
Submitted:
30 January 2025
Posted:
30 January 2025
You are already at the latest version
Abstract
Background: Escherichia albertii is a lactose-negative escherichia that causes gastritis and enteritis in humans. Analysis of possible sources of infection points out that poultry may be a significant reservoir for this pathogen. The question of whether E. albertii can cause infections in poultry is still unanswered. Our article describes the isolation of E. albertii, for the first time in Russia, from the intestines of birds on a quail farm and characterization of obtained cultures. Methods: We isolated different bacteria from pathological poultry material using bacteriological methods and ruled out them as probable causes for enteritis. Biochemical identification of E. albertii and antibiotic sensitivity were performed using a Vitek-2 Compact instrument. Bacterial identification was done using the MALDI-TOF Biotyper instrument (Bruker, Germany). E. albertii-specific genes, virulence factor genes and microcin genes were determined real-time PCR. Results: Concluded that E. albertii isolated from sites of intestinal inflammation are a potential cause of enteritis and high poultry mortality – up to 15% of total livestock for 10- to 20-days quails. One of the E. albertii culture differed from the main group of Escherichia by its biochemical properties, and subsequent PCR analysis showed a lack of the intimin gene (eae). However, we have to perform model experiments, which we will conduct later, to prove that the bird is not just a reservoir for the pathogen but is also sick itself. Conclusions: We describing the first occasion of infection caused by E. albertii in industrial quails. During the study it was found that according to the molecular-genetic and phenotypic properties of isolated strains in quails there were at least 2 clonal groups of E. albertii differing in antibiotic resistance, biochemical indices and presence of the eae (intimin) gene.
Keywords:
Escherichia albertii
; eae genes
; bird infections
; APEC
; antibiotic resistance
1. Introduction
Escherichia albertii is a Gram-negative, immobile, nonspore-forming, facultatively anaerobic, lactose-negative bacillus belonging to the family Enterobacteriaceae [1]. E. albertii was first isolated in diarrhea cases in children from Bangladesh, tentatively misidentified as Hafnia alvei, and later classified as a new Escherichia species in 2003. [2].
Often E. albertii is misidentified as enteropathogenic E. coli (EPEC) or enterohemorrhagic E. coli (EHEC) because of its genetic and phenotypic similarities to these pathogens. For example, E. albertii usually carries the eae gene encoding intimin, an important virulence factor also shared by pathogenic subgroups of E. coli [3]. This result probably led to an erroneous underestimation of the number of infections caused by E. albertii. For example, in several outbreaks of gastroenteritis, the causative agent was misdiagnosed as EPEC instead of eae-positive E. albertii [4].
The lack of a clear differential diagnosis remains a major problem, as there is no simple and accurate diagnos-tic protocol, especially for PCR typing, such as multilocus sequence typing (MLST) and O-genotyping. [5].
To date, three biogroup of E. albertii have been distinguished according to their phenotypic features: biogroup 1 – indole-negative, lysine-positive; biogroup 2 – indole-positive, lysine-negative; biogroup 3 – indole-positive, lysine-positive. This method may be useful for the identification of E. albertii in diagnostic laboratories or in the context of phylogeny [6].
Strains of E. albertii have been isolated from various animal sources, such as poultry, pigs, cats, dogs, bats, and raccoons, and raw meat of animal origin [7], but its natural reservoirs and routes of transmission to humans remain uncertain.
Nevertheless, an analysis of possible sources indicates that birds are the most significant reservoir for this pathogen [1]. Thus, the isolation of E. albertii cultures from migratory birds of the Asian-Australasian flyway (EAAF), which affects the territory of Russia, has been reported [8]. The genetic diversity of E. albertii isolates from migratory birds was demonstrated, and some isolates can potentially cause disease in humans. A large-scale study conducted on 9 farms in Mississippi and Alabama showed that of 270 cloacal swabs (30 per farm) tested by PCR diagnostics, 43 were positive for the presence of E. albertii, and 12 strains of E. albertii were isolated with varying degrees of isolation on individual farms from 0% to 23.3% [9]. Phylogenetically, isolates from different farms were distant but clonally similar from the same farm, including those with antibiotic and plasmid resistance. Importantly, 9 out of 12 E. albertii strains exhibited multiple drug resistance; one strain was even resistant to imipenem, a clinically important carbapenem antibiotic. In addition, comparative genomics analysis showed that two clusters of E. albertii strains isolated from chickens exhibited very close evolutionary relationships and similar virulence gene profiles to E. albertii strains isolated from humans.
A study in Switzerland on the detection of E. albertii in wild birds showed that out of 280 fecal samples (collected from 26 bird species), E. albertii isolates were obtained from 11 bird species belonging to eight families. Four of the eleven species were waterfowl. The other 7 species, including raptors and raven, often inhabit agricultural lands [10]. In the same study, pooled fecal samples from 150 broiler flocks (more than one million birds) did not reveal the presence of E. albertii in any of the samples.
It should be noted that the review studies cited were aimed at detecting E. albertii carriage, but not at detecting infection caused by this bacterium. Whether E. albertii can cause infection in poultry remains an open question.
Our paper describes the isolation of E. albertii isolates, for the first time in the Russian Federation, from the intestines of birds of a quail farm and the phenotypic and molecular genetic characterization of the obtained cultures. In addition, we show that E. albertii is the cause of lethal infection in birds. The listed provisions were the objective of our work.
2. Materials and Methods
2.1. Object of the Study
Live quails came from a quail farm in the Moscow region with 25 animals. Anamnesis indicated a significant loss of livestock (up to 15% of the total livestock) at the age of 10 to 20 days of life with signs of gastrointestinal tract damage. Keeping birds on the farm is cage-based. Crosses of three lines – Japanese, Pharaoh, and Manchurian gold. Quails were killed under chloroform anesthesia, after which they were isolated using a nutrient me-dia.
2.2. Nutrient Media and Seeding
Using dactyloscopic smears (see below), quail parenchymatous organs were seeded on nutrient agar #1 (Nutrient Media, Obolensk), Endo agar (Nutrient Media, Obolensk), BHI agar (brain-heart infusion, HiMedia M-211, India) and addition of 10 μg/ml NADH (A1393.0001, AppliChem, Spain) 5% (w/ml), fresh yeast extract, 1% (wt/volume) glucose (A0883.0500, AppliChem, Spain) and 0.001% (wt/volume) L cysteine (LLC NPP "PanEco"). To obtain parenchymatous smears, parenchymatous organs were briefly cauterized with ethanol, the organ was excised, and the cut surface was pressed against agar several times. To obtain scrapings from the suborbital sinuses, the upper part of the beak was excised with sterile scissors and scraped with a 2-μl loop. The loop content was seeded on Petri dishes containing dense agar. The trachea and oviducts were scraped with loops and the loop contents were seeded onto agar dishes. The intestinal contents were sown with a 10 µl loop on the agar surface and further spread on the surface of the medium with new loops using the Drigalsky method to isolate colonies. Clostridium perfringens agar (HiMedia, India) supplemented with nutrients was used to isolate clostridia. The seeded Petri dishes were placed in an anaerostat (Schuett-biotec GmbH D-37079, Göttingen, Germany) and cultured under anaerobic con-ditions using anaerobic bags cultured at 42 °C for 24 h.
Cups of crops were also duplicated. Part of the dishes was placed in an anaerostat under microaerophilic conditions (with a candle) at 37 °C. The other part was cultured under anaerobic conditions, using anaerobic bags, at 42 °C for 24 h.
Culture typing. The grown colonies were evaluated culturally and morphologically and then analyzed by mass spectrometry.
2.3. Mass Spectrometric Analysis
Sample preparation was performed as previously described [11]. Protein extraction was performed according to the protocol proposed by Bruker (Daltonics GmbH, Germany). The results were summarized using the FlexAnalysis software (Bruker Daltonics GmbH, Bremen, Germany).
2.4. PCR Analysis of Escherichia coli
DNA was isolated from bacterial culture grown on nutrient agar dishes (HiMedia, Mumbai, India) using simple cell lysis. The PCR-mix for 20 μL reactions contained 0.4 mM dNTP (Thermo Fisher Scientific, USA), 2.5 μL DreemTaq Green Buffer 10x (Thermo Fisher Scientific, USA), 10 mM of each primer, 0.1 units/μL DreemTaq DNA polymerase (Thermo Fisher Scientific, USA). The amplification products were analyzed using the ChemiDoc XRS+ system (Bio-Rad, USA).
E. albertii strains were identified by searching for E. albertii-specific genes such as mdh, lysP, and clpX (Table 1). We amplified the malate dehydrogenase gene (mdh), lysine-specific transporter gene (lysP) and heat shock protein gene (clpX) using multiplex PCR (Table 1) [12].
In addition, we used real-time PCR differentiation of E. coli pathogroups (EPEC, ETEC, EIEC, EHEC, EAgEC) with the AmpliSens® Escherichiose-FL kit (AmpliSens, Moscow, Russia) for isolated E. albertii and E. coli cultures.
Identification of the intimin gene – eae (common between E. albertii and EPEC) in E. albertii cultures was performed according to the European Union reference laboratory methodology for E. coli "Identification and characterization of verocytotoxin-producing Escherichia coli (VTEC) by real-time PCR amplification of major virulence genes and genes associated with serogroup mainly associated with severe human infections" [13].
2.5. CDT Typing
Cytolethal cell distension toxin (CDT) and cell cycle inhibitory factor (cif), types of cyclomodulins of pathogenic Escherichia coli that are not associated with any specific phylogroup, were investigated using specific primers (Table 1) [14].
Virulence factor genes of APEC (avian pathogenic Escherichia coli) were investigated according to the protocols proposed in the articles (Table 1) [15,16].
Microcin genes were determined according to the conditions proposed in [17].
2.6. Serotyping of Escherichia coli
The serotyping of Escherichia coli was performed according to the standard protocol recommended by the manufacturer (JSC "Biomed" named after. I.I. Mechnikov, Russian Federation).
2.7. Biochemical Identification and Susceptibility to Antimicrobials
Biochemical identification of E. albertii and antibiotic sensitivity were performed using GN cards on a Vitek-2 Compact instrument.
In addition, antibiotic sensitivity was studied by the disk-diffusion method using NICF disks (St. Petersburg), with the results recorded automatically on a Scan 500 device (Interscience).
3. Results
3.1. Characteristics of E. albertii and Other Microorganisms
The study aim was to determine the causes of increased mortality of quails 10-20 days old with signs of gastrointestinal disorders and diarrhea. The mortality rate, depending on the workshop, ranged from 8% to 20%. Antibiotic therapy with a complex preparation including enrofloxacin and colistin before laboratory testing showed no obvious improvement in quail health.
According to the results of the study, most quails carried Salmonella enterica variant bredeney, which is the causative agent of avian salmonellosis. Salmonella cultures were not isolated from the gastrointestinal tract. There was respiratory salmonellosis, which was localized mainly in the trachea and suborbital sinuses. The identity of Salmonella was confirmed using PCR and agglutinating serum. Cultures of Streptococcus pluranimalium causing avian streptococcosis, Bordetella hinzii and Gallibacterium anatis, the causative agents of pasteurell-like infection in birds, were isolated from the same localization.
At the same time, viral infections such as avian influenza, infectious bronchitis, and Newcastle virus were excluded by PCR analysis. In isolated cultures of Clostridium perfringens, PCR analysis did not detect the NetB toxin, which has a leading role in the development of necrotizing enteritis, and the cultures were classified as toxotype A according to the table proposed by Van Immersee [16].
E. coli cultures isolated from the intestine were not assigned to any of the Escherichia pathogroups (EPEC, ETEC, EIEC, EHEC, EAgEC) by real-time PCR-RT, and only 4 isolated cultures had genes that could be assigned to the APEC pathogroup. 75% of cultures of this pathogroup were isolated from the tracheas.
The intestine was found to have the following lesions: greenish feces, injected intestinal and mesenteric vessels, wall thickening, necrotic, hemorrhagic and ulcerated areas (Figure 1). Cultures were isolated from poultry with these features in one of the workshops and classified them as E. albertii during the study. The results of the bacteriological examination are summarized in Table 2.
3.2. Identification of E. albertii
The cultures isolated on Endo agar were lactose-negative and were initially considered to be E. coli/Salmonella spp. because no significant cultural and morphological differences from colonies of E. coli strains were detected and the bird was a carrier of Salmonella spp. The mass spectral analysis did not provide an unambiguous interpretation, with close ID values for both E. albertii and E. coli.
Vitek2 analysis using GN maps showed that all cultures were E. coli with a probability of more than 90%. At the same time, it was determined that, conditionally, they are divided into two groups according to the atypical features of E. coli. Uncharacteristic for E. coli features of the first group: AGAL (Alpha-galactosidase), BGAL (Beta-galactosidase), BGUR (Beta-glucuronidase) – most cultures, for the second group – AGAL, BGAL, BGUR, PHOS (Phosphotase). Further characterization of antibiotic resistance using Vitek2 device and disk diffusion method showed that they differed in antibiotic sensitivity as well (Table 3, Figure 2). Thus, culture-genetic differences were observed in strains of poultry from the same housing.
It should be noted the discrepancies in the readings of the two methods for cefepime and tigecycline, for the other antibiotics the readings were the same. The difference between the two clonal groups E. albertii was in their relationship to the antibiotics ampicillin and amoxicillin/clavulonic acid – in the first group the cultures were sensitive to them, whereas in the second group they were resistant. Otherwise, the antibiotic resistance profile coincided, and cultures were resistant to fluoroquinolones (ciprofloxacin, norfloxacin), polymyxins (colistin), aminoglycosides (amikacin, gentamicin, tigecycline), and fosfomycin. Given the widespread use of fluoroquinolones and colistin in poultry farming (including on the livestock of this farm), the resistance of cultures to these groups is natural.
3.3. Results of PCR Analysis
The AmpliSens® Escherichiosis-FL PCR-RT kit showed that all isolated cultures of E. albertii belonged to the EPEC (enteropathogenic Escherichiae) pathogroup and contained the eae gene (encoding intimin), except for one culture, which we designated as group 2, because it also differed from the others in biochemical parameters and antibiotic resistance.
According to the multiplex PCR data for the presence of E. albertii-specific genes mdh, lysP and clpX, all tested cultures had all three genes and thus it was confirmed that they belonged to the species E. albertii.
O-serotyping of cultures. All tested E. albertii cultures reacted simultaneously with sera for O18 and O101.
CDT- and cif- typing. According to the results, both strains contained cdtB-II, cdtB-III, cdtB-V, cdtB-I, cdtB-IV genes. The presence of cdtB-II, cdtB-III, cdtB-V genes was confirmed by positive results for all three primer pairs – CDT-s1+CDT-as1, CDT-s1+CDT-IIas, CDT-s1+CDT-IIIas (Table 1). The presence of the cdtB-I, cdtB-IV genes was con-firmed by positive results only using the primer pair CDT-s2+CDT-as2 multiplex (cdtB-I, cdtB-IV). Specific primers for these types separately did not provide results. The presence of cdtB-I, cdtB-IV genes homologous to E. coli are doubt-ful and should be verified by full-genome sequencing. The cdtC-V and cif genes were not detected.
In addition, the obtained E. albertii cultures were tested for the presence of genes specific to pathogenic APEC and microcin genes (Table 1).
According to the results of the study, genes of the APEC pathogenicity group, such as the fimbriae gene (fim) and episomal serum survival enhancement gene (iss) were detected in E. albertii strains of both groups. The microcin M4 and microcin B17 genes were also identified, which apparently increased the competitive advantage of E. albertii among closely related species.
4. Discussion
We examined quails from an industrial farm for bacterial infections to determine the causes of mass mortality based on signs of gastrointestinal tract damage. A significant proportion of quails (48% of the total number of quails examined) were carriers of salmonellosis caused by Salmonella enterica serovariant bredeney (serovar O:4 H:Lv 1,7). However, the localization of all isolated Salmonella cultures in the upper respiratory tract indicates that they are not the cause of gastrointestinal tract lesions in poultry. For industrial poultry, the respiratory tract is often the "entry gate" for Salmonella. Moreover, it is possible that the respiratory route of Salmonella infection in birds is preferable over the fecal-oral route [19,20].
Although in the above studies, in experimental infections with Salmonella by intratracheal or aerosolized methods, during the course of time, Salmonella colonization also occurred in the cecum, but not in organs (except lungs). In our case, we observed the only respiratory carriage of Salmonellae. Salmonella bredeney serovariant itself is not described as a causative agent of salmonellosis infection in birds, whereas consumption of poultry meat contaminated with this serovariant can cause gastroenteritis outbreaks in humans [21]. Thus, we excluded Salmonella as a possible cause of poultry mortality in this case.
Other isolated bacteria of importance in the poultry industry include Gallibacterium anatis. This bacterium is widely spread in poultry at industrial poultry farms in the Russian Federation [22,23]. However, the cultures G. anatis of isolated by us, although they have the RTX-toxin gene (GtxA) – the key virulence factor, are not hemolytic (do not belong to the haemolytica biovar), and it is known that hemolytic variants of G. anatis are capable of causing enteritis [24]. Therefore, G. anatis was not an etiologic agent of GI lesions in our case.
Another possible cause of enteritis is E. coli. However, this study determined that the isolated E. coli strains did not belong to any of the pathogroups leading to GI lesions. Avirulent E. coli strains and APEC pathogens were detected. It is known that APEC are extraintestinal E. coli with a leading intratracheal route of infection [25]. This was confirmed in our studies, as three of 4 cultures of the APEC pathogroup were isolated from the trachea (Table 2).
Thus, we hypothesized that E. albertii is a possible pathogen responsible for GI lesions in quail. Many cases of human gastroenteritis caused by E. albertii have been described, but information on avian diseases caused by this pathogen is scarce. For example, the authors [26], describe the death of wild birds of the Vesjurk family (Carduelis flammea) caused by E. albertii. According to their data, the cause of death was more than 8000 birds (out of those identified) in Alaska.
Previously, in the late 1990s, E. albertii was determined to be the cause of wild bird mortality in Scotland, but the authors incorrectly identified it as E. coli O86:K61[27].
We herein describe the first case of infection caused by E. albertii in industrial quail and industrial poultry in general. In the process of typing E. albertii, the biochemical properties of the cultures were determined. In general, the results showed that the differences in the biochemical properties of E. coli/E. albertii were characteristic of previously described E. albertii cultures [26]. At the same time, one isolated E. albertii culture exhibited a slight difference compared with the other cultures. Further analysis of antibiotic resistance and diagnosis using molecular genetic methods confirmed the heterogeneity of this culture with the others. Thus, it can be stated that there were two clonal groups of E. albertii in birds – containing and not containing the intimin eae gene. CDT-toxin genes were present in both groups, but no shigatoxin genes were detected. Apparently, the strain not carrying the intimin gene (and in the absence of shigatoxin genes) is avirulent in humans. For the remaining E. albertii strains, the virulence potential will be studied later after full-genome sequencing data is performed. In addition, it is necessary to obtain in vivo data on laboratory animals for a definitive conviction that they are virulent to birds because the mechanisms of pathogenesis in humans and birds may be different [27]. This work will be continued in the future.
It is interesting to note the presence of microcin M4 and B17 genes in the isolated cultures. M4 microcin of class IIb (siderophore peptide) [28] has antimicrobial activity against Escherichia coli and Salmonellae which may explain the fact that S. enterica is not found in the intestine. B17 is a class I microcin, a post-translationally modified peptide that inhibits bacterial DNA gyrase [29] has an even broader spectrum of antimicrobial activity affecting most Enterobacteriaceae. It is possible that the presence of these microcins may increase the distribution of E. albertii in the intestine ultimately increasing the risk of infection caused by it.
5. Conclusions
Significant mortality of poultry on quail farms with signs of GI lesions is caused, in our opinion, by E. albertii. During the study it was found that according to the molecular-genetic and phenotypic properties of isolated strains in quails there were at least 2 clonal groups of E. albertii differing in antibiotic resistance, biochemical indices and presence of the eae (intimin) gene. CDT-toxin genes were present in all strains and shigatoxin genes were absent. The described infection is the first evidence of E. albertii as a cause of death in industrial poultry.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, MGT and NNK; methodology, MGT and NNK; software, MGT and AAA; validation, MGT, NNK and AAA; formal analysis, MGT and YPS; investigation, MGT, AAA and OIT; resources, MGT and OEK; data curation, MGT, AAA, OIT and NNK; writing—original draft preparation, MGT and AAA; writing—review and editing, MGT, NNK, YPS and OEK; visualization, MGT and AAA; supervision, MGT; project administration, MGT and NNK; funding acquisition, MGT and NNK All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by framework of the industry program of Rospotrebnadzor.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
None declared.
Acknowledgments
We are grateful to Razifa Salikhovna Sherstyukova and Yuri Ivanovich Khatyushin for comprehensive assistance at the stage of preparation of microbiological works.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Oaks, J.L.; Besser, T.E.; Walk, S.T.; Gordon, D.M.; Beckmen, K.B.; Burek, K.A. Escherichia albertii in wild and domestic birds. Emerg Infect Dis. 2010, 16, 638–46. [Google Scholar] [CrossRef]
- Albert, M.J.; Alam, K.; Islam, M.; Montanaro, J.; Rahaman, A.S.; Haider, K. Hafnia alvei, a probable cause of diarrhea in humans. Infect Immun. 1991, 59, 1507–13. [Google Scholar] [CrossRef] [PubMed]
- Masuda, K.; Ooka, T.; Akita, H.; Hiratsuka, T.; Takao, S.; Fukada, M. Epidemiological Aspects of Escherichia albertii Outbreaks in Japan and Genetic Characteristics of the Causative Pathogen. Foodborne Pathog Dis. 2020, 17, 144–150. [Google Scholar] [CrossRef]
- Tokuoka, E.; Furukawa, K.; Nagamura, T.; Harada, F.; Ekinaga, K.; Tokunaga, H. Food poisoning outbreak due to atypical EPEC OUT:HNM, May 2011-Kumamoto. Infect. Agents Surveill. Rep. 2012, 33, 8–9. [Google Scholar]
- Murakami, K.; Maeda-Mitani, E.; Kimura, H.; Honda, M.; Ikeda, T.; Sugitani, W. Non-biogroup 1 or 2 Strains of the Emerging Zoonotic Pathogen Escherichia albertii, Their Proposed Assignment to Biogroup 3, and Their Commonly Detected Characteristics. Front Microbiol. 2019, 10, 1543. [Google Scholar] [CrossRef] [PubMed]
- Muchaamba, F.; Barmettler, K.; Treier, A.; Houf, K.; Stephan, R. Microbiology and Epidemiology of Escherichia albertii-An Emerging Elusive Foodborne Pathogen. Microorganisms. 2022, 10, 875. [Google Scholar] [CrossRef]
- Liu, B.; Knirel, Y.A.; Feng, L.; Perepelov, A.V.; Senchenkova, S.N.; Reeves, P.R. Structural diversity in Salmonella O antigens and its genetic basis. FEMS Microbiol Rev. 2014, 38, 56–89. [Google Scholar] [CrossRef]
- Liu, Q.; Bai, X.; Yang, X.; Fan, G.; Wu, K.; Song, W. Identification and Genomic Characterization of Escherichia albertii in Migratory Birds from Poyang Lake, China. Pathogens. 2022, 12, 9. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, L.; Cao, L.; Zeng, X.; Gillespie, B.; Lin, J. Isolation and characterization of Escherichia albertii originated from the broiler farms in Mississippi and Alabama. Vet Microbiol. 2022, 267, 109379. [Google Scholar] [CrossRef]
- Barmettler, K.; Biggel, M.; Treier, A.; Muchaamba, F.; Vogler, B.R.; Stephan, R. Occurrence and Characteristics of Escherichia albertii in Wild Birds and Poultry Flocks in Switzerland. Microorganisms. 2022, 10, 2265. [Google Scholar] [CrossRef]
- Alispahic, M.; Hummel, K.; Jandreski-Cvetkovic, D.; Nöbauer, K.; Razzazi-Fazeli, E.; Hess, M. Species-specific identification and differentiation of Arcobacter, Helicobacter and Campylobacter by full-spectral matrix-associated laser desorption/ionization time of flight mass spectrometry analysis. J Med Microbiol. 2010, 59, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, M.; Yoshioka, S.; Ito, M.; Ahsan, C.R. Characterization of the Emerging Enteropathogen Escherichia Albertii Isolated from Urine Samples of Patients Attending Sapporo Area Hospitals, Japan. Int J Microbiol. 2022, 6, 4236054. [Google Scholar] [CrossRef]
- EU-RL VTEC_Method_02_Rev 0, EU Reference Laboratory VTEC, Rome, Italy.
- Dubois, D.; Delmas, J.; Cady, A.; Robin, F.; Sivignon, A.; Oswald, E. Cyclomodulins in urosepsis strains of Escherichia coli. J Clin Microbiol. 2010, 48, 2122–9. [Google Scholar] [CrossRef]
- van der Westhuizen, W.A.; Bragg, R.R. Multiplex polymerase chain reaction for screening avian pathogenic Escherichia coli for virulence genes. Avian Pathol. 2012, 41, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of minimal predictors of avian pathogenic Escherichia coli virulence for use as a rapid diagnostic tool. J Clin Microbiol. 2008, 46, 3987–96. [Google Scholar] [CrossRef]
- Telhig, S.; Ben Said, L.; Zirah, S.; Fliss, I.; Rebuffat, S. Bacteriocins to Thwart Bacterial Resistance in Gram Negative Bacteria. Front Microbiol. 2020, 11, 586433. [Google Scholar] [CrossRef] [PubMed]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe. 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Kallapura, G.; Morgan, M.J.; Pumford, N.R.; Bielke, L.R.; Wolfenden, A.D.; Faulkner, O.B. Evaluation of the respiratory route as a viable portal of entry for Salmonella in poultry via intratracheal challenge of Salmonella Enteritidis and Salmonella Typhimurium. Poult Sci. 2014, 93, 340–6. [Google Scholar] [CrossRef]
- Camba, S.I.; Del Valle, F.P.; Shirota, K.; Sasai, K.; Katoh, H. Evaluation of 3-week-old layer chicks intratracheally challenged with Salmonella isolates from serogroup c1 (O:6,7) and Salmonella Enteritidis. Avian Pathol. 2020, 49, 305–310. [Google Scholar] [CrossRef]
- Moore, J.E.; Murray, L.; Fanning, S.; Cormican, M.; Daly, M.; Delappe, N. Comparison of phenotypic and genotypic characteristics of Salmonella bredeney associated with a poultry-related outbreak of gastroenteritis in Northern Ireland. J Infect. 2003, 47, 33–9. [Google Scholar] [CrossRef]
- Teymurazov, M.G.; Platonov, M.E.; Tazina, O.I.; Manin, T.B. The biological properties of the strains Avibacterium paragallinarum and Gallybacterium anatis isolated in the Russian Federation in 2015. Veterinary, 6.
- Manin, TB.; Маximov, Т.P.; Teimurazov, M.G.; Platonov, М.Е. The new data of spread of infectious coryza in Russia and some CIS countries in 2013 – 2015 years. Veterinary, 4.
- Persson, G.; Bojesen, A.M. Bacterial determinants of importance in the virulence of Gallibacterium anatis in poultry. Vet Res. 2015, 46, 57. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Jiang, H.; Qi, Z.; Shen, X.; Xue, M.; Hu, J. APEC infection affects cytokine-cytokine receptor interaction and cell cycle pathways in chicken trachea. Res Vet Sci. 2020, 130, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Oaks, J.L.; Besser, T.E.; Walk, S.T.; Gordon, D.M.; Beckmen, K.B.; Burek, K.A. Escherichia albertii in wild and domestic birds. Emerg Infect Dis. 2010, 16, 638–46. [Google Scholar] [CrossRef] [PubMed]
- La Ragione, R.M.; McLaren, I.M.; Foster, G.; Cooley, W.A.; Woodward, M.J. Phenotypic and genotypic characterization of avian Escherichia coli O86:K61 isolates possessing a gamma-like intimin. Appl Environ Microbiol. 2002, 68, 4932–42. [Google Scholar] [CrossRef]
- Vassiliadis, G.; Destoumieux-Garzón, D.; Lombard, C.; Rebuffat, S.; Peduzzi, J. Isolation and characterization of two members of the siderophore-microcin family, microcins M and H47. Antimicrob Agents Chemother. 2010, 54, 288–97. [Google Scholar] [CrossRef]
- Li, Y.M.; Milne, J.C.; Madison, L.L.; Kolter, R.; Walsh, C.T. From peptide precursors to oxazole and thiazole-containing peptide antibiotics: microcin B17 synthase. Science. 1996, 274, 1188–93. [Google Scholar] [CrossRef]
Figure 1.
Pathological anatomical image of the dissection of the affected intestine with the site from which cultures of E. albertii were isolated.
Figure 1.
Pathological anatomical image of the dissection of the affected intestine with the site from which cultures of E. albertii were isolated.
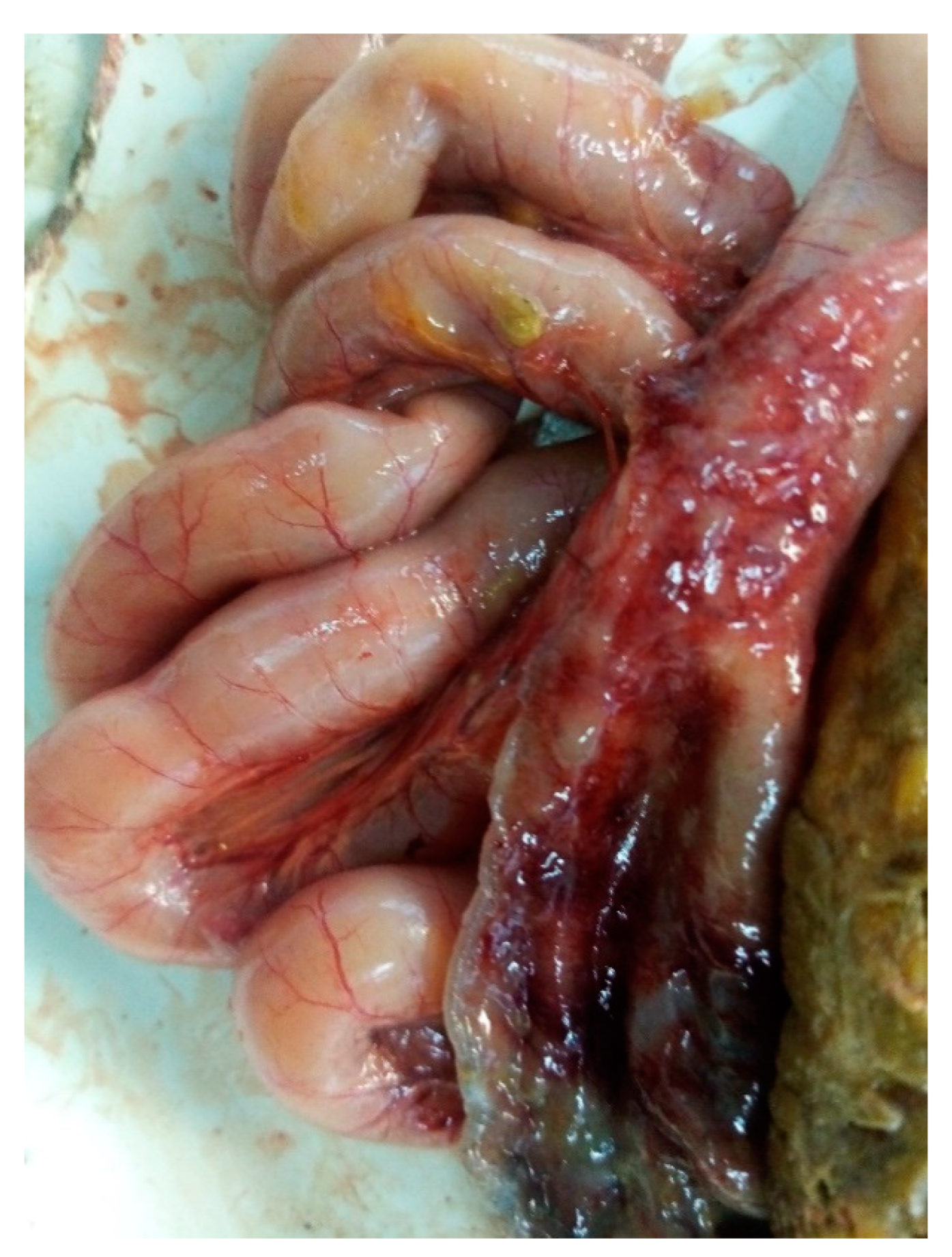
Figure 2.
Antibiotic resistance of E. albertii cultures using the disk diffusion method (automatic detection using Scan 500 Intersciense).
Figure 2.
Antibiotic resistance of E. albertii cultures using the disk diffusion method (automatic detection using Scan 500 Intersciense).


Table 1.
List of primers used in the study.
| The mdh, lysP and clpX genes are specific to E. albertii | |||||
|---|---|---|---|---|---|
| Gene | Primer | 5’-3’ | Annealing, °C | Product size (bp) | Reference |
|
Mdh malatedehydrogenase |
mdh-F | CTGGAAGGCGCAGATGTGGTACTGATT | 55 | 115 | [Ошибка! Истoчник ссылки не найден.] |
| mdh-R | CTTGCTGAACCAGATTCTTCACAATACCG | ||||
|
Lys Lysine-specific transporter |
lysP-F | GGGCGCTGCTTTCATATATTCTT | 55 | 252 | [Ошибка! Истoчник ссылки не найден.] |
| lysP-R | TCCAGATCCAACCGGGAGTATCAGGA | ||||
|
Clp Heat shock protein |
clpX-F | TGGCGTCGAGTTGGGCA | 55 | 384 | [Ошибка! Истoчник ссылки не найден.] |
| clpX-R | TCCTGCTGCGGATGTTTACG | ||||
| Pre-denaturation 95°C – 5 min; 25 cycles: denaturation 95°C – 1 min, annealing 55°C – 1 min, elongation 72°C – 1 min-. Final elongation 72°C – 3 min. | |||||
| E. coli APEC virulence genes | |||||
|
cva colicin V plasmid |
cvaF | CACACACAAACGGGAGCTGTT | 63 | 672 | [Ошибка! Истoчник ссылки не найден.] |
| cvaR | CTTCCGCAGCATAGTTCCAT | ||||
|
omp episomal outer membrane protease |
ompF | TCATCCCGGAAGCCTCCCTCACTACTAT | 63 | 496 | [Ошибка! Истoчник ссылки не найден.] |
| ompR | TAGCGTTTGCTGCACTGGCTTCTGATAC | ||||
|
iroN salmochelinsiderophore receptor |
ironF | AAGTCAAAGCAGGGGTTGCCCG | 63 | 667 | [Ошибка! Истoчник ссылки не найден.] |
| ironR | GATCGCCGACATTAAGACGCAG | ||||
|
fim fimbriae |
fimF | GGATAAGCCGTGGCCGGTGG | 63 | 331 | [Ошибка! Истoчник ссылки не найден.] |
| fimR | CTGCGGTTGTGCCGGAGAGG | ||||
|
iut Aerobactinsiderophore receptor |
iutF | GGCTGGACATCATGGGAACTGG | 63 | 302 | [Ошибка! Истoчник ссылки не найден.] |
| iutR | CGTCGGGAACGGGTAGAATCG | ||||
|
iss Episomal gene for increased survival in serum |
issF | CAGCAACCCGAACCACTTGATG | 63 | 323 | [Ошибка! Истoчник ссылки не найден.] |
| issR | AGCATTGCCAGAGCGGCAGAA | ||||
|
hly hemolysin |
hlyF | GGCCACAGTCGTTTAGGGTGCTTACC | 63 | 450 | [Ошибка! Истoчник ссылки не найден.] |
| hlyR | GGCGGTTTAGGCATTCCGATACTCAG | ||||
|
eae intimin |
eaeF | CATTGATCAGGATTTTTCTGGTGATA | 63 | 102 | [Ошибка! Истoчник ссылки не найден.] |
| eaeR | CTCATGCGGAAATAGCCGTTA | ||||
| Pre-denaturation 95°C – 5 min; 35 cycles: denaturation 95°C – 30 sec, annealing 63°C – 45 sec, elongation 72°C – 1 min 45 sec. Final elongation 72°C – 5 min. | |||||
| E. colimicrocin genes | |||||
| Microcin B17 | mcc B17-F | TCACGCCAGTCTCCATTAGGTGTTGGCATT | 60 | 135 | [Ошибка! Истoчник ссылки не найден.] |
| mcc B17-R | TTCCGCCGCTGCCACCGTTTCCACCACTAC | ||||
| Microcin C7 | mcc C7-F | CGTTCAACTGTTGCAATGCT | 60 | 134 | |
| mcc C7-R | AGTTGAGGGGCGTGTAATTG | ||||
| Microcin E492 | mcc E492-F | GTCTCTCCTGCACCAAAAGC | 60 | 291 | |
| mcc E492-R | TTTTCAGTCATGGCGTTCTG | ||||
| Microcin H47 | mcc H47-F | CACTTTCATCCCTTCGGATTG | 60 | 227 | |
| mcc H47-R | AGCTGAAGTCGCTGGCGCACCTCC | ||||
| Microcin J25 | mcc J25-F | TCAGCCATAGAAAGATATAGGTGTACCAAT | 60 | 175 | |
| mcc J25-R | TGATTAAGCATTTTCATTTTAATAAAGTGT | ||||
| Microcin L | mcc L-F | GGTAAATGATATATGAGAGAAATAACGTTA | 60 | 233 | |
| mcc L-R | TTTCGCTGAGTTGGAATTTCCTGCTGCATC | ||||
| Microcin V | mcc V-F | CACACACAAAACGGGAGCTGTT | 60 | 680 | |
| mcc V-R | TTTCGCTGAGTTGGAATTTCCTGCTGCATC | ||||
| Microcin M | micM-4-F | CGTTTATTAGCCCGGGATTT | 60 | 166 | |
| micM-4-R | GCAGACGAAGAGGCACTTG | ||||
| Pre-denaturation 95°C – 5 min; 35 cycles: denaturation 95°C – 30 sec, annealing 60°C – 30 sec, elongation 72°C – 30 sec. Final elongation 72°C – 5 min. | |||||
| Primers encoding different types of cdt genes and cif gene in E. coli | |||||
| cdtB-II, cdtB-III, cdtB-V | CDT-s1 | GAAAGTAAATGGAATATAAATGTCCG | 60 | 467 | [Ошибка! Истoчник ссылки не найден.] |
| CDT-as1 | AAATCACCAAGAATCATCCAGTTA | ||||
| cdtB-II* | CDT-IIas | TTTGTGTTGCCGCCGCTGGTGAAA | 62 | 556 | |
| cdtB-III, cdtB-V* | CDT-IIIas | TTTGTGTCGGTGCAGCAGGGAAAA | 62 | 555 | |
| cdtB-I, cdtB-IV | CDT-s2 | GAAAATAAATGGAACACACATGTCCG | 56 | 467 | |
| CDT-as2 | AAATCTCCTGCAATCATCCAGTTA | ||||
| cdtB-I | CDT-Is | CAATAGTCGCCCACAGGA | 56 | 411 | |
| CDT-Ias | ATAATCAAGAACACCACCAC | ||||
| cdtB-IV | CDT-IVs | CCTGATGGTTCAGGAGGCTGGTTC | 56 | 350 | |
| CDT-IVas | TTGCTCCAGAATCTATACCT | ||||
| cdtC-V | P105 | GTCAACGAACATTAGATTAT | 49 | 748 | |
| c2767r | ATGGTCATGCTTTGTTATAT | ||||
| cif | cif-int-s | AACAGATGGCAACAGACTGG | 55 | 383 | |
| cif-int-as | AGTCAATGCTTTATGCGTCAT | ||||
| Pre-denaturation 95°C – 5 min; 35 cycles: denaturation 95°C – 30 sec, annealing – 30 sec, elongation 72°C – 30 sec. Final elongation 72°C – 5 min. | |||||
* Reverse primers, used with the CDT-s1 primer.
Table 2.
Summary of the results of the bacteriological isolation of pathogenic microorganisms from poultry.
Table 2.
Summary of the results of the bacteriological isolation of pathogenic microorganisms from poultry.
| No. of sample, workshop, age | Parenchymatous organs (liver, lungs, heart, spleen) | The contents of the sinuses, trachea | Intestines | |
|---|---|---|---|---|
| 1 line, 21d | 1 | Staphylococcus piscifermentans | Salmonella bredeney, Streptococcus pluranimalium, Bordetella hinzii | Clostridium perfringens |
| 2 | Staphylococcus piscifermentans | Salmonella bredeney, Bordetella hinzii | Clostridium perfringens | |
| 3 | Staphylococcus piscifermentans | Salmonella bredeney | Clostridium perfringens | |
| 4 | Staphylococcus piscifermentans | Salmonella bredeney, Streptococcus pluranimalium, Bordetella hinzii | ||
| 5 | Salmonella bredeney, Bordetella hinzii | |||
| 2 line, 21d | 1 | Staphylococcus piscifermentans | Salmonella bredeney, Streptococcus pluranimalium | |
| 2 | Staphylococcus piscifermentans | Gallibacterium anatis, Staphylococcus piscifermentans | Clostridium perfringens | |
| 3 | Staphylococcus piscifermentans | Streptococcus pluranimalium | ||
| 4 | Staphylococcus piscifermentans | Gallibacterium anatis, Staphylococcus piscifermentans | ||
| 5 | Staphylococcus piscifermentans | Gallibacterium anatis, Staphylococcus piscifermentans | Clostridium perfringens | |
| 3 line, 18d | 1 | Staphylococcus piscifermentans | Gallibacterium anatis, Streptococcus pluranimalium, Bordetella hinzii | |
| 2 | Salmonella bredeney, Streptococcus pluranimalium, Bordetella hinzii | |||
| 3 | Staphylococcus piscifermentans | Salmonella bredeney, Streptococcus pluranimalium, Bordetella hinzii, Gallibacterium anatis | ||
| 4 | Staphylococcus piscifermentans | Streptococcus pluranimalium, Bordetella hinzii | ||
| 5 | Gallibacterium anatis, Salmonella bredeney | |||
| 4 line, 18 d | 1 | Gallibacterium anatis, Streptococcus pluranimalium, E.coli | ||
| 2 | Staphylococcus piscifermentans | Gallibacterium anatis, Streptococcus pluranimalium, Staphylococcus piscifermentans | ||
| 3 | Gallibacterium anatis | |||
| 4 | Staphylococcus piscifermentans | Gallibacterium anatis, Streptococcus pluranimalium, Staphylococcus piscifermentans | ||
| 5 | Staphylococcus piscifermentans | Staphylococcus piscifermentans | Escherichia albertii | |
| m/f, 18 d | 1 | Salmonella bredeney | Escherichia albertii, Clostridium perfringens | |
| 2 | Staphylococcus piscifermentans | E.coli | ||
| 3 | E.coli | Escherichia albertii | ||
| 4 | Salmonella bredeney | Escherichia albertii | ||
| 5 | Salmonella bredeney | Escherichia albertii | ||
Table 3.
Summary of the results of the bacteriological isolation of pathogenic microorganisms from poultry.
Table 3.
Summary of the results of the bacteriological isolation of pathogenic microorganisms from poultry.
| Antibiotics by Vitek2 | E. albertii group 1 with the eae gene (in parentheses MPC mg/l) | E. albertii group 2 without eae gene |
|---|---|---|
| BLRS | - | - |
| Ampicillin | S (8) | R (16) |
| Amoxicillin/clavulonic acid | S (8) | R (16) |
| Cefotaxime | S (≤ 0,25) | S (≤ 0,25) |
| Ceftazidime | S (≤ 0,12) | S (≤ 0,12) |
| Cefipime | S (≤ 0,12) | S (≤ 0,12) |
| Eatapenem | S (≤ 0,12) | S (≤ 0,12) |
| Meropenem | S (≤ 0,25) | S (≤ 0,25) |
| Amikacin | S (≤ 2) | S (≤ 2) |
| Gentamicin | S (≤ 1) | S (≤ 1) |
| Netilmicin | ND (≤ 1) | ND (≤ 1) |
| Ciprofloxacin | R (1) | R (1) |
| Tigecycline | S (≤ 0,5) | S (≤ 0,5) |
| Fosfomycin | ≤ 16 | ≤ 16 |
| Nitrofurantoin | S (≤ 16) | S (≤ 16) |
| Trimethoprim/sulfamethoxazole | S (≤ 20) | S (≤ 20) |
| Antibiotics using the disk-diffusion method | ||
| Amoxicillin/clavulonic acid, 30 | S | R |
| Ampicillin, 25 | S | R |
| Ceftazidime, 10 | S | S |
| Gentamicin, 10 | R | R |
| Colistin, 10. | R | R |
| Cefepime, 30. | R | R |
| Chloramphenicol, 30 | S | S |
| Imipenem, 10 | S | S |
| Trimethoprim/sulfamethoxazole | S | S |
| Amikacin, 30 | R | R |
| Fosfomycin, 50 | R | R |
| Norfloxacin, | R | R |
| Tigecycline | R | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



