
Submitted:
08 November 2024
Posted:
08 November 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Garlic (Allium sativum L.) is a species of the onion family, Alliaceae that has been widely used as a food and a folk medicine. The objective of this study was to determine the effects of AGE (Aged garlic extract) on proinflammatory genes relevant to COVID-19. To this aim, we treated bronchial epithelial IB3-1 cells with SARS-CoV-2 Spike protein (S-protein) or with the COVID-19 BNT162b2 vaccine in the absence or in the presence of AGE. The results obtained demonstrated that AGE is a potent inhibitor of the S-protein in-duced expression of the IL-1beta, IL-6 and IL-8 genes. Bio-plex analysis demonstrate that AGE reduced release of IL-6 and IL-8, that were highly induced by S-protein. No inhibition of cells growth, toxicity and proapoptotic effects were found in AGE-treated cells. The effects of one of the major AGE constituents (S-allyl cysteine, SAC) were studied on the same experimental model systems. SAC was able to inhibit the S-protein induced expression of IL-1beta, IL-6 and IL-8 genes and extracellular release of IL-6 and IL-8, con-firming that S-allyl cysteine is one of the constituents of AGE that are responsible for inhibiting S-protein induced pro-inflammatory genes. Docking experiments suggest that a possible mechanism of action of SAC is an interference with the activity of Toll-like receptors (TLRs), particularly TLR4, thereby inhibiting NF-B and NF-B regulated genes, such as IL-1beta, IL-6 and IL-8 genes. These results suggest that both AGE and SAC deserve further experimental efforts to verify their effects on pro-inflammatory genes in SARS-CoV-2 infected cells.
Keywords:
garlic
; natural products
; toll‐like receptor4
; NF‐B
; pro‐inflammatory genes
1. Introduction
Garlic (Allium sativum L.) is a species of the onion family (Alliaceae) that has been not only used as a food, but also as a medicinal plant in folk medicine [1,2,3,4,5]. A number of epidemiological studies have strongly suggested that garlic is effective in the prevention and treatment of several human diseases. In fact, this medicinal plant exhibited multiple pharmacological functions, such as anticarcinogenic, antithrombotic, hypolipidemic and hepatoprotective activity [6,7,8,9]. For instance, an interesting is that this herb exerts cytotoxic effects on multidrug-resistant human cancer cells by altering mitochondrial permeability [10] and by inducing apoptosis [11].
Among a large variety of commercial garlic supplements, AGE (Aged garlic extract) is well-known and has been studied in detail by different research groups [10]. AGE is a commercially available odorless preparation obtained by immersing fresh garlic in 15% aqueous ethanol solution over a prolonged period of time (up to 20 months) at room temperature [10,12,13,14,15]. This natural product has been shown to possess immunomodulatory and anticancer properties [10]. Several of the beneficial effects of garlic have been attributed to several bioactive compounds isolated from garlic, including the lipid-soluble allyl sulfur compounds (e.g., diallyl sulfide, diallyl disulfide and diallyl trisulfide) and water-soluble compounds, such as S-allyl cysteine (SAC), S-allylmercaptocysteine (SAMC) and S-1-propenylcysteine (S1PC) [16,17,18]. These bioactive compounds might be extracted from AGE by unique manufacturing process [11]. Most of the available data support the concept that AGE and AGE-related compounds retain anti-inflammatory properties. This is highly relevant for a possible treatment of the pandemic coronavirus disease 2019 (COVID-19), since inflammatory complications are a key factor, determining the severity of COVID-19 [19,20,21,22]. In fact, it is firmly established that COVID-19, caused by Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) is characterized by two major clinical phases: (a) a SARS-CoV-2 infection of target cells and tissues and (b) a deep inflammatory state, known as “cytokine storm” [21,22,23,24]. The interaction between the SARS-CoV-2 S-protein and ACE2 is a key step for inducing the “cytokine storm”. The interaction of SARS-CoV-2 S-protein with cellular receptors is followed by deep cellular changes including the hyperactivation of Nuclear Factor kappa-B (NF-κB) by IL-6/STATs axis [25]. This induces ARDS (Acute Respiratory Distress Syndrome) in lungs, that has been frequently observed in severe COVID-19 patients [26] and is clearly associated with the severity of the pathology [27,28], demonstrated by the fact that patients with COVID-19-related ARDS have high mortality rates compared to COVID-19 patients without any ARDS symptoms [28]. The impact of anti-inflammatory protocols for anti-SARS-CoV-2 pharmacological strategies is clear, as recently demonstrated by the effective treatments targeting IL-6 [29] and IL-8 [30]. Notably, the pharmacological approach for treating ARDS steadily needs novel anti-inflammatory reagents as different COVID-19 patients might respond in a different way to these treatments [31]. Several anti-inflammatory strategies to reduce COVID-19 “cytokine storm” and associated ARDS have been proposed using biomolecules derived from herbal medicinal extracts and reviewed by several authors [32,33]. This was judged to be a key strategy at the beginning of the pandemic, in consideration of the unknown nature of the disease and the lack of effective treatment protocols and approved vaccines [34,35]. Repurposing of known plant-derived reagents for anti-inflammatory activity against the COVID-19 “cytokine storm” might be of great interest [36,37]. The main objective of the present study was to verify whether AGE and SAC inhibit the expression of pro-inflammatory genes relevant in COVID-19. To this aim, human bronchial epithelial IB3-1 cells [38] have been exposed to SARS-CoV-2 Spike protein [38,39] and then cultured for 72 hours in the presence of AGE and SAC. Cells were then harvested and RNA isolated for RT-qPCR analysis. Supernatants were taken for Bioplex analysis.
2. Results
2.1. The Experimental Model System: Induction of Pro-Inflammatory Genes in Bronchial Epithelial IB3-1 Cells After Exposure to the SARS-CoV-2 Spike Protein and the COVID-19 BNT162b2 Vaccine
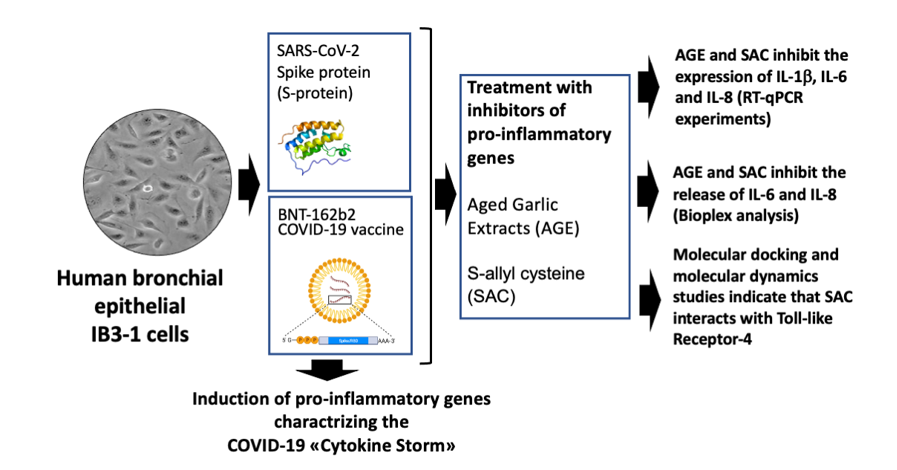
Two experimental model system have been considered in the present study. The first is constituted by human bronchial epithelial cells exposed to SARS-CoV-2 Spike protein and elsewhere described [38,39]. The exposure to S-protein leads to a NF-κB-dependent increase in the expression of pro-inflammatory genes, such as those coding Interleukin-6 (IL-6), IL-8, IL-1β. This experimental system (Figure 1A) has been already employed for studying the effects of sulforaphane (SFN) [38] and the agomiR-93-5p [39] on the expression of pro-inflammatory genes [38,39]. The second experimental model system (Figure 1B) is based on the treatment with the COVID-19 BNT162b2 vaccine [40,41]. In our present study, IB3-1 cells have been treated with 5, 15 and 50 nM S-protein, or 1μg/ml of BNT162b2. These treatments were performed in the absence or in the presence of increasing concentrations of AGE and SAC. After treatments, RNA was purified from the treated cells for RT-qPCR analysis, and cellular supernatants were isolated for the analysis of secreted cytokines, chemokines and growth factors in order to identify alteration(s) of SARS-CoV-2 Spike protein-induced secretome profile.
2.2. GC-MS Analysis of AGE and SAC
Figure 2 displays GC-MS analysis of AGE (Figure 2, A–C) and SAC (Figure 2, D–F). In both figures, Total Ion Chromatogram (TIC) and Selected Ion Monitoring (SIM) modes. These analyses reveal the presence of SAC within the aged garlic extracts powder, as evidenced by the overlaid SIM chromatograms (Figure 2C). SAC, as a di-TBDMS derivative, elutes at 10.49 minutes (Figure 2C, inset), and its identity is confirmed by the EI-mass spectrum (Figure 2D), wherein the loss of the t-butyl group yields a characteristic [M – t-Bu]+ ion (m/z 332). As expected, the presence of S-1-propenyl-l-cysteine (S1PC) was also noted in the AGE solution (Figure 2C).
2.3. The Treatment of IB3-1 Cells with AGE Reverses Upregulation of Pro-Inflammatory Genes Induced by SARS-CoV-2 Spike Protein
In the experiment shown in Figure 3, the effects of increasing amounts of AGE were studied on the S-protein induced expression of IL-1β, IL-6 and IL-8 genes. This experiment is the proof-of-principle experiment demonstrating that treatment with 0.1-0.5 mg/ml of AGE leads to a sharp inhibition of the expression of IL-1β (Figure 3A), IL-6 (Figure 3B) and IL-8 (Figure 3C) genes. This finding was highly reproducible and observed using different housekeeping internal control sequences, such as RPL13A (Figure 3), or β-actin and GAPDH (data not shown). We avoided the use of higher concentrations of AGE, since it is known that this extract is able to induce also pro-apoptotic effects at high concentrations, due to mitochondrial membrane depolarization process already observed in other cell types [10,11]. No major effects of AGE on IB3-1 cell growth were observed.
2.4. Effects of AGE on Toxicity and Apoptosis
The experiment shown in Figure 4 was performed in oder to verify whether AGE treatment of IB3-1 cells (Figure 4A) was to some extent cytotoxic. The inhibitory effect of the treatment with AGE on S-protein induced IL-6 mRNA accumulation was highly significant (p < 0.001). We employed the Annexin V assay, that differentiate cells in “live cells”, “early apoptotic cells”, “late apoptotic cells” and “dead cells” [42,43]. The results obtained (panels B-D of Figure 4) demonstrated that AGE, when maintained in contact with IB3-1 cells for 72 h, did not reduce the extent of viable cells and did not induce their apoptosis (Figure 4, B,D). In these experiments, IB3-1 cells were cultured for 72 h in the presence of 0.1, 0.5 and 1 mg/l of AGE.
These data sustain the concept that inhibitory effects of AGE of the expression of pro-inflammatory genes should not be ascribed to unspecific cytotoxic and/or pro apoptotic effects, at least at the concentrations used in our experiments. We expect however that higher concentrations of AGE might causes some pro-apoptotic effects, considering that AGE, and its constituents SAC and S1PC have been shown to exert potential anti-tumor effects caused by activation of the apoptotic pathway [11].
2.5. The Treatment of IB3-1 Cells with S-allyl-cysteine (SAC) Inhibits the Expression of Pro-Inflammatory Genes Induced by SARS-CoV-2 Spike Protein
During the process of aging of garlic, γ-glutamyl-S-allylcysteine is converted to S-allylcysteine (SAC) by a γ-glutamyltransferase, as reviewed by Colín-González et al., 2012) [44]. In the experiment shown in Figure 5, the effects of increasing amounts of SAC were studied on the S-protein induced expression of IL-1β, IL-6 and IL-8 genes. The results obtained indicate that treatment with 10-100 μM SAC leads to a sharp inhibition of the expression of IL-1β (Figure 5A), IL-6 (Figure 5B) and IL-8 (Figure 5C) genes.
The data presented in Figure 5 indicate that 10 μM SAC are sufficient to inhibit S-protein induced expression of the proinflammatory genes IL-1β, IL-6 and IL-3, suggesting that S-allyl-cysteine should be considered a compound responsible for the inhibitory effects of AGE on pro-inflammatory genes.
2.6. Effects of AGE and SAC on the Release of IL-6 and IL-8 by IB3-1 Cells Induced to Express Pro-Inflammatory Genes by Exposure to SARS-CoV-2 S-Protein
We have previously reported that S-protein induced IB3-1 release higher levels of cytokines, chemokines and growth factors when compared to control untreated cells. This can be studied by the Bio-plex technology, that allows the parallel analysis of 27 different released proteins, including cytokines, chemokines and growth factors [45]. We have also previously published that IL-1β is expressed at very low levels in IB3-1 cells [38,39]. For this reason, we limited the analysis of secreted proteins to IL-6 and IL-8. Figure 6 shows that treatment with both AGE (0.5 mg/l) and SAC (50 μM) leads to inhibition of the release of IL-6 (Figure 6A) and IL-8 (Figure 6B) by IB3-1. The Bioplex assay was performed on supernatants isolated from 72 hours treated IB3-1 cell cultures, either untreated or treated with the S-protein in the presence of AGE and SAC.
2.7. Effects of AGE and SAC on the Expression of Proinflammatory Genes Induced by the BNT162b2 COVID-19 Vaccine
In the experiments based on the use of the BNT162b2 vaccine, the effects of AGE and SAC were studied on the expression of the IL-8 gene. This was done because IL-8 gene expression was induced by the BNT162b2 vaccine at very high levels compared to the S-protein induction (data not shown). As expected, the intracellular content of SARS-CoV-2 Spike protein mRNA increases depending on the employed concentrations in BNT-162b2 treated IB3-1 cells. A significant increase (p < 0.01) was observed when 1 μg/mL vaccine was used (data not shown). As expected from previously published observations [40,41], production of S-protein was detectable when Western blotting was performed using cellular lysates from IB3-1 cells treated with the BNT162b2 vaccine (Papi et al., manuscript in preparation). As expected from the notion that in many cellular systems the S-protein induces the expression of pro-inflammatory genes through up-regulation of NF-κB [54,55,56], increase of expression of NF-κB was found in IB3-1 cells treated with the BNT-162b2 vaccine (Papi et al., manuscript in preparation). In agreement with the NF-κB upregulation, Figure 7 shows that treatment of IB3-1 cells with the BNT162b2 vaccine causes >30-fold increase in the content of IL-8 RNA. This increase was much lower (1.5-3 folds with respect to control untreated IB3-1 cells) when the SARS-CoV-2 S-protein was employed to induce NF-κB dependent increase in the expression of pro-inflammatory genes [38,39]. In agreement with the findings reported in Figure 2 and Figure 4, treatment of IB3-1 cells with AGE or SAC inhibits the expression of IL-8 gene induced by the BNT162b2 vaccine.
2.8. S-allyl-cysteine (SAC) Efficiently Interacts with Toll-Like Receptor 4 (TLR4): Molecular Docking and Molecular Dynamics Analyses
Preliminary analyses demonstrated a lack of binding of SAC to NF-κB. Binding of low-molecular weight drugs were reported by our group studying, trimethylangelicin and analogues [46], corilagin [47] and sulforaphane [48]. In these cases, efficient interations with NF-κB were found. On the contrary, no evidence of molecular interaction between SAC and NF-κB was found in our docking analyses (data not shown). Since Toll-like receptor are upstream regulators of the NF-κB signaling [49,50,51,52], we simulated the interaction between SAC and the TIR domain of TLR-4 using the well-known docking software AutoDock Vina (Figure 8 A,B). To further strength the reliability of the proposed interaction, the computed model was submitted to 50 ns of all-atom unbiased molecular dynamics simulation. During the simulation, the interactions between SAC and the chain A of TIR was well retained (average number of H-bonds between SAC and TIR-A: 2.97; Figure 8C), whereas a marked reduction of the interchain number of hydrogen bonds was noted (average number of H-bond between apo-TIR-A and apo-TIR-B: 7.75; average number of H-bond between SAC-bound-TIR-A and SAC-bound-TIR-B: 4.04; Figure 8D). We thus argued that the binding of SAC led to a destabilization of the TIR-TLR4 dimer, as can be seen by the comparison between Cα-RMSD values of apo-TIR and SAC-bound-TIR dimers (Figure 8E,F), and that this effect can be the leading cause of the anti-inflammatory effect of SAC.
3. Discussion
One of the clinical features of COVID-19 is the hyperinflammatory activity that is characterized by high expression of IL-6, IL-8 and several other cytokines, chemokines and growth factors [24]. This hyperinflammatory activity is associated with severe forms of COVID-19 and poor prognosis for patients [20]. For instance, Del Valle et al. found that high serum IL-6, IL-8 and TNF-α levels at the time of hospitalization are strong and independent predictors of patient survival [21]. In another study, Burke et al. found that inflammatory phenotyping (revealing upregulation of IL-6 and IL-8 gene expression) predicts clinical outcome in COVID-19 subjects [19]. Therefore, anti-inflammatory compounds and specific clinical protocols are highly needed.Concerning this issue, differential approaches targeting IL-6 and IL-8 cytokines has been proposed in several studies as well as in clinical trials [29,31,53,54,55,56].
The results presented in our study are a proof-of-principle that the expression of IL-1β, IL-6 and IL-8 genes, coding key proteins of the COVID-19 “cytokine storm” [22] can be inhibited by aged garlic extract (AGE) and the AGE bioactive compound S-allyl-cysteine (SAC). Since the control of the “cytokine storm” is a major issue in the management of COVID-19 patients [56], our study could stimulate research activity that can contribute to the development of protocols useful to control hyperinflammatory state associated with SARS-CoV-2 infection. This paper is expected to sustain research activity on plant extracts and food supplement containing S-allyl-cysteine and related bioactive molecules, in order to support the integration of 'phytopreparations' into conventional/official medicine focusing on COVID-19 treatment.
One of the limits of our study is the activity of AGE and SAC was studied on only three pro-inflammatory genes, IL-1β, IL-6 and IL-8. In this respect, it should be noted that the COVID-19 “Cytokine storm” is composed by a large number of pro-inflammatory proteins [19,20,21,22,24]. Therefore, our study is expected to stimulate research activity focusing on the effects of AGE and AGE-related compound on other “Cytokine storm” actors, such as IL-2, IL-7, IL-10, G-CSF, GM-CSF, IP10, MCP-1, MIP1A and TNF-α. In addition, only SAC, among the large number of constituents of AGE, was considered. In future experimental efforts, other AGE constituents should be studied, such as allicin, S-allylmercaptocysteine (SAMC) and S-1-propenylcysteine (S1PC) [16,17,18].
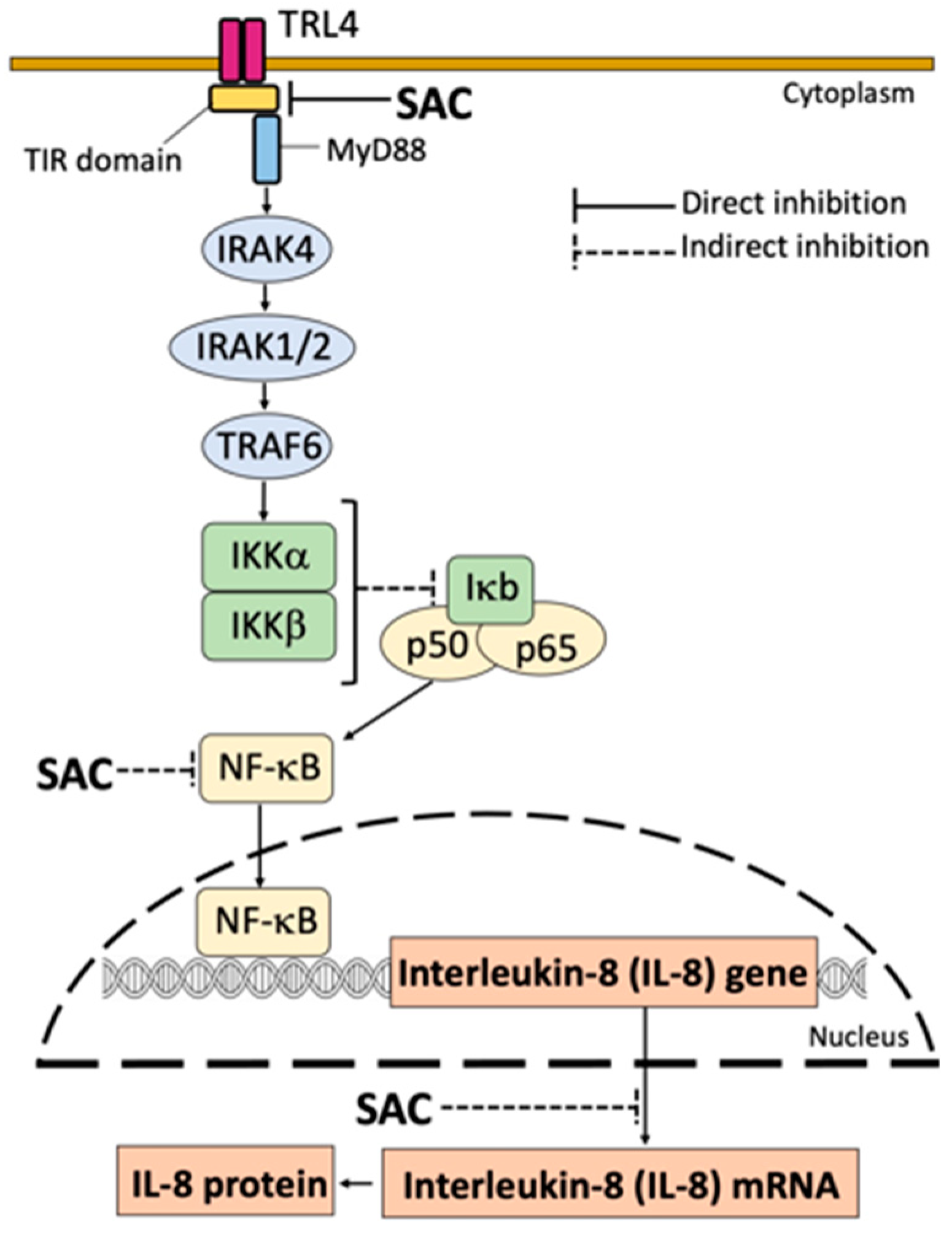
A final limit of our study is that the SAC mechanism of action was not fully analyzed and validated. This should be considered a major issue of the research on this topic in the future, since it could help in finding new targets for therapeutic protocols. Among several possibilities (that in any case are not mutually exclusive), SAC can exert its anti-inflammatory activity by NF-κB inhibition. In this context, we have hypothesized that SAC interacts with Toll-like receptors (mainly TLR4). This is shown in Figure 9, that suggest that SAC interacts with, and possibly inhibits TLR4 by destabilizing the dimer interactions. Since TLRs are involved in the NF-κB signal transduction [49,50,51,52], SAC might indirectly inhibit NF-κB through a direct inhibition of TLR activity.
In unstimulated cells (i.e. IB3-1 cells cultured In the absence of external stimulation), an inactive trimer is formed in the cytoplasm, due to the fact that the NF-κB protein p50/p65 binds the inhibitory protein IκB. In this condition, NF-κB is not translocated to the nucleus. By sharp contrast, when external stimuli act on the corresponding receptors (for example, when TLR4 is activated by SARS-CoV-2 through S-protein/TLT4 interactions) activation of IκK occurs leading to phosphorylation of IκB. Consequently, phospho-IκB dissociates from the trimer, and NF-κB is “activated”. In these conditions, the p50/p65 NF-κB proteins translocate to the nucleus and specifically interact with NF-κB binding sites present in the promoters of NF-κB regulated genes, such as the IL-8 gene (and other genes coding pro-inflammatory proteins)[57,58,59], thus causing transcriptional activation. In addition, upregulated inflammatory factors further activate NF-κB, thereby forming a positive feedback loop [68]. Our hypothesis is that SAC indirectly inhibits the NF-κB pathway through direct inhibition of TRL4. (Figure 9).
Full understanding the mechanism of action of SAC will be a very important step to identify novel molecular targets and to develop combined treatments with other anti-inflammatory agents.
4. Materials and Methods
4.1. Materials
All reagents and chemicals were analytical grade. SARS-Cov-2 Spike recombinant glycoprotein (ab49046) was purchased by Abcam (Cambridge, UK). The purity was > 90% as determined by SDS-PAGE. The BNT162b2 vaccine (COMIRNATYTM, Lot. FP8191) was obtained from the Hospital Pharmacy of University of Padova.
4.2. AGE Extraction and Chemical Characterization
AGE and SAC were provided by Wakunaga Pharmaceutical Co. Ltd. (Hiroshima, Japan) and manufactured as described by Kanamori et al. [11]. Briefly, Garlic cloves were sliced, immersed in a water-ethanol mixture solution and naturally extracted for >10 months at room temperature, as previously described [28]. The AGE powder used in our experiments was prepared by lyophilization. It contained approximately 28.6% (w/v, 286 mg/ml) solid material, 0.63% (6.3 mg/ml) arginine and 0.1% SAC (calculated on a dry weight basis) as a marker compound for standardization [29]. Both SAC and AGE powders were freshly dissolved in complete RPMI-1640 medium prior to each experiment. For chemical characterization, AGE and SAC were analyzed by GC-MS as TBDMS derivatives according Jiménez-Martín et al. [60]. S-allyl cysteine (SAC) was dissolved in 0.1 N HCl at a final concentration of 4 mg/mL. 5 µL of these solutions were spiked with 10 μL of internal standard (3,4-dimethoxybenzoic acid, 0.1 mg/mL) and dried under N2. Then, 30 μL of pure MTBSTFA, followed by 30 μL of pyridine, was added. The mixture was heated at 95 °C for 2 hours. The sample was then neutralized with sodium bicarbonate and subjected to GC-MS analysis. The same derivatization protocol was used for AGE power (4 mg/mL 0.1 M HCl). GC-MS analyses were performed with an Agilent 7890B gas chromatograph coupled to a 5977B quadrupole mass selective detector (Agilent Technologies, Palo Alto, CA, USA). Chromatographic separations were carried out with an Agilent HP5ms fused-silica capillary column (30 m × 0.25 mm i.d.) coated with 5%- phenyl-95%-dimethylpolysiloxane (film thickness 0.25 μm) as stationary phase. Injection mode: splitless at a temperature of 280 °C. Column temperature program: 70 °C (1 min) then to 300 °C at a rate of 20 °C/min and held for 10 min. The carrier gas was helium at a constant flow of 1.0 ml/min. The spectra were obtained in the electron impact mode at 70 eV ionization energy; ion source 280 °C; ion source vacuum 10-5 Torr.MS analysis was performed simultaneously in TIC (mass range scan from m/z 50 to 600 at a rate of 0.42 scans s-1) and SIM mode. GC-SIM-MS analysis was performed selecting the following ions: m/z 332 for SAC/S1-PC and m/z 239 for 3,4-dimethoxybenzoic acid (internal standard).
4.3. Cell Culture Conditions
The human bronchial epithelial IB3-1 cell line [38] was cultured in humidified atmosphere of 5% CO2/air in LHC-8 medium (Gibco, Thermo Fischer Scientific, Waltham, MA, USA) supplemented with 5% fetal bovine serum (FBS, Biowest, Nuaillé, France) in the absence of gentamycin. To verify the effect on proliferation, cell growth was monitored by determining the cell number/ml using a Z2 Coulter Counter (Coulter Electronics, Hialeah, FL, USA).
4.4. Stimulation of Cells with SARS-CoV-2 Spike Protein
SARS-CoV-2 Spike protein (139 KDa; stock concentration = 7.2 M in 9% urea, 0.32% Tris-HCl pH 7.2, 50% glycerol) was diluted in 200 µl of LHC-8 medium to achieve the final concentrations used to treat IB3-1 cells [39,40]. Briefly, cells seeded at 50% of confluence, were treated with Spike protein (5-50 nM) and incubated for 30 min at 4°C, then for 30 min at 37°C (this procedure is expected to maximize S-protein interaction with the receptor and the S-protein cellular uptake) [38,39]. After this incubation, LHC-8 medium supplemented with 5% (final concentration) FBS was added to a final 500 l volume and the cultures were further incubated at 37°C and for 24h.
4.5. Treatment of IB3-1 Cells with the BNT162b2 Vaccine
4.6. RNA Extraction
Cultured cells were trypsinized (0,05% trypsin and 0,02% EDTA; Sigma-Aldrich) and collected by centrifugation at 1,000 x g for 8 min at 4 °C, washed twice with DPBS 1X (Gibco, Thermo Fischer Scientific) and lysed with Tri-Reagent (Sigma Aldrich), according to manufacturer’s instructions. The isolated RNA was washed once with cold 75% ethanol, dried and dissolved in nuclease-free pure water before use. Obtained RNA was stored at -80°C until use [38,61].
4.7. Quantitative Analyses of mRNAs
For ILs mRNA analysis, 500 ng of total RNA were reverse transcribed to complementary DNA (cDNA) using the Taq-Man Reverse Transcription PCR Kit and random hexamers (Applied Biosystems, Thermo Fischer Scientific) in a final reaction volume of 50 µl. Real-time-qPCR experiments were carried out using an assay composed by a primer pair and a fluorescently labeled 5′ nuclease probe purchased from IDT (Integrated DNA Technologies, Coralville, IO, USA; Assays ID: Hs.PT.58.38869678.g for IL-8 and Hs.PT.58.40226675 for IL-6). An amount of 2 µl of cDNA were amplified, in presence of 2x PrimeTime Gene Expression Master Mix for 40 PCR cycles using CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Relative expression was calculated using the comparative cycle threshold method (ΔΔCT method) and the endogenous control human β-actin was used as normalizer. Negative controls (no template cDNA and RT-minus control) were also run in every experimental plate to assess specificity and to rule out contamination. RT-qPCR reactions were performed in duplicate for both target and normalizer genes [38,61].
4.8. Analysis of Cytokines, Chemokines and Growth Factors
Proteins released into culture supernatants were measured using Bio-Plex Human Cytokine 27-plex Assay (Bio-Rad) as suggested by the manufacturer. The assay allows the multiplexed quantitative measurement of 27 cytokines/chemokines (including FGF basic, Eotaxin, G-CSF, GM-CSF, IFN-γ, IL-1β, IL-1ra, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12 (p70), IL-13, IL-15, IL-17A, IP-10, MCP-1 (MCAF), MIP-1α, MIP-1β, PDGF-BB, RANTES, TNF-α, VEGF) in a single well. An amount of 50 μl of cytokine standards or samples (diluted supernatants recovered from IB3-1 cells) was incubated with 50 μl of anti-cytokine conjugated beads in 96-well filter plate for 30 min at room temperature with shaking. The plate was washed by vacuum filtration three times with 100 μl of Bio-Plex Wash Buffer, 25 μl of diluted detection antibody were added, to each well and plate was incubated for 30 min at room temperature with shaking. After three filter washes, 50 μl of streptavidin-phycoerythrin were added, and plate was incubated for 10 min at room temperature with shaking. Finally, plate was washed by vacuum filtration three times, beads were suspended in Bio-Plex Assay Buffer, plate was read by Bio-Rad 96-well plate reader. Collected data were analyzed by the Bio-Plex Manager Software (Bio-Rad) [45].
4.9. Effects on Cellular Viability and Apoptosis
Annexin V and Dead Cell assay was performed using Muse Cell Analyzer (Merck Millipore) instrument, according to the instructions supplied by the manufacturer. Cells were washed with sterile DPBS 1X, trypsinized, suspended and diluted (1:2) with Muse Annexin V & Dead Cell reagent (Merck Millipore). After an incubation of 15 minutes at room temperature in the dark, samples were acquired and data were analyzed using Annexin V and Dead Cell Software Module (Merck Millipore) [42,43].
4.10. Computational Studies
All the computational methodologies were carried out on a 32 Core AMD Ryzen 93,905×, 3.5 GHz Linux Workstation (O.S. Ubuntu 20.04) equipped with GPU (Nvidia Quadro RTX 4000, 8 GB). The structure of intracellular domains (TIR) of TLR4 dimer was derived from literature [62]. The structure of SAC was prepared with Avogadro software [63]. Blind docking simulation was performed on the entire TIR dimer surface using the AutoDock Vina software [64]. The top scored complex was submitted to all-atom unbiased molecular dynamics (MDs) simulation using the GROMACS software [65] patched with Plumed ver 2.6.5 [66] under the Charmm36 force field [67], as described in Zurlo et al. [40]. The complex was included in a rectangula box of 8 x 10 x 7 nanometers length, solvated and neutralized using 0.15M sodium chloride. The full system was submitted to energy minimization and equilibrated under NVT and NPT conditions. Long range electrostatic interactions were modelled using the Particle Mesh Ewald algorithm. LINCS, Nosé-Hoover and Parrinello-Rahman algorithms were used in the simulations for restraints, and as thermostat and barostat respectively. MDs were conducted under the NPT conditions for 50 ns with 2 fs time steps. Root-mean-squared deviation (RMSD) and number of hydrogen bonds were obtained through the “rms” and “hbond” routines implemented in Gromacs.
4.11. Statistics
All the data were normally distributed and presented, unless otherwise stated, as mean ± S.D. Statistical differences between groups were compared using one-way ANOVA (analyses of variance between groups) followed by Dunnett’s multiple comparison or paired t-test employing Prism (v. 9.02) by GraphPad software. Statistical differences were considered significant when p < 0.05 (*), and highly significant when p < 0.01 (**) and p < 0.001 (***).
5. Conclusions
We have developed simple experimental systems and analytical protocols for the screening of molecules interfering with the expression of proteins known to be involved in the COVID-19 “cytokine storm” (Figure 1). The results here presented demonstrate that exposure of epithelial IB3-1 cells to the SARS-CoV-2 spike protein or COVID-19 BNT162b2 vaccine induces increased expression of NF-κB and increased expression of pro-inflammatory genes, particularly of IL-6 and IL-8. Treatment with aged garlic extracts (AGE) and the AGE constituent S-allyl-cysteine (SAC) reverses IL-6 and IL-8 upregulation induced by SARS-CoV-2 Spike protein and BNT162b2 vaccine. in IB3-1 cells. Therefore, AGE and AGE constituents should be further evaluated as potential inhibitors of the COVID-19 “cytokine storm”. Further experiments should be programmed to identify other AGE derived agents able to inhibit changes in gene expression induced by SARS-CoV-2 Spike and COVID-19 BNT162b2 vaccine. Understanding the SAC mechanism of action will help in designing combined therapies with other anti-inflammatory agents.
Author Contributions
Conceptualization, Giovanni Marzaro, Enzo Agostinelli and Roberto Gambari; Data curation, Jessica Gasparello, Chiara Papi, Alberto Macone, Alessia Finotti, Enzo Agostinelli and Roberto Gambari; Formal analysis, Jessica Gasparello, Chiara Papi, Giovanni Marzaro, Alberto Macone, Matteo Zurlo, Alessia Finotti, Enzo Agostinelli and Roberto Gambari; Funding acquisition, Alessia Finotti and Roberto Gambari; Investigation, Jessica Gasparello and Chiara Papi; Methodology, Jessica Gasparello, Chiara Papi, Giovanni Marzaro, Alberto Macone, Matteo Zurlo, Alessia Finotti and Roberto Gambari; Project administration, Alessia Finotti; Resources, Giovanni Marzaro, Alberto Macone, Alessia Finotti, Enzo Agostinelli and Roberto Gambari; Software, Giovanni Marzaro; Supervision, Alessia Finotti, Enzo Agostinelli and Roberto Gambari; Validation, Jessica Gasparello, Chiara Papi, Matteo Zurlo and Roberto Gambari; Writing – original draft, Chiara Papi, Giovanni Marzaro, Alberto Macone, Enzo Agostinelli and Roberto Gambari; Writing – review & editing, Jessica Gasparello, Chiara Papi, Giovanni Marzaro, Alberto Macone, Matteo Zurlo, Alessia Finotti, Enzo Agostinelli and Roberto Gambari. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the MUR-FISR COVID-miRNAPNA Project (FISR2020IP_04128) (to R.G. and A.F.), by the Interuniversity Consortium for Biotechnologies, Italy (C.I.B) (CIB-Unife-2020 to R.G. and A.F.), and by the CARIPARO Foundation (MARZ_CARIVARI20_01 C94I20002500007, to G.M.). JG is funded by FIRC-AIRC “Michele e Carlo Ardizzone” fellowship (ID: 25528).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data will be made available upon request to the corresponding authors.
Acknowledgments
We thank Wakunaga Pharmaceutical Co., Ltd. (Japan) for providing the AGE and SAC samples used in the present study.
Conflicts of Interest
The authors declare that they have no competing interests. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Hahn, G. History, folk medicine, and legendary uses of garlic. In: Koch HP, Lawson LD, editors. Garlic: The Science and Therapeutic Application of Allium Sativum L and Related Species. Baltimore, Md, USA: Williams & Wilkins; 1996. pp. 1–24.
- Tudu, C.K.; Dutta, T.; Ghorai, M.; Biswas, P.; Samanta, D.; Oleksak, P.; Jha, N.K.; Kumar, M.; Radha Proćków, J.; Pérez de la Lastra, J.M.; Dey, A. Traditional uses, phytochemistry, pharmacology and toxicology of garlic (Allium sativum), a storehouse of diverse phytochemicals: A review of research from the last decade focusing on health and nutritional implications. Front. Nutr. 2022, 9, 949554. [Google Scholar] [CrossRef] [PubMed]
- Giang, T.V.; Hoa, L.N.M.; Hien, T.T.; Cuong, Q.D.; Cap, N.T.; Lam Vuong, N.; Thach, P.N. Traditional Vietnamese Medicine Containing Garlic Extract for Patients With Non-severe COVID-19: A Phase-II, Double-Blind, Randomized Controlled Trial. Cureus 2023, 15, e42484. [Google Scholar] [CrossRef] [PubMed]
- de Vahl E, Svanberg I. Traditional uses and practices of edible cultivated Allium species (fam. Amaryllidaceae) in Sweden. J. Ethnobiol Ethnomed. 2022, 18, 14. [Google Scholar]
- Melguizo-Rodríguez, L.; García-Recio, E.; Ruiz, C.; De Luna-Bertos, E.; Illescas-Montes, R.; Costela-Ruiz, V.J. Biological properties and therapeutic applications of garlic and its components. Food Funct. 2022, 13, 2415–2426. [Google Scholar] [CrossRef]
- Marni, R.; Kundrapu, D.B.; Chakraborti, A.; Malla, R. Insight into drug sensitizing effect of diallyl disulfide and diallyl trisulfide from Allium sativum L. on paclitaxel-resistant triple-negative breast cancer cells. J. Ethnopharmacol. 2022, 29, 115452. [Google Scholar] [CrossRef]
- Mollahosseini, M.; Hosseini-Marnani, E.; Panjeshahin, A.; Panbehkar-Jouybari, M.; Gheflati, A.; Mozaffari-Khosravi, H. A systematic review of randomized controlled trials related to the effects of garlic supplementation on platelet aggregation. Phytother. Res. 2022, 36, 4041–4050. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, X.; Sun, J.; Chen, Y.; Zhang, M.; Tang, X.; Wang, W.; Simal-Gandara, J.; Xu, H.; Li, N.; Liu, C. Evaluating the hypolipidemic effect of garlic essential oil encapsulated in a novel double-layer delivery system. Colloids Surf B Biointerfaces. 2024, 237, 113835. [Google Scholar] [CrossRef]
- Zhao, X.; Yin, F.; Fu, L.; Ma, Y.; Ye, L.; Huang, Y.; Fan, W.; Gao, W.; Cai, Y.; Mou, X. Garlic-derived exosome-like nanovesicles as a hepatoprotective agent alleviating acute liver failure by inhibiting CCR2/CCR5 signaling and inflammation. Biomater. Adv. 2023, 154, 213592. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, S.; Dalla Via, L.; Grancara, S.; Kanamori, Y.; García-Argáez, A.N.; Canettieri, G.; Arcari, P.; Toninello, A.; Agostinelli, E. The antioxidant, aged garlic extract, exerts cytotoxic effects on wild-type and multidrug-resistant human cancer cells by altering mitochondrial permeability. Int. J. Oncol. 2018, 53, 1257–1268. [Google Scholar] [CrossRef]
- Kanamori, Y.; Via, L.D.; Macone, A.; Canettieri, G.; Greco, A.; Toninello, A.; Agostinelli, E. Aged garlic extract and its constituent, S-allyl-L-cysteine, induce the apoptosis of neuroblastoma cancer cells due to mitochondrial membrane depolarization. Exp. Ther. Med. 2020, 19, 1511–1521. [Google Scholar] [CrossRef]
- Imai, J.; Ide, N.; Nagae, S.; Moriguchi, T.; Matsuura, H.; Itakura, Y. Antioxidant and radical scavenging effects of aged garlic extract and its constituents. Planta Medica 1994, 60, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.C.E.; Castro-Boqué, E.; García-Carrasco, A.; Morán-Valero, M.I.; González-Hedström, D.; Bermúdez-López, M.; Valdivielso, J.M.; Espinel, A.E.; Portero-Otín, M. Antihypertensive Effects of an Optimized Aged Garlic Extract in Subjects with Grade I Hypertension and Antihypertensive Drug Therapy: A Randomized, Triple-Blind Controlled Trial. Nutrients 2023, 15, 3691. [Google Scholar] [CrossRef]
- Yang, Q.; Li, F.; Jia, G.; Liu, R. Aged black garlic extract inhibits the growth of estrogen receptor-positive breast cancer cells by downregulating MCL-1 expression through the ROS-JNK pathway. PLoS One 2023, 18, e0286454. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; MK; Mitra, A. ; Zaky, M.Y.; Pathak, S.; Banerjee, A. A Review on the Efficacy of Plant-derived Bio-active Compounds Curcumin and Aged Garlic Extract in Modulating Cancer and Age-related Diseases. Curr. Rev. Clin. Exp. Pharmacol. 2024, 19, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, N.; He, Z.; Chen, C.; Ma, J.; Liu, X.; Deng, S.; Xie, L. Diallyl trisulfide inhibits osteosarcoma 143B cell migration, invasion and EMT by inducing autophagy. Heliyon 2024, 10, e26681. [Google Scholar] [CrossRef]
- Ferguson, D.T.; Taka, E.; Messeha, S.; Flores-Rozas, H.; Reed, S.L.; Redmond, B.V.; Soliman, K.F.A.; Kanga, K.J.W.; Darling-Reed, S.F. The Garlic Compound, Diallyl Trisulfide, Attenuates Benzo[a]Pyrene-Induced Precancerous Effect through Its Antioxidant Effect, AhR Inhibition, and Increased DNA Repair in Human Breast Epithelial Cells. Nutrients 2024, 16, 300. [Google Scholar] [CrossRef]
- Bentke-Imiolek A, Szlęzak D, Zarzycka M, Wróbel M, Bronowicka-Adamska P. S-Allyl-L-Cysteine Affects Cell Proliferation and Expression of H2S-Synthetizing Enzymes in MCF-7 and MDA-MB-231 Adenocarcinoma Cell Lines. Biomolecules 2024, 14, 188.
- Burke, H.; Freeman, A.; Cellura, D.C.; Stuart, B.L.; Brendish, N.J.; Poole, S.; Borca, F.; Phan, H.T.T.; Sheard, N.; Williams, S.; Spalluto, C.M.; Staples, K.J.; Clark, T.W.; Wilkinson, T.M.A. Inflammatory phenotyping predicts clinical outcome in COVID-19. Respir. Res. 2020, 21, 245. [Google Scholar] [CrossRef]
- Zeng, Z. , Yu, H., Chen, H., Qi, W., Chen, L., Chen, G., Yan, W., Chen, T., Ning, Q., Han, M., Wu, D., Longitudinal changes of inflammatory parameters and their correlation with disease severity and outcomes in patients with COVID-19 from Wuhan, China. Crit. Care. 2020, 24, 525. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; Marron, T.U.; Xie, H.; Patel, M.; Tuballes, K.; Van Oekelen, O.; Rahman, A.; Kovatch, P.; Aberg, J.A.; Schadt, E.; Jagannath, S.; Mazumdar, M.; Charney, A.W.; Firpo-Betancourt, A.; Mendu, D.R.; Jhang, J.; Reich, D.; Sigel, K.; Cordon-Cardo, C.; Feldmann, M.; Parekh, S.; Merad, M.; Gnjatic, S. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Soy, M. , Keser, G., Atagündüz, P., Tabak, F., Atagündüz, I., Kayhan, S., 2020. Cytokine in COVID-19: pathogenesis and overview of anti-inflammatory agents used in treatment. Clin. Rheumatol. 2020, 39, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Pascarella, G. , Strumia, A., Piliego, C., Bruno, F., Del Buono, R., Costa, F., Scarlata, S., Agrò, F.E. COVID-19 diagnosis and management: a comprehensive review. J. Intern. Med. 2020, 288, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Pelaia, C.; Tinello, C.; Vatrella, A.; De Sarro, G.; Pelaia, G. Lung under attack by COVID-19-induced cytokine storm: pathogenic mechanisms and therapeutic implications. Ther. Adv. Respir. Dis. 2020, 14, 1753466620933508. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z. , Bujko, K., Ciechanowicz, A., Sielatycka, K., Cymer, M., Marlicz, W., Kucia, M. SARS-CoV-2 entry receptor ACE2 is expressed on very small CD45(-) precursors of hematopoietic and endothelial cells and in response to virus Spike protein activates the Nlrp3 inflammasome. Stem Cell Rev. Rep. 2021, 17, 266–277. [Google Scholar]
- Soumagne, T.; Winiszewski, H.; Besch, G.; Mahr, N.; Senot, T.; Costa, P.; Grillet, F.; Behr, J.; Mouhat, B.; Mourey, G.; Fournel, A.; Meneveau, N.; Samain, E.; Capellier, G.; Piton, G.; Pili-Floury, S. Pulmonary embolism among critically ill patients with ARDS due to COVID-19. Respir. Med. Res. 2020, 78, 100789. [Google Scholar] [CrossRef]
- Grasselli, G.; Tonetti, T.; Protti, A.; Langer, T.; Girardis, M.; Bellani, G.; Laffey, J.; Carrafiello, G.; Carsana, L.; Rizzuto, C.; Zanella, A.; Scaravilli, V.; Pizzilli, G.; Grieco, D.L.; Di Meglio, L.; de Pascale, G.; Lanza, E.; Monteduro, F.; Zompatori, M.; Filippini, C.; Locatelli, F.; Cecconi, M.; Fumagalli, R.; Nava, S.; Vincent, J.L.; Antonelli, M.; Slutsky, A.S.; Pesenti, A.; Ranieri, V.M. Pathophysiology of COVID-19-associated acute respiratory distress syndrome: a multicentre prospective observational study. Lancet Respir. Med. 2020, 8, 1201–1208. [Google Scholar] [CrossRef]
- Matthay, M.A.; Leligdowicz, A.; Liu, K.D. Biological mechanisms of COVID-19 Acute Respiratory Distress Syndrome. Am. J. Respir. Crit. Care Med. 2020, 202, 1489–1491. [Google Scholar] [CrossRef] [PubMed]
- Nasonov, E.; Samsonov, M. The role of Interleukin 6 inhibitors in therapy of severe COVID-19. Biomed. Pharmacother. 2020, 131, 110698. [Google Scholar] [CrossRef]
- Andreakos, E.; Papadaki, M.; Serhan, C.N. Dexamethasone, pro-resolving lipid mediators and resolution of inflammation in COVID-19. Allergy 2021, 76, 626–628. [Google Scholar] [CrossRef]
- de Simone, G. , Mancusi, C., 2020. Finding the right time for anti-inflammatory therapy in COVID-19. Int. J. Infect. Dis. 2020, 101, 247–248. [Google Scholar]
- Khalifa, S.A.M.; Yosri, N.; El-Mallah, M.F.; Ghonaim, R.; Guo, Z.; Musharraf, S.G.; Du, M.; Khatib, A.; Xiao, J.; Saeed, A.; El-Seedi, H.H.R.; Zhao, C.; Efferth, T.; El-Seedi, H.R. Screening for natural and derived bio-active compounds in preclinical and clinical studies: One of the frontlines of fighting the coronaviruses pandemic. Phytomedicine 2021, 85, 153311. [Google Scholar] [CrossRef]
- Matveeva, T.; Khafizova, G.; Sokornova, S. In search of herbal anti-SARS-Cov2 compounds. Front. Plant. Sci. 2020, 11, 589998. [Google Scholar] [CrossRef] [PubMed]
- Adem, Ş.; Eyupoglu, V.; Sarfraz, I.; Rasul, A.; Zahoor, A.F.; Ali, M.; Abdalla, M.; Ibrahim, I.M.; Elfiky, A.A. Caffeic acid derivatives (CAFDs) as inhibitors of SARS-CoV-2: CAFDs-based functional foods as a potential alternative approach to combat COVID-19. Phytomedicine 2021, 85, 153310. [Google Scholar] [CrossRef]
- Ngwa, W.; Kumar, R.; Thompson, D.; Lyerly, W.; Moore, R.; Reid, T.E.; Lowe, H.; Toyang, N. Potential of flavonoid-inspired phytomedicines against COVID-19. Molecules 2020, 25, 2707. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K. , Chiririwa, H., Dandara, C., Dzobo, W., Coronavirus Disease-2019 treatment strategies targeting interleukin-6 signaling and herbal medicine. OMICS 2020, 25, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Y.; Vilekar, P.; Yang, S.P.; Gupta, M.; Oh, M.I.; Meek, A.; Doyle, L.; Villar, L.; Brennecke, A.; Liyanage, I.; Reed, M.; Barden, C.; Weaver, D.F. Small molecule therapeutics for COVID-19: repurposing of inhaled furosemide. Peer J. 2020, 8, e9533. [Google Scholar] [CrossRef]
- Gasparello, J.; D'Aversa, E.; Papi, C.; Gambari, L.; Grigolo, B.; Borgatti, M.; Finotti, A.; Gambari, R. Sulforaphane inhibits the expression of interleukin-6 and interleukin-8 induced in bronchial epithelial IB3-1 cells by exposure to the SARS-CoV-2 Spike protein. Phytomedicine 2021, 87, 153583. [Google Scholar] [CrossRef]
- Gasparello, J.; d'Aversa, E.; Breveglieri, G.; Borgatti, M.; Finotti, A.; Gambari, R. In vitro induction of interleukin-8 by SARS-CoV-2 Spike protein is inhibited in bronchial epithelial IB3-1 cells by a miR-93-5p agomiR. Int. Immunopharmacol. 2021, 101, 108201. [Google Scholar] [CrossRef]
- Zurlo, M.; Gasparello, J.; Verona, M.; Papi, C.; Cosenza, L.C.; Finotti, A.; Marzaro, G.; Gambari, R. The anti-SARS-CoV-2 BNT162b2 vaccine suppresses mithramycin-induced erythroid differentiation and expression of embryo-fetal globin genes in human erythroleukemia K562 cells. Exp. Cell. Res. 2023, 433, 113853. [Google Scholar] [CrossRef]
- Cosenza, L.C.; Marzaro, G.; Zurlo, M.; Gasparello, J.; Zuccato, C.; Finotti, A.; Gambari, R. Inhibitory effects of SARS-CoV-2 spike protein and BNT162b2 vaccine on erythropoietin-induced globin gene expression in erythroid precursor cells from patients with β-thalassemia. Exp. Hematol. 2024, 129, 104128. [Google Scholar] [CrossRef]
- Gasparello, J.; Gambari, L.; Papi, C.; Rozzi, A.; Manicardi, A.; Corradini, R.; Gambari, R.; Finotti, A. High Levels of Apoptosis Are Induced in the Human Colon Cancer HT-29 Cell Line by Co-Administration of Sulforaphane and a Peptide Nucleic Acid Targeting miR-15b-5p. Nucleic Acid Ther. 2020, 30, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Gasparello, J.; Papi, C.; Zurlo, M.; Gambari, L.; Rozzi, A.; Manicardi, A.; Corradini, R.; Gambari, R.; Finotti, A. Treatment of Human Glioblastoma U251 Cells with Sulforaphane and a Peptide Nucleic Acid (PNA) Targeting miR-15b-5p: Synergistic Effects on Induction of Apoptosis. Molecules 2022, 27, 1299. [Google Scholar] [CrossRef] [PubMed]
- Colín-González, A.L.; Santana, R.A.; Silva-Islas, C.A.; Chánez-Cárdenas, M.E.; Santamaría, A.; Maldonado, P.D. The antioxidant mechanisms underlying the aged garlic extract- and S-allylcysteine-induced protection. Oxid Med Cell Longev. 2012, 2012, 907162. [Google Scholar] [CrossRef] [PubMed]
- Penolazzi, L.; Lambertini, E.; Tavanti, E.; Torreggiani, E.; Vesce, F.; Gambari, R.; Piva, R. Evaluation of chemokine and cytokine profiles in osteoblast progenitors from umbilical cord blood stem cells by BIO-PLEX technology. Cell Biol. Int. 2008, 32, 320–25. [Google Scholar] [CrossRef] [PubMed]
- Marzaro, G.; Lampronti, I.; D'Aversa, E.; Sacchetti, G.; Miolo, G.; Vaccarin, C.; Cabrini, G.; Dechecchi, M.C.; Gambari, R.; Chilin, A. Design, synthesis and biological evaluation of novel trimethylangelicin analogues targeting nuclear factor kB (NF-kB). Eur J. Med. Chem. 2018, 151, 285–293. [Google Scholar] [CrossRef]
- Milani, R.; Brognara, E.; Fabbri, E.; Finotti, A.; Borgatti, M.; Lampronti, I.; Marzaro, G.; Chilin, A.; Lee, K.K.; Kok, S.H.; Chui, C.H.; Gambari, R. Corilagin Induces High Levels of Apoptosis in the Temozolomide-Resistant T98G Glioma Cell Line. Oncol. Res. 2018, 26, 1307–1315. [Google Scholar] [CrossRef]
- Gasparello, J.; Marzaro, G.; Papi, C.; Gentili, V.; Rizzo, R.; Zurlo, M.; Scapoli, C.; Finotti, A.; Gambari, R. Effects of Sulforaphane on SARS-CoV-2 infection and NF-κB dependent expression of genes involved in the COVID-19 'cytokine storm'. Int. J. Mol. Med. 2023, 52, 76. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- McKiel, L.A.; Fitzpatrick, L.E. Toll-like Receptor 2-Dependent NF-κB/AP-1 Activation by Damage-Associated Molecular Patterns Adsorbed on Polymeric Surfaces. ACS Biomater. Sci. Eng. 2018, 4, 3792–3801. [Google Scholar] [CrossRef]
- Kumar, V. Toll-like receptors in sepsis-associated cytokine storm and their endogenous negative regulators as future immunomodulatory targets. Int Immunopharmacol. 2020, 89, 107087. [Google Scholar] [CrossRef]
- Jiang H, Wang HY, Wang JW, Lou DY, Niu N, Li GH, Qu P. NF-κB inhibitor on Toll-like receptor 4 signal-induced expression of angiotensinogen and AT1a receptor in neonatal rat left ventricular myocytes. Exp. Ther. Med. 2018, 16, 3875–3882. [Google Scholar]
- Xiao, F.; Wu, L.; Zhu, X.; Zhang, L.; Liu, D.; Wu, L.; Zou, H.; Lu, L. Interleukin-6 blocking therapy for COVID-19: From immune pathogenesis to clinical outcomes. Rheumatol Immunol Res. 2022, 3, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Gavioli, E.; Landoni, G.; Piemonti, L.; Grossi, P.; Toya, S.; Minnella, E.; Allegretti, M.; Mantelli, F. A Phase 2 Multicenter, Randomized, Controlled Study to Evaluate the Efficacy and Safety of Reparixin in Hospitalized Adult Patients with COVID-19 Pneumonia. Am J Resp. Crit. Care Med. 2022, 205. [Google Scholar]
- Gritti, G.; Raimondi, F.; Bottazzi, B.; Ripamonti, D.; Riva, I.; Landi, F.; Alborghetti, L.; Frigeni, M.; Damiani, M.; Micò, C.; Fagiuoli, S.; Lorini, F.L.; Gandini, L.; Novelli, L.; Morgan, J.P.; Owens, B.M.J.; Kanhai, K.J.K.; Reljanovic, G.T.; Rizzi, M.; Di Marco, F.; Mantovani, A.; Rambaldi, A. Siltuximab downregulates interleukin-8 and pentraxin 3 to improve ventilatory status and survival in severe COVID-19. Leukemia 2021, 35, 2710–2714. [Google Scholar] [CrossRef]
- Mustafa, M.I. , Abdelmoneim, A.H., Mahmoud, E.M., Makhawi, A.M., 2020. Cytokine Storm in COVID-19 patients, its impact on organs and potential treatment by QTY Code-Designed Detergent-Free Chemokine Receptors. Mediators Inflamm. 2020; 8198963. [Google Scholar]
- Brasier, A. The nuclear factor-kappaB-interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–8. [Google Scholar] [CrossRef] [PubMed]
- Libermann, T.A.; Baltiore, D. Activation of Interleukin-6 Gene Expression through the NF-KB Transcription Factor. Mol. & Cell Biol. 1990; 2327–2334. [Google Scholar]
- Bezzerri, V.; Borgatti, M.; Finotti, A.; Tamanini, A.; Gambari, R.; Cabrini, G. Mapping the transcriptional machinery of the IL-8 gene in human bronchial epithelial cells. J. Immunol. 2011, 187, 6069–81. [Google Scholar] [CrossRef]
- Jiménez-Martín, E.; Ruiz, J.; Pérez-Palacios, T.; Silva, A.; Antequera, T. Gas chromatography-mass spectrometry method for the determination of free amino acids as their dimethyl-tert-butylsilyl (TBDMS) derivatives in animal source food. J. Agric. Food Chem. 2012, 60, 2456–63. [Google Scholar] [CrossRef]
- Gasparello, J.; Lomazzi, M.; Papi, C.; D'Aversa, E.; Sansone, F.; Casnati, A.; Donofrio, G.; Gambari, R.; Finotti, A. Efficient Delivery of MicroRNA and AntimiRNA Molecules Using an Argininocalix [4]arene Macrocycle. Mol. Ther. Nucleic Acids. 2019, 18, 748–763. [Google Scholar] [CrossRef]
- Patra, M.C.; Kwon, H.K.; Batool, M.; Choi, S. Computational Insight Into the Structural Organization of Full-Length Toll-Like Receptor 4 Dimer in a Model Phospholipid Bilayer. Front. Immunol. 2018, 9, 489. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: an advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf.Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Abraham MJ, Murtola T, Schulz R, Páll S, Smith JC, Hess B, Lindahl E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar]
- Bonomi M, Branduardi D, Bussi G, Camilloni C, Provasi D, Raiteri P, Donadio D, Marinelli F, Pietrucci F, Broglia R, Parrinello M. PLUMED: A portable plugin for free-energy calculations with molecular dynamics. Computer Physics Comm. 2009, 180, 1961–1972. [Google Scholar] [CrossRef]
- Huang J, MacKerell AD. CHARMM36 all-atom additive protein force field: Validation based on comparison to NMR data. J. Comput. Chem. 2013, 34, 2135–2145. [Google Scholar] [CrossRef]
Figure 1.
Pictorial representation of the experimental model systems of the present study.
Figure 2.
GC-MS analysis of AGE and SAC. A, total ion current (TIC) chromatogram of the AGE sample derivatized with TBDMS; B, total ion current (TIC) chromatogram of the SAC sample derivatized with TBDMS; C, overlaid selected ion chromatograms (SIM) of AGE and SAC samples derivatized with TBDMS acquiring the following ions: m/z 239 (3,4-dimethoxybenzoic acid, internal standard) and m/z 332 (SAC); D, electron impact mass-spectrum of S-allylcysteine as di-TBDMS derivative (retention time 10.49 min). Both AGE and the SAC powders were freshly dissolved in complete RPMI-1640 medium prior to each experiment.
Figure 2.
GC-MS analysis of AGE and SAC. A, total ion current (TIC) chromatogram of the AGE sample derivatized with TBDMS; B, total ion current (TIC) chromatogram of the SAC sample derivatized with TBDMS; C, overlaid selected ion chromatograms (SIM) of AGE and SAC samples derivatized with TBDMS acquiring the following ions: m/z 239 (3,4-dimethoxybenzoic acid, internal standard) and m/z 332 (SAC); D, electron impact mass-spectrum of S-allylcysteine as di-TBDMS derivative (retention time 10.49 min). Both AGE and the SAC powders were freshly dissolved in complete RPMI-1640 medium prior to each experiment.
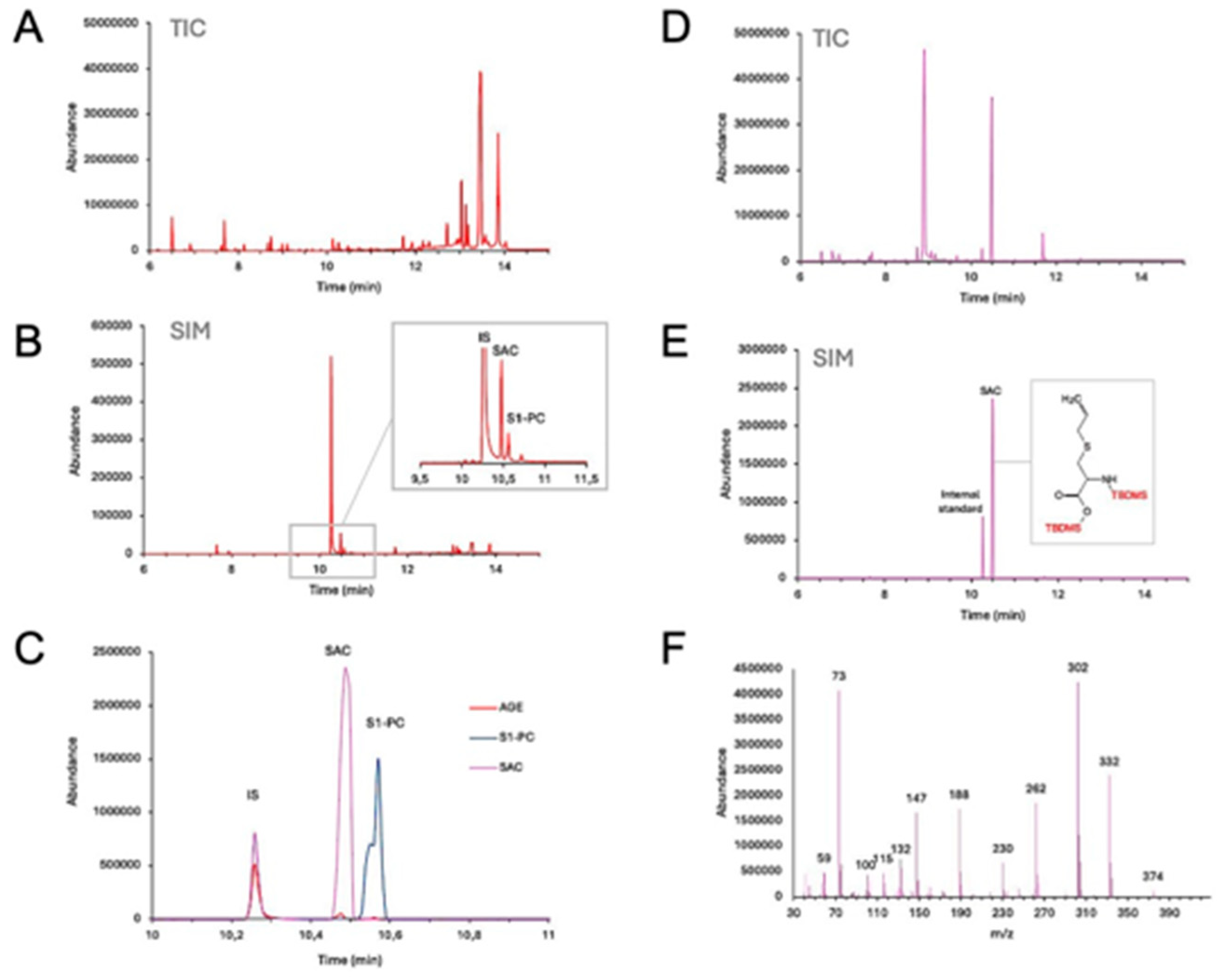
Figure 3.
Representative examples of the effects of increasing amounts of AGE on S-protein induced expression of IL-1β, IL-6 (B) and IL-8 (C) genes. The mRNA content was analyzed by RT-qPCR using IB3-1 cells exposed to 20 ng/l of SARS-CoV-2 S-protein and treated for 72 hours as described.
Figure 3.
Representative examples of the effects of increasing amounts of AGE on S-protein induced expression of IL-1β, IL-6 (B) and IL-8 (C) genes. The mRNA content was analyzed by RT-qPCR using IB3-1 cells exposed to 20 ng/l of SARS-CoV-2 S-protein and treated for 72 hours as described.
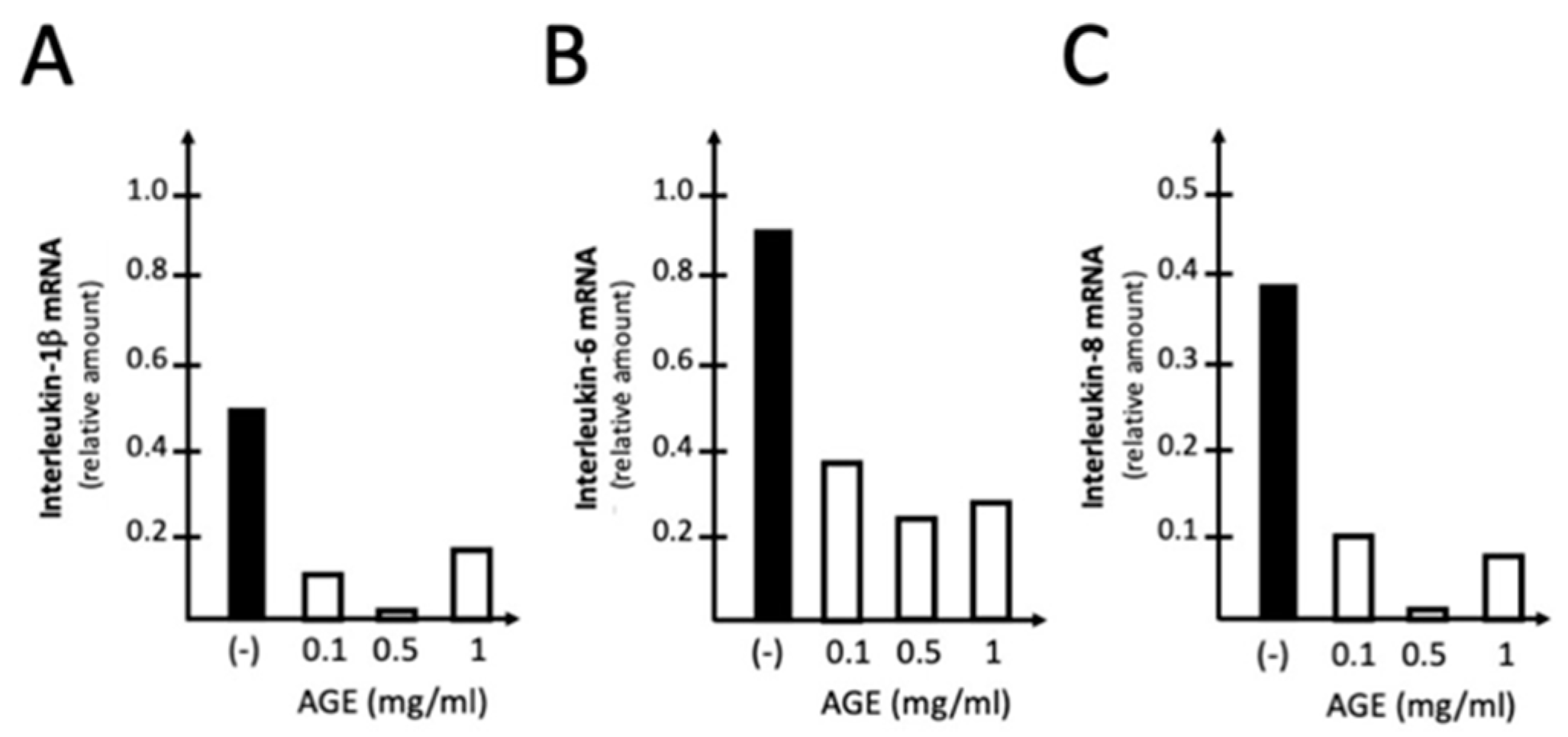
Figure 4.
Effects of AGE on cell vitality and apoptosis. IB3-1 cells were cultured in the absence or the presence of the indicated concentrations of AGE for 72h. Vitality and apoptosis were determined using the Muse Annexin V & Dead Cell kit. A. Effects of AGE on the IL-6 mRNA content in treated IB3-1 cells. B-E. Representative Annex V assay performed on control untreated IB3-1 cells (B) or IB3-1 cells treated for three days with AGE, used at 0.1 g/l (C), 0.5 g/l (D) and 1 μg/ml (F). Summary of the Annexin V assay data shown in panels B-E.
Figure 4.
Effects of AGE on cell vitality and apoptosis. IB3-1 cells were cultured in the absence or the presence of the indicated concentrations of AGE for 72h. Vitality and apoptosis were determined using the Muse Annexin V & Dead Cell kit. A. Effects of AGE on the IL-6 mRNA content in treated IB3-1 cells. B-E. Representative Annex V assay performed on control untreated IB3-1 cells (B) or IB3-1 cells treated for three days with AGE, used at 0.1 g/l (C), 0.5 g/l (D) and 1 μg/ml (F). Summary of the Annexin V assay data shown in panels B-E.
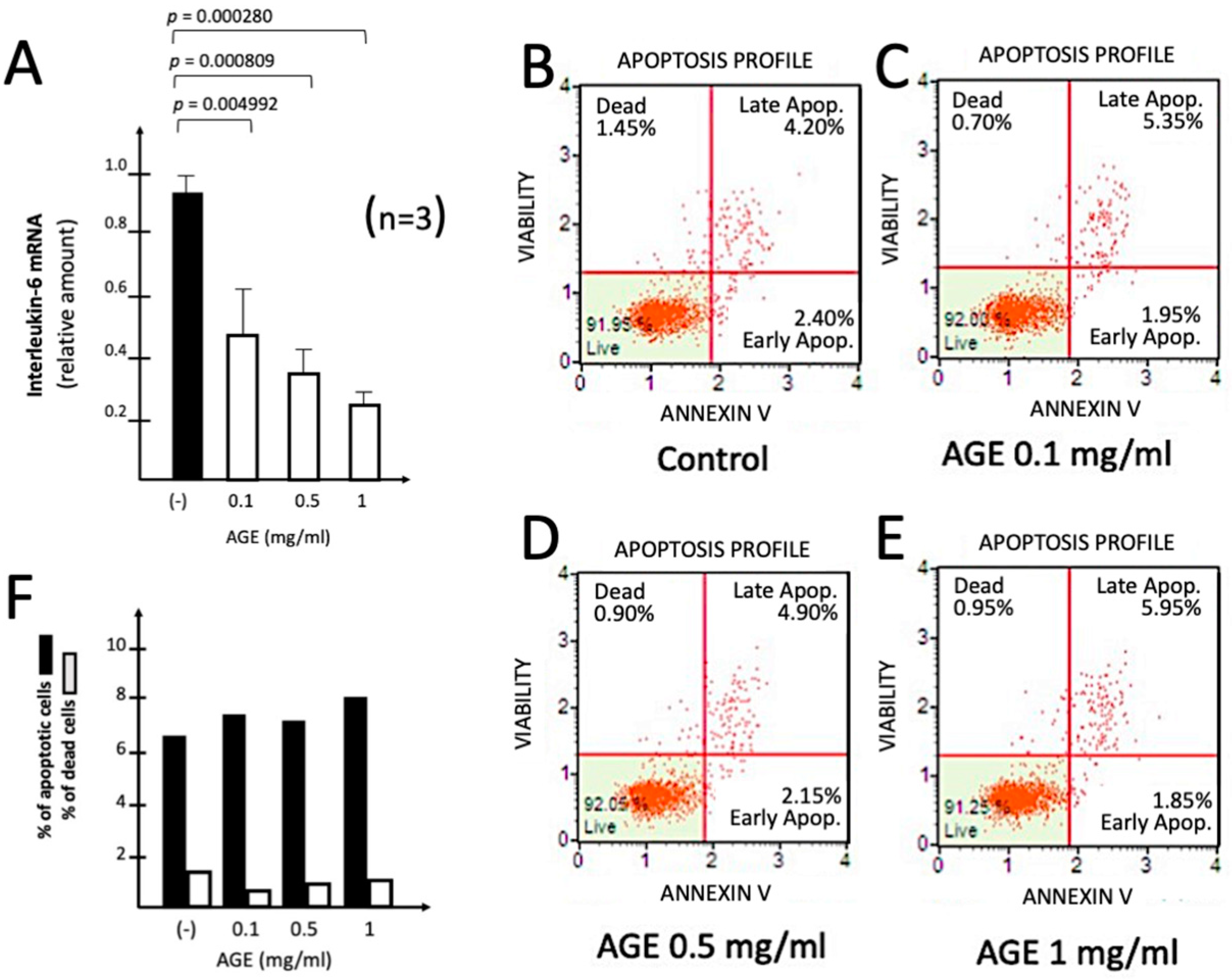
Figure 5.
Representative examples of the effect of increasing concentrations of SAC on S-protein induced expression of IL-1β (A), IL-6 (B) and IL-8 (C) genes. The mRNA content was analyzed by RT-qPCR using IB3-1 cells exposed to 20 ng/l of SARS-CoV-2 S-protein and treated for 72 hours as described.
Figure 5.
Representative examples of the effect of increasing concentrations of SAC on S-protein induced expression of IL-1β (A), IL-6 (B) and IL-8 (C) genes. The mRNA content was analyzed by RT-qPCR using IB3-1 cells exposed to 20 ng/l of SARS-CoV-2 S-protein and treated for 72 hours as described.
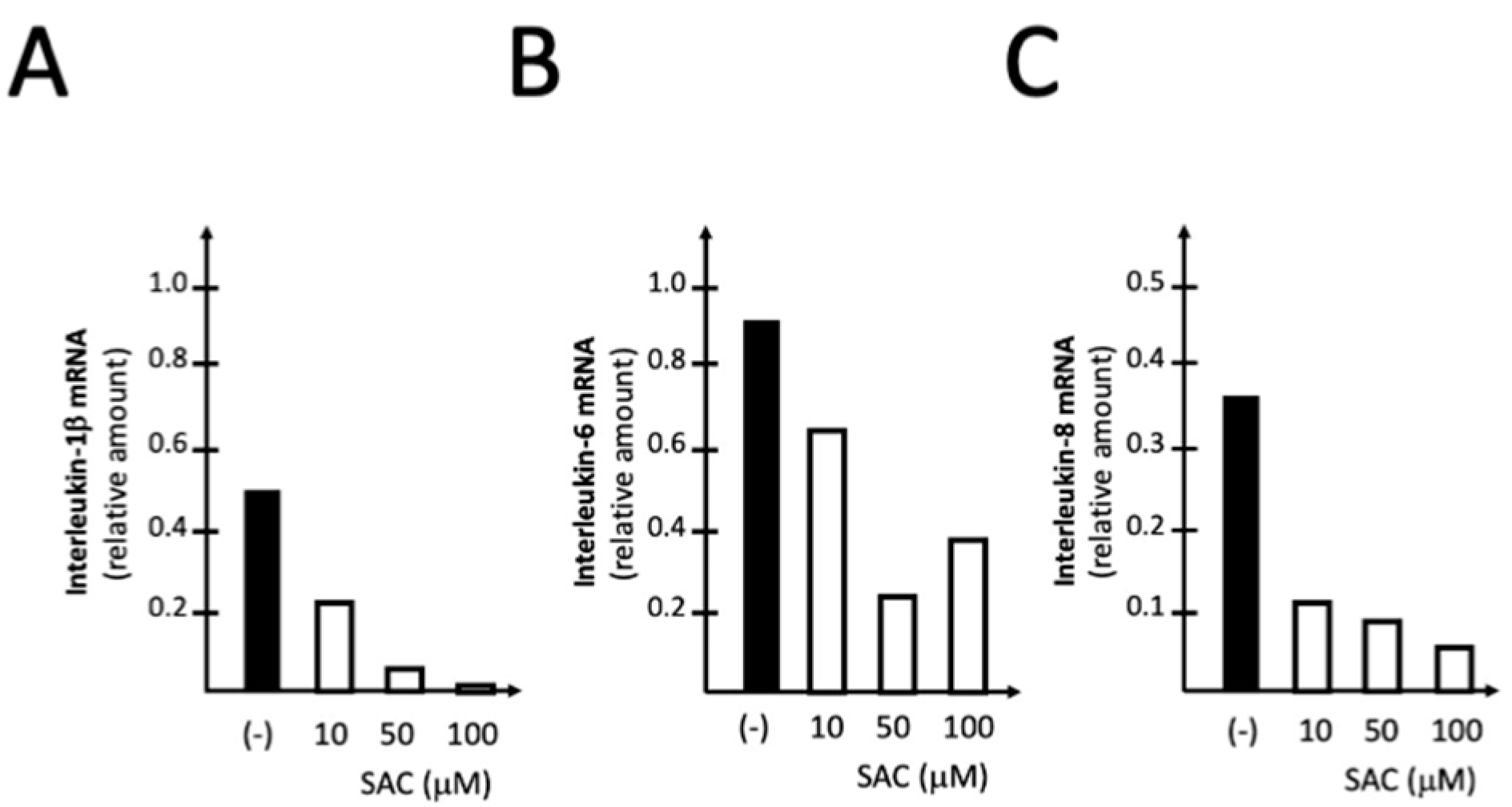
Figure 6.
Effects of AGE and SAC on release of IL-6 and IL-8 by S-protein stimulated IB3-1 cells. A panel of 27 cytokines/chemokines/growth factors was analyzed by the Bio-plex technology in IB3-1 infected cells 24h after the exposure to the S-protein. Results represent the average ± SD (n= 3).
Figure 6.
Effects of AGE and SAC on release of IL-6 and IL-8 by S-protein stimulated IB3-1 cells. A panel of 27 cytokines/chemokines/growth factors was analyzed by the Bio-plex technology in IB3-1 infected cells 24h after the exposure to the S-protein. Results represent the average ± SD (n= 3).
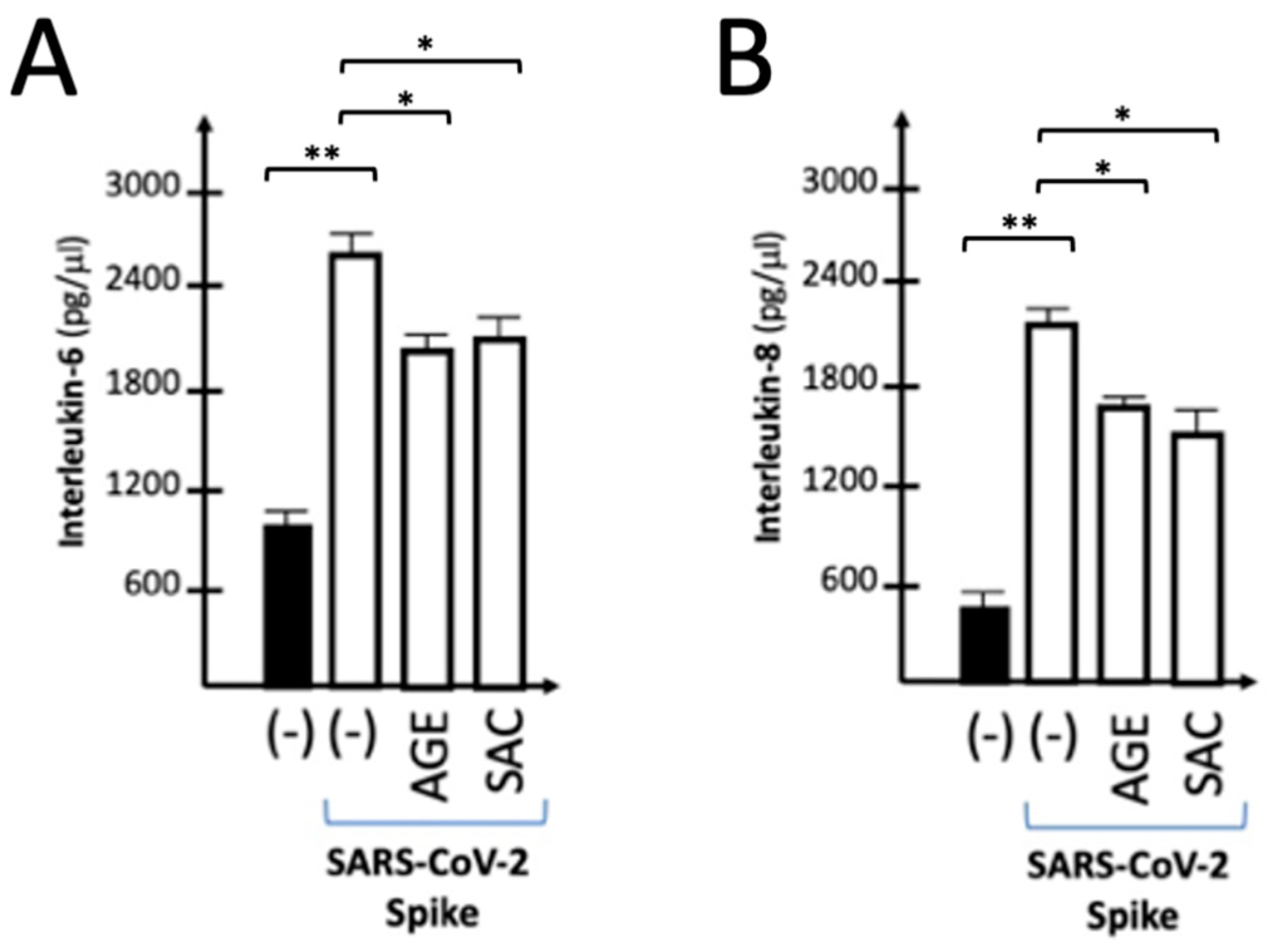
Figure 7.
Effects of AGE and SAC on IL-8 gene expression induced in IB3-1 cells by treatment with BNT162b2 COVID-19 vaccine.
Figure 7.
Effects of AGE and SAC on IL-8 gene expression induced in IB3-1 cells by treatment with BNT162b2 COVID-19 vaccine.
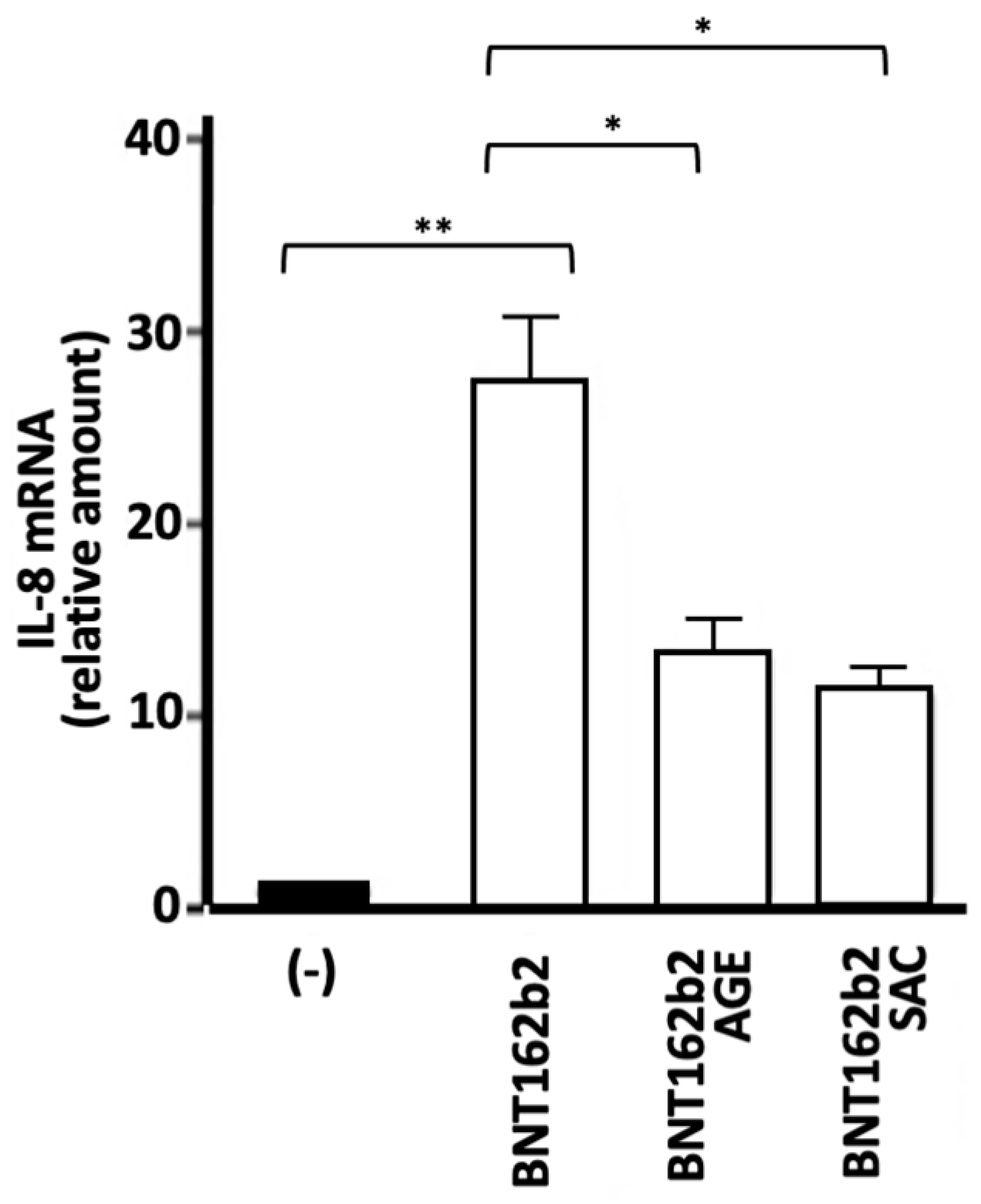
Figure 8.
simulation of SAC binding to TLR-4. Binding mode predicted for SAC (A) with the TLR4-TIR dimer and details of interactions (B). TLR4 monomers are depicted in blue (chain A) and green (chain B) respectively; SAC is depicted as stick (magenta colored carbon). Hydrogen bonds are depicted as dashed black lines. Number of H-bonds formed between chains A and B during 50ns of MDs. Black line represents the simulations of apo-TLR-4; red line represents the simulation of SAC-bound-TLR-4. Note a marked reduction in interchain interactions as a consequence of the interaction with SAC. Number of H-bonds formed between SAC and TLR-4 chain A (black) and chain B (red). Root mean squared deviation (RMSD, nm) of Cα-atoms in SAC-bound-TLR-4. Black line represents the total RMSD; blue line represents the chain A RMSD; red line represents the chain B RMSD. Root mean squared deviation (RMSD, nm) of SAC heavy atoms with respect to Cα-atoms. Black line represents the SAC RMSD relative to the dimer; blue line represents the SAC RMSD relative to chain A; red line represents the SAC RMSD relative to chain B. Note that the binding between SAC and chain A remains constant during the 50ns of MDs.
Figure 8.
simulation of SAC binding to TLR-4. Binding mode predicted for SAC (A) with the TLR4-TIR dimer and details of interactions (B). TLR4 monomers are depicted in blue (chain A) and green (chain B) respectively; SAC is depicted as stick (magenta colored carbon). Hydrogen bonds are depicted as dashed black lines. Number of H-bonds formed between chains A and B during 50ns of MDs. Black line represents the simulations of apo-TLR-4; red line represents the simulation of SAC-bound-TLR-4. Note a marked reduction in interchain interactions as a consequence of the interaction with SAC. Number of H-bonds formed between SAC and TLR-4 chain A (black) and chain B (red). Root mean squared deviation (RMSD, nm) of Cα-atoms in SAC-bound-TLR-4. Black line represents the total RMSD; blue line represents the chain A RMSD; red line represents the chain B RMSD. Root mean squared deviation (RMSD, nm) of SAC heavy atoms with respect to Cα-atoms. Black line represents the SAC RMSD relative to the dimer; blue line represents the SAC RMSD relative to chain A; red line represents the SAC RMSD relative to chain B. Note that the binding between SAC and chain A remains constant during the 50ns of MDs.
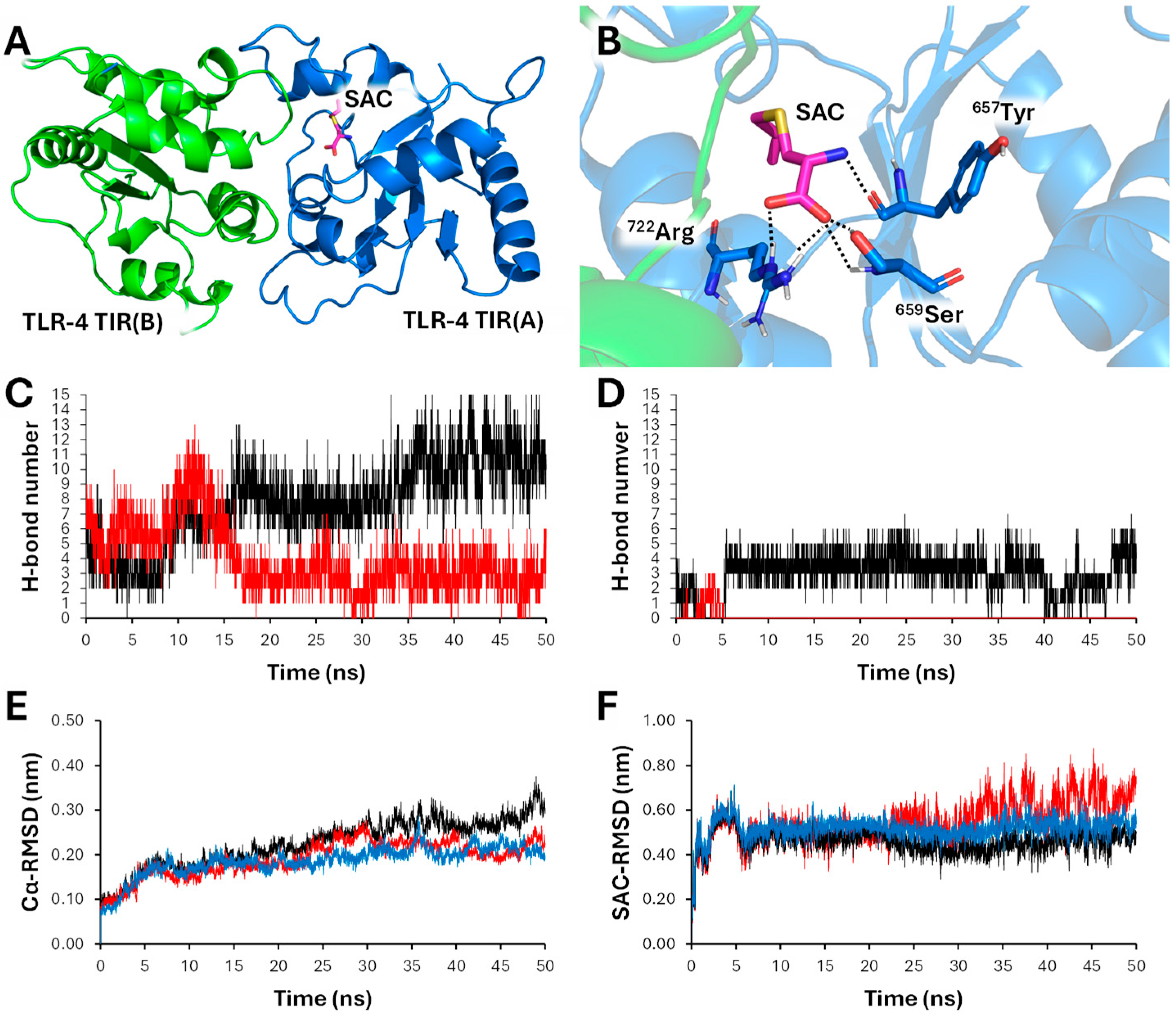
Figure 9.
Proposed mechanism of action of SAC. Indirect inhibition of NF-κB might be caused by a direct inhibition of TLR4 (and/or other TLRs, including intracellular TLRs). NFκB inhibition is causative of the inhibitory effects of NF-κB regulated genes, such as IL-1κ, IL-6, IL-8 and others.
Figure 9.
Proposed mechanism of action of SAC. Indirect inhibition of NF-κB might be caused by a direct inhibition of TLR4 (and/or other TLRs, including intracellular TLRs). NFκB inhibition is causative of the inhibitory effects of NF-κB regulated genes, such as IL-1κ, IL-6, IL-8 and others.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



