Submitted:
07 March 2025
Posted:
10 March 2025
You are already at the latest version
Abstract
Heart failure (HF) is a global health challenge characterized by the heart’s inability to satisfy metabolic demands, driven by renin-angiotensin-aldosterone system (RAAS) overactivation, neurohormonal imbalance, and emerging mechanisms like the gut-heart axis and mitochondrial dysfunction. Affecting over 6 million adults in the US alone, HF incurs a 5-year mortality rate of 50% and escalating costs projected to double by 2030. This review examines HF’s molecular paradigms, integrating established pathways with advances in omics, stem cell therapy, genetic modification, and personalized medicine. RAAS blockade remains central, yet its efficacy is limited in HF with preserved ejection fraction (HFpEF). Stem cell therapies (mesenchymal and induced pluripotent stem cells) show regenerative potential but face poor retention (10% survival at 30 days). CRISPR/Cas9 offers precision, though off-target effects persist. The gut microbiome, via trimethylamine N-oxide, exacerbates inflammation, while omics technologies promise biomarkers for tailored treatments. Challenges include translating these innovations into practice, particularly for HFpEF. Future directions involve novel HFpEF therapies, enhanced stem cell delivery, precise genetic tools, and microbiome interventions, supported by artificial intelligence. By 2030, these advances could shift HF management toward regeneration, contingent on overcoming translational barriers through global collaboration.
Keywords:
Heart Failure
; Renin-Angiotensin System
; Microbiota
; Genomics
; Stem Cell Therapy
; Genetic Therapy
; Personalized Medicine
; Oxidative Stress
1. Introduction
Heart failure (HF) is a complex clinical syndrome characterized by the heart's inability to maintain adequate cardiac output to meet peripheral metabolic demands, often requiring elevated ventricular filling pressures to compensate [1]. This condition imposes a significant global health burden with profound epidemiological and economic implications. In the United States, HF affects approximately 6.2 million adults (2013-2016), with an annual incidence of 915,000 new cases and a projected 46% increase by 2030, driven by an aging population and persistent risk factors such as systemic arterial hypertension and type 2 diabetes mellitus [2]. In Europe, prevalence ranges from 1-2% in adults, rising to over 10% among those aged 70 and older, with elevated rates in Germany and the United Kingdom due to ischemic heart disease [3]. Japan reports a prevalence of 1-1.2%, while Brazil’s is 1-2%, reflecting a shift toward Western cardiovascular risk profiles [4,5]. HF’s morbidity is marked by recurrent acute decompensation episodes necessitating frequent hospitalizations, with a 5-year mortality rate approaching 50%, comparable to certain aggressive malignancies [6]. Economically, HF-associated costs in the US exceeded $30.7 billion in 2012, projected to double by 2030, while Europe faces an annual burden of approximately €29 billion, highlighting the need for optimized therapeutic strategies [7,8].
The pathophysiology of HF involves myocardial contractile dysfunction, oxidative stress, and adverse ventricular remodeling, primarily mediated by overactivation of the renin-angiotensin-aldosterone system (RAAS) and the release of proinflammatory cytokines like tumor necrosis factor-alpha (TNF-α) [9]. Emerging research has identified novel mechanisms, including epigenetic regulation of cardiomyocyte gene expression, the gut-heart axis via the microbiome, and mitochondrial dysfunction, all currently under investigation as potential therapeutic targets [10]. This review provides an overview of HF’s molecular paradigms, integrating established knowledge with cutting-edge research and exploring their applicability across diverse global contexts.
2. Classic and Contemporary Metabolic Pathways in Heart Failure
The most widely recognized pathophysiological mechanism in HF is the overactivation of the RAAS, a neurohormonal system pivotal to cardiovascular homeostasis [11]. In the short term, RAAS activation compensates for reduced cardiac output by inducing adaptive changes at cardiac, renal, and vascular levels, such as increased contractility and fluid retention. However, its chronic overactivation generates hemodynamic stress, leading to deleterious effects including myocardial hypertrophy, fibrosis, and systemic vasoconstriction, all of which exacerbate HF progression [11]. This sustained neurohormonal activation, first comprehensively integrated by Packer in 1992, remains a primary driver of HF, positioning pharmacological RAAS blockade—via ACE inhibitors (ACEIs), angiotensin II receptor blockers (ARBs), and mineralocorticoid receptor antagonists—as a cornerstone of contemporary treatment [12].
Reduced cardiac output triggers a cascade of compensatory responses detected by peripheral arterial baroreceptors, signaling an underfilled state [11]. These responses encompass increased heart rate and myocardial contractility (via sympathetic nervous system [SNS] activation), sodium and water retention (via RAAS), and peripheral vasoconstriction to maintain blood pressure [11]. In heart failure with reduced ejection fraction (HFrEF), these mechanisms initially preserve cardiovascular homeostasis but become maladaptive over time [12]. Under normal conditions, counter-regulatory systems, such as parasympathetic tone and natriuretic peptides, mitigate SNS and RAAS activity. However, in HF, parasympathetic tone diminishes, and resistance to natriuretic peptides increases, amplifying neurohormonal activation [12]. Originally termed "neurohormones" due to their neuroendocrine origins, these molecules are now recognized to also function via paracrine and autocrine pathways, broadening their role in HF pathophysiology [12].
2.1. Neurohormonal Activation
Circulating volume is tightly regulated by baroreceptors in the aortic arch and carotid sinus (sensing high pressure) and pulmonary mechanoreceptors (detecting low pressure), which relay inhibitory signals to the central nervous system (CNS) to modulate sympathetic outflow [14]. In HF, reduced cardiac output diminishes baroreceptor activity, leading to heightened SNS activation, increased heart rate, and enhanced contractility, alongside peripheral vasoconstriction to redistribute blood flow [14]. Neurogenic signaling further disrupts the sympathetic-parasympathetic balance, with HF patients exhibiting increased chemosensitivity to hypoxia and hypercapnia, as well as exaggerated ergoreceptor reflexes triggered by metabolic byproducts of muscle work [14,15]. These alterations impair functional capacity, reduce exercise tolerance (evidenced by decreased peak oxygen consumption; VO₂), and correlate with disease severity and poorer survival prognosis [14]. Elevated circulating neurohormone levels, sufficient to induce ventricular dysfunction and remodeling, underscore the therapeutic efficacy of SNS and RAAS inhibitors, which significantly improve clinical outcomes [12].
SNS overactivation, a hallmark of early HF, elevates circulating norepinephrine levels, amplifying adrenergic signaling through sustained release and reduced reuptake at nerve terminals [19]. This chronic stimulation promotes cardiomyocyte apoptosis, myocardial fibrosis, and endothelial dysfunction, exerting long-term deleterious effects on the heart, kidneys, and peripheral vasculature [20]. Concurrently, RAAS activation amplifies these effects by increasing angiotensin II and aldosterone, further driving vasoconstriction and fluid overload [11]. Over time, these changes contribute to adverse cardiac remodeling, including left ventricular hypertrophy and dilatation, worsening HF progression [19].
2.2. Renal Function
Renal sodium and water retention, resulting in peripheral and pulmonary edema, is a central feature of HF, orchestrated by SNS and RAAS activation rather than intrinsic renal pathology [21]. Sympathetic stimulation constricts the renal afferent arteriole, reducing glomerular blood flow and triggering renin release from the juxtaglomerular apparatus, an effect enhanced by β-adrenergic receptor activation [21]. Renin catalyzes the conversion of hepatic angiotensinogen to angiotensin I, which ACE transforms into angiotensin II, a potent vasoconstrictor that binds to type 1 receptors (AT1) in the adrenal glomerulosa, stimulating aldosterone production [21]. Angiotensin II also promotes proximal tubule sodium reabsorption and triggers vasopressin release from the hypothalamus, increasing water retention via non-osmotic pathways that override plasma osmolarity regulation, leading to hyponatremia [22]. Aldosterone, acting on distal nephrons, further enhances sodium retention, with levels rising 30-40% in HF patients despite ACEI or ARB use, a phenomenon termed “aldosterone escape” [22].
Counter-regulatory mechanisms involve natriuretic peptides, secreted in response to atrial and myocardial stretch, which promote cyclic guanosine monophosphate (cGMP)-mediated vasodilation and natriuresis [22]. However, peripheral resistance to these peptides in HF limits their efficacy, compounded by neprilysin-mediated degradation [22]. Additional RAAS derivatives, such as angiotensin III (stimulating aldosterone) and angiotensin 1-7 (counteracting ventricular remodeling), modulate this axis, though their therapeutic potential remains underexplored [22]. These renal and systemic effects highlight the intricate interplay of neurohormonal pathways in HF, informing targeted interventions like neprilysin inhibitors (for example, sacubitril) combined with RAAS blockers.
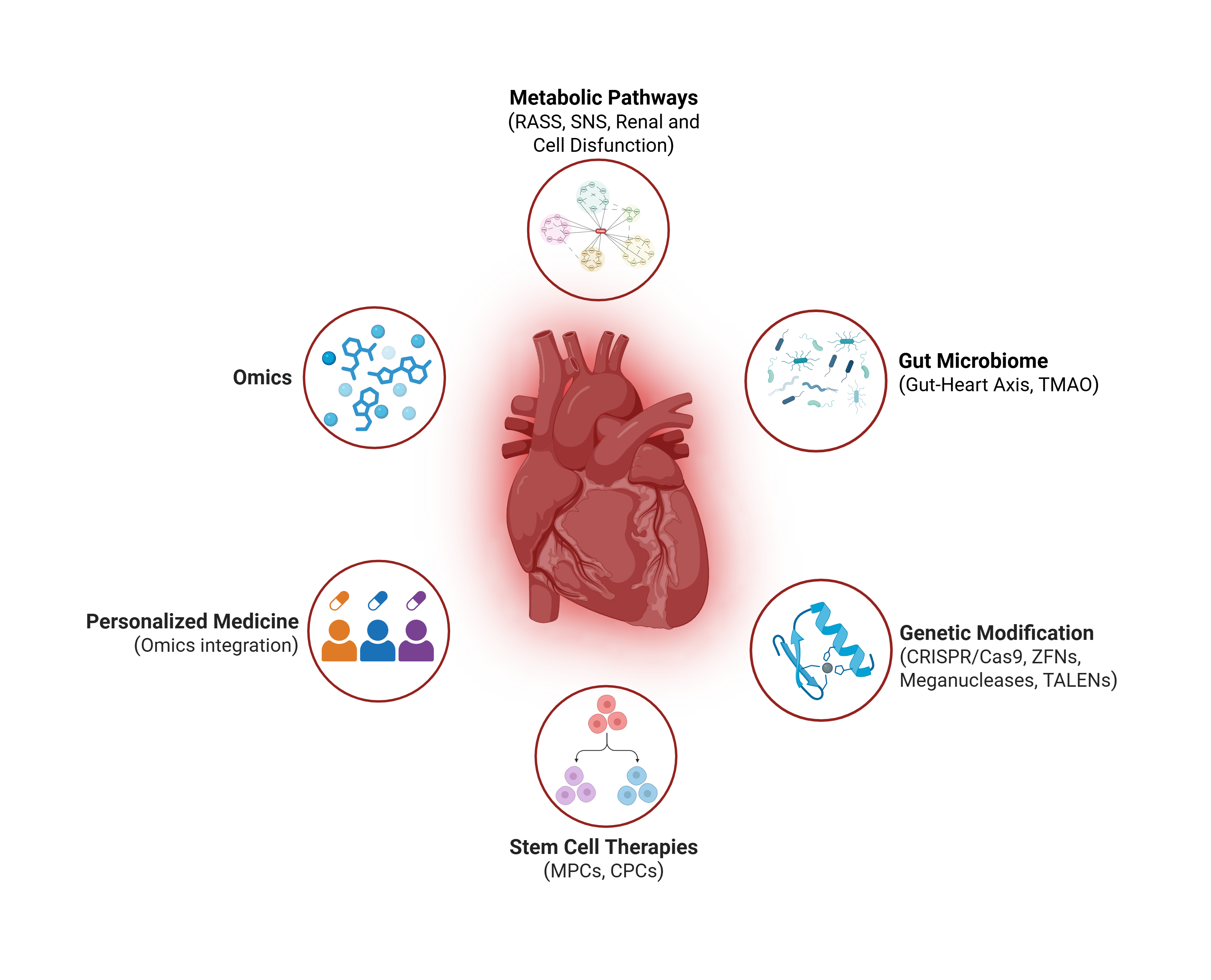
Figure 1.
Pathways in Heart Failure (Simplified View). Note that, for explanatory purposes, the pathways have been separated; however, there is complex crosstalking between all of them.
Figure 1.
Pathways in Heart Failure (Simplified View). Note that, for explanatory purposes, the pathways have been separated; however, there is complex crosstalking between all of them.

3. Microbiome in Heart Failure: The Gut-Heart Axis
The gut microbiome comprises a diverse array of microorganisms, including bacteria, viruses, archaea, bacteriophages, protozoa, and fungi, that play critical roles in metabolism, facilitating digestion, nutrient absorption, and immune system development [27,28]. Its composition exhibits significant interindividual variability, even among genetically related individuals, and is heavily shaped by environmental factors such as diet, medications, and lifestyle [29]. To characterize this diversity, microbiome research employs three key metrics: alpha diversity, assessing taxonomic richness (number of taxa) and evenness (relative abundance) within a sample; beta diversity, evaluating variability in microbial community composition across samples from the same habitat; and gamma diversity, representing total taxonomic richness across all samples from a habitat [30]. Disruptions in microbiome composition, both qualitatively and quantitatively, are implicated in the onset and progression of prevalent diseases, including obesity, metabolic syndrome, chronic kidney disease, hepatic steatosis, and cardiovascular disorders such as acute myocarditis, coronary artery disease, atrial fibrillation, and HF [31,32,33].
3.1. Gut Microbiome and Heart-Gut Axis
In HF, cardiac dysfunction compromises intestinal barrier function, leading to ischemia and edema of the gut wall [34]. This increased permeability permits translocation of microorganisms and their metabolic byproducts (e.g., lipopolysaccharides) into the portal and systemic circulation, eliciting immune and inflammatory responses [34]. The "gut hypothesis" of HF suggests that this process drives chronic low-grade inflammation, worsening cardiac dysfunction and establishing a bidirectional feedback loop between the heart and gut [29]. This dysbiosis often precedes clinical HF, influenced by factors such as Western diets rich in processed foods, sedentary lifestyles, circadian rhythm disruptions, and aging [35,36]. Notably, microbiome profiles in HF patients mirror those in dysmetabolic individuals prior to cardiovascular disease onset, suggesting a preclinical role in disease progression [35]. In advanced HF, pathogenic genera such as Shigella, Campylobacter, and Salmonella proliferate, linked to persistent T-cell activation and increased susceptibility to Clostridioides infections, particularly in hospitalized patients receiving antibiotics [29]. Concurrently, bacteria like Bacteroides/Prevotella, Eubacterium rectale, and Fusobacterium prausnitzii increase, while anti-inflammatory taxa, including Coriobacteriaceae, Erysipelotrichaceae, Ruminococcaceae, Faecalibacterium, Eubacterium, Dorea, and Bifidobacterium decline, reducing butyrate production, a key anti-inflammatory metabolite that modulates cytokine cascades and regulatory T-cell activity [29,37].
3.2. Disorders of Intestinal Metabolism in HF
HF-induced intestinal hypoxia and visceral venous congestion, particularly in right-sided HF, impair blood flow to epithelial cells, triggering cellular hypoxia, anaerobic metabolism, and overexpression of the sodium/hydrogen exchanger 3, which increases sodium transport and lowers luminal pH [38]. Given that up to 40% of total blood volume resides in the gastrointestinal tract, these metabolic shifts have significant clinical impact [39]. This functional dysbiosis disrupts nutrient digestion, vitamin synthesis, and mucosal immunity, with a notable reduction in Bacteroides and Bifidobacteria and an increase in Firmicutes and Proteobacteria [40]. Fecal samples from chronic HF patients reveal higher concentrations of enteric pathogens (e.g., Salmonella, Shigella, Campylobacter), correlating with disease severity and systemic inflammation [40]. These changes exacerbate HF by amplifying inflammatory signaling and immune dysregulation, further straining cardiac function [29].
3.3. Trimethylamine N-Oxide
Trimethylamine N-Oxide (TMAO), a metabolite produced by gut microbiota from dietary precursors like choline, betaine, L-carnitine, and phosphatidylcholine (found in seafood, dairy, eggs, meat, and organ meats), is increasingly recognized as a cardiovascular risk factor, dubbed the “missing link” between Western diets and HF [43]. Gut bacteria, particularly Firmicutes and Proteobacteria (e.g., Providencia rettgeri, Clostridioides sporogenes), hydrolyze these nutrients via enzymes like choline-TMA lyase (cutC/D) and carnitine monooxygenase (cntA/B), producing trimethylamine (TMA) [43]. TMA is absorbed and oxidized to TMAO by hepatic flavin-containing monooxygenase (FMO3), with mutations in FMO3 causing trimethylaminuria due to TMA accumulation [44,45,46,47]. In HF, elevated TMAO levels correlate with increased inflammation, oxidative stress, and adverse ventricular remodeling, amplifying disease progression [43]. Excess TMA may also degrade into dimethylamine or methane, though TMAO’s role in HF is most pronounced [46,47].
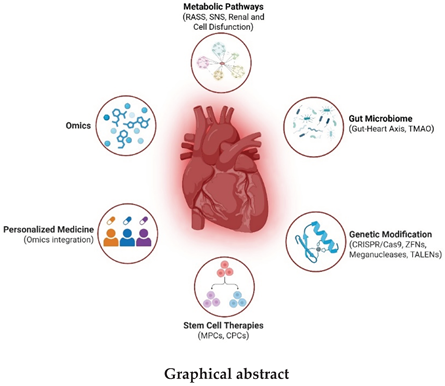
Figure 2.
Gut-Heart Axis and Its Perpetuation. Intestinal dysbiosis, further sustained by dysfunction secondary to heart failure (HF), leads to an increase in TMA, which is converted into TMAO by hepatic FMO3. TMAO, along with other external agents, enhances adverse cardiac remodeling, perpetuating the cycle. Adapted from original article by M. Karmazyn y X. T. Gan, 2023 [110].
Figure 2.
Gut-Heart Axis and Its Perpetuation. Intestinal dysbiosis, further sustained by dysfunction secondary to heart failure (HF), leads to an increase in TMA, which is converted into TMAO by hepatic FMO3. TMAO, along with other external agents, enhances adverse cardiac remodeling, perpetuating the cycle. Adapted from original article by M. Karmazyn y X. T. Gan, 2023 [110].
4. The Evolution of Omics in Heart Failure Research
Recent advances in omics technologies (genomics, transcriptomics, proteomics, and metabolomics) have revolutionized HF research by providing comprehensive insights into its molecular mechanisms across multiple biological layers [48]. These tools have shifted cardiac research from broad pathophysiological models to detailed molecular profiles, enabling precise diagnostic and therapeutic strategies that pave the way for personalized medicine [48]. This section explores how omics approaches elucidate HF’s complexity and their potential to transform clinical practice.
4.1. Genomic Foundations and Environmental Interactions
Genomic studies reveal that HF susceptibility extends beyond single nucleotide polymorphisms to include structural variants such as deletions, duplications, and inversions, particularly in inherited conditions like hypertrophic cardiomyopathy (HCM) and dilated cardiomyopathy (DCM) [49]. Rare genetic disorders underpin these cardiomyopathies, with mutations in genes like MYH7 and TTN driving disease progression [50]. Environmental factors, including diet, lifestyle, and toxin exposure, interact with genetic predispositions, adding complexity to HF development [49]. Understanding these gene-environment dynamics has improved risk stratification, enabling family screening protocols and targeted preventive strategies, though predicting individual disease trajectories remains challenging [51].
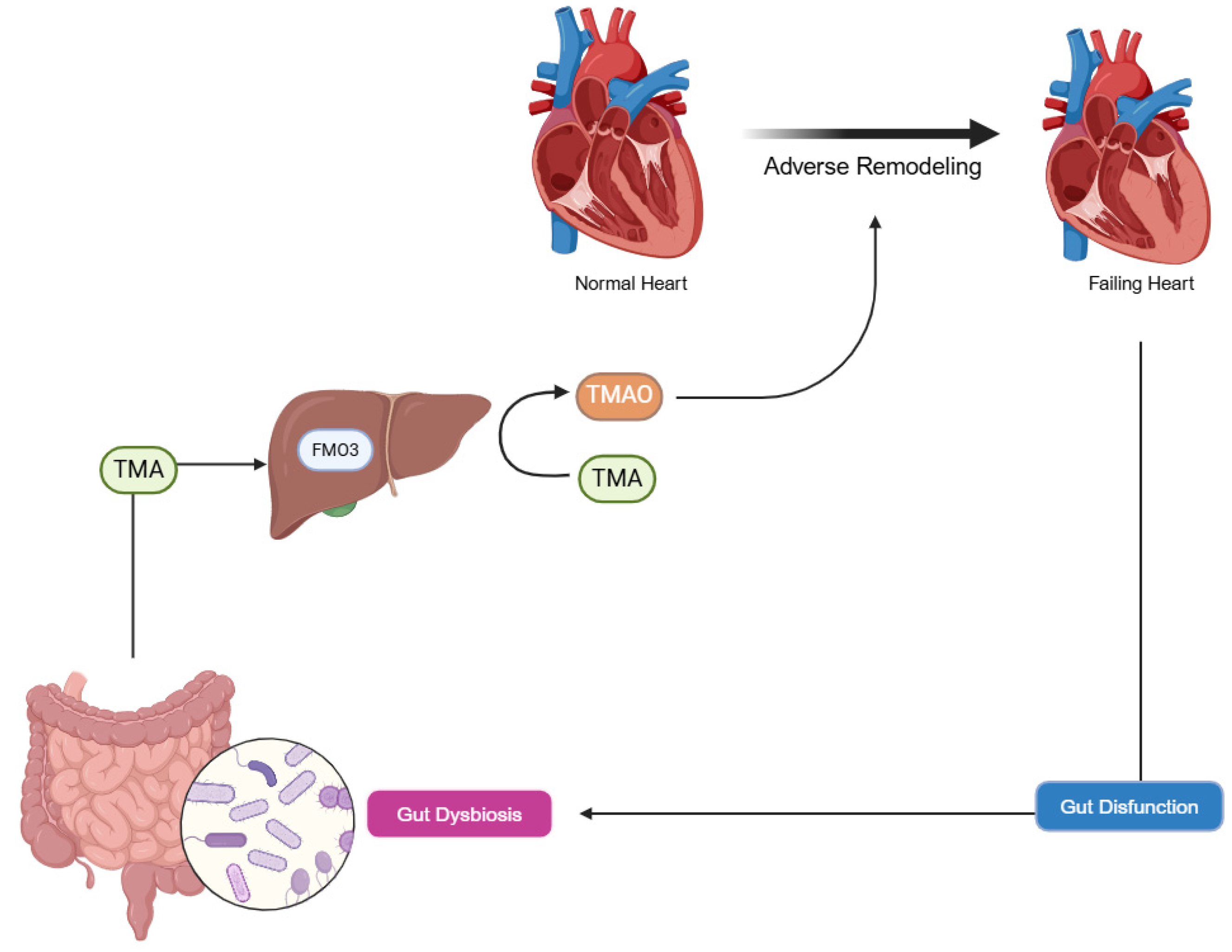
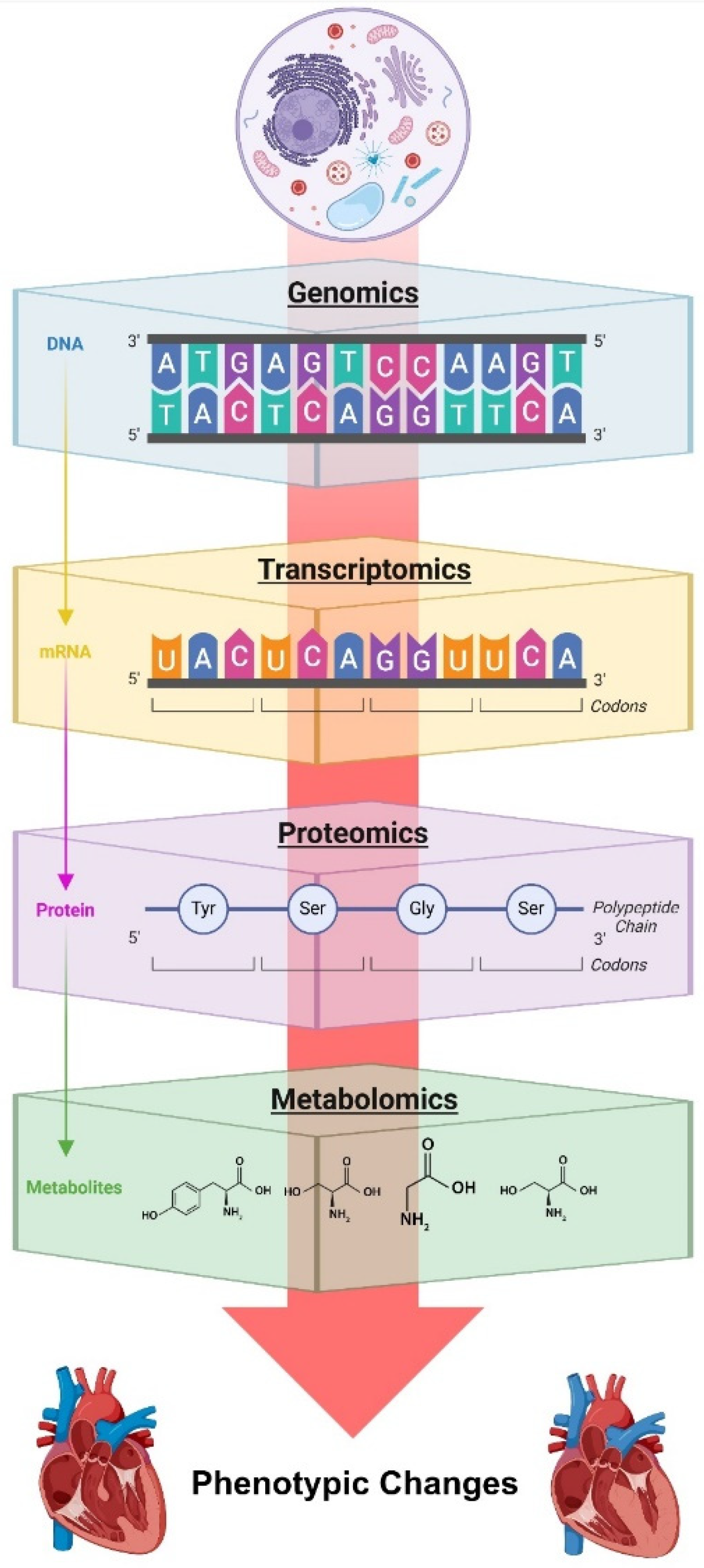
Figure 3.
Basic concepts of the relationship between the genome, transcriptome, proteome, and metabolome. Together with other elements (such as the epigenome), these factors shape phenotypic changes, such as hypertrophic cardiomyopathy and dilated cardiomyopathy.
Figure 3.
Basic concepts of the relationship between the genome, transcriptome, proteome, and metabolome. Together with other elements (such as the epigenome), these factors shape phenotypic changes, such as hypertrophic cardiomyopathy and dilated cardiomyopathy.
4.2. Advances in Transcriptomics and Regulatory RNA Networks
Transcriptomics has uncovered regulatory mechanisms involving long non-coding RNAs (lncRNAs) and circular RNAs (circRNAs), which act as scaffolds for protein complexes and modulate alternative splicing in the failing heart [52]. The advent of spatial transcriptomics allows mapping of gene expression heterogeneity at cellular and tissue levels, revealing distinct transcriptional signatures across cardiac regions [53]. For instance, studies have identified upregulated fibrotic genes in peri-infarct zones post-myocardial infarction (MI), explaining regional dysfunction in HF [54]. These RNA networks offer potential therapeutic targets, with early preclinical efforts exploring RNA-based interventions to modulate cardiac remodeling [55].
4.3. Proteomic Insights and Post-Translational Modifications
Quantitative and interaction-based proteomics have expanded understanding of HF protein networks, identifying key players in contractile dysfunction and signaling cascades [56]. Post-translational modifications, such as acetylation and methylation, critically regulate protein function, influencing cardiac remodeling, energy metabolism, and cellular signaling [57]. Advanced techniques have uncovered novel interactions, such as acetylation-driven changes in mitochondrial proteins, which underlie energy deficits in HF, and elucidated mechanisms of existing therapies like beta-blockers [58]. These findings highlight proteomics as a tool for identifying therapeutic targets and biomarkers, though translating these insights into clinical applications requires further validation [56].
4.4. Metabolomic Alterations and Cellular Energetics
Metabolomics has detailed perturbations in lipid metabolism, amino acid processing, and nucleotide pathways, reflecting altered cellular energetics, protein turnover, and cell death in HF [59,60]. These changes underscore the energy crisis in the failing heart, with reduced ATP production linked to mitochondrial dysfunction [61]. For example, studies have identified elevated branched-chain amino acid levels in HFrEF, correlating with disease severity, and altered lipid profiles as early markers of cardiac stress [62]. Mapping these metabolic shifts has revealed targets like ketone metabolism for improving energy efficiency, alongside potential biomarkers for early detection [63].
Thus, integrating multi-omic datasets using artificial intelligence (AI) and deep learning has identified precise biomarkers for HF diagnosis and prognosis, such as circulating microRNAs and proteomic signatures predictive of acute decompensation [64]. However, challenges include validating these biomarkers in diverse cohorts, interpreting complex data, and addressing high costs and limited accessibility [64]. Ethical considerations, including genomic data privacy and informed consent, also pose hurdles, necessitating robust frameworks to ensure equitable application [64]. The future of omics in HF hinges on overcoming these barriers, with potential to deliver personalized treatments by combining genomic risk scores with metabolic and proteomic profiles [65].
5. Advanced Molecular Therapies
Advanced molecular therapies aim to regenerate myocardium or correct underlying defects in HF, leveraging stem cells and genetic modification techniques with promising yet evolving applications.
5.1. Stem Cell Therapy in Heart Failure
Stem cell therapy seeks to repair lost myocardium or stimulate endogenous repair using mesenchymal stem cells (MSCs), induced pluripotent stem cells (iPSCs), and cardiac progenitor cells (CPCs) [66]. MSCs, derived from bone marrow or adipose tissue, exert therapeutic effects primarily through paracrine mechanisms, releasing cytokines such as vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF), and interleukin-10 (IL-10) to promote angiogenesis, reduce inflammation, and attenuate fibrosis [66]. A 2022 meta-analysis of 34 clinical trials in post-myocardial infarction (MI) HF patients reported a modest left ventricular ejection fraction (LVEF) improvement of 3.8% (95% CI 1.2-6.4) at 6 months, though 12-month mortality remained unchanged (OR 0.89; p=0.34) [67]. Lineage-tracing studies confirm that differentiation into functional cardiomyocytes is minimal (<1%), emphasizing paracrine effects as the dominant mechanism [68].
iPSCs, generated by reprogramming somatic cells (for example, fibroblasts), offer scalable production of autologous cardiomyocytes [69]. In a 2023 porcine MI model, bioengineered cardiac patches with iPSC-derived cardiomyocytes achieved electrical integration with native myocardium, boosting LVEF by 12% at 12 weeks, as assessed by cardiac magnetic resonance imaging (MRI) [69]. However, risks include teratoma formation (5-10% incidence in animal models) and ventricular arrhythmias (15% incidence) due to electrical immaturity, though co-culture with endothelial cells or electrical stimulation reduced arrhythmia rates by 50% in preliminary studies [70,71].
CPCs, isolated from human cardiac tissue, provide tissue-specific repair potential [72]. The ALLSTAR trial (2017-2022) evaluated intramyocardial delivery of allogeneic CPCs in chronic post-MI HF, reporting an 8.1 mL reduction in left ventricular end-systolic volume (p=0.03) at 6 months, though no significant differences in major adverse events (e.g., death, hospitalization) were observed at 12 months versus placebo [72]. Advances in bioengineering, such as three-dimensional (3D) hydrogel scaffolds with MSCs, have increased post-transplantation cell survival from 25% to 65% at 4 weeks in murine models, enhancing neovascularization and tissue integration [73]. Despite these advances, challenges persist, including poor cell retention (<10% survival at 30 days post-injection), variability in delivery methods (intracoronary, intramyocardial, or intravenous), and limited differentiation into functional cardiomyocytes [74]. Ongoing research for 2024-2025 explores preconditioning with hypoxia or growth factors like insulin-like growth factor-1 (IGF-1), improving viability by 30% in preclinical models [75].
5.2. Genetic Modification in Heart Failure
Genetic modification offers strategies to correct mutations or enhance repair, encompassing targeted genome editing (CRISPR/Cas9, zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), meganucleases) and non-integrative gene delivery (viral vectors, mRNA, transposons) [76]. CRISPR/Cas9, valued for its simplicity, corrected a MYH7 mutation in hypertrophic cardiomyopathy (HCM) patient-derived iPSCs, restoring 90% of cardiomyocyte contractility in vitro via calcium transients [76]. In a murine laminopathy-associated dilated cardiomyopathy (DCM) model, AAV9-delivered CRISPR reduced fibrosis by 45% and improved LVEF by 14% (p<0.01) at 8 weeks [77]. Post-MI, silencing TGF-β1 with CRISPR decreased scar formation by 30% and enhanced LVEF by 10% at 6 weeks in mice [78]. However, off-target effects (4% in 2023 porcine trials) and low delivery efficiency (15-25% of cardiomyocytes transduced) limit clinical use, with base editing and prime editing (95% precision in 2024 murine models) under evaluation [79,80].
ZFNs corrected a TTN mutation in iPSC-derived cardiomyocytes, improving contractility by 70% in vitro, while silencing connective tissue growth factor (CTGF) in ischemic HF models reduced fibrosis by 20% [81,82]. Their specific design limits flexibility compared to CRISPR [83]. TALENs overexpressed VEGF-A in post-MI murine models, increasing capillary density by 35% and LVEF by 8% at 8 weeks, while transposons like PiggyBac promote cardiomyocyte proliferation [84,85]. AAV9 vectors delivering VEGF-A in porcine HF models boosted capillary density by 40% and LVEF by 11% at 12 weeks, though limited cargo capacity (4.7 kb) and immunity in 50% of humans pose barriers [86,87]. Synthetic mRNA via lipid nanoparticles increased capillary density by 45% and LVEF by 11% at 4 weeks in post-MI mice, with storage and cost challenges remaining [88,89].

Table 1.
Summary of Advanced Molecular Therapies in Heart Failure.
| Therapy | Method | Advantages | Disadvantages | Current Use |
|---|---|---|---|---|
| Stem Cell Therapy | ||||
| Mesenchymal Stem Cells (MSCs) | Derived from bone marrow or adipose tissue; delivered via intracoronary, intramyocardial, or intravenous routes; exert paracrine effects via cytokines (VEGF, HGF, IL-10). | Promotes angiogenesis, reduces inflammation and fibrosis; modest LVEF improvement (3.8% at 6 months) in post-MI HF [67]. | Poor cell retention (<10% survival at 30 days); minimal differentiation into cardiomyocytes (<1%); no mortality reduction [67,68,74]. | Clinical trials; experimental preconditioning with hypoxia or IGF-1 [67,75]. |
| Induced Pluripotent Stem Cells (iPSCs) | Reprogrammed somatic cells (fibroblasts) into cardiomyocytes; used in bioengineered cardiac patches [69]. | Scalable autologous production; significant LVEF increase (12% at 12 weeks) in porcine MI models with electrical integration [69]. | Risks of teratoma formation (5-10%) and arrhythmias (15%) due to electrical immaturity [70]. | Preclinical models; maturation strategies reducing arrhythmias by 50% [71]. |
| Cardiac Progenitor Cells (CPCs) | Isolated from human cardiac tissue; intramyocardial delivery of allogeneic cells [72]. | Tissue-specific repair; reduces LV end-systolic volume (-8.1 mL at 6 months) in chronic post-MI HF [72]. | No significant impact on major adverse events (death, hospitalization) at 12 months [72]. | Clinical trials. |
| Genetic Modification | ||||
| CRISPR/Cas9 | Targeted genome editing to correct mutations or silence genes; delivered via AAV9 or iPSCs [78]. | High precision; restores contractility (90% in vitro), reduces fibrosis (45%), improves LVEF (14%) in models [76,77,78]. | Off-target effects (4% in porcine trials); low delivery efficiency (15-25% of cardiomyocytes) [79]. | Preclinical; base/prime editing (95% precision) under evaluation [80]. |
| Zinc Finger Nucleases | Early genome-editing tool to correct TTN mutations or silence CTGF; delivered to iPSC-derived cardiomyocytes [81]. | Improves contractility (70% in vitro); reduces fibrosis (20%) in ischemic HF models [81,82]. | Labor-intensive design; less flexible than CRISPR [83]. | Limited use; largely superseded by CRISPR [81]. |
| TALENs & Meganucleases | Precision editing to overexpress VEGF-A or promote proliferation; delivered in murine models [84,85,86,87]. | Increases capillary density (35%) and LVEF (8%) in post-MI models; alternative to CRISPR [84]. | Complex design; limited scalability compared to newer tools [86]. | Preclinical |
| Viral Vectors (e.g., AAV9) | Non-integrative gene delivery of VEGF-A or SERCA2a; intracoronary or intramyocardial injection [88]. | Boosts angiogenesis (40% capillary density) and LVEF (11%) in porcine HF; robust expression [88]. | Limited cargo capacity (4.7 kb); pre-existing immunity in 50% of humans [89]. | Clinical trials (CUPID for SERCA2a); ongoing optimization [91]. |
| Synthetic mRNA | Transient gene expression (VEGF-A) via lipid nanoparticles; intramyocardial injection [92]. | Enhances capillary density (45%) and LVEF (11%) in mice; safe, no genomic integration [92,93,94]. | High cost, storage challenges; transient effect limits duration [93]. | Preclinical; adapted from mRNA vaccine technology [94]. |
| Transposons (e.g., PiggyBac) | Non-viral gene integration to promote cardiomyocyte proliferation [95]. | Versatile; enhances regeneration in murine models [97]. | Potential mutagenicity; less precise than CRISPR [96]. | Early preclinical; combined with stem cells [97]. |
| Synergistic Approaches | Combines stem cells with genetic editing (e.g., CRISPR-iPSCs with GATA4/TBX5, TALEN-MSCs with HGF) [93,94]. | High differentiation efficiency (92%); reduces apoptosis (35%), increases angiogenesis (40%) [93,94]. | Combines limitations of both approaches (e.g., retention, off-target effects) [93]. | Preclinical; advancing toward phase II/III trials by 2025 [95]. |
Synergistic approaches, such as CRISPR-edited iPSCs overexpressing GATA4 and TBX5 (92% differentiation efficiency) or TALEN-modified MSCs with HGF (35% less apoptosis, 40% more angiogenesis), enhance regeneration [93,94]. Advances like bioengineered scaffolds and safer delivery systems aim to overcome scalability and regulatory hurdles by 2025 [95,96,97,98,99,100].
6. Personalized Medicine in Heart Failure
Personalized medicine leverages molecular profiling via genomics, transcriptomics, epigenomics, and proteomics to enhance HF diagnosis and treatment, tailoring therapies to individual profiles [101].
One of the main aspects of personalized medicine and HF remains between therapies and oxidative stress. Reactive oxygen species (ROS), such as superoxide and hydrogen peroxide, regulate cardiac myocyte growth and apoptosis, with mitochondria as both a source and target of oxidative damage [101]. Proinflammatory cytokines like TNF-α drive mitochondrial ROS production, contributing to HF progression, while imbalanced mitochondrial fission and fusion exacerbate ischemia-reperfusion injury [102,103,104].
Figure 4.
The key difference between personalized and standard medicine lies in identifying similar characteristics in phenotypic aspects (lipidome, genome, microbiome, metabolome, etc.), which influence specific responses and advantages. This approach adjusts treatments not only based on clinical or demographic characteristics but also on cellular and molecular factors.
Figure 4.
The key difference between personalized and standard medicine lies in identifying similar characteristics in phenotypic aspects (lipidome, genome, microbiome, metabolome, etc.), which influence specific responses and advantages. This approach adjusts treatments not only based on clinical or demographic characteristics but also on cellular and molecular factors.
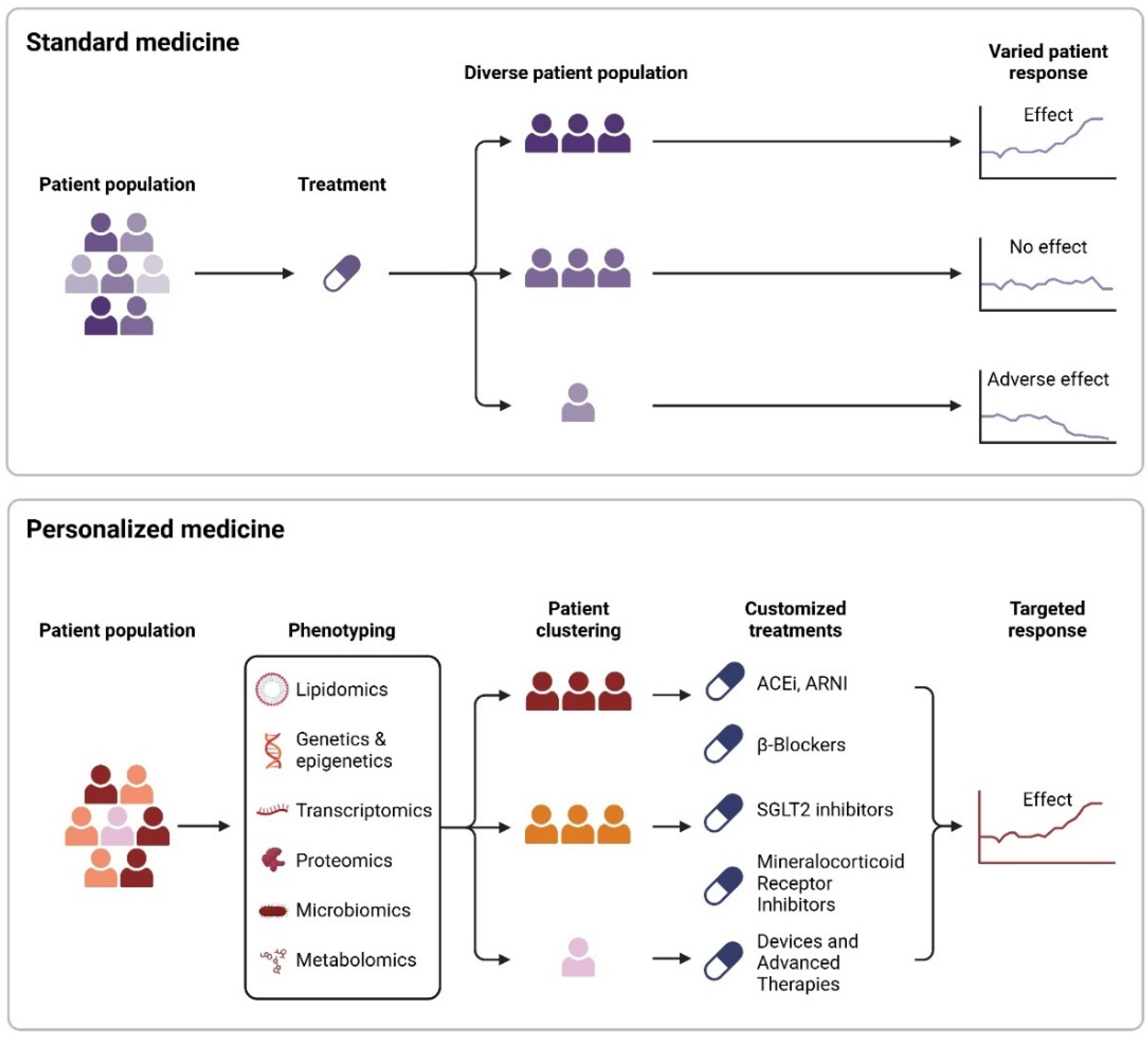
6.1. Therapeutic Interventions
Structured physical training (aerobic, resistance, or respiratory) improves nitric oxide (NO) bioavailability by optimizing endothelial nitric oxide synthase (eNOS) function and antioxidant enzyme expression, mobilizing endothelial progenitor cells and reducing TNF-α, IL-10, and IL-6 levels [106]. ACEIs and ARBs reduce mortality, enhancing endothelial function via increased flow-mediated dilation and arterial elasticity, with perindopril lowering E-selectin and boosting NO in HFrEF and HFpEF [101,105,106]. Sacubitril/valsartan, an angiotensin receptor-neprilysin inhibitor (ARNI), inhibits natriuretic peptide degradation, improving vasodilation and natriuresis; a 12-week study in 80 HFrEF patients showed increased LVEF, NO, and flow-mediated dilation, with reduced endothelin-1 (ET-1) [106,107].
Beta-blockers like carvedilol (acting on β2 and α1 receptors) reduce mitochondrial oxygen consumption and ROS, achieving a 64% mortality reduction in 1996, unlike metoprolol, while improving L-arginine, L-citrulline, and vascular cell adhesion molecule-1 (VCAM-1) levels [101,108]. Sodium-glucose cotransporter 2 inhibitors (SGLT2i), such as empagliflozin, counter anaerobic glycolysis in HF by increasing ketone production, inhibiting Na+/H+ exchanger activity, and reducing ROS and arrhythmogenesis, with benefits in HFpEF via eNOS-dependent pathways [109]. Mineralocorticoid receptor antagonists (e.g., spironolactone with olmesartan) mitigate oxidative stress in ischemic HF, increasing endothelial progenitor cells (VEGFR2+/CD34+) [101,103,106].
In diabetic cardiomyopathy, metformin enhances autophagy via AMPK activation, increasing LC3-II and mitochondrial respiration [101]. Statins offer anti-inflammatory and antioxidant effects, stabilizing eNOS mRNA, reducing TNF-α, and promoting CD34+ cell activation for neovascularization and LVEF improvement [106]. Pharmacological inhibition of dynamin-related protein-1 (DRP-1) reduces cell death post-ischemia-reperfusion, while antioxidants (e.g., allopurinol) and Szeto-Schiller peptides (e.g., SS-31 in PROGRESS-HF) show limited structural benefits [101,102].
These therapies target endothelial dysfunction and oxidative stress, with ongoing research needed to optimize clinical outcomes [106].
7. Conclusions, Challenges and Future Directions
HF remains a complex syndrome driven by RAAS overactivation and neurohormonal imbalance, with chronic SNS and aldosterone effects amplifying remodeling, as detailed earlier [9,12]. Despite progress, challenges persist. RAAS inhibitors excel in HFrEF but show limited efficacy in HFpEF [12]. Stem cell therapies (MSCs, iPSCs, CPCs) offer regenerative potential, yet poor retention (<10% survival at 30 days) and variable delivery methods (e.g., intracoronary, intramyocardial, intravenous) restrict impact, with modest LVEF gains (3.8-12%) and unresolved risks like arrhythmias [67,69,74]. CRISPR/Cas9 and viral vectors target mutations, but off-target effects (4% in 2023 porcine trials) and low transduction efficiency (15-25% of cardiomyocytes) impede translation [79]. The gut-heart axis links dysbiosis and TMAO to inflammation, amplifying HF via immune activation, yet lacks actionable therapies [29]. Omics technologies promise biomarkers (e.g., microRNAs, metabolic profiles), yet face validation, cost, and ethical hurdles [64]. Personalized therapies (e.g., SGLT2i, ARNI) improve endothelial function, though structural benefits remain inconsistent [106].
Future research targets HFpEF with novel inflammation and fibrosis therapies, building on RAAS insights [12]. Stem cell advances, such as preconditioning with IGF-1 (30% viability boost) and 3D scaffolds, aim to enhance efficacy, potentially synergizing with genetic modification [75,93]. Genetic tools like base editing (95% precision in 2024) and lipid nanoparticles promise precision [80]. Microbiome interventions, including probiotics or TMAO inhibitors, are anticipated for 2025 trials to mitigate inflammation [43]. AI-driven omics could validate biomarkers like lncRNAs and lipid profiles, supporting cost-effective care models [64].
In conclusion, HF’s molecular complexity, from RAAS-driven remodeling to microbiome and omics-derived insights, is increasingly understood, yet therapeutic gaps, especially in HFpEF, remain unsolved [10]. Advances in stem cells, genetics, and personalized medicine, bolstered by microbiome and omics research, signal a shift toward regenerative care by 2030, contingent on overcoming translational barriers through global collaboration [65].
Author Contributions
Conceptualization, Manuel Mallol-Simmonds; Investigation, Manuel Mallol-Simmonds, Cristian Avila, Ivan Canete, Alfredo Parra, Josseline Pena; Writing – Original Draft Preparation, Manuel Mallol-Simmonds, Cristian Avila, Ivan Canete, Alfredo Parra, Josseline Pena; Writing – Review & Editing, Manuel Mallol-Simmonds; Supervision, Sergio Bustamante
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest
Abbreviations
The following abbreviations are used in this manuscript:
| ACE | Angiotensin-converting enzyme |
| ACEIs | ACE inhibitors |
| AI | Artificial intelligence |
| ARBs | Angiotensin II receptor blockers |
| ARNI | Angiotensin receptor-neprilysin inhibitor |
| AT1 | Angiotensin II type 1 receptor |
| cGMP | Cyclic guanosine monophosphate |
| CNS | Central nervous system |
| CPCs | Cardiac progenitor cells |
| CRISPR | Clustered regularly interspaced short palindromic repeats |
| DCM | Dilated cardiomyopathy |
| DRP-1 | Dynamin-related protein-1 |
| eNOS | Endothelial nitric oxide synthase |
| ET-1 | Endothelin-1 |
| FMO | Flavin-containing monooxygenase |
| HF | Heart failure |
| HFrEF | Heart failure with reduced ejection fraction |
| HFpEF | Heart failure with preserved ejection fraction |
| HCM | Hypertrophic cardiomyopathy |
| HGF | Hepatocyte growth factor |
| IGF-1 | Insulin-like growth factor-1 |
| IL-10 | Interleukin-10 |
| iPSCs | Induced pluripotent stem cells |
| lncRNAs | Long non-coding RNAs |
| LVEF | Left ventricular ejection fraction |
| MI | Myocardial infarction |
| MSCs | Mesenchymal stem cells |
| NO | Nitric oxide |
| RAAS | Renin-angiotensin-aldosterone system |
| ROS | Reactive oxygen species |
| SGLT2i | Sodium-glucose cotransporter 2 inhibitors |
| SNS | Sympathetic nervous system |
| TALENs | Transcription activator-like effector nucleases |
| TMA | Trimethylamine |
| TMAO | Trimethylamine N-oxide |
| TNF-α | Tumor necrosis factor-alpha |
| VCAM-1 | Vascular cell adhesion molecule-1 |
| VEGF | Vascular endothelial growth factor |
| VO₂ | Peak oxygen consumption |
| ZFNs | Zinc finger nucleases |
References
- McMurray, J.J.; Adamopoulos, S.; Anker, S.D.; Auricchio, A.; Böhm, M.; Dickstein, K.; Falk, V.; Filippatos, G.; Fonseca, C.; Gomez-Sanchez, M.A.; et al. ESC Committee for Practice Guidelines. ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure 2012: The Task Force for the Diagnosis and Treatment of Acute and Chronic Heart Failure 2012 of the European Society of Cardiology. Developed in collaboration with the Heart Failure Association (HFA) of the ESC. Eur. Heart J. 2012, 33, 1787–1847. [Google Scholar] [CrossRef] [PubMed]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; et al. Heart disease and stroke statistics—2020 update: a report from the American Heart Association. Circulation. 2020, 141, e139–e596. [Google Scholar] [PubMed]
- Mosterd, A.; Hoes, A.W. Clinical epidemiology of heart failure. Heart. 2007, 93, 1137–46. [Google Scholar] [PubMed]
- Okamoto, R.; Kario, K.; Inoue, T.; Kanai, H.; Ohashi, N.; Ito, H.; et al. Epidemiology of heart failure in Japan: A systematic review. J Cardiol. 2021, 77, 447–55. [Google Scholar]
- Bocchi, E.A.; Arias, A.; Verdejo, H.; Diez, M.; Gómez, E.; Castro, P. The Reality of Heart Failure in Latin America. J Am Coll Cardiol. 2013, 62, 949–958. [Google Scholar] [CrossRef]
- Ho, K.K.; Pinsky, J.L.; Kannel, W.B.; Levy, D. The epidemiology of heart failure: The Framingham Study. Circ. 1993, 22, A6–A13. [Google Scholar] [CrossRef]
- Heidenreich, P.A.; Albert, N.M.; Allen, L.A.; Bluemke, D.A.; Butler, J.; Fonarow, G.C.; et al. Forecasting the impact of heart failure in the United States: a policy statement from the American Heart Association. Circ Heart Fail. 2013, 6, 606–19. [Google Scholar] [CrossRef]
- Cook, C.; Cole, G.; Asaria, P.; Jabbour, R.; Francis, D.P. The annual global economic burden of heart failure. Int J Cardiol. 2014, 171, 368–76. [Google Scholar]
- Mann, D.L. Mechanisms and models in heart failure: A combinatorial approach. Circulation 1999, 100, 999–1008. [Google Scholar]
- Yang, J.; Xu, W.W.; Hu, S.J. Heart failure and the microbiome: A new frontier in cardiovascular disease. World J Cardiol. 2021, 13, 447–58. [Google Scholar]
- Mann, D.L. Innate immunity and the failing heart: the cytokine hypothesis revisited. Circ. Res. 2015, 116, 1254–1268. [Google Scholar] [CrossRef] [PubMed]
- Rouleau, J.L.; Moyé, L.A.; de Champlain, J.; Klein, M.; Bichet, D.; Packer, M.; Sussex, B.; Arnold, J.; Sestier, F.; Parker, J.O.; et al. Activation of neurohumoral systems following acute myocardial infarction. Am. J. Cardiol. 1991, 68, 80–86. [Google Scholar] [CrossRef]
- Piepoli, M.; et al. Contribution of muscle afferents to the hemodynamic, autonomic, and ventilatory responses to exercise in patients with chronic heart failure: effects of physical training. Circulation 1996, 93, 940–952. [Google Scholar] [CrossRef]
- Giannoni, A.; Emdin, M.; Bramanti, F.; Iudice, G.; Francis, D.P.; Barsotti, A.; Piepoli, M.; Passino, C. Combined Increased Chemosensitivity to Hypoxia and Hypercapnia as a Prognosticator in Heart Failure. Circ. 2009, 53, 1975–1980. [Google Scholar] [CrossRef] [PubMed]
- Ponikowski, P.P.; Chua, T.P.; Francis, D.P.; Capucci, A.; Coats, A.J.; Piepoli, M.F. Muscle Ergoreceptor Overactivity Reflects Deterioration in Clinical Status and Cardiorespiratory Reflex Control in Chronic Heart Failure. Circulation 2001, 104, 2324–2330. [Google Scholar] [CrossRef]
- Cohn, J.N.; Levine, T.B.; Olivari, M.T.; Garberg, V.; Lura, D.; Francis, G.S.; Simon, A.B.; Rector, T. Plasma Norepinephrine as a Guide to Prognosis in Patients with Chronic Congestive Heart Failure. New Engl. J. Med. 1984, 311, 819–823. [Google Scholar] [CrossRef]
- Dzau, V.J.; Colucci, W.S.; Hollenberg, N.K.; Williams, G.H. Relation of the renin-angiotensin-aldosterone system to clinical state in congestive heart failure. Circulation 1981, 63, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.S.; Benedict, C.; E Johnstone, D.; Kirlin, P.C.; Nicklas, J.; Liang, C.S.; Kubo, S.H.; Rudin-Toretsky, E.; Yusuf, S. Comparison of neuroendocrine activation in patients with left ventricular dysfunction with and without congestive heart failure. A substudy of the Studies of Left Ventricular Dysfunction (SOLVD). Circulation 1990, 82, 1724–1729. [Google Scholar] [CrossRef]
- Hartupee, J.; Mann, D.L. Neurohormonal activation in heart failure with reduced ejection fraction. Nat. Rev. Cardiol. 2017, 14, 30–38. [Google Scholar] [CrossRef]
- Notarius, C.F.; Millar, P.J.; Floras, J.S. Muscle sympathetic activity in resting and exercising humans with and without heart failure. Appl. Physiol. Nutr. Metab. 2015, 40, 1107–1115. [Google Scholar] [CrossRef]
- Weinberger, M.H.; Aoi, W.; Henry, D.P. Direct effect of beta-adrenergic stimulation on renin release by the rat kidney slice in vitro. Circ. Res. 1975, 37, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Bekheirnia, M.R.; Schrier, R.W. Pathophysiology of water and sodium retention: edematous states with normal kidney function. Curr. Opin. Pharmacol. 2006, 6, 202–207. [Google Scholar] [PubMed]
- Shah, A.M.; Mann, D.L. In search of new therapeutic targets and strategies for heart failure: recent advances in basic science. Lancet 2011, 378, 704–712. [Google Scholar]
- Mann, D.L.; Bristow, M.R. Mechanisms and models in heart failure: the biomechanical model and beyond. Circulation 2005, 111, 2837–2849. [Google Scholar] [PubMed]
- Arai, M.; Alpert, N.R.; MacLennan, D.H.; Barton, P.; Periasamy, M. Alterations in sarcoplasmic reticulum gene expression in human heart failure. Circ. Res. 1993, 72, 463–469. [Google Scholar] [CrossRef]
- Hasenfuss, G.; Reinecke, H.; Studer, R.; Meyer, M.; Pieske, B.; Holtz, J.; Holubarsch, C.; Posival, H.; Just, H.; Drexler, H. Relation between myocardial function and expression of sarcoplasmic reticulum Ca(2+)-ATPase in failing and nonfailing human myocardium. Circ. Res. 1994, 75, 434–442. [Google Scholar] [CrossRef]
- Tang, W.W.; Hazen, S.L. The contributory role of gut microbiota in cardiovascular disease. J. Clin. Investig. 2014, 124, 4204–4211. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-Gut Microbiota Metabolic Interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
- Trøseid, M.; Andersen, G.; Broch, K.; et al. The gut microbiome in heart failure. J Am Coll Cardiol. 2023, 81, 371–382. [Google Scholar]
- Costello, E.K.; Stagaman, K.; Dethlefsen, L.; et al. Diversity and stability of the human gut microbiota. PLoS One. 2020, 15, e0233872. [Google Scholar]
- Brunkwall, L.; Orho-Melander, M. The gut microbiome in cardiovascular disease and health. Endocr Rev. 2023, 44, 144–156. [Google Scholar]
- Tang, W.W.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; et al. Trimethylamine-N-oxide and risk of atherosclerosis. Eur J Prev Cardiol. 2023, 30, 892–900. [Google Scholar]
- Dorrestein, P.C.; Mazmanian, S.K.; Knight, R. Gut microbiome and host immunity. Mucosal Immunol. 2020, 13, 39–49. [Google Scholar]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut-brain axis and mental health. Microbiome Res. 2021, 14, 285–299. [Google Scholar]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; et al. Gut microbiota in obesity and metabolic disorders. Nutr Metab Cardiovasc Dis. 2023, 33, 687–699. [Google Scholar]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2021, 184, 4168–4180. [Google Scholar]
- Wang, B.; Yao, M.; Lv, L.; et al. The gut microbiota's role in cardiovascular disease. Transl Res. 2020, 226, 83–92. [Google Scholar]
- Keller, A.; Chiu, S.; Jenkins, N.; et al. Anesthesia and gut microbiota. Br J Anaesth. 2002, 89, 50–56. [Google Scholar]
- Knapp, S. Bacterial infections and diabetes. JCHF. 2015, 3, 345–354. [Google Scholar]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Haiser, H.J.; Seim, K.L.; Balskus, E.P.; et al. Molecular basis for gut microbiome interactions with digoxin therapy. Science. 2013, 341, 295–298. [Google Scholar]
- Romano, K.A.; Martinez-Del Campo, A.; Kasahara, K.; et al. Metabolic, epigenetic, and transgenerational effects of gut microbiota. Sci China Life Sci. 2018, 61, 1109–1121. [Google Scholar]
- Schugar, R.C.; Gliniak, C.M.; Osborn, L.J.; et al. Gut microbiota metabolism and cardiovascular disease. J Am Heart Assoc. 2019, 8, e011347. [Google Scholar]
- Reuter, S.E.; O’Mara, M. Mechanisms of drug metabolism in heart failure patients. Clin Pharmacokinet. 2004, 43, 127–139. [Google Scholar]
- Krishnan, S.; Alden, N.; Lee, K. Metabolomics and cardiovascular disease. Cell Metab. 2012, 16, 19–30. [Google Scholar]
- Trøseid, M.; Ueland, T.; Hov, J.R.; Svardal, A.; Gregersen, I.; Dahl, C.P.; Aakhus, S.; Gude, E.; Bjørndal, B.; Halvorsen, B.; et al. Microbiota-dependent metabolite trimethylamine-N-oxide is associated with disease severity and survival of patients with chronic heart failure. Am J Clin Nutr. 2022, 114, 571–580. [Google Scholar]
- Tebani, A.; Afonso, C.; Marret, S.; Bekri, S. Omics-Based Strategies in Precision Medicine: Toward a Paradigm Shift in Inborn Errors of Metabolism Investigations. Int. J. Mol. Sci. 2016, 17, 1555. [Google Scholar] [CrossRef]
- Li, C.; Gill, R.; Lu, D.; Eres, I.; Lu, J.; Cui, J.; et al. Dissecting regulatory non-coding GWAS loci reveals fibroblast causal genes with pathophysiological relevance to heart failure.
- Gill, R.; Lu, D.; Eres, I.; Lu, J.; Cui, J.; Yu, Z.; et al. Dissecting regulatory non-coding heart disease GWAS loci with high-resolution 3D chromatin interactions reveals causal genes with pathophysiological relevance to heart failure.
- Aiyer, S.; Kalutskaya, E.; Agdamag, A.C.; Tang, W.H.W. Genetic Evaluation and Screening in Cardiomyopathies: Opportunities and Challenges for Personalized Medicine. J. Pers. Med. 2023, 13, 887. [Google Scholar] [CrossRef]
- Yi, Q.; Feng, J.; Lan, W.; Shi, H.; Sun, W.; Sun, W. CircRNA and lncRNA-encoded peptide in diseases, an update review. Mol. Cancer 2024, 23, 214. [Google Scholar] [CrossRef]
- Zhou, R.; Yang, G.; Zhang, Y.; Wang, Y. Spatial transcriptomics in development and disease. Mol. Biomed. 2023, 4, 1–22. [Google Scholar] [CrossRef]
- Kuppe, C.; Flores, R.O.R.; Li, Z.; Hayat, S.; Levinson, R.T.; Liao, X.; Hannani, M.T.; Tanevski, J.; Wünnemann, F.; Nagai, J.S.; et al. Spatial multi-omic map of human myocardial infarction. Nature 2022, 608, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Dzau, V.J.; Hodgkinson, C.P. RNA Therapeutics for the Cardiovascular System. Circulation 2024, 149, 707–716. [Google Scholar] [CrossRef]
- Shah, A.M.; Myhre, P.L.; Arthur, V.; Dorbala, P.; Rasheed, H.; Buckley, L.F.; Claggett, B.; Liu, G.; Ma, J.; Nguyen, N.Q.; et al. Large scale plasma proteomics identifies novel proteins and protein networks associated with heart failure development. Nat. Commun. 2024, 15, 528. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, K.; Zhao, Y.; Wang, K. Research progress on post-translational modification of proteins and cardiovascular diseases. Cell Death Discov. 2023, 9, 275. [Google Scholar] [PubMed]
- Karpov, O.A.; Stotland, A.; Raedschelders, K.; Chazarin, B.; Ai, L.; Murray, C.I.; Van Eyk, J.E. Proteomics of the heart. Physiol. Rev. 2024, 104, 931–982. [Google Scholar] [CrossRef]
- Wende, A.R.; Brahma, M.K.; McGinnis, G.R.; Young, M.E. Metabolic Origins of Heart Failure. JACC: Basic Transl. Sci. 2017, 2, 297–310. [Google Scholar] [CrossRef]
- Ng, S.M.; Neubauer, S.; Rider, O.J. Myocardial metabolism in heart failure. Curr Heart Fail Rep. 2023, 20, 63–75. [Google Scholar]
- He, H.; Huang, W.; Pan, Z.; Wang, L.; Yang, Z.; Chen, Z. Intercellular Mitochondrial transfer: Therapeutic implications for energy metabolism in heart failure. Pharmacol. Res. 2024, 211, 107555. [Google Scholar] [CrossRef]
- Lin, C.; Tian, Q.; Guo, S.; Xie, D.; Cai, Y.; Wang, Z.; Chu, H.; Qiu, S.; Tang, S.; Zhang, A. Metabolomics for Clinical Biomarker Discovery and Therapeutic Target Identification. Molecules 2024, 29, 2198. [Google Scholar] [CrossRef]
- Lopaschuk, G.D.; Karwi, Q.G.; Tian, R.; Wende, A.R.; Abel, E.D. Cardiac Energy Metabolism in Heart Failure. Circ. Res. 2021, 128, 1487–1513. [Google Scholar] [CrossRef]
- Bonomi, L.; Huang, Y.; Ohno-Machado, L. Privacy challenges and research opportunities for genomic data sharing. Nat. Genet. 2020, 52, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Molla, G.; Bitew, M. Revolutionizing Personalized Medicine: Synergy with Multi-Omics Data Generation, Main Hurdles, and Future Perspectives. Biomedicines 2024, 12, 2750. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.R.; Samadani, U.; Han, R.; et al. Stem cell therapy for heart failure: a meta-analysis of clinical trials. J Am Coll Cardiol. 2022, 79, 873–85. [Google Scholar]
- Nguyen, P.K.; Rhee, J.-W.; Wu, J.C. Adult stem cell therapy and heart failure, 2000 to 2016: A systematic review. JAMA Cardiol. 2016, 1, 831–841. [Google Scholar] [CrossRef]
- Kawamura, T.; Miyagawa, S.; Fukushima, S.; et al. Cardiomyocyte maturation in human iPSC-derived cardiac tissue constructs. Nat Commun. 2021, 12, 3525. [Google Scholar]
- Chong, J.J.H.; Yang, X.; Don, C.W.; Minami, E.; Liu, Y.W.; Weyers, J.J.; Mahoney, W.M.; Van Biber, B.; Cook, S.M.; Palpant, N.J.; et al. Human embryonic-stem-cell-derived cardiomyocytes regenerate non-human primate hearts. Nature 2014, 510, 273–277. [Google Scholar] [CrossRef]
- Zhang, J.; Bolli, R. Preconditioning of stem cells for cardiac repair: a new frontier. J Mol Cell Cardiol. 2023, 174, 45–53. [Google Scholar]
- Makkar, R.R.; Kereiakes, D.J.; Aguirre, F.; et al. Allogeneic cardiosphere-derived cells for chronic ischemic heart failure: results of the ALLSTAR trial. J Am Coll Cardiol. 2022, 79, 1445–56. [Google Scholar]
- Li, Y.; Zhang, J.; Chen, W.; et al. Engineering 3D hydrogels for cardiac tissue repair: advances and challenges. Bioact Mater. 2023, 15, 123–34. [Google Scholar]
- Madonna, R.; Van Laake, L.W.; Davidson, S.M.; et al. Position paper of the European Society of Cardiology Working Group Cellular Biology of the Heart: cell-based therapies for heart disease. Eur Heart J. 2021, 42, 1626–33. [Google Scholar]
- Mosqueira, D.; Mannhardt, I.; Bhagwan, J.R.; et al. CRISPR/Cas9 editing in human iPSCs corrects MYH7 mutation and restores cardiomyocyte function in hypertrophic cardiomyopathy. Circulation. 2022, 145, 927–30. [Google Scholar]
- Gao, E.; Lei, Y.H.; Shang, J.Y.; et al. CRISPR/Cas9-mediated correction of LMNA mutation attenuates fibrosis in dilated cardiomyopathy mice. Mol Ther. 2022, 30, 1834–45. [Google Scholar]
- Wang, X.; Liu, Z.; Zhang, L.; et al. CRISPR/Cas9-mediated TGF-β1 knockdown reduces myocardial fibrosis post-infarction in mice. J Clin Invest. 2023, 133, e165432. [Google Scholar]
- Musunuru, K.; Chadwick, A.C.; Mizoguchi, T.; et al. In vivo CRISPR editing for cardiovascular disease: promises and perils. Nat Rev Cardiol. 2023, 20, 245–58. [Google Scholar]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR-Cas nucleases, base editors, transposases and prime editors. Nat Biotechnol. 2020, 38, 824–44. [Google Scholar] [PubMed]
- Maeder, M.L.; Thibodeau-Beganny, S.; Osiak, A.; Wright, D.A.; Anthony, R.M.; Eichtinger, M.; Jiang, T.; Foley, J.E.; Winfrey, R.J.; Townsend, J.A.; et al. Rapid “Open-Source” Engineering of Customized Zinc-Finger Nucleases for Highly Efficient Gene Modification. Mol. Cell 2008, 31, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, S.; Chen, Y.; et al. ZFN-mediated CTGF silencing attenuates fibrosis in ischemic heart failure models. Cardiovasc Res. 2019, 115, 1345–55. [Google Scholar]
- Carroll, D. Genome engineering with zinc-finger nucleases. Genetics 2011, 188, 773–782. [Google Scholar] [CrossRef]
- Joung, J.K.; Sander, J.D. TALENs: a widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Biol. 2013, 14, 49–55. [Google Scholar] [CrossRef]
- Sun, N.; Zhao, H.; Ren, X.; et al. TALEN-mediated VEGFA overexpression enhances angiogenesis in post-myocardial infarction mice. Circ Res. 2021, 129, 412–25. [Google Scholar]
- Bogdanove, A.J.; Voytas, D.F. TAL Effectors: Customizable Proteins for DNA Targeting. Science 2011, 333, 1843–1846. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, B.L. Homing Endonucleases: From Microbial Genetic Invaders to Reagents for Targeted DNA Modification. Structure 2011, 19, 7–15. [Google Scholar] [CrossRef]
- Silva, G.; Poirot, L.; Galetto, R.; Smith, J.; Montoya, G.; Duchateau, P.; Paques, F. Meganucleases and Other Tools for Targeted Genome Engineering: Perspectives and Challenges for Gene Therapy. Curr. Gene Ther. 2011, 11, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Tilemann, L.; Ishikawa, K.; Weber, T.; Hajjar, R.J. Gene Therapy for Heart Failure. Circ. Res. 2012, 110, 777–793. [Google Scholar] [CrossRef]
- Zacchigna, S.; Zentilin, L.; Giacca, M. Adeno-Associated Virus Vectors as Therapeutic and Investigational Tools in the Cardiovascular System. Circ. Res. 2014, 114, 1827–1846. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, C.T.; Deshpande, P.S.; Dever, D.P.; et al. Identification of pre-existing adaptive immunity to Cas9 proteins in humans. Nat Med. 2019, 25, 249–54. [Google Scholar]
- Wang, D.; Tai, P.W.L.; Gao, G. Lentiviral vector platforms for gene therapy. Mol Ther Methods Clin Dev. 2019, 12, 245–57. [Google Scholar]
- Jessup, M.; Greenberg, B.; Mancini, D.; et al. Calcium upregulation by percutaneous administration of gene therapy in cardiac disease (CUPID): a phase 2 trial of intracoronary gene therapy of sarcoplasmic reticulum Ca2+-ATPase in patients with advanced heart failure. Circulation. 2011, 124, 304–13. [Google Scholar]
- Zangi, L.; Lui, K.O.; von Gise, A.; et al. Modified mRNA directs the fate of heart progenitor cells in myocardial infarction models. Nat Biotechnol. 2013, 31, 898–907. [Google Scholar]
- Sahin, U.; Karikó, K.; Türeci, Ö. mRNA-based therapeutics—Developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; et al. mRNA vaccines—a new era in vaccinology. Nat Rev Drug Discov. 2018, 17, 261–79. [Google Scholar] [PubMed]
- Yusa, K.; Zhou, L.; Li, M.A.; Bradley, A.; Craig, N.L. A hyperactive piggyBac transposase for mammalian applications. Proc. Natl. Acad. Sci. 2011, 108, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Ivics, Z.; Izsvák, Z. Transposons for gene therapy! Curr Gene Ther. 2006, 6, 593–607. [Google Scholar]
- Wang, H.; Yang, Y.; Liu, J.; et al. Cardiomyocyte-specific transposon-mediated gene delivery enhances cardiac repair in mice. Mol Ther. 2022, 30, 2134–45. [Google Scholar]
- Sena-Esteves, M.; Gao, G. Introducing Genes into Mammalian Cells: Viral Vectors. Mol Ther Methods Clin Dev. 2020, 17, 431–438. [Google Scholar]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; et al. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature. 2017, 551, 464–71. [Google Scholar]
- Shiba, Y.; Gomibuchi, T.; Seto, T.; et al. CRISPR-enhanced iPSC-derived cardiomyocytes for cardiac tissue engineering. Sci Transl Med. 2023, 15, eabn4215. [Google Scholar]
- Knowlton, A.A.; Chen, L.; Malik, Z.A. Heart failure and mitochondrial dysfunction: The role of mitochondrial fission/fusion abnormalities and new therapeutic strategies. J Cardiovasc Pharmacol. 2014, 63, 196–206. [Google Scholar]
- Bisaccia, G.; Ricci, F.; Gallina, S.; Di Baldassarre, A.; Ghinassi, B. Mitochondrial Dysfunction and Heart Disease: Critical Appraisal of an Overlooked Association. Int. J. Mol. Sci. 2021, 22, 614. [Google Scholar] [CrossRef]
- Gallo, G.; Rubattu, S.; Volpe, M. Mitochondrial Dysfunction in Heart Failure: From Pathophysiological Mechanisms to Therapeutic Opportunities. Int. J. Mol. Sci. 2024, 25, 2667. [Google Scholar] [CrossRef] [PubMed]
- Schwinger, R.H.G. Pathophysiology of heart failure. Cardiovasc Diagn Ther. 2021, 11. [Google Scholar]
- Sabbah, H.N. Targeting the Mitochondria in Heart Failure: A Translational Perspective. JACC Basic Transl Sci. 2020, 5, 88–106. [Google Scholar] [PubMed]
- Tsigkou, V.; Oikonomou, E.; Anastasiou, A.; Lampsas, S.; Zakynthinos, G.E.; Kalogeras, K.; Katsioupa, M.; Kapsali, M.; Kourampi, I.; Pesiridis, T.; et al. Molecular Mechanisms and Therapeutic Implications of Endothelial Dysfunction in Patients with Heart Failure. Int. J. Mol. Sci. 2023, 24, 4321. [Google Scholar] [CrossRef]
- Li, B.-H.; Fang, K.-F.; Lin, P.-H.; Zhang, Y.-H.; Huang, Y.-X.; Jie, H. Effect of sacubitril valsartan on cardiac function and endothelial function in patients with chronic heart failure with reduced ejection fraction. Clin. Hemorheol. Microcirc. 2021, 77, 425–433. [Google Scholar] [CrossRef]
- Packer, M.; Bristow, M.R.; Cohn, J.N.; Colucci, W.S.; Fowler, M.B.; Gilbert, E.M.; Shusterman, N.H. The Effect of Carvedilol on Morbidity and Mortality in Patients with Chronic Heart Failure. New Engl. J. Med. 1996, 334, 1349–1355. [Google Scholar] [CrossRef]
- Kolijn, D.; Pabel, S.; Tian, Y.; Lódi, M.; Herwig, M.; Carrizzo, A.; Zhazykbayeva, S.; Kovács, Á.; Fülöp, G.Á.; Falcão-Pires, I.; et al. Empagliflozin improves endothelial and cardiomyocyte function in human heart failure with preserved ejection fraction via reduced pro-inflammatory-oxidative pathways and protein kinase Gα oxidation. Cardiovasc. Res. 2020, 117, 495–507. [Google Scholar] [CrossRef]
- Karmazyn, M.; Gan, X.T. Probiotics as potential treatments to reduce myocardial remodelling and heart failure via the gut-heart axis: State-of-the-art review. Mol. Cell. Biochem. 2023, 478, 2539–2551. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



