
Submitted:
31 January 2025
Posted:
05 February 2025
You are already at the latest version
Abstract
Bluehead gilia or bluefield gilia (Gilia capitata Sims) from the family Polemoniaceae is an annual herbaceous plant widely distributed in the western regions of North America, but cultivated as an ornamental flower in various regions to support pollinators. The chemical composition of the plant has not been studied before. For the first time, essential oil (EO) was obtained by hydrodistillation from the different parts of gilia, harvested in Estonia. The yield of EOs was from 0.42 to 1.97 mL/kg, with the greatest yields from seeds, flowers, fruits, and leaves. Its component composition was studied using the GC-MS method. 118 compounds have been identified. The EO of flowers was dominated by the hexahydrofarnesyl acetone (19.1%), followed by palmitic acid (12.2%), γ-decalactone (7.1%), (Z)-2-p-menthen-1-ol (3.6%). Hexahydrofarnesyl acetone was also dominant in most parts of the G. capitata. The predominant compounds in the EO of leaves were diterpene alcohols: phytol (23.3%), isomanool (12.1%) and sclareol (4.5%), apocarotenoids, C13-norisoprenoid (E)-β-ionone (4.5%). The most dominant compounds in EO of stems are isomanool (8.3 %), sclareol (6.7%) and (E,E)-2,4-decadienal (5.3%). The dominant in the fruits were hexahydrofarnesyl acetone (18.2%), palmitic acid (8.0%), heptacosane (5.3%) and γ-decalactone (4.6 %). The dominant in the shell were (+)-epi-bicyclosesquiphellandrene (15.4%), β-elemene (8.5%) and pentadecanal (6.3%). The dominant in the seeds were hexahydrofarnesyl acetone (15.2%), palmitic acid (11.1%), 1,3-d’dimethylbenzene (10.3%) and heptacosane (5.0%). The dominant in the root were (-)-myrtenol (25.7%), (E)-myrtanol (16.4%), palmitic acid (8.1%) and 2-pentylfuran (2.9%). The results hint at further study options on the antimicrobial and anti-inflammatory effects of G. capitata.
Keywords:
Gilia capitata
; essential oil
; ornamental flower
; volatile compounds
1. Introduction
Gilia capitata Sims, with the common name bluehead gilia or blue field gilia (bule gilia), belongs to the family Polemoniaceae, and is a close relative to floxes and polemonium [1]. G. capitata Sims belongs to the subgenus Gilia and the Eugilia section. The plants that make up this section are herbaceous annuals with blue and purple flowers gathered in loose false umbrellas, corymbs or heads. The section consists of 9 species, among which are G. achilleaefolia, G. capitata, G. millefoliata, G. clivorum, G. laciniata, G. tricolor, G. angelensis, G. nevinii, and G. valdiviensis [2,3].
G. capitata is native to all western states of the USA [4,5]. The native habitats of G. capitata are sunny rocky hillslopes and open fields with well-drained soils at an elevation range of up to 2000 m [6,7]. It is introduced widely throughout western North America, Alaska, northern Mexico, and Europe as an ornamental flower [8].
G. capitata is a polytypic species. It is phenotypically plastic in response to environmental conditions [9,10]. It has a wide morphological diversity because of the ecotypic and genotypic variability [6]. Eight defined subspecies form a morphological gradation throughout the Pacific coast of North America [6]. Chromosomes: 2n=18. The flagship sub-species, G. capitata subsp. capitata, is an herbaceous annual plant with a slender 20-100 cm erect stem. Stem can be simple or branched in the upper third, glabrous, glandular or slightly floccose. The root system consists of a relatively shallow main root. On base and stem, cauline leaves are bipinnately dissected, usually glabrous, 4–10 cm long, and with 5–20 mm lobes; upper leaves are more straightforward linear. Blue (or blue-violet) campanulate flowers are clustered into globose to conoidal inflorescences with (25) 50–100 flowers at the terminal position of each stem or branch. Inflorescence diameter varies between 14–40 mm. Individual flowers therein are 0.6-2.5 mm [9] with acute lobes 0.6–1 mm wide and 2 mm long, straight-tipped and with a 6-8 mm corolla. Pollen has a light blue colour. The flowers of the gilia are protandrous, i.e. the anthers mature first, and stigmas rise a few days later [2,11]. Gilia is mostly self-compatible; only some populations are facultatively outcrossing [2,11], though [9] reports that gilia is self-incompatible. The fruit is a three-celled globose or obovoid capsule, which splits at maturity only partly as three valves (i.e. it has an indehiscent capsule)). The capsule contains 1-10(25) seeds, one or two in each cell. The capsule remains indehiscent in dry weather and splits only partly open with the rain. When dry, capsules break off easily by touch. Seeds are hard, brown, ovoid or ovoid-angled, 1–2.3 mm long. When wet, the verrucose seed coat becomes a sticky, mucilaginous coat; this feature probably serves to stick seeds to animals for dispersal and later to substrate [9].
G. capitata is easily grown from seed. No stratification or scarification is required (though two weeks of cold stratification has been suggested to speed up germination). Seeds can be sown directly into the garden in spring. Still, we observed that they reseed reasonably well in autumn and overwinter in the Estonian winter to germinate in early spring. Minimal maintenance is required except for weeding. Soil should be moist when seeds are planted, but adult plants are drought-tolerant. Cutting deadhead flowers will encourage reflowering. In nature, gilia profits from wildfires, removing mature vegetation and opening the ground for light [12]. Its germination probably reacts to the signals from fire residues [13]. It grows in gardens alone or with other low-competition annuals in sunny open locations. Grows well in well-drained medium to coarse soil with a neutral pH (6.0-7.0). In Estonia, G. capitata is cultivated as an ornamental plant.
The gilia is declared one of the pollinator magnets (e.g., Blog of Oregon Native Plants for Pollinators [14] ), and it is included in many wildflower seed mixes for bees. The flowering can be long-term continuous depending on water and temperature conditions. G. capitata needs an insect vector for pollination; flowers are visited by a wide range of insects, including small-sized pollinators and wild bees, bumblebees, long-tongued flies, and butterflies [15,16]. The attractiveness has been partly explained by its tight capitate inflorescence structure, composed of up to 80 flowers and forms a convenient landing pad for all insects [15]. Some of its essential oils could also be one of the causes of flower visit attraction of pollinators [17].
The chemical composition of G. capitata has not been studied before. Therefore, this work aimed to study the component composition of essential oil (EO) of different morphological groups of raw materials of G. capitata.
2. Materials and Methods
2.1. Plant Material
Seeds, obtained from different sources, were mixed and sown in spring. Plants were grown in field conditions at three sites with different soil conditions around Tartu, Estonia. Later, all the collected plant material was mixed in relatively equal proportions before analysis. During the growth, plant growth was supported with complex fertilizers. Plant material was collected twice. The main collection of plant material was done at the time of mass-flowering. We observed two waves of flowering – the first inflorescence formed at the top of the main stem and, several weeks later, on lateral stems – the material was collected in both rounds and later mixed. Depending on the site phenology, the first forage was done at the end of June 2023 and early weeks of July. Plant parts (flowers, leaves, roots, fruits) were collected separately in the field. Later, after drying, the leaves and stems were separated. The second collection was done in late summer in August 2023 (the date depended on the site and weather), at the time of fruit and seed ripening – the heads and clusters of fruit capsules were collected and dried. Later, fruit capsules were crushed, and seeds were separated. Capsule residues containing mainly fruit valves and walls were kept as a separate fraction as shells of the fruit wall (hereafter shells). The plant material was dried for 14 days at room temperature in a well-ventilated room. It was stored at room temperature in paper bags before distillation of essential oil. The voucher sample specimens (No Polemon/Gil1-7) are available at the Institute of Pharmacy, University of Tartu, Estonia.
2.2. Hydrodistillation of Essential Oil
The EO hydrodistilled from the different dried parts of G. capitata using the method described in the European Pharmacopoeia [18]. The plant materials (20 g) with 400 mL of purified water were hydrodistilled in a 1000 mL round-bottom flask for 2 hours (3–4 ml/min). Hexane (0.5 mL) was added to a graduated tube to remove the distilled oil [19,20]. The yield of EO was measured using European Pharmacopoeia with xylol [18]. Due to the content of saponins, intense foam was produced during the distillation of most plant parts, to reduce which 30 g of potassium chloride was added to the flask (except for the roots and stems).
2.3. Gas Chromatography/Mass Spectrometry
The samples of EO were analysed by gas chromatography-mass spectrometry (GC/MS), using an Agilent 6890/5973 GCMS system controlled by MSD Chemstation. 1 µL of the sample was injected at an injector temperature of 280 oC in split mode (10:1), using He as the carrier gas onto Agilent HP-5MSI column (30 m length, 0.25 mm inner diameter, 0.25 µm film thickness). The carrier gas was held at the constant flow rate of 1 mL/min. The oven was held at 50 oC for 2 min, followed by a ramp of 4 oC/min to a final temperature of 280 oC and held at 280 oC for 5 minutes. The MSD was operated in EI mode at 70 eV. After a delay time of 4 min mass spectra were recorded in the range of 29 – 400 m/z at a rate of 3.8 scans per second. The data were analysed by Agilent Masshunter Data Analysis Software using a deconvolution algorithm with different window size factors. Obtained compounds were identified by using NIST23 library with Match Factor ≥ 90 and by retention indexes (relative to n-alkanes C8 – C30) or obtained by the analysis of the reference compounds. The area percentages of each peak were calculated from the total areas in the chromatograms without using correction factors [19,20].
2.4. Data Analysis
The similarity of plant parts in their composition of EO was analysed using the cluster analysis, implemented in PC-ord v7 [21]. The Sorensen (Bray-Curtis) distance measure of similarity was used to build the cluster tree by applying the Flexible beta linkage method with parameter -0.25 (a default value). The log-transformed proportion of oils was used, and only components with at least 1% were included in the data set to reduce noise.
3. Results
As a result of the study, it was found that different parts of G. capitata accumulated from 0.42 ml/kg (shells) to 1.97 ml/kg (seeds) of EO (Figure 1). It should be noted that the flowers contain less essential oil than the seeds. According to the content of EO, raw materials are arranged in the following order: seeds > flowers > fruits > leaves > stems > roots > shells. Seeds contain a bit less than twice as many essential oils as flowers and leaves.
The composition of hydrodistillates was studied by GC-MS (Table 1). In total, 118 compounds were identified, representing 81.31-93.74 % in the oil (Table 1). All parts of the plant contain benzeneacetaldehyde, nonanal, (E)-2-nonenal, (-)-myrtenol, (E,E)-2,4-decadienal, tetradecanal, pentadecanal, hexahydrofarnesyl acetone, and di-2-methylpropylphtalate.
The composition of essential oils in different parts of G. capitata was illustrated with a cluster tree (Figure 2). Fruits and seeds were most similar in composition; flowers were next similar ones. Leaves and stems formed another narrow cluster. The most distant were shells of the fruit. The composition of roots was the most distinct from all other parts of a plant (Figure 2).
4. Discussion
As mentioned above, the yield of EO in different parts of G. capitata was 0.42-1.97 ml/kg (Figure 1). We compare that with results obtained in studies of some well-known medicinal plants studied. The aerial parts of Origanum vulgare from different countries contained 2-11 ml/kg of EO [22], in Thymus vulgaris herb the yield of EO was 3-28 ml/kg [23], in commercial samples of Valeriana officinalis roots from various countries different origin 2-10 ml/kg [24], and in the herb of Artemisia absinthium from different origin 2-4 ml/kg [25]. The branches of Juniperus communis shrubs were collected from 27 different habitats in Estonia and contained 0.3-6 ml/kg of EO [26]. Therefore, the oil content in G. capitata is modest compared to classic EO plants.
The essential oil of flowers was dominated by hexahydrofarnesyl acetone (19.1%), followed by palmitic acid (12.2%), γ-decalactone (7.1%) and (Z)-2-p-menthen-1-ol (3.6%) (Figure 3). In addition to the flowers, hexahydrofarnesyl acetone is also the dominant component in fruits (18.2%) and seeds (15.2%), and it is found in significant quantities in the stems (4.9%) of the G. capitata (Figure 3). Hexahydrofarnesyl acetone has been proven to exhibit a potent antimicrobial, anti-inflammatory, and cytotoxic activity and is used in pain relief research [27,28,29,30]. Palmitic acid, present also in seeds (11.1%) and fruits (8.0%), has antioxidant and antibacterial activity [31,32].
Lactones, including γ-decalactone, constitute an essential group of fatty acid-derived volatile organic compounds conferring peach-like aroma to a few essential oils and fruits including peach, plum, pineapple and strawberry [33]. γ-Decalactone inhibits strawberry pathogen growth and achene germination [34].
Menthane monoterpenoids (Z)-2-p-menthen-1-ol and (E)-p-2-menthen-1-ol are found in all organs of the aboveground part of the plant. They make up 5.7% of flowers, 2.2% of leaves, 6.0% of stems, 5.6% of fruits, 3.3% of shells and 4.6% of seeds. Menthenol derivatives have potentially different biological properties. 1-Methyl-4-(1-methylethenyl)-2-cyclohexen-1-ol is an acetal reagent used in the synthesis of desoxy cannabidiols and THC related psychoactive compounds. It is formed from (+)-Limonene using a photosynthesized O2 transfer [35].
In addition to the dominant compounds in the flowers, (Z)-γ-bisabolene was also found (though, it is similarly abundant in stems and fruits). The antitumor activity of γ-bisabolene in human neuroblastoma cells has been proven through the induction of p53-mediated mitochondrial apoptosis [36]. γ-Bisabolene has demonstrated antiproliferative activities against several human cancer cell lines. Another study [29] disclosed the antiproliferative and apoptosis induction activities of γ-bisabolene to human neuroblastoma TE671 cells, and a CC50 value of γ-bisabolene was 8.2 μM to TE671 cells [37].
Caryophyllene oxide is found in flowers (0.9%), while caryophyllene was distributed more widely in flowers, leaves, fruits and shells (1.3, 0.5, 2.8%, respectively). Its anticancer, antioxidant and antimicrobial properties have been proven [38].
Linalool is found in flowers (0.3%), leaves (0.5%), stems (0.3%), shells (0.6%) and seeds (0.3%). Linalool and (E)-caryophyllene exhibited high cytotoxic activity against the amelanotic melanoma and renal adenocarcinoma cells [39].
The characteristic terpenoids of G. capitata are shown in Figure 4.
The predominant compounds in the EOs of the leaves were phytol (23.3%) and diterpene alcohols isomanool (12.1%) and sclareol (4.5%), (E)-β-ionone (4.5%).
Phytol has antibacterial and antioxidant activity, inhibiting the growth of Staphylococcus aureus [40,41]. Phytol is used in fragrance and cosmetics to produce shampoos, toilet cleaners, household cleaners and detergents [42].
Labdane diterpenoids are the most common types of diterpenoids isolated in minute amounts from higher plants [43]. Labdane diterpenoids are interesting for their cytotoxic, antifungal, anti-inflammatory, antiparasitic and analgesic properties [44,45,46]. Labdane diterpenoid sclareol has a wide range of bioactivity, including anti-tumor, anti-inflammation and anti-pathogenic microbes, and even anti-diabetes and hypertension [47]. Sclareol also can kill human leukemic cells and colon cancer cells in vitro by apoptosis [48,49]. Labdanum-type diterpenoids are usually found in the plant as a mixture of components. Therefore, we found it necessary to provide all the structural formulas of labdanum diterpenoids contained in G. capitata raw materials (Figure 4). Labdan-type diterpenes, such as manoyl oxide, 13-epi-manoyl oxide, epi-13-manool, kolavelool, isomanool, sclareol make up a significant part of leaves and stems oils – 22.63% and 20.2%, respectively. Fruits and seeds essential oils contain less– 3.85% and 2.99%, respectively. The main component among labdan diterpenes is isomanool (labda-8,14-diene-13β-ol), it is found in flowers 0.39%, leaves 12.07%, stems 8.29%, fruits 3.23% and seeds 2.48%. It is followed by manoyl oxide, found in leaves 2.28%, stems 2.96%, fruits 0.62%, seeds 0.51%. They are absent in shells and roots. Nor-labdane diterpenoids ambrial is found in trace amounts only in stems.
C-13-isoprenoid, apocarotenoid β-ionone had fungicidal activities against Aspergillus fumigatus. It also ameliorated fungal keratitis in mice by reducing inflammation, which LOX-1, p-p38MAPK and p-JNK regulated [50]. Literature data indicate that β-ionone and its derivatives have a lot of important pharmacological activities, including antibacterial, antifungal, antioxidant, anti-inflammatory, antiproliferative, and anti-cancer [51].
The most dominant compounds in EO of stems are isomanool (8.3 %), sclareol (6.7%) and 2,4-decadienal, (E,E)- (2.9%). Known about nematicidal activity 2,4-decadienal [52] and insecticidal against the barn bug [53].
In addition to hexahydrofarnesyl acetone, the dominant in the fruits were palmitic acid (8.0%), heptacosane (5.3%) and γ-decalactone (4.6%). The dominant in the shell were (+)-epi-bicyclosesquiphellandrene (15.4%), β-elemene (8.5%) and pentadecanal- (6.3%). β-Elemene exhibits anti-tumour properties, and anti-inflammatory and antioxidant effects [54,55]. β-Elemene and β-elemene piperazine derivatives have been shown to inhibit tumour cell growth in vitro and in vivo [56]. The anti-bacterial authorities of the pentadekanal were established [57,58].
In addition to hexahydrofarnesyl acetone, the dominant in the seeds were palmitic acid (11.1%), 1,3-dimethylbenzene (10.3%) and heptacosane (5.0%). Heptacosan can improve P-glycoprotein-mediated drug transport, demonstrating the ability to retain the substrate doxorubicin inside the cell and enhancing its cytotoxic effects [59].
The dominant in the root were (-)-myrtenol (25.7%), (E)-myrtanol (16.4%), palmitic acid (8.1%) and 2-pentylfuran (2.9%). Several reports have demonstrated the pharmacological properties of myrtenol, including its antioxidant, antibacterial, antifungal, antidiabetic, anxiolytic, and gastroprotective activities [60,61]. (-)-(E)-Myrtanol is an antimicrobial and acaricide agent [62,63]. 2-Pentylfuran has been suggested as a repellent for spotted-wing drosophila, as it can significantly reduce fruit infestations under field conditions [64]. In the roots, cuparene (0.3%) was found. Cuparene derivatives show moderate to high activity levels on lung cancer cell lines NSCLC-N6 and A549 [65].
A significant number of volatile compounds are aliphatic (24.37 – 48.56%). They are the dominant compounds in flowers, stems, fruits, and seeds (Figure 5).
At the same time, diterpenoids are the dominant group in the composition of volatile compounds in leaves (45.89%), sesquiterpenoids in shells (34.68%), and monoterpenoids in roots (46.43%).
It should also be noted that the content of mono- and sesquiterpenoids is quite high in flowers (12.78% and 26.73%, respectively), stems (9.88% and 5.71%), fruits (9.57% and 18.88%), seeds (8.08% and 15.84%), monoterpenoids in shells (7.93%).
Sesquiterpenoids are represented by derivatives of the naphthalene, such as copaene, humulene, (+)-epi-bicyclosesquiphellandrene, β-selinene, τ-muurolol, α-cadinol and azulene, such as 7-epi-α-cedrene series. Antiviral activity against dengue virus (VDEN) was established for a few sesquiterpenoids, including isomers of copaene, β-caryophillene, caryophillene oxide and (+)-epi-bicyclosquiphellandrene [66]. Norsesquiterpenoids or apocarotenoids are represented by two compounds such as (E)-β-ionone and (E)-β-damascenone. They are absent in the roots. β-Damascenone inhibits the expression of pro-inflammatory cytokines and leukocyte adhesion molecules [67]. β-Damascenol has been shown to be effective in preventing skin sunburn [68].
Aromatic compounds represent an insignificant part of the composition of volatile compounds, are found in all parts of the plant and accumulate in fruits to the maximum extent (11.0%).
5. Conclusions
The composition of volatile compounds of various parts of G. capitata was studied for the first time. The results of the analysis of Gilia essential oil made it possible to detect and identify important potentially active compounds with various biological properties: antioxidant, anti-cancer, antidepressant and others, which indicates the prospects for further phytochemical and pharmacological studies of the genus Gilia. Future studies should focus on Gilia saponins and polyphenolic substances and their biological activity.
6. Patents
No patents.
Author Contributions
Conceptualization, A.R., J.L. and O.K.; methodology, A.R., M.L., T.I., A.K., and O.K.; software, T.I., A.K., and A.R.; validation, A.R., M.L., T.I. and A.K.; formal analysis, A.R., M.L., P.S. and O.K.; investigation, A.R., M.L., P.S. and O.K.; resources, A.R. and J.L.; data curation, A.R., M.L., P.S. and O.K.; writing—original draft preparation, A.R., J.L., T.I., A.K. and O.K.; writing—review and editing, A.R., J.L., M.L., T.I., A.K. and O.K.; visualization, J.L., T.I. and A.K.; supervision, A.R.; project administration, A.R.; funding acquisition, A.R. and O.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Estonian Research Council grant PRG1223 and the European Union in the MSCA4Ukraine project “Design and development of 3D-printed medicines for bioactive materials of Ukrainian and Estonian medicinal plants origin” (ID number 1232466).
Data Availability Statement
The data supporting the results of this study can be obtained from the corresponding authors upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gilia Capitata. Calscape. California Native Plant Society. 2024.
- Grant, V. Genetic and Taxonomic Studies in Gilia: VI. Interspecific Relationships in the Leafy-Stemmed Gilias. aliso 1954, 3, 35–49. [Google Scholar] [CrossRef]
- Grant, V.; Grant, A. Genetic and Taxonomic Studies in Gilia: X. Conspectus of the Subgenus Gilia. Aliso: A Journal of Systematic and Evolutionary Botany 1956, 3, 297–300. [Google Scholar] [CrossRef]
- Jaramillo, Z.; Leigh, J. Revision of the Genus Gilia of Utah. Journal of Undergraduate Research 2019, 105. [Google Scholar]
- Baskin, Je.M.; Baskin, C.C. Propagation Protocol for Production of Container (Plug) Gilia Capitata Sims Plants 2002.
- Grant, V. Genetic and Taxonomic Studies in Gilia: I. Gilia Capitata. aliso 1950, 239–316. [Google Scholar] [CrossRef]
- Porter, J.M. Gilia Capitata Subsp. Capitata, in Jepson Flora Project (Eds.). Jepson eFlora 2023 2023.
- Gilia Capitata Sims. Kew Plants of the World.
- Nagy, E.S.; Rice, K.J. Local Adaptation in Two Subspecies of an Annual Plant: Implications for Migration and Gene Flow. Evolution 1997, 51, 1079. [Google Scholar] [CrossRef]
- Kruckeberg, A.R. INTRASPECIFIC VARIABILITY IN THE RESPONSE OF CERTAIN NATIVE PLANT SPECIES TO SERPENTINE SOIL. American J of Botany 1951, 38, 408–419. [Google Scholar] [CrossRef]
- Brown, H.S. Differential Chiasma Frequencies in Self-Pollinating and Cross-Pollinating Species of the Genus Gilia. Aliso: A Journal of Systematic and Floristic Botany 1961, 67–81.
- Grant, V. Seed Germination in Gilia Capitata and Its Relatives. Madroño 1949, 87–93.
- Keeley, J.E.; Keeley, S.C. Role of Fire in the Germination of Chaparral Herbs and Suffrutescents. Madroño 1987, 240–249.
- Hayes, Je. Top 10 Oregon Native Plants for Pollinators: Week 8.
- Grant, V.; Grant, A. Flower Pollination in the Phlox Family. Columbia Univ. Press, N.Y. 1965.
- Nagy, E.S. Selection for Native Characters in Hybrids Between Two Locally Adapted Plant Subspecies. Evolution 1997, 51, 1469. [Google Scholar] [CrossRef]
- Cseke, L.J.; Kaufman, P.B.; Kirakosyan, A. The Biology of Essential Oils in the Pollination of Flowers. Natural Product Communications 2007, 2, 1934578X0700201225. [Google Scholar] [CrossRef]
- European Pharmacopoeia; 11th ed.; Council of Europe: Strasbourg, 2022.
- Raal, A.; Ilina, T.; Kovalyova, A.; Koshovyi, O. Volatile Compounds in Distillates and Hexane Extracts from the Flowers of Philadelphus Coronarius and Jasminum Officinale. ScienceRise: Pharmaceutical Science 2024, 37–46. [CrossRef]
- Hrytsyk, Y.; Koshovyi, O.; Lepiku, M.; Jakštas, V.; Žvikas, V.; Matus, T.; Melnyk, M.; Grytsyk, L.; Raal, A. Phytochemical and Pharmacological Research in Galenic Remedies of Solidago Canadensis L. Herb. Phyton 2024, 93, 2303–2315. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data. 2018.
- Raal, A.; Gontova, T.; Ivask, A.; Orav, A.; Koshovyi, O. Yield, Composition, and Chemotypes of Essential Oils from Origanum Vulgare L. Aerial Parts Cultivated in Different European Countries. Agronomy 2024, 14, 3046. [Google Scholar] [CrossRef]
- Raal, A.; Gontova, T.; Palmeos, M.; Orav, A.; Sayakova, G.; Koshovyi, O. Comparative Analysis of Content and Composition of Essential Oils of Thymus Vulgaris L. from Different Regions of Europe. PEAS 2024, 73, 332. [Google Scholar] [CrossRef]
- Raal, A.; Kokitko, V.; Odyntsova, V.; Orav, A.; Koshovyi, O. Comparative Analysis of the Essential Oil of the Underground Organs of Valeriana Spp. from Different Countries. Phyton 2024, 93, 1365–1382. [Google Scholar] [CrossRef]
- Raal, A.; Ilina, T.; Kovaleva, A.; Orav, A.; Karileet, M.; Džaniašvili, M.; Koliadzhyn, T.; Grytsyk, A.; Koshovyi, O. Variation in the Composition of the Essential Oil of Commercial Artemisia Absinthium L. Herb Samples from Different Countries. ScienceRise: Pharmaceutical Science 2024, 19–28. [CrossRef]
- Raal, A.; Komarov, R.; Orav, A.; Kapp, K.; Grytsyk, A.; Koshovyi, O. Chemical Composition of Essential Oil of Common Juniper (Juniperus Communis L.) Branches from Estonia. SR: PS 2022, 66–73. [CrossRef]
- Filipowicz, N.; Kamiński, M.; Kurlenda, J.; Asztemborska, M.; Ochocka, J.R. Antibacterial and Antifungal Activity of Juniper Berry Oil and Its Selected Components. Phytotherapy Research 2003, 17, 227–231. [Google Scholar] [CrossRef]
- Razavi, S.M.; Nejad-Ebrahimi, S. Phytochemical Analysis and Allelopathic Activity of Essential Oils of Ecballium Elaterium A. Richard Growing in Iran. Natural Product Research 2010, 24, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, S.; Shi, J.; Sun, Z.; Lei, Z.; Yin, Z.; Qian, Z.; Tang, H.; Xie, H. Genotypic and Environmental Effects on the Volatile Chemotype of Valeriana Jatamansi Jones. Front. Plant Sci. 2018, 9, 1003. [Google Scholar] [CrossRef]
- Avoseh, O.N.; Mtunzi, F.M.; Ogunwande, I.A.; Ascrizzi, R.; Guido, F. Albizia Lebbeck and Albizia Zygia Volatile Oils Exhibit Anti-Nociceptive and Anti-Inflammatory Properties in Pain Models. Journal of Ethnopharmacology 2021, 268, 113676. [Google Scholar] [CrossRef]
- Ganesan, T.; Subban, M.; Christopher Leslee, D.B.; Kuppannan, S.B.; Seedevi, P. Structural Characterization of N-Hexadecanoic Acid from the Leaves of Ipomoea Eriocarpa and Its Antioxidant and Antibacterial Activities. Biomass Conv. Bioref. 2024, 14, 14547–14558. [Google Scholar] [CrossRef]
- Johannes, E.; Litaay, M.; Syahribulan, S. The Bioactivity of Hexadecanoic Acid Compound Isolated from Hydroid Aglaophenia Cupressina Lamoureoux as Antibacterial Agent against Salmonella Typhi. Int. J. Biol. Med. Res. 2016, 5469–5472. [Google Scholar]
- Sánchez-Sevilla, J.F.; Cruz-Rus, E.; Valpuesta, V.; Botella, M.A.; Amaya, I. Deciphering Gamma-Decalactone Biosynthesis in Strawberry Fruit Using a Combination of Genetic Mapping, RNA-Seq and eQTL Analyses. BMC Genomics 2014, 15, 218. [Google Scholar] [CrossRef]
- Chambers, A.H.; Evans, S.A.; Folta, K.M. Methyl Anthranilate and γ-Decalactone Inhibit Strawberry Pathogen Growth and Achene Germination. J. Agric. Food Chem. 2013, 61, 12625–12633. [Google Scholar] [CrossRef]
- Gong, X.; Sun, C.; Abame, M.A.; Shi, W.; Xie, Y.; Xu, W.; Zhu, F.; Zhang, Y.; Shen, J.; Aisa, H.A. Synthesis of CBD and Its Derivatives Bearing Various C4′-Side Chains with a Late-Stage Diversification Method. J. Org. Chem. 2020, 85, 2704–2715. [Google Scholar] [CrossRef] [PubMed]
- Jou, Y.-J.; Hua, C.-H.; Lin, C.-S.; Wang, C.-Y.; Wan, L.; Lin, Y.-J.; Huang, S.-H.; Lin, C.-W. Anticancer Activity of γ-Bisabolene in Human Neuroblastoma Cells via Induction of P53-Mediated Mitochondrial Apoptosis. Molecules 2016, 21, 601. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.K.; Ali, A.Y.; Hayward, O.A.; Turnham, D.; Jackson, T.; Bowen, I.D.; Clarkson, R. β-Bisabolene, a Sesquiterpene from the Essential Oil Extract of Opoponax ( Commiphora Guidottii ), Exhibits Cytotoxicity in Breast Cancer Cell Lines. Phytotherapy Research 2016, 30, 418–425. [Google Scholar] [CrossRef]
- Dahham, S.; Tabana, Y.; Iqbal, M.; Ahamed, M.; Ezzat, M.; Majid, A.; Majid, A. The Anticancer, Antioxidant and Antimicrobial Properties of the Sesquiterpene β-Caryophyllene from the Essential Oil of Aquilaria Crassna. Molecules 2015, 20, 11808–11829. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Tundis, R.; Menichini, F.; Saab, A.M.; Statti, G.A.; Menichini, F. Cytotoxic Activity of Essential Oils from Labiatae and Lauraceae Families against in Vitro Human Tumor Models. Anticancer Res 2007, 27, 3293–3299. [Google Scholar]
- Yoshihiro, I.; Toshiko, H.; Akiko, S.; Kazuma, H.; Hajime, H.; Shigeki, K. Biphasic Effects of Geranylgeraniol, Terpenone and Phytolon Thegrowth of Staphylococcus Aureus. 2005, 1770–1774.
- Sabudak, T.; Ozturk, M.; Goren, A.C.; Kolak, U.; Topcu, G. Fatty Acids and Other Lipid Composition of Five Trifolium Species with Antioxidant Activity. Pharmaceutical Biology 2009, 47, 137–141. [Google Scholar] [CrossRef]
- McGinty, D.; Letizia, C.S.; Api, A.M. Fragrance Material Review on Phytol. Food and Chemical Toxicology 2010, 48, S59–S63. [Google Scholar] [CrossRef]
- Demetzos, C.; Dimas, K.S. Labdane-Type Diterpenes: Chemistry and Biological Activity. In Studies in Natural Products Chemistry; Elsevier, 2001; Vol. 25, pp. 235–292 ISBN 978-0-08-044001-9.
- Tran, Q.T.N.; Wong, W.S.F.; Chai, C.L.L. Labdane Diterpenoids as Potential Anti-Inflammatory Agents. Pharmacological Research 2017, 124, 43–63. [Google Scholar] [CrossRef]
- Villamizar, Jo.E.; Juncosa, Jo.; Pittelaud, Je.; Hernández, M.; Canudas, N.; Tropper, E.; Salazar, F.; Fuentes, Ju. Facile Access to Labdane-Type Diterpenes: Synthesis of Coronarin C, Zerumin B, Labda-8(17), 13(14)-Dien-15,16-Olide and Derivatives from (+)-Manool. Journal of chemical research, 2007, 6, 342-346. 2007, 342–346.
- Cyr, A.; Wilderman, P.R.; Determan, M.; Peters, R.J. A Modular Approach for Facile Biosynthesis of Labdane-Related Diterpenes. J Am Chem Soc 2007, 129, 6684–6685. [Google Scholar] [CrossRef]
- Zhou, J.; Xie, X.; Tang, H.; Peng, C.; Peng, F. The Bioactivities of Sclareol: A Mini Review. Front. Pharmacol. 2022, 13, 1014105. [Google Scholar] [CrossRef]
- Jameel, S.; Bhat, K.A. Sclareol: Isolation, Structural Modification, Biosynthesis, and Pharmacological Evaluation – A Review. Pharm Chem J 2024, 57, 1568–1579. [Google Scholar] [CrossRef]
- Park, J.; Lee, K.; Jung, E.; Kang, S.; Kim, Y.J. Sclareol Isolated from Salvia Officinalis Improves Facial Wrinkles via an Antiphotoaging Mechanism. J of Cosmetic Dermatology 2016, 15, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Li, C.; Zhang, L.; Lin, Ji.; Jiang, N.; Wang, Qa.; Xu, Qi.; Zheng, H.; Gu, L.; Jia, Yi.; et al. Mechanism of Antifungal Activity and Therapeutic Action of β-Ionone on Aspergillus Fumigatus Keratitis via Suppressing LOX1 and JNK/P38 MAPK Activation. International Immunopharmacology 2022, 108992. [Google Scholar] [CrossRef]
- Kang, C.-H.; Jayasooriya, R.G.P.T.; Choi, Y.H.; Moon, S.-K.; Kim, W.-J.; Kim, G.-Y. β-Ionone Attenuates LPS-Induced pro-Inflammatory Mediators Such as NO, PGE2 and TNF-α in BV2 Microglial Cells via Suppression of the NF-κB and MAPK Pathway. Toxicology in Vitro 2013, 27, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Caboni, P.; Ntalli, N.G.; Aissani, N.; Cavoski, I.; Angioni, A. Nematicidal Activity of ( E, E )-2,4-Decadienal and ( E )-2-Decenal from Ailanthus Altissima against Meloidogyne Javanica. J. Agric. Food Chem. 2012, 60, 1146–1151. [Google Scholar] [CrossRef]
- Mirek, J.; Walkowiak-Nowicka, K.; Słocińska, M. The Effect of (E,E)-2,4-Decadienal, (E)-2-Decenal, 2-Undecanone and Furfural on Reproduction of Tenebrio Molitor. In Proceedings of the 1st International Electronic Conference on Entomology; MDPI: Sciforum.net, July 1 2021; p. 10540.
- Feng, Ye.; An, Qi.; Zhao, Zh.; Wu, M.; Yang, Ch.; Liang, W.Y.; Xu, Xu.; Jiang, T.; Zhang, Gu. Beta-Elemene: A Phytochemical with Promise as a Drug Candidate for Tumor Therapy and Adjuvant Tumor Therapy. Biomedicine & Pharmacotherapy 2024, 116266.
- Chen, X.; Huang, C.; Li, K.; Liu, J.; Zheng, Y.; Feng, Y.; Kai, G. Recent Advances in Biosynthesis and Pharmacology of β-Elemene. Phytochem Rev 2023, 22, 169–186. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, R.; Xu, L.; Xie, S.; Dong, J.; Jing, Y. β-Elemene Piperazine Derivatives Induce Apoptosis in Human Leukemia Cells through Downregulation of c-FLIP and Generation of ROS. PLoS ONE 2011, 6, e15843. [Google Scholar] [CrossRef]
- Ricciardelli, A.; Casillo, A.; Papa, R.; Monti, D.M.; Imbimbo, P.; Vrenna, G.; Artini, M.; Selan, L.; Corsaro, M.M.; Tutino, M.L.; et al. Pentadecanal Inspired Molecules as New Anti-Biofilm Agents against Staphylococcus Epidermidis. Biofouling 2018, 34, 1110–1120. [Google Scholar] [CrossRef]
- Casillo, A.; Papa, R.; Ricciardelli, A.; Sannino, F.; Ziaco, M.; Tilotta, M.; Selan, L.; Marino, G.; Corsaro, M.M.; Tutino, M.L.; et al. Anti-Biofilm Activity of a Long-Chain Fatty Aldehyde from Antarctic Pseudoalteromonas Haloplanktis TAC125 against Staphylococcus Epidermidis Biofilm. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef]
- Labbozzetta, M.; Poma, P.; Tutone, M.; McCubrey, J.A.; Sajeva, M.; Notarbartolo, M. Phytol and Heptacosane Are Possible Tools to Overcome Multidrug Resistance in an In Vitro Model of Acute Myeloid Leukemia. Pharmaceuticals 2022, 15, 356. [Google Scholar] [CrossRef]
- Mrabti, H.N.; Jaouadi, I.; Zeouk, I.; Ghchime, R.; El Menyiy, N.; Omari, N.E.; Balahbib, A.; Al-Mijalli, S.H.; Abdallah, E.M.; El-Shazly, M.; et al. Biological and Pharmacological Properties of Myrtenol: A Review. CPD 2023, 29, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Barbhuiya, P.A.; Pathak, M.P. Myrtenol: A Promising Terpene with Potent Pharmacological Properties. Pharmacological Research - Natural Products 2024, 4, 100067. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Lee, H.-S. Verbenone Structural Analogues Isolated from Artemesia Aucheri as Natural Acaricides against Dermatophagoides Spp. and Tyrophagus Putrescentiae. J. Agric. Food Chem. 2013, 61, 12292–12296. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-Y.; Lee, H.-W.; Lee, H.-S. Growth Inhibitory Activities of Myrtanol and Structural Analogues from Thymus Tosevii against Intestinal Bacteria. Food Sci Biotechnol 2015, 24, 169–174. [Google Scholar] [CrossRef]
- Cha, D.H.; Roh, G.H.; Hesler, S.P.; Wallingford, A.; Stockton, D.G.; Park, S.K.; Loeb, G.M. 2-Pentylfuran: A Novel Repellent of Drosophila Suzukii. Pest Management Science 2021, 77, 1757–1764. [Google Scholar] [CrossRef]
- Kladi, M.; Vagias, C.; Furnari, G.; Moreau, D.; Roussakis, C.; Roussis, V. Cytotoxic Cuparene Sesquiterpenes from Laurencia Microcladia. Tetrahedron Letters 2005, 46, 5723–5726. [Google Scholar] [CrossRef]
- Pajaro-Castro, N.; Flechas, M.C.; Ocazionez, R.; Stashenko, E.; Olivero-Verbel, J. Potential Interaction of Components from Essential Oils with Dengue Virus Proteins. Boletin Latinoamericano y del Caribe de plantas Medicinales y Aromaticas 2015, 14, 141–155. [Google Scholar]
- Pan, S.-P.; Pirker, T.; Kunert, O.; Kretschmer, N.; Hummelbrunner, S.; Latkolik, S.L.; Rappai, J.; Dirsch, V.M.; Bochkov, V.; Bauer, R. C13 Megastigmane Derivatives From Epipremnum Pinnatum: β-Damascenone Inhibits the Expression of Pro-Inflammatory Cytokines and Leukocyte Adhesion Molecules as Well as NF-κB Signaling. Front. Pharmacol. 2019, 10, 1351. [Google Scholar] [CrossRef]
- Uddin, A.N.; Labuda, I.; Burns, F.J. A Novel Mechanism of Filaggrin Induction and Sunburn Prevention by β-Damascenone in Skh-1 Mice. Toxicology and Applied Pharmacology 2012, 265, 335–341. [Google Scholar] [CrossRef]
Figure 1.
Yield of essential oil (mL/kg (d/w)) in different parts of G. capitata.
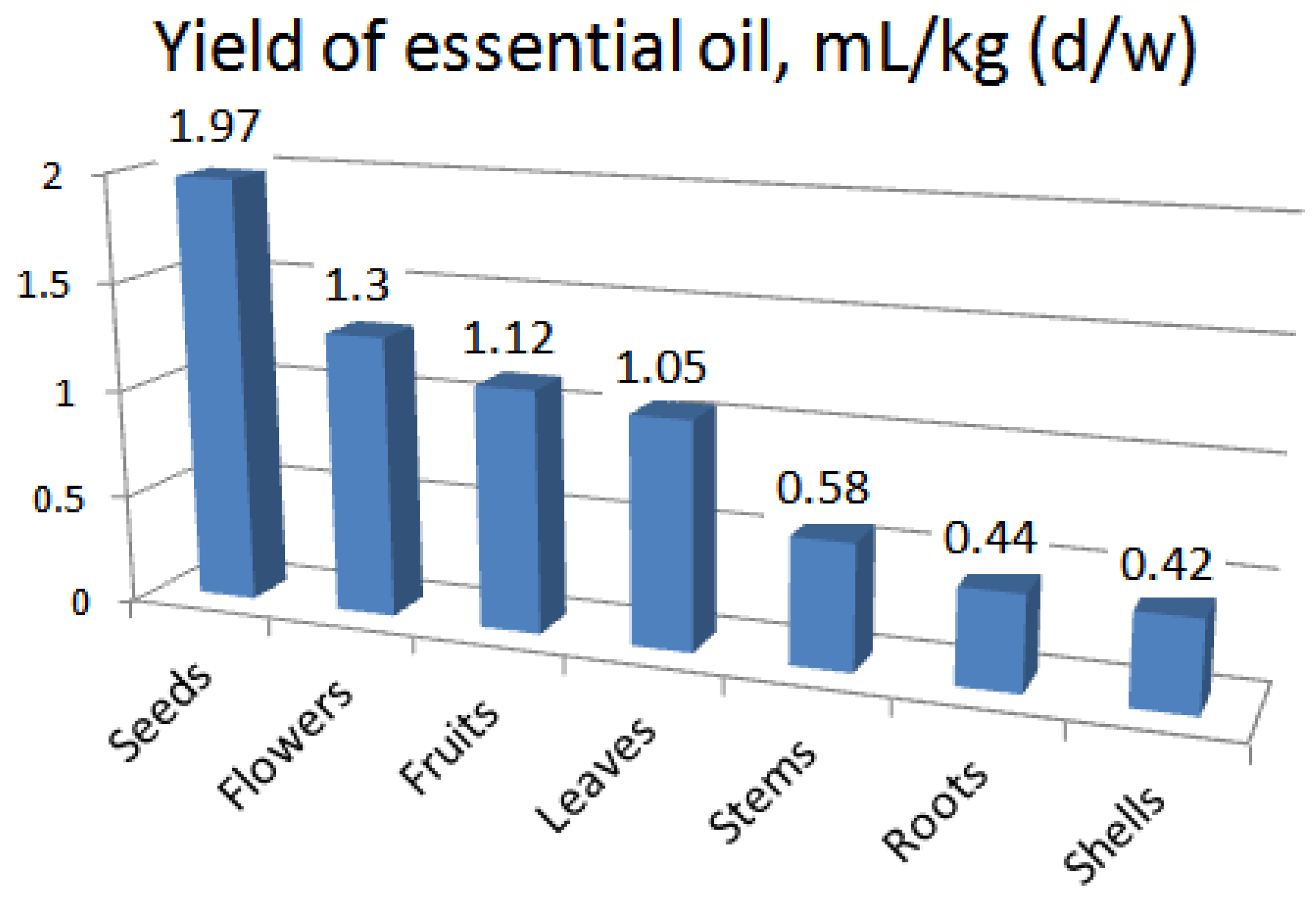
Figure 2.
Cluster tree characterizing the essential oil composition between different parts of G. capitata.
Figure 2.
Cluster tree characterizing the essential oil composition between different parts of G. capitata.
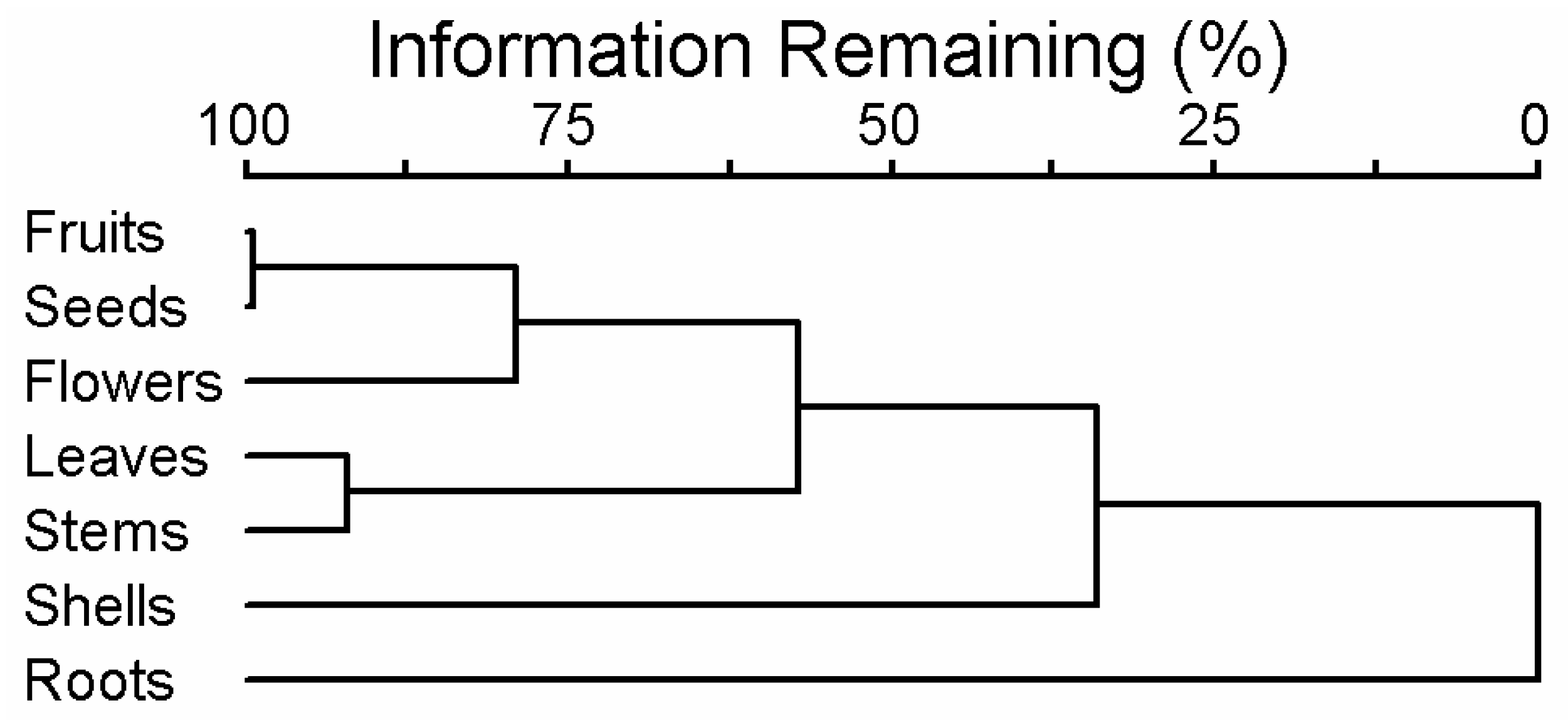
Figure 3.
Dominant substances (%) in different part of G. capitata. Compounds with at least 5% are presented.
Figure 3.
Dominant substances (%) in different part of G. capitata. Compounds with at least 5% are presented.
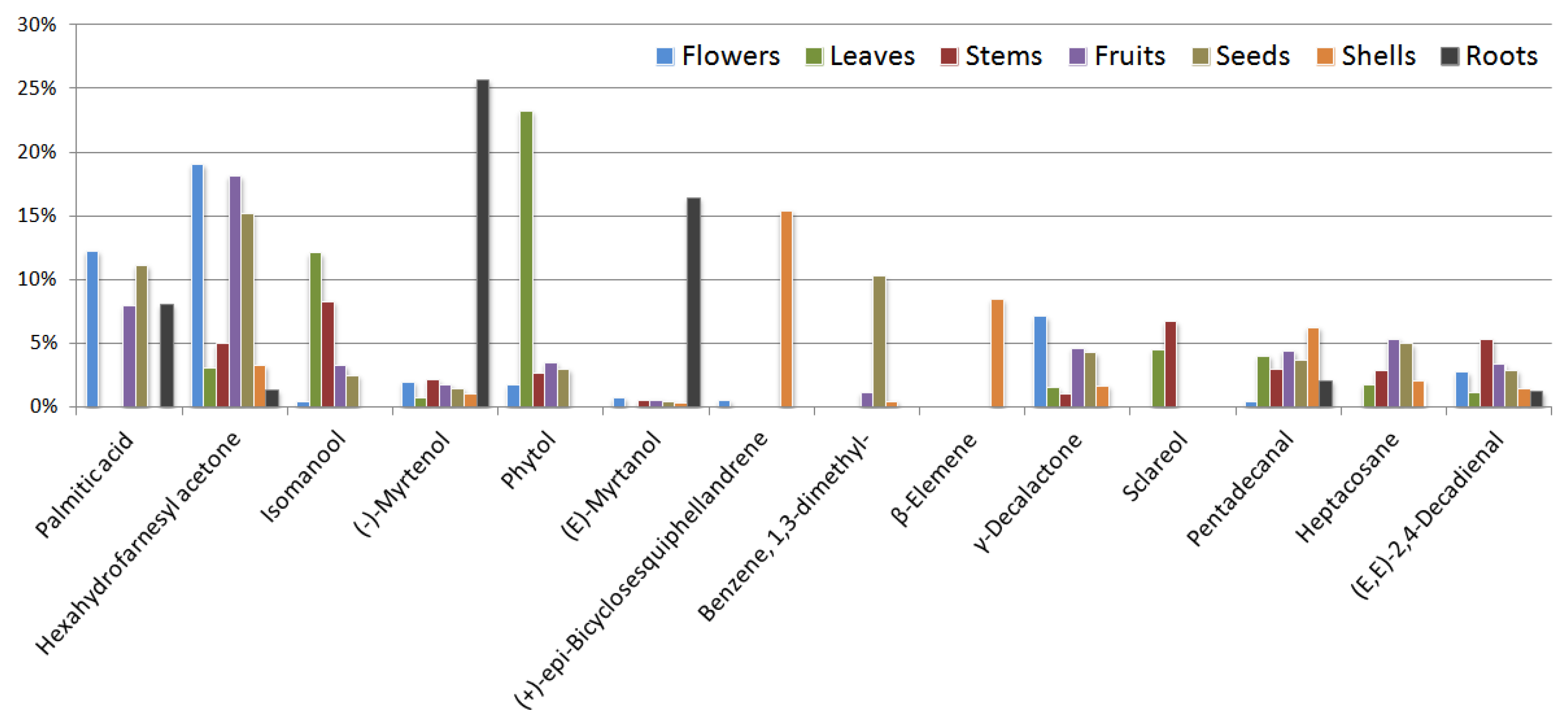
Figure 4.
Characteristics terpenoids of Gilia capitata Sims.
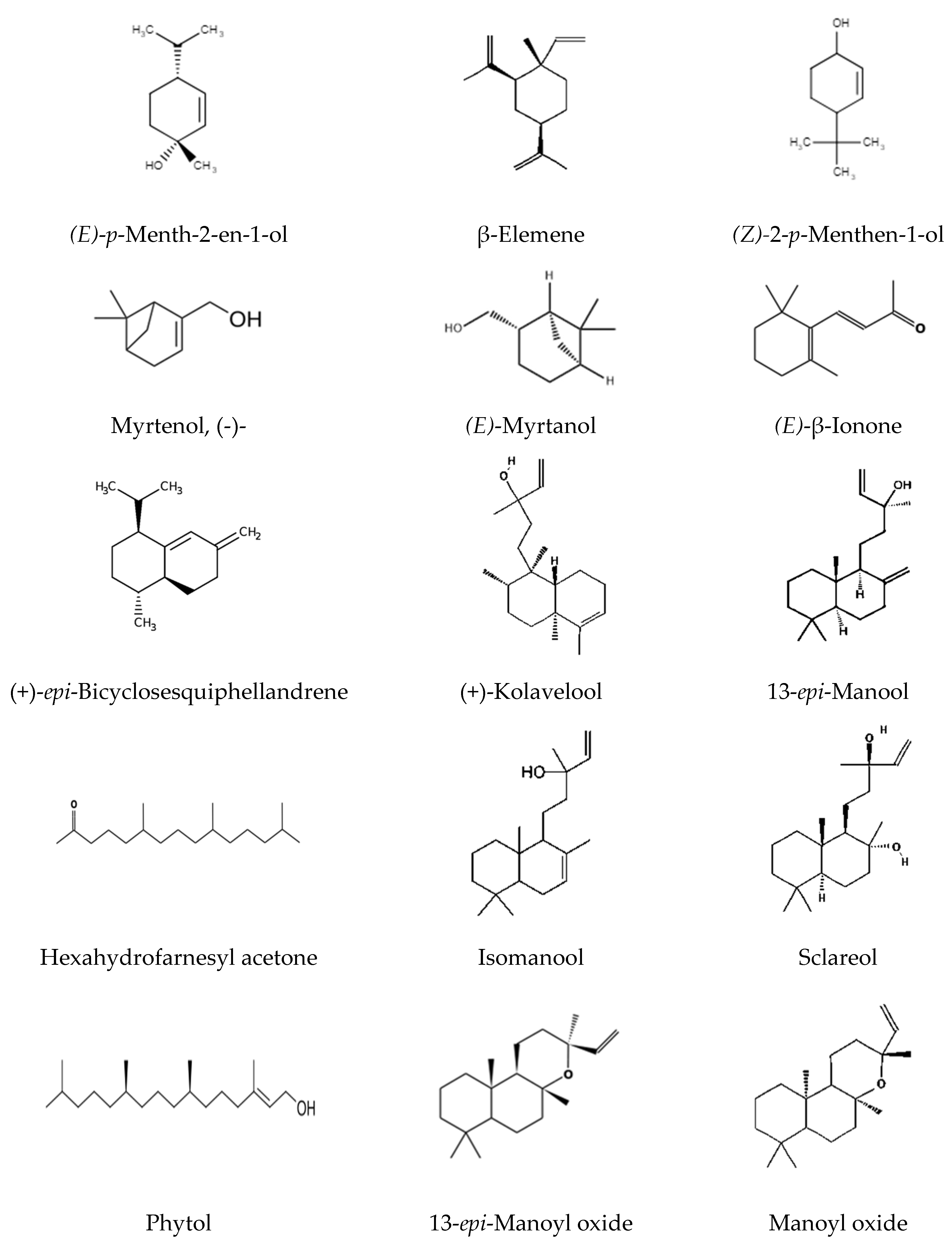
Figure 5.
The content (%) group compounds in volatile components of different parts of G. capitata. The total % from Table 1 has been rescaled to 100%.
Figure 5.
The content (%) group compounds in volatile components of different parts of G. capitata. The total % from Table 1 has been rescaled to 100%.
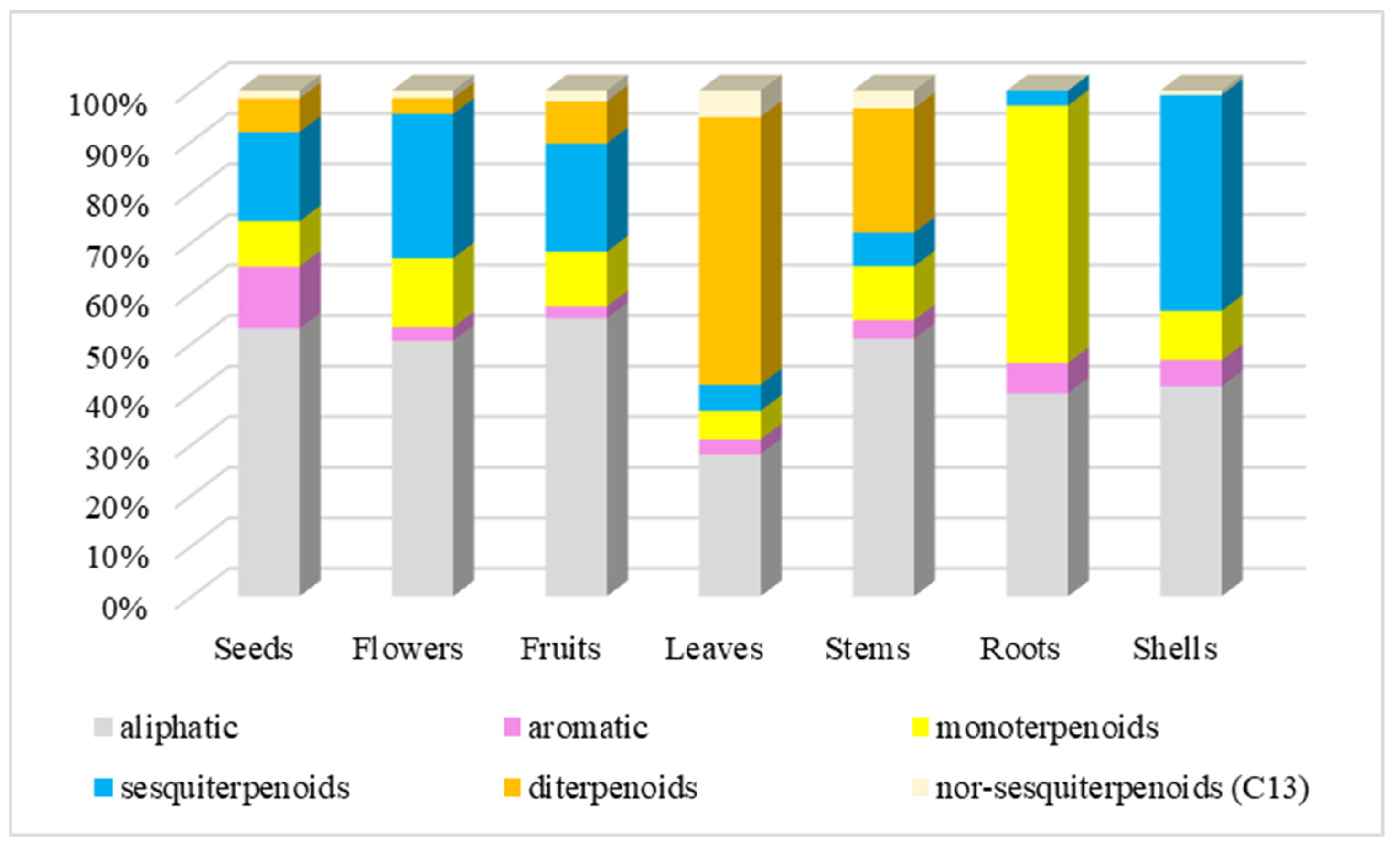

Table 1.
The content (%) of volatile compounds (>0.1%) in different parts of G. capitata.
| Compound | RI | Library NIST23 | Content (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Flowers | Leaves | Stems | Fruits | Shells | Seeds | Roots | |||
| Hexanal | 799 | 798 | 3.35 | 1.42 | 3.20 | nd | 4.49 | 0.20 | 2.31 |
| (E)-2-Hexenal | 848 | 848 | 0.88 | 2.09 | 0.43 | nd | nd | nd | nd |
| 1-Hexanol | 864 | 864 | 2.69 | 0.28 | nd | 0.19 | 0.93 | 0.04 | 0.08 |
| 1,3-d’Dimethyl-benzene, | 866 | 866 | nd | nd | nd | 1.11 | 0.44 | 10.27 | nd |
| (E,Z)-4-Ethylidenecyclohexene | 876 | 877 | 0.36 | nd | 0.07 | nd | nd | nd | nd |
| 2-Heptanone | 890 | 889 | 0.29 | 0.07 | 0.13 | nd | 0.74 | nd | 0.08 |
| Nonane | 900 | 900 | 0.28 | nd | 0.19 | nd | nd | nd | nd |
| Heptanal | 902 | 901 | 0.26 | 0.17 | nd | nd | 0.50 | nd | 0.28 |
| (E)-2-Heptenal | 954 | 954 | 0.17 | 0.23 | 3.23 | nd | 0.46 | nd | 0.15 |
| Benzaldehyde | 958 | 958 | 0.28 | 0.41 | 0.99 | nd | 0.41 | nd | 1.03 |
| 1-Heptanol | 968 | 968 | 0.05 | nd | nd | nd | 0.14 | nd | nd |
| 1-Octen-3-ol | 978 | 978 | 0.14 | nd | 1.65 | 0.21 | 0.46 | 0.19 | 0.22 |
| 2,3-Octanedione | 983 | 983 | 0.26 | nd | 0.34 | nd | nd | nd | 0.14 |
| 6-Methyl-5-hepten-2-one | 986 | 986 | 0.11 | nd | 0.10 | nd | nd | nd | 0.09 |
| 2-Pentylfuran | 991 | 991 | 2.90 | 0.76 | 1.37 | nd | 1.20 | nd | 2.89 |
| (Z)-2-(2-Pentenyl)furan | 1001 | 1001 | 0.18 | nd | 0.21 | nd | nd | nd | 0.20 |
| Octanal | 1003 | 1002 | 0.37 | 0.53 | 0.30 | nd | nd | nd | 0.26 |
| (E,E)-2,4-Heptadienal, | 1010 | 1010 | 0.17 | 0.34 | 0.74 | nd | nd | nd | nd |
| α-Terpinene | 1016 | 1016 | 0.40 | nd | nd | nd | nd | nd | nd |
| p-Cymene | 1023 | 1023 | 0.12 | nd | nd | nd | nd | nd | nd |
| 2-Ethyl-1-hexanol | 1028 | 1027 | nd | nd | nd | nd | nd | nd | 0.49 |
| β-Phellandrene | 1028 | 1028 | 0.67 | nd | nd | nd | nd | nd | nd |
| 3-Octen-2-one | 1038 | 1038 | 0.13 | nd | 1.22 | nd | nd | nd | 0.09 |
| Benzeneacetaldehyde | 1042 | 1042 | 0.65 | 1.27 | 0.98 | 0.39 | 2.45 | 0.31 | 1.43 |
| (E)-2-Octenal | 1057 | 1056 | 0.80 | nd | 2.11 | nd | 0.62 | nd | 1.04 |
| γ-Terpinene | 1057 | 1057 | 0.47 | nd | nd | nd | nd | nd | nd |
| Acetophenone | 1065 | 1064 | nd | nd | nd | nd | 0.12 | nd | 0.10 |
| (Z)-2-Octen-1-ol | 1066 | 1067 | nd | nd | 0.51 | nd | nd | nd | nd |
| 1-Octanol | 1069 | 1069 | 2.89 | 2.02 | nd | 2.53 | 0.88 | 2.26 | 0.37 |
| (E)-β-Terpinolene | 1087 | 1087 | 0.16 | nd | nd | nd | nd | nd | nd |
| 3,5-Octadien-2-one | 1092 | 1093 | 0.25 | nd | 0.49 | nd | nd | nd | 0.12 |
| Linalool | 1098 | 1098 | 0.34 | 0.50 | 0.28 | nd | 0.58 | 0.29 | nd |
| Nonanal | 1103 | 1102 | 2.17 | 1.25 | 1.49 | 1.96 | 1.18 | 1.78 | 1.38 |
| (Z)-2-p-Menthen-1-ol | 1119 | 1120 | 3.62 | 1.43 | 3.82 | 3.28 | 1.98 | 2.70 | nd |
| (E)-p-2-Menthen-1-ol | 1138 | 1138 | 2.07 | 0.81 | 2.14 | 2.36 | 1.32 | 1.91 | nd |
| (E)-Verbenol | 1143 | 1143 | 0.20 | nd | nd | nd | nd | nd | nd |
| (R,S)-5-Ethyl-6-methyl-3E-hepten-2-one | 1145 | 1145 | nd | nd | 0.41 | nd | nd | nd | 0.19 |
| (E,Z)-2,6-Nonadienal | 1152 | 1152 | 0.27 | 0.35 | 0.47 | nd | nd | nd | 0.18 |
| (E)-2-Nonenal | 1158 | 1158 | 0.94 | 0.44 | 0.92 | 0.46 | 0.57 | 0.42 | 0.95 |
| α-Phellandren-8-ol | 1165 | 1165 | 0.08 | 0.03 | nd | 0.03 | 0.09 | nd | 0.05 |
| 1-Nonanol | 1170 | 1170 | nd | 0.15 | 0.56 | 0.32 | 0.70 | 0.29 | 0.22 |
| L-α-Terpineol | 1189 | 1189 | 0.39 | nd | nd | nd | nd | nd | nd |
| (-)-Myrtenol | 1195 | 1195 | 1.94 | 0.67 | 2.12 | 1.73 | 0.97 | 1.44 | 25.72 |
| Decanal | 1204 | 1204 | 1.06 | 1.16 | 2.21 | 1.34 | nd | 1.03 | 1.89 |
| (E)-Piperitol | 1206 | 1206 | 1.21 | nd | 1.05 | 1.63 | 1.39 | 1.31 | nd |
| (E,E)-2,4-Nonadienal | 1212 | 1212 | nd | nd | 0.74 | 0.25 | nd | 0.23 | 0.39 |
| Bicyclo [3.3.0]octan-2-one, 7-methylene-6(or 8)-methyl- | 1220 | 1220 | nd | nd | nd | nd | nd | nd | 2.20 |
| Carvone | 1244 | 1243 | nd | nd | nd | nd | nd | nd | 1.47 |
| p-Mentha-1(7),8(10)-dien-9-ol | 1245 | 1246 | nd | nd | nd | nd | nd | nd | 1.19 |
| Geraniol | 1253 | 1254 | nd | nd | nd | nd | nd | nd | 1.01 |
| (E)-Myrtanol | 1258 | 1258 | 0.70 | nd | 0.47 | 0.54 | 0.34 | 0.43 | 16.44 |
| 1-Decanol | 1270 | 1271 | 0.46 | 0.19 | nd | 0.64 | 0.88 | 0.53 | nd |
| Nonanoic acid | 1271 | 1272 | 1.04 | nd | nd | nd | nd | nd | nd |
| (E)-Bornyl acetate | 1285 | 1285 | nd | nd | nd | nd | 0.73 | nd | nd |
| Thymol | 1289 | 1290 | nd | nd | nd | nd | nd | nd | 0.55 |
| (E,Z)-2,4-Decadienal | 1292 | 1292 | 0.77 | nd | 1.61 | 0.87 | nd | 0.71 | 0.32 |
| (E)-Undec-4-enal | 1298 | 1296 | nd | nd | 0.33 | nd | nd | nd | nd |
| Undecanal | 1306 | 1305 | 0.19 | 0.25 | 0.33 | 0.43 | nd | 0.39 | 0.32 |
| 2-Methoxy-4-vinylphenol | 1312 | 1312 | 0.41 | 0.51 | nd | nd | 0.53 | nd | nd |
| (E,E)-2,4-Decadienal | 1315 | 1315 | 2.71 | 1.10 | 5.29 | 3.36 | 1.43 | 2.85 | 1.18 |
| Eugenol | 1357 | 1357 | nd | 0.97 | nd | nd | nd | nd | nd |
| Dihydro-5-pentyl-2(3H)-furanone | 1362 | 1362 | nd | nd | nd | nd | nd | nd | 0.25 |
| 2-Undecenal | 1363 | 1363 | nd | nd | 3.89 | 1.26 | 0.83 | 1.12 | nd |
| n-Decanoic acid | 1373 | 1372 | nd | nd | nd | nd | nd | nd | 1.32 |
| 2-Butyl-2-octenal | 1372 | 1373 | 0.34 | nd | nd | 0.32 | nd | 0.26 | nd |
| (E)-β-Damascenone | 1385 | 1385 | 0.27 | nd | nd | 0.58 | 0.72 | 0.45 | nd |
| β-Elemene | 1393 | 1393 | nd | nd | nd | nd | 8.46 | nd | nd |
| 6,10-Dimethyl-2-undecanone, | 1404 | 1404 | nd | nd | 0.25 | nd | nd | nd | nd |
| Dodecanal | 1408 | 1408 | 0.29 | 0.26 | 0.28 | 0.36 | nd | 0.30 | 0.50 |
| 7-epi-α-Cedrene | 1416 | 1417 | nd | nd | nd | nd | nd | nd | 0.56 |
| Copaene | 1421 | 1420 | 1.28 | nd | nd | nd | 2.87 | nd | nd |
| Caryophyllene | 1421 | 1421 | 1.34 | 0.54 | nd | nd | 2.77 | nd | nd |
| (E)-Geranylacetone | 1453 | 1453 | 0.54 | 0.85 | 0.52 | 0.72 | nd | 0.63 | 0.86 |
| Humulene | 1455 | 1456 | nd | nd | nd | nd | 1.52 | nd | nd |
| γ-Decalactone | 1469 | 1469 | 7.09 | 1.50 | 1.04 | 4.62 | 1.66 | 4.23 | nd |
| 1-Dodecanol | 1474 | 1474 | nd | 0.20 | 0.13 | nd | 0.43 | nd | 0.94 |
| (+)-epi-Bicyclosesquiphellandrene | 1484 | 1484 | 0.48 | nd | nd | nd | 15.41 | nd | nd |
| (E)-β-Ionone | 1488 | 1488 | 1.14 | 4.54 | 3.24 | 1.30 | nd | 0.97 | nd |
| β-Selinene | 1488 | 1489 | nd | nd | nd | nd | 0.41 | nd | nd |
| α-(3-Methylbutylidene)- benzeneacetaldehyde | 1492 | 1492 | nd | nd | 0.21 | nd | nd | nd | nd |
| (+)-Cuparene | 1510 | 1508 | nd | nd | nd | nd | nd | nd | 0.29 |
| α-Farnesene | 1510 | 1510 | 1.01 | nd | nd | nd | nd | nd | nd |
| Tridecanal | 1511 | 1511 | nd | 0.53 | 0.34 | 0.53 | nd | 0.44 | 0.29 |
| (Z)-γ-Bisabolene | 1517 | 1518 | 0.57 | nd | nd | nd | nd | nd | nd |
| n-Tridecan-1-ol | 1575 | 1575 | nd | nd | nd | nd | nd | nd | 0.25 |
| Caryophyllene oxide | 1587 | 1587 | 0.89 | nd | nd | nd | nd | nd | nd |
| 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | 1600 | 1599 | 0.38 | nd | nd | nd | nd | nd | 0.92 |
| Tetradecanal | 1613 | 1613 | 0.24 | 0.76 | 0.54 | 1.03 | 1.55 | 0.88 | 0.50 |
| Benzophenone | 1629 | 1629 | nd | 0.20 | 0.14 | nd | nd | nd | nd |
| τ-Muurolol | 1644 | 1646 | 0.62 | nd | nd | nd | nd | nd | nd |
| α-Cadinol | 1658 | 1658 | 0.41 | nd | nd | nd | nd | nd | nd |
| 1-Tetradecanol | 1677 | 1677 | nd | nd | 0.49 | 1.01 | 1.81 | 0.97 | 1.52 |
| Pentadecanal | 1715 | 1715 | 0.37 | 4.01 | 2.99 | 4.38 | 6.25 | 3.65 | 2.06 |
| 1-Tetradecene | 1736 | 1736 | nd | nd | 0.65 | 0.54 | nd | 0.50 | nd |
| Myristic acid | 2765 | 1764 | nd | nd | nd | nd | nd | nd | 0.85 |
| n-Pentadecanol | 1779 | 1778 | nd | nd | nd | 0.43 | 1.63 | 0.44 | 0.38 |
| Ambrial | 1804 | 1804 | nd | nd | 0.48 | nd | nd | nd | nd |
| Farnesyl acetaldehyde | 1845 | 1845 | nd | nd | 0.25 | nd | nd | nd | nd |
| Hexahydrofarnesyl acetone | 1851 | 1851 | 19.09 | 3.10 | 4.94 | 18.16 | 3.24 | 15.21 | 1.27 |
| Di-2-methylpropyl phthalate | 1878 | 1878 | 1.58 | 0.64 | 0.72 | 0.55 | 0.81 | 0.42 | 2.61 |
| 1-Hexadecanol | 1889 | 1889 | nd | 1.05 | 0.78 | 1.47 | 0.83 | 1.55 | 0.74 |
| Manoyl oxide | 1896 | 1897 | nd | 2.28 | 2.96 | 0.62 | nd | 0.51 | nd |
| Roughanic acid | 1904 | 1904 | nd | nd | 0.70 | nd | nd | nd | nd |
| Farnesyl acetone | 1928 | 1928 | 0.50 | nd | nd | nd | nd | nd | nd |
| Cembrene A | 1969 | 1969 | 0.82 | nd | nd | nd | nd | nd | nd |
| Palmitic acid | 1976 | 1974 | 12.24 | nd | nd | 7.95 | nd | 11.13 | 8.05 |
| 13-epi-Manoyl oxide | 2018 | 2018 | nd | 1.15 | 0.51 | nd | nd | nd | nd |
| Epi-13-Manool | 2061 | 2061 | nd | 1.01 | 0.77 | nd | nd | nd | nd |
| Kolavelool | 2070 | 2070 | nd | 1.67 | 0.97 | nd | nd | nd | nd |
| Isomanool | 2093 | 2094 | 0.39 | 12.07 | 8.29 | 3.23 | nd | 2.48 | nd |
| γ-Palmitolactone | 2103 | 2104 | nd | nd | 0.41 | nd | nd | nd | nd |
| Phytol | 2116 | 2114 | 1.72 | 23.26 | 2.63 | 3.51 | nd | 2.97 | nd |
| Sclareol | 2225 | 2227 | nd | 4.45 | 6.70 | nd | nd | nd | nd |
| Nonadecane | 2300 | 2300 | 0.29 | 0.53 | 0.17 | 1.95 | nd | 1.71 | nd |
| Pentacosane | 2500 | 2500 | nd | 0.99 | 0.64 | 3.34 | nd | 3.10 | nd |
| Bis(2-ethylhexyl) phthalate | 2551 | 2550 | nd | nd | 0.38 | nd | nd | nd | nd |
| Heptacosane | 2700 | 2701 | nd | 1.74 | 2.84 | 5.34 | 2.07 | 4.95 | nd |
| Octacosane | 2800 | 2800 | nd | nd | 0.52 | 1.47 | 1.51 | 1.47 | nd |
| TOTAL | 93.74 | 86.73 | 92.87 | 88.30 | 81.31 | 89.92 | 91.18 | ||
Note. “nd” – not detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



