
Submitted:
18 February 2025
Posted:
19 February 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The persistent threat of emerging respiratory RNA viruses like SARS-CoV-2 and Influenza A virus (IAV) necessitates the continuous development of effective, safe, broadly acting and gener-ally accessible antiviral agents. Current treatments often face limitations such as early admin-istration requirements, resistance development, and limited global access. Natural products like European black elderberry (Sambucus nigra L.) fruit extract and quinine have been used histori-cally against viral infections. In this study, we investigated the antiviral efficacy of a standard-ized black elderberry fruit extract containing 3.2 % anthocyanins (EC 3.2) and, as a second natu-ral antiviral product, quinine, against IAV and SARS-CoV-2 in vitro. Madin-Darby Canine Kid-ney II (MDCKII) cells were infected with IAV PR-8, while human Calu-3 lung epithelial cells were infected with SARS-CoV-2 Wuhan type. Cells were treated with varying concentrations of EC 3.2 and quinine as either mono– or combinational therapy. Viral replication was assessed using quantitative RT-PCR, and cell viability was evaluated using WST-1 assays. Our results demonstrate for the first time that both EC 3.2 and quinine individually inhibited IAV replica-tion in a dose-dependent manner, with IC₅₀ values of approximately 1:400 for EC 3.2 and 250 nM for quinine. Most importantly, the combinational treatment exhibited a strong synergistic anti-viral effect, as confirmed by the Bliss independence model (synergy scores of 14.7 for IAV and 27.8 for SARS-CoV-2) without affecting cell viability. These findings suggest that the combined use of black elderberry extract and quinine may serve as an effective antiviral strategy against IAV and SARS-CoV-2. The synergistic effect allows for lower doses of each product while keep-ing therapeutic efficacy. In summary, this combinational approach should be expanded to other respiratory RNA viruses, and thus might offer a promising avenue for pandemic preparedness. Moreover, the in vitro data obtained in this study have to be confirmed in clinical studies.
Keywords:
European black elderberry fruit extract
; quinine
; natural substance
; anthocyanins
; phenolic compounds
; SARS-CoV-2
; Influenza A Virus
; antiviral
; pandemic preparedness
; broad antiviral activity
1. Introduction
Even after its transition to an endemic spread, the SARS-CoV-2 remains a public health menace, mainly in the light of emerging variants of concern (VoCs) and the necessity for pandemic preparedness to encounter newly emerging and/or reemerging viruses in the future [1]. Several vaccines and monoclonal antibodies have been developed and globally approved, which could moderate the course of infection, although sterile immunity, which would prevent SARS-CoV-2 virus transmission and thus, progressive adaptation of the pathogen, could not be achieved [2,3,4]. In addition, the protease inhibitor Paxlovid® as well as the polymerase inhibitors Molnupiravir and Remdesivir have been available for antiviral treatment [5]. However, given the fact that these drugs are required to become administered in the initial stages of infection, operational and prophylactically acting substances with subtle adverse effects have still not been made available. For these reasons, and with respect to potential future pandemic events, there is still an unmet medical need for broadly acting antiviral drugs against SARS-CoV-2 VoCs, as well as other respiratory viruses, which should be safe, readily available and inexpensive.
Similar to the situation with SARS-CoV-2, Influenza A is a highly contagious respiratory virus causing seasonal epidemics with significant morbidity and mortality worldwide. Every year about 10% of the world’s population is infected with Influenza and each year about 500.000 people die due to infection [6]. Characterized by symptoms such as fever, cough, sore throat, muscle aches, and fatigue, it poses a recurring public health challenge [7]. Current prevention and treatment options include annual vaccinations, antiviral medications like the neuraminidase inhibitors Oseltamivir, Zanamivir and Peramivir, and symptomatic care [6]. However, issues such as antiviral resistance, variable vaccine efficacy, and limited global access to treatments highlight the ongoing medical need for improved vaccines, new effective antiviral drugs, rapid diagnostics, and equitable healthcare strategies [8]. Altogether, pandemic preparedness against upcoming emerging highly pathogenic IAVs with pandemic potential is inevitable.
The coevolution of humankind and viruses headed to the customarily uncharted and thus more or less intuitional practice of environmental products in prevention and treatment of infectious diseases [9,10]. Natural substances have been applied for epochs in order to oppose infections, even though the precise antiviral mode of action have not been established in most cases [9]. Natural antivirals have been used as prophylaxis and treatment to avoid deterioration of diseases persuaded by viral infection, principally in cases of newly emerging or reemerging viruses [11]. Given the fact that they ordinarily represent broadly acting antivirals they exhibit immense potential to improve our pandemic preparedness [9]. Additionally, in most cases, they are favorably targeting host cell structures and thus odds for occurrence of drug resistances are low. Furthermore, for the reason of their natural origin, they have lower side effects than synthetically intended drugs [9,10,11].
European Black elderberry (Sambucus nigra L.) has been used for centuries in traditional medicine to treat cold diseases mostly due to respiratory viruses. Recent in vivo studies, conducted in animal and humans, have demonstrated that extracts from the berries of black elderberry possess antiviral properties and can shorten the duration and severity of upper respiratory infections in humans [12,13,14,15]. Very recently, the strong antiviral activity of black elderberry fruit extract, standardized to anthocyanins and phenolic compounds, against SARS-CoV-2 and its variants was demonstrated, suggesting its potential as an effective treatment option for COVID-19 [16].
Moreover, crude ethanol extract of Sambucus nigra has shown significant antiviral properties against Infectious Bronchitis Virus (IBV), reducing viral titers by four to six orders of magnitude. The extract works by compromising viral envelopes and inducing membrane vesicles, rendering the virus non-infectious early in the infection process [17]. In addition, cyanidin-3-sambubiocide, a main anthocyanin of black elderberry extract, has been shown to be a potent inhibitor of Influenza neuraminidase. The anthocyanin interacts specifically with the active center of the neuraminidase and thereby inhibits virus release [18].
Roschek et al. identified anti-viral components in elderberry (Sambucus nigra L.) extract, which inhibited human Influenza A Virus (H1N1) in vitro. Key flavonoids, including 5,7,3’,4’-tetra-O-methylquercetin, demonstrated potent anti-H1N1 activity, comparable to the neuraminidase inhibitors Oseltamivir and Amantadine [19].
Quinine can be extracted from the bark of the Cinchona tree and was used worldwide for centuries to treat feverish infections, especially malaria. As a matter of principle, Quinine can be anticipated as the very first ancient medicine that was applied in form of a purified drug since the 17th century [20,21,22]. It’s medical use was first documented in 1630, where quinine was used to successfully treat the countess of Chinchon from malaria using an extract of the bark of the fever tree, later termed as Cinchona bark [21,22]. The structure of quinine was discovered in 1820 and it was first chemically synthesized in 1944 as the only available antimalarial drug. Quinine was extensively used in the 19th century by British citizens and soldiers as prevention against malaria [23]. It served as a template for the synthesis of Hydroxy-Chloroquine (H-CQN) in 1946, which was then mainly used for the treatment of malaria. Until now it is still a treatment option for severe and H-CQN-resistant cases of malaria tropica [21,22,23].
In addition, quinine is approved for the treatment of calf cramps, and is widely used as an aromatic agent in beverages like tonic water, with its use subjected to regulatory limits. Recently, we have shown for the first time that quinine efficiently inhibits the replication of SARS-CoV-2 in vitro [20]. Moreover, antiviral activity of quinine against Human Immunodeficiency Virus, Zika Virus, Herpes Simplex Virus, Ebola and Dengue virus was described [21,24,25,26,27]. Interestingly, in 1946 Seeler et al. reported first enigmatic hints, that quinine exhibits a benefit on Influenza infections in mice [28].
Here, we first describe that a black elderberry fruit extract containing 3.2% anthocyanins (Elder Craft 3.2; EC 3.2) efficiently inhibits the replication of the Influenza A H1N1 isolate PR-8 (IAVPR-8) in vitro. Moreover, this is the first-time report that quinine inhibits the replication of IAVPR-8 with an IC50 of 250 nM in vitro. Most importantly, we could show that the combinational treatment of quinine with EC 3.2 exhibits a synergistic antiviral activity against IAV and SARS-CoV-2. Our cumulative data suggest that combined treatment with quinine and black elderberry extract might be considered as a potentially effective treatment of SARS-CoV-2 or IAV infections leveraging their synergistic effects.
2. Materials and Methods
2.1. Inhibitors
Liquid European black elderberry (Sambucus nigra L.) fruit extract, branded as ElderCraft® and designated EC 3.2, was provided by Iprona Lana SpA (Lana, Italy). EC 3.2 is a water-based liquid extract standardized to contain a minimum of 3.2% anthocyanins. Dry European black elderberry extract, standardized to 15% anthocyanins and designated EC 15, was also supplied by Iprona Lana SpA. Additionally, a combination product containing anthocyanins and quinine, designated ElderCraft® Q-Complex, was obtained from the same supplier. The exact composition of EC 3.2 and EC 15 is provided in Supplementary Table S1. The anthocyanin stability in EC 3.2 was measured for eleven months by monthly total anthocyanin quantification using the pH differential method (Supplementary Figure S1). Quinine was obtained as quinine-sulfate from Sigma-Aldrich (St. Louis, MO, USA) and dissolved in DMSO.
2.2. Viruses
The “Wuhan type” virus SARS-CoV-2PR-1, isolated from a 61-year-old patient, was amplified in Vero B4 cells as described previously [20]. Viral titers were determined by an endpoint titration assay. For the generation of new virus stock, virus containing cell culture supernatant was harvested 72 hours post infection (hpi) and passed through a 0.45 μm pore-size filter. All virus stocks were stored at −80 °C until further usage.
The Influenza A virus isolate (IAV) A/Puerto Rico/8/34 (PR8) was a provided by Prof. Dr. Matthias Tenbusch (Institute of Clinical and Molecular Virology, Erlangen, Germany). Viral titers were determined by a plaque assay. For the generation of new virus stock, virus containing cell culture supernatant was harvested 48 hpi and passed through a 0.45 μm pore-size filter. All virus stocks were stored at −80 °C until further usage.
2.3. Infection Experiments
Calu-3 cells were inoculated with SARS-CoV-2PR-1 (Wuhan type) with a multiplicity of infection (MOI) of 2x10-2 for 1 h, washed and further treated with interventions. 72 hpi, virus-containing cell culture supernatants were incubated for 10 min at 95°C and finally used for qRT-PCR analysis. For titer determination of SARS-CoV-2 virus stocks, Calu-3 cells were infected with serial dilutions of the virus stock over 72 h. Afterwards cells were fixed (4% PFA), permeabilized (0.5 % Triton/PBS), blocked (1% BSA/PBS-T) and finally stained with a SARS-CoV-2 NP antibody (Biozol, Eching, Germany). Endpoint of virus infection was analyzed via fluorescence microscopy and viral titer was calculated by the method of Reed and Muench [29].
MDCKII (Madin-Darby Canine Kidney II) cells were inoculated with IAVPR8 (MOI: 0.01) for 30 minutes, washed and further treated with the indicated interventions. 48 hpi, virus-containing cell culture supernatants were incubated for 10 min at 95°C and finally used for qRT-PCR analysis. For titer determination of IAVPR-8 virus stocks, MDCKII cells were infected with serial dilutions of the virus stock for 3 h. Afterwards cells were overlayed with 1% purified agar (Oxoid, Wesel, Germany) and incubated at 37°C for 4 days. Endpoint of virus infection was analyzed by counting plaque forming units (PFU) visualized following incubation of the cells with MTT (3-(4,5-Dimethylthiazol-2-yl) for 3 hours. Viral titer was calculated by the method of Reed and Muench [29].
2.4. Cell Culture
Calu-3 cells were maintained in Minimal Essential Medium (MEM) containing 20% (v/v) inactivated fetal calf serum (FCS), 1 mM l-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin and 1 mM sodium pyruvate.
MDCKII cells were maintained in Dulbecco’s Minimal Essential Medium (DMEM) containing 10% (v/v) inactivated FCS, 1 mM l-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin.
2.5. Assessment of Cell Viability
Viability of uninfected but treated cells was assessed by the water-soluble tetrazolium salt (WST)-1 assay (Cat.: 5015944001, Roche, Penzberg, Germany) according to the manufacturer’s instructions.
2.6. Determination of the Amount of Viral RNA Copies from Released Viruses by qRT-PCR
The amount of viral SARS-CoV-2 RNA copies in the virus–containing samples was quantified by real-time PCR Luna Universal Probe One-Step RT-PCR Kit from New England Biolabs (Cat: E3006L, Ipswich, MA, USA). This kit allows the reverse transcription, cDNA synthesis and PCR amplification in a single step. Samples were analyzed by 7500 software v2.3 (Applied Biosystems, Waltham, MA, USA). PCR primers were designed and used as described previously in [30]. Thereby, the polynucleotide sequence contains parts of the SARS-CoV-2 Envelope (E) and RNA-dependent RNA-polymerase (RdRp) genes and was used as standard for the determination of viral RNA copies in the experiments. The sequences of the used primers were: RdRp_forward (fwd): 5′-GTG-ARA-TGG-TCA-TGT-GTG-GCG-G-3′ and RdRp_reverse (rev) 5′-CAR-ATG-TTA-AAS-ACA-CTA-TTA-GCA-TA-C-3′. Probe was 5′--CAG-GTG-GAA-/ZEN/CCT-CAT-CAG-GAG-ATG-C -3′ (Label: FAM/IBFQ Iowa Black FQ). A dsDNA-polynucleotide sequence (Integrated DNA Technologies, Coralville, IA, USA) was used as a positive control: 5’-TAA-TAC-GAC-TCA-CTA-TAG-GGT-ATT-GAG-TGA-AAT-GGT-CAT-GTG-TGG-CGG-TTC-ACT-ATA-TGT-TAA-ACC-AGG-TGG-AAC-CTC-ATC-AGG-AGA-TGC-CAC-AAC-TGC-TTA-TGC-TAA-TAG-TGT-TTT-TAA-CAT-TTG-GAA-GAG-ACA-GGT-ACG-TTA-ATA-GTT-AAT-AGC-GTA-CTT-CTT-TTT-CTT-GCT-TTC-GTG-GTA-TTC-TTG-CTA-GTT-ACA-CTA-GCC-ATC-CTT-ACT-GCG-CTT-CGA-TTG-TGT-GCG-TAC-TGC-TGC-AAT-ATT-GTT-3’. Generating a series of dilutions (104, 105, 106 and 107 copies/mL) of this standard, the experiments were quantified using a standard curve to obtain absolute values of RNA copies in the sample.
The amount of viral IAV RNA copies in the virus–containing samples was quantified by real-time PCR GoTaq® Probe qPCR one step Kit from Promega (Cat: A6120, Madison, Wisconsin, USA). This kit allows the reverse transcription, cDNA synthesis and PCR amplification in a single step. Samples were analyzed by 7500 software v2.3 (Applied Biosystems, Waltham, MA, USA). PCR primers (Integrated DNA Technologies, Coralville, Iowa, USA) contains parts of the IAV Matrix gene. The sequences of the used primers were: 5’Inf-A-M: 5’-AGA TGA GTC TTC TAA CCG AGG TCG-3’, 3’Inf-A-M: 5’-TGC AAA AAC ATC TTC AAG TCT CTG-3’ and 3’Inf-A-SW-M: 5’-TGC AAA GAC ATC TTC CAG TCT CTG-3’-.
2.7. Software and Statistics
GraphPad Prism 9.0 was used for statistical analyses and to generate graphs. 7500 software v2.3 was used to evaluate the results obtained by qRT-PCR. To determine the combinatory effects of the treatment with EC 3.2. and quinine, the open-source and free web application SynergyFinder was used [31] and the drug interactions were analyzed by the commonly used Bliss independence [32].
3. Results
3.1. European Black Elderberry Fruit Extract and Quinine Exhibits Antiviral Activity Against Influenza A Virus in MDCKII Cells
Although there are several clinical as well as animal studies, which demonstrates its potential anti IAV activity in vivo [12,13,14,15], experiments showing an antiviral activity of black elderberry fruit extract against IAV in vitro have not been reported yet. Concerning quinine, first hints for in vivo activity against IAV was reported in 1946 [28]. To analyze if liquid European black elderberry fruit extract from Sambucus nigra L. (EC 3.2), a water-based liquid extract standardized to contain a minimum of 3.2% anthocyanins, and quinine exhibits antiviral activity against IAV, Madin-Darby Canine Kidney (MDCKII) cells [33], the standard cell culture model for studying IAV replication in vitro, were infected with the IAV isolate A/Puerto Rico/8/34 (PR8) [34] (Figure 1). 30 minutes post infection, serial dilutions of EC 3.2, or various concentrations of quinine were added to the cell cultures and incubated continuously. Two days post infection (dpi), cell culture supernatants were harvested, and virus production was analyzed by quantitative RT-PCR (qRT-PCR) (Figure 1).
Treatment with EC 3.2 or quinine led to a dose dependent inhibition of IAV replication, reducing the production of progeny virions by 80 % at a dilution of 1:200 EC 3.2, and by 90 % with a concentration of 10 µM quinine. DMSO, a solvent control for quinine, had no influence on the replication of IAV (Figure 1B).
Water-soluble tetrazolium salt (WST)-1 assays were conducted in uninfected MDCKII cells under otherwise identical conditions as for the virus infection experiments in order to control for potential unspecific effects of EC 3.2, or quinine, treatment on cell viability. Neither EC 3.2 at dilutions up to 1:100 nor quinine at up to 100 µM, both at concentrations fully blocking IAVPR8 replication, affected cell viability (Figure 2). Staurosporine (1 µM) served as a positive control.
3.2. Combination Treatment with Black Elderberry Fruit Extract and Quinine Exhibits Synergistic Antiviral Activity Against IAV
It has been shown previously, that the combination of two different natural products exerts synergistic antiviral activity, exemplified by testing different extracts from seaweed against measles virus [35]. It was, therefore, legitimate to analyze whether or not combinational treatment with EC 3.2 and quinine has additive or even synergistic antiviral activity against IAVPR-8. MDCKII cells were infected with IAVPR-8 and 30 minutes post infection, different dilutions of EC 3.2 or concentrations quinine alone, or in combination, were added to the cell cultures (Figure 3). Two dpi cell culture supernatants were harvested, and virus production was analyzed by qRT-PCR (Figure 3).
Following treatment with increasing amounts of EC 3.2 (1:800-1:200) in combination with 100 nM of quinine, a concentration representing the IC50 value for inhibition of IAV replication as monotherapy (Figure 1), significant and dose-dependent reduction in replication capacity ranging from 58 - 98 %, from the lowest to highest concentration of EC 3.2 was detected. Treatment with the identical concentration series of EC 3.2 in the presence of 1 µM quinine resulted in even higher antiviral efficacy leading to a reduction of viral replication ranging from 93 - 99 %, from the lowest to highest concentration of EC 3.2 (Figure 3A,B).
In contrast, individual treatment with EC 3.2 or quinine alone had only minor effects on virus replication at concentrations tested (Figure 3A). Single treatment with the lowest tested dilution of EC 3.2 (1:800) has no significant influence on virus replication (Figure 3A), while the highest concentration of quinine (1 µM) reduced viral replication by max. 64 % (Figure 3B). However, combination treatment (1:800 EC 3.2 and 1 µM quinine) almost completely inhibits viral replication, which points towards a synergistic antiviral effect.
To quantify the effects of combination treatment, synergy scoring was conducted using the Bliss independence model [32]. By using this model, independent effects of different small molecules assumed by stochastic processes were analyzed [31,32]. Thereby, a Bliss synergy score >10 indicates synergistic activity, -10 to 10 represents additive effects, and < -10 suggests antagonism [31]. The conducted small molecule interaction analysis resulted in a Bliss synergy score of 14.7 for the combination of EC 3.2 and quinine for the inhibition of the replication of IAVPR-8 (Figure 4).
In summary, the results clearly show that EC 3.2 and quinine exhibits a synergistic effect to inhibit the replication of IAV. Moreover, these data suggest that the combination of these two natural substances may show substantial efficacy when administered in combination at relatively low doses.
Water-soluble tetrazolium salt (WST)-1 assays were conducted in uninfected MDCKII cells under otherwise identical conditions as for the virus infection experiments in order to control for potential unspecific effects of EC 3.2, or quinine, treatment on cell viability. Thereby, it could be demonstrated that the combinational treatment with EC 3.2 and quinine had no impact on cell viability in all tested dilutions or concentrations (Figure 5).
3.3. Treatment with a Combination of European Black Elderberry Fruit Extract and Quinine Exhibits Synergistic Antiviral Activity Against SARS-CoV-2
We previously showed that the single treatment of Calu-3 cells with EC 3.2 or quinine efficiently inhibits the replication capacity of SARS-CoV-2 in a dose-dependent manner [16,20]. Thus, we next wanted to analyze if the combinational treatment with these two natural substances has, as shown for IAV (Figure 3 and Figure 4), also an additive or even synergistic antiviral activity against SARS-CoV-2. Therefore, Calu-3 cells, exemplifying the most comprehensively studied surrogate lung cell infection model that expresses ACE2 and TMPRSS2 endogenously [36], were infected with SARS-CoV-2 Wuhan type. Various dilutions of EC 3.2 or concentrations of quinine alone, or in combination, were added to the cell cultures one hour post infection, (Figure 6). Two dpi cell culture supernatants were harvested, and virus production was analyzed by qRT-PCR (Figure 6).
Following treatment with increasing amounts of EC 3.2 (1:3200-1:800) in combination with 1 µM of quinine, a concentration representing the IC50 value for inhibition of SARS-CoV-2 replication [16,20] results in a complete block of the replication capacity, independent from the dilution of EC 3.2. Treatment with the identical dilution series of EC 3.2 in the presence of 100 nM quinine although resulted in a very high antiviral efficacy leading to a reduction of viral replication ranging from 80 - 97 %, from the lowest to highest concentration of EC 3.2 (Figure 6A,B).
By contrast, treatment with EC 3.2 (1:3200) alone reduced replication by only 19%, and 10 nM quinine alone had no significant effect. (Figure 6A). However, combination treatment (1:3200 EC 3.2 and 10 nM quinine) inhibits viral replication by 65-87%, which again clearly points towards a strong synergistic antiviral effect.
To analyze if the combinational treatment of EC 3.2 and quinine exhibits synergistic antiviral activity on SARS-CoV-2 Wuhan type the Bliss independence model was used again [6]. The conducted small molecule interaction analysis resulted in a Bliss synergy score of 27.8 for the combination of EC 3.2 and quinine for the inhibition of the replication of SARS-CoV-2 Wuhan type (Figure 7).
In summary, the results clearly show that the combinatorial treatment of EC 3.2 and quinine, in addition to IAV (Figure 3 and Figure 4), although exhibits a very strong synergistic effect in the inhibition of the replication of SARS-CoV-2.
To verify that the combined treatment of EC 3.2 and quinine did not exert nonspecific cytotoxic effects, WST-1 assays were performed on uninfected Calu-3 cells under the same conditions as the infection experiments. No loss of cell viability was observed for any of the tested dilutions or concentrations (Figure 8). Staurosporine (1 µM) served as a positive control.
3.4. Antiviral Activity of ElderCraft® Q-Complex Against IAV and SARS-CoV-2 in Comparison to ElderCraft® Without Quinine
Based on our findings that both, an anthocyanin-rich elderberry fruit extract (EC 3.2) and quinine possess antiviral properties against IAV and SARS-CoV-2 (Figure 1 and [16,20]) and that these agents acting synergistically (Figures 3, 4, 6 and 7), we aimed to analyze a commercially available product that integrates these two compounds into a single formulation, christened ElderCraft® Q-Complex. By evaluating ElderCraft® Q-Complex, we aimed to determine its efficacy as a practical and accessible antiviral agent, potentially offering enhanced synergistic effects in the inhibition of viral replication.
Therefore, we compared the antiviral activity of ElderCraft® Q-Complex with ElderCraft® (EC 15, 15% anthocyanins) alone following infection of MDCKII cells with IAVPR-8 (Figure 9A,B) or Calu-3 cells with SARS-CoV-2 Wuhan type (Figure 9C,D). ElderCraft® Q-Complex contains similar amount of anthocyanins as EC 15, however, in addition, quinine was added in an amount of 10 mg per 15 mg anthocyanins. The infection with IAVPR-8 or SARS-CoV-2 Wuhan type was performed as described before (Figure 1, Figure 3 and Figure 6) and qRT-PCR analysis were conducted (Figure 9).
Following treatment with increasing amounts of EC 15 (1:3200-1:200) a dose-dependent reduction of the replication of IAV (Figure 9A) or SARS-CoV-2 (Figure 9C) was detectable resuling in an IC50 of ~ 1:1000 for both, IAV and SARS-CoV-2. Treatment with the identical dilution series of ElderCraft® Q-Complex, were additionally quinine was added resulted in an IC50 of ~ 1:3000 for the inhibition of the replication of IAV (Figure 9B) and an IC50 ~1:3200 for SARS-CoV-2 (Figure 9D). Thus, it could be concluded that an combinational formulation of black elderberry fruit extract and quinine resulted in an about 3-fold better reduction of virus replication in comparison to the treatement with black elderberry fruit extract alone.
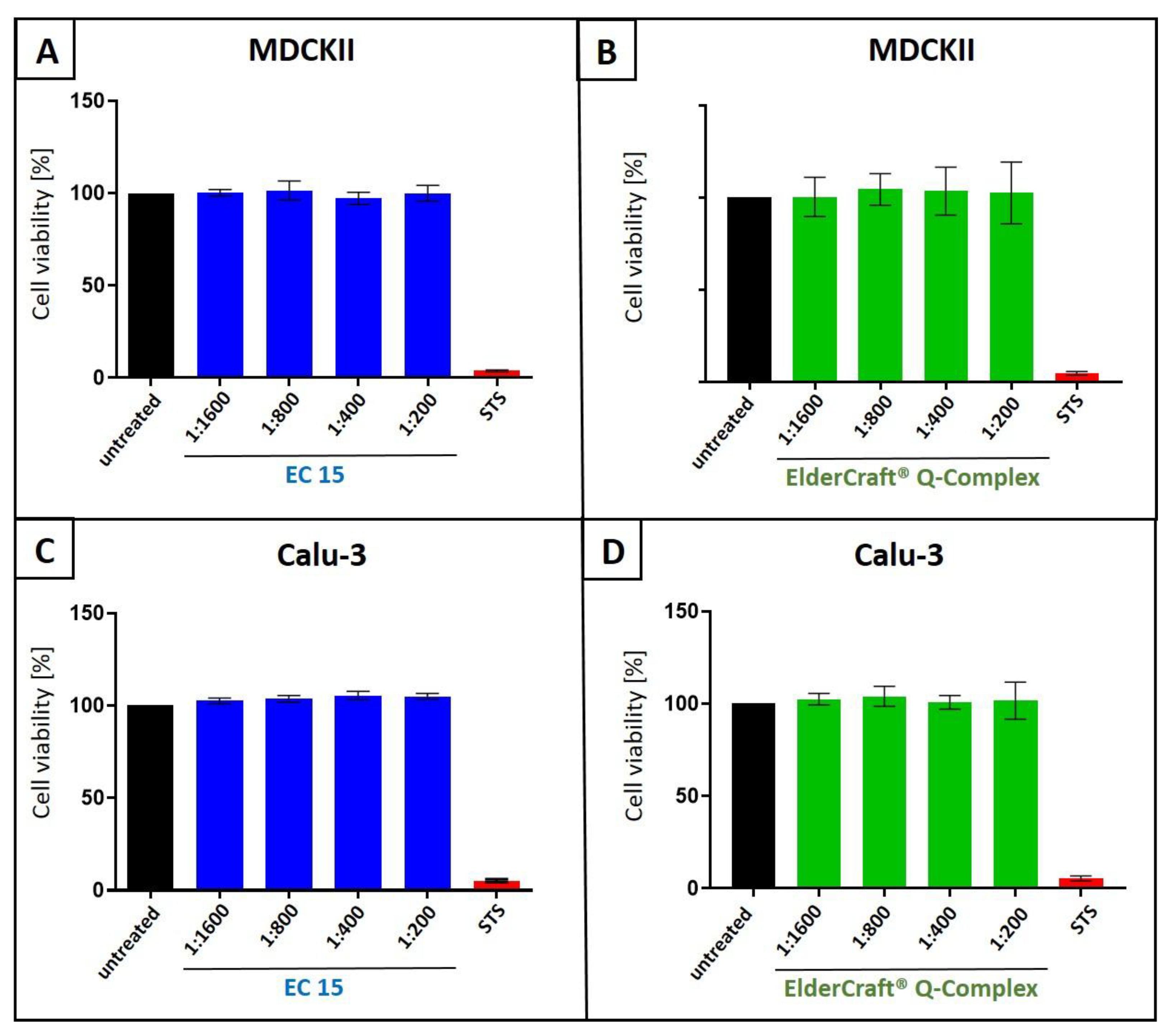
To assess potential nonspecific effects of EC 15 or ElderCraft® Q-Complex treatment on cell viability, WST-1 assays were performed on uninfected MDCKII (Figure 10A,B) and Calu-3 cells (Figure 10C,D) under conditions identical to the infection experiments. No cytotoxicity was observed at any tested dilution (Figure 10). Staurosporine (1 µM) served as a positive control.
4. Discussion
The continuous emergence of new variants of SARS-CoV-2 as well as IAV, have been posing serious global health and socioeconomic challenges. Moreover, it remains highly likely that new viruses or virus variants capable of causing pandemic threats will emerge in the future, as it was previously the case for MERS, SARS-CoV-2, and various historical IAV pandemics, including the Spanish, Asian, Hong Kong, and Russian flu. In particular, RNA viruses such as corona- and orthomyxoviruses exhibit high mutation rates due to the error-prone viral RNA-dependent RNA polymerase or the shift of viral genome sequence, providing one reason for the extremely high evolutionary dynamic of both, SARS-CoV-2 and IAV [37]. This situation underscores the urgent need for the development of new antiviral agents as part of a comprehensive pandemic preparedness strategy.
Although effective, existing antiviral treatment options for SARS-CoV-2 as well as IAV infections encounter some remaining problems: They have to be administered early after infection and, moreover, development of drug resistance have been observed frequently as well as side effects. Additionally, those drugs are expensive and therefore unavailable to a large part of the world’s population [38,39]. In the last years, antiviral activity in vitro and in vivo was reported for several natural products, and some of them entered clinical studies [38,40,41]. One example is iota-carrageenan, a high molecular weight sulfated polymer extracted from red seaweed. It was shown that iota-carrageenan exhibits antiviral activity against several respiratory viruses, among them IAV and SARS-CoV-2 [42,43,44,45,46,47]. Most importantly, during a randomized, placebo-controlled, double-blinded, multicenter clinical study a relative risk reduction of 79.8% for SARS-CoV-2 infections could achieved, when healthcare workers in a COVID-19 station used a nasal spray containing iota-carrageenan [48]. Moreover, another clinical trial with a nasal spray containing iota-carrageenan and Ivermectin demonstrated a reduction of disease severity following SARS-CoV-2 infection [49]. Similar to carrageenans, most natural products are easily distributable, have significantly fewer side effects than chemically synthesized drugs, are broadly active and exhibit a low risk for the development of resistance as they mostly inhibit host cell targets [38].
This study demonstrates that the combination of the two natural substances—European black elderberry fruit extract and quinine—exhibits a strong synergistic antiviral effect against SARS-CoV-2 and IAV in vitro without affecting cell viability. The results suggests that this combinational treatment might represent an effective treatment option against various respiratory RNA viruses and variants thereof.
Juices and different fruit extracts and have previously been demonstrated to represent potential sources for antiviral agents. Thereby, extracts from fruits such as blackberry, blackcurrant, mulberry, and pomegranate show antiviral activity against a variety of viruses, including Dengue virus, SARS-CoV-2, Hepatitis C virus, Poliovirus, IAV, Zika virus, and Human immunodeficiency virus type 1 [50].
Sambucus nigra, commonly known as European black elderberry, has been utilized in traditional medicine for centuries to alleviate symptoms associated with viral infections. Historically, its use has been particularly prevalent in managing upper respiratory infections [51]. Over the last three decades, the therapeutic potential of black elderberry has gained scientific validation through several clinical trials. These studies have consistently demonstrated that extracts from black elderberry fruits are effective in reducing both the duration and severity of upper respiratory infections [13,14,52]. In parallel with clinical findings, numerous in vitro studies have sought to elucidate the mechanisms underlying properties of black elderberry.
Three main hypotheses have emerged from this body of research: (i) specific components within black elderberry, such as flavonoids and phenolic acids, are believed to exert direct antiviral effects by interfering with the viral lifecycle [16,19,53,54]; (ii) another proposed mechanism involves the inhibition of viral enzymes critical for replication and proliferation, thereby impeding the virus’s ability to multiply within the host [18,55]; and (iii) polysaccharides present in black elderberry are thought to play a crucial role in stimulating the immune system, enhancing the body’s natural defense mechanisms against viral pathogens [56,57].
Quinine has been previously shown to exert antiviral activity against various different viruses [16,24,25,26,27,28]. Mechanistically, mainly three different hypotheses for its antiviral activity are discussed: (i) quinine was shown to be a weak base and thus is able to increase intracellular the pH of acidic organelles, e.g., endosomes or lysosomes [58]. This step is crucial for the receptor-mediated endocytosis of different viruses and could therefore be one explanation for the antiviral activity of quinine in the early stages of virus replication [21,59]. (ii) It was demonstrated that quinine enhances the synthesis and thus the release of IFN-α. The released IFN-α then binds to the IFN-α –receptor and thereby stimulates the 2-5 (A) synthetase that activates RNAse L. The RNAse L then degrades viral RNA and thus inhibits the replication of different RNA viruses, such as SARS-CoV-2 and IAV [21,60]. (iii) Quinine also exhibits immunoregulatory properties, which can contribute to its antiviral activity. It was shown that quinine causes a reduction of inflammatory response by inhibiting the production of pro-inflammatory factors [21,61].
As quinine is one of the oldest used drugs, its pharmacokinetics have been well elaborated. In the European Union its actually allowed to add quinine up to 100 mg/kg to food or up to 85 mg/L to beverages and up to 250 mg/L to alcoholic beverages [62]. For example, in 1 l of tonic water, 85 mg quinine are present, which leads to a plasma concentration of ~0.5 µg/mL, which is equal to a molarity of ~1.5 µM [63,64]. One tablet of quinine, which is approved for the treatment of calf cramps, contains 200 mg quinine sulfate, which correlates to a plasma concentration of ~2.9 µM quinine. However, our combinational experiments clearly show, that ~100 nM quinine in combination with elderberry fruit extract is able to completely block the replication of SARS-CoV-2 or IAV in vitro (Figure 3 and 6). Thus, very low amounts of quinine in combination with elderberry fruit extract would be sufficient to reach values that exhibits antiviral efficacy in our in vitro systems.
A possible explanation for the strong synergetic antiviral activity of the combinational treatment of elderberry fruit extract and quinine (Figure 3, Figure 4 and Figure 9) is the fact that they target different steps of viral life cycle. Synergistic activity of chemical synthesized drugs has been shown previously for SARS-CoV-2 and IAV [65]. For instance, in the case of IAV, Favipiravir, a viral RNA-polymerase inhibitor was tested in combination with Oseltamivir, a neuroaminidase-inhibitor. Thereby, it was shown that these two small molecules exhibit synergistic antiviral activity, most likely by acting at different stages of the IAV replication cycle [66]. For SARS-CoV-2 a promising synergistic antiviral activity was shown for the polymerase inhibitor Molnupiravir in combination with Tilorone, an interferon-inductor [67]. However, a synergistic activity using two natural products against IAV or SARS-CoV-2 has not been shown yet.
For black elderberry extract, it was demonstrated before, that e.g., flavonoids, which are present at significant amounts in the extract used in this study [16], are able to inhibit the entry of IAV by directly binding to the virus [19]. In addition, isoquercetin, another component of the black elderberry extract, blocks the attachment of IAV at the cell [68]. Both of these mechanisms acting at stages of virus replication earlier than these from quinine. On the other hand, in the case of SARS-CoV-2 it is unlikely that black elderberry extract act at early stages of virus replication, as we could show previously, that the extract has no influence on SARS-CoV-2 replication at early stage [16]. Some reports showing that anthocyanins or phenolic compounds present in black elderberry extract can inhibit later steps in virus replication. For instance, it was described that these compounds block the active pocket of the neuraminidase of IAV or the papain-like protease of SARS-CoV-2 and thus act during late stages of virus replication [18,69]. In addition to its antiviral activity, both natural products, black elderberry extract as well as quinine were shown to modulate the immune system and thereby might have an influence on the spread of infection in vivo [21,56,57,61].
The combination of standardized elderberry fruit extract and quinine offers several additional advantages, in terms of cost and efficacy. Standardized extracts rich in anthocyanins and polyphenols, derived from fruits, are relatively expensive. Human studies indicate that a daily dose of 60 mg to 90 mg of anthocyanins is required for effective treatment and prevention, making these natural products costly and often unaffordable for the general population. In general, Elderberry fruit extract is well tolerated, even at high doses, with no known issues related to tolerability. However, the stability of elderberry extract in liquid formulations presents a challenge. Anthocyanins degrade over time in aqueous conditions, necessitating over-dosing by manufacturers to compensate for the degradation [70]. This practice further increases the cost per dose. While our data suggests that EC 3.2 is chemically stable under storage conditions and likely under the experimental conditions used, we acknowledge that anthocyanin degradation could occur to some extent and this issue warrants further investigation in future studies.
In addition to elderberry fruit extract, similar antiviral activities have to be expected in extracts obtained from other plant components of Sambucus nigra, like flower or bark extracts, which will be subject of further studies.
The search for solutions to reduce the daily required dose of polyphenols has led to exploring the synergy between quinine and black elderberry fruit extract. Our findings demonstrate that the combination of quinine with elderberry extract can enhance the efficacy of the antiviral effect (Figure 3, Figure 4 and Figure 9). This synergy allows for a reduction in the required daily dose of polyphenols while maintaining, or even improving, the therapeutic benefits. This reduction not only makes the treatment more cost-effective but also enhances its accessibility for a broader population. Additionally, by improving stability and reducing the need for over-dosing, the combination minimizes the economic burden on manufacturers and consumers alike.
5. Patents
Iprona Lana SpA has filed a PCT and EP patent entitled “Combination of Elderberry Extract and Quinine for the Prevention and Treatment of RNA Virus Infections” claiming the priority date of 31.08.2023.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org.
Author Contributions
Conceptualization, U.S., C.S., S.P. and S.B.; methodology, C.S., P.R. and M.S.; validation, C.S., U.S. and M.S.; investigation, C.S., P.R., M.S. and U.S.; data curation, U.S.; writing—original draft preparation, C.S., M.S, S.B., S.P and U.S; writing—review and editing, C.S., P.R., M.S. and U.S; visualization, C.S. and S.P.; supervision, U.S.; project administration, C.S., S.B., S.P. and U.S.; funding acquisition, U.S and S.B.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation)—401821119/GRK2504 to U.S.. Iprona Lana SpA partially provide funding for consumable costs.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data from each individual experiment carried out in this study is included in Supplementary Table S2.
Acknowledgments
We thank Matthias Tenbusch for providing reagents and protocols, Dennis Lapuente for the fruitful discussions and advices regarding methodology and Pascal Irrgang for the technical advices regarding the generation of IAV stocks and infection experiments.
Conflicts of Interest
The authors have read the journal’s policy and declare that the authors Stephan Plattner and Stephan Breitenberger are employed by Iprona Lana SpA. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data or in the decision to publish the results. All other authors declare no conflict of interest.
References
- Williams, B. A.; Jones, C. H.; Welch, V.; True, J. M. , Outlook of pandemic preparedness in a post-COVID-19 world. npj Vaccines 2023, (1), 178. [Google Scholar] [CrossRef] [PubMed]
- Wibmer, C. K.; Ayres, F.; Hermanus, T.; Madzivhandila, M.; Kgagudi, P.; Oosthuysen, B.; Lambson, B. E.; de Oliveira, T.; Vermeulen, M.; van der Berg, K.; Rossouw, T.; Boswell, M.; Ueckermann, V.; Meiring, S.; von Gottberg, A.; Cohen, C.; Morris, L.; Bhiman, J. N.; Moore, P. L., SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nature Medicine 2021, 27, (4), 622-625.
- Blumental, S.; Debré, P. , Challenges and Issues of Anti-SARS-CoV-2 Vaccines. Frontiers in medicine 2021, 8, 664179. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, R.; Singh, A.; Kumar, S., SARS-CoV-2: Immunity, Challenges with Current Vaccines, and a Novel Perspective on Mucosal Vaccines. Vaccines 2023, 11, (4).
- Chan, J. F.-W.; Yuan, S.; Chu, H.; Sridhar, S.; Yuen, K.-Y. , COVID-19 drug discovery and treatment options. Nature Reviews Microbiology 2024, (7), 391–407. [Google Scholar] [CrossRef] [PubMed]
- Javanian, M.; Barary, M.; Ghebrehewet, S.; Koppolu, V.; Vasigala, V.; Ebrahimpour, S. , A brief review of influenza virus infection. Journal of medical virology 2021, (8), 4638–4646. [Google Scholar] [CrossRef]
- Uyeki, T. M.; Hui, D. S.; Zambon, M.; Wentworth, D. E.; Monto, A. S. , Influenza. The Lancet 2022, (10353), 693–706. [Google Scholar] [CrossRef]
- Comber, L.; E, O. M.; Jordan, K.; Hawkshaw, S.; Marshall, L.; O’Neill, M.; Teljeur, C.; Ryan, M.; Carnahan, A.; Pérez Martín, J. J.; Robertson, A. H.; Johansen, K.; de Jonge, J.; Krause, T.; Nicolay, N.; Nohynek, H.; Pavlopoulou, I.; Pebody, R.; Penttinen, P.; Soler-Soneira, M.; Wichmann, O.; Harrington, P. , Systematic review of the efficacy, effectiveness and safety of high-dose seasonal influenza vaccines for the prevention of laboratory-confirmed influenza in individuals ≥18 years of age. Reviews in medical virology 2023, (3), e2330. [Google Scholar] [CrossRef]
- Chaachouay, N.; Zidane, L., Plant-Derived Natural Products: A Source for Drug Discovery and Development. In Drugs and Drug Candidates, 2024; Vol. 3, pp 184-207.
- Dias, D. A.; Urban, S.; Roessner, U. , A historical overview of natural products in drug discovery. Metabolites 2012, (2), 303–36. [Google Scholar] [CrossRef]
- Atanasov, A. G.; Zotchev, S. B.; Dirsch, V. M.; Orhan, I. E.; Banach, M.; Rollinger, J. M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E. A.; et al. Natural products in drug discovery: advances and opportunities. Nature Reviews Drug Discovery 2021, (3), 200–216. [Google Scholar] [CrossRef]
- Hawkins, J.; Baker, C.; Cherry, L.; Dunne, E. , Black elderberry (Sambucus nigra) supplementation effectively treats upper respiratory symptoms: A meta-analysis of randomized, controlled clinical trials. Complementary therapies in medicine 2019, 42, 361–365. [Google Scholar] [CrossRef]
- Tiralongo, E.; Wee, S. S.; Lea, R. A. , Elderberry Supplementation Reduces Cold Duration and Symptoms in Air-Travellers: A Randomized, Double-Blind Placebo-Controlled Clinical Trial. Nutrients 2016, (4), 182. [Google Scholar] [CrossRef]
- Zakay-Rones, Z.; Thom, E.; Wollan, T.; Wadstein, J. , Randomized study of the efficacy and safety of oral elderberry extract in the treatment of influenza A and B virus infections. The Journal of international medical research 2004, (2), 132–40. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, E.; Hayashi, K.; Katayama, H.; Hayashi, T.; Obata, A., Anti-influenza virus effects of elderberry juice and its fractions. Bioscience, biotechnology, and biochemistry 2012, 76, (9), 1633-8.
- Setz, C.; Fröba, M.; Große, M.; Rauch, P.; Auth, J.; Steinkasserer, A.; Plattner, S.; Schubert, U. , European Black Elderberry Fruit Extract Inhibits Replication of SARS-CoV-2 In Vitro. Nutraceuticals 2023, (1), 91–106. [Google Scholar] [CrossRef]
- Chen, C.; Zuckerman, D. M.; Brantley, S.; Sharpe, M.; Childress, K.; Hoiczyk, E.; Pendleton, A. R. , Sambucus nigra extracts inhibit infectious bronchitis virus at an early point during replication. BMC veterinary research 2014, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, K.; Dyason, J. C.; Maggioni, A.; von Itzstein, M.; Downard, K. M. , Binding of a natural anthocyanin inhibitor to influenza neuraminidase by mass spectrometry. Analytical and bioanalytical chemistry 2013, (20), 6563–72. [Google Scholar] [CrossRef]
- Roschek, B., Jr.; Fink, R. C.; McMichael, M. D.; Li, D.; Alberte, R. S. , Elderberry flavonoids bind to and prevent H1N1 infection in vitro. Phytochemistry 2009, (10), 1255–61. [Google Scholar] [CrossRef]
- Große, M.; Ruetalo, N.; Layer, M.; Hu, D.; Businger, R.; Rheber, S.; Setz, C.; Rauch, P.; Auth, J.; Fröba, M.; Brysch, E.; Schindler, M.; Schubert, U., Quinine Inhibits Infection of Human Cell Lines with SARS-CoV-2. Viruses 2021,13, (4).
- Latarissa, I. R.; Barliana, M. I.; Meiliana, A.; Lestari, K., Potential of Quinine Sulfate for COVID-19 Treatment and Its Safety Profile: Review. Clinical pharmacology : advances and applications 2021, 13, 225-234.
- Miller, L. H.; Rojas-Jaimes, J.; Low, L. M.; Corbellini, G. , What Historical Records Teach Us about the Discovery of Quinine. The American journal of tropical medicine and hygiene 2023, (1), 7–11. [Google Scholar] [CrossRef]
- Achan, J.; Talisuna, A. O.; Erhart, A.; Yeka, A.; Tibenderana, J. K.; Baliraine, F. N.; Rosenthal, P. J.; D’Alessandro, U. , Quinine, an old anti-malarial drug in a modern world: role in the treatment of malaria. Malaria journal 2011, 10, 144. [Google Scholar] [CrossRef]
- Khan, S. A.; Al-Balushi, K. , Combating COVID-19: The role of drug repurposing and medicinal plants. Journal of infection and public health 2021, (4), 495–503. [Google Scholar] [CrossRef]
- Malakar, S.; Sreelatha, L.; Dechtawewat, T.; Noisakran, S.; Yenchitsomanus, P.-t.; Chu, J. J. H.; Limjindaporn, T. , Drug repurposing of quinine as antiviral against dengue virus infection. Virus Research 2018, 255, 171–178. [Google Scholar] [CrossRef]
- D’Alessandro, S.; Scaccabarozzi, D.; Signorini, L.; Perego, F.; Ilboudo, D. P.; Ferrante, P.; Delbue, S., The Use of Antimalarial Drugs against Viral Infection. Microorganisms 2020, 8, (1).
- Baroni, A.; Paoletti, I.; Ruocco, E.; Ayala, F.; Corrado, F.; Wolf, R.; Tufano, M. A.; Donnarumma, G. , Antiviral effects of quinine sulfate on HSV-1 HaCat cells infected: Analysis of the molecular mechanisms involved. Journal of Dermatological Science 2007, (3), 253–255. [Google Scholar] [CrossRef]
- Seeler, A. O.; Graessle, O.; Ott, W. H. , Effect of Quinine on Influenza Virus Infections in Mice. The Journal of Infectious Diseases 1946, (2), 156–158. [Google Scholar] [CrossRef] [PubMed]
- Reed, L. J. M., H. , A Simple Method Of Estimating Fifty Per Cent Endpoints. American Journal of Epidemiology 1936, (3), 493–497. [Google Scholar]
- Corman, V. M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D. K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M. L.; Mulders, D. G.; Haagmans, B. L.; van der Veer, B.; van den Brink, S.; Wijsman, L.; Goderski, G.; Romette, J. L.; Ellis, J.; Zambon, M.; Peiris, M.; Goossens, H.; Reusken, C.; Koopmans, M. P.; Drosten, C., Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro surveillance : bulletin Europeen sur les maladies transmissibles = European communicable disease bulletin 2020, 25, (3).
- Ianevski, A.; He, L.; Aittokallio, T.; Tang, J., SynergyFinder: a web application for analyzing drug combination dose-response matrix data. Bioinformatics (Oxford, England) 2017, 33, (15), 2413-2415.
- Liu, Q.; Yin, X.; Languino, L. R.; Altieri, D. C. , Evaluation of drug combination effect using a Bliss independence dose-response surface model. Statistics in biopharmaceutical research 2018, (2), 112–122. [Google Scholar] [CrossRef] [PubMed]
- Gaush, C. R.; Hard, W. L.; Smith, T. F., Characterization of an established line of canine kidney cells (MDCK). Proceedings of the Society for Experimental Biology and Medicine. Society for Experimental Biology and Medicine (New York, N.Y.) 1966, 122, (3), 931-5.
- Wit, E. d.; Spronken, M. I. J.; Bestebroer, T. M.; Rimmelzwaan, G. F.; Osterhaus, A. D. M. E.; Fouchier, R. A. M. , Efficient generation and growth of influenza virus A/PR/8/34 from eight cDNA fragments. Virus Research 2004, (1), 155–161. [Google Scholar] [CrossRef]
- Morán-Santibañez, K.; Peña-Hernández, M. A.; Cruz-Suárez, L. E.; Ricque-Marie, D.; Skouta, R.; Vasquez, A. H.; Rodríguez-Padilla, C.; Trejo-Avila, L. M., Virucidal and Synergistic Activity of Polyphenol-Rich Extracts of Seaweeds against Measles Virus. Viruses 2018, 10, (9).
- Aguiar, J. A.; Tremblay, B. J.; Mansfield, M. J.; Woody, O.; Lobb, B.; Banerjee, A.; Chandiramohan, A.; Tiessen, N.; Cao, Q.; Dvorkin-Gheva, A.; Revill, S.; Miller, M. S.; Carlsten, C.; Organ, L.; Joseph, C.; John, A.; Hanson, P.; Austin, R. C.; McManus, B. M.; Jenkins, G.; Mossman, K.; Ask, K.; Doxey, A. C.; Hirota, J. A. , Gene expression and in situ protein profiling of candidate SARS-CoV-2 receptors in human airway epithelial cells and lung tissue. The European respiratory journal.
- Owen, L.; Laird, K.; Shivkumar, M. , Antiviral plant-derived natural products to combat RNA viruses: Targets throughout the viral life cycle. Letters in Applied Microbiology 2022, (3), 476–499. [Google Scholar] [CrossRef]
- Musarra-Pizzo, M.; Pennisi, R.; Ben-Amor, I.; Mandalari, G.; Sciortino, M. T., Antiviral Activity Exerted by Natural Products against Human Viruses. Viruses 2021, 13, (5).
- Antonelli, G.; Turriziani, O. , Antiviral therapy: old and current issues. International Journal of Antimicrobial Agents 2012, (2), 95–102. [Google Scholar] [CrossRef]
- Özçelik, B.; Kartal, M.; Orhan, I. , Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharmaceutical Biology 2011, (4), 396–402. [Google Scholar] [CrossRef]
- Mohammadi Pour, P.; Fakhri, S.; Asgary, S.; Farzaei, M. H.; Echeverría, J. , The Signaling Pathways, and Therapeutic Targets of Antiviral Agents: Focusing on the Antiviral Approaches and Clinical Perspectives of Anthocyanins in the Management of Viral Diseases. Frontiers in pharmacology 2019, 10, 1207. [Google Scholar] [CrossRef]
- Grassauer, A.; Weinmuellner, R.; Meier, C.; Pretsch, A.; Prieschl-Grassauer, E.; Unger, H. , Iota-Carrageenan is a potent inhibitor of rhinovirus infection. Virol J 2008, 5, 107. [Google Scholar] [CrossRef]
- Leibbrandt, A.; Meier, C.; König-Schuster, M.; Weinmüllner, R.; Kalthoff, D.; Pflugfelder, B.; Graf, P.; Frank-Gehrke, B.; Beer, M.; Fazekas, T.; Unger, H.; Prieschl-Grassauer, E.; Grassauer, A. , Iota-carrageenan is a potent inhibitor of influenza A virus infection. PloS one 2010, (12), e14320. [Google Scholar] [CrossRef]
- Morokutti-Kurz, M.; Graf, C.; Prieschl-Grassauer, E. , Amylmetacresol/2,4-dichlorobenzyl alcohol, hexylresorcinol, or carrageenan lozenges as active treatments for sore throat. Int J Gen Med 2017, 10, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Morokutti-Kurz, M.; Fröba, M.; Graf, P.; Große, M.; Grassauer, A.; Auth, J.; Schubert, U.; Prieschl-Grassauer, E. , Iota-carrageenan neutralizes SARS-CoV-2 and inhibits viral replication in vitro. PLOS ONE 2021, (2), e0237480. [Google Scholar] [CrossRef] [PubMed]
- Schütz, D.; Conzelmann, C.; Fois, G.; Groß, R.; Weil, T.; Wettstein, L.; Stenger, S.; Zelikin, A.; Hoffmann, T. K.; Frick, M.; Müller, J. A.; Münch, J., Carrageenan-containing over-the-counter nasal and oral sprays inhibit SARS-CoV-2 infection of airway epithelial cultures. American Journal of Physiology-Lung Cellular and Molecular Physiology 2021, 320, (5), L750-L756.
- Fröba, M.; Große, M.; Setz, C.; Rauch, P.; Auth, J.; Spanaus, L.; Münch, J.; Ruetalo, N.; Schindler, M.; Morokutti-Kurz, M.; Graf, P.; Prieschl-Grassauer, E.; Grassauer, A.; Schubert, U., Iota-Carrageenan Inhibits Replication of SARS-CoV-2 and the Respective Variants of Concern Alpha, Beta, Gamma and Delta. International journal of molecular sciences 2021, 22, (24).
- Figueroa, J. M.; Lombardo, M. E.; Dogliotti, A.; Flynn, L. P.; Giugliano, R.; Simonelli, G.; Valentini, R.; Ramos, A.; Romano, P.; Marcote, M.; Michelini, A.; Salvado, A.; Sykora, E.; Kniz, C.; Kobelinsky, M.; Salzberg, D. M.; Jerusalinsky, D.; Uchitel, O. , Efficacy of a Nasal Spray Containing Iota-Carrageenan in the Postexposure Prophylaxis of COVID-19 in Hospital Personnel Dedicated to Patients Care with COVID-19 Disease. Int J Gen Med 2021, 14, 6277–6286. [Google Scholar] [CrossRef]
- Chahla, R. E.; Medina Ruiz, L.; Ortega, E. S.; Morales Rn, M. F.; Barreiro, F.; George, A.; Mancilla Rn, C.; S, D. A. R.; Barrenechea, G.; Goroso, D. G.; Peral de Bruno, M. , Intensive Treatment With Ivermectin and Iota-Carrageenan as Pre-exposure Prophylaxis for COVID-19 in Health Care Workers From Tucuman, Argentina. Am J Ther 2021, (5), e601–e604. [Google Scholar] [CrossRef]
- Santhi, V. P.; Sriramavaratharajan, V.; Murugan, R.; Masilamani, P.; Gurav, S. S.; Sarasu, V. P.; Parthiban, S.; Ayyanar, M., Edible fruit extracts and fruit juices as potential source of antiviral agents: a review. Food Measure. 2021;15(6):5181-90. Epub 2021 Aug 3. [CrossRef]
- Ulbricht, C.; Basch, E.; Cheung, L.; Goldberg, H.; Hammerness, P.; Isaac, R.; Khalsa, K. P.; Romm, A.; Rychlik, I.; Varghese, M.; Weissner, W.; Windsor, R. C.; Wortley, J. , An evidence-based systematic review of elderberry and elderflower (Sambucus nigra) by the Natural Standard Research Collaboration. Journal of dietary supplements 2014, (1), 80–120. [Google Scholar] [CrossRef]
- Zakay-Rones, Z.; Varsano, N.; Zlotnik, M.; Manor, O.; Regev, L.; Schlesinger, M.; Mumcuoglu, M., Inhibition of several strains of influenza virus in vitro and reduction of symptoms by an elderberry extract (Sambucus nigra L.) during an outbreak of influenza B Panama. Journal of alternative and complementary medicine (New York, N.Y.) 1995, 1, (4), 361-9.
- Krawitz, C.; Mraheil, M. A.; Stein, M.; Imirzalioglu, C.; Domann, E.; Pleschka, S.; Hain, T. , Inhibitory activity of a standardized elderberry liquid extract against clinically-relevant human respiratory bacterial pathogens and influenza A and B viruses. BMC complementary and alternative medicine 2011, 11, 16. [Google Scholar] [CrossRef]
- Porter, R. S.; Bode, R. F., A Review of the Antiviral Properties of Black Elder (Sambucus nigra L.) Products. Phytotherapy research : PTR 2017, 31, (4), 533-554.
- Swaminathan, K.; Müller, P.; Downard, K. M. , Substituent effects on the binding of natural product anthocyanidin inhibitors to influenza neuraminidase with mass spectrometry. Analytica chimica acta 2014, 828, 61–9. [Google Scholar] [CrossRef]
- Stich, L.; Plattner, S.; McDougall, G.; Austin, C.; Steinkasserer, A., Polysaccharides from European Black Elderberry Extract Enhance Dendritic Cell Mediated T Cell Immune Responses. International journal of molecular sciences 2022, 23, (7).
- Ho, G. T.; Ahmed, A.; Zou, Y. F.; Aslaksen, T.; Wangensteen, H.; Barsett, H. , Structure-activity relationship of immunomodulating pectins from elderberries. Carbohydrate polymers 2015, 125, 314–22. [Google Scholar] [CrossRef]
- Bray, P. G.; Mungthin, M.; Hastings, I. M.; Biagini, G. A.; Saidu, D. K.; Lakshmanan, V.; Johnson, D. J.; Hughes, R. H.; Stocks, P. A.; O’Neill, P. M.; Fidock, D. A.; Warhurst, D. C.; Ward, S. A. , PfCRT and the trans-vacuolar proton electrochemical gradient: regulating the access of chloroquine to ferriprotoporphyrin IX. Molecular microbiology 2006, (1), 238–51. [Google Scholar] [CrossRef]
- Mauthe, M.; Orhon, I.; Rocchi, C.; Zhou, X.; Luhr, M.; Hijlkema, K. J.; Coppes, R. P.; Engedal, N.; Mari, M.; Reggiori, F. , Chloroquine inhibits autophagic flux by decreasing autophagosome-lysosome fusion. Autophagy 2018, (8), 1435–1455. [Google Scholar] [CrossRef]
- Nugraha, R. V.; Ridwansyah, H.; Ghozali, M.; Khairani, A. F.; Atik, N., Traditional Herbal Medicine Candidates as Complementary Treatments for COVID-19: A Review of Their Mechanisms, Pros and Cons. Evidence-Based Complementary and Alternative Medicine 2020, 2020, (1), 2560645.
- Savarino, A.; Boelaert, J. R.; Cassone, A.; Majori, G.; Cauda, R. , Effects of chloroquine on viral infections: an old drug against today’s diseases? The Lancet. Infectious diseases 2003, (11), 722–7. [Google Scholar] [CrossRef]
- law, E. U. Regulation (EC) No 1334/2008 of the European Parliament and of the Council of 16 December 2008 on flavourings and certain food ingredients with flavouring properties for use in and on foods and amending Council Regulation (EEC) No 1601/91, Regulations (EC) No 2232/96 and (EC) No 110/2008 and Directive 2000/13/EC. https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:02008R1334-20190521.
- Hall, A. P.; Czerwinski, A. W.; Madonia, E. C.; Evensen, K. L. , Human plasma and urine quinine levels following tablets, capsules, and intravenous infusion. Clinical pharmacology and therapeutics 1973, (4), 580–5. [Google Scholar] [CrossRef]
- Soyinka, J. O.; Onyeji, C. O.; Omoruyi, S. I.; Owolabi, A. R.; Sarma, P. V.; Cook, J. M. , Effects of concurrent administration of nevirapine on the disposition of quinine in healthy volunteers. The Journal of pharmacy and pharmacology 2009, (4), 439–43. [Google Scholar] [CrossRef] [PubMed]
- Shyr, Z. A.; Cheng, Y.-S.; Lo, D. C.; Zheng, W. , Drug combination therapy for emerging viral diseases. Drug Discovery Today 2021, (10), 2367–2376. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fan, G.; Salam, A.; Horby, P.; Hayden, F. G.; Chen, C.; Pan, J.; Zheng, J.; Lu, B.; Guo, L.; Wang, C.; Cao, B.; Network, C.-A. P. C. , Comparative Effectiveness of Combined Favipiravir and Oseltamivir Therapy Versus Oseltamivir Monotherapy in Critically Ill Patients With Influenza Virus Infection. The Journal of Infectious Diseases 2019, (10), 1688–1698. [Google Scholar] [CrossRef] [PubMed]
- Boulon, R.; Mazeaud, C.; Farahani, M. D.; Broquière, M.; Iddir, M.; Charpentier, T.; Anton, A.; Ayotte, Y.; Woo, S.; Lamarre, A.; Chatel-Chaix, L.; LaPlante, S. R., Repurposing Drugs and Synergistic Combinations as Potential Therapies for Inhibiting SARS-CoV-2 and Coronavirus Replication. ACS Pharmacology & Translational Science 2024.
- Cho, W.-K.; Lee, M.-M.; Ma, J. Y. , Antiviral Effect of Isoquercitrin against Influenza A Viral Infection via Modulating Hemagglutinin and Neuraminidase. International journal of molecular sciences 2022, (21), 13112. [Google Scholar] [CrossRef]
- Srinivasan, V.; Brognaro, H.; Prabhu, P. R.; de Souza, E. E.; Günther, S.; Reinke, P. Y. A.; Lane, T. J.; Ginn, H.; Han, H.; Ewert, W.; Sprenger, J.; Koua, F. H. M.; Falke, S.; Werner, N.; Andaleeb, H.; Ullah, N.; Franca, B. A.; Wang, M.; Barra, A. L. C.; Perbandt, M.; Schwinzer, M.; Schmidt, C.; Brings, L.; Lorenzen, K.; Schubert, R.; Machado, R. R. G.; Candido, E. D.; Oliveira, D. B. L.; Durigon, E. L.; Niebling, S.; Garcia, A. S.; Yefanov, O.; Lieske, J.; Gelisio, L.; Domaracky, M.; Middendorf, P.; Groessler, M.; Trost, F.; Galchenkova, M.; Mashhour, A. R.; Saouane, S.; Hakanpää, J.; Wolf, M.; Alai, M. G.; Turk, D.; Pearson, A. R.; Chapman, H. N.; Hinrichs, W.; Wrenger, C.; Meents, A.; Betzel, C. , Antiviral activity of natural phenolic compounds in complex at an allosteric site of SARS-CoV-2 papain-like protease. Communications Biology 2022, (1), 805. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N. P.; O’Donnell, C.; Tiwari, B. K., Effect of thermal processing on anthocyanin stability in foods; mechanisms and kinetics of degradation. Trends in Food Science & Technology 2010, 21, (1), 3-11.
Figure 1.
Liquid European black elderberry (EC 3.2) extract (A) and quinine (B) inhibit the replication of Influenza A virus (IAV) in MDCKII cells. MDCKII cells were infected with the IAV isolate A/Puerto Rico/8/34 (PR8) at a MOI of 0.01. 30 minutes after infection and removal of input virus, cells were treated with indicated dilution steps of EC 3.2 (A) or concentrations of quinine-sulfate (B). DMSO as a solvent control was tested at the same concentration as used in the highest concentration of quinine (10 µM). Cell culture supernatants were harvested at 2 days post infection (dpi). The virions were purified and analyzed by qRT-PCR. Bars show mean values of three independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (*p <0.00312; **** p <0.0001 and ns = not significant versus the untreated control).
Figure 1.
Liquid European black elderberry (EC 3.2) extract (A) and quinine (B) inhibit the replication of Influenza A virus (IAV) in MDCKII cells. MDCKII cells were infected with the IAV isolate A/Puerto Rico/8/34 (PR8) at a MOI of 0.01. 30 minutes after infection and removal of input virus, cells were treated with indicated dilution steps of EC 3.2 (A) or concentrations of quinine-sulfate (B). DMSO as a solvent control was tested at the same concentration as used in the highest concentration of quinine (10 µM). Cell culture supernatants were harvested at 2 days post infection (dpi). The virions were purified and analyzed by qRT-PCR. Bars show mean values of three independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (*p <0.00312; **** p <0.0001 and ns = not significant versus the untreated control).
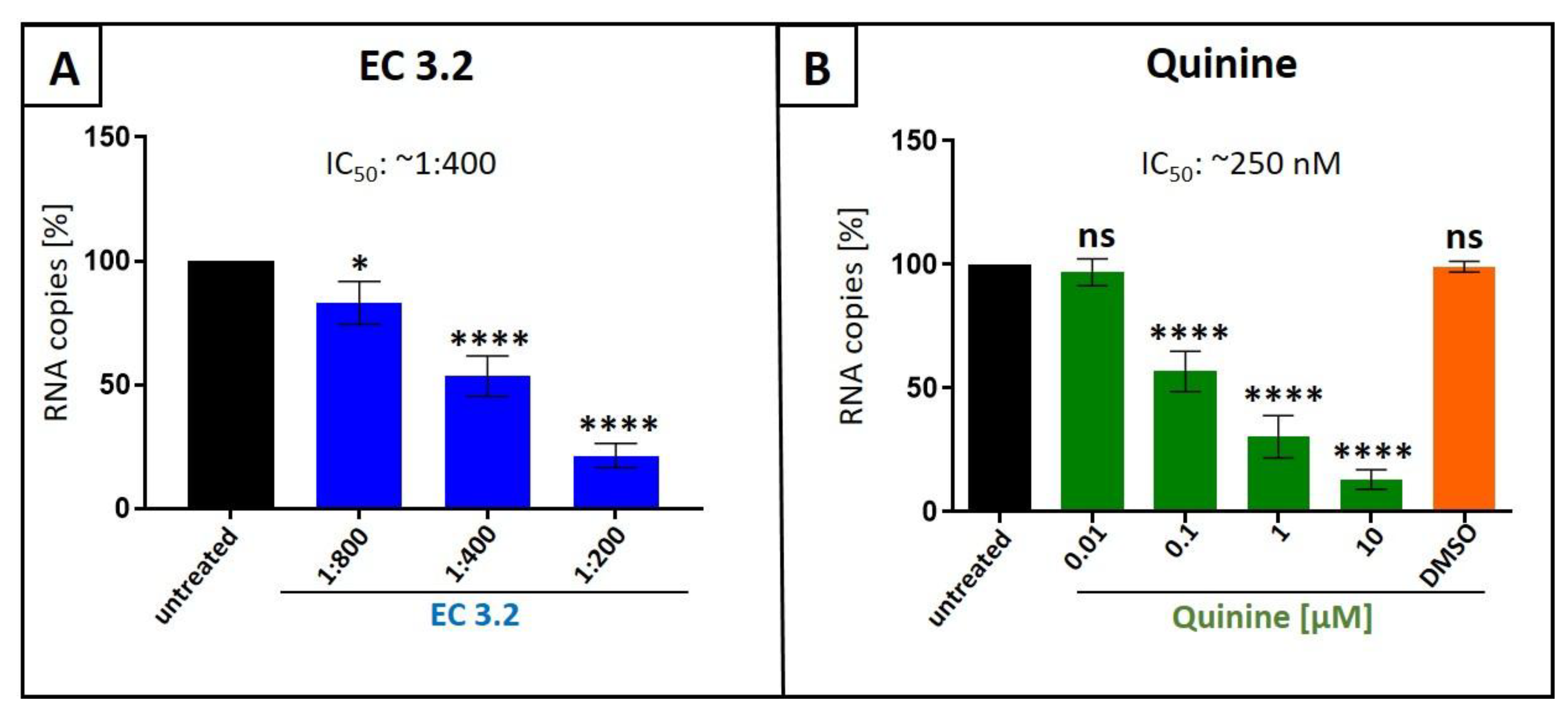
Figure 2.
Influence of EC 3.2 (A) or quinine (B) on the cell viability of MDCKII cells. Following treatment with different dilutions of EC 3.2 or concentrations of quinine-sulfate (indicated at the x-axis) for two days, the influence on cell viability was measured by water-soluble tetrazolium salt (WST)-1 assay. Bars represent means of three independent experiments ±SD. Staurosporine (StS, 1 µM) was used as a positive control.
Figure 2.
Influence of EC 3.2 (A) or quinine (B) on the cell viability of MDCKII cells. Following treatment with different dilutions of EC 3.2 or concentrations of quinine-sulfate (indicated at the x-axis) for two days, the influence on cell viability was measured by water-soluble tetrazolium salt (WST)-1 assay. Bars represent means of three independent experiments ±SD. Staurosporine (StS, 1 µM) was used as a positive control.
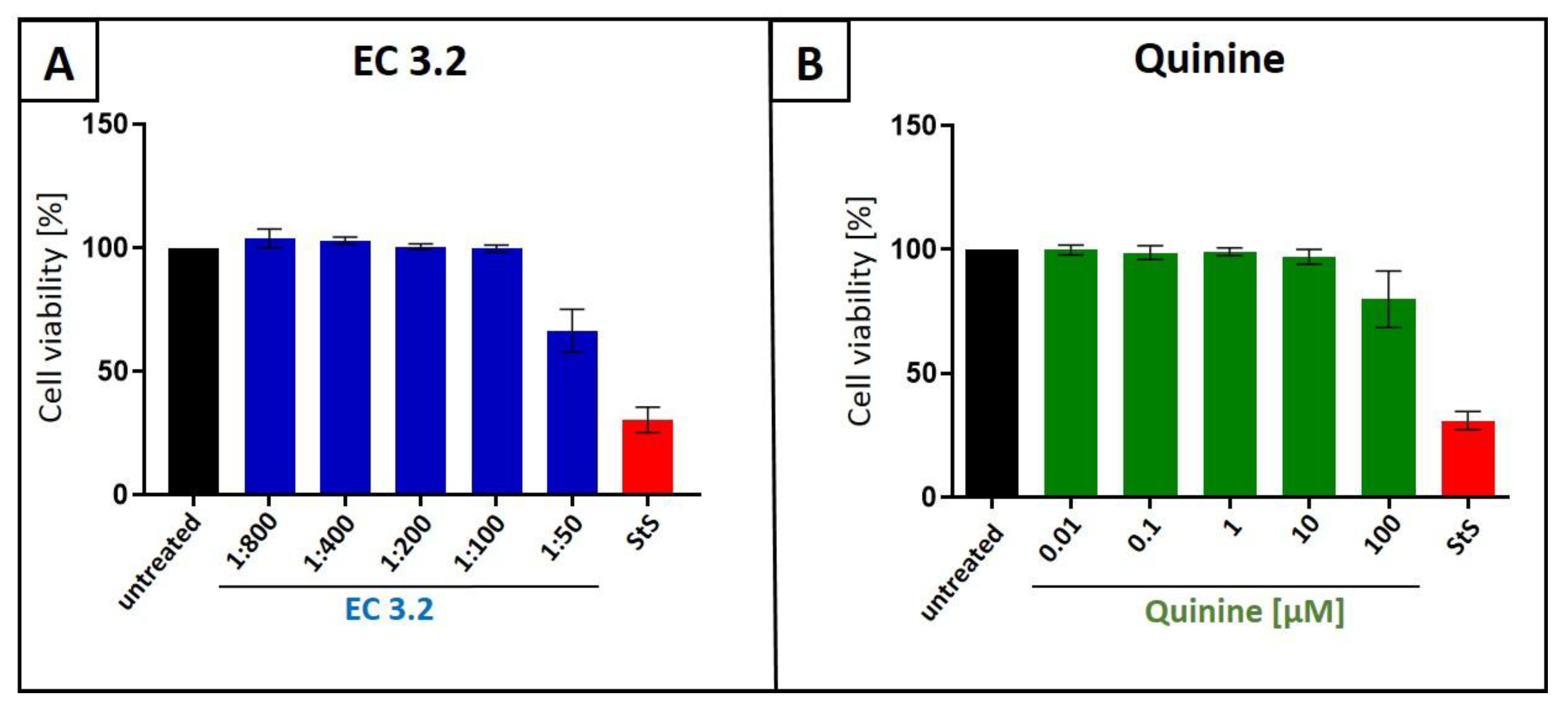
Figure 3.
Antiviral activity of the combinatory treatment of quinine with EC 3.2 following infection with IAVPR-8. A: MDCKII cells were infected with IAVPR-8 at a MOI of 0.01. 30 minutes after infection and removal of input virus, cells were treated with indicated concentrations of quinine (green), dilution steps of EC 3.2 (blue) or the combinatory treatment of quinine and EC 3.2 (orange). Cell culture supernatants were harvested at 2 dpi. The virions were purified and analyzed by qRT-PCR. Bars show mean values of three independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (**p <0.01; **** p <0.0001 and ns = not significant versus the untreated control). B: percent of the inhibition of viral replication following combined treatment with quinine and EC 3.2 and infection with IAVPR-8. The tables were created using the open-source and free web application SynergyFinder [31].
Figure 3.
Antiviral activity of the combinatory treatment of quinine with EC 3.2 following infection with IAVPR-8. A: MDCKII cells were infected with IAVPR-8 at a MOI of 0.01. 30 minutes after infection and removal of input virus, cells were treated with indicated concentrations of quinine (green), dilution steps of EC 3.2 (blue) or the combinatory treatment of quinine and EC 3.2 (orange). Cell culture supernatants were harvested at 2 dpi. The virions were purified and analyzed by qRT-PCR. Bars show mean values of three independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (**p <0.01; **** p <0.0001 and ns = not significant versus the untreated control). B: percent of the inhibition of viral replication following combined treatment with quinine and EC 3.2 and infection with IAVPR-8. The tables were created using the open-source and free web application SynergyFinder [31].
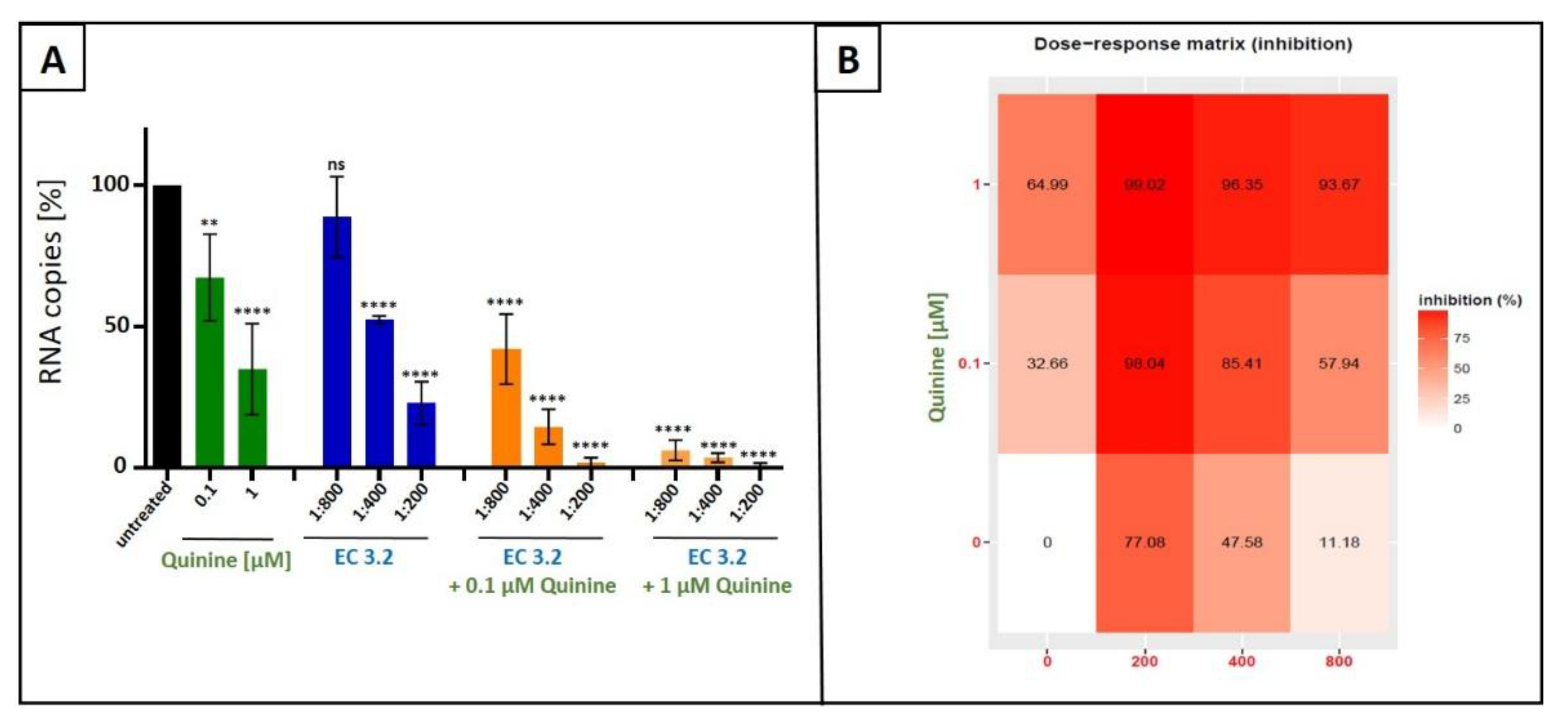
Figure 4.
Interaction profile of quinine and EC 3.2 for determining the synergy in the inhibition of the replication capacity of IAVPR-8. Drug interactions were analyzed using the reference model Bliss independence. The illustrations were created using the open-source and free web application SynergyFinder. The synergy calculations were performed on data derived from the experiments in MDCKII cells. The data represent means of three independent experiments. A color-coded interaction graphic was used to illustrate the Bliss synergy scores. High synergy scores are colored in red.
Figure 4.
Interaction profile of quinine and EC 3.2 for determining the synergy in the inhibition of the replication capacity of IAVPR-8. Drug interactions were analyzed using the reference model Bliss independence. The illustrations were created using the open-source and free web application SynergyFinder. The synergy calculations were performed on data derived from the experiments in MDCKII cells. The data represent means of three independent experiments. A color-coded interaction graphic was used to illustrate the Bliss synergy scores. High synergy scores are colored in red.
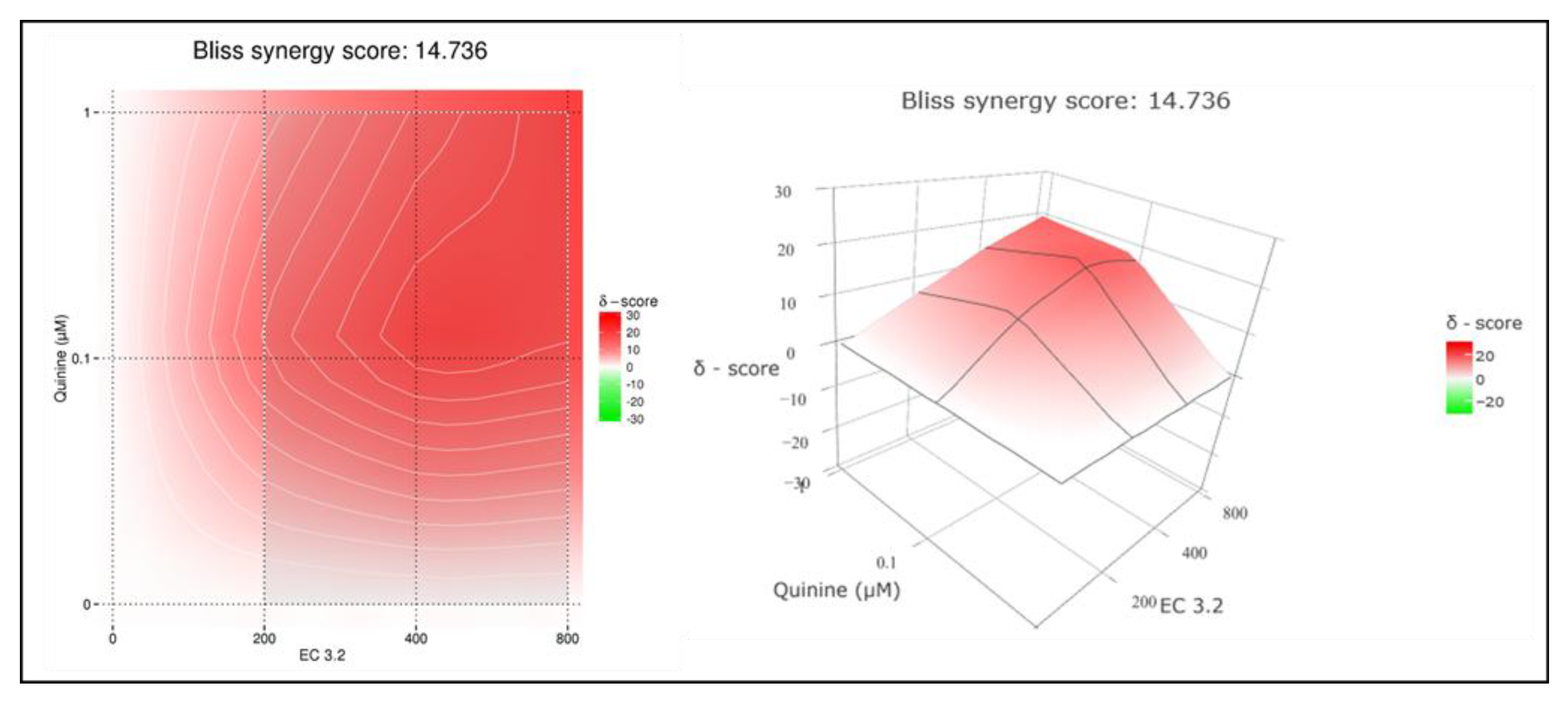
Figure 5.
Influence of the combinatory treatment of quinine with EC 3.2 on the cell viability of MDCKII cells. Following treatment with different dilutions of EC 3.2 or concentrations of quinine-sulfate (indicated at the x-axis) for two days, the influence on cell viability was measured by water-soluble tetrazolium salt (WST)-1 assay. Bars represent means of three independent experiments ±SD. Staurosporine (StS, 1 µM) was used as a positive control.
Figure 5.
Influence of the combinatory treatment of quinine with EC 3.2 on the cell viability of MDCKII cells. Following treatment with different dilutions of EC 3.2 or concentrations of quinine-sulfate (indicated at the x-axis) for two days, the influence on cell viability was measured by water-soluble tetrazolium salt (WST)-1 assay. Bars represent means of three independent experiments ±SD. Staurosporine (StS, 1 µM) was used as a positive control.
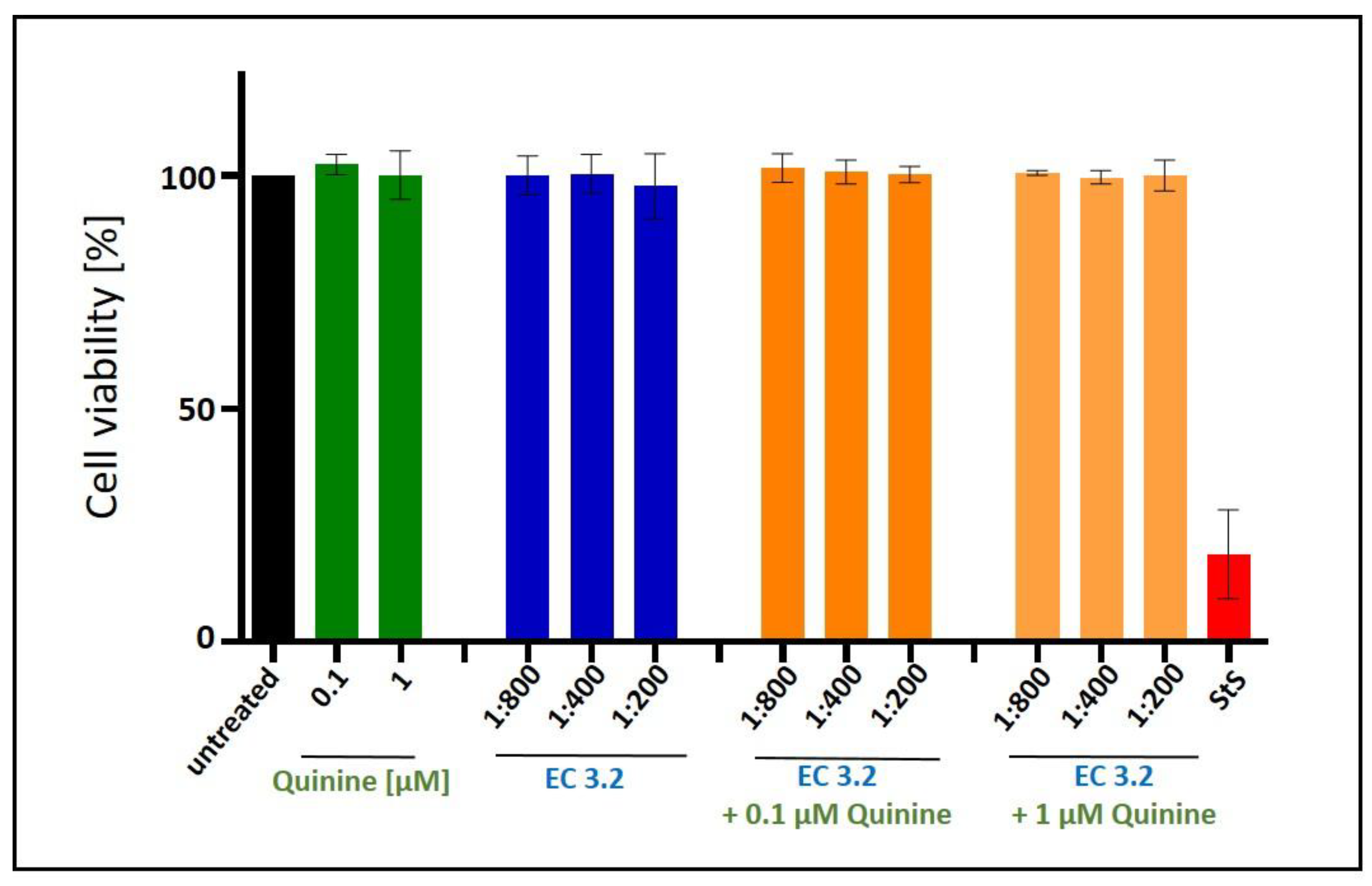
Figure 6.
Antiviral activity of the combinatory treatment of quinine with EC 3.2 following infection with SARS-CoV-2 Wuhan type. A: Calu-3 cells were infected with the clinical isolate SARS-CoV-2PR-1 at a MOI of 2x10-2. 1 hour after infection and removal of input virus, cells were treated with indicated concentrations of quinine (green), dilution steps of EC 3.2 (blue) or the combinatory treatment of quinine and EC 3.2 (orange). Cell culture supernatants were harvested at 3 dpi. The virions were purified and analyzed by qRT-PCR. Bars show mean values of three independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (**p <0.057 **** p <0.0001 and ns = not significant versus the untreated control). B: percent of the inhibition of viral replication following combined treatment with quinine and EC 3.2 and infection with IAVPR-8. The tables were created using the open-source and free web application SynergyFinder [31].
Figure 6.
Antiviral activity of the combinatory treatment of quinine with EC 3.2 following infection with SARS-CoV-2 Wuhan type. A: Calu-3 cells were infected with the clinical isolate SARS-CoV-2PR-1 at a MOI of 2x10-2. 1 hour after infection and removal of input virus, cells were treated with indicated concentrations of quinine (green), dilution steps of EC 3.2 (blue) or the combinatory treatment of quinine and EC 3.2 (orange). Cell culture supernatants were harvested at 3 dpi. The virions were purified and analyzed by qRT-PCR. Bars show mean values of three independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (**p <0.057 **** p <0.0001 and ns = not significant versus the untreated control). B: percent of the inhibition of viral replication following combined treatment with quinine and EC 3.2 and infection with IAVPR-8. The tables were created using the open-source and free web application SynergyFinder [31].
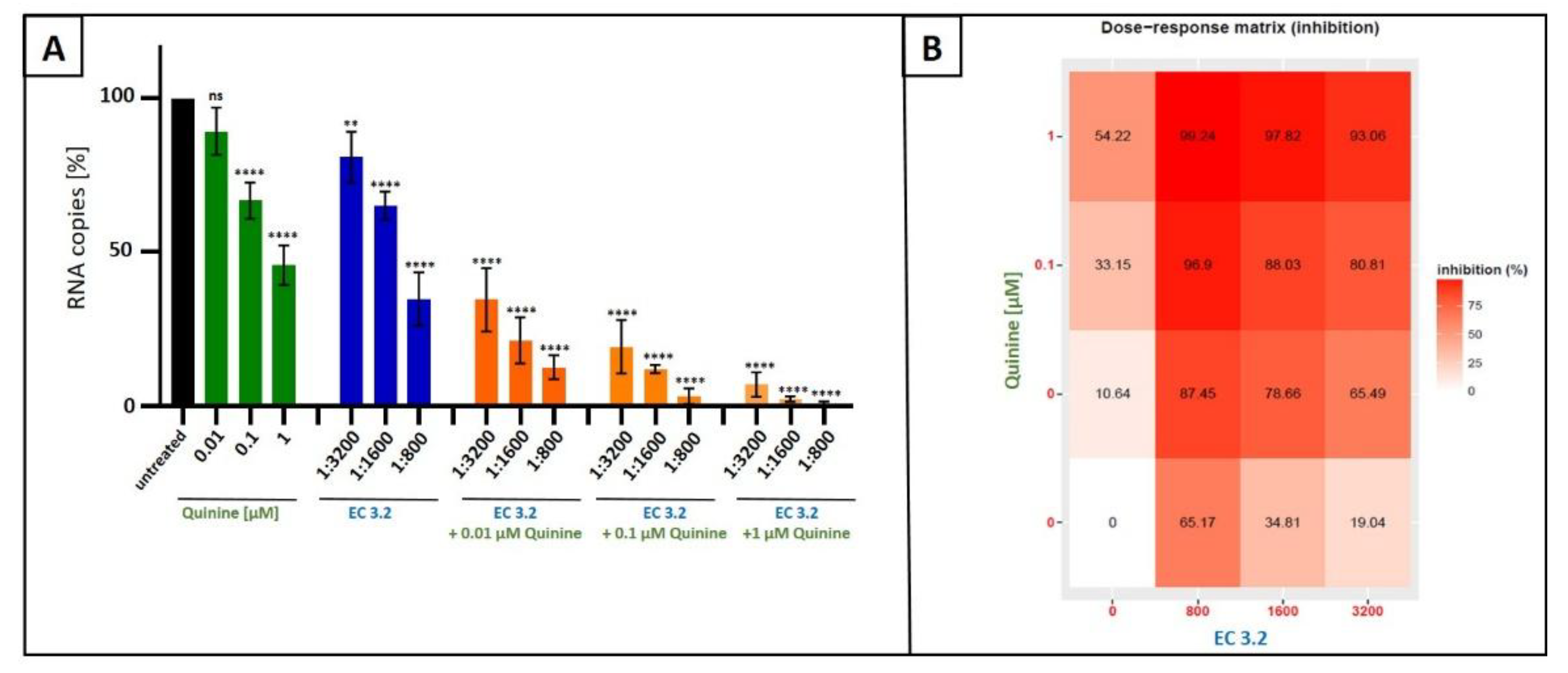
Figure 7.
Interaction profile of quinine and EC 3.2 for determining the synergy in the inhibition of the replication capacity of SARS-CoV-2 Wuhan type. Drug interactions were analyzed using the reference model Bliss independence. The illustrations were created using the open-source and free web application SynergyFinder. The synergy calculations were performed on data derived from the experiments in Calu-3 cells. The data represent means of three independent experiments. A color-coded interaction graphic was used to illustrate the Bliss synergy scores. High synergy scores are colored in red.
Figure 7.
Interaction profile of quinine and EC 3.2 for determining the synergy in the inhibition of the replication capacity of SARS-CoV-2 Wuhan type. Drug interactions were analyzed using the reference model Bliss independence. The illustrations were created using the open-source and free web application SynergyFinder. The synergy calculations were performed on data derived from the experiments in Calu-3 cells. The data represent means of three independent experiments. A color-coded interaction graphic was used to illustrate the Bliss synergy scores. High synergy scores are colored in red.
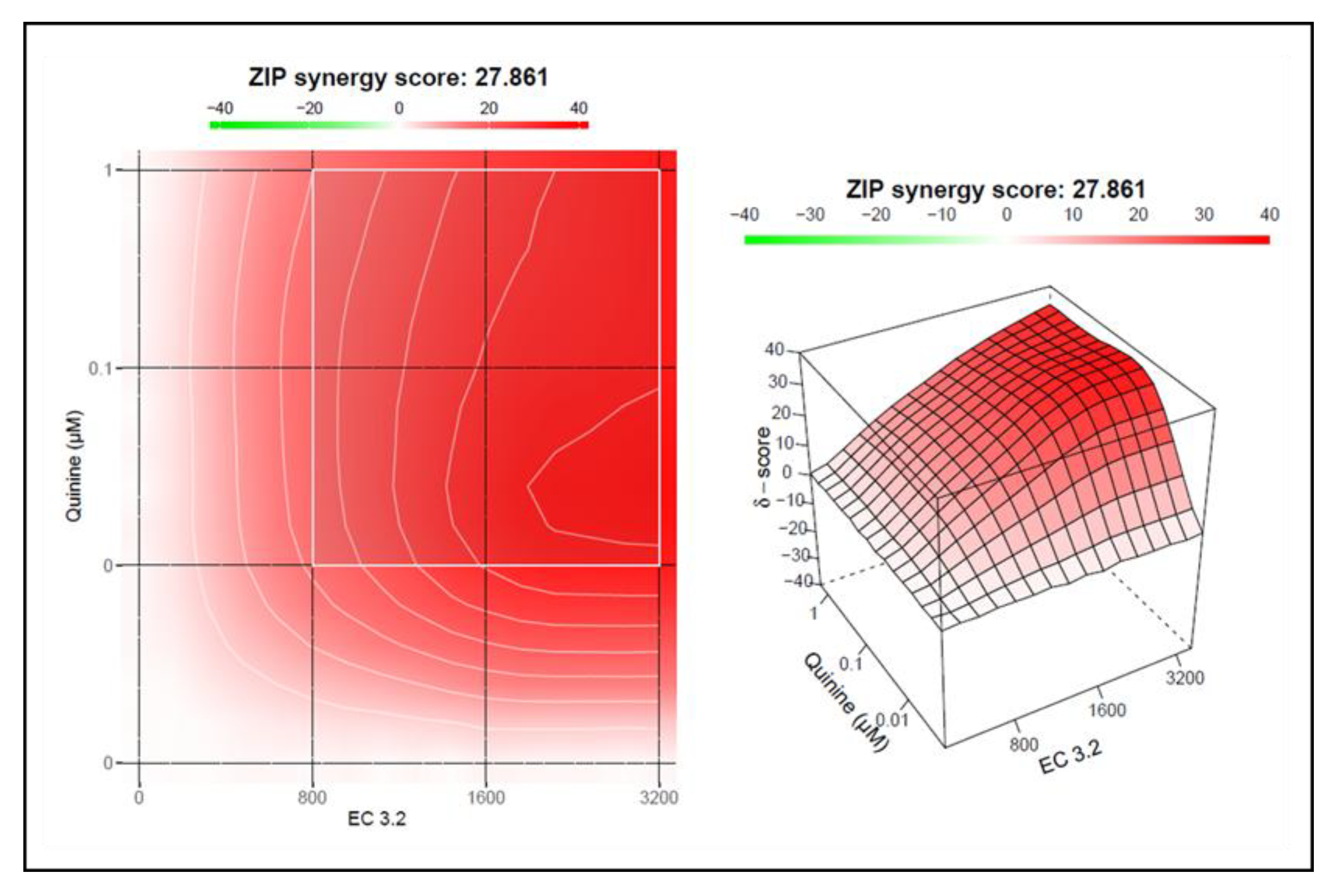
Figure 8.
Influence of the combinatory treatment of quinine with EC 3.2 on the cell viability of Calu-3 cells. Following treatment with different dilutions of EC 3.2 or concentrations of quinine-sulfate (indicated at the x-axis) for three days, the influence on cell viability was measured by water-soluble tetrazolium salt (WST)-1 assay. Bars represent means of three independent experiments ±SD. Staurosporine (StS, 1 µM) was used as a positive control.
Figure 8.
Influence of the combinatory treatment of quinine with EC 3.2 on the cell viability of Calu-3 cells. Following treatment with different dilutions of EC 3.2 or concentrations of quinine-sulfate (indicated at the x-axis) for three days, the influence on cell viability was measured by water-soluble tetrazolium salt (WST)-1 assay. Bars represent means of three independent experiments ±SD. Staurosporine (StS, 1 µM) was used as a positive control.
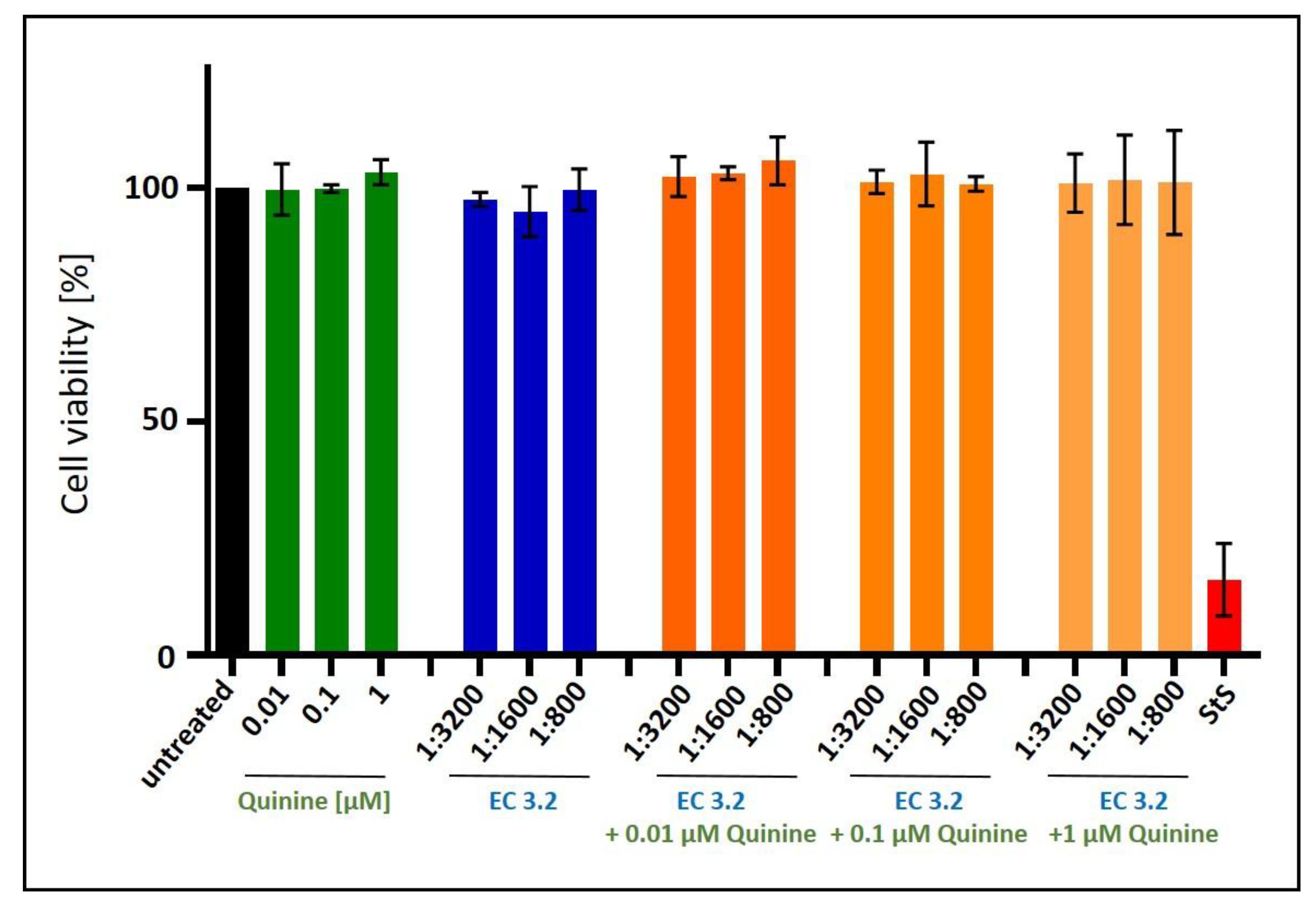
Figure 9.
Antiviral activity of ElderCraft® Q-Complex against IAV and SARS-CoV-2 compared to EC 15. A+B: MDCKII cells were infected with the IAV isolate A/Puerto Rico/8/34 (PR8) at a MOI of 0.01. 30 minutes after infection and removal of input virus, cells were treated with indicated dilution steps of EC 15 (A) or ElderCraft® Q-Complex (B). Cell culture supernatants were harvested at 2 days post infection (dpi). The virions were purified and analyzed by qRT-PCR. Bars show mean values of three independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (***p <0.0002; **** p <0.0001 and ns = not significant versus the untreated control). C+D: Calu-3 cells were infected with the clinical isolate SARS-CoV-2PR-1 at a MOI of 2x10-2. 1 hour after infection and removal of input virus, cells were treated with indicated concentrations EC 15 (C) or ElderCraft® Q-Complex (D). Cell culture supernatants were harvested at 3 dpi. The virions were purified and analyzed by qRT-PCR. Bars show mean values of four independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (*p <0.0192 and **** p <0.0001 versus the untreated control).
Figure 9.
Antiviral activity of ElderCraft® Q-Complex against IAV and SARS-CoV-2 compared to EC 15. A+B: MDCKII cells were infected with the IAV isolate A/Puerto Rico/8/34 (PR8) at a MOI of 0.01. 30 minutes after infection and removal of input virus, cells were treated with indicated dilution steps of EC 15 (A) or ElderCraft® Q-Complex (B). Cell culture supernatants were harvested at 2 days post infection (dpi). The virions were purified and analyzed by qRT-PCR. Bars show mean values of three independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (***p <0.0002; **** p <0.0001 and ns = not significant versus the untreated control). C+D: Calu-3 cells were infected with the clinical isolate SARS-CoV-2PR-1 at a MOI of 2x10-2. 1 hour after infection and removal of input virus, cells were treated with indicated concentrations EC 15 (C) or ElderCraft® Q-Complex (D). Cell culture supernatants were harvested at 3 dpi. The virions were purified and analyzed by qRT-PCR. Bars show mean values of four independent experiments ± standard deviation. Statistical analysis was performed using a multiple comparison Kruskal-Wallis test (Anova) followed by Dunn’s post hoc test (*p <0.0192 and **** p <0.0001 versus the untreated control).
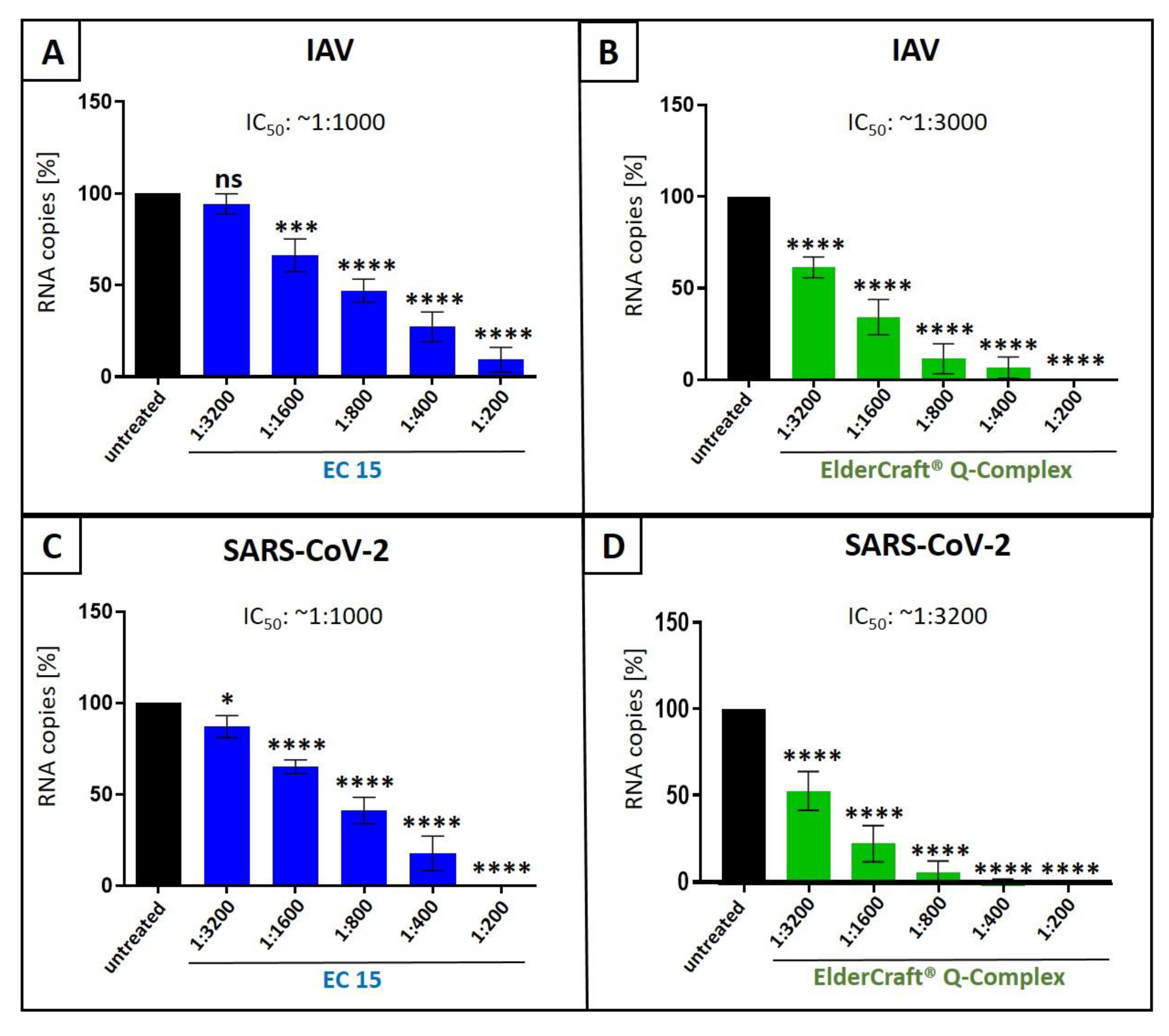
Figure 10.
Influence of EC 15 (A+C) or ElderCraft® Q-Complex (B+D) on the cell viability of MDCKII (A+B) and Calu-3 (C+D) cells. Following treatment with different dilutions of EC 15 (A+C) or ElderCraft® Q-Complex (B+D) (indicated at the x-axis) for two (A+B) or three (C+D) days, the influence on cell viability was measured by water-soluble tetrazolium salt (WST)-1 assay. Bars represent means of three independent experiments ±SD. Staurosporine (StS, 1 µM) was used as a positive control.
Figure 10.
Influence of EC 15 (A+C) or ElderCraft® Q-Complex (B+D) on the cell viability of MDCKII (A+B) and Calu-3 (C+D) cells. Following treatment with different dilutions of EC 15 (A+C) or ElderCraft® Q-Complex (B+D) (indicated at the x-axis) for two (A+B) or three (C+D) days, the influence on cell viability was measured by water-soluble tetrazolium salt (WST)-1 assay. Bars represent means of three independent experiments ±SD. Staurosporine (StS, 1 µM) was used as a positive control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



