, 143, 97–107. [Google Scholar] [CrossRef]
- Ueno, Y. Toxicological Features of T-2 Toxin and Related Trichothecenes. Fundam. Appl. Toxicol. 1984, 4, S124–S132. [Google Scholar] [CrossRef]
- Wu, Q.-H.; Wang, X.; Yang, W.; Nüssler, A.K.; Xiong, L.-Y.; Kuča, K.; Dohnal, V.; Zhang, X.-J.; Yuan, Z.-H. Oxidative Stress-Mediated Cytotoxicity and Metabolism of T-2 Toxin and Deoxynivalenol in Animals and Humans: An Update. Arch. Toxicol. 2014, 88, 1309–1326. [Google Scholar] [CrossRef]
Figure 1.
Na (+) -dependent glucose co-transporter2 transmembrane transport in bird kidneys.
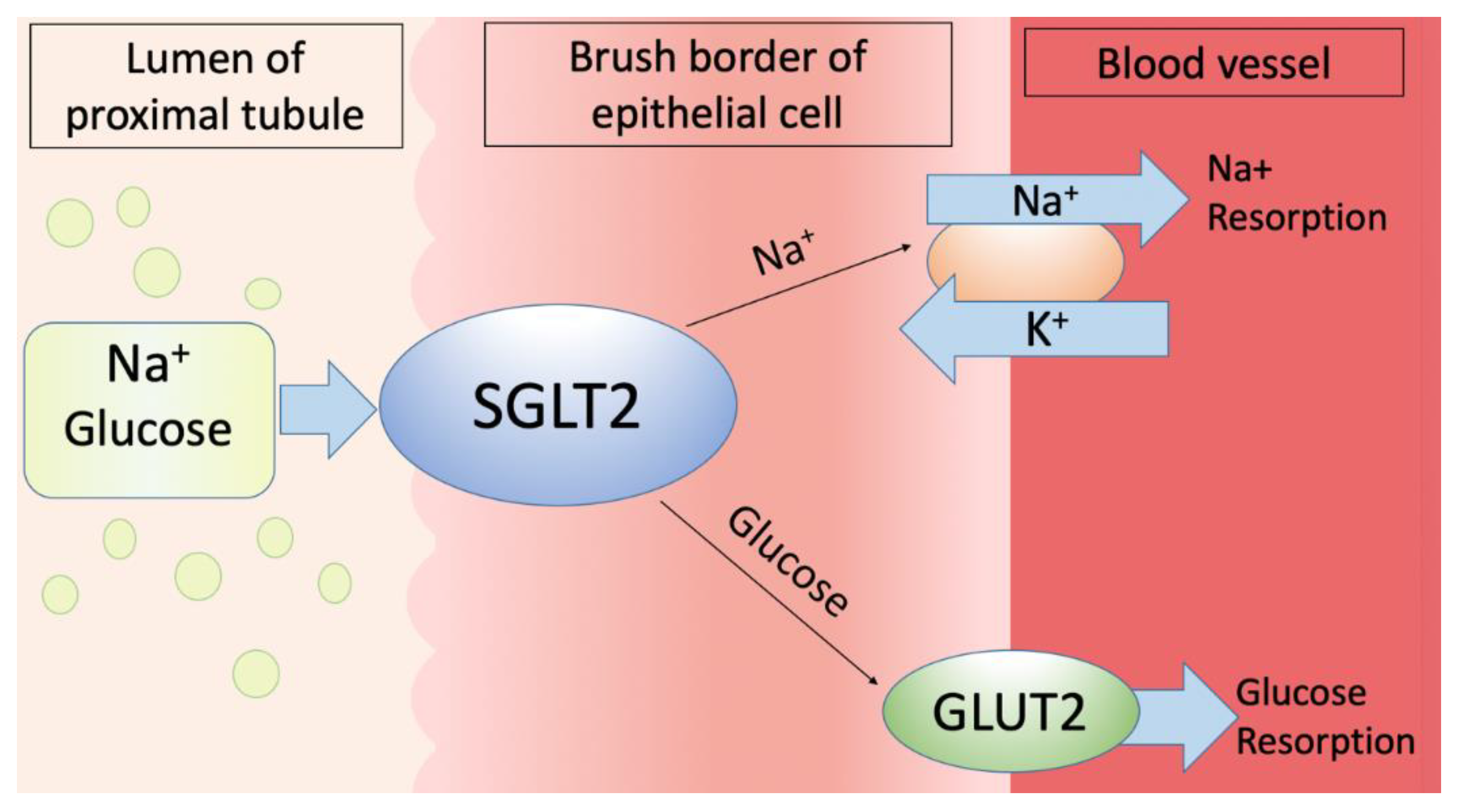
Figure 2.
Normal kidney morphology of a 7-day old chicken: proximal (arrowheads) and distal tubules (arrows) in the cortex of the kideney. Hematoxylin and eosin, 400x.
Figure 2.
Normal kidney morphology of a 7-day old chicken: proximal (arrowheads) and distal tubules (arrows) in the cortex of the kideney. Hematoxylin and eosin, 400x.
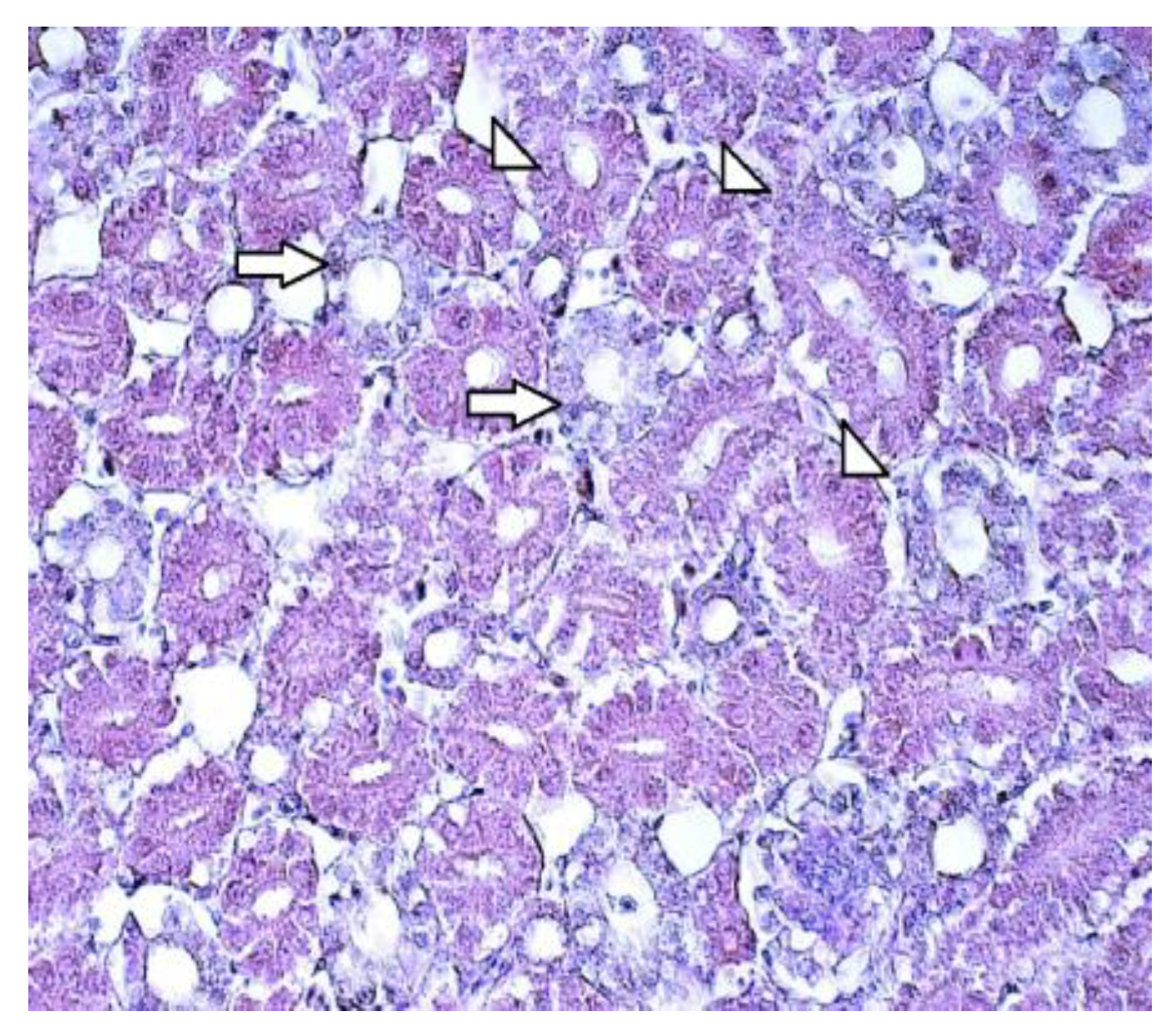
Figure 3.
Immunolocalization of the sodium-dependent glucose co-transporter-1 (SGLT1) in kidney tissue: (a) in healthy 7 days old chicken note the strong expression of SGLT1 in the apical part of the epithelial cells of renal proximal tubules (arrowheads), 400 x; (b) damaged brush border membranes of proximal tubule’s epithelial cells in intoxicated chicken (arrows) in T-2 mycotoxicated birds kidney tissue (arrows), 400 x.
Figure 3.
Immunolocalization of the sodium-dependent glucose co-transporter-1 (SGLT1) in kidney tissue: (a) in healthy 7 days old chicken note the strong expression of SGLT1 in the apical part of the epithelial cells of renal proximal tubules (arrowheads), 400 x; (b) damaged brush border membranes of proximal tubule’s epithelial cells in intoxicated chicken (arrows) in T-2 mycotoxicated birds kidney tissue (arrows), 400 x.
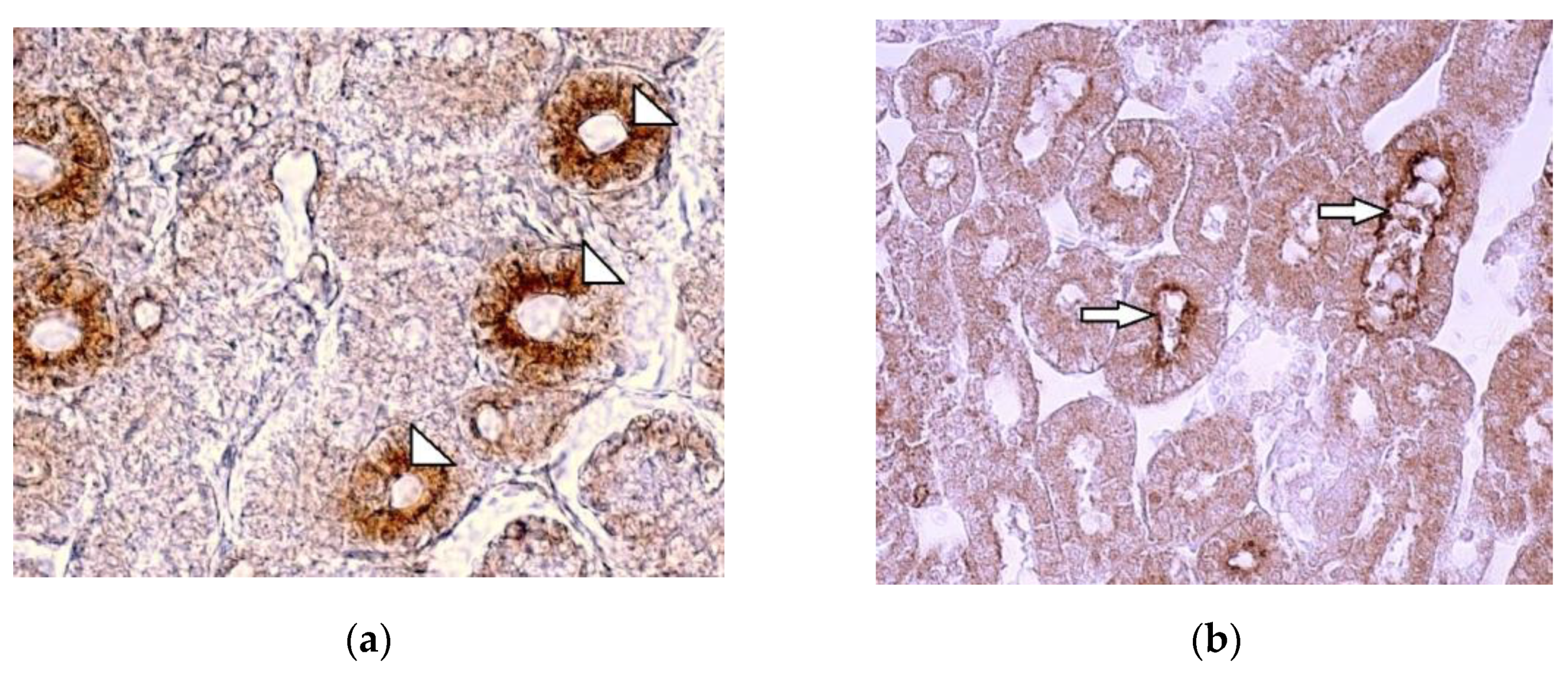
Figure 4.
Immunolocalization of sodium-dependent glucose co-transporter-2 (SGLT2) in: (a) healthy chicken kidney strongly stained prximal tubules (arrowheads) and weakly stained distal tubules (arrows) are noted 400 x; (b) intoxicated birds’ group is noted the pale staining of proximal renal tubules (arrows), 400 x.
Figure 4.
Immunolocalization of sodium-dependent glucose co-transporter-2 (SGLT2) in: (a) healthy chicken kidney strongly stained prximal tubules (arrowheads) and weakly stained distal tubules (arrows) are noted 400 x; (b) intoxicated birds’ group is noted the pale staining of proximal renal tubules (arrows), 400 x.
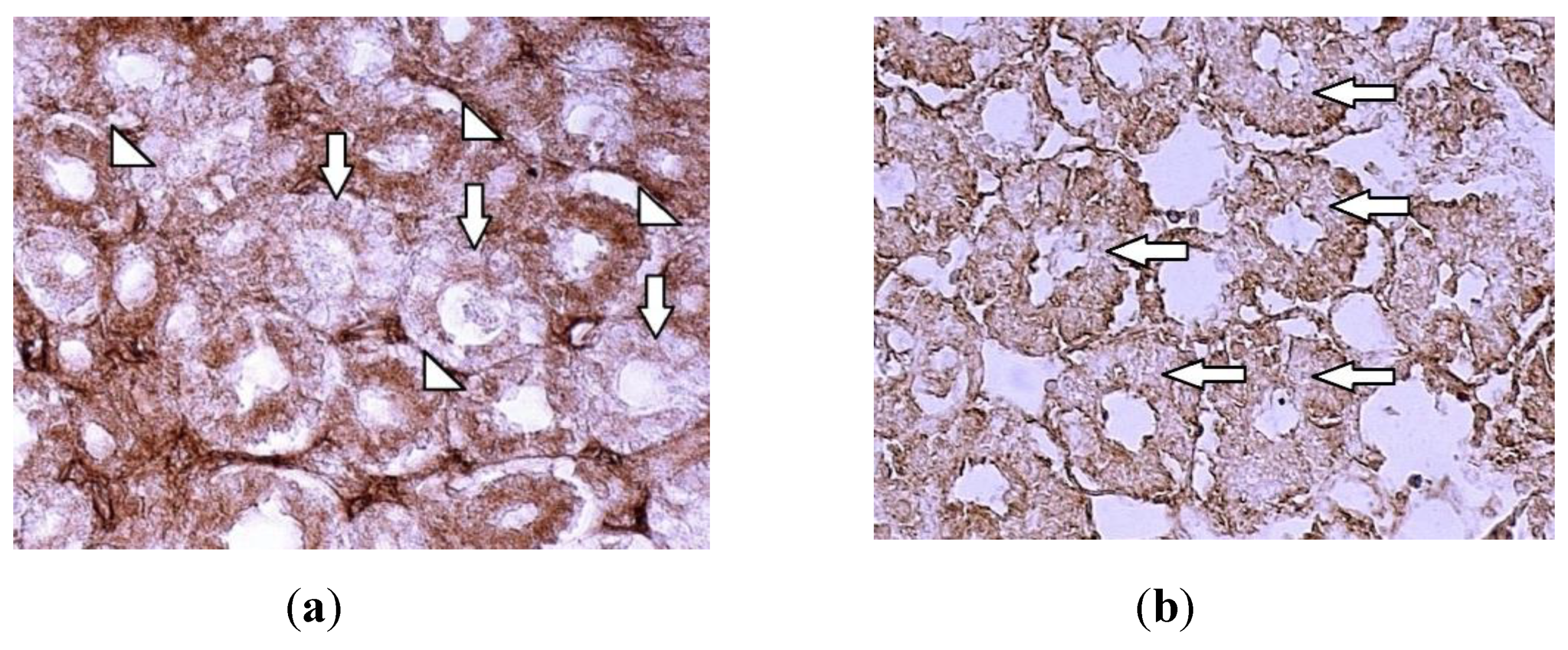
Figure 5.
SGLT2 in healthy 7 days old chicken kidneys. Note the unstained collecting ducts (arrows), Immunohistochemistry (IHC), 400 x.
Figure 5.
SGLT2 in healthy 7 days old chicken kidneys. Note the unstained collecting ducts (arrows), Immunohistochemistry (IHC), 400 x.
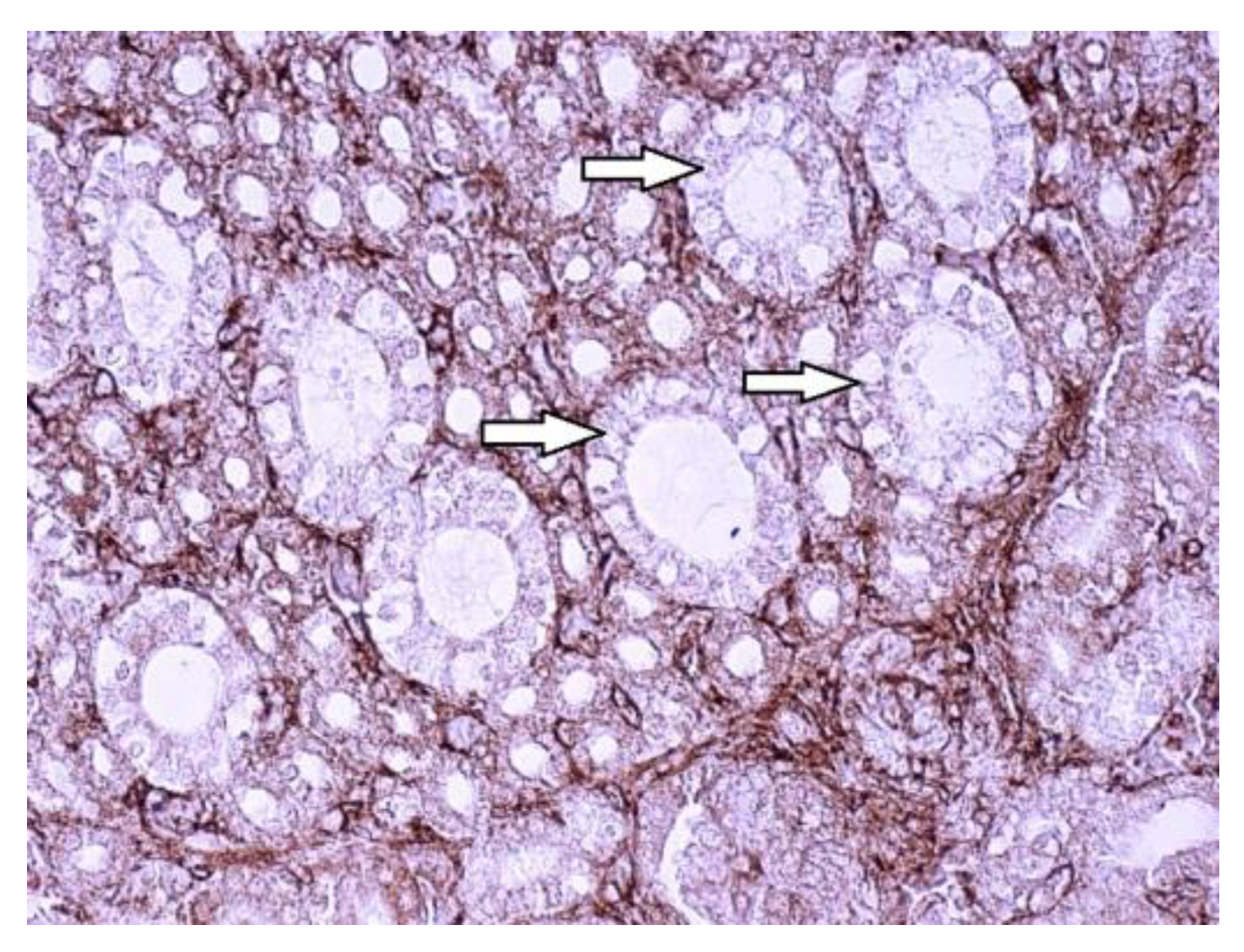

Table 1.
Expression of SGLT1 and SGLT2 in the proximal tubules of the chicken kidneys.
| Antibody | Chicken nr. | Control group | T-2 toxin group |
|---|---|---|---|
| SGLT1* | 1 2 3 4 5 6 7 |
++ +++ ++ ++ ++ ++ ++ |
++ + + + ++ + + |
| SGLT2* | 1 | +++ | + |
| 2 | +++ | ++ | |
| 3 | ++ | ++ | |
| 4 | +++ | + | |
| 5 | ++ | + | |
| 6 | +++ | + | |
| 7 | +++ | + |
* The intensity of staining is shown as: + weak; + + moderate; + + + strong.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



