
Submitted:
05 February 2025
Posted:
06 February 2025
You are already at the latest version
Abstract
A combination of morphological analysis and DNA barcoding (partial sequences of the cytochrome c oxidase I (COI) gene) was used to differentiate four gobiid fish species in the family Oxudercidae. Rhinogobius chiengmaiensis and Rhinogobius mekongianus were found in Thailand, alongside Eugnathogobius siamensis and Pseudogobiopsis oligactis. Morphological identification relied on appearances, counts, and measurements. The 707-base pair COI sequences from eleven samples of four gobiid species were compared with reference sequences in public databases to confirm their scientific names. The average AT content was 51.8 ± 0.5% and the GC content was 48.2 ± 0.5%. Intraspecific genetic distances ranged from 0.00% to 0.28%, while interspecific genetic distances ranged from 0.86% to 16.63%. A maximum likelihood (ML) phylogenetic tree depicted the relationships among the COI sequences of these species. Morphological analysis and COI sequences successfully identified the four gobiid species. Notably, the COI sequences of R. chiengmaiensis, R. mekongianus, and E. siamensis were previously unreported, hence, this study is the first report to add their sequences to public databases. These results can serve as valuable information for the management of aquatic resources, conservation, and aquaculture efforts.
Keywords:
morphological identification
; DNA barcoding
; cytochrome c oxidase I
; species identification
; Rhinogobius
; Oxudercidae
1. Introduction
The family Oxudercidae (or Gobionellidae) belongs to the order Gobiiformes, and comprises approximately 86 genera and 598 species that are found worldwide from freshwater to marine habitats [1]. Fish species in this family were previously classified in the subfamily Oxudercinae of the family Gobiidae according to their taxonomic arrangement following Nelson [2]. Later, when the taxonomy of fish in this taxa was verified, it was revised that the subfamily Oxudercinae was separated from the family Gobiidae with the status of the family Oxudercidae [1]. One distinguishing feature of the family Oxudercidae, as compared to the family Gobiidae, is the presence of elongated and slender suspensoria structures [1,3].
Rhinogobius, a genus within the family Oxudercidae, contains the most species of freshwater gobies found in the lakes and streams across East and Southeast Asia, including Thailand [4,5,6]. Rhinogobius species exhibit characteristics that are commonly observed in gobies, such as their small body size, the fusion of their pelvic fins to form a disc-like structure on the ventral side, and an elongated and rounded body [7,8]. Moreover, this genus exhibits an elongated form of the head, characterized by a long snout. Presently, the genus Rhinogobius is recognized to encompass more than 80 distinct species, and ongoing taxonomic research is consistently discovering and documenting new species such as R. sangenloensis in Southern China [5], R. maxillivirgatus in Eastern China [9], R. yangminshanensis in Taiwan [10], and R. aonumai aonumai and R. aonumai ishigakiensis in Japan [11]. In addition, two new species, R. rong and R. nami, have recently been described from central Vietnam [12]. However, Panitvong [13] notes that only three Rhinogobius species are currently recognized in Thailand: R. chiengmaiensis, R. mekongianus, and R. giurinus.
Rhinogobius chiengmaiensis exhibits a broad distribution in the upper region of the Chao Phraya river, whereas R. mekongianus is found inhabiting the Mekong river basin [14]. In 2013, Panitvong [13] found R. giurinus within a stream located in Chiang Rai province, Northern Thailand. Nevertheless, there remains limited documentation of the presence of this species in Thailand. Therefore, this study focused on two species, R. chiengmaiensis, and R. mekongianus. Although these two species are found in different natural habitats, they share some similarities in appearance, such as small size, elongated body shape, and the pattern of colors. They are currently becoming popular as ornamental fish, causing the natural population trend to decrease, especially in the case of R. chiengmaiensis. The conservation statuses of R. chiengmaiensis and R. mekongianus were assessed by the International Union for Conservation of Nature (IUCN) as vulnerable (VU) and least concern (LC), respectively (as of February 2025). Hence, both the morphological and molecular identification methods were required to distinguish between these two species.
The morphological identification of fish species involves the assessment of meristic and morphometric characteristics. Meanwhile, the molecular identification, DNA barcoding or the standard sequences of the cytochrome c oxidase I (COI) gene have been commonly employed for global species bioidentification [15], including the case of fishes [16,17,18,19,20]. Both morphological identification and DNA barcoding have distinct advantages and disadvantages. Morphological identification is typically conducted by experienced fish taxonomists, while DNA barcoding is employed for identifying fish species even in cases of incomplete samples such as fish fillets [21], or the various developmental stages such as the eggs [22] or larval stage [23]. Collaboration between morphological identification and DNA barcoding enhances the accuracy of fish species identification and contributes to the expansion of the COI gene sequences in public databases for further analysis.
The objective of this research was to differentiate between two species of the genus Rhinogobius, namely R. chiengmaiensis and R. mekongianus, through the application of both morphological identification and DNA barcoding techniques. Furthermore, the research also focused on studying two other species within the family Oxudercidae, namely Eugnathogobius siamensis and Pseudogobiopsis oligactis. These two species were analyzed in comparison to two species of the genus Rhinogobius. This research provided the first report of the COI sequences of R. chiengmaiensis, R. mekongianus, and E. siamensis. The COI sequences of all four species were expanded and deposited into the GenBank database to serve as reference sequences for aiding in the identification of unknown species and establishing a database for specifying of new species within the family Oxudercidae, especially the genus Rhinogobius.
2. Materials and Methods
2.1. Sample Collection
Seven live samples of two species belonging to the genus Rhinogobius, R. chiengmaiensis and R. mekongianus were obtained from collectors of fish from nature with known sources (Table 1). In addition, four samples of two other species of the family Oxudercidae, E. siamensis and P. oligactis, were also received. All samples were photographed and anesthetized with 0.2 g/L of MS-222 (Sigma, Missouri, USA) solution. Whole samples were preserved in absolute ethanol at room temperature for morphology identification. This animal use protocol has been approved by the Institutional Animal Care and Use Committee, University of Phayao (No. UP-AE 1-011-67).
2.2. Morphological Identification
All samples were first identified based on morphological appearances including body shape, mouth and eye size. Counts and measurements following Nagao Natural Environment Foundation [24] were performed on the left side of the samples. Fin rays were counted for the dorsal, pectoral, pelvic, and anal fins. The longitudinal, predorsal, and circumcaudal scales were also recorded. Measurements were taken point to point with digital vernier calipers to the nearest 0.01 mm. Subunits of the head were presented as proportions of head length (HL). Measurements of head and body parts were provided as proportions of standard length (SL).
2.3. Molecular Identification
After morphological identification, a small piece of right pectoral fin from each sample was used for the extraction of the genomic DNA using the GF-1 Nucleic Acid Extraction kit (Vivantis Technologies Sdn. Bhd., Selangor Darul Ehsan, Malaysia). The quantity and quality of the extracted DNA was determined using a NanoDrop Microvolume Spectrophotometer (Thermo Fisher Scientific Inc., MA, USA) and 1.2% agarose gel electrophoresis at 100 V for 30 min, respectively.
A 707-bp fragment of the COI gene was amplified for each sample using the equal volume of each 10 µM primer, namely FishF1 (5´-TCAACCAACCACAAAGACATTGGCAC-3´), Fish F2 (5´-TCGACTAATCATAAAGATATCGGCAC-3´), FishR1 (5´-TAGACTTCTGGGTGGCCAAAGAATCA-3´), and FishR2 (5´-ACTTCAGGGTGACCGAAGAATCAGAA-3´) [20], with the PCR technique. Each PCR reaction mixture included of 1 µl of extracted DNA (approximately 100 ng/µl), 2.25 µl of 10x Taq buffer, 0.8 µl of 50 mM MgCl2, 1 µl of 2.5 mM dNTP mix, 1 µl of 10 µM mixed primers, 0.2 µl of 1 U Taq DNA polymerase (BIO-HELIX Co., Ltd., New Taipei City, Taiwan), and nuclease-free water to 25 µl. The thermal cycling conditions included an initial denaturation at 95 °C for 2 min followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 54 °C for 30 s, and extension at 72 °C for 1 min, with a final extension at 72 °C for 10 min. The PCR products were analyzed using 1.2% agarose gel electrophoresis stained with Novel Juice (BIO-HELIX Co., Ltd., New Taipei City, Taiwan). The target bands were then purified to remove unwanted reagents using the HiYield™ Gel/PCR DNA Fragments Extraction kit (RBC Bioscience Corp., New Taipei City, Taiwan), following the manufacturer's instructions.
The purified PCR products were sequenced bidirectionally at ATGC Co., Ltd. (Thailand) using primer FishF1/FishF2 for the 3' end and FishR1/FishR2 for the 5' end. Two sequences of each sample were aligned and assembled using the Clustal Omega program [25]. All sequences covering the primer sequences were searched for the stop codon, deletion, and insertion using the Open Reading Frame Finder website (ORF finder; https://www.ncbi.nlm.nih.gov/orffinder/). The scientific name of each sample was investigated by comparing its sequence with reference sequences in two public databases, namely the GenBank (https://www.ncbi.nlm.nih.gov/) using the Basic Local Alignment Search Tool (BLAST) program [26] and BOLD (https://boldsystems.org/). Each sample was assigned a scientific name based on the top match with a minimum of 99% sequence similarity.
The percentage of base composition of the four gobiid species was calculated using the MEGA11 program [27]. The intraspecific and interspecific genetic distances were calculated using the MEGA11 program with the Kimura-2-parameter (K2P) distance model [28]. Because the COI sequences of R. chiengmaiensis, R. mekongianus, and E. siamensis have not been reported in any databases, several sequences of other fish species of the genus Rhinogobius and P. oligactis were downloaded from GenBank database for construction of a maximum likelihood (ML) tree [29] to analyze their evolutionary relationship. The tree was generated using the MEGA11 program, employing 1000 bootstrap replications. A COI sequence from Oxyeleotris marmorata (OK670610), a member of the family Butidae, was used as an outgroup.
3. Results
3.1. Morphological Identification
Although these two species of the genus Rhinogobius showed very similar characteristics, their appearances were clearly different (Table 2), including two other members of the family Oxudercidae, E. siamensis and P. oligactis. A short description of the four gobiid species examined in this study is provided below.
Rhinogobius chiengmaiensis Fowler, 1934
D1. vi-vii; D2. i,7-8; P1. i,14; P2. 12 (total); A. i,6-7; C. branched 10
First dorsal fin with 6-7 single rays; i,7-8 second dorsal fin: i,14 pectoral fin rays: 12 (total) pelvic fin rays: i,6-7 anal fin rays: 10 caudal branched rays; ctenoid scale; longitudinal scales 29-30; predorsal scales 3-4; circumcaudal scales 12; body elongate, moderate slender (15.85 ± 0.31 %SL); pelvic fins origin slightly in front of opercula margin (29.71 ± 2.56 vs. 31.82 ± 1.67 %SL); pelvic fins length 19.37 ± 1.15 %SL; mouth moderate large (35.50 ± 3.86 %HL), maxillary extending to middle of eyes; eyes diameter 20.16 ± 1.39 %HL.
Rhinogobius mekongianus (Pellegrin & Fang, 1940)
D1. vi; D2. i,7-8; P1. i,14; P2. 12 (total); A. i,6-7; C. branched 10
First dorsal fin with 6-7 single rays; i,7-8 second dorsal fin: i,14 pectoral fin rays: 10 (total) pelvic fin rays: i,6 anal fin rays: 12 caudal branched rays; ctenoid scale; longitudinal scales 29-30; predorsal scales 3-4; circumcaudal scales 12; body elongate, very slender (13.28 ± 0.54 %SL); pelvic fins origin slightly in front of opercula margin (29.33 ± 0.18 vs. 30.50 ± 0.38 %SL); pelvic fins length 17.88 ± 1.17 %SL; mouth moderate large (39.19 ± 2.06 %HL), maxillary extending to middle of eyes; eyes diameter 19.74 ± 0.57 %HL.
Eugnathogobius siamensis (Fowler, 1934)
D1. vi; D2. i,6-7; P1. i,16-18; P2. 10 (total); A. i,6; C. branched 12-14
First dorsal fin with 6 single rays; i,6-7 second dorsal fin: i,16-18 pectoral fin rays: 10 (total) pelvic fin rays: i,6 anal fin rays: 12-14 caudal branched rays; large ctenoid scale; longitudinal scales 24; predorsal scales 7; circumcaudal scales 12; body elongate, moderate slender (18.08 ± 2.29 %SL); pelvic fins origin slightly behind opercula margin (33.31 ± 1.54 vs. 32.60 ± 1.24 %SL); pelvic fins length 24.85 ± 0.35 %SL; caudal fin length 22.61 ± 3.17 %SL; mouth large (48.33 ± 12.41 %HL), maxillary extending well beyond posterior margin of eyes in male and extending to middle of eyes in female; small eyes, diameter 13.16 ± 0.72 %HL.
Pseudogobiopsis oligactis (Bleeker, 1875)
D1. vi; D2. i,6-7; P1. i,14; P2. 12-14 (total); A. i,7-8; C. branched 12-14
First dorsal fin with 6 single rays; i,6-7 second dorsal fin: i,14 pectoral fin rays: 12-14 (total) pelvic fin rays: i,7-8 anal fin rays: 12-14 caudal branched rays; large ctenoid scale; longitudinal scales 24-25; predorsal scales 7; circumcaudal scales 10; body elongate, moderate slender (15.88 ± 0.51 %SL); pelvic fins origin slightly behind opercula margin (36.06 ± 0.37 vs. 35.86 ± 3.28 %SL); pelvic fins length 22.75 ± 0.36 %SL; mouth large (63.24 ± 0.80 %HL), maxillary extending well beyond posterior margin of eyes; small eyes, diameter 12.63 ± 0.19 %HL. The diagnostic key to all four species gobiid in this study was provided as follows:
Diagnostic key to species of four gobiid in this study
- 1a
- large ctenoid scale, longitudinal scales 24-25, predorsal scales 7; pelvic fins origin slightly behind opercula margin .…………………………………..….…………..…..... 2
- 1b
- moderate ctenoid scale, longitudinal scales 29-30, predorsal scales 3-4; pelvic fins origin slightly behind opercula margin .……………………………….......................... 3
- 2a
- big head, head length 35.86 ± 3.28 %SL, head width 62.30 ± 3.15 %HL; mouth large (63.24 ± 0.80 %HL), maxillary extending well beyond posterior margin of eyes both gender; pelvic fins length 22.75 ± 0.36 %SL …………..……… Pseudogobiopsis oligactis
- 2b
- big head, head length 32.60 ± 1.24 %SL, head width 55.61 ± 3.85 %HL; mouth large (48.33 ± 12.41 %HL), in male maxillary extending well beyond posterior margin of eyes (57.10 %HL), and extending to middle of eyes in female (39.56 %HL), pelvic fins length 24.85 ± 0.35 %SL ……………...………….…..….. Eugnathogobius siamensis
- 3a
- mouth moderate large, maxillary extending to middle of eyes in both gender (35.50 ± 3.86 %HL); body moderate slender, body depth at pelvic fins origin 15.85 ± 0.31 %SL; pelvic fins length 19.37 ± 1.15 %SL; caudal fin length 24.77 ± 0.54 %SL ………………………………………………………………… Rhinogobius chiengmaiensis
- 3b
-
mouth moderate large, maxillary extending to middle of eyes in both gender (39.19 ± 2.06 %HL); body very slender, body depth at pelvic fins origin 13.28 ± 0.54 %SL; pelvic fins length 17.88 ± 1.17 %SL; caudal fin length 27.34 ± 2.28 %SL …...……..…..…………………………….……………………………………….. Rhinogobius mekongianus
3.2. Molecular Identification
The 707-bp fragments of the COI gene were successfully amplified from all samples using the PCR technique. The 235 amino acid residues were translated without stop codon, deletion, and insertion for any sequences. From sequence alignment, two sequences of R. chiengmaiensis showed 100% similarity (1 haplotype). While five sequences of R. mekongianus showed 99.4% similarity (4 haplotypes), four bases were different. Two COI sequences of R. chiengmaiensis were compared to reference sequences in the GenBank and BOLD databases, which top matched with Rhinogobius virgigena at 96.24% and 96.21% identities, respectively. Likewise, five sequences of R. mekongianus also top matched with R. virgigena at 95.92-96.24% and 95.89-96.21% identities in the GenBank and BOLD databases, respectively.
The COI sequences of E. siamensis and P. oligactis demonstrated complete similarity, with each species showing a haplotype. Sixteen bases were different from each species. For comparison with the reference sequences in the GenBank and BOLD databases, two COI sequences of E. siamensis were top matched with P. oligactis at 98.14% and 98.92% identities, respectively. Meanwhile, two COI sequences of P. oligactis were top matched with P. oligactis at 98.14% and 98.92% identities in the GenBank and BOLD databases, respectively.
The average base compositions of eleven COI sequences were T (29.0 ± 1.3%), C (28.6 ± 0.1%), A (22.9 ± 0.9%), and G (19.5 ± 0.5%), as shown in Table 3. The GC content was 48.2 ± 0.5% at all sites while the AT content was 51.8 ± 0.5%. Pseudogobiopsis oligactis presented the highest GC content (49.1 ± 0.0%), while R. mekongianus had the lowest (47.8 ± 0.1%).
The intraspecific genetic distances were 0.00% for R. chiengmaiensis, E. siamensis and P. oligactis, and 0.28% for R. mekongianus (Table 4), while the average intraspecific distance was 0.07%. In contrast, the interspecific genetic distances ranged from 0.86% to 16.63%. The lowest distance was between R. chiengmaiensis and R. mekongianus (0.86%). The highest distances were between R. chiengmaiensis and P. oligactis (16.63%) and R. chiengmaiensis and E. siamensis (15.38%), followed by R. mekongianus and P. oligactis (12.00%) and R. mekongianus and E. siamensis (11.02%), respectively. Meanwhile, the distance between P. oligactis and E. siamensis was 1.64%. The average interspecific distance was 9.59%. Thus, the interspecific genetic distance was 137-fold greater than the intraspecific genetic distance.
The ML phylogenetic tree presented the relationship between the COI sequences of the four gobiid species of this study and other species retrieved from the GenBank database (Figure 1). Two major clades were clearly delineated, one comprising Rhinogobius species and the other including E. siamensis and P. oligactis. Within these groups, R. chiengmaiensis showed the greatest similarity to R. mekongianus, while E. siamensis and P. oligactis clustered together. Additionally, the COI sequence of Oxyeleotris marmorata was distinctly separated from the other sequences.
4. Discussion
The morphometric characters of R. chiengmaiensis and R. mekongianus provided in this study aligned with findings from previous research [4,14,30,31]. Rhinogobius chiengmaiensis and R. mekongianus exhibited quite similar appearances; however, they differed in certain characteristics such as the number of rays in the pelvic fin, anal fin, and caudal branch, as well as the diameter of the eyes. Additionally, a notable distinguishing characteristic was the body shape, with R. mekongianus being more slender compared to R. chiengmaiensis.
The concise characterizations of E. siamensis and P. oligactis presented in this study were also consistent with the descriptions provided in the previous research [32,33]. Considerable confusion has arisen in accurately distinguishing between these two gobiid fish species due to their similar appearances, particular the coloration of their head and body [24] and greatly enlarged jaws [34]. In addition, these two species have been greatly confused, as evidenced by their synonyms being swapped between genera in FishBase, a global fish species database (https://www.fishbase.se/). At one time, the invalid names Pseudogobiopsis siamensis and Eugnathogobius oligactis were used for E. siamensis and P. oligactis, respectively. In 2009, Larson [32] conducted a review of the gobiid fish genera Eugnathogobius and Pseudogobiopsis, which significantly enhanced the ability to distinguish between these two species.
To accurately identify these four gobiid species, which exhibit highly similar external characteristics, it is essential to employ molecular data and DNA barcoding, alongside traditional taxonomic methods. This integrative approach ensures precise identification of these closely related fish species. Xia et al. [9] examined the COI sequences of the newly discovered species R. maxillivirgatus, revealed that it is closely related to its nearest species, R. wuyanlingensis, while still being distinct.
Due to low identity percentages, the COI sequences of two fish species belonging to the genus Rhinogobius in Thailand have not yet been reported in any databases. Thus, this study was the first investigation to report the COI sequences of R. chiengmaiensis and R. mekongianus. Additionally, the COI sequences of E. siamensis have also been initially provided. However, the top match identity percentages for the COI sequences of P. oligactis were below 99%, likely due to genetic variation influenced by their diverse habitats [35]. A total of eleven COI sequences were deposited in the GenBank database with accession numbers PQ193904-PQ193914 (Table 1).
The average AT content (51.8 ± 0.5%) was greater than the average GC content (48.2 ± 0.5%) due to the average T being the highest in the base composition of eleven sequences, followed by C and A, respectively. The highest average thymine base composition was exhibited in several fishes including freshwater fishes of Bangladesh [16], marine and coastal fishes of Bangladesh [17], fish species in the Taiwan Strait [18], four fish species in the family Notopteridae [19] and Wallago attu [35]. Furthermore, the average G content was the lowest, representing a clear pattern of anti-G bias [16,17].
Generally, the intraspecific genetic distances based on the COI gene for each animal species are usually less than 2% [36], including fishes [20]. The average intraspecific genetic distances of each species in this study ranged from 0.00% to 0.28%. Conversely, the average interspecific genetic distances were higher, ranging from 0.86% to 16.63%. The lower intraspecific and higher interspecific genetic distances suggested that the COI sequences were effective in distinguishing between the four gobiid species examined in this study. This investigation made use of observations in several studies, including fish species in the Taiwan Strait [18], four fish species in the family Notopteridae [19], and Cyprinidae fish in the midstream of the Yangtze river [37]. Furthermore, the 137-fold differences between the average interspecific and intraspecific genetic distance exhibited the species delimitation of the four species. According to previous studies, the difference was greater than the 214-fold difference observed in four fish species in the family Notopteridae [19]. Hebert et al. [38] suggested that a 10-fold COI sequence difference between the average interspecific and intraspecific differences serve as a criterion for animal species differences. These investigations demonstrated that the COI gene sequences were effective in distinguishing the same species from other species. Nevertheless, the small sample size of this study was one of its limitations.
The evolutionary relationship of the COI sequences among the four fish species and the other species is shown in Figure 1, according to the fish taxonomy of Nelson et al. [1]. The species in the family Oxudercidae, which consisted of Rhinogobius spp., E. siamensis, and P. oligactis, were clearly separated from O. marmorata, that belongs to the family Butidae. One major clade was the Rhinogobius species. The COI sequences of R. chiengmaiensis and R. mekongianus were the closest species, which agreed with their morphological identification, followed by R. virgigena and other Rhinogobius species. The second clade comprised E. siamensis and P. oligactis, indicating a close evolutionary relationship between these two species. The genera Eugnathogobius and Pseudogobiopsis were categorized as a part of the Mugilogobius-lineage, whereas the genus Rhinogobius was classified to the Acanthogobius-lineage, for which the phylogenetic analysis was similar to studies conducted by Agorreta et al. [39].
At present, these four gobiid species are increasingly popular as ornamental fish. They are often collected from the wild, which may lead to a decline in their natural populations. Furthermore, the IUCN has assessed one species, R. chiengmaiensis, as vulnerable (VU), indicating that this species is at considerable risk of becoming endangered in its natural habitat unless effective conservation measures are implemented [40]. The remaining species were assessed as least concern (LC). Therefore, promoting the commercial breeding of these fish can resolve this problem. A few research studies have been conducted on the breeding of these four gobiid fish species, including R. chiengmaiensis [41]. However, these fish have very similar appearances, making accurate species identification necessary before breeding. Furthermore, the results of this study will contribute to species identification and serve as information for future research on new fish species in the genus Rhinogobius.
5. Conclusions
The species R. chiengmaiensis and R. mekongianus have been identified through both morphological and molecular methods. Additionally, E. siamensis and P. oligactis, members of the family Oxudercidae, were also identified. The morphology of the four gobiid species exhibited distinctive characteristics, although some species displayed considerable similarity. For molecular identification, DNA barcoding proves to be an effective approach for distinguishing between these four species, considering both intraspecific and interspecific genetic distances as well as phylogenetic analysis. Moreover, this study is the first to report the COI sequences for R. chiengmaiensis, R. mekongianus, and E. siamensis. These findings can serve as a valuable database for future research, including breeding programs and the discovery of new species.
Author Contributions
Conceptualization, S.T. and D.P.; methodology, S.T., N.P. and D.P.; software, S.T. and D.P.; validation, S.T. and D.P.; formal analysis, S.T., N.P. and D.P.; investigation, S.T., P.P., N.P., E.W., S.R., K.M. and D.P.; resources, S.T., N.P. and D.P.; data curation, S.T. and D.P.; writing—original draft preparation, S.T. and D.P.; writing—review and editing, S.T., P.P., N.P., E.W., S.R., K.M. and D.P.; visualization, S.T. and D.P.; supervision, D.P.; project administration, D.P.; funding acquisition, D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by University of Phayao and Thailand Science Research and Innovation Fund (Fundamental Fund 2025, Grant No. 5051/2567).
Institutional Review Board Statement
This study was approved by the Institutional Animal Care and Use Committee, University of Phayao (No. UP-AE 1-011-67) on 1 August 2024.
Informed Consent Statement
Not applicable.
Data Availability Statement
All sequences of the COI gene have been deposited under GenBank accession numbers PQ193904-PQ193914.
Acknowledgments
We are grateful to Assoc. Prof. Chitnarong Sirisathitkul and Assis. Prof. Yaowarat Sirisathitkul for their suggestions on this manuscript. Additionally, we would like to thank Ms. Kanokwan Timkon, Mr. Auttpon Timkon, and Mr. Ketjarin Kheawnil for providing some samples.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons, Inc.: New Jersey, Canada, 2016. [Google Scholar]
- Nelson, J.S. Fishes of the World, 4th ed.; John Wiley & Sons, Inc.: New Jersey, Canada, 2006. [Google Scholar]
- Thacker, C.E. Phylogenetic placement of the European sand gobies in Gobionellidae and characterization of gobionellid lineages (Gobiiformes: Gobioidei). Zootaxa 2013, 3619, 369–382. [Google Scholar] [CrossRef]
- Chen, I.S.; Kottelat, M.; Miller, P.J. Freshwater gobies of the genus Rhinogobius from the Mekong basin in Thailand and Laos, with descriptions of three new species. Zool. Stud. 1999, 38, 19–32. [Google Scholar]
- Chen, I.S.; Miller, P.J. A new freshwater goby of Rhinogobius (Teleostei: Gobiidae) from Hainan Island, southern China. J. Mar. Sci. Technol. 2013, 21, 124–129. [Google Scholar]
- Kottelat, M. Zoogeography of the fishes from Indochinese inland waters with an annotated check-list. Bull. ZooI. Mus. Univ. Amsterdam 1989, 12, 1–55. [Google Scholar]
- McCraney, W.T.; Thacker, C.E.; Alfaro, M.E. Supermatrix phylogeny resolves goby lineages and reveals unstable root of Gobiaria. Mol. Phylogenet. Evol. 2020, 151, 106862. [Google Scholar] [CrossRef]
- Thacker, C.E.; Roje, D.M. Phylogeny of Gobiidae and identification of gobiid lineages. Syst. Biodivers. 2011, 9(4), 329–347. [Google Scholar] [CrossRef]
- Xia, J.H.; Wu, H.L.; Li, C.H.; Wu, Y.Q.; Liu, S.H. A new species of Rhinogobius (Pisces: Gobiidae), with analyses of its DNA barcode. Zootaxa 2018, 4407, 553. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.S.; Wang, S.C.; Shao, K.T. A new freshwater gobiid species of Rhinogobius Gill, 1859 (Teleostei: Gobiidae) from northern Taiwan. Zootaxa 2022, 5189, 29–44. [Google Scholar] [CrossRef]
- Suzuki, T.; Oseko, N.; Yamasaki, Y.Y.; Kimura, S.; Shibukawa, K. A new species with two new subspecies of Rhinogobius (Teleostei: Gobiidae) from Yaeyama Group, the Ryukyu Islands, Japan. Bull. Kanagawa Pref. Mus. (Nat. Sci.) 2022, 51, 9–34. [Google Scholar]
- Maeda, K.; Kobayashi, H.; Iida, M.; Tran, H.D. Taxonomy of freshwater gobies of the genus Rhinogobius (Oxudercidae, Gobiiformes) from central Vietnam, with descriptions of two new species. Zootaxa 2024, 5493(5), 507–541. [Google Scholar] [CrossRef]
- Panitvong, N. Freshwater Fishes of Thailand; Parbpim Ltd.: Bangkok, Thailand, 2020. [Google Scholar]
- Suvarnaraksha, A.; Utsugi, K. A Field Guild of the Northern Thai Fishes; Maejo University, Chiang Mai, Thailand, and Nagao Natural Environment Foundation: Tokyo, Japan, 2023. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.S.; Datta, S.K.; Zhilik, A.A. Molecular diversity of freshwater fishes of Bangladesh assessed by DNA barcoding. Bangladesh J. Zool. 2020, 48, 1–19. [Google Scholar] [CrossRef]
- Ahmed, M.S.; Datta, S.K.; Saha, T.; Hossain, Z. Molecular characterization of marine and coastal fishes of Bangladesh through DNA barcodes. Ecol. Evol. 2021, 11, 3696–3709. [Google Scholar] [CrossRef] [PubMed]
- Bingpeng, X.; Heshan, L.; Zhilan, Z.; Chunguang, W.; Yanguo, W.; Jianjun, W. DNA barcoding for identification of fish species in the Taiwan Strait. PLoS ONE 2018, 13, e0198109. [Google Scholar] [CrossRef]
- Seetapan, K.; Panprommin, N.; Wangkahart, E.; Ruenkoed, S.; Panprommin, D. COI-high resolution melting analysis for discrimination of four fish species in the family Notopteridae in Thailand. Zool. Anz. 2024, 309, 90–97. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia's fish species. Philos. Trans. R. Soc. B. 2005, 360, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Panprommin, D.; Manosri, R. DNA barcoding as an approach for species traceability and labeling accuracy of fish fillet products in Thailand. Food Control 2022, 136, 108895. [Google Scholar] [CrossRef]
- Breitbart, M.; Kerr, M.; Schram, M.J.; Williams, I.; Koziol, G.; Peebles, E.; Stallings, C.D. Evaluation of DNA metabarcoding for identifying fish eggs: a case study on the West Florida Shelf. PeerJ 2023, 11, e15016. [Google Scholar] [CrossRef]
- Chen, W.; Li, C.; Li, X.; Li, J.; Li, Y. Unraveling the drifting larval fish community in a large spawning ground in the Middle Pearl River using DNA barcoding. Animals 2022, 12, 2555. [Google Scholar] [CrossRef]
- Nagao Natural Environment Foundation. Fishes of the Indochinese Mekong; Nagao Natural Environment Foundation: Tokyo, 2021. [Google Scholar]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol Biol. 2014, 1079, 105–116. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Chen, I.S.; Cheng, Y.H.; Shao, K.T. A new species of Rhinogobius (Teleostei: Gobiidae) from the Julongjiang Basin in Fujian Province, China. Ichthyol. Res. 2008, 55, 335–343. [Google Scholar] [CrossRef]
- Kottelat, M. Fishes of Laos; WHT Publications Ltd.: Colombo, 2001. [Google Scholar]
- Larson, H.K. Review of the gobiid fish genera Eugnathogobius and Pseudogobiopsis (Gobioidei: Gobiidae: Gobionellinae), with descriptions of three new species. Raffles Bull. Zool. 2009, 57, 127–181. [Google Scholar]
- Tan, H.H.; Lim, K.K.P. Rediscovery of the bigmouth stream goby, Pseudogobiopsis oligactis (Actinopterygii: Gobiiformes: Gobionellidae) in Singapore. Nat. Singap. 2011, 4, 363–367. [Google Scholar]
- Smith, H.M. The freshwater fishes of Siam, or Thailand. Bull. U.S. Natl. Mus. 1945, 188, 1–622. [Google Scholar]
- Sajjad, A.; Jabeen, F.; Ali, M.; Zafar, S. DNA barcoding and phylogenetics of Wallago attu using mitochondrial COI gene from the River Indus. J. King Saud Univ. - Sci. 2023, 35, 102725. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. B 2003, 270 (Suppl_1), S96–S99. [Google Scholar] [CrossRef]
- Shen, Y.; Guan, L.; Wang, D.; Gan, X. DNA barcoding and evaluation of genetic diversity in Cyprinidae fish in the midstream of the Yangtze River. Ecol. Evol. 2016, 6, 2702–2713. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Stoeckle, M.Y.; Zemlak, T.S.; Francis, C.M. Identification of birds through DNA barcodes. PLoS Biol. 2004, 2, e312. [Google Scholar] [CrossRef] [PubMed]
- Agorreta, A.; San Mauro, D.; Schliewen, U.; Van Tassell, J.L.; Kovačić, M.; Zardoya, R.; Rüber, L. Molecular phylogenetics of Gobioidei and phylogenetic placement of European gobies. Mol. Phylogenet. Evol. 2013, 69, 619–633. [Google Scholar] [CrossRef] [PubMed]
- International Union for Conservation of Nature. The IUCN Red List of Threatened Species. 2023. Available online: https://www.iucnredlist.org/.
- Pornsopin, P.; Sirisuksa, T.; Kantiyawong, S.; Surajit, T. Study on cultivation of Chiangmai stream goby (Rhinogobius chiengmaiensis fowler, 1934); Inland Aquaculture Research and Development Division: Bangkok, 2022. [Google Scholar]
Figure 1.
A maximum likelihood (ML) phylogenetic tree was constructed using eleven COI nucleotide sequences from four gobiid species, highlighted in bold, in comparison with multiple sequences sourced from the GenBank database. The bootstrap values are presented above the branches. A COI sequence of Oxyeleotris marmorata (OK670610) was designated as the outgroup species. The scale bar corresponds to the number of substitutions per site.
Figure 1.
A maximum likelihood (ML) phylogenetic tree was constructed using eleven COI nucleotide sequences from four gobiid species, highlighted in bold, in comparison with multiple sequences sourced from the GenBank database. The bootstrap values are presented above the branches. A COI sequence of Oxyeleotris marmorata (OK670610) was designated as the outgroup species. The scale bar corresponds to the number of substitutions per site.
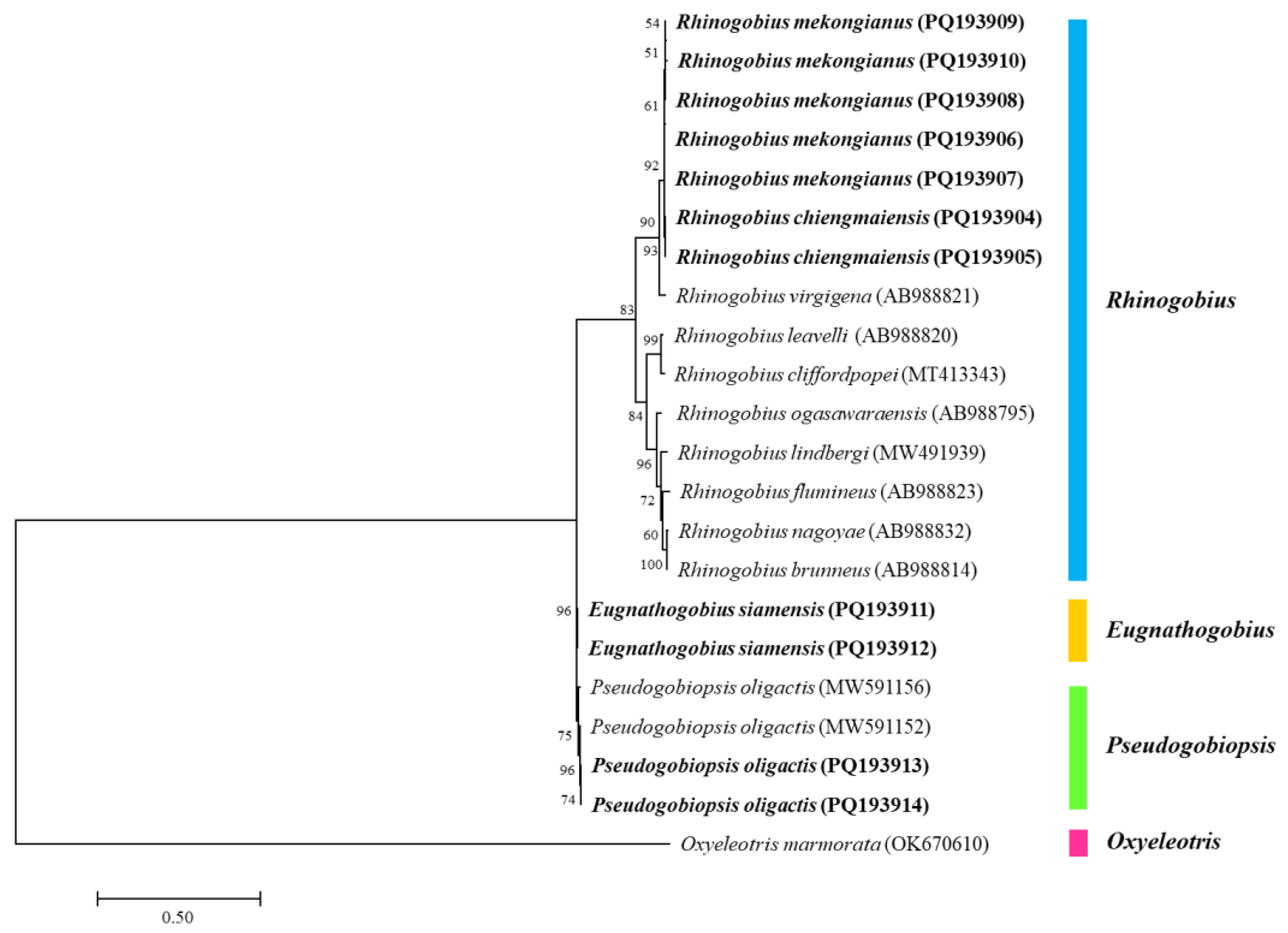

Table 1.
Primary information about eleven COI sequences in this study.
| Species | Accession no. | Collection site |
|---|---|---|
| Rhinogobius chiengmaiensis | PQ193904-PQ193905 | Ping river basin, Chiang Mai province |
| Rhinogobius mekongianus | PQ193906-PQ193910 | Kok River, Chiang Mai province |
| Eugnathogobius siamensis | PQ193911-PQ193912 | Surat Thani province |
| Pseudogobiopsis oligactis | PQ193913-PQ193914 | Satun province |
Table 2.
Morphometry of R. chiengmaiensis, R. mekongianus, E. siamensis, and P. oligactis.
| Characters | R. chiengmaiensis | R. mekongianus | E. siamensis | P. oligactis |
|---|---|---|---|---|
| SL (mm) | 28.82 ± 0.51 | 29.16 ± 2.95 | 30.48 ± 2.98 | 29.42 ± 3.68 |
| As % in SL | ||||
| Head length | 31.82 ± 1.67 | 30.50 ± 0.38 | 32.60 ± 1.24 | 35.86 ± 3.28 |
| Head width | 16.46 ± 3.06 | 15.23 ± 1.90 | 18.11 ± 0.57 | 22.39 ± 3.17 |
| Body depth at P2 | 15.85 ± 0.31 | 13.28 ± 0.54 | 18.08 ± 2.29 | 15.88 ± 0.51 |
| Body depth at A | 14.80 ± 0.24 | 11.78 ± 0.37 | 16.49 ± 1.94 | 13.52 ± 0.72 |
| Snout to D1 | 37.28 ± 1.59 | 39.66 ± 2.02 | 39.56 ± 3.06 | 45.14 ± 1.75 |
| Snout to D2 | 59.50 ± 2.57 | 59.01 ± 3.47 | 61.91 ± 2.21 | 58.13 ± 0.89 |
| Snout to A | 65.08 ± 1.89 | 64.27 ± 0.31 | 65.00 ± 1.75 | 63.29 ± 2.38 |
| Snout to P2 | 29.71 ± 2.56 | 29.33 ± 0.18 | 33.31 ± 1.54 | 36.06 ± 0.37 |
| D1 base length | 15.24 ± 2.40 | 14.46 ± 1.18 | 13.16 ± 0.13 | 8.84 ± 0.50 |
| D1 base length | 20.62 ± 2.94 | 17.39 ± 0.98 | 16.95 ± 0.29 | 16.31 ± 1.47 |
| A base length | 14.19 ± 0.01 | 13.72 ± 0.17 | 15.85 ± 1.16 | 13.31 ± 3.12 |
| C length | 24.77 ± 0.54 | 27.34 ± 2.28 | 22.61 ± 3.17 | - |
| Caudal peduncle length | 20.60 ± 1.03 | 19.72 ± 0.15 | 20.53 ± 1.52 | 13.95 ± 2.88 |
| Caudal peduncle depth | 11.55 ± 0.38 | 9.62 ± 0.54 | 11.38 ± 0.44 | 10.56 ± 0.56 |
| P1 length | 20.74 ± 6.94 | 25.48 ± 1.84 | 28.63 ± 0.56 | 22.70 ± 0.37 |
| P2 length | 19.37 ± 1.15 | 17.88 ± 1.17 | 24.85 ± 0.35 | 22.75 ± 0.36 |
| As % in HL | ||||
| Snout length | 29.76 ± 1.44 | 27.92 ± 2.71 | 27.98 ± 0.75 | 27.79 ± 0.01 |
| Eye diameter | 20.16 ± 1.39 | 19.74 ± 0.57 | 13.16 ± 0.72 | 12.63 ± 0.19 |
| Postorbital length | 50.08 ± 0.04 | 52.34 ± 2.13 | 58.87 ± 0.03 | 59.58 ± 0.17 |
| Interorbital space | 21.93 ± 0.01 | 16.35 ± 1.22 | 19.13 ± 0.03 | 19.99 ± 1.23 |
| snout to maxilla | 35.50 ± 3.86 | 39.19 ± 2.06 | 48.33 ± 12.41 | 63.24 ± 0.80 |
| Head width | 51.54 ± 6.93 | 49.97 ± 6.87 | 55.61 ± 3.85 | 62.30 ± 3.15 |
SL: standard length; HL: head length; D1: first dorsal fin; D2: second dorsal fin; P1: pectoral fin; P2: pelvic fin; A: anal fin; C: caudal fin. The values are presented as the mean ± standard error.
Table 3.
The average nucleotide composition, GC content, and AT content (%) of all sites in the COI sequences of the four fish species.
Table 3.
The average nucleotide composition, GC content, and AT content (%) of all sites in the COI sequences of the four fish species.
| Species | Nucleotide composition (%) | %GC content |
%AT content |
||||
|---|---|---|---|---|---|---|---|
| T | C | A | G | ||||
| R. chiengmaiensis | 29.7 ± 0.0 | 28.8 ± 0.0 | 22.3 ± 0.0 | 19.2 ± 0.0 | 48.0 ± 0.0 | 52.0 ± 0.0 | |
| R. mekongianus | 30.0 ± 0.1 | 28.5 ± 0.0 | 22.2 ± 0.1 | 19.3 ± 0.1 | 47.8 ± 0.1 | 52.2 ± 0.1 | |
| E. siamensis | 27.4 ± 0.0 | 28.6 ± 0.0 | 24.3 ± 0.0 | 19.7 ± 0.0 | 48.3 ± 0.0 | 51.7 ± 0.0 | |
| P. oligactis | 27.3 ± 0.0 | 28.7 ± 0.0 | 23.6 ± 0.0 | 20.4 ± 0.0 | 49.1 ± 0.0 | 50.9 ± 0.0 | |
| Average | 29.0 ± 1.3 | 28.6 ± 0.1 | 22.9 ± 0.9 | 19.5 ± 0.5 | 48.2 ± 0.5 | 51.8 ± 0.5 | |
The values are presented as the mean ± standard error.
Table 4.
The average K2P intraspecific and interspecific genetic distances (%) were determined for the four fish species.
Table 4.
The average K2P intraspecific and interspecific genetic distances (%) were determined for the four fish species.
| Species | R. chiengmaiensis | R. mekongianus | E. siamensis | P. oligactis |
|---|---|---|---|---|
| R. chiengmaiensis | 0.00 | |||
| R. mekongianus | 0.86 | 0.28 | ||
| E. siamensis | 15.38 | 11.02 | 0.00 | |
| P. oligactis | 16.63 | 12.00 | 1.64 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



