
Submitted:
06 December 2024
Posted:
06 December 2024
You are already at the latest version
Abstract
Buchwaldoboletus xylophilus is an edible boletes species belonging to the family Boletaceae and the genus Buchwaldoboletus, is found in tropical and subtropical regions, and is known for its rare wild resources. In this study, wild B. xylophilus was isolated and cultured, and its biological characteristics and artificial cultivation techniques were studied. The results showed that the optimal carbon source, nitrogen source and inorganic salt for the mycelium growth of B. xylophilus were maltose, ammonium tartrate and magnesium sulfate, respectively. The most appropriate temperature is 28 ℃ and the pH value is between 5 and 6. The most effective combination was determined by orthogonal experiment as follows: glucose, ammonium nitrate, potassium dihydrogen phosphate and 28 ℃. The results of artificial cultivation in mushroom houses showed that the mycelium of B. xylophilus was strong and grew well on the culture medium. The mycelial growth rate is 4.54 mm/d, and the fungus bags are filled about 50 days after inoculation. The primordium is formed 9 to 14 days after covering with soil and the fruiting body matured in 6 ~ 8 days. The average yield of fresh mushrooms reached 131.07±29.38 g/bag, and the average biological efficiency reached 28.48±6.39 %. In this study, the artificial cultivation technology of B. xylophilus in mushroom house was reported for the first time. The fruiting bodies obtained through cultivation were identified by morphological and molecular biological methods. This technology offers benefits such as affordability, a brief cultivation cycle, substantial yields, and superior quality, making it ideal for industrial-scale and extensive cultivation.
Keywords:
artificial cultivation
; Bolete
; Buchwaldoboletus xylophilus
; fruiting body
; edible mushrooms
1. Introduction
Boletes is one of the important groups of macrofungi [1,2,3]. China has abundant resources of boletes, and nearly 500 species have been reported [4,5,6]. The majority of boletes species are mycorrhizal fungi, which can establish a mutually beneficial symbiotic relationship with various higher plants, including Pinaceae, Salicaceae, Betulaceae, Casuarinaceae, Fabaceae and Dipterocarpaceae [7,8,9,10,11]. However, the artificial cultivation of boletus remains challenges. At present, there are only a few types of boletes that can be artificially cultivated. In 1969, Pilat took Buchwaldoboletus lignicola as a model species and formally established the genus Buchwaldoboletus and classified it into the Boletaceae family [12]. This genus distributed all over the world, and 12 species have been described so far [13]. Predominantly, Buchwaldoboletus species are wood-rot fungi, typically found growing on decomposed wooden stakes, decomposed rubber sawdust, and in the bamboo forests [14,15,16].
Buchwaldoboletus xylophilus (Petch) Both & B. Ortiz, is an infrequently encountered boletes species native to tropical regions with the wild fruiting bodies which are exceptionally rare and are known to occur in select locations within China, including Xishuangbanna, Hainan and Hong Kong [13,17]. They are also distributed in Sri Lanka, Malaysia, India, and the Philippines [13,18,19,20]. In 2020, we collected a rare species of boletes in the rubber woodland of Xishuangbanna Dai Autonomous Prefecture, Yunnan Province. Through morphological and molecular biological identification, it was determined that the species is a member of the family Boletaceae and the genus Buchwaldoboletus, namely B. xylophilus (Petch) Both & B. Ortiz [17]. B. xylophilus is an edible mushroom with a delicious taste and a crispy texture. In the Yunnan region, it can be sold at a price ranging from CNY 60 to 100 per kilogram (approximately to USD 8 - 14). Additionally, the market demand for it is expected to continuously expand.
The fruiting body of B. xylophilus, is moderate size and can be found either solitary or in clusters, boasts an aesthetically pleasing mushroom form. This species also displays a yellow-brown to brown cap and a stipe that ranges from yellow-brown to reddish-brown. This species also displays a distinctive yellow mycelium at the stipe’ s base. Additionally, its spores are diminutive, typically oval or spherical in shape [17]. The flesh is yellowish and turns blue rapidly after injury. As time passes, the color of the injured part gradually darkens [17].
B. xylophilus is an endangered species with few wild resources and low wild yield. Therefore, the biological characteristics and cultivation of B. xylophilus were studied, laying a foundation for the resource protection and development and utilization of B. xylophilus.
2. Materials and Methods
2.1. Isolation
Fruiting bodies of wild B. xylophilus were collected from the rubber forest of Jinghong City, Xishuangbanna Dai Autonomous Prefecture, Yunnan Province (Figure 1). The morphological structure was recorded. The soil particles at the root of the fruiting body were removed. The mycelia were isolated from the fruiting bodies and cultured on PD medium (potato 200.0 g, glucose 20.0 g, yeast extract 2.0 g, MgSO4 1.0 g, KH2PO4 1.0 g, agar 18.0 g, water up to 1 L, pH not adjusted). The isolated strains were named YITC-BU001, YITC-BU002 and YITC-BU003 respectively. And YITC-BU001 strain was used for the experiments of biological characteristics. Mycelial cultures were incubated at 28 ℃ for 15 days. The cultures were kept on PD slants at 15 ℃.
2.2. Effect of Carbon and Nitrogen Source
To assess mycelial growth, six different carbon source media were utilized. Potato dextrose PD medium, devoid of glucose, was added with 20 g/L carbon source comprising one of the following: glucose, fructose, sucrose, mannitol, maltose, or soluble starch. The medium was sterilized by autoclaving at 121 ℃ for 30 minutes. 20 mL of the media was dispensed into each 9-cm-diameter Petri dish. A 5-mm-diameter plug of mycelium from the edge of a 15-day-old culture was inoculated on the media. Each Petri dish was sealed with Parafilm to mitigate contamination risks. Incubation of the cultures was carried out in the dark at 28 ℃. The Colony diameters were measured at 5-day intervals, and each treatment was repeated 10 times.
Six distinct nitrogen source media were employed to evaluate mycelial growth. PD medium (without yeast extract) was added with 2 g/L nitrogen source comprising one of the following: tryptone, beef extract, yeast extract, ammonium tartrate, KNO3 and NH4NO3. The subsequent procedures were identical to those of the carbon source experiment.
2.3. Effect of Inorganic Salt
Six inorganic salt media were used to test the growth ability of mycelia. PD medium (without MgSO4, KH2PO4) was added with 1 g/L inorganic salt comprising one of the following: NaCl, MgSO4, KH2PO4, MnSO4, CaSO4, NaNO3. Cultures were incubated in darkness at 28 ℃. The colony diameter was measured every 5 days, and each treatment was repeated 10 times.
2.4. Effect of Temperature and pH
A 5-mm-diameter plug of mycelium was inoculated on PD medium plates from the edge of a 15-day-old culture, then incubated at 15, 20, 23, 25, 28, 30 and 35 ℃ in the darkness. The colony diameter was measured every 5 days, and each treatment was repeated 10 times.
A 5-mm-diameter plug of mycelium was inoculated on PD medium plates from the edge of a 15-day-old culture with different pH. The pH was adjusted to 4.0, 5.0, 6.0, 7.0 and 8.0 with 0.1 mol/L NaOH or 0.1 mol/L HCL. The cultures were incubated in darkness at 28 ℃. The colony diameter was measured every 5 days, and each treatment was repeated 10 times.
2.5. Orthogonal Experiment
According to the screening results of carbon source, nitrogen source, inorganic salt and temperature conditions, the best three levels were selected in each condition, and the orthogonal experiment of four factors and three levels was carried out. In the orthogonal experiment, the mother culture medium was replaced the corresponding carbon source, nitrogen source and inorganic salt, and the experiment was carried out at the corresponding temperature.
2.6. Preparation Of Liquid Culture
200 g potato and 20 g bran were boiled with 1 liter of water for 20 minutes, and the filtrate was collected. 15 g glucose, 2 g yeast extract, 1 g MgSO4, 1 g KH2PO4, and 2 tablets of vitamin B1 (included 10 mg of Vitamin B1) were added to the filtrate and dissolved, pH was not adjusted. The liquid medium was added to 1 L and sub-packed into a 500 mL triangular flask, each containing 250 mL of liquid medium. The media were autoclaved at 121 ℃ for 30 minutes. Four or five 0.5 cm3 plugs of mycelium were inoculated in the liquid media. Cultures were incubated on a shaker (140 rpm/min) in darkness at 28 ℃ for 7 days.
2.7. Solid Substrate Inoculums
The cultivation medium contained 40% rubber sawdust, 35% rubber wood bits, 15% bagasse, 8% corn flour, 1% sucrose and 1% calcium carbonate. The cultivation medium components were mixed in their respective proportions, and water was added to moisten the medium, adjusting its water content to 60~65%. Each polypropylene bag was filled with 1.0-1.1 kg of the cultivation medium. The media were autoclaved at 121 ℃ for 2 hours and cooled to ambient temperature before inoculation. A volume of 10~15 mL of liquid inoculum was inoculated into each cultivation bag (17×35×0.07 cm). The cultures were cultured in darkness at 28 ℃. The mycelium grew from the inoculation surface as the starting point, and the mycelium grew downward to the full bag as the end point. The distance from the inoculation site to the point of full colonization was measured to calculate the mycelial growth rate within the cultivation medium.
2.8. Fructification
The casing materials were composed of peat and vegetable garden soil mixed in a 1:1 volume ratio, casing layer with a thickness of about 3-4 cm. Once the bags were fully colonized by mycelium, their surfaces were cased with the soil mixture. The cased mushroom bags were transferred to the mushroom cultivation house. The temperature in the mushroom cultivation house was maintained at 28-30 ℃, and the relative humidity was 80%~85%.
When the mycelium grows to the surface of the casing layer and the primordia is formed, the cultures were subjected to a temperature of 27-29 ℃ and a light intensity of 1000 lx in a 12 hour light/dark cycle. The quantities of primordia formed within each bag as well as those of the mature fruiting bodies were duly recorded. Subsequently, the yield of fresh mushrooms was accurately measured and the biological conversion rate was calculated.
2.9. Morphological Observation and Molecular Identification
The morphological characteristics of the fruiting bodies of artificially cultivated B. xylophilus were observed and described. The morphology of mycelia and basidiospores were observed under Zeiss microscope (ZEISS AxioScope A1), at a magnification of 40 times (40X).
Genome DNA of fruiting bodies of artificially cultivated B. xylophilus strains YITC-BU001, YITC-BU002 and YITC-BU003 was extracted by plant genome extraction kit (Beijing Tsingke Biotech Co., Ltd.). Using ITS5/ITS4 and LR0R/LR5 as primer pairs, the ITS and 28S rDNA fragments of the strain were amplified by PCR. The parameters of PCR reaction were: pre-denaturation at 98 ℃ for 2 min; 98 ℃ for 10 s, 56 ℃ for 10 s and 72 ℃ for 10 s, totally 35 cycles. After amplification at 72 ℃, it was extended for 5 minutes. After the PCR amplification product was detected by 1.2% agarose gel electrophoresis, the DNA was purified and recovered by purification kit, and the amplification products were sent to Qingke Biotechnology Co., Ltd. for gene sequencing. The sequencing results were spliced by Contig Express software and submitted to GenBank. Based on the outcomes of the BLAST online comparison, the strains featuring high coverage and similarity were chosen, and the phylogenetic tree was constructed through cluster analysis using the software MEGA 6.0.
3. Results
3.1. Strain Isolation and Mycelial Growth
The preserved slant cultures were inoculated onto a PD petri dishes (9.0 cm). Upon cultivation on the medium, the colonies achieved a diameter of 7.0 cm within 15 days. The periphery of the mycelium exhibited hues ranging from pale yellow to bright yellow, presenting a robust and well-defined appearance (Figure 2). The mycelium near the inoculation point was yellow and white fluffy. There was no pigment or sclerotia formation during the culture process.
3.2. Effect of Carbon and Nitrogen Source on Mycelial Growth
B. xylophilus was observed to grow on all media. The mycelia grew fastest on the medium with maltose as carbon source, with a growth rate of 1.70 ± 0.07 mm/d (Table 1). The mycelium was thick, dense, and yellow. The mycelium also grew well on the medium with glucose as carbon source, with a growth rate of 1.63 ± 0.07 mm/d. When cultivated on a medium with sucrose or soluble starch serving as the carbon source, the mycelial growth was observed to be slower, sparse, and lacking vigor, recording growth rates as low as 0.64 ± 0.14 mm/d and 0.91 ± 0.09 mm/d, respectively. The selection of carbon source is very important in the artificial cultivation of B. xylophilus, and maltose is the better carbon source.
Ammonium tartrate can promote the mycelial growth of B. xylophilus (Table 2). The most rapid mycelial growth was noted on a medium where ammonium tartrate was utilized as the nitrogen source, achieving a growth rate of 1.77 ± 0.04 mm/d. Additionally, the mycelium developed the greatest density on this medium. The mycelium thrived on the medium supplemented with ammonium nitrate as a nitrogen source, exhibiting a growth rate of 1.58 ± 0.13 mm/d. However, the edge of the mycelium grew sparsely. The mycelial growth rate was the slowest on the medium with potassium nitrate as the nitrogen source. Ammonium nitrate was the most suitable nitrogen source.
3.3. Effect of Inorganic Salt on Mycelial Growth
The mycelial growth was most vigorous on the medium containing MgSO4 as inorganic salt, with a growth rate of 1.49±0.06 mm/d (Table 3). The mycelial growth rate was the slowest on the medium with MnSO4 as inorganic salt, with a growth rate of 0.95±0.09 mm/d. There was no significant difference on the NaCl, KH2PO4, CaSO4, and NaNO3 inorganic salt media.
3.4. Effect of Temperature and pH on Mycelial Growth
The mycelium did not grow at 15 ℃. At a temperature of 20 ℃, the mycelium exhibited significantly slow growth rate of 0.49 ± 0.08 mm/d. With the temperature increased, the growth rate of mycelium also increased. When the temperature reached 28 ℃, the mycelial growth rate was the fastest, with a growth rate of 1.63 ± 0.10 mm/d, and the mycelium was thick and dense. The fast rate of mycelial growth was observed at 30 ℃, yet the mycelium was found to be sparse with irregularly shaped edges. When the temperature reached 35 ℃, the mycelium underwent fragmentation, adopted a white appearance, and exhibited irregular growth patterns. Considering mycelial growth rate and mycelial growth vigor, the optimal temperature was 28 ℃.

Table 4.
Effect of Temperature on mycelial growth.
| Temperature(℃) | Mycelial growth rate(mm/d) | Mycelial growth vigor |
| 15 | 0.00±0.00 | - |
| 20 | 0.49±0.08 | + |
| 23 | 0.99±0.07 | + |
| 25 | 1.23±0.04 | +++ |
| 28 | 1.63±0.10 | ++++ |
| 30 | 1.57±0.12 | +++ |
| 35 | 1.31±0.06 | + |
The pH had a certain influence on the mycelial growth of B. xylophilus. The mycelial growth rate was the fastest at pH values of 5 and 6, reaching 1.42 ± 0.14 mm/d and 1.41 ± 0.07 mm/d respectively. There was no significant disparity between the two, therefore the optimal pH range was from 5 to 6. When the pH was 7 or above, the mycelium became weakened and turned yellow white. B. xylophilus was more favorably adapted for growth in acidic environments.
Table 5.
Effect of pH on mycelial growth.
| pH | Mycelial growth rate(mm/d) | Mycelial growth vigor |
| 4 | 1.12±0.11 | ++ |
| 5 | 1.42±0.14 | +++ |
| 6 | 1.41±0.07 | +++ |
| 7 | 1.23±0.09 | ++ |
| 8 | 0.84±0.13 | + |
3.5. Orthogonal Experiment
Temperature had the greatest impact on the mycelial growth of B. xylophilus (Table 6). The temperature range was the largest, at 0.67. The second was the nitrogen source, with a range of 0.31. The influence of carbon sources and inorganic salts on the mycelial growth of B. xylophilus was minimal. The optimal conditions for the mycelia growth of B. xylophilus are glucose, NH4NO3, KH2PO4, and 28 ℃.
3.6. Cultivation of B. xylophilus
The mycelium of B. xylophilus was robust and appeared light yellow or yellow on the cultivation medium (Figure 3). The mycelial growth rate was 4.54 mm/d and the mycelium filled the bags about 50 days after inoculation.
After casing the soil for 7-12 days, the yellow mycelium grew to the surface of soil layer (Figure 4A).Then the cultures were incubated at 27~29 ℃ with light of 1000Lx (12h light/12h dark). After 2 days, the primordia were formed on the surface (Figure 4B). When the primordia grew to 1-2 cm, the pileus and the stipe began to differentiate and enter the stage of young mushroom growth (Figure 4C~F). About 6-8 days after the formation of the primordia, the fruiting body matured (Figure 4G~H). When the height of the fruiting body reached 8-12 cm and the pileus had not yet flattened, the fruiting bodies could be harvested.
The average time for the formation of B. xylophilus primordia after covering with soil is 11.28±1.50 days. A large number of primordia are densely spread on the surface of the overlying soil layer. The average number of young mushrooms and mature fruiting bodies per bag were 5.70±3.69 and 4.13±2.21, respectively. The average fresh mushroom yield of B. xylophilus is 131.07±29.38 g/bag, and the average biological efficiency is 28.48±6.39 % (Table 7).
3.7. Identification of Cultivated Mushrooms
3.7.1. Morphological Analysis
Compared with the wild fruiting body, the cultivated fruiting bodies of B. xylophilus had a similar morphology, yet they possessed a more vibrant hue (Figure 5A~C). The height of the fruiting body was 6~19 cm. The pileus was orange, with a diameter of 5~13 cm. The context and tubes were yellow. The thickness of context was 1~2.5 cm. The stipe was yellowish brown, with yellow villous hyphae at the base of the stipe (Figure 5D). The fruiting body quickly turned light blue or blue after injury (Figure 5E).
3.7.2. Characteristics of Phylogeny
BLAST was used for homology search, and similarity comparison with the sequences of near-source strains in the database (Table 8). Strain YITC-BU001's 28S rDNA gene sequence exhibits the highest similarity, with a value of 99.77%, to the wild B. xylophilus voucher FHMU5931-1 from Yunnan. The 28S rDNA gene sequences of strains YITC-BU002 and YITC-BU003 have the highest similarity with B. xylophilus voucher X.H. Deng1 (FHMU5848) from Hainan, with 99.88% and 99.76%, respectively. The 28S gene sequence of the effectively published strains with high coverage and similarity was selected for phylogenetic analysis. From the clustering results, it could be observed that the cultivated strains YITC-BU001, YITC-BU002 and YITC-BU003 in this study were clustered on the same branch as the wild B. xylophilus reported from Hainan and Yunnan (Figure 7).
The ITS gene sequences of strains YITC-BU001, YITC-BU003 and YITC-BU002 exhibited the highest similarity with wild B. xylophilus voucher FHMU5932 and FHMU5933-1 from Yunnan, which were 99.14%, 100%, and 99.12%, respectively. The phylogenetic tree was constructed by using ITS genes of strains with high ITS similarity and effective publication. The results showed that YITC-BU001, YITC-BU002 and YITC-BU003 were clustered on the same branch as the wild B. xylophilus reported from Hainan and Yunnan (Figure 8).
4. Discussion
The genus Buchwaldoboletus does not engage in mycorrhizal associations, instead, it is characterized as saprophytic or lignicolous [12,13]. In this study, the different carbon sources, nitrogen sources and inorganic salts had effects on the mycelium growth of B. xylophilus. The best combination was determined by orthogonal experiment as follows: glucose, ammonium nitrate and potassium dihydrogen phosphate. The results are consistent with the optimal carbon sources for mycelial growth of Phlebopus portentosus and Boletus edulis, yet different from nitrogen sources and inorganic salts [29,30]. Cheng et al [31] reported that glucose was the best carbon source for most macrofungi. B. xylophilus preferred ammonium nitrogen, such as ammonium tartrate and ammonium nitrate, while other edible fungi (such as Clitocybe fragrans, Polyporus tuberaster, Flammulina filiformis) preferred organic nitrogen sources such as peptone, beef extract and yeast powder [32,33]. This indicates that various mushroom species have distinct nutritional requirements.
In this study, the optimal growth temperature for B. xylophilus was determined to be 28 ℃ and the optimal pH was within the range of 5 to 6. When the temperature is below 15 ℃, the mycelium cannot grow, and when the temperature is above 30 ℃, the mycelium is sparse, white, and grows irregularly. This is similar to the optimal growth temperature and pH value for P. portentosus and B. edulis mycelium [29,34]. Phlebopus spongiosus has the highest mycelial growth rate and biomass at 30 ℃ and a pH value of 5 [35], while some edible mushrooms have lower mycelial growth temperatures, such as Leccinum aurantiacum and Lepista sordida [36,37].
Mycorrhizal edible fungi, such as B. edulis, Tricholoma matsuake, Tuber indicum and Thelephora ganbajun, are highly valued for their unique flavors and potential health benefits [38,39,40,41]. These fungi form symbiotic relationships with the roots of specific host plants, a process that is essential for their growth and reproduction [42]. The complexity of this symbiotic relationship poses distinctive challenges in the field of artificial cultivation. Artificial cultivation represents a crucial approach for the preservation, advancement, and utilization of wild edible fungi. It can not only protect endangered species but also fulfill the sampling needs for scientific research. Some scholars have been trying to research the cultivation of wild bolete mushrooms [43]. However, the majority of boletes fall into the category of ectomycorrhiza fungi. They form a mycorrhizal structure in association with higher plants like those in the Pinaceae and Fagaceae families. This characteristic makes them difficult to cultivate artificially in mushroom houses [44,45,46,47].
The genus Phlebopus is a tropical and subtropical unique edible fungus, which usually grows with Citrus maxima, Mangifera indica, Eriobotrya japonica and other fruit trees in the wild [48,49,50]. Among them, P. portentosus and Phlebopus roseus can form a special insect gall structure with plant roots and scale insects [28,51,52,53]. P. portentosus is the first edible bolete in the world to achieve industrial cultivation and currently. At present, China has realized year-round industrial cultivation and production, with the daily output of fresh mushrooms reaching up to 16 tons [29,54]. The method of artificial cultivation of P. spongiosus was reported, and the yield was increased through high-voltage pulsed stimulation [35,55]. P. portentosus is not a typical saprophytic edible fungus, it has certain saprophytic properties, however, its genome lacks or only has a small amount of genes related to lignin and cellulose degradation [56,57]. However, the cultivation research of other boletes species is still in the initial stage, such as B. edulis, B. tomentipes, and B. albidus, which are in the screening and optimization stage of mother culture medium and mycelial growth conditions, and the mature fruiting bodies have not yet been cultivated in mushroom houses [58,59].
Buchwaldoboletus is different from Phlebopus, which can be distinguished by habitats [15], and the molecular data from this study confirm their genetic remoteness from one another. In addition, P. portentosus and P. spongiosus will form a large number of sclerotia in the wild and pure culture [35,60], while B. xylophilus has not been observed to form sclerotia, neither in its natural environment nor under artificial cultivation conditions.
Separation from the host plants is a necessary condition for industrial year-round cultivation and production of edible fungi. In this study, the artificial cultivation of B. xylophilus in mushroom house was successfully achieved. The mycelium of B. Xylophilus grows strong in a culture medium containing of 40% rubber sawdust, 35% rubber wood bits, 15% bagasse, 8% corn flour, 1% sucrose and 1% calcium carbonate. The mycelium of P. portentosus and P. spongiosus grows well in rice and sawdust medium, while the former can also grow rapidly in sorghum and sawdust medium [29,35,61]. Adding 25% to 50% rice husk in eucalyptus sawdust can effectively increase the yield of Pleurotus ostreatus [62], suggesting that edible fungi varieties exhibit diverse utilization and requirements for cultivation substrates.
B. xylophilus has strong saprophytic properties. During the mycelial growth process, the cultivation substrates are gradually degraded, and the culture medium become soft. After casing with soil, one mushroom bag can mature 1-11 fruiting bodies, and the average fresh mushroom yield of B. xylophilus is 131.07±29.38 g/bag, with a biological efficiency of 28.48±6.39%. Inoculating Buchwaldoboletus hemichrysus or B. lignicola on wood alone, the mass of wood blocks decreased by 35~65% after 6 months of continuous culture [63], which indicated that the genus Buchwaldoboletus had strong ability to degrade lignin and cellulose.
5. Conclusions
In this paper, the biological characteristics of carbon source, nitrogen source, inorganic salt, temperature and pH of mycelial growth of B. xylophilus were identified. The artificial cultivation of B. xylophilus within a mushroom house has been achieved for the first time, which is a new breakthrough in the cultivation of wild edible mushroom, and great significance to the development of edible mushroom industry. This technology presents numerous advantages, such as affordability, a brief cultivation cycle, substantial yields, and superior quality. Thereby, it is highly suitable for industrial-scale and extensive cultivation applications.
Author Contributions
Conceptualization, T.W.Y., H.J.M. and C.X.Z.; methodology, T.W.Y., X.J.X. and C.X.Z.; software, L.M.D., Y.W.F. and T.W.Y.; validation, J.L., X.J.X. and F.G.; formal analysis, T.W.Y., S.P.J. and L.M.D.; investigation, M.X.H., J.L., F.G. and C.X.Z.; data curation, T.W.Y. and C.X.Z.; writing—original draft preparation, T.W.Y. and L.M.D.; writing—review and editing, H.J.M. and C.X.Z.; visualization, T.W.Y. and C.X.Z.; project administration, H.J.M. and C.X.Z.; funding acquisition, H.J.M., T.W.Y. and C.X.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Agricultural basic research joint special project of Yunnan province (No. 202301BD070001-067, No. 202101BD070001-084, and No. 202401BD070001-125), the National Natural Science Foundation of China (No. 32060707) and the Funds of Sci-Tech Innovation System Construction for Tropical Crops of Yunnan Province (No. 655-4-3).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequence data generated in this study can be obtained from NCBI GenBank (http://www.ncbi.nlm.nih.gov/, accessed on 22 May 2023). The data included in this study are available through contacting the authors.
Acknowledgments
We are very grateful to Zhenquan He for his help in sample collection and field investigation. And grateful to Wangguang Qi, Nengqiu Fan and Feng Wang for their help in the artificial cultivation of B. xylophilus in mushroom house. We are deeply grateful to Dr. Yuanzhong Wang and Dr. Yan Li for their modifications to the paper's content and English language expressions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Li, T.H.; Song, B. Species and Distributions of Chinese Edible Boletes. Acta Edulis Fungi. 2002, 9, 22-30. [CrossRef]
- Mao, X.L. The macrofungi in China. Henan Sci. Technol. Press. 2000, 1-322.
- Wu, G.; Feng, B.; Xu, J; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Zhu, L.Y. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93-115. [CrossRef]
- Zang, M. Flora fungorum sinicorum. Vol. 22, Boletaceae (I). Sci. Press, Beijing. 2006, 1-215.
- Zang, M. Flora fungorum sinicorum. Vol. 44, Boletaceae (II). Sci. Press, Beijing. 2013, 1-156.
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Cui, Y.Y.; Li, F.; Xu, J.P.; Yang, Z.L. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25-188. [CrossRef]
- Gu, C.D. Key to the Species of Boletus. Korean J. mycology. 1993, 21, 146-156.
- den Bakker, H.C.; Zuccarello, G.C.; Kuyper, T.W.; Noordeloos, M.E. Evolution and host specificity in the ectomycorrhizal genus Leccinum. New Phytol. 2004, 163, 201–215.
- Sato, H.; Yumoto, T.; Murakami, N. Cryptic species and host specificityin the ectomycorrhizal genus Strobilomyces (Strobilomycetaceae). Am. J. Bot. 2007, 94, 1630–1641.
- Smith, M.E.; Pfister, D.H. Tuberculate ectomycorrhizae of angiosperms: The interaction between Boletus rubropunctus (Boletaceae) and Quercus species (Fagaceae) in the United States and Mexico. Am. J. Bot. 2009, 96, 1665–1675.. [CrossRef]
- Husbands, D.R.; Henkel, T.W.; Bonito, G.; Vilgalys, R.; Smith, M.E. New species of Xerocomus (Boletales) from the Guiana Shield, with notes on their mycorrhizal status and fruiting occurrence. Mycologia. 2013, 105, 422–435. [CrossRef]
- Pilát A. Buchwaldoboletus, genus novum Boletacearum. Friesia. 1969, 9, 217−218.
- Ortiz-Santana, B.; Both, E.E. A preliminary survey of the genus Buchwaldoboletus. Bulletin of the Buffalo Society of Natural Sciences, 2011, 40, 1–14.
- Venturella G. Buchwaldoboletus lignicola (Boletaceae), a rare basidiomycete from Europe. Plant Biosyst. 2017, 1-3. [CrossRef]
- Watling, R. New combinations in boletaceae and gomphidiaceae (boletales). Edinb. J. Bot. 2004, 61, 41-47. [CrossRef]
- Jo, J.; Kwag, Y.N.; Cho, S.E.; Han, S.K.; Han, J.G.; Lim, Y.; Sung, G.; Oh, S.H.; Kim, C. First Report of Buchwaldoboletus lignicola (Boletaceae), a Potentially Endangered Basidiomycete Species, in South Korea. Mycobiology. 2019, 47, 521-526. [CrossRef]
- Xie, H.J.; Zhang, C. X.; He, M.X.; Liang, Z.Q.; Deng, X.H.; Zeng N.K. Buchwaldoboletus xylophilus and Phlebopus portentosus, two non-ectomycorrhizal boletes from tropical China. Phytotaxa. 2021, 520, 137–154.
- Pegler, D.N. Agaric flora of Sri Lanka. Kew Bulletin Additional Series. 1986, 12, 1–519.
- Corner, E.J.H. Boletus in Malaysia. Government Printing Office, Singapore. 1972, 263.
- Nanu, S.; Kumar, T.K.A. Buchwaldoboletus xylophilus, a boletoid fungus new to India. Studies in Fungi. 2022, 7, 13.. [CrossRef]
- Bidartondo, M.I.; Ameri, G.; Dring, H. Closing the mycorrhizal DNA sequence gap. Mycol. Res. 2009, 113, 1025-1026. [CrossRef]
- Halling, R.E.; Nuhn, M.; Fechner, N.A.; Osmundson, T.W.; Soytong, K.; Arora, D.; Hibbett, D.S.; Binder, M. Sutorius, a new genus for Boletus eximius. Mycologia. 2012, 104: 951–961. [CrossRef]
- Zhang, M.; Wang, C.Q.; Li, T.H.; Song, B. A new species of Chalciporus (Boletaceae, Boletales) with strongly radially arranged pores. Mycoscience. 2016, 57, 20-25. [CrossRef]
- Chai, H.; Liang, Z.Q.; Xue, R.; Jiang, S.; Luo, S.H.; Wang, Y.; Wu, L.L.; Tang, L.P.; Chen, Y.; Hong, D.; Zeng, N.K. New and noteworthy boletes from subtropical and tropical China. MycoKeys. 2019, 46: 55–96. [CrossRef]
- Binder, M.; Hibbett, D.S. Molecular systematics and biological diversification of Boletales. Mycologia. 2006, 98, 971–981. [CrossRef]
- Baroni, T.J.; Cifuentes, J.; Santana, B.O.; Cappello, S. A new species of Phlebopus (Boletales, Basidiomycota) from Mexico. North American Fungi. 2015, 7, 1–13. [CrossRef]
- Wilson, A.W.; Binder, M.; Hibbett, D.S. Diversity and evolution of ectomycorrhizal host associations in the Sclerodermatineae (Boletales, Basidiomycota). New Phytologist. 2012, 194, 1079–1095. [CrossRef]
- Mei, Y.; Liu, C.Y.; Li, S.H.; Guerin-Laguette, A.; Xiao, Y.J.; Tang, P.; Wan, S.P.; Bonito, G.; Wang, Y. Phlebopus roseus, a new edible bolete from China, is associated with insects and plants. Mycologia. 2020, 113, 33–42. [CrossRef]
- Ji, K.P.; Cao, Y.; Zhang, C.X.; He M.X.; Liu, J.; Wang, W.B.; Wang, Y. Cultivation of Phlebopus portentosus in southern China. Mycol Progress. 2011, 10, 293-300. [CrossRef]
- Cai, X.L.; He, W. Research progress on biological chearacteristics and cultivation of Boletus edulis. Hortic. Seed. 2019, 39, 30-31. [CrossRef]
- Cheng, G.H.; An, X.Y.; Wang, X.; Zhang, B.; Li, Y. Cultural characteristics and domestic cultivation of Polyporus tuberaster. Mycosystema. 2018, 37, 712-721. [CrossRef]
- Gu, D.D.; Shi, L.Y.; Liu, H.X.; Zhang, J.X.; Yao, Q.G.; Wang, L.A. Biological characteristics and cultivation of Clitocybe fragrans. Mycosystema. 2022, 41, 647-657. [CrossRef]
- He, J.; Luo, Z.L.; Deng, R.M.; Su, K.M.; Li S.H.; Su, H.Y. Isolation, biological characteristics and domestication of wild Flammulina rossica and F. filiformis. Mycosystema. 2022, 41, 630-646. [CrossRef]
- Chen, J.Z.; Qin, J.S.; Wang, H.C.; Tan, Y.Z.; Dai, X. Study on the Nutritional Characteristics of Boletus edulis Bull. : Fr Mycelium. Hubei Agricultural Sciences. 2013, 52, 607-609. [CrossRef]
- Kumla, J.; Suwannarach, N.; Lumyong, S. A New Report on Edible Tropical Bolete, Phlebopus spongiosus in Thailand and Its Fruiting Body Formation without the Need for a Host Plant. Mycobiology. 2020, 48, 263-275.. [CrossRef]
- He, Y.L.; Liu, H.L.; Lu, S.; Li, B.; Zhang, X.; Wang, S.M. Isolation and culture of Leccinum aurantiacum from Xinjiang. Edible Fungi. 2010, 32, 17-19.
- Alim, Y.; Sidhoum, W.; Dib, S. First Record of the Edible Mushroom Lepista sordida in Western Algerian Forest: Nutritional Value and Physicochemical Parameters of Mycelial Culture. J. Fungi. 2023, 9, 858. [CrossRef]
- Mortimer, P.E.; Karunarathna, S.C.; Li, Q.H.; Gui, H.; Yang, X.Q.; Yang, X.F.; He, J.; Ye, L.; Guo, J.Y.; Li, H.L.; Sysouphanthong, P.; Zhou D.Q.; Xu, J.C.; Hyde, K.D. Prized edible Asian mushrooms: ecology, conservation and sustainability. Fungal Divers. 2012, 56, 31–47. [CrossRef]
- Liu, Q.M.; Sun, L.P.; Ding, Y.Y.; Zhuang Y.L. Chemical composition, health benefits, food processing effects and applications of Boletus: a review. Crit. Rev. Food Sci. 2023, 1–23. [CrossRef]
- Guerin-Laguette, A. Successes and challenges in the sustainable cultivation of edible mycorrhizal fungi – furthering the dream. Mycoscience. 2021, 62, 10-28. [CrossRef]
- Li, W.; Zi. L.; Xu, N.; Yang, H.; Dong, S.; Qin, F.; Guo, L. Identification of characteristic flavor compounds of boletus edulis from different regions based on by E-nose, HS-GC-IMS and HS-SPME-GC–MS. Food Chem. X. 2024, 23. [CrossRef]
- Branco, S.; Liao, H.L.; Ruytinx, J.; Nguyen, N.; Ke, Y.H.; Colpaert, J.; Vilgalys, R.; Kennedy, P.; Lofgren, L. Suillus as an Emerging Model for Ecology and Evolution of Plant-fungal Mutualisms. Abstract book of the 11th international conference on mycorrhiza. 2022. DOI:10.26914/c.cnkihy.2022.042862.
- Ohta, A.; Fujiwara, N. Fruit-body production of an ectomycorrhizal fungus in genus Boletus in pure culture. Mycoscience. 2003, 44, 295–300. [CrossRef]
- Zang, M. Taxonomy, mycogeography and ectomycorrhizal association with the Boletales from China I. Family Strobilomycetaceae. Mycosystema. 1997, 16, 264–269. [CrossRef]
- Yamada, A.; Furukawa, H.; Yamanaka, T. Cultivation of edible ectomycorrhizal mushrooms in Japan. Rev. Fitotec. Mex. 2017, 40, 379-398.
- Hirose, D.; Shirouzu, T.; Tokumasu, S. Host range and potential distribution of ectomycorrhizal basidiomycete Suillus pictus in Japan. Fungal Ecol. 2010, 3: 255–260. [CrossRef]
- Tan, Q.; You, L.; Hao, C.; Wang, J.; Liu, Y. Effects of four bolete species on ectomycorrhizae formation and development in Pinus thunbergii and Quercus acutissima. BMC Ecol. Evol. 2024, 24, 54. [CrossRef]
- Fang, Y.W.; Wang, W.B.; He, M.X.; Xu X.J.; Gao, F.; Yang, T.W.; Liu, J.; Cao, Y.; Wang, Y.; Zhang, C.X. Investigation on host plants and scale insects species of fungus-insect gall of Phlebopus portentosus in Yunnan,Sichuang,and Guangxi Provinces. J. Environ. Entomol. 2020, 42, 684-694. [CrossRef]
- Zhang, C.X.; Fang, Y.W.; Yang, T.W.; Xu X.J.; Gao, F.; Liu, J.; He, M.X.; Wang, W.B.; Wang, Y. Research progress on nutritional relationship of Phlebopus portentosus, root mealybug and plant. Edible Med. Mushrooms. 2023, 31, 97-101.
- Yuwa-amornpitak T.; Butkhup, L.; Yeunyaw, P.N.; Chookietwattana, K. Cultivation of Edible Ectomycorrhizal Mushrooms (Phelobopus portentosus) Associated with Sesbania javanica Miq. Jordan J. Biol. Sci. 2023, 16, 125-130. [CrossRef]
- Zhang, C.X.; He, M.X.; Cao, Y.; Liu, J.; Gao, F.; Wang, W.B.; Ji, K.P.; Shao, S.C.; Wang, Y. Fungus-insect gall of Phlebopus portentosus. Mycologia. 2015, 107, 12-20. [CrossRef]
- Zhang, C.X.; He, M.X.; Liu, J.; Xu, X.J.; Cao, Y.; Gao, F.; Fang, Y.W.; Wang, W.B.; Wang, Y. Brief introduction to a Unique edible bolete Phlebopus portentosus in southern China. J. of Agri. Sci. Technol. B. 2017, 7, 386-394. [CrossRef]
- Fang, Y.W.; Wang, W.B.; He, M.X.; Xu X.J.; Gao, F.; Liu, J.; Yang, T.W.; Cao, Y.; Yang T.; Wang, Y.; Zhang, C.X. Relationship Between the Honeydew of Mealy Bugs and the Growth of Phlebopus Portentosus. PLoS ONE. 2020, 15, e0233710. [CrossRef]
- Cao, Y.; Ji, G.Y.; Luo, S.Z.; Gao, L.X.; Tao, L.; Wang, Q.L.; Yang, R.H.; Bao, D.P.; Ji, K.P. Domestication and artificially cultivation of Phlebopus portentosus: retrospect and prospect. Mycosystema. 2021, 40, 3064-3080. [CrossRef]
- Kumla, J.; Suwannarach, N.; Lumyong, S. Cultivation of Edible Tropical Bolete, Phlebopus spongiosus, in Thailand and Yield Improvement by High-Voltage Pulsed Stimulation. Agronomy. 2022, 12, 115. [CrossRef]
- Cao, Y.; Zhang, Y.; Yu, Z.F.; Wang, P.F.; Tang, X.Z.; He, XX.; Mi, F.; Liu C.L.; Yang, D.; Xu, J.P. Genome Sequence of Phlebopus portentosus Strain PP33, a Cultivated Bolete. Genome Announc. 2015, 3, e00326-15. [CrossRef]
- Zhang, C.X.; Wang, Y.; He, M.X.; Liu, J.; Cao, Y.; Gao, F.; Yang, T.W.; Wang, W.B.; Xu, X.J.; Fang, Y.W.; Li, G.H.; Ni, S.B.; Chen, K.N.; Wu, Y.; Xu, T. Amazing delicious black boletus Phlebopus portentosus. Yunnan Sci.Tech. Press. 2018, 1-140.
- He, R.; Liu, S.X.; Liu, C.L.; Zhang, W.S. The Research Progress of Boletus edulis. Food Ferment. Sci. Tech. 2016, 52, 75-77. [CrossRef]
- Tang, C.; Chen, Y.L.; Liu, R.J. Research progress of edible mycorrhizal fungi. Mycosystema. 2011, 30, 367-378. [CrossRef]
- Yang, T.W.; Liu, J.; Xu, X.J.; He, M.X.; Gao, F.; Fang, Y.W.; Wang, W.B.; Dai, L.M.; Wang, Y.; Zhang, C.X. Sclerotia formation of Phlebopus portentosus under natural and artificial conditions. Forests. 2023, 14, 1096. [CrossRef]
- Kumla, J.; Danell, E.; Lumyong, S. Improvement of yield for a tropical black bolete, Phlebopus portentosus, cultivation in northern Thailand. Mycoscience. 2015, 114-117. [CrossRef]
- Costa, A.F.P.; Steffen, G.P.K.; Steffen, R.B.; Portela, V.O.; Santana, N.A.; Richards, N.S.P.S.; Jacques, R.J.S. The use of rice husk in the substrate composition increases Pleurotus ostreatus mushroom production and quality. Sci. Hortic. 2023, 321, 112372.. [CrossRef]
- Caiafa, M.V.; Smith, M.E. Polyphyly, asexual reproduction and dual trophic mode in Buchwaldoboletus. Fungal Ecol. 2022, 56, 101141. [CrossRef]
Figure 1.
The fruiting bodies of wild B. xylophilus.

Figure 2.
The strain of B. xylophilus. A: slant cultures, B: petri dishes cultures.
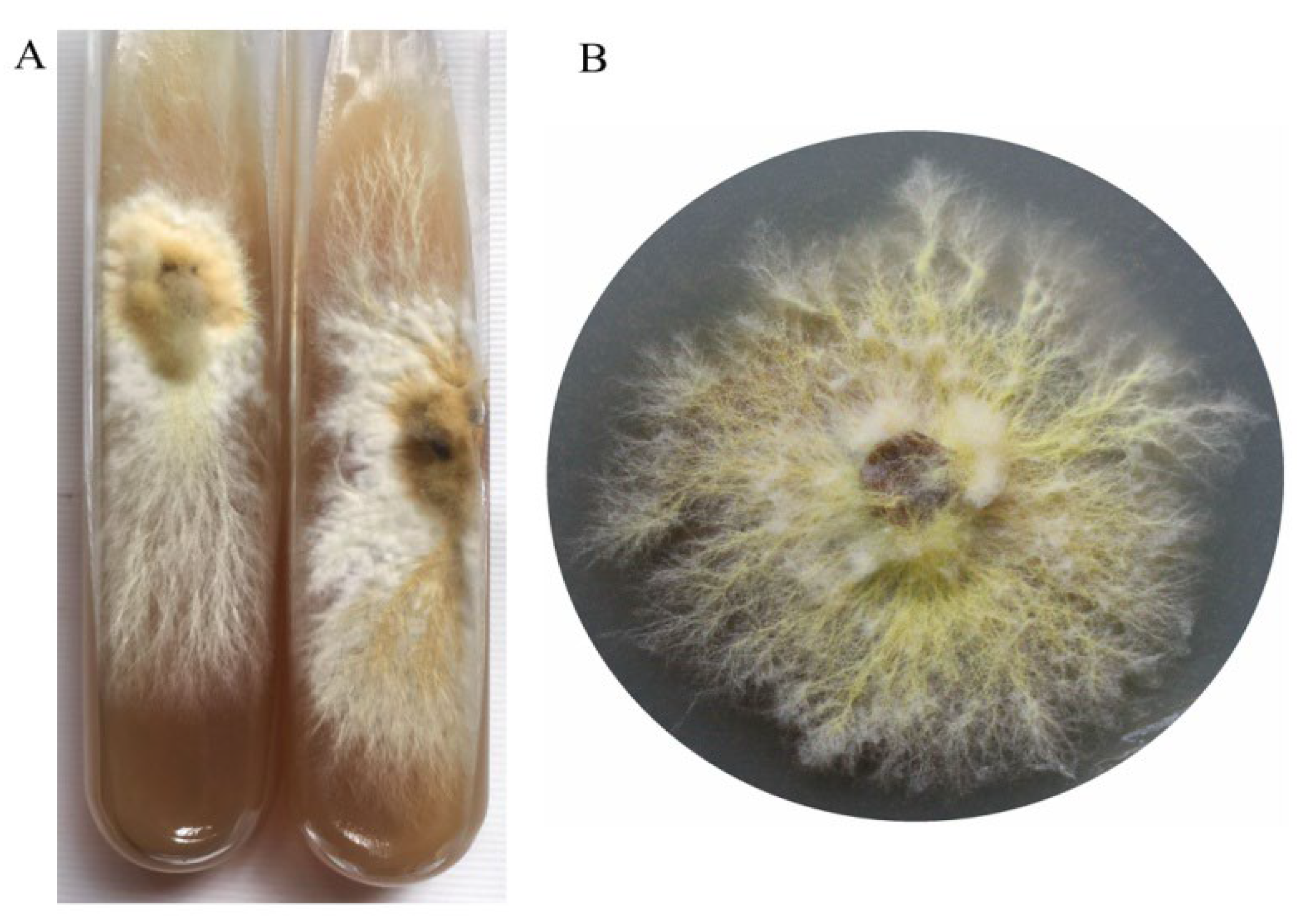
Figure 3.
The growth mycelium of B. xylophilus on the cultivation substrate.

Figure 4.
Growth process of fruiting bodies of B. xylophilus. A: Mycelia grows to the surface of covering soil layer,B: primordium, C~F: young mushroom, D~G: Changes of fruiting body growth at intervals of 24 h,G~H: mature fruiting bodies.
Figure 4.
Growth process of fruiting bodies of B. xylophilus. A: Mycelia grows to the surface of covering soil layer,B: primordium, C~F: young mushroom, D~G: Changes of fruiting body growth at intervals of 24 h,G~H: mature fruiting bodies.

Figure 5.
Morphology of fruiting bodies of artificially cultivated B. xylophilus. A~B: growing young mushrooms, C~D: mature fruiting bodies,E: longitudinal section of fruiting body.
Figure 5.
Morphology of fruiting bodies of artificially cultivated B. xylophilus. A~B: growing young mushrooms, C~D: mature fruiting bodies,E: longitudinal section of fruiting body.

Figure 6.
Microstructure of the mycelium and basidiospores of B. xylophilus. A: the mycelium of B. xylophilus, without a clamp connection,B: Basidiospores.
Figure 6.
Microstructure of the mycelium and basidiospores of B. xylophilus. A: the mycelium of B. xylophilus, without a clamp connection,B: Basidiospores.
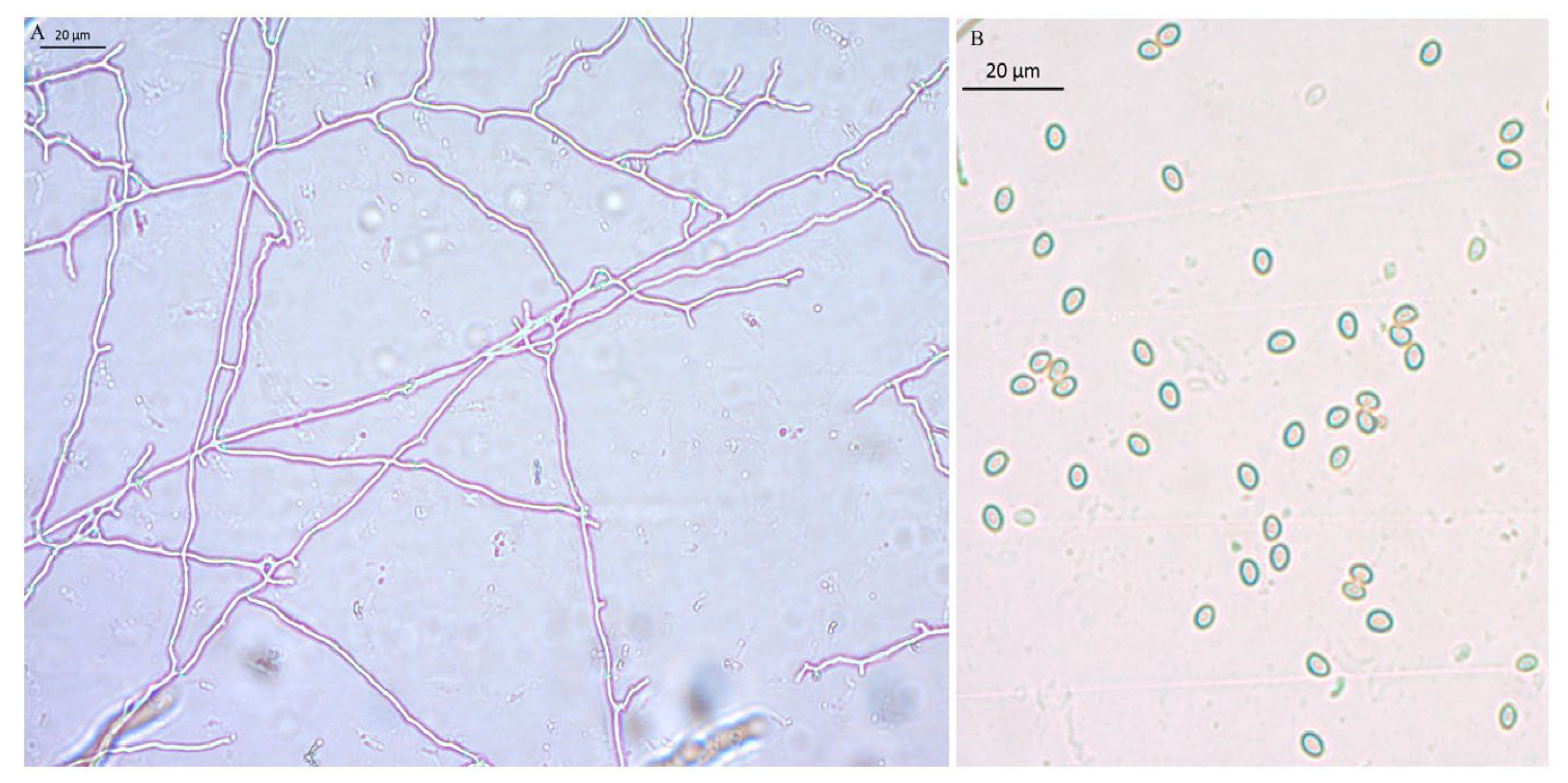
Figure 7.
Phylogenetic analysis based on 28S gene sequence.

Figure 8.
Phylogenetic tree constructed by ITS genes.
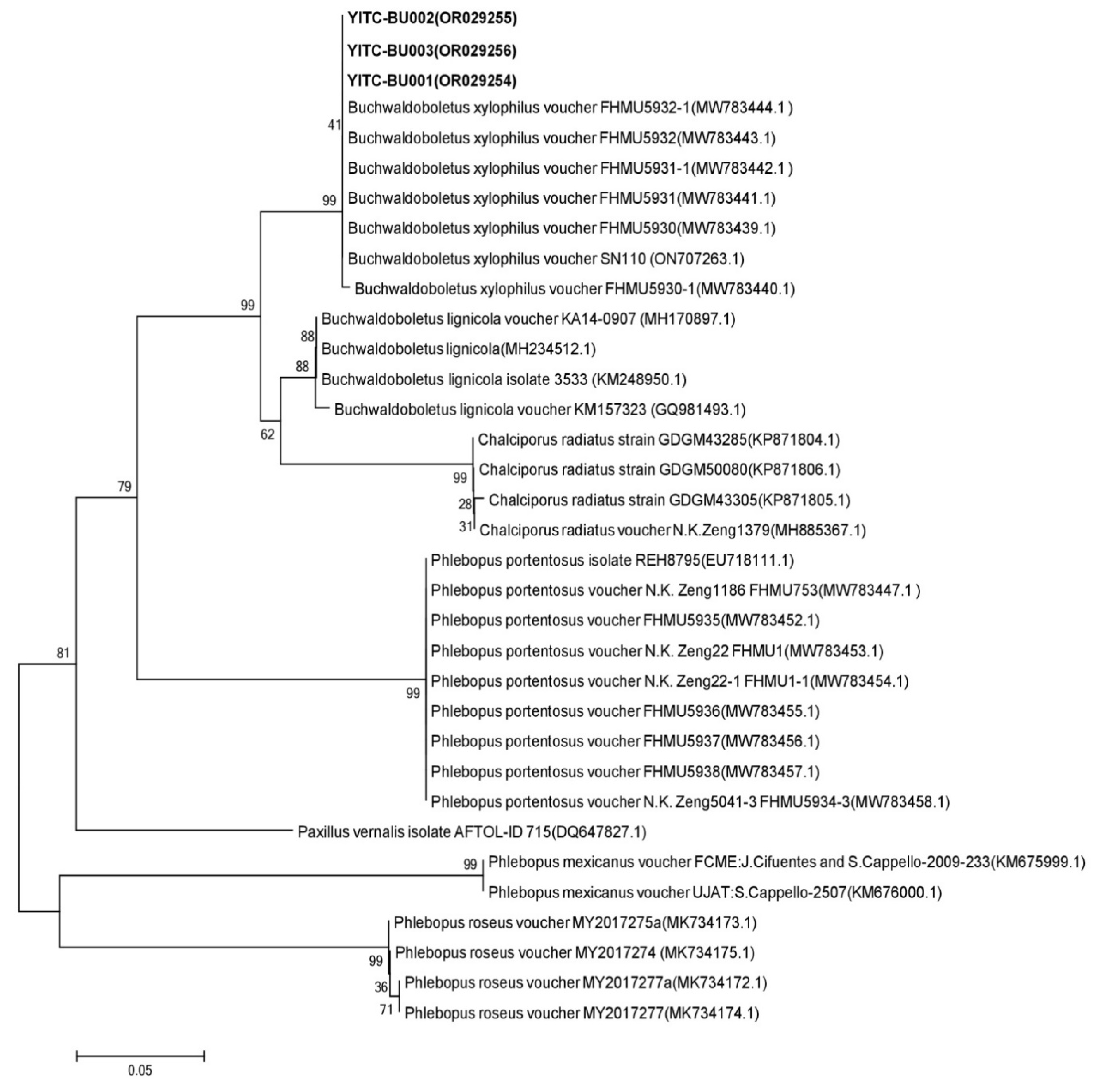
Table 1.
Effect of Different Carbon Sources on the Mycelial Growth of B. xylophilus.
| Carbon sources | Mycelial growth rate(mm/d) | Mycelial growth vigor |
| Glucose | 1.63±0.07 | +++ |
| Fructose | 1.58±0.06 | ++++ |
| Sucrose | 0.64±0.14 | ++ |
| Mannitol | 1.53±0.09 | +++ |
| Maltose | 1.70±0.07 | ++++ |
| Soluble starch | 0.91±0.09 | + |
Note: “+” means the mycelium is sparse and grows less, “++” means the mycelium is sparse and the growth is poor, “+++” means the mycelium is dense and thick, “++++” means the mycelium is thick and robust, and grow well. The same below.
Table 2.
Effect of Different nitrogen Sources on the Mycelial Growth of B. xylophilus.
| Nitrogen Sources | Mycelial growth rate(mm/d) | Mycelial growth vigor |
| Tryptone | 1.46±0.05 | +++ |
| Beef extract | 1.41±0.09 | +++ |
| Yeast extract | 1.44±0.09 | +++ |
| Ammonium Tartrate | 1.77±0.04 | ++++ |
| KNO3 | 1.27±0.10 | ++ |
| NH4NO3 | 1.58±0.13 | ++++ |
Table 3.
Effect of Different inorganic salt on the Mycelial Growth of B. xylophilus.
| Inorganic Salt | Mycelial growth rate(mm/d) | Mycelial growth vigor |
| NaCl | 1.42±0.06 | +++ |
| MgSO4 | 1.49±0.06 | +++ |
| KH2PO4 | 1.43±0.13 | +++ |
| MnSO4 | 0.95±0.09 | ++ |
| CaSO4 | 1.42±0.08 | +++ |
| NaNO3 | 1.27±0.08 | ++ |
Table 6.
Orthogonal experiment on mycelial growth of B. xylophilus.
| Number | Carbon sources | Nitrogen Sources | Inorganic Salt | Temperature | Mycelial growth rate(mm/d) | Mycelial growth vigor |
| 1 | 1(maltose) | 1(NH4NO3) | 1(MgSO4) | 1(25℃) | 1.02±0.14 | ++ |
| 2 | 1(maltose) | 2(ammonium tartrate) | 2(KH2PO4) | 2(28℃) | 1.76±0.08 | ++++ |
| 3 | 1(maltose) | 3(tryptone) | 3(CaSO4) | 3(30℃) | 1.29±0.11 | ++ |
| 4 | 2(glucose) | 1(NH4NO3) | 2(KH2PO4) | 3(30℃) | 1.73±0.11 | +++ |
| 5 | 2(glucose) | 2(ammonium tartrate) | 3(CaSO4) | 1(25℃) | 1.13±0.05 | ++ |
| 6 | 2(glucose) | 3(tryptone) | 1(MgSO4) | 2(28℃) | 1.48±0.19 | +++ |
| 7 | 3(fructose) | 1(NH4NO3) | 3(CaSO4) | 2(28℃) | 1.81±0.07 | ++++ |
| 8 | 3(fructose) | 2(ammonium tartrate) | 1(MgSO4) | 3(30℃) | 1.62±0.12 | +++ |
| 9 | 3(fructose) | 3(tryptone) | 2(KH2PO4) | 1(25℃) | 0.87±0.13 | ++ |
| K1 | 4.07 | 4.56 | 4.12 | 3.02 | ||
| K2 | 4.34 | 4.51 | 4.36 | 5.05 | ||
| K3 | 4.30 | 3.64 | 4.23 | 4.64 | ||
| X1 | 1.36 | 1.52 | 1.37 | 1.01 | ||
| X2 | 1.45 | 1.50 | 1.45 | 1.68 | ||
| X3 | 1.43 | 1.21 | 1.41 | 1.55 | ||
| R | 0.09 | 0.31 | 0.08 | 0.67 |
Note: Kn: the sum of mycelial growth rates at the n level. Xn: the mean mycelial growth rate at the n level. R: Range.
Table 7.
The average fresh mushroom yield and biological efficiency of B. xylophilus.
| No. | Time of primordial formation (d)* | Primordial number (per bag) | Number of young mushrooms (per bag)* | Number of mature fruiting bodies (per bag)* | Fresh mushroom yield (g/bag)* | Biological efficiency (%)* |
| 1 | 11.28±1.50 | densely packed | 5.70±3.69 | 4.13±2.21 | 131.07±29.38 | 28.48±6.39 |
| 9~14 | 1~19 | 1~11 | 73.25~186.43 | 15.92~40.53 |
Note: “*” Results are means ± SD and scope of 40 replicates.
Table 8.
Sequences used in the analysis.
| Taxon | Voucher | Locality | GenBank accession Nos. | References | |
| 28S | ITS | ||||
| Buchwaldoboletus lignicola | HKAS76674 | Heilongjiang, China | KF112350 | — | [6] |
| Buchwaldoboletus lignicola | HKAS84904 | Germany | KT990538 | — | |
| Buchwaldoboletus lignicola | KA14-0907 | South Korea | — | MH170897 | [16] |
| Buchwaldoboletus lignicola | 3533 | — | — | KM248950 | Unpublished |
| Buchwaldoboletus lignicola | — | — | — | MH234512 | Unpublished |
| Buchwaldoboletus lignicola | KM157323 | — | — | GQ981493 | [21] |
| Buchwaldoboletus lignicola | Pul1 | Germany | JQ326997 | [22] | |
| Buchwaldoboletus xylophilus | FHMU5930 | Yunnan, SW China | MW783417 | MW783439 | [17] |
| Buchwaldoboletus xylophilus | FHMU5930-1 | Yunnan, SW China | MW783418 | MW783440 | |
| Buchwaldoboletus xylophilus | FHMU5931 | Yunnan, SW China | MW783419 | MW783441 | |
| Buchwaldoboletus xylophilus | FHMU5931-1 | Yunnan, SW China | MW783420 | MW783442 | |
| Buchwaldoboletus xylophilus | FHMU5932 | Yunnan, SW China | MW783421 | MW783443 | |
| Phlebopus portentosus | N.K.Zeng22 (FHMU1) | Hainan, southern China | MW783433 | MW783453 | |
| Phlebopus portentosus | N.K.Zeng22-1 (FHMU1-1) | Hainan, southern China | MW783434 | MW783454 | |
| Phlebopus portentosus | N.K.Zeng1186 (FHMU753) | Hainan, southern China | MW783427 | MW783447 | |
| Phlebopus portentosus | N.K.Zeng5041-3 (FHMU5934-3) | Hainan, southern China | MW783438 | MW783458 | |
| Phlebopus portentosus | FHMU5935 | Yunnan, SW China | MW783432 | MW783452 | |
| Phlebopus portentosus | FHMU5936 | Yunnan, SW China | MW783435 | MW783455 | |
| Phlebopus portentosus | FHMU5937 | Yunnan, SW China | MW783436 | MW783456 | |
| Phlebopus portentosus | FHMU5938 | Yunnan, SW China | MW783437 | MW783457 | |
| Buchwaldoboletus xylophilus | FHMU5932-1 | Yunnan, SW China | MW783422 | MW783444 | |
| Buchwaldoboletus xylophilus | — | India | — | ON707263 | [20] |
| Buchwaldoboletus xylophilus | YITC-BU001 | Yunnan, China (Artificial cultivation) | OR030332 | OR029254 | this study |
| Buchwaldoboletus xylophilus | YITC-BU002 | Yunnan, China (Artificial cultivation) | OR030333 | OR029255 | this study |
| Buchwaldoboletus xylophilus | YITC-BU003 | Yunnan, China (Artificial cultivation) | OR030334 | OR029256 | this study |
| Chalciporus radiatus | GDGM43285 | Hunan, Central China | KP871800 | KP871804 | [23] |
| Chalciporus radiatus | GDGM50080 | Hunan, Central China | KP871801 | KP871806 | |
| Chalciporus radiatus | GDGM43305 | Guangdong, China | KP871802 | KP871805 | |
| Chalciporus radiatus | N.K.Zeng1379 (FHMU930) | Fujian, SE China | MH879710 | MH885367 | [24] |
| Paxillus vernalis | AFTOL-ID 715 | — | AY645059 | DQ647827 | [25] |
| Phlebopus mexicanus | FCME:J.Cifuentes and S.Cappello-2009-233 | Mexico | KM676001 | KM675999 | [26] |
| Phlebopus mexicanus | UJAT:S.Cappello-2507 | — | KM676002 | KM676000 | |
| Phlebopus portentosus | REH8795 | Thailand | FJ153623 | EU718111 | [27] |
| Phlebopus roseus | MY2017274 | Sichuan, China | MK734306 | MK734175 | [28] |
| Phlebopus roseus | MY2017277_a | Sichuan, China | MK734305 | MK734174 | |
| Phlebopus roseus | MY2017275_a | Sichuan, China | MK734304 | MK734173 | |
| Phlebopus roseus | MY2017277 | Sichuan, China | MK734303 | MK734172 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



