
Submitted:
16 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
Pituitary cells are specialized cells located within the pituitary gland, a small, pea-sized gland situated at the base of the brain. Through the use of cellular electrophysiological techniques, the electrical properties of these cells have been progressively revealed. This review paper aims to introduce the ion currents that are known to be functionally expressed in pituitary cells. These currents include voltage-gated Na+ current (INa), erg-mediated K+ current (IK(erg)), M-type K+ current (IK(M)), hyperpolarization-activated cation current (Ih), and large-conductance Ca2+-activated K+ (BKCa) channel. The biophysical characteristics of the respective ion current were described. Additionally, we also provide explanations for the effect of various drugs or compounds on each of these currents. GH3-cell exposure to GV-58 can increase the magnitude of INa with concurrent raise in the inactivation time constant of the current. The presence of esaxerenone, an antagonist of aldosterone receptor, directly suppress the magnitude of peak and late INa. Risperidone, an atypical antipsychotic agent, is effective at suppressing IK(erg) amplitude directly, and di(2-ethylhexyl)-phthalate suppressed IK(erg). Solifenacin and kynurenic acid can interact with KM channel to stimulate IK(M), while carisbamate and cannabidiol inhibit Ih activated by sustained hyperpolarization. Moreover, either the presence of either rufinamide or QO-40 can enhance the activity of single BKCa channels. To sum it up, alterations in ion currents within native pituitary cells or pituitary tumor cells can influence their functional activity, particularly in processes like stimulus-secretion coupling. The effects of small-molecule modulators, as demonstrated here, bear significance in clinical, therapeutic, and toxicological contexts.
Keywords:
pituitary cell
; voltage-gated ionic currents
; small-molecule modulators
1. Introduction
The pituitary gland, often referred to as the “master gland”, is essential for regulating numerous physiological processes in the body by releasing release hormones. It consists of two main sections, the anterior and posterior pituitary. The anterior pituitary contains various cell types, including somatotrophs, thyrotrophs, corticotrophs, gonadotrophs, melanotrophs, and lactotrophs. Among these, lactotrophs are a specific cell population responsible for secreting prolactin.
By utilizing specialized separation techniques [52], it is possible to immortalize pituitary cells derived from pituitary tumors, enabling in-depth academic research into the functional properties of these cells. Additionally, unlike conventional intracellular recordings that involve impaling or piercing the cell, the patch-clamp technique, which uses a relatively large electrode tip with a pipette resistance of 2-4 MΩ, can be used on small cells with minimal damage, making it suitable for cellular electrophysiological experiments [46,121]. In order words, the electrode used in patch-clamp recordings applies negative pressure to generate suction, which holds the small cell, such as pituitary cell, in place or keeps in a relatively suspended position without displacing or distorting it. This method eliminates the need to impale or puncture the cell, reducing cell damage and enhancing electrophysiological recordings. Figure 1 illustrates a typical population of pituitary tumor cells, specifically GH3 cells.
In this paper, we review the main ion currents in pituitary cells that are influenced by changes in cell membrane voltage, exploring their physiological and pharmacological implications. Figure 2 depicts the major ionic currents commonly observed in pituitary cells. We also provide examples of various drugs or compounds that affect the amplitude and gating kinetics of these ion currents.
Physiological importance and pharmacological impact on voltage-gated ionic currents in pituitary cells
In this review paper, we will provide a detailed description of the potential voltage-gated currents that may occur inn pituitary cells and discuss various drugs or compounds that are currently known to influence these currents (Table 1).
A. Voltage-Gated Na+ Current (INa)
The NaV channels exist in nine isoforms (NaV1.1-1.9, also known as SCN1A-SCN5A and SCN8A-SCN11A) and are found in various mammalian excitable tissues, including the central and peripheral nervous systems, as well as the neuroendocrine or endocrine systems [17,43,110,122]. When activated, NaV channel activity generates a macroscopic INa, which is characterized by its rapid activation and inactivation, both occurring within milliseconds. This current temporarily depolarizes the membrane, providing positive feedback that triggers the upstroke of the action potential (AP). As a result, changes in the magnitude of INa can influence the amplitude, frequency, and patterns of APs in various electrically active cells [17,122]. Previous studies have shown that NaV channels are present in all secretory pituitary cells, including pituitary GH3 cells [122]. Additionally, late (or persistent) INa has been progressively identified, and its presence exerts a significant influence on electrical behavior and firing patterns of pituitary cells [21,30,56,122,143].
- 1. GV-58 ((2R)-2-[(6-{[(5-methylthiophen-2-yl)methyl]amino}-9-propyl-9H-purin-2-yl)amino]butan-1-ol)
GV-58 was developed as a modification of (R)-roscovitine. It has been viewed as an opener of N- and P/Q-type Ca2+ channels [12,81,92,126]. This compound was presumably thought to slow the closing of the voltage-gated Ca2+ (CaV) channel, resulting in a large increase in total Ca2+ entry during motor nerve AP activity [126]. Its presence was reported to enhance spontaneous and evoked activity from the cultures of murine ventral horn of the spinal cord on microelectrode arrays [12]. Earlier studies have demonstrated that this compound is effective for the management of neuromuscular weakness, such as Lambert-Eaton myasthenic syndrome [27, 81, 92; 101, 117, 125].
However, recent studies have demonstrated that, as pituitary GH3 cells were continually exposed to GV-58, the peak and late components of INa activated by abrupt step depolarization were increased in a concentration-, time-, and state-dependent manner [27]. The INa activated by brief depolarizing pulse was sensitive to either block by tetrodotoxin or stimulation by GV-58, but it failed to be affected by ω-conotoxin MVIID. ω-Conotoxin MVIID, a small, disulfide-rich peptide purified from the venoms of predatory cone snails, was reported to be an inhibitor of N- and P/Q-type Ca2+ currents in adrenal chromaffin cells [40]. The recovery of INa inactivation induced with varying interpulse intervals was overly enhanced in the presence of GV-58. The decline of peak INa during rapid repetitive stimuli was slowed during cell exposure to GV-58. This compound stimulated peak INa in a tonic and use-dependent manner. GH3-cell exposure to GV-58 was also found to enhance the magnitude of instantaneous resurgent and window INa. Under current-clamp conditions, GV-58 effectively increased the frequency and spontaneous APs. The NSC-34 motor neuron-like cells could be enhanced by adding GV-58. The mRNA transcripts for the α-subunit of NaV1.1, NaV1.2, and NaV1.6 were reported to be expressed in pituitary GH3 cells [122]. Findings from this study can be interpreted to reflect that GV-58 can interact with NaV channels to stimulate the magnitude and to alter the gating of INa [27]. These results would engage in the modification of spontaneous APs in electrically excitable cells like GH3 and NSC-34 cells, presuming that similar in vitro or in vivo findings occur.
Concentration-dependent stimulation of both peak and late INa was observed with GV-58, and it exhibited effective half-maximal concentration (EC50) values of 8.9 and 2.6 μM for peak and late INa, respectively [27]. Continuous exposure to GV-58 resulted in an increase in the amplitude of peak INa in response to abrupt step depolarization, accompanied by a significant increase in the value of slow component of INa inactivation time constant. It is therefore important to emphasize caution when ascribing the expanding use of GV-58 to its selective agonistic effects on N- and P/Q-type CaV channels [92,101,117].
- 2.
- Esaxerenone (ESAX, Minnebro®, CS-3150, XL-550, (4S)-4-(5,6-dimethyl-2-oxo-1,2-dihydropyridin-3-yl)-2-fluoro-N-(1-methyl-1H-pyrazol-4-yl)benzamide)
ESAX, known to be a newly oral, non-steroidal selective blocker on the activity of mineralocorticoid receptor, has been growingly used for the management of varying pathologic disorders, such as primary aldosteronism, refractory hypertension, chronic kidney disease, diabetic nephropathy, and heart failure [5,64,77,98,148]. Alternatively, the activity of mineralocorticoid receptor has been previously demonstrated in pituitary cells including GH3 cells or in various brain regions [32,41,82,105].
However, earlier studies have shown the ability of ESAX to suppress the peak and late component of INa elicited during short depolarizing pulse in pituitary GH3 cells [21]. The subsequent addition of ESAX reversed tefluthrin-mediated increase in the strength of voltage-dependent hysteresis of persistent INa activated by the isosceles-triangular ramp pulse. Tefluthrin was known to be an activator of INa [83]. In pituitary MMQ cells, the presence of ESAX was effective at decreasing the amplitude and gating of INa.
The IC50 value needed for ESAX-mediated inhibition of peak or late INa observed in GH3 cells was yielded to be 13.2 or 3.2 μM, respectively, the value of which was distinguishable between its suppressive effects on these two components of the current [21]. The presence of neither dexamethasone nor aldosterone affected the magnitude or gating of INa. In continued exposure to aldosterone, further addition of ESAX was still able to suppress peak INa. It therefore seems unlikely that ESAX-mediated inhibition of INa amplitude together with changes in the gating kinetics of the current was predominantly associated with its blockade of mineralocorticoid receptor. Moreover, as shown in Figure 3, the presence of ESAX (10 μM) can effectively diminish the strength in voltage-dependent hysteresis of persistent INa (INa(P)) activated by prolonged isosceles-triangular ramp voltage observed in GH3 cells (21].
B. erg-Mediated K+ Current (IK(erg))
The erg (ether-à-go-go related gene)-mediated K+ current (IK(erg)), which is encoded by three different subfamilies of the KCNH gene, is enabled to generate the pore-forming α-subunit of erg-mediated K+ (i.e., Kerg or KV11) channels [9,51]. These K+ currents are widely believed to represent the cloned equivalent of the rapidly activating delayed-rectifying K+ currents found in cardiac myocytes, and the KCNH2 gene encodes the α-subunit responsible for pore formation in KV11.1 channels, commonly referred to as the human erg K+ (HERG) channels [20,129]. The intrinsic presence of IK(erg) extends beyond excitable cells to various types of epithelial or neoplastic cells, as reported previously [17,129]. Earlier work has also shown the effectiveness of IK(erg) magnitude in regulating the apoptosis and proliferation of various types of neoplastic or stem cells [152].
- 1
- Risperidone (Risperdal®, 3-[2-[4-(6-fluoro-1,2-benzisoxazol-3-yl)piperidin-1-yl]ethyl]-2-methyl-6,7,8,9-tetrahydro-4H-pyrido[1,2-a]pyrimidin-4-one)
Risperidone, a benzisoxazole compound that has been approved for use in the United States in 1994, is known to be effective at terminating acute psychotic episodes and preventing recurrence of psychotic episodes in schizophrenics [11,13]. However, the significant and consistent neuroendocrine effect of neuroleptic drugs including risperidone is to stimulate prolactin secretion and cause galactorrhea, although these untoward effects vary greatly in potency and chemical structure [13,78,123]. The important site of action has been thought to be due to the blockade of dopamine D2 and 5HT receptors [11,93]. However, other evidence suggests that these neuroleptics, including risperidone, may cause a significant prolongation of electrocardiographic QTc interval [18,118].
In a previous study made in pituitary GH3 cells, the exposure to risperidone effectively suppressed the magnitude of IK(erg), together with a slowing in the rate of activation [139]. The results also showed that there was the difference in reciprocal time constants of IK(erg) decays observed at different voltages, suggesting that risperidone may increase the rate of deactivation [139]. It is therefore possible that the different level of membrane potential can exert an interaction with erg-mediated K+ (Kerg) channels to modify the magnitude and gating of whole-cell IK(erg) in GH3 cells. In other words, the sensitivity to risperidone in pituitary lactotrophs would be dependent on the preexisting level of resting membrane potential, the firing rate of AP, or the concentration of risperidone used, assuming that the risperidone action in pituitary lactotrophs is similar to that on GH3 cells [139].
It is worth noting that neither dopamine nor metoclopramide affected the magnitude of IK(erg) in GH3 cells, although haloperidol or thioridazine mimicked the risperidone-mediated inhibition of IK(erg). Metoclopramide was reported to antagonize at dopamine receptor. In GH3 cells preincubated with dopamine, the inhibitory effect of risperidone on IK(erg) remained unaffected. Several tyrosine kinase inhibitors were also found to suppress the IK(erg) magnitude [53]. Therefore, the effect of risperidone on IK(erg) appear to be direct and independent of its binding to dopamine receptors. The carefulness may need to be made in ascribing the risperidone-mediated prolactin release or QT prolongation to those caused by the blockade of dopamine receptors residing in vivo or in vitro [11,16,75]. These findings imply that the risperidone-mediated stimulation of prolactin release could be partly, if not entirely, ascribed to the direct blockade of IK(erg) functionally expressed in pituitary lactotrophs [75,123,139].
- 2
- Di(2-ethylhexyl)-phthalate (DEHP)
Phthalates are a group of chemicals that are mainly used as plasticizers to allow stiff plastics, such as polyvinyl chloride, to become more flexible. Because phthalate plasticizers are not chemically bound to polyvinyl chloride, they may leach, migrate, or evaporate into air and atmosphere, foodstuffs, and other materials [3,70]. Due to their suitable properties and low cost, the general population will become significantly exposed to these compounds [63]. One of the phthalate plasticizers used in a wide variety of medical devices is di(2-ethylhexyl)-phthalate (DEHP), which is recognized to be an endocrine-disrupting chemical [42,94,128,134,154].
Previous work has shown that high doses of DEHP could change cell size or function in anterior pituitary gland [94,154]. DEHP was demonstrated to suppress tamoxifen-induced apoptosis possibly linked to its estrogenic effects as well as to influence signaling pathways in pituitary GH3 cells [38,66]. This compound was also noted to cause an age-dependent influence on the pituitary-adrenocortical axis in vivo [154]. A previous report unveiled that the concentration of phthalate esters, including DEHP, in semen or serum samples were positively associated with circulating prolactin levels in adult men [79]. DEHP was found to impair the electrical and mechanical behavior of cardiac cell network [44].
The study conducted by Wu et al. [142] revealed that DEHP has been observed to reduce the amplitude of IK(erg) in pituitary GH3 cells in a concentration-dependent manner, with an IC50 value of 16.3 μM. This IC50 value is notably lower than the typical concentration of DEHP found in human blood or blood components, which has been reported to range from 10 to 650 μg/ml (equivalent to 27 μM to 1.6 mM) [59]. Exposure of GH3 cells to DHEP was found to alter the activation kinetics of IK(erg) without affecting in the deactivation kinetics of the current. Additionally, the presence of DEHP led to an increase in the firing of spontaneous APs in these pituitary cells.
We conducted a more in-depth exploration of the interaction between the HERG protein and DEHP using PyRx software. Figure 4 illustrates the predicting docking sites of the DEHP molecule. Notably, during the docking process with the HERG channel, DEHP was noted to establish a hydrogen bond with residue Arg537 with a distance of 3.03 Å. Furthermore, DEHP exhibited hydrophobic interactions with several residues, including Lys407, Asp411, Asn470, Thr474, His492, Tyr493, Trp497, Arg541, Lys538, Asp540, and Arg541. These findings suggest a strong binding affinity between DEHP and the amino acid residues of the HERG channel, estimated at -5.9 kcal/mol. This interaction predominantly occurs in the vicinity of the transmembrane region, specifically around positions 496-516 or 521-541 of the channel. The predicted interaction raises concerns about the potential impact on DEHP-mediated alterations in the magnitude and gating kinetics of IK(erg).
Taken together, these results suggest that the inhibition of Kerg channels by DHEP or other chemically related compounds may play a role in the changes observed in the functional activities of pituitary cells, including processes like hormonal release, assuming that similar effects can be replicated in a live, in vivo context [142].
C. M-Type K+ Current (IK(M))
The KCNQ2, KCNQ3, and KCNQ5 genes are responsible for encoding the core subunits of KV7.2, KV7.3, and KV7.5 channels, respectively [1,15]. These K+ channels, when activated, give rise to the IK(M), which is found in various electrically excitable cells, including pituitary cells [15,113,144]. This current is characterized by its low threshold voltage activation and displays a slow activation and deactivation profile [15,113]. The regulation of IK(M) has garnered considerable interest as an adjunctive therapeutic strategy for addressing neurological disorders marked by excessive neuronal activity. These disorders encompass conditions such as cognitive dysfunction, neuropathic pain, and epilepsy [90]. Furthermore, it is believed that the magnitude of IK(M) play a role in regulating the availability of NaV channels during extended periods high-frequency firing [150,153].
- 1. Solifenacin (SOL, Vesicare®, (R)-1-phenyl-3-(1-piperidin-4-ylpropyl)oxy-1,1-diphenyl-4-ylbutan-1-amine)
SOL, a member of isoquinoline, has been viewed as an oral anticholinergic (i.e., a competitive muscarinic [M1 and M3] receptor antagonist) and antispasmodic agent used to treat the symptoms of overactive bladder, neurogenic detrusor overactivity, or urinary incontinence [69,89,108]. It has been reported to be a muscarinic (M2 and M3) receptor antagonist that has anticholinergic effects such relaxation of the detrusor muscle in urinary bladder [2].
Earlier clinical investigations have revealed the efficacy and safety of the antimuscarinic, SOL, for treating patients with overactive bladder or neurogenic detrusor overactivity [2,69]. However, recent evidence has been reported to demonstrate that the treatment with SOL could be linked to an increased risk of the impairment in cognitive functions [48,135]. It is therefore pertinent to reappraise the mechanism of SOL actions on electrical behaviors in varying excitable cells, given that its growing clinical use occurs.
Many types of anterior pituitary cells have been shown to secrete acetylcholine [72]. Earlier studies have also revealed that pituitary GH3 cells could exhibit the activity of muscarinic receptors and that muscarinic agonists were able to inhibit hormonal secretion through a reduction in intracellular cyclic AMP [114]. In these pituitary cells, the binding of acetylcholine to M2-muscarinic receptor might induce a weak stimulation on the hydrolysis of phosphatidylinositol 4,5-bisphosphate [4].
A previous report has shown that in pituitary GH3 cells, during exposure to SOL, the IK(M) amplitude elicited upon membrane depolarization was concentration-dependently increased with an EC50 value of 0.34 μM [28]. The activation time course of IK(M) concurrently became shorted, and the value of dissociation constant (KD) obtained on the basis of minimal reaction scheme was estimated to 0.55 μM. The value of EC50 required for SOL-mediated effect on IK(M) was similar, reflecting that SOL has the propensity to bind to the open or activated state of the channel. There was a leftward shift of the quasi-steady-state activation curve of IK(M) in the presence of SOL. The strength of voltage-dependent hysteresis of IK(M) activated by isosceles-triangular ramp pulse became increased during cell exposure to SOL. Furthermore, the KM-channel activity was elevated by the addition of SOL, without affecting single channel conductance of the channel. However, the mean open time of KM channel after exposure to SOL was increased. Under current-clamp conditions, the firing frequency of spontaneous APs present in GH3 cells was found to be effectively decreased in the presence of SOL. Collectively, findings from these results provide an unanticipated and yet non-canonical ionic mechanisms through which the SOL molecule can interact with KM channel to enhance whole-cell IK(M) and, consequently, to diminish the firing rate of spontaneous APs [28]. It turns out that whether the action of SOL or other structurally similar compounds like darifenacin) on overactive bladder or neurogenic detrusor overactivity [2,89,103] are related to its enhanced actions on KM-channel activity, warrants further investigations, despite its high-affinity binding to muscarinic receptors [2].
- 2.
- Kynurenic acid (KYNA, 4-hydroxyquinoline-2-carboxylic acid)
KYNA is a naturally occurring product of the normal metabolism of amino acid L-tryptophan that has been reported to inhibit N-methyl-D-aspartate receptor (NMDAR) and nicotinic α7 receptors [67,96]. This compound, together with L-kynurenine, is thought to be an endogenous metabolite of L-tryptophan known to block NMDAR, and it has been frequently shown to exert neuroprotective or anticonvulsant properties in the brain [33,55,62,67,96,104,145]. This compound has been disclosed to inhibit NMDARs at the glycine-binding site and it can noncompetitively inhibit α7-nicotinic acetylcholine receptor, and through this action, it might modulate glutamate release presynaptically [62,96,97]. For instance, as administered systemically, a KYNA analog (SZR104) was shown to decrease population spike activity from the pyramidal layer of area CA1 of the hippocampus [33].
Previous studies have shown that the reduction in the astrocytic formation of KYNA could enhance glutamatergic tone in the hippocampus as well as cognitive abilities and synaptic plasticity [55,62,130]. KYNA is also recognized to be a target molecule in neuroendocrinology [130]. The KYNA derivatives have been increasing noticed to exert various biological actions [29]. Earlier work has also shown that KYNA-induced hypotension is strongly linked to the stimulation of IK(M) magnitude [111].
Of interest, recent studies have demonstrated that GH3-cell exposure to KYNA can result in the stimulation of IK(M) with an EC50 value of 18.1 μM. The EC50 value of KYNA-stimulated IK(M) appeared to be lower than that for its inhibition of NR1a/NR2A receptors or AMPA-evoked currents [95]. The relationship of IK(M)-conductance versus membrane potential during cell exposure to KYNA was noted to produce a leftward shift along the voltage axis by approximately 4 mV. Therefore, it is anticipated to be a pertinent link between KYNA effects on endocrine or neuroendocrine cells and the stimulatory effect on IK(M) magnitude.
In addition to the increased IK(M) amplitude, the presence of KYNA can shorten the activation time constant of the current. Stimulation of IK(M) caused by KYNA thus does not become instantaneous, yet it develops with time, once the KM channels are opened upon membrane depolarization, thereby leading to an increase in current activation. In keeping with these observations, single-channel current recordings were found to prolong mean open time of KM channels in the presence of KYNA. Therefore, the increase in both open-state probability and mean open time of KM channels produced by KYNA or its amide derivatives would be responsible for its increase of macroscopic IK(M) carried through these channels, despite their ineffectiveness in changing the single-channel amplitude. In this regard, KYNA or its structural similar compounds would be expected to be valuable tools for probing the structure and function of KM channels [99]
D. Hyperpolarization-Activated Cation Current (Ih)
The hyperpolarization-activated cation current (Ih), also known as the “funny current” (If), play a crucial role in regulating repetitive electrical activity in cardiac cells, various types of central neurons, and endocrine or neuroendocrince cells [50,116,151,155,156]. This type of ionic currents exhibits unique characteristics, including slow voltage-dependent activation kinetics and a mixed Na+/K+ current that flows inwardly, and it can be blocked by CsCl or ivabradine [35,58,116]. Activation of Ih may lead to depolarization of the resting potential, reaching the threshold required for generating or triggering an AP. Consequently, it influences pacemaker activity and impulse propagation [35,133].
Additionally, the inwardly directed Ih, triggered by sustained hyperpolarization and recognized for its gradual activation, can lead to persistent, activity-dependent adjustments in membrane excitability in diverse types of excitable cells [151,155,156,157]. Furthermore, the unique voltage-dependent hysteresis of Ih evoked by double triangular ramp pulse has been demonstrated [24,91,146]. The Ih is mediated by channels encoded by members of the hyperpolarization-activated cyclic nucleotide-gated (HCN) gene family, and earlier studies have demonstrated that the activity of these channels underlies the ionic mechanisms associated with both convulsive disorders and inflammatory pain disorders [26,31,100,149,156,157].
- 1. Carisbamate (CRS, RW1-333369, Vimpat®, (S)-2-Oxo-1-pyrrolidineacetamide)
CRS, a bioactive, orally administered neuromodulator, has been shown to be beneficial for the treatment of different types of convulsive disorders, including drug-resistant focal epilepsy and partial-onset seizure [36,88,107]. CRS is thought to work by affecting the activity of certain neurotransmitters in the brain, helping to reduce the occurrence of seizures [74]. This compound was also reported to be effective in the treatment of alcoholism [109]. Another study demonstrated that CRS prevented the development and production of epilepsy-like discharges and exerted a neuroprotective effect after epilepticus-like injury [37].
A recent study has produced an intriguing result, demonstrating that CRS can induce an inhibitory effect on Ih intrinsically in GH3 cells, and the extent of this effect varies with the applied concentration [57]. Using the modified Hill equation, the IC50 value required for CRS to suppress the Ih amplitude as seen in GH3 cells were estimated to be 38 μM. With the use of double triangular ramp pulse, the voltage-dependent hysteresis of Ih can be robustly activated. When the GH3 cells were continually exposed to CRS, the hysteretic strength of Ih evoked by the inverted triangular ramp pulse became progressively decreased [57]. The anticipated docking interaction between CRS and a model of the HCN channel also underscores CRS’s capacity to form hydrogen bonds and engage in hydrophobic interactions with amino acid residues in HCN channel [57]. Therefore, in addition to the inhibition of INa, CRS can interact directly with HCN channel to alter the magnitude, gating kinetics, and hysteretic strength of Ih present in pituitary cells. The extent to which CRS-mediated inhibition of Ih affects brain or endocrine function requires further investigation.
- 2
- Cannabidiol (CBD, 2-[(1R,6R)-3-methyl-6-prop-1-en-2-ylcyclohex-2-en-1-yl]-5-pentylbenzene-1,3-diol)
CBD is a non-psychoactive cannabinoid derived from the Cannabis plant, known for its potential therapeutic and medicinal properties. It is among over 100 cannabinoids present in the plant and has been demonstrated to be effective at treating various medical conditions, such as epilepsy, bipolar disorder, inflammation, and cancer [14,39]. Recent work has demonstrated that CBD can modify the activity in the hypothalamic-pituitary-adrenal axis [87,131].
A current study showed the effectiveness of CBD in suppressing the magnitude of Ih in GH3 cells and in increasing the activation time constant of the current [85]. The IC50 value for CBD-mediated inhibition of Ih was calculated to be 3.3 μM, and the decrease was reversed by oxaliplatin. Oxaliplatin, a platinum-based chemotherapeutic agent, was reported to stimulate Ih [19]. The quasi-steady-state activation curve of Ih was shifted in the leftward direction with no changes in the steepness of the curve. CBD also diminished the strength of voltage-dependent hysteresis on Ih elicited by double triangular ramp pulse. Findings from recent results suggest that CBD’s modification of Ih is independent of the binding to cannabinoid or opioid receptors and may exert a significant impact on the functional activities of electrically active cells occurring in vitro or in vivo.
E. Large-conductance Ca2+-activated K+ (BKCa) channel
The large-conductance Ca2+-activated K+ (BKCa or BK) channels (KCa1.1, KCNMA1, Slo1) belong to a family of voltage-gated K+ channels, and they are activated by an increase in either the intracellular concentration of Ca2+, the membrane potential, or both. The activation of BKCa channels can result in conducting large amounts of K+ ions across the cell membrane. Due to its high-conductance state and a single-channel conductance of about 150-250 pS, the BKCa channel is also thought to be a maxi- or large-K+ channel. This family of K+ channels is functionally expressed in pituitary cells, and its activity can affect the magnitude of whole-cell Ca2+-activated K+ currents (IK(Ca)) in pituitary cells, consequently impacting the membrane potential and stimulus-secretion coupling of these cells.
- 1. Rufinamide (RFM, Banzel®, Inovelon®, ethyl 1-(2,6-difluorophenyl)-1H-1,2,3-triazole-4-carboxylate)
RFM is recognized as a unique anticonvulsant drug, because, as a triazole derivative, its structure is dissimilar to other currently marked antiepileptic drugs [7,60]. It is increasingly being used in combination with other medications and therapies to treat Lennox-Gaustaut syndrome, severe epileptic encephalopathy, and other seizure disorders [10,102,112,115]. Lennox-Gaustaut syndrome is a rare and severe form of epilepsy that typically begins in childhood and it is characterized by multiple types of seizures and intellectual and developmental disabilities.
Although the mechanism of RFM action as an antiepileptic drug is still unclear, RFM was reported to modulate the activity of NaV channels by prolonging the inactive state of these channels [124,132]. Recent studies have also shown that RFM can interact with BKCa channels to enhance whole-cell Ca2+-activated K+ currents (IK(Ca)) effectively. As shown in Figure 5, the application of 10 μM RPM significantly enhances the amplitude of IK(Ca) across the entire voltage-clamp step. The effective EC50 value of RFM required for stimulating IK(Ca) was estimated to be 3.9 μM, with a Hill coefficient of 1.2 [73]. The maximum plasma concentrations of RFM at dosages of 10 mg/kg/day and 30 mg/kg/day have been reported as 4.01 μg/ml (16.8 μM] and 8.68 μg/ml (36.4 μM], respectively [136]. Consequently, the EC50 value is observed to be within the range of clinically achieved concentrations.
Additionally, the docking study showed that RFM can bind to the intracellular domain of KCa1.1 channel at certain amino-acid residues and that the RFM-induced docking site is not located in the pore regions of the channels [73]. Indeed, in pituitary GH3 cells, the addition of RFM to the cytosolic surface of the detached patch of membrane resulted in the enhanced activity of BKCa channels, with no modification in single-channel conductance of the channel [73]. The mean closed time of BKCa channels was decreased by the application of RFM to the cytosolic leaflet of the channel. Overall, aside from its ability to block INa, similar to riluzole as demonstrated previously [138,140], RFM has been shown to effectively enhance activity of BKCa channels within excitable cells in in vivo settings.
- 2 QO-40 ((5-(chloromethyl)-3-(naphthalen-1-yl)-2-(trifluoromethyl)pyrazolo [1,5-a]pyrimidin-7(4 H)-one)
QO-40 is a highly pure, synthetic, and biologically active compound. This compound has been previously reported to enhance KCNQ2/KCNQ3 heteromeric currents expressed in Xenopus oocytes [61]. QO58-lysine, a compound structurally similar to QO-40, can also activate neuronal KCNQ channels and exert antinociceptive effects on inflammatory pain [127]. The QO-58-induced amelioration of inflammatory pain observed in rodents was previously viewed as being accompanied by the activation of KCNQ-encoded K+ currents [127,152].
In a recent study [22], as pituitary GH3 cells were exposed to QO-40, the magnitude of IK(Ca) was observed to be notably increased with an EC50 value of 2.3 μM. QO-40-stimulated IK(Ca) was attenuated by further addition of paxilline, yet not by linopirdine or TRAM-34. It is worth noting that paxilline is a tremorgenic mycotoxin known to suppress the activity of BKCa channels [68], while linopirdine inhibits the IK(M) magnitude, and TRAM-34 can suppress the activity of intermediate-conductance Ca2+-activated K+ channels [147]. In inside-out single-channel recordings, it was observed that QO-40 not only produced a 14 mV shift towards a less positive potential in the steady-state activation curve of BKCa channels but also increased the gating charge by 1.4-fold. However, it is important to highlight that QO-40 did not alter the single-channel conductance of the channel, despite causing a reduction in the mean closed time of BKCa channels when it was present. Additionally, with the long-lasting isosceles-triangular ramp pulse, the presence of QO-40 enhanced the voltage-dependent hysteretic strength of BKCa channels. Although the detailed mechanism of the stimulatory actions of QO-40 on BKCa channels is not yet known, experimental observations suggest that QO-40 can enhance the activity of BKCa channels in a voltage-dependent manner [22]. As a result, its interaction with the BKCa channel can vary significantly based on factors such as the resting potential, AP firing pattern, the concentration of QO-40 used, or any combination of these variables.
The maximal concentration of QO-58, a synthesized compound that is structurally similar to QO-40, following oral administration at 25, 50, or 100 mg/kg, has been reported to reach 8.25, 16.29, or 18.27 mg/liter (approximately 18.6, 37, or 41 μM), respectively [84]. In this scenario, the stimulatory effect of QO-40 on BKCa channels would be of pharmacological or therapeutic relevance, as this compound at lower concentrations is effective at stimulating IK(Ca) and enhancing BKCa channel activity. However, it remains to be answered whether the rank order for QO-40 or other chemically related agents in activating BKCa channels would share a similar magnitude for their stimulation of neuronal KCNQ currents.
2. Conclusions
This paper does not focus voltage-independent currents, such as those mediated by transient receptor potential (TRP) channels, including TRPC, TRPM, and TRPV [65]. However, it provides a proof-of-concept for understanding the pathophysiological and pharmacological roles of pituitary cells, with a particular emphasis on ion channel functionality. This review also examines how specific drugs or compounds influence the intrinsic voltage-gated ionic currents within pituitary cells. These insights are crucial for regulating the function of electrically active cells and advancing our understanding of pituitary neuroendocrine tumors (PitNets). Notably, rats can also develop PitNets, or pituitary tumors, in their pituitary glands, much like humans and some other animals.
It is important to note that the current pituitary cell lines primarily used in research are derived from rats, such as GH3 cells, GH4C1 cells, MMQ, R1220, and AtT-20 cells [22,49,54,76,83,119,158]. Detailed descriptions of these cell lines can be found in Table 2. However, there are limited reports on immortalized human pituitary cell lines. Whether the electrical properties observed in these cell lines, as well as their responses to various drugs, are preserved in human pituitary cells remains an area that requires further in-depth investigation. In addition to pituitary cells, other endocrine cell types―such as those responsible for insulin and glucagon secretion, as well as Leydig cells―have also been shown to exhibit similar ionic currents [8,25,28,47,71]. However, further research is needed to determine whether the drugs or compounds mentioned here impact the electrical activity of these different endocrine cell types and their functional implications. Understanding the regulation of ion currents discussed in this paper is crucial for unraveling the molecular mechanisms underlying PiNets and their potential for advancing therapeutic approaches [23,159,160,161].
In clinical practice, it is common to observe that many patients with PitNets develop tumors in other endocrine glands, leading to an occurrence of an endocrine tumor syndrome known as multiple endocrine neoplasia type 1 (Wermer syndrome) [6,34,45,106,159,160]. Computerized tomography-guided radiofrequency ablation has become a key treatment option for removing PiNets removal, helping to minimize the risk of damage to surrounding deep brain tissues [162]. Additionally, the development of different medications aimed at preventing the recurrence of such tumors or other forms of multiple endocrine neoplasia will be an important topic for future research. At the same time, it is also an important task to conduct in-depth investigations of potential genetic abnormalities in patient samples of these PiNets [163]. Therefore, the comprehensive research presented in this paper will play a crucial role in advancing our comprehension of the origin and management of these conditions.
Although techniques like polymerase chain reaction (PCR) and Western blotting can be used to measure gene and protein expression abnormalities, respectively, in PitNets, performing patch-clamp experiments on different pituitary cells enables direct investigation of the biophysical properties of individual or whole ionic currents across the cell membrane, as well as changes in membrane potential, as presented herein. Therefore, it remains a crucial approach for functional studies. Additionally, performing direct measurements on PitNets’ organoid (165) will open up further opportunities for detailed pituitary research in the future.
Funding
The research described in this paper was supported by grants from the Nation Science and Technology Council (NSTC-111-2314-B-006-028, and NSTC-112-2923-B-006-001 to S-N Wu), Taiwan, and An Nan Hospital (ANHR-112-43 and ANHR-113-44), Taiwan. The funders of this research had no role in the study design, data collection, analyses, or interpretation of the results.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data were available by request.
Acknowledgments
This research is deeply grateful for the encouragement and support of the late Dr. Huai-hu Chuang (https://www.imb.sinica.edu.tw/ch/faculty/memoriam.html, accessed on 20 March 2025), a research professor at the Institute of Biomedical Sciences, Academia Sinica, Taiwan, and would like to express sincere thanks.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AP | action potential |
| BKCa channel | large-conductance Ca2+-activated K+ channel |
| HERG channel | human erg K+ channel |
| Ih | hyperpolarization-activated cationic current |
| IK(Ca) | Ca2+-activated K+ current |
| IK(erg) | erg-mediated K+ current |
| IK(M) | M-type K+ current |
| INaINa(P) | voltage-gated Na+ currentpersistent Na+ current |
| Kerg channel | erg-mediated KV channel |
| NaV channel | voltage-gated Na+ channel |
| PitNet | pituitary neuroendocrine tumor |
References
- Abbott, G.W. KCNQs: Ligand- and Voltage-Gated Potassium Channels. Front Physiol 2020, 11, 583. [Google Scholar] [CrossRef] [PubMed]
- Abrams, P.; Andersson, K.E. Muscarinic receptor antagonists for overactive bladder. BJU Int 2007, 100, 987–1006. [Google Scholar] [CrossRef]
- Anderson, D.; Yu, T.W.; Hinçal, F. Effect of some phthalate esters in human cells in the comet assay. Teratog Carcinog Mutagen 1999, 19, 275–280. [Google Scholar] [CrossRef]
- Aragay, A.M.; Katz, A.; Simon, M.I. The G alpha q and G alpha 11 proteins couple the thyrotropin-releasing hormone receptor to phospholipase C in GH3 rat pituitary cells. J Biol Chem 1992, 267, 24983–24988. [Google Scholar] [CrossRef] [PubMed]
- Arai, K.; Tsuruoka, H.; Homma, T. CS-3150, a novel non-steroidal mineralocorticoid receptor antagonist, prevents hypertension and cardiorenal injury in Dahl salt-sensitive hypertensive rats. Eur J Pharmacol 2015, 769, 266–273. [Google Scholar] [CrossRef]
- Armeni, E.; Grossman, A. The Spectrum of Familial Pituitary Neuroendocrine Tumors. Endocr Pathol 2023, 34, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, S. Rufinamide. Neurotherapeutics 2007, 4, 155–162. [Google Scholar] [CrossRef]
- Barg, S.; Galvanovskis, J.; Göpel, S.O.; Rorsman, P.; Eliasson, L. Tight coupling between electrical activity and exocytosis in mouse glucagon-secreting alpha-cells. Diabetes 2000, 49, 1500–1510. [Google Scholar] [CrossRef]
- Bauer, C.K.; Schwarz, J.R. Ether-à-go-go K+ channels: effective modulators of neuronal excitability. J Physiol 2018, 596, 769–783. [Google Scholar] [CrossRef]
- Besag, F.M.C.; Vasey, M.J.; Chin, R.F.M. Current and emerging pharmacotherapy for the treatment of Lennox-Gastaut syndrome. Expert Opin Pharmacother 2023, 24, 1249–1268. [Google Scholar] [CrossRef]
- Bhat, A.A.; Gupta, G.; Afzal, O.; Kazmi, I.; Al-Abbasi, F.A.; Alfawaz Altamimi, A.S.; Almalki, W.H.; Alzarea, S.I.; Singh, S.K.; Dua, K. Neuropharmacological effect of risperidone: From chemistry to medicine. Chem Biol Interact 2023, 369, 110296. [Google Scholar] [CrossRef] [PubMed]
- Black, B.J.; Atmaramani, R.; Pancrazio, J.J. Spontaneous and evoked activity from murine ventral horn cultures on microelectrode arrays. Front Cell Neurosci 2017, 11, 304. [Google Scholar] [CrossRef] [PubMed]
- Breier, A.F.; Malhotra, A.K.; Su, T.P.; Pinals, D.A.; Elman, I.; Adler, C.M.; Lafargue, R.T.; Clifton, A.; Pickar, D. Clozapine and risperidone in chronic schizophrenia: effects on symptoms, parkinsonian side effects, and neuroendocrine response. Am J Psychiatry 1999, 156, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Britch, S.C.; Babalonis, S.; Walsh, S.L. Cannabidiol: pharmacology and therapeutic targets. Psychopharmacology (Berl) 2021, 238, 9–28. [Google Scholar] [CrossRef]
- Brown, D.A.; Passmore, G.M. Neural KCNQ (Kv7) channels. Br J Pharmacol 2009, 156, 1185–1195. [Google Scholar] [CrossRef]
- Carboni, E.; Rolando, M.T.; Silvagni, A.; Di Chiara, G. Increase of dialysate dopamine in the bed nucleus of stria terminalis by clozapine and related neuroleptics. Neuropsychopharmacology 2000, 22, 140–147. [Google Scholar] [CrossRef]
- Catterall, W.A.; Lenaeus, M.J.; Gamal El-Din, T.M. Structure and pharmacology of voltage-gated sodium and calcium channels. Annu Rev Pharmacol Toxicol 2020, 60, 133–154. [Google Scholar] [CrossRef]
- Chan, H.Y.; Lin, W.W.; Lin, S.K.; Hwang, T.J.; Su, T.P.; Chiang, S.C.; Hwu, H.G. Efficacy and safety of aripiprazole in the acute treatment of schizophrenia in Chinese patients with risperidone as an active control: a randomized trial. J Clin Psychiatry 2007, 68, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Gao, Z.H.; Li, S.W.; Liu, P.Y.; Lo, Y.C.; Wu, S.N. Characterization in dual activation by oxaliplatin, a platinum-based chemotherapeutic agent of hyperpolarization-activated cation and electroporation-induced currents. Int J Mol Sci 2020, 21, 396. [Google Scholar] [CrossRef]
- Chang, W.T.; Wu, S.N. Activation of voltage-gated sodium current and inhibition of erg-mediated potassium current caused by telmisartan, an antagonist of angiotensin II type-1 receptor, in HL-1 atrial cardiomyocytes. Clin Exp Pharmacol Physiol 2018, 45, 797–807. [Google Scholar] [CrossRef]
- Chang, W.T.; Wu, S.N. Characterization of direct perturbations on voltage-gated sodium current by esaxerenone, a nonsteroidal mineralocorticoid receptor blocker. Biomedicines 2021, 9, 549. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Wu, S.N. Effective activation of BKCa channels by QO-40 (5-(chloromethyl)-3-(Naphthalen-1-yl)-2-(Trifluoromethyl)Pyrazolo [1,5-a]pyrimidin-7(4H)-one), known to be an opener of KCNQ2/Q3 channels. Pharmaceuticals (Basel) 2021, 14, 388. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.; Das, U.; Jabbar, S.; Gangisetty, O.; Rousseau, B.; Hanft, S.; Sarkar, D.K. Developmental pluripotency-associated 4 increases aggressiveness of pituitary neuroendocrine tumors by enhancing cell stemness. Neuro Oncol 2025, 27, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S.; So, E.C.; Wu, S.N. Modulating hyperpolarization-activated cation currents through small molecule perturbations: magnitude and gating Control. Biomedicines 2023, 11, 2177. [Google Scholar] [CrossRef]
- Chen, L.; Cho, H.Y.; Chuang, T.H.; Ke, T.L.; Wu, S.N. The effectiveness of isoplumbagin and plumbagin in regulating amplitude, gating kinetics, and voltage-dependent hysteresis of erg-mediated K+ currents. Biomedicines 2022, 10, 780. [Google Scholar] [CrossRef]
- Chen, Y.; Li, D.; Li, N.; Loh, P.; Guo, Y.; Hu, X.; Zhang, J.; Dou, B.; Wang, L.; Yang, C.; Guo, T.; Chen, S.; Liu, Z.; Chen, B.; Chen, Z. Role of nerve signal transduction and neuroimmune crosstalk in mediating the analgesic effects of acupuncture for neuropathic pain. Front Neurol 2023, 14, 1093849. [Google Scholar] [CrossRef]
- Cho, H.Y.; Chen, P.C.; Chuang, T.H.; Yu, M.C.; Wu, S.N. Activation of voltage-gated Na+ current by GV-58, a known activator of CaV channels. Biomedicines 2022, 10, 721. [Google Scholar] [CrossRef]
- Cho, H.Y.; Chuang, T.H.; Wu, S.N. Effective perturbations on the amplitude and hysteresis of erg-mediated potassium current caused by 1-octylnonyl 8-[(2-hydroxyethyl)[6-oxo-6(undecyloxy)hexyl]amino]-octanoate (SM-102), a cationic lipid. Biomedicines 2021, 9, 1367. [Google Scholar] [CrossRef]
- Chojnacki, C.; Gąsiorowska, A.; Popławski, T.; Konrad, P.; Chojnacki, M.; Fila, M.; Blasiak, J. Beneficial Effect of Increased Tryptophan Intake on Its Metabolism and Mental State of the Elderly. Nutrients 2023, 15, 847. [Google Scholar] [CrossRef]
- Chuang, T.H.; Cho, H.Y.; Wu, S.N. Effective Accentuation of Voltage-Gated Sodium Current Caused by Apocynin (4'-Hydroxy-3'-methoxyacetophenone), a Known NADPH-Oxidase Inhibitor. Biomedicines 2021, 9, 1146. [Google Scholar] [CrossRef]
- Crunelli V, David F, Morais TP, Lorincz ML. HCN channels and absence seizures. Neurobiol Dis 2023, 181, 106107. [Google Scholar] [CrossRef]
- De Alcubierre, D.; Ferrari, D.; Mauro, G.; Isidori, A.M.; Tomlinson, J.W.; Pofi, R. Glucocorticoids and cognitive function: a walkthrough in endogenous and exogenous alterations. J Endocrinol Invest 2023, 46, 1961–1982. [Google Scholar] [CrossRef] [PubMed]
- Demeter, I.; Nagy, K.; Gellért, L.; Vécsei, L.; Fülöp, F.; Toldi, J. A novel kynurenic acid analog (SZR104) inhibits pentylenetetrazole-induced epileptiform seizures. An electrophysiological study : special issue related to kynurenine. J Neural Transm (Vienna) 2012, 119, 151–154. [Google Scholar] [CrossRef]
- Deng, J.; Liao, X.; Cao, H. Neuroendocrine tumors in a patient with multiple endocrine neoplasia type 1 syndrome: A case report and review of the literature. Medicine (Baltimore) 2023, 102, e34350. [Google Scholar] [CrossRef]
- DiFrancesco, D. The role of the funny current in pacemaker activity. Circ Res 2010, 106, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Elkommos, S.; Mula, M. Current and future pharmacotherapy options for drug-resistant epilepsy. Expert Opin Pharmacother 2022, 23, 2023–2034. [Google Scholar] [CrossRef]
- Fernandes, M.J.; Carneiro, J.E.; Amorim, R.P.; Araujo, M.G.; Nehlig, A. Neuroprotective agents and modulation of temporal lobe epilepsy. Front Biosci (Elite Ed) 2015, 7, 79–93. [Google Scholar] [CrossRef]
- Fortunati, N.; Guaraldi, F.; Zunino, V.; Penner, F.; D'Angelo, V.; Zenga, F.; Pecori Giraldi, F.; Catalano, M.G.; Arvat, E. Effects of environmental pollutants on signaling pathways in rat pituitary GH3 adenoma cells. Environ Res 2017, 158, 660–668. [Google Scholar] [CrossRef]
- Fraguas-Sánchez, A.I.; Torres-Suárez, A.I. Medical use of cannabinoids. Drugs 2018, 78, 1665–1703. [Google Scholar] [CrossRef]
- Gandía, L.; Lara, B.; Imperial, J.S.; Villarroya, M.; Albillos, A.; Maroto, R.; García, A.G.; Olivera, B.M. Analogies and differences between omega-conotoxins MVIIC and MVIID: binding sites and functions in bovine chromaffin cells. Pflugers Arch 1997, 435, 55–64. [Google Scholar] [CrossRef]
- Gaudenzi, C.; Mifsud, K.R.; Reul, J.M.H.M. Insights into isoform-specific mineralocorticoid receptor action in the hippocampus. J Endocrinol 2023, 258, e220293. [Google Scholar] [CrossRef] [PubMed]
- Ghisari, M.; Bonefeld-Jorgensen, E.C. Effects of plasticizers and their mixtures on estrogen receptor and thyroid hormone functions. Toxicol Lett 2009, 189, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Ghovanloo, M.R.; Ruben, P.C. Cannabidiol and sodium channel pharmacology: General overview, mechanism, and clinical implications. Neuroscientist 2022, 28, 318–334. [Google Scholar] [CrossRef]
- Gillum, N.; Karabekian, Z.; Swift, L.M.; Brown, R.P.; Kay, M.W.; Sarvazyan, N. Clinically relevant concentrations of di (2-ethylhexyl) phthalate (DEHP) uncouple cardiac syncytium. Toxicol Appl Pharmacol 2009, 236, 25–38. [Google Scholar] [CrossRef]
- Halperin, R.; Tirosh, A. Progress report on multiple endocrine neoplasia type 1. Fam Cancer 2025, 24, 15. [Google Scholar] [CrossRef] [PubMed]
- Hamill, O.P.; Marty, A.; Neher, E.; Sakmann, B.; Sigworth, F.J. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch 1981, 391, 85–100. [Google Scholar] [CrossRef]
- Hamilton, A.; Zhang, Q.; Gao, R.; Hill, T.G.; Salehi, A.; Knudsen, J.G.; Draper, M.B.; Johnson, P.R.V.; Rorsman, P.; Tarasov, A.I. Nicotinic Signaling Stimulates Glucagon Secretion in Mouse and Human Pancreatic α-Cells. Diabetes 2025, 74, 53–64. [Google Scholar] [CrossRef]
- Harnod, T.; Yang, Y.C.; Chiu, L.T.; Wang, J.H.; Lin, S.Z.; Ding, D.C. Use of bladder antimuscarinics is associated with an increased risk of dementia: a retrospective population-based case-control study. Sci Rep 2021, 11, 4827. [Google Scholar] [CrossRef] [PubMed]
- Haug, T.M.; Hafting, T.; Sand, O. Inhibition of BK channels contributes to the second phase of the response to TRH in clonal rat anterior pituitary cells. Acta Physiol Scand 2004, 180, 347–357. [Google Scholar] [CrossRef]
- He, C.; Chen, F.; Li, B.; Hu, Z. Neurophysiology of HCN channels: from cellular functions to multiple regulations. Prog Neurobiol 2014, 112, 1–23. [Google Scholar] [CrossRef]
- He, F.Z.; McLeod, H.L.; Zhang, W. Current pharmacogenomic studies on hERG potassium channels. Trends Mol Med 2013, 19, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Hinkle PM, Tashjian AH Jr. Thyrotropin-releasing hormone regulates the number of its own receptors in the GH3 strain of pituitary cells in culture. Biochemistry 1975, 14, 3845–3851. [Google Scholar] [CrossRef]
- Hu, C.H.; Wu, S.N.; So, E.C. Tyrosine kinase inhibitors, ionic currents, and cardiac arrhythmia. Front Oncol 2023, 13, 1218821. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.H.; So, E.C.; Liu, Y.C.; Wu, S.N. Glucocorticoids stimulate the activity of large-conductance Ca2+-activated K+ channels in pituitary GH3 and AtT-20 cells via a non-genomic mechanism. Steroids 2006, 71, 129–140. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, M.; Chen, X.; Zhang, R.; Le, A.; Hong, M.; Zhang, Y.; Jia, L.; Zang, W.; Jiang, C.; Wang, J.; Fan, X.; Wang, J. Tryptophan metabolism in central nervous system diseases: Pathophysiology and potential therapeutic strategies. Aging Dis 2023, 14, 858–878. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.Y.; Wu, S.N.; Huang, C.W. The Modulation of Ubiquinone, a Lipid Antioxidant, on Neuronal Voltage-Gated Sodium Current. Nutrients 2022, 14, 3393. [Google Scholar] [CrossRef]
- Hung, T.Y.; Wu, S.N.; Huang, C.W. Concerted suppressive effects of carisbamate, an anti-epileptic alkyl-carbamate drug, on voltage-gated Na+ and hyperpolarization-activated cation currents. Front Cell Neurosci 2023, 17, 1159067. [Google Scholar] [CrossRef]
- Irisawa, H.; Brown, H.F.; Giles, W. Cardiac pacemaking in the sinoatrial node. Physiol Rev 1993, 73, 197–227. [Google Scholar] [CrossRef]
- Jaeger, R.J.; Rubin, R.J. Migration of a phthalate ester plasticizer from polyvinyl chloride blood bags into stored human blood and its localization in human tissues. N Engl J Med 1972, 287, 1114–1118. [Google Scholar] [CrossRef]
- Jeong, H.J.; Min, S.; Baek, J.; Kim, J.; Chung, J.; Jeong, K. Real-time reaction monitoring of azide-alkyne cycloadditions using benchtop NMR-based signal amplification by reversible exchange (SABRE). ACS Meas Sci Au 2023, 3, 134–142. [Google Scholar] [CrossRef]
- Jia, C.; Qi, J.; Zhang, F.; Mi, Y.; Zhang, X.; Chen, X.; Liu, L.; Du, X.; Zhang, H. Activation of KCNQ2/3 potassium channels by novel pyrazolo[1,5-a]pyrimidin-7(4H)-one derivatives. Pharmacology 2011, 87, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Jorratt, P.; Ricny, J.; Leibold, C.; Ovsepian, S.V. Endogenous modulators of NMDA receptor control dendritic field expansion of cortical neurons. Mol Neurobiol 2023, 60, 1440–1452. [Google Scholar] [CrossRef]
- Jurewicz, J.; Hanke, W. Exposure to phthalates: reproductive outcome and children health. A review of epidemiological studies. Int J Occup Med Environ Health 2011, 24, 115–141. [Google Scholar] [CrossRef] [PubMed]
- Kario, K.; Nishizawa, M.; Kato, M.; Ishii, H.; Uchiyama, K.; Nagai, M.; Takahashi, N.; Asakura, T.; Shiraiwa, T.; Yoshida, T.; Kaneshiro, M.; Taguchi, T.; Shiosakai, K.; Sugimoto, K. Nighttime home blood pressure lowering effect of esaxerenone in patients with uncontrolled nocturnal hypertension: the EARLY-NH study. Hypertens Res 2023, 46, 1782–1794. [Google Scholar] [CrossRef]
- Kelly, M.J.; Wagner, E.J. Canonical transient receptor potential channels and hypothalamic control of homeostatic functions. J Neuroendocrinol. 2024, 36, e13392. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Ishizaka, M.; Kazusaka, A.; Fujita, S. Di-(2-ethylhexyl) phthalate suppresses tamoxifen-induced apoptosis in GH3 pituitary cells. Arch Toxicol 2007, 81, 27–33. [Google Scholar] [CrossRef]
- Klaessens, S.; Stroobant, V.; De Plaen, E.; Van den Eynde, B.J. Systemic tryptophan homeostasis. Front Mol Biosci 2022, 9, 897929. [Google Scholar] [CrossRef]
- Knaus HG, McManus OB, Lee SH, Schmalhofer WA, Garcia-Calvo M, Helms LM, Sanchez M, Giangiacomo K, Reuben JP, Smith AB 3rd, et al. Tremorgenic indole alkaloids potently inhibit smooth muscle high-conductance calcium-activated potassium channels. Biochemistry 1994, 33, 5819–5828. [Google Scholar] [CrossRef]
- Kreder, K.J. Solifenacin. Urol Clin North Am 2006, 33, 483–490. [Google Scholar] [CrossRef]
- Krithivasan, R.; Miller, G.Z.; Belliveau, M.; Gearhart, J.; Krishnamoorthi, V.; Lee, S.; Kannan, K. Analysis of ortho-phthalates and other plasticizers in select organic and conventional foods in the United States. J Expo Sci Environ Epidemiol 2023, 33, 778–786. [Google Scholar] [CrossRef]
- Kuo, P.C.; Yang, C.J.; Lee, Y.C.; Chen, P.C.; Liu, Y.C.; Wu, S.N. The comprehensive electrophysiological study of curcuminoids on delayed-rectifier K+ currents in insulin-secreting cells. Eur J Pharmacol 2018, 819, 233–241. [Google Scholar] [CrossRef]
- Kushmerick, C.; Romano-Silva, M.A.; Gomez, M.V.; Prado, M.A. Muscarinic regulation of Ca2+ oscillation frequency in GH3 cells. Brain Res 1999, 851, 39–45. [Google Scholar] [CrossRef]
- Lai, M.C.; Wu, S.N.; Huang, C.W. Rufinamide, a triazole-derived antiepileptic drug, stimulates Ca2+-activated K+ currents while inhibiting voltage-gated Na+ currents. Int J Mol Sci 2022, 23, 13677. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Lee, M.L.; Shih, C.C.; Liou, H.H. Carisbamate (RWJ-333369) inhibits glutamate transmission in the granule cell of the dentate gyrus. Neuropharmacology 2011, 61, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Choi, J.S.; Choi, B.H.; Hahn, S.J. Inhibition of cloned hERG potassium channels by risperidone and paliperidone. Naunyn Schmiedebergs Arch Pharmacol 2017, 390, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Leng, L.; Zhang, Y. Effects of an estrogen receptor antagonist on proliferation, prolactin secretion and growth factor expression in the MMQ pituitary prolactinoma cell line. J Clin Neurosci 2011, 18, 1694–1698. [Google Scholar] [CrossRef]
- Lerma, E.; White, W.B.; Bakris, G. Effectiveness of nonsteroidal mineralocorticoid receptor antagonists in patients with diabetic kidney disease. Postgrad Med 2023, 135, 224–233. [Google Scholar] [CrossRef]
- Leucht, S.; Schneider-Thoma, J.; Burschinski, A.; Peter, N.; Wang, D.; Dong, S.; Huhn, M.; Nikolakopoulou, A.; Salanti, G.; Davis, J.M. Long-term efficacy of antipsychotic drugs in initially acutely ill adults with schizophrenia: systematic review and network meta-analysis. World Psychiatry 2023, 22, 315–324. [Google Scholar] [CrossRef]
- Li, S.; Dai, J.; Zhang, L.; Zhang, J.; Zhang, Z.; Chen, B. An association of elevated serum prolactin with phthalate exposure in adult men. Biomed Environ Sci 2011, 24, 31–39. [Google Scholar] [CrossRef]
- Li, Y.; Patel, M.; Baroudi, J.; Wu, M.; Gatti, S.; Liang, M.; Wipf, P.; Badawi, Y.; Meriney, S.D. A cross-sectional study of ageing at the mouse neuromuscular junction and effects of an experimental therapeutic approach for dynapenia. J Physiol 2023, 601, 4135–4150. [Google Scholar] [CrossRef]
- Liang, M.; Tarr, T.B.; Bravo-Altamirano, K.; Valdomir, G.; Rensch, G.; Swanson, L.; DeStefino, N.R.; Mazzarisi, C.M.; Olszewski, R.A.; Wilson, G.M.; Meriney, S.D.; Wipf, P. Synthesis and biological evaluation of a selective N- and p/q-type calcium channel agonist. ACS Med Chem Lett 2012, 3, 985–990. [Google Scholar] [CrossRef]
- Lin, D.S.; Lin, F.J.; Lin, Y.S.; Lee, J.K.; Lin, Y.H. The effects of mineralocorticoid receptor antagonists on cardiovascular outcomes in patients with end-stage renal disease and heart failure. Eur J Heart Fail 2023, 25, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.H.; Lin, J.F.; Yu, M.C.; Wu, S.N.; Wu, C.L.; Cho, H.Y. Characterization in potent modulation on voltage-gated Na+ current exerted by deltamethrin, a pyrethroid insecticide. Int J Mol Sci 2022, 23, 14733. [Google Scholar] [CrossRef]
- Liu, C.; Jinlong, Q.I.; Zhang, H.; Jia, Q. Pharmacokinetic study of QO-58, A new potassium channel opener. Chin Pharmacol Bull 2014, 30, 574–577. [Google Scholar]
- Liu, Y.C.; So, E.C.; Wu, S.N. Cannabidiol modulates M-type K+ and hyperpolarization-activated cation currents. Biomedicines 2023, 11, 2651. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.K.; Wu, S.N.; Lee, C.T.; Li, H.F.; Chiang, H.T. Characterization of action potential waveform-evoked L-type calcium currents in pituitary GH3 cells. Pflugers Arch 2001, 442, 547–557. [Google Scholar] [CrossRef]
- Lopresti, A.L.; Smith, S.J.; Drummond, P.D. Modulation of the hypothalamic-pituitary-adrenal (HPA) axis by plants and phytonutrients: a systematic review of human trials. Nutr Neurosci 2022, 25, 1704–1730. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zheng, J.; Cao, Y.; Bresnahan, R.; Martin-McGill, K.J. Carisbamate add-on therapy for drug-resistant focal epilepsy. Cochrane Database Syst Rev 2021, 12, CD012121. [Google Scholar] [CrossRef]
- Majdinasab, N.; Orakifar, N.; Kouti, L.; Shamsaei, G.; Seyedtabib, M.; Jafari, M. Solifenacin versus posterior tibial nerve stimulation for overactive bladder in patients with multiple sclerosis. Front Neurosci 2023, 17, 1107886. [Google Scholar] [CrossRef]
- Maljevic, S.; Wuttke, T.V.; Lerche, H. Nervous system KV7 disorders: breakdown of a subthreshold brake. J Physiol 2008, 586, 1791–1801. [Google Scholar] [CrossRef]
- Männikkö, R.; Pandey, S.; Larsson, H.P.; Elinder, F. Hysteresis in the voltage dependence of HCN channels: conversion between two modes affects pacemaker properties. J Gen Physiol 2005, 125, 305–326. [Google Scholar] [CrossRef]
- Maria, S.A.; Kumar, A.; Wilfred, P.M.; Shanthi, M.; Peedicayil, J. Inhibition of contractility of isolated caprine detrusor by the calcium channel blocker cilnidipine and reversal by calcium channel openers. Curr Ther Res Clin Exp 2023, 99, 100717. [Google Scholar] [CrossRef]
- Martínez, A.; García-Gutiérrez, P.; Zubillaga, R.A.; Garza, J.; Vargas, R. Main interactions of dopamine and risperidone with the dopamine D2 receptor. Phys Chem Chem Phys 2021, 23, 14224–14230. [Google Scholar] [CrossRef]
- Masutomi, N.; Shibutani, M.; Takagi, H.; Uneyama, C.; Lee, K.Y.; Hirose, M. Alteration of pituitary hormone-immunoreactive cell populations in rat offspring after maternal dietary exposure to endocrine-active chemicals. Arch Toxicol 2004, 78, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Mok, M.H.; Fricker, A.C.; Weil, A.; Kew, J.N. Electrophysiological characterisation of the actions of kynurenic acid at ligand-gated ion channels. Neuropharmacology 2009, 57, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Moroni, F.; Cozzi, A.; Sili, M.; Mannaioni, G. Kynurenic acid: a metabolite with multiple actions and multiple targets in brain and periphery. J Neural Transm (Vienna) 2012, 119, 133–139. [Google Scholar] [CrossRef]
- Mourão, A.A.; de Mello, A.B.S.; Dos Santos Moreira, M.C.; Rodrigues, K.L.; Lopes, P.R.; Xavier, C.H.; Gomes, R.M.; Freiria-Oliveira, A.H.; Blanch, G.T.; Colombari, E.; Pedrino, G.R. Median preoptic nucleus excitatory neurotransmitters in the maintenance of hypertensive state. Brain Res Bull 2018, 142, 207–215. [Google Scholar] [CrossRef]
- Munkhjargal, U.; Fukuda, D.; Ganbaatar, B.; Suto, K.; Matsuura, T.; Ise, T.; Kusunose, K.; Yamaguchi, K.; Yagi, S.; Yamada, H.; Soeki, T.; Wakatsuki, T.; Sata, M. A selective mineralocorticoid receptor blocker, esaxerenone, attenuates vascular dysfunction in diabetic C57BL/6 mice. J Atheroscler Thromb 2023, 30, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Nagy, K.; Plangár, I.; Tuka, B.; Gellért, L.; Varga, D.; Demeter, I.; Farkas, T.; Kis, Z.; Marosi, M.; Zádori, D.; Klivényi, P.; Fülöp, F.; Szatmári, I.; Vécsei, L.; Toldi, J. Synthesis and biological effects of some kynurenic acid analogs. Bioorg Med Chem 2011, 19, 7590–7596. [Google Scholar] [CrossRef]
- Ohashi, N.; Uta, D.; Ohashi, M.; Baba, H. Analgesic effect of ivabradine against inflammatory pain mediated by hyperpolarization-activated cyclic nucleotide-gated cation channels expressed on primary afferent terminals in the spinal dorsal horn. Pain 2022, 163, 1356–1369. [Google Scholar] [CrossRef]
- Ojala, K.S.; Kaufhold, C.J.; Davey, M.R.; Yang, D.; Liang, M.; Wipf, P.; Badawi, Y.; Meriney, S.D. Potentiation of neuromuscular transmission by a small molecule calcium channel gating modifier improves motor function in a severe spinal muscular atrophy mouse model. Hum Mol Genet 2023, 32, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, M.; Prabhakar, H.; Marson, A.G. Rufinamide add-on therapy for refractory epilepsy. Cochrane Database Syst Rev 2018, 4, CD011772. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.; Robinson, D.; Cardozo, L. Darifenacin in the treatment of overactive bladder. Int J Clin Pract 2005, 59, 831–838. [Google Scholar] [CrossRef]
- Paul, E.R.; Schwieler, L.; Erhardt, S.; Boda, S.; Trepci, A.; Kämpe, R.; Asratian, A.; Holm, L.; Yngve, A.; Dantzer, R.; Heilig, M.; Hamilton, J.P.; Samuelsson, M. Peripheral and central kynurenine pathway abnormalities in major depression. Brain Behav Immun 2022, 101, 136–145. [Google Scholar] [CrossRef]
- Pearce, P.T.; McNally, M.; Funder, J.W. Nuclear localization of type 1 aldosterone binding sites in steroid-unexposed GH3 cells. Clin Exp Pharmacol Physiol 1986, 13, 647–654. [Google Scholar] [CrossRef]
- Pieterman, C.R.C.; Valk, G.D. Update on the clinical management of multiple endocrine neoplasia type 1. Clin Endocrinol (Oxf) 2022, 97, 409–423. [Google Scholar] [CrossRef] [PubMed]
- Pong, A.W.; Ross, J.; Tyrlikova, I.; Giermek, A.J.; Kohli, M.P.; Khan, Y.A.; Salgado, R.D.; Klein, P. Epilepsy: expert opinion on emerging drugs in phase 2/3 clinical trials. Expert Opin Emerg Drugs 2022, 27, 75–90. [Google Scholar] [CrossRef]
- Raman, G.; Tunnicliffe, D.; Lai, E.; Bennett, T.; Caldwell, P. Safety and tolerability of solifenacin in children and adolescents with overactive bladder- a systematic review. J Pediatr Urol 2023, 19, 19.e1–19.e13. [Google Scholar] [CrossRef]
- Rezvani, A.H.; Lawrence, A.J.; Arolfo, M.P.; Levin, E.D.; Overstreet, D.H. Novel medication targets for the treatment of alcoholism: preclinical studies. Recent Pat CNS Drug Discov 2012, 7, 151–162. [Google Scholar] [CrossRef]
- Rodrigues, T.C.M.L.; de Moura, J.P.; Dos Santos, A.M.F.; Monteiro, A.F.M.; Lopes, S.M.; Scotti, M.T.; Scotti, L. Epileptic targets and drugs: A mini-review. Curr Drug Targets 2023, 24, 212–224. [Google Scholar] [CrossRef]
- Sakakibara, K.; Feng, G.G.; Li, J.; Akahori, T.; Yasuda, Y.; Nakamura, E.; Hatakeyama, N.; Fujiwara, Y.; Kinoshita, H. Kynurenine causes vasodilation and hypotension induced by activation of KCNQ-encoded voltage-dependent K+ channels. J Pharmacol Sci. 2015, 129, 31–37. [Google Scholar] [CrossRef]
- Sankar, R.; Chez, M.; Pina-Garza, J.E.; Dixon-Salazar, T.; Flamini, J.R.; Hyslop, A.; McGoldrick, P.; Millichap, J.J.; Resnick, T.; Rho, J.M.; Wolf, S. Proposed anti-seizure medication combinations with rufinamide in the treatment of Lennox-Gastaut syndrome: Narrative review and expert opinion. Seizure 2023, 110, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, S.; Simasko, S.M. Characterization of an M-like current modulated by thyrotropin-releasing hormone in normal rat lactotrophs. J Neurosci 1996, 16, 1668–1678. [Google Scholar] [CrossRef]
- Secondo, A.; De Mizio, M.; Zirpoli, L.; Santillo, M.; Mondola, P. The Cu-Zn superoxide dismutase (SOD1) inhibits ERK phosphorylation by muscarinic receptor modulation in rat pituitary GH3 cells. Biochem Biophys Res Commun 2008, 376, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Sills, G.J. Pharmacological diversity amongst approved and emerging antiseizure medications for the treatment of developmental and epileptic encephalopathies. Ther Adv Neurol Disord 2023, 16, 17562864231191000. [Google Scholar] [CrossRef] [PubMed]
- Simasko, S.M.; Sankaranarayanan, S. Characterization of a hyperpolarization-activated cation current in rat pituitary cells. Am J Physiol 1997, 272, E405–14. [Google Scholar] [CrossRef]
- Singh, M.; Sapkota, K.; Sakimura, K.; Kano, M.; Cowell, R.M.; Overstreet-Wadiche, L.; Hablitz, J.J.; Nakazawa, K. Maturation of GABAergic synaptic transmission from neocortical parvalbumin interneurons involves N-methyl-D-aspartate receptor recruitment of Cav2.1 channels. Neuroscience 2023, 513, 38–53. [Google Scholar] [CrossRef]
- Sinvani, L.; Afroz-Hossain, A.; Muran, A.; Strunk, A.; Williams, M.S.; Qiu, M.; Zeltser, R.; Makaryus, A.N.; Wolf-Klein, G.; Pekmezaris, R. Electrocardiogram monitoring practices for hospitalized adults receiving antipsychotics: A retrospective cohort study. J Psychiatr Pract 2022, 28, 108–116. [Google Scholar] [CrossRef]
- So, E.C.; Foo, N.P.; Ko, S.Y.; Wu, S.N. Bisoprolol, known to be a selective β₁-receptor antagonist, differentially but directly suppresses IK(M) and IK(erg) in pituitary cells and hippocampal neurons. Int J Mol Sci 2019, 20, 657. [Google Scholar] [CrossRef]
- So, E.C.; Wu, S.N.; Lo, Y.C.; Su, K. Differential regulation of tefluthrin and telmisartan on the gating charges of INa activation and inactivation as well as on resurgent and persistent INa in a pituitary cell line (GH3). Toxicol Lett 2018, 285, 104–112. [Google Scholar] [CrossRef]
- Stojilkovic, S.S. Pituitary cell type-specific electrical activity, calcium signaling and secretion. Biol Res 2006, 39, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Stojilkovic SS, Tabak J, Bertram. Ion channels and signaling in the pituitary gland. Endoc Rev 2010, 31, 845–915. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, M.; Radmanovic, B.; Jovanovic, M.; Janjic, V.; Muric, N.; Ristic, D.I. Risperidone induced hyperprolactinemia: From basic to clinical studies. Front Psychiatry 2022, 13, 874705. [Google Scholar] [CrossRef]
- Suter, M.R.; Kirschmann, G.; Laedermann, C.J.; Abriel, H.; Decosterd, I. Rufinamide attenuates mechanical allodynia in a model of neuropathic pain in the mouse and stabilizes voltage-gated sodium channel inactivated state. Anesthesiology 2013, 118, 160–172. [Google Scholar] [CrossRef]
- Tarr, T.B.; Lacomis, D.; Reddel, S.W.; Liang, M.; Valdomir, G.; Frasso, M.; Wipf, P.; Meriney, S.D. Complete reversal of Lambert-Eaton myasthenic syndrome synaptic impairment by the combined use of a K+ channel blocker and a Ca2+ channel agonist. J Physiol 2014, 592, 3687–3696. [Google Scholar] [CrossRef]
- Tarr, T.B.; Valdomir, G.; Liang, M.; Wipf, P.; Meriney, S.D. New calcium channel agonists as potential therapeutics in Lambert-Eaton myasthenic syndrome and other neuromuscular diseases. Ann N Y Acad Sci 2012, 1275, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Teng, B.C.; Song, Y.; Zhang, F.; Ma, T.Y.; Qi, J.L.; Zhang, H.L.; Li, G.; Wang, K. Activation of neuronal Kv7/KCNQ/M-channels by the opener QO58-lysine and its anti-nociceptive effects on inflammatory pain in rodents. Acta Pharmacol Sin 2016, 37, 1054–1062. [Google Scholar] [CrossRef]
- Ullah, S.; Ahmad, S.; Guo, X.; Ullah, S.; Ullah, S.; Nabi, G.; Wanghe, K. A review of the endocrine disrupting effects of micro and nano plastic and their associated chemicals in mammals. Front Endocrinol (Lausanne) 2023, 13, 1084236. [Google Scholar] [CrossRef]
- Vandenberg JI, Perry MD, Perrin MJ, Mann SA, Ke Y, Hill AP. HERG K+ channels: structure, function, and clinical significance. Physiol Rev 2012, 92, 1393–1478. [Google Scholar] [CrossRef]
- Vécsei, L.; Horváth, Z.; Tuka, B. Old and new neuroendocrine molecules: somatostatin, cyseamine, pantethine and kynurenine. Ideggyogy Sz 2014, 67, 107–112. [Google Scholar]
- Viudez-Martínez, A.; García-Gutiérrez, M.S.; Manzanares, J. Cannabidiol regulates the expression of hypothalamus-pituitary-adrenal axis-related genes in response to acute restraint stress. J Psychopharmacol 2018, 32, 1379–1384. [Google Scholar] [CrossRef]
- Vohora, D.; Saraogi, P.; Yazdani, M.A.; Bhowmik, M.; Khanam, R.; Pillai, K.K. Recent advances in adjunctive therapy for epilepsy: focus on sodium channel blockers as third-generation antiepileptic drugs. Drugs Today (Barc) 2010, 46, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Wahl-Schott C, Fenske S, Biel M. HCN channels: new roles in sinoatrial node function. Curr Opin Pharmacol 2014, 15, 83–90. [Google Scholar] [CrossRef]
- Weiss, B. Endocrine disruptors as a threat to neurological function. J Neurol Sci 2011, 305, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Welk, B.; McClure, J.A. The impact of anticholinergic use for overactive bladder on cognitive changes in adults with normal cognition, mild cognitive impairment, or dementia. Eur Urol Open Sci 2022, 46, 22–29. [Google Scholar] [CrossRef]
- Wier, H.A.; Cerna, A.; So, T.Y. Rufinamide for pediatric patients with Lennox-Gastaut syndrome: a comprehensive overview. Paediatr Drugs 2011, 13, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; White, H.V.; Boehm, B.A.; Meriney, C.J.; Kerrigan, K.; Frasso, M.; Liang, M.; Gotway, E.M.; Wilcox, M.R.; Johnson, J.W.; Wipf, P.; Meriney, S.D. New Cav2 calcium channel gating modifiers with agonist activity and therapeutic potential to treat neuromuscular disease. Neuropharmacology 2018, 131, 176–189. [Google Scholar] [CrossRef]
- Wu, S.N.; Chiang, H.T.; Shen, A.Y.; Lo, Y.K. Differential effects of quercetin, a natural polyphenolic flavonoid, on L-type calcium current in pituitary tumor (GH3) cells and neuronal NG108-15 cells. J Cell Physiol 2003, 195, 298–308. [Google Scholar] [CrossRef]
- Wu, S.N.; Jan, C.R.; Li, H.F.; Chiang, H.T. Characterization of inhibition by risperidone of the inwardly rectifying K+ current in pituitary GH3 cells. Neuropsychopharmacology 2000, 23, 676–689. [Google Scholar] [CrossRef]
- Wu, S.N.; Li, H.F. Characterization of riluzole-induced stimulation of large-conductance calcium-activated potassium channels in rat pituitary GH3 cells. J Investig Med 1999, 47, 484–495. [Google Scholar]
- Wu, S.N.; Lo, Y.K.; Li, H.F.; Shen, A.Y. Functional coupling of voltage-dependent L-type Ca2+ current to Ca2+-activated K+ current in pituitary GH3 cells. Chin J Physiol 2001, 44, 161–167. [Google Scholar]
- Wu, S.N.; Yang, W.H.; Yeh, C.C.; Huang, H.C. The inhibition by di(2-ethylhexyl)-phthalate of erg-mediated K⁺ current in pituitary tumor (GH₃) cells. Arch Toxicol 2012, 86, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Yu, M.C. Inhibition of Voltage-Gated Na+ Currents Exerted by KB-R7943 (2-[2-[4-(4-nitrobenzyloxy)phenyl]ethyl]isothiourea), an Inhibitor of Na+-Ca2+ Exchanging Process. Int J Mol Sci 2023, 24, 1805. [Google Scholar] [CrossRef] [PubMed]
- Wulfsen, I.; Hauber, H.P.; Schiemann, D.; Bauer, C.K.; Schwarz, J.R. Expression of mRNA for voltage-dependent and inward-rectifying K channels in GH3/B6 cells and rat pituitary. J Neuroendocrinol 2000, 12, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Wyant GA, Yu W, Doulamis IP, Nomoto RS, Saeed MY, Duignan T, McCully JD, Kaelin WG Jr. Mitochondrial remodeling and ischemic protection by G protein-coupled receptor 35 agonists. Science 2022, 377, 621–629. [Google Scholar] [CrossRef]
- Xiao, Y.F.; Chandler, N.; Dobrzynski, H.; Richardson, E.S.; Tenbroek, E.M.; Wilhelm, J.J.; Sharma, V.; Varghese, A.; Boyett, M.R.; Iaizzo, P.A.; Sigg, D.C. Hysteresis in human HCN4 channels: a crucial feature potentially affecting sinoatrial node pacemaking. Sheng Li Xue Bao 2010, 62, 1–13. [Google Scholar]
- Yeh, P.S.; Wu, S.J.; Hung, T.Y.; Huang, Y.M.; Hsu, C.W.; Sze, C.I.; Hsieh, Y.J.; Huang, C.W.; Wu, S.N. Evidence for the Inhibition by temozolomide, an imidazotetrazine family alkylator, of intermediate-conductance Ca2+-activated K+ channels in glioma cells. Cell Physiol Biochem 2016, 38, 1727–1742. [Google Scholar] [CrossRef]
- Yoshida, Y.; Fujiwara, M.; Kinoshita, M.; Sada, K.; Miyamoto, S.; Ozeki, Y.; Iwamoto, M.; Mori, Y.; Nagai, S.; Matsuda, N.; Noguchi, T.; Okamoto, M.; Gotoh, K.; Masaki, T.; Shibata, H. Effects of esaxerenone on blood pressure, urinary albumin excretion, serum levels of NT-proBNP, and quality of life in patients with primary aldosteronism. Hypertens Res 2024, 47, 157–167. [Google Scholar] [CrossRef]
- Yu, C.; Deng, X.J.; Xu, D. Gene mutations in comorbidity of epilepsy and arrhythmia. J Neurol 2023, 270, 1229–1248. [Google Scholar] [CrossRef]
- Yue C, Yaari Y. KCNQ/M channels control spike afterdepolarization and burst generation in hippocampal neurons. J Neurosci 2004, 24, 4614–4624. [Google Scholar] [CrossRef]
- Zaborska, K.E.; Jordan, K.L.; Thorson, A.S.; Dadi, P.K.; Schaub, C.M.; Nakhe, A.Y.; Dickerson, M.T.; Lynch, J.C.; Weiss, A.J.; Dobson, J.R.; Jacobson, D.A. Liraglutide increases islet Ca2+ oscillation frequency and insulin secretion by activating hyperpolarization-activated cyclic nucleotide-gated channels. Diabetes Obes Metab 2022, 24, 1741–1752. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Mi, Y.; Qi, J.L.; Li, J.W.; Si, M.; Guan, B.C.; Du, X.N.; An, H.L.; Zhang, H.L. Modulation of K(v)7 potassium channels by a novel opener pyrazolo[1,5-a]pyrimidin-7(4H)-one compound QO-58. Br J Pharmacol. 2013, 168, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Zhang Y, Li D, Darwish Y, Fu X, Trussell LO, Huang H. KCNQ channels enable reliable presynaptic spiking and synaptic transmission at high frequency. J Neurosci 2022, 42, 3305–3315. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Y.; Tao, Y.; Guo, X.; Cui, Y.; Li, Z. Phthalates (PAEs) and reproductive toxicity: Hypothalamic-pituitary-gonadal (HPG) axis aspects. J Hazard Mater 2023, 459, 132182. [Google Scholar] [CrossRef]
- Zhou, X.; Li, A.; Mi, X.; Li, Y.; Ding, Z.; An, M.; Chen, Y.; Li, W.; Tao, X.; Chen, X.; Li, Y. Hyperexcited limbic neurons represent sexual satiety and reduce mating motivation. Science 2023, 379, 820–825. [Google Scholar] [CrossRef]
- Mishra, P.; Narayanan, R. The enigmatic HCN channels: A cellular neurophysiology perspective. Proteins. 2025, 93, 72–92. [Google Scholar] [CrossRef]
- Chen, L.; Chehade, H.D.; Chu, H.Y. Motor cortical neuronal hyperexcitability associated with α-synuclein aggregation. NPJ Parkinsons Dis 2025, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Cui, W.; Zhu, D.; Gao, N.; Zhu, Y. Common tools for pituitary adenomas research: cell lines and primary cells. Pituitary 2020, 23, 182–188. [Google Scholar] [CrossRef]
- Villa, C.; Birtolo, M.F.; Perez-Rivas, L.G.; Righi, A.; Assie, G.; Baussart, B.; Asioli, S. Grading and staging for pituitary neuroendocrine tumors. Brain Pathol 2025, 35, e13299. [Google Scholar] [CrossRef]
- Cui, R.; Duan, H.; Hu, W.; Li, C.; Zhong, S.; Liang, L.; Chen, S.; Hu, H.; He, Z.; Wang, Z.; Guo, X.; Chen, Z.; Xu, C.; Zhu, Y.; Chen, Y.; Sai, K.; Yang, Q.; Guo, C.; Mou, Y.; Jiang, X. Establishment of Human Pituitary Neuroendocrine Tumor Derived Organoid and Its Pilot Application for Drug Screening. J Clin Endocrinol Metab 2025, 110, e827–e840. [Google Scholar] [CrossRef]
- Calvanese, F.; Jannelli, G.; Feuvret, L.; Vasiljevic, A.; Manet, R.; Sergeant, C.; Raverot, G.; Jouanneau, E. Aggressive Pituitary Tumors and Pituitary Carcinomas: definition, management and overview for clinical practice. Neuro-Oncology Advances 2025, vdae114. [Google Scholar] [CrossRef]
- Perrier, N.D. From Initial Description by Wermer to Present-Day MEN1, What have We Learned? World J Surg 2018, 42, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Gedvilaite-Vaicechauskiene, G.; Kriauciuniene, L.; Tamasauskas, A.; Rovite, V.; Mandrika, I.; Wu, S.N.; Huang, C.W.; Poskiene, L.; Liutkeviciene, R. Pituitary Adenoma: SSTR2 rs2236750, SSTR5 rs34037914, and AIP rs267606574 Genetic Variants, Serum Levels, and Ki-67 Labeling Index Associations. Medicina (Kaunas) 2024, 60, 1252. [Google Scholar] [CrossRef]
- Cui, R.; Duan, H.; Hu, W.; Li, C.; Zhong, S.; Liang, L.; Chen, S.; Hu, H.; He, Z.; Wang, Z.; Guo, X.; Chen, Z.; Xu, C.; Zhu, Y.; Chen, Y.; Sai, K.; Yang, Q.; Guo, C.; Mou, Y.; Jiang, X. Establishment of Human Pituitary Neuroendocrine Tumor Derived Organoid and Its Pilot Application for Drug Screening. J Clin Endocrinol Metab 2025, 110, e827–e840. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphology of GH3 pituitary tumor cells. Live cell imaging was captured using an inverted microscope at a magnification of ×200. The shadow in the lower right corner indicates the position of the measuring electrode we applied. To perform patch-clamp recordings, the patch electrode was slowly positioned aganist the pituitary cells using a micromanipulator. Despite being dispersed in the recording chamber, the pituitary cells exhibit a tendency to aggregate.
Figure 1.
Morphology of GH3 pituitary tumor cells. Live cell imaging was captured using an inverted microscope at a magnification of ×200. The shadow in the lower right corner indicates the position of the measuring electrode we applied. To perform patch-clamp recordings, the patch electrode was slowly positioned aganist the pituitary cells using a micromanipulator. Despite being dispersed in the recording chamber, the pituitary cells exhibit a tendency to aggregate.
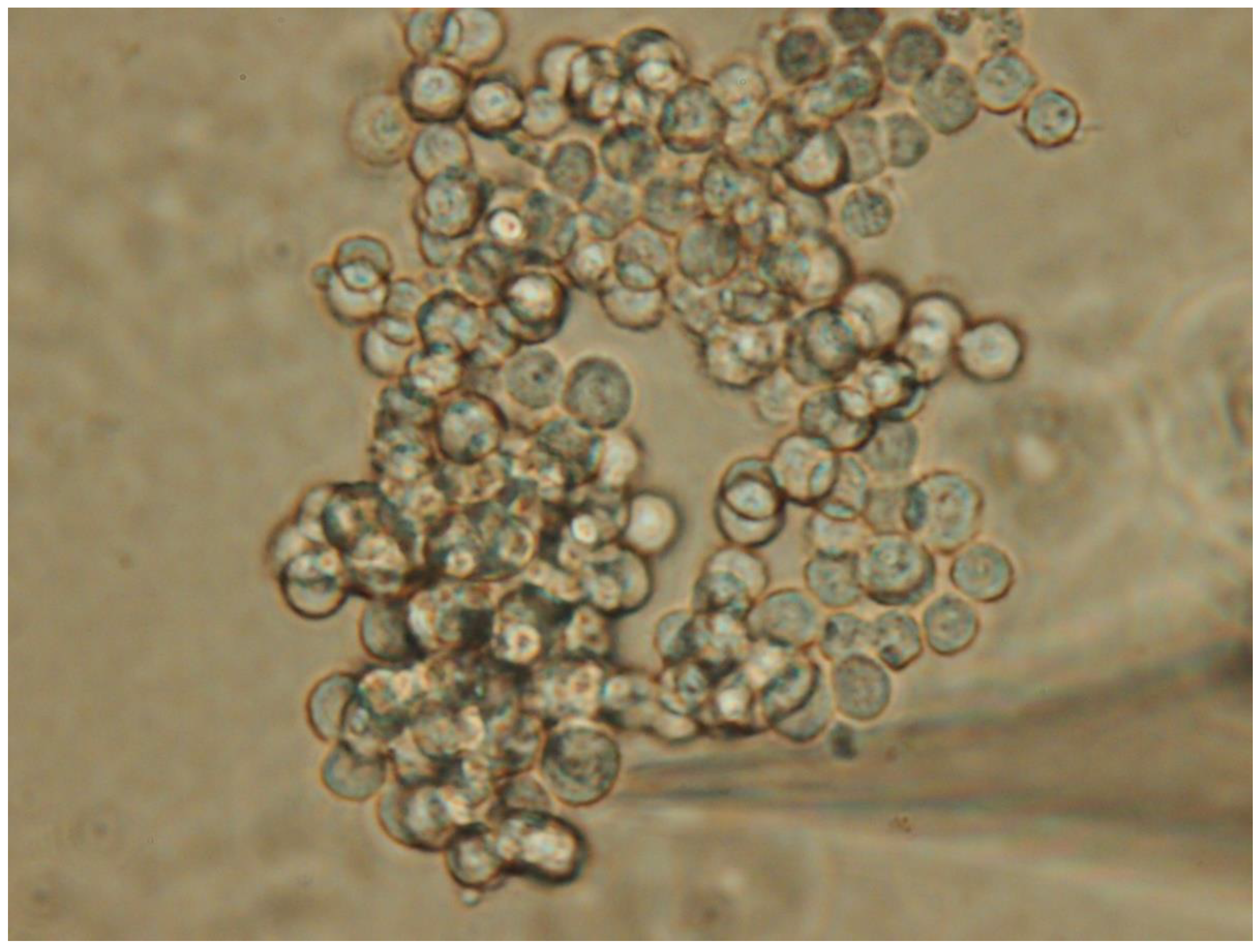
Figure 2.
Main ionic currents observed in pituitary cells, along with their corresponding voltage and current traces. The full names of these ion currents can be found in the abbreviation section of the main text. The black and red traces represent voltage and the respective ion currents. The y-axis scales for each current are indicated below the respective traces. For measurements of IK(erg) or IK(M), the cells are bathed in a high K+ solution [145 mM K+). As shown in black color, an upward voltage step denotes depolarization, while a downward step represents hyperpolarization. The detailed voltage-step protocol applied to the membrane potential requires referencing of various ionic currents. Downward current (red color) indicates inward current, meaning Na+ or K+ ions are entering the cell. Of note, “BKCa” refers to single channel activity, reflecting the opening or closing of BKCa channel in the cell membrane), while the other currents represent whole-cell ionic currents.
Figure 2.
Main ionic currents observed in pituitary cells, along with their corresponding voltage and current traces. The full names of these ion currents can be found in the abbreviation section of the main text. The black and red traces represent voltage and the respective ion currents. The y-axis scales for each current are indicated below the respective traces. For measurements of IK(erg) or IK(M), the cells are bathed in a high K+ solution [145 mM K+). As shown in black color, an upward voltage step denotes depolarization, while a downward step represents hyperpolarization. The detailed voltage-step protocol applied to the membrane potential requires referencing of various ionic currents. Downward current (red color) indicates inward current, meaning Na+ or K+ ions are entering the cell. Of note, “BKCa” refers to single channel activity, reflecting the opening or closing of BKCa channel in the cell membrane), while the other currents represent whole-cell ionic currents.
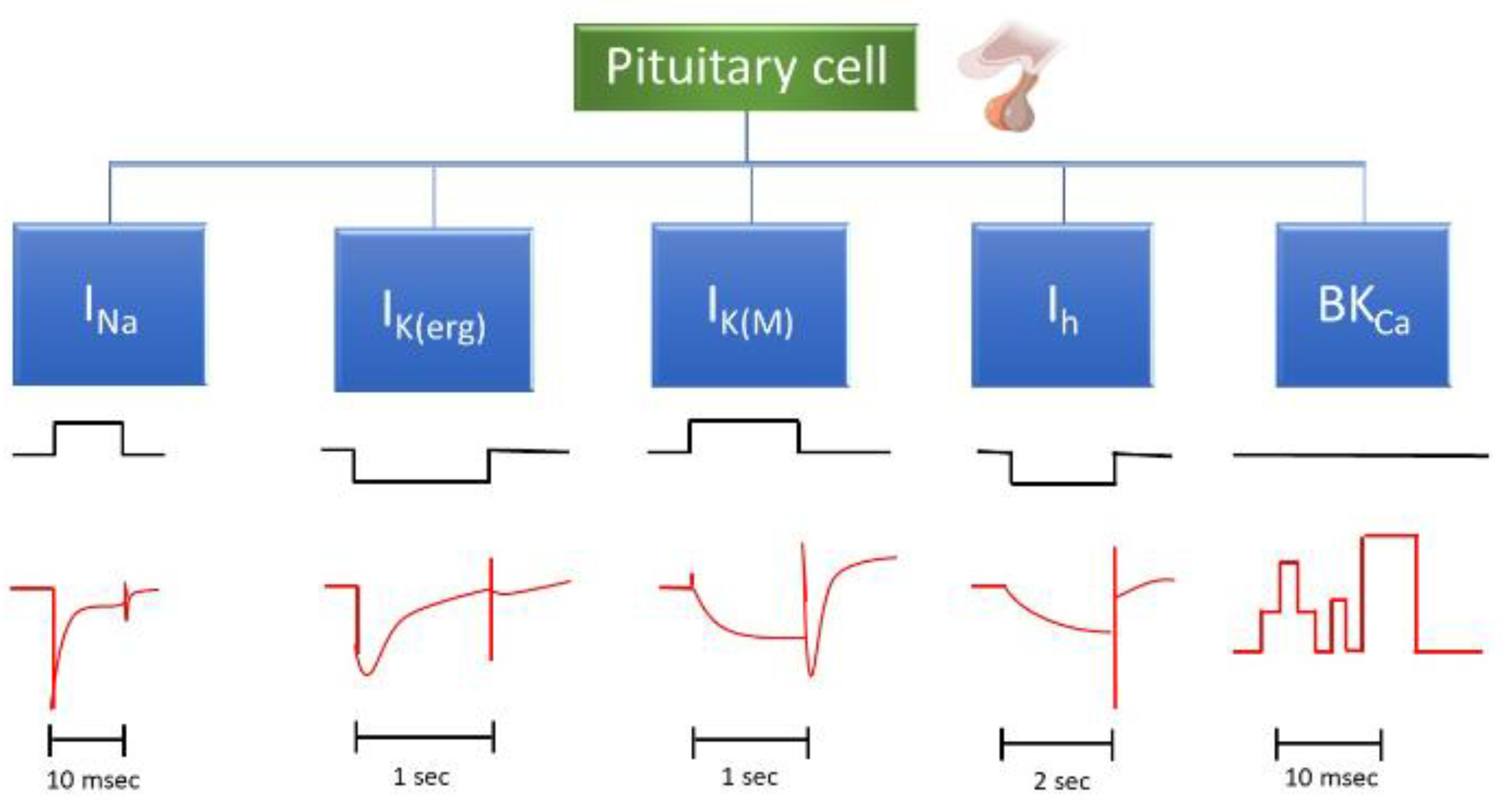
Figure 3.
Effect of ESAX on voltage-dependent hysteresis of persistent INa (INa(P)) in GH3 cells. (A) Figure-of-eight pattern in voltage-dependent hysteresis of INa(P) activated by isosceles-triangular ramp voltage with a total ramp duration of 3.2 s (or a ramp speed of ±0.094 mV/ms) in the presence of 10 mM tefluthrin (Tef), an activator of INa. The ascending limb is shown in blue, while the descending one is shown in orange. An inset at the top illustrates the applied double ramp voltage. The dashed arrow indicates the direction of the current trajectory as time progresses. (B) Summary bar graph showing the effect of Tef (10 μM) and Tef (10 μM) plus ESAX (10 μM) on INa(P) amplitude activated by the upsloping and downsloping limb of triangular ramp pulse (mean ± SEM; n = 7 for each bar). The current amplitude on the left side was taken at the level of −40 mV, corresponding to the forward (upsloping) limb of triangular pulse, which was used to evoke INa(P) (i.e., high-threshold INa(P)). On the right side, the current amplitude was measured at −80 mV, during the backward (downsloping) phase of the pulse, corresponding to low-threshold INa(P). * Significantly different from control (p < 0.05) and + significantly different from Tef (10 μM) alone group (p < 0.05). This figure is adapted from Chang and Wu (2020, ref. 20) and published under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Figure 3.
Effect of ESAX on voltage-dependent hysteresis of persistent INa (INa(P)) in GH3 cells. (A) Figure-of-eight pattern in voltage-dependent hysteresis of INa(P) activated by isosceles-triangular ramp voltage with a total ramp duration of 3.2 s (or a ramp speed of ±0.094 mV/ms) in the presence of 10 mM tefluthrin (Tef), an activator of INa. The ascending limb is shown in blue, while the descending one is shown in orange. An inset at the top illustrates the applied double ramp voltage. The dashed arrow indicates the direction of the current trajectory as time progresses. (B) Summary bar graph showing the effect of Tef (10 μM) and Tef (10 μM) plus ESAX (10 μM) on INa(P) amplitude activated by the upsloping and downsloping limb of triangular ramp pulse (mean ± SEM; n = 7 for each bar). The current amplitude on the left side was taken at the level of −40 mV, corresponding to the forward (upsloping) limb of triangular pulse, which was used to evoke INa(P) (i.e., high-threshold INa(P)). On the right side, the current amplitude was measured at −80 mV, during the backward (downsloping) phase of the pulse, corresponding to low-threshold INa(P). * Significantly different from control (p < 0.05) and + significantly different from Tef (10 μM) alone group (p < 0.05). This figure is adapted from Chang and Wu (2020, ref. 20) and published under the terms and conditions of the Creative Commons Attribution (CC BY) license.
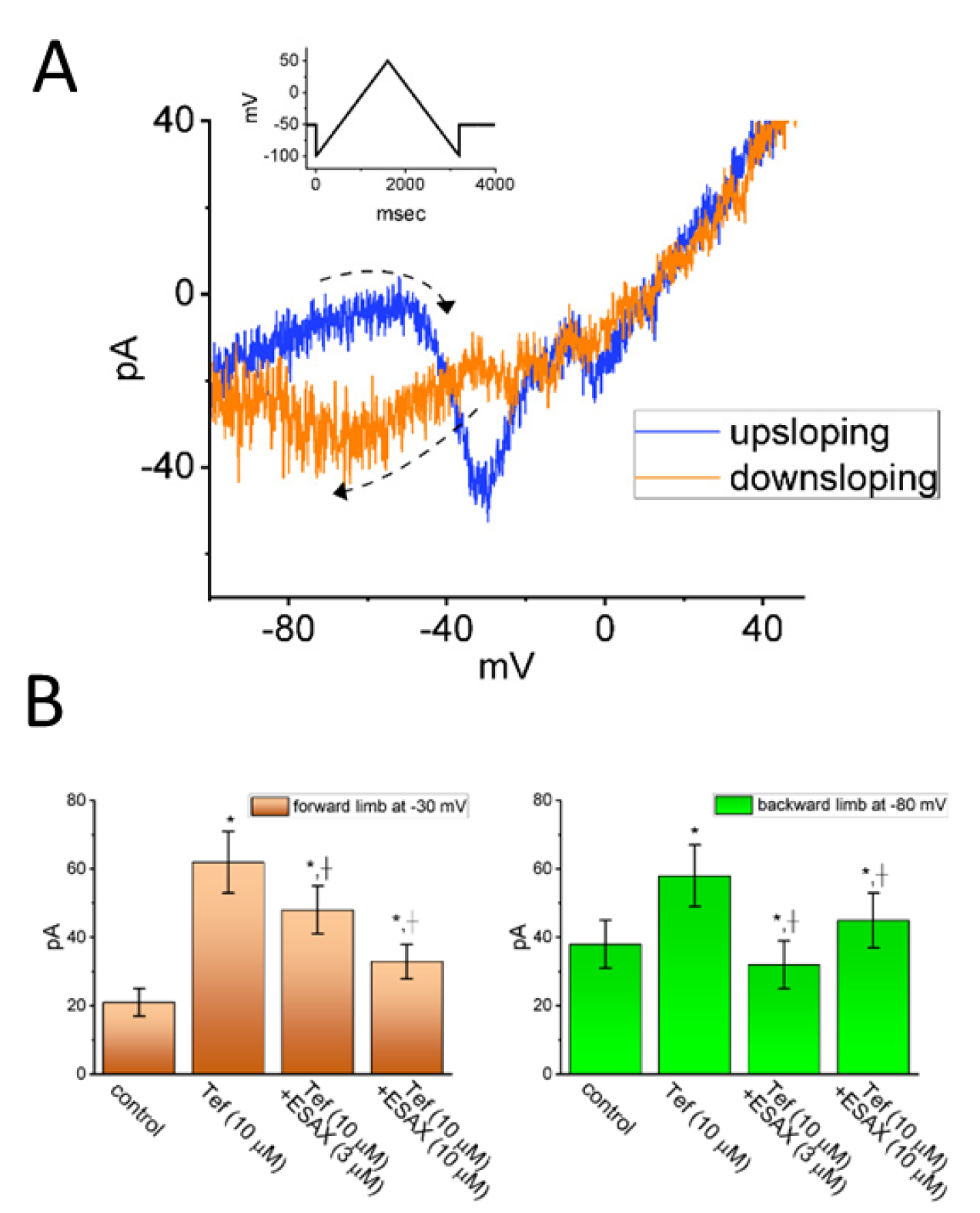
Figure 4.
Docking interaction between the di(2-ethylhexyl)-phthalate (DEHP) molecule and the HERG channel. The left graph refers to the docking prediction between DEHP and the HERG channel. The protein structure of HERG was acquired from PDB (PDB ID: 5VA1), while the chemical structure of DEHP was from PubChem (compound CID: 8343). On the right side, the yellow box and curved arrow indicates a snapshot, illustrating the hydrophobic interactions and hydrogen bond formation between DEHP and the channel. Note that the red arcs, with spokes directed towards the ligand (such as the DEHP molecule), indicates the presence of hydrophobic contacts. The green dashed line represents the formation of hydrogen bond, with a distance of 3.00 Å.
Figure 4.
Docking interaction between the di(2-ethylhexyl)-phthalate (DEHP) molecule and the HERG channel. The left graph refers to the docking prediction between DEHP and the HERG channel. The protein structure of HERG was acquired from PDB (PDB ID: 5VA1), while the chemical structure of DEHP was from PubChem (compound CID: 8343). On the right side, the yellow box and curved arrow indicates a snapshot, illustrating the hydrophobic interactions and hydrogen bond formation between DEHP and the channel. Note that the red arcs, with spokes directed towards the ligand (such as the DEHP molecule), indicates the presence of hydrophobic contacts. The green dashed line represents the formation of hydrogen bond, with a distance of 3.00 Å.
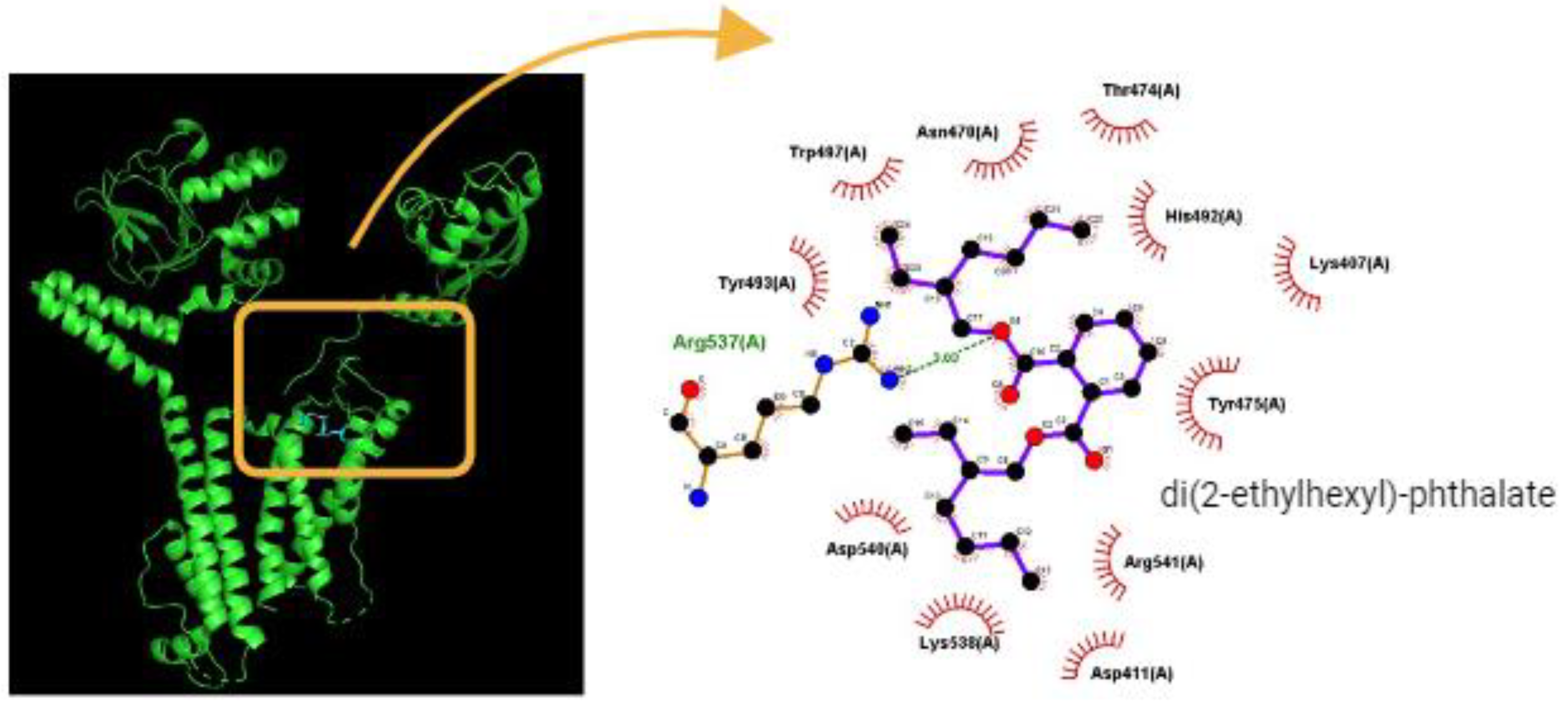
Figure 5.
Effect of rufinamide (RPM) on mean current-voltage (I-V) relationships of Ca2+-activated K+ current (IK(Ca)) identified in GH3 cells. (A) Representative current traces obtained in the absence (upper) and presence (lower) of 10 μM RFM. The uppermost part shows the voltage-clamp protocol applied. The potential traces labeled in different colors correspond with current ones acquired without or with the RPM presence. The duration in each depolarizing step is different for better illustrations, and the blue solid arrow indicates the outwardly rectifying properties of IK(Ca) with increasing positive voltage. (B) Mean I-V relationships of IK(Ca) amplitude acquired in the control (blue squares), during exposure to 10 μM RPM (red squares), and following washout of RFM (black squares) (mean ± SEM for each point). Current amplitude was measured at the end of each depolarizing step. This figure is adapted from Lai el al. (2022) (ref. 73) and published under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Figure 5.
Effect of rufinamide (RPM) on mean current-voltage (I-V) relationships of Ca2+-activated K+ current (IK(Ca)) identified in GH3 cells. (A) Representative current traces obtained in the absence (upper) and presence (lower) of 10 μM RFM. The uppermost part shows the voltage-clamp protocol applied. The potential traces labeled in different colors correspond with current ones acquired without or with the RPM presence. The duration in each depolarizing step is different for better illustrations, and the blue solid arrow indicates the outwardly rectifying properties of IK(Ca) with increasing positive voltage. (B) Mean I-V relationships of IK(Ca) amplitude acquired in the control (blue squares), during exposure to 10 μM RPM (red squares), and following washout of RFM (black squares) (mean ± SEM for each point). Current amplitude was measured at the end of each depolarizing step. This figure is adapted from Lai el al. (2022) (ref. 73) and published under the terms and conditions of the Creative Commons Attribution (CC BY) license.
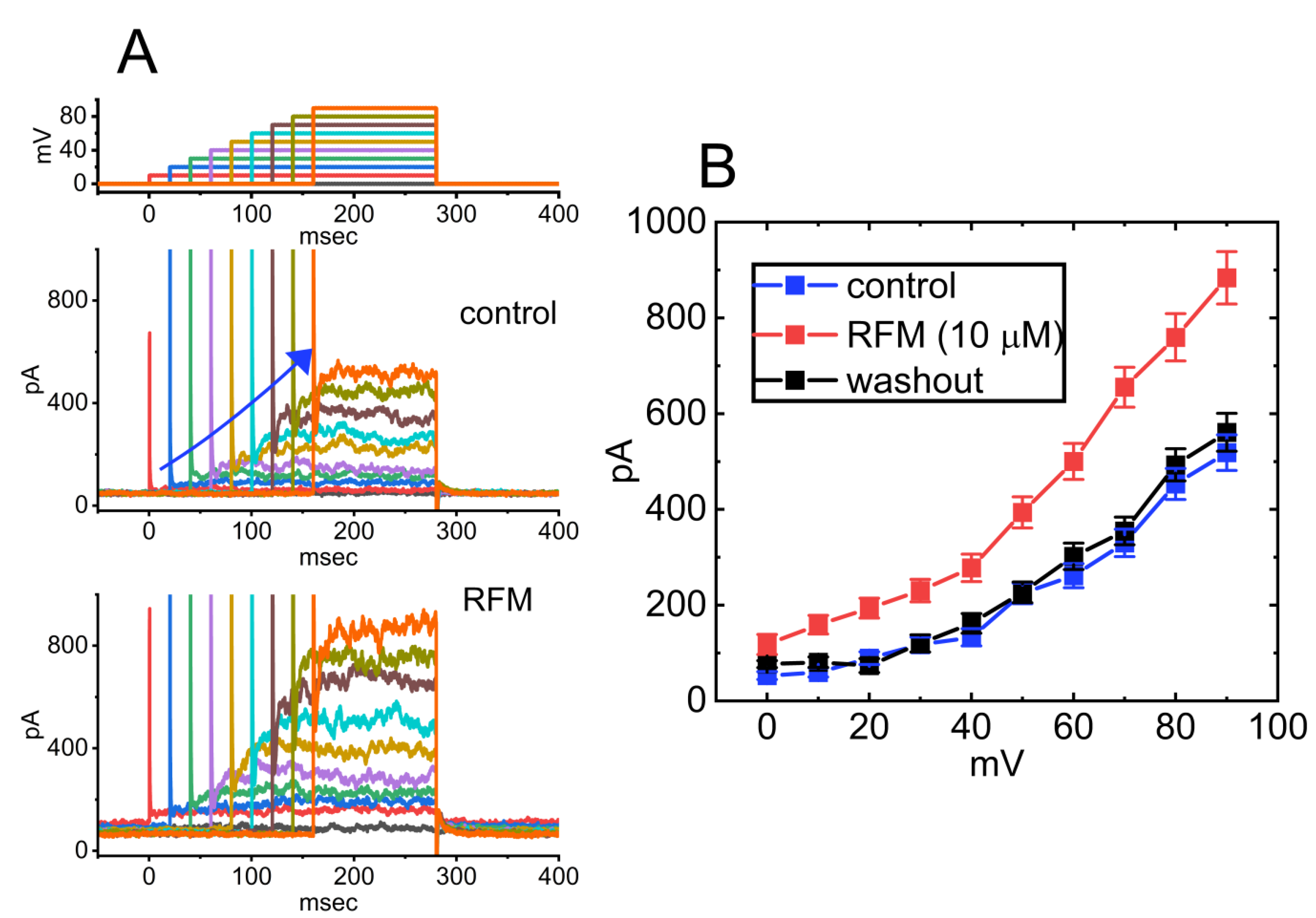

Table 1.
A brief description of the transmembrane ionic currents known to exist in pituitary cells, along with the effects of various drugs or compounds with their respective chemical structures on these ion currents demonstrated in this paper.
Table 1.
A brief description of the transmembrane ionic currents known to exist in pituitary cells, along with the effects of various drugs or compounds with their respective chemical structures on these ion currents demonstrated in this paper.
| Ionic current | Chemical or drug | Abbreviated name | Chemical structure* | Reference |
| INa | GV-58 | NA | 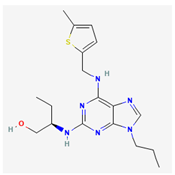 |
Cho et al., 2022 |
| Esaxerenone | ESAX | 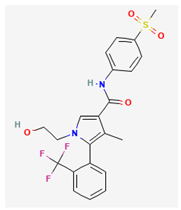 |
Chang and Wu, 2021a | |
| IK(erg) | Risperidone | NA | 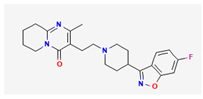 |
Lee et al., 2017, Wu et al., 2000 |
| Di(2-ethylhexyl)-phthalate | DEHP |  |
Wu et al., 2012 | |
| IK(M) | Solifenacin | SOL |  |
Cho et al., 2021 |
| Kynurenic acid | KYNA |  |
Sakakibara et al., 2015, Lo et al., 2021 | |
| Ih | Carisbamate | CRS |  |
Chen et al., 2023, Hung et al., 2023 |
| Cannabidiol | CBD | 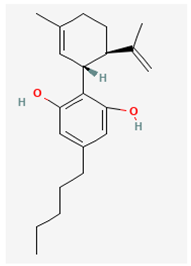 |
Liu et al., 2023 | |
| BKCa channel | Rufinamide | RFM | 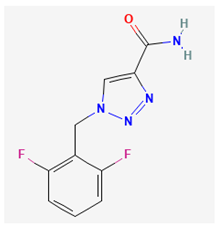 |
Lai et al., 2022 |
| QO-40 | NA | 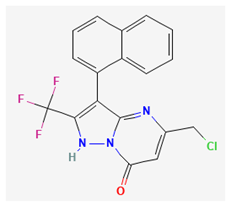 |
Chang and Wu, 2021b |
Table 2.
Pituitary cell lines cited in this article, along with their sources and links to websites for the detailed information.
Table 2.
Pituitary cell lines cited in this article, along with their sources and links to websites for the detailed information.
| Cell line | ATCC* website | BCRC* website | ScienCellTM website |
| AtT-20 | https://www.atcc.org/products/ccl-89 (CCL-89) | https://catalog.bcrc.firdi.org.tw/BcrcContent?bid=60244&rowid=1 | |
| GH3 | https://www.atcc.org/products/ccl-82.1 (CCL-82.1) | https://catalog.bcrc.firdi.org.tw/BcrcContent?bid=60015&rowid=1 | |
| GH4C1 | https://www.atcc.org/products/ccl-82.2 (CCL-82.2) | NA | |
| MMQ | https://www.atcc.org/products/crl-10609 (CRL-10609) | NA | |
| R1220 | https://sciencellonline.com/rat-pituitary-cel |
* ATCC stands for the American type culture collection (Manassas, VA), while BCRC refers to the Bioresource Collection and Research Center (Hsinchu, Taiwan). NA = non-available.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



