
Submitted:
05 December 2024
Posted:
06 December 2024
You are already at the latest version
Abstract
Reproductive characteristics are important tools for sustainable forestry and to transmit of present gene diversity to future generations by forestry practices. Knowledge and estimation of fertility variation and its linkage parameters such as population size, gene diversity in seed crop calculated by reproductive traits is used widely because of its many advantages. Estimation of these parameters are used for many purposes in forestry practices (i.e., natural regenerations, establishment and management of seed sources). In this study, cone and seed production and their effect on fertility variation were examined in two natural populations sampled at southern part of Türkiye of Taurus cedar (Cedrus libani A. Rich.). Numbers of mature cone which of two-years, and filled seed were counted from fifty trees selected phenotypic of each population in 2023. The averages of cone and seed number were 90 and 33, and 5321 and 3115 per tree, respectively. Among individual trees within population, and between populations showed large differences for the cone and seed productions. Percentage of filled seed were 94% and 83% in the populations. There were significant differences (p<0.05) between populations for the productions and percentage of filled seeds according to results of analysis of variance. Estimated fertility variations (Ψ<2) were well accordance with target (Ψ<3). Effective number of parents was ranged from 30.1 (60% of number of individuals) to 41.4 (83%). Besides, data sets can be used to fill the FLR-Library.

Keywords:
gene diversity
; population size
; reproductive
; sibling coefficient
; stand
; FLR-Library
Abstract. Reproductive characteristics are important tools for sustainable forestry and to transmit of present gene diversity to future generations by forestry practices. Knowledge and estimation of fertility variation and its linkage parameters such as population size, gene diversity in seed crop calculated by reproductive traits is used widely because of its many advantages. Estimation of these parameters are used for many purposes in forestry practices (i.e., natural regenerations, establishment and management of seed sources). In this study, cone and seed production and their effect on fertility variation were examined in two natural populations sampled at southern part of Türkiye of Taurus cedar (Cedrus libani A. Rich.). Numbers of mature cone which of two-years, and filled seed were counted from fifty trees selected phenotypic of each population in 2023. The averages of cone and seed number were 90 and 33, and 5321 and 3115 per tree, respectively. Among individual trees within population, and between populations showed large differences for the cone and seed productions. Percentage of filled seed were 94% and 83% in the populations. There were significant differences (p<0.05) between populations for the productions and percentage of filled seeds according to results of analysis of variance. Estimated fertility variations (Ψ<2) were well accordance with target (Ψ<3). Effective number of parents was ranged from 30.1 (60% of number of individuals) to 41.4 (83%). Besides, data sets can be used to fill the FLR-Library.
Introduction
Taurus cedar (Cedrus libani A. Rich.) occupy widely on the Taurus Mountains in southern Türkiye together with some local populations in the Black Sea region of Anatolia, and remain populations in Lebanon and Syria in the world (Boydak 2003). The species occurs mainly between 800 and 2100 m asl. Taurus cedar has also natural distribution as small population, group or individuals both at 500-600 m and 2400 m as lower and higher altitude (Boydak 2003; Odabaşı 1990). Taurus cedar which is a remarkable evergreen conifer grows as individual tree up to 1000 years or more than that as monumental tree, 3 m stem diameter with 50 m height (Boydak 2003). Monoecious Taurus cedar has 3-5 cm male and 1-1.5 cm female strobili with mature cone which of size 8-10 cm length and 4-6 cm diameter (Boydak 2003; Odabaşı 1990) (Figure 1).
Pollination is carried out in September or early of October according to its elevation. Between April and June of following year, cone lets develop to mature cone in about 25-26 months after flowering (Evcimen 1963; Bilir, Kang 2021). Cedrus libani is classified as one of the important species ecologically and commercially for Turkish forestry. It is also a target species of the “National Tree Breeding and Seed Production Programme” (Koski, Antola 1993) because of its valuable wood and non-wood products, social-cultural importance for rural area, and adaptation ability to various and changeable ecological conditions such as climate. Taurus cedar is used widely plantation, afforestation and other forestry practices such as forest restoration as natural or exotic species due to these characteristics (Bilir, Kang 2021). Fertility data is one of the important guides to establish successful plantation to different environmental conditions, and to give future direction to breeding programme and other forestry practices. Fertility is also one of the major tools used for various purposes in theoretical and applied forestry (Griffin 1982; Xie, Knowles 1992; Bila 2000; Kang et al. 2003; Kang, Bilir 2021). However, fertility variation and linkage parameters have been estimated by reproductive traits (i.e., strobili, cone, flower, and seed) in various plant species (Xie, Knowles 1992; Roeder et al. 1989; Savolainen et al. 1993). While many studies have been carried out on fertility variation mainly by individual strobili production, estimations of cone and seed production in different plant species such as Cedrus libani were very limited (i.e., Bilir, Kang 2014; Yazıcı, Bilir 2017; Bilir, Özel 2017; Bilir, Kang 2021; Yazıcı et al. 2023). Additionally, variation in cone and seed production, and their fertility in Cedrus libani has not yet been comparatively investigated.
It is getting importance of quantity and quality of seed supply. Seed procurement and its quality using frontier techniques (Bernardes et al. 2022; Novikova et al. 2022; Novikova et al. 2023; Novikov et al. 2021a; Novikov et al. 2021b; Novikov et al. 2019) is an important stage of the program and practices. It is also known that quality of seed crop genetically improved together with other morphological (i.e., grading) and physiological characteristics play important roles in successful forestry practices (Yazıcı et al. 2023) such as plantation.
The aims of this study are to estimate variations of cone and seed productions, to calculate the fertility variation and linkage parameters (i.e., population size, gene diversity) in two natural populations of Cedrus libani based on cone and seed productions, and to discuss on possible forestry practices of the species.
Materials and Methods
Data Collection
Two natural populations of Taurus cedar were sampled at southern part of Türkiye (Figure 2, Table 1). Data of numbers of mature cone which of two-years (NC) and filled seed (NS) were collected from fifty trees selected phenotypic and 100 m distance each other in 2023. Four cones were harvested from each direction of sampled trees for seed extraction. Extracted seeds were floated to separate empty and filled seeds on water during 12-16 hours. After that, the number of empty and filled seeds was counted (Figure 3), and the percentage of filled seed (NS%) was calculated.
Data Analysis
The ANOVA-test of the SPSS statistical package (SPSS 2011) was used as a tool for analyzing the productivity indicators of cones and seeds in the Taurus cedar populations studied in this research. If we define the letter l for the number of each surveyed tree, and the letter k for the population number, then the indicator variant is expressed by the symbol Ykl(C) or Ykl(S) for the productivity of cones (C) or productivity of seeds (S), respectively. A variant of the productivity indicator Ykl includes the terms μ + Rl + ekl, denoting for each specific Taurus cedar population k the average value of μ, the random effect of Rl in the k-th population (in this study k = 2, the populations were designated as P1 and P2), and the error ekl of determination of the model. Guided by the methodology for calculating the fertility of cones (ΨC) and seed fertility (ΨS), presented by Kang and Lindgren (1999), and Bilir (2011), the fertility of the l-th tree was determined as the proportion of individual cones or individual seeds in the k population. In this case (ΨC) is the sum of one and the square of the coefficient of variation CVCF of the fertility values of cones, and (ΨS) is the sum of one and the square of the coefficient of variation CVS of the fertility values of seeds of each l-th (l = 1...N) tree. Further, as shown in Kang, Lindgren 1998, Park et al. 2017, Genetic Diversity (GD) was calculated as the difference between one (1) and half (1/2) of the Np index characterized by Kang et al. (2003) a quotient of dividing the census number N by the corresponding fertility Ψ.
Results and Discussion
Cone and Seed Production
The averages of cone and seed number in P1 and P2 populations were 90 and 33, and 5321 and 3115 per tree, respectively. Averages of cone production were 46 ranged from 19 to 76 for years of pooled aspects (Yazıcı, Bilir 2017), and 20 and 22 in two populations of Taurus cedar (Bilir, Kang 2014). Percentage of filled seed were 94% and 83% in the populations. Among individual trees within population, and between populations for the numbers of cone, seed and filled seeds showed large differences (Table 2). For instance, there were more than twenty times differences among individuals in both populations for number of filled seed (Table 2). Populations had significant differences (p<0.05) for cone and seed production, and percentage of filled seeds depending on results of analysis of variance. The results showed importance of selection of individual tree and populations for higher reproductivity. The most productive 10 trees (20% of total trees) produced 40% and 35% of total cone production in P1 and P2, respectively, while they were 48% and 41% for number of filled seeds in the population. There could be many biotic (i.e., population) and a-biotic (i.e., altitude, crown closure) factors effective on these differences (i.e., Bila, Lindgren 1998; Bilir et al. 2005; Yazıcı, Bilir 2017 and 2023; Çatal et al. 2018; Çerçioğlu, Bilir 2018; Kang, Bilir 2021; Yazıcı et al. 2023; Bilir, Yazıcı 2024). Parental-balance curves in the populations were shown by means of cumulative gamete contribution in Figure 4. In both populations, individual seed productions were much closer to equal contribution than cone production (Figure 4). However, both characteristics could be balanced by forestry practices.
Large differences in reproductivity were reported among trees within population and among populations in Taurus cedar (i.e., Evcimen 1963; Odabaşı 1990; Bilir, Kang 2014; Çatal et al. 2018; Bilir, Kang 2021; Yazıcı, Bilir 2023), and in various plant species (e.g., Shea 1987; Kang et al. 2003; Bilir et al. 2005; Kang, Bilir 2021). Estimated coefficient of variations (CV) of the productions (Table 2) could be acceptable for natural populations CV ≤ 40% (Kang, Bilir 2021).
The average of mature cone number was reported 21 in the species (Bilir, Kang 2014), while averages of cone number were 47 (Yazıcı, Bilir 2017), while result of the study had higher cone productions (Table 2). The results indicated importance of populations and years in cone production. Good seed year had once in two-three years depending on altitude in natural populations of the species (Boydak 2003). However, present study had one-year data. Differences in biotic and a-biotic characteristics of populations may have impact on reproductive traits in the natural forest. Besides, similar differences were reported among individuals within population, and among populations for cone production and number of filled seeds in natural populations of Taurus cedar (Evcimen 1963).
Fertility Variation
Estimations of the cone and seed fertility, and linkage parameters were given in Table 3. As seen from Table 3, seed fertility (ΨS) was higher than cone fertility (ΨC) in both populations.
Cone and seed fertility variations (ΨC &ΨS) estimated of the proportion of individual production in the population were acceptable for ideal population suggested for natural stands (Ψ<3) (Kang 2001; Kang, Bilir 2021). Seed fertility (ΨS) was lower than that of cone (ΨC) in both populations (Table 3). Fertility variations varied according to populations, years and reproductive traits in Taurus cedar (Bilir, Kang 2014 and 2021; Yazıcı, Bilir 2023; Yazıcı et al. 2023).
Effective number of parents (Np) which of mirror of the fertility variation was ranged from 30.1 (for Ns in P1, for 60% of census number) to 41.4 (for Nc in P2 for 83%) (Table 3). The results of the present study showed that about 83% of individuals for Nc in the P2 behavior as they are under the ideal population. The size, equivalent to the target populations, was 12% (Nc) larger than Ns in P2. Difference of gene diversity was 0.004 between Nc and Ns (0.987 and 0.983) in P1 (Table 3). The results indicated importance of reproductive traits used in the estimations. Gene diversity was higher in P2 than P1 in both traits (Table 3). However, it could be increased by forestry practices to harvest seed crop which had higher gene diversity for sustainable and adaptable forestry to different environmental conditions such as climate change.
Various reproductive traits such as cone, strobili, fruit and seeds production have been used to estimate fertility variation and linkage parameters in many plant species (Shea 1987; Roeder et al. 1989; Savolainen et al. 1993; Kang 2001; Bilir 2011; Kang, Bilir 2021). However, cone and seed were the last stage of reproductivity after pollination. Besides, data collection on cone and seed productions had many advantages than that of strobilus count such as easy, cheap and more accurate because of longer staying and bigger size of cone for researcher as also emphasized in different studies (Bilir, Kang 2014; Bilir, Kang 2021). Reproductive data of the study belonged to only two populations and one year. Therefore, more data is needed from different populations and years including good and poor seed year for estimation fertility variation and linkage parameters in the species.
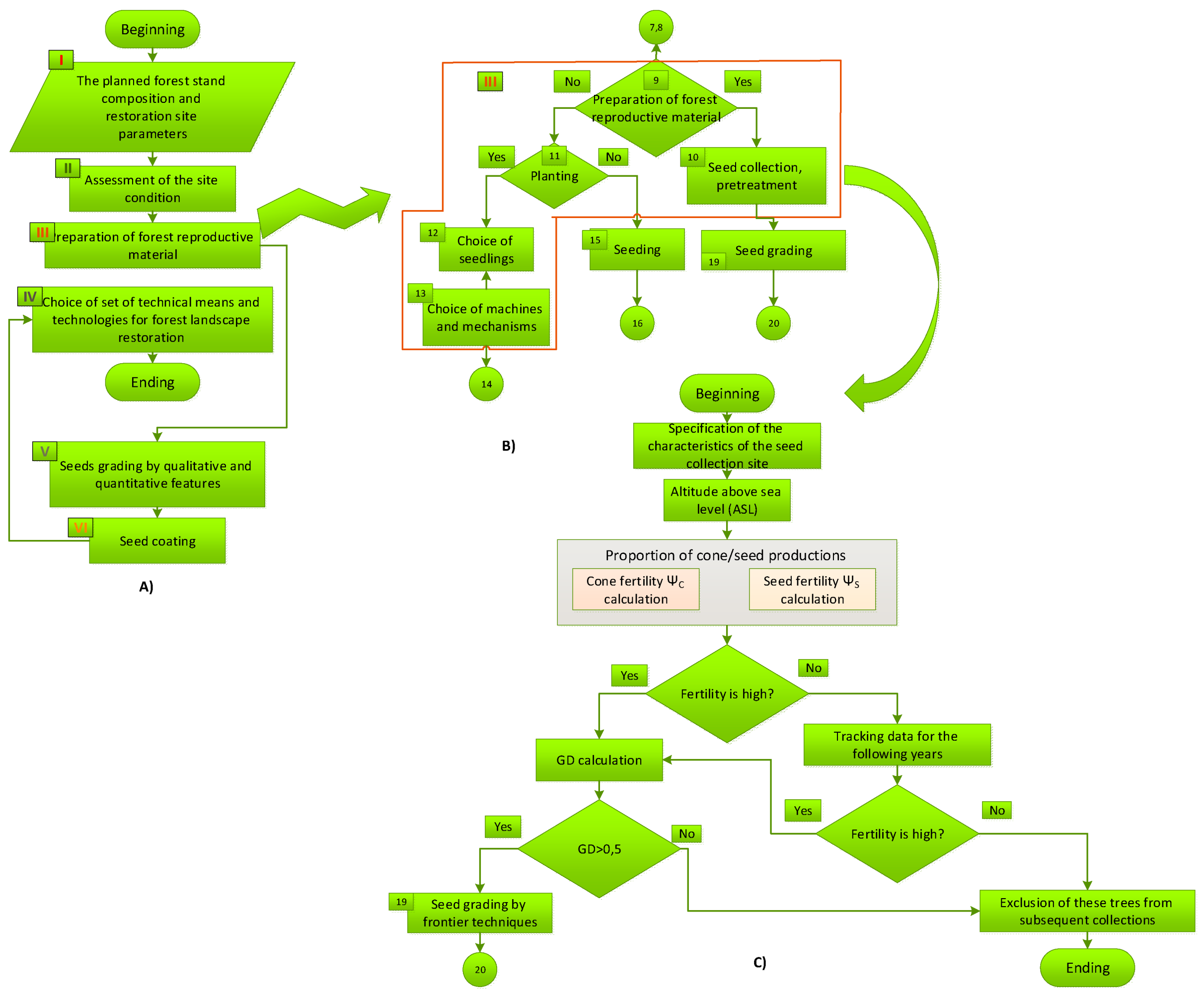
For the practical application of the data from this study, the following must be done. In the tenth block (Novikova, 2022a) (Appendix A, Figure A.1, b) of the third group of the FLR-algorithm, the structure of which consists of six basic groups (Novikova 2022b) (Appendix A, Figure A.1, a) on the technological operations of the forest restoration process, it is necessary to additionally include data from this study (Appendix A, fig. A.1, c), along with data from a previous study (Yazıcı et al. 2023).
Conclusions
Results of the present study belong to one year, and limited area and number of populations of Taurus cedar. Future studies should be carried to give large conclusions depending of geographic variation in natural distribution of Taurus cedar. However, results of the study could be used local area of the species for different purposes such as balancing of variation in seed crop for higher gene diversity by various forestry practices. Fertility variation should be observed by local foresters for transmit gene diversity to future generations as an environmental friendly.
Acknowledgements
The synthesis of samples and genetic research was carried out by N.B. The development, conceptualization and validation of the FLR-model was carried out by T.P.N. and A.I.N. within the framework of the grant of Russian Science Foundation (RSF), grant number 23-26-00102, https://rscf.ru/project/23-26-00102/.
Conflicts of Interest
The authors reported no potential conflicts of interest.
Appendix A
Figure A1.
FLR-algorithm of the forest restoration technology (A), decomposing of the «Preparation of forest reproductive material» module (B), modification of block III of the module 10 (Yazıcı et al. 2023) by evaluation algorithm operators of Taurus cedar cone and seed fertility (C) used in this study. Figures A) and B) are adapted and modification from the paper by co-author T.N. (Novikova 2022a), figure (C) is the T.N. own composition.
Figure A1.
FLR-algorithm of the forest restoration technology (A), decomposing of the «Preparation of forest reproductive material» module (B), modification of block III of the module 10 (Yazıcı et al. 2023) by evaluation algorithm operators of Taurus cedar cone and seed fertility (C) used in this study. Figures A) and B) are adapted and modification from the paper by co-author T.N. (Novikova 2022a), figure (C) is the T.N. own composition.
References
- Bernardes, R.C.; De Medeiros, A.; da Silva, L.; Cantoni, L.; Martins, G.F.; Mastrangelo, T.; Novikov, A.; Mastrangelo, C.B. Deep-Learning Approach for Fusarium Head Blight Detection in Wheat Seeds Using Low-Cost Imaging Technology. Agriculture 2022, 12, 1801. [Google Scholar] [CrossRef]
- Bila, A.D. (2000). Fertility variation and its effects on gene diversity in forest tree populations [. Ph.D. Thesis, ] Umeå, Sweden, Swedish University of Agricultural Science, Acta Universitatis Agriculturae Sueciae.
- Bila A.D. and Lindgren D. (1998). Fertility variation in Milletia stuhlmannii, Brachystegia spiciformis, Brachystegya bohemii and Leucaena leucocephala and its effects on relatedness in seeds. For. Genet. 5, 119–129.
- Bilir, N. (1997). Nursery stage of provanence on Taurus cedar (Cedrus libani A. Rich) in Eastern Black Sea region [. MSc. Thesis, ] Trabzon, Türkiye.
- Bilir, N.; Kang, K.-S.; Lindgren, D. Fertility variation in six populations of Brutian pine (Pinus brutia Ten.) over altitudinal ranges. Euphytica 2005, 141, 163–168. [Google Scholar] [CrossRef]
- Bilir, N.; Prescher, F.; Lindgren, D.; Kroon, J. Variation in cone and seed characters in clonal seed orchards of Pinus sylvestris. New For. 2008, 36, 187–199. [Google Scholar] [CrossRef]
- Bilir, N. (2011). Fertility variation in wild rose (Rosa canina) over habitat classes. Int. J. Agric. Biol., 13, 110–114.
- Bilir, N. and Kang K.-S. (2014). Estimation of fertility variation by strobili and cone productions in Taurus cedar (Cedrus libani A. Rich.) populations. In: Proceedings of the Proceedings of the IUFRO Forest Tree Breeding Conference; Prague, Czech Republic, 25-29 August, 2014.
- Bilir, N. and Özel H.B. (2017). Fertility variation in a natural stand of Taurus cedar (Cedrus libani A. Rich.). In: Proceedings of the International Forestry and Environment Symposium (IFES); Trabzon, Türkiye, 7-10 November, 2017.
- Bilir, N.; Kang, K.-S. Fertility variation, seed collection and gene diversity in natural stands of Taurus cedar (Cedrus libani). Eur. J. For. Res. 2021, 140, 199–208. [Google Scholar] [CrossRef]
- Bilir, N.; Yazici, N. Effects of climatic factors on strobilus production of Taurus cedar (Cedrus libani A. Rich.) populations. Theor. Appl. Clim. 2023, 155, 2151–2159. [Google Scholar] [CrossRef]
- Boydak, M. Regeneration of Lebanon cedar (Cedrus libani A. Rich.) on karstic lands in Turkey. For. Ecol. Manag. 2003; 178, 231–243. [Google Scholar] [CrossRef]
- atal, Y., Bilir N. and Özel H.B. (2018). Effect of growth characteristics on cone and seed production in Taurus cedar (Cedrus libani A. Rich.). Fresenius Environmental Bulletin, 27, 3832-3836.
- erçioğlu, M. and Bilir N. (2018). Altitudinal fertility variation in natural populations of Anatolian black pine [Pinus nigra Arnold. Subsp. pallasiana (Lamb.) Holmboe]. Human Journals, 4(8), 136-142.
- Eler, U. (1990). Seed yield in Calabrian cluster pine (Pinus brutia Ten.) by age. In Forest Research Institute, Technical Bulletin; Forest Research Institute: Antalya. Türkiye; 53–78. [Google Scholar] [CrossRef]
- Evcimen, B. S (1963). Commercial importance and management of Turkish Taurus cedar. Ankara, Türkiye, General Directorate of Forestry Press.
- Griffin, A.R. (1982). Clonal variation in radiata pine seed orchards. I. Some flowering, cone, and seed production traits. Australian Forest Research, 12, 4, 295-302.
- Kamalakannan, R.; Varghese, M.; Park, J.-M.; Kwon, S.-H.; Song, J.-H.; Kang, K.-S. Fertility Variation and its Impact on Effective Population Size in Seed Stands of Tamarindus indica and Azadirachta indica. Silvae Genet. 2015, 64, 91–99. [Google Scholar] [CrossRef]
- Kang, K.-S. Kang K.-S. and Lindgren D. (1998). Fertility variation and its effect on the relatedness of seeds in Pinus densiflora, Pinus thunbergii and Pinus koraiensis clonal seed orchards. Silvae Genet., 47, 196–201.
- Kang, K.-S. and Lindgren D. (1999). Fertility variation among clones of Korean pine (Pinus koraiensis S. et Z.) and its implications on seed orchard management. For Genet 6:191–200.
- Kang, K.-S. (2001). Genetic gain and gene diversity of seed orchard crops [PhD Thesis], Umeå, Sweden: Swedish University of Agricultural Science.
- Kang, K.-S.; Bila, A.; Harju, A.; Lindgren, D. Estimation of fertility variation in forest tree populations. For. Int. J. For. Res. 2003, 76, 329–344. [Google Scholar] [CrossRef]
- Kang, K.-S. and Bilir N. (2021). Seed orchards (Establishment, Management and Genetics. Ankara, Türkiye, OGEM-VAK Press, 189.
- Koski, V. and Antola J. (1993). National tree breeding and seed production programme for Türkiye 1994-2003. [online] Available at: https://ortohum.ogm.gov.tr [accessed on May 12, 2024].
- I Novikov, A.; Sokolov, S.V.; Drapalyuk, M.; Zelikov, V.; Ivetić, V. Performance of Scots Pine Seedlings from Seeds Graded by Colour. Forests 2019, 10, 1064. [Google Scholar] [CrossRef]
- Novikov, A.I.; Zolnikov, V.K.; Novikova, T.P. Grading of Scots Pine Seeds by the Seed Coat Color: How to Optimize the Engineering Parameters of the Mobile Optoelectronic Device. Inventions 2021, 6, 7. [Google Scholar] [CrossRef]
- Novikov, A.; Lisitsyn, V.; Tigabu, M.; Tylek, P.; Chuchupal, S. Detection of Scots Pine Single Seed in Optoelectronic System of Mobile Grader: Mathematical Modeling. Forests 2021, 12, 240. [Google Scholar] [CrossRef]
- Novikova, T.P.; Mastrangelo, C.B.; Tylek, P.; Evdokimova, S.A.; Novikov, A.I. How Can the Engineering Parameters of the NIR Grader Affect the Efficiency of Seed Grading? Agriculture 2022, 12, 2125. [Google Scholar] [CrossRef]
- Novikova, T. Study of a set of technological operations for the preparation of coniferous seed material for reforestation. For. Eng. J. 2022, 11, 150–160. [Google Scholar] [CrossRef]
- Novikova, T.P. The Choice of a Set of Operations for Forest Landscape Restoration Technology. Inventions 2022, 7, 1. [Google Scholar] [CrossRef]
- Novikova, T.P.; Tylek, P.; Mastrangelo, C.B.; Drapalyuk, M.V.; Kharin, S.V.; Novikov, A.I. The Root Collar Diameter Growth Reveals a Strong Relationship with the Height Growth of Juvenile Scots Pine Trees from Seeds Differentiated by Spectrometric Feature. Forests 2023, 14, 1164. [Google Scholar] [CrossRef]
- Odabaşı, T. (1990). Research on cone and seed characteristics of Cedrus libani. [online] Available at: https://www.ogm.gov.tr [accessed on Jun 12, 2024].
- Park, J.-M.; Kwon, S.-H.; Lee, H.-J.; Na, S.-J.; El-Kassaby, Y.A.; Kang, K.-S. Integrating fecundity variation and genetic relatedness in estimating the gene diversity of seed crops:Pinus koraiensisseed orchard as an example. Can. J. For. Res. 2017, 47, 366–370. [Google Scholar] [CrossRef]
- Roeder, K.; Devlin, B.; Lindsay, B.G. Application of Maximum Likelihood Methods to Population Genetic Data for the Estimation of Individual Fertilities. Biometrics 1989, 45, 363–379. [Google Scholar] [CrossRef]
- Savolainen, O.; Karkkainen, K.; Harju, A.; Nikkanen, T.; Rusanen, M. Fertility Variation in Pinus sylvestris: A Test of Sexual Allocation Theory. Am. J. Bot. 1993, 80, 1016. [Google Scholar] [CrossRef]
- Savolainen, O.; Karkkainen, K.; Harju, A.; Nikkanen, T.; Rusanen, M. Fertility variation in Pinus sylvestris: a test of sexual allocation theory. Am. J. Bot. 1993, 80, 1016–1020. [Google Scholar] [CrossRef]
- SPSS (2011). IBM SPSS Statistics for Windows, Version 20.0., NY: IBM Corp.
- Yazici, N.; Bilir, N. Aspectual Fertility Variation and Its Effect on Gene Diversity of Seeds in Natural Stands of Taurus Cedar (Cedrus libani A. Rich.). Int. J. Genom. 2017, 2017, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Yazici, N.; Bilir, N. Impact of Crown Closure on Cone Production and Effective Number of Parents in Natural Stands of Taurus Cedar (Cedrus libani A. Rich.). Forests 2023, 14, 1130. [Google Scholar] [CrossRef]
- Yazici, N.; Novikova, T.P.; Novikov, A.I.; Bilir, N. Gene Diversity In Seed Crop Of Taurus Cedar (Cedrus Libani A. Rich.) Over An Altitudinal Range. Geogr. Environ. Sustain. 2023; 16, 63–71. [Google Scholar] [CrossRef]
- Xie, C.Y.; Knowles, P. Male fertility variation in an open-pollinated plantation of Norway spruce (Piceaabies). Can. J. For. Res. 1992, 22, 1463–1468. [Google Scholar] [CrossRef]
Figure 1.
Male strobili and cones of Taurus cedar.

Figure 2.
Sampled P1 (a) and P2 (b) Taurus cedar populations.


Figure 3.
Mature cones (a) and filled seeds (b) of the species used in this study.


Figure 4.
Parental-balance curves in the P1 (a) and P2 (b) populations.
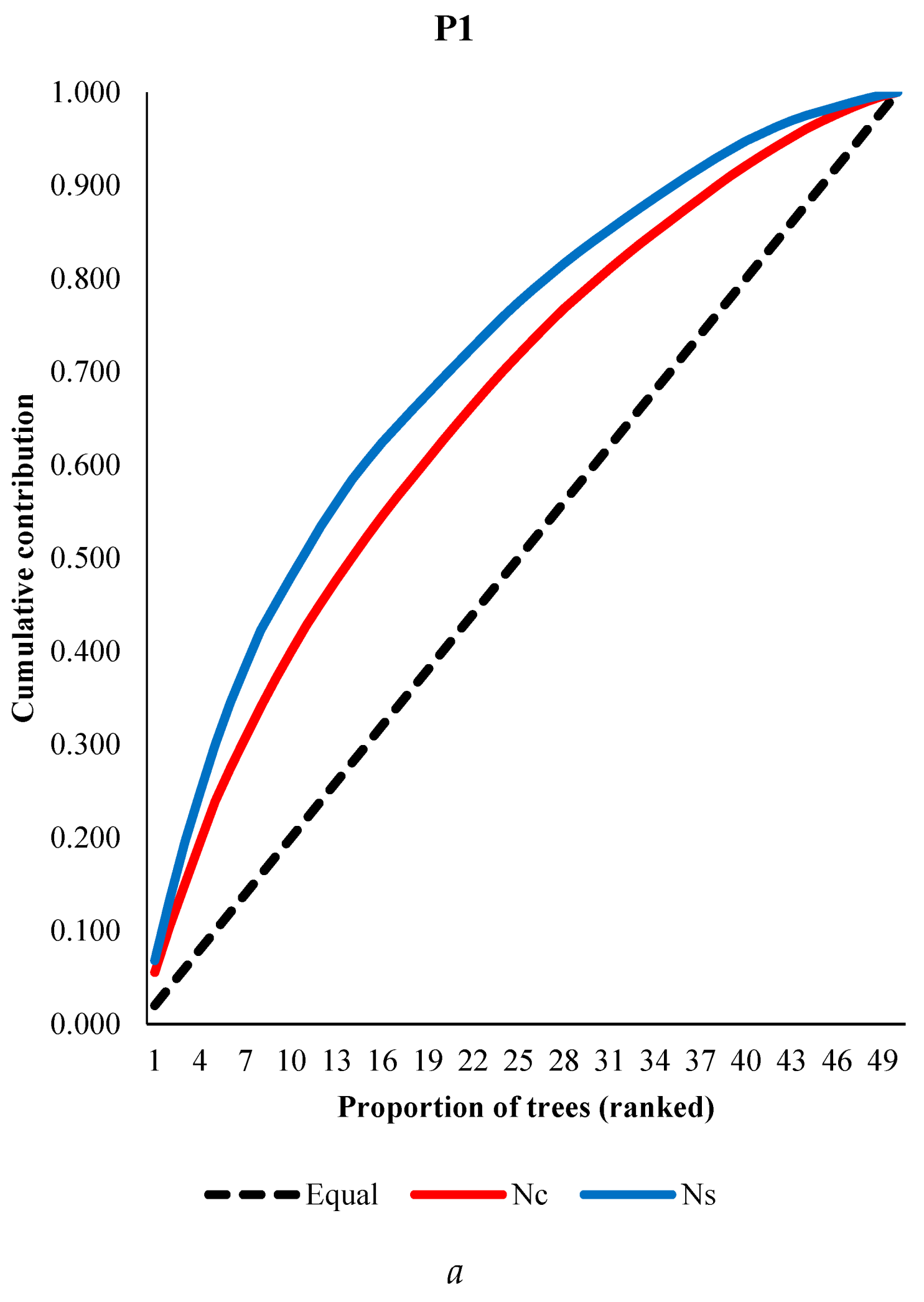
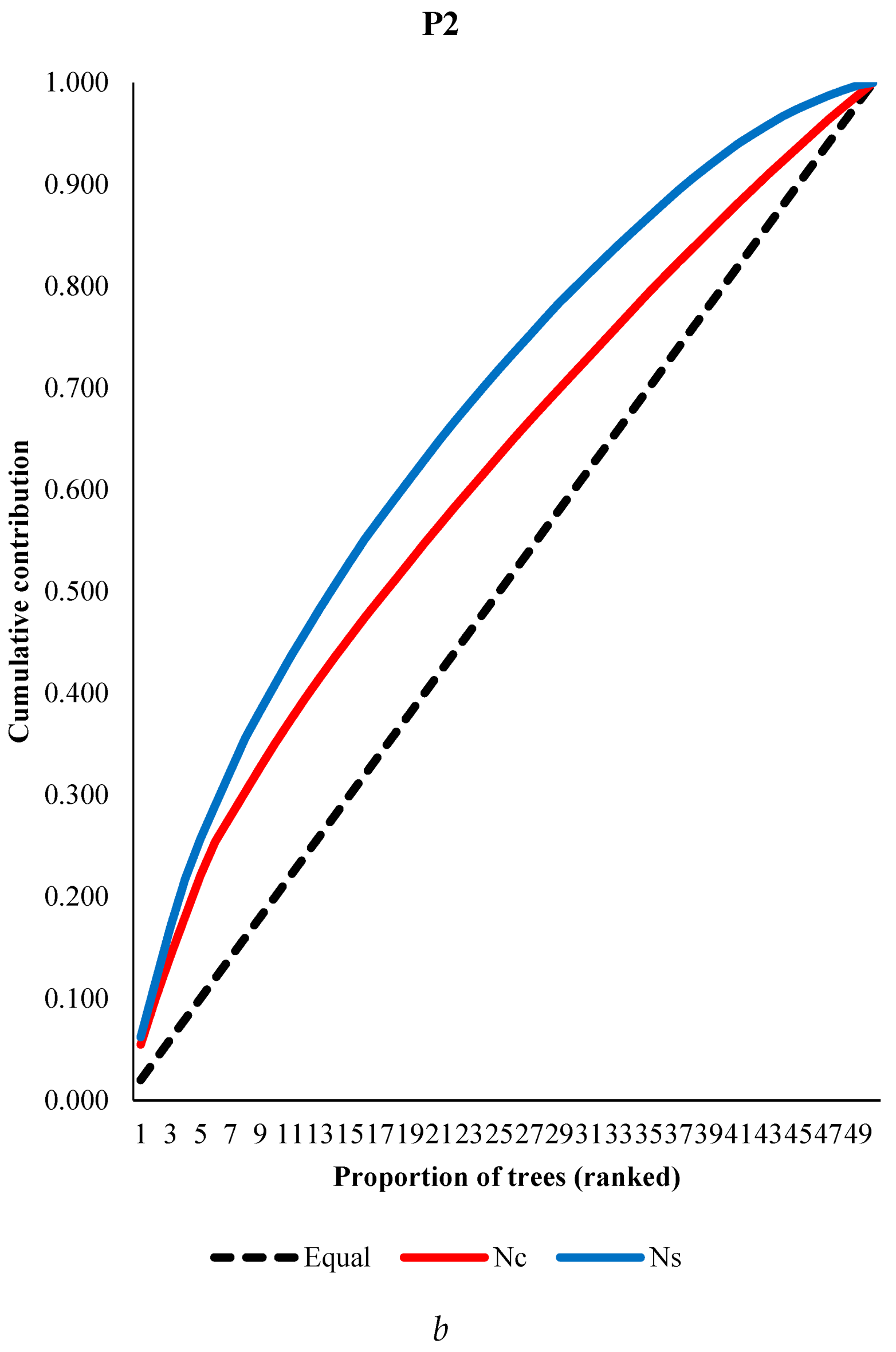

Table 1.
Geographic details of studied Taurus cedar populations.
| Populations | Age (year) |
Latitude (N) | Longitude (E) | Average Altitude (m) | Aspect |
|---|---|---|---|---|---|
| P1 | 60 | 38°49′ | 30°45′′ | 1500 | North |
| P2 | 120 | 37°40′ | 30°51′ | 1580 | Northwest |
Table 2.
Basic statistical values for cone and seed production in the populations.
| Statistics | P1 | P2 | ||||
|---|---|---|---|---|---|---|
| NC | NS | NS% | NC | NS | NS% | |
| Mean | 90.2 | 5320.5 | 93.5 | 32.9 | 3114.9 | 83.3 |
| Range | 20-250 | 750-18144 | 78.3-99.7 | 20-90 | 418-9685 | 39.5-97.2 |
| CV% | 60.1 | 82.3 | 5.2 | 46.1 | 64.2 | 16.9 |
Table 3.
Cone and seed fertility (ΨC &ΨS), effective number of parents (Np(c) & Np(s)) and gene diversity (GD) in the populations.
Table 3.
Cone and seed fertility (ΨC &ΨS), effective number of parents (Np(c) & Np(s)) and gene diversity (GD) in the populations.
| P1 | P2 | |||
| NC | NS | NC | NS | |
| ΨC &ΨS | 1.35 | 1.66 | 1.21 | 1.41 |
| Np(c) & Np(s) | 36.97 (0.74)* | 30.06 (0.6) | 41.40 (0.83) | 35.41 (0.71) |
| GD | 0.987 | 0.983 | 0.988 | 0.986 |
*; relative effective number of parents in the parenthesis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



