
Submitted:
31 March 2025
Posted:
01 April 2025
You are already at the latest version
Abstract
Epigenetic modifications play a crucial role in gene regulation and have been implicated in various physiological processes and disease conditions. DNA methylation (DNAm) has been implicated in the etiology and progression of many stress-related psychiatric behaviors such as depression. The ability to manipulate DNAm may provide a means to reverse and treat such disorders. Although CRISPR-based technologies have enabled locus-specific DNAm editing, their clinical applicability may be limited due to immunogenicity concerns and off-target effects. In this study, we introduce a novel approach for targeted DNAm manipulation using single-stranded, methylated DNA probes. We designed probes targeting the glucocorticoid response element (GRE) within the FKBP5 (FK506 binding protein 5) gene, a key regulator of stress response and depressive symptoms, and the promoter region of the MAOA (monoamine oxidase A) gene. In both HEK293 human embryonic kidney and mouse pituitary AtT-20 cells, transfection with their respective methylated probes significantly increased DNAm at targeted CpG sites in a persistent and dose-dependent manner. Importantly, the induced methylation effectively attenuated glucocorticoid-induced upregulation of FKBP5 gene expression. Alteration of methylation was specific to single-stranded probes, as double-stranded methylated probes and unmethylated probes showed no significant effects. Our findings suggest that methylated DNA probes have the potential to function as a simple tool for targeted epigenetic manipulation and serve as a safer alternative to CRISPR-based epigenome editing tools for the treatment of stress-related disorders such as depression.
Keywords:
Epigenetics
; DNA methylation (DNAm)
; gene expression
; cortisol
; FK506 Binding Protein 5 (FKBP5)
Introduction
Epigenetic modifications, such as DNA methylation (DNAm), can lead to alterations in nuclear architecture and landscape, which can in turn affect the accessibility of transcription and regulatory factors to genes. DNAm typically occurs at 5’ end cytosine residues within CpG dinucleotides and is generally associated with transcriptional repression, which is mediated by the binding of methyl-CpG binding proteins and recruitment of histone-modifying enzymes [1]. Aberrant DNAm has been linked to many diseases from cancer to neurodegenerative disorders, but also psychiatric diseases such as depression, anxiety, and PTSD [2,3,4,5,6,7].
Epigenetic mechanisms can also mediate the impact of adverse environmental conditions on gene function. For instance, environmental stressors can cause epigenetic changes in the brain [8,9]. In such instances, it has been shown that the glucocorticoid (GC) receptor that binds to cortisol can directly alter DNAm of genes that are targets of GC signaling, thus, raising the possibility of mitigating disease symptoms by potentially reversing these epigenetic marks.
Targeted manipulation of DNAm at specific gene loci can modulate gene function and provide new therapeutic strategies for disorders associated with aberrant epigenetic regulation. In plants, small RNAs have been shown to direct DNAm and gene silencing [10]. Studies have shown the possibility of using pharmacological methods, such as DNMT inhibitors, to impact DNAm in mammalian systems. However, these drugs target the epigenetic machinery and are not locus-specific [11]. Although recent advances in CRISPR-based technologies have enabled locus-specific DNAm editing in mammalian systems [12,13,14], these methods may have limited clinical applicability due to their immunogenicity [15,16,17] and off targeting [18,19]. However, CRISPR-based therapies are now on the horizon for the treatment of debilitating diseases such as sickle cell-disease [20] and cancer [21]. Until these technologies are further refined to minimize their own disease burden and gain traction for the treatment of more common disorders, safer methods need to be explored.
The ability of the glucocorticoid receptor to induce loss of DNAm provides a useful model for testing alternative epigenetic tools. In particular, the FK506 binding protein 5 (FKBP5) gene encodes a co-chaperone of the glucocorticoid receptor and has been identified as a key regulator of the stress response [22]. It is thought that GC-induced increase in FKBP5 levels leads to attenuated intracellular signaling and GC resistance, which are comorbid in more than 50% of cases of depression [23]. As such, genetic and epigenetic variations have been linked to depressive symptoms [24,25]. At the molecular levels, chronic exposure to stress or excess glucocorticoids can induce the persistent demethylation of intronic glucocorticoid response elements (GREs) in the FKBP5 gene [26]. This demethylation allows for increased binding of the GR to the GREs and a more robust transcription of FKBP5, which in turn leads to decreased sensitivity to GCs and GC resistance [27,28]. Epigenetic alterations in FKBP5 have been linked to several stress-related psychiatric disorders such as PTSD, anxiety, and alcohol abuse [5,25,29,30].
In this study, we sought to develop a simpler tool for site-specifically altering DNAm. We asked whether a single-stranded, methylated DNA probe can induce DNAm of its complementary target. Specifically, we sought to reverse the persistent loss of DNAm caused by chronic exposure to GCs. Our findings suggest that methylated DNA probes may serve as a promising tool for targeted epigenetic manipulation and have potential therapeutic applications for mitigating the impact of excess stress or GC exposure in psychiatric disorders such as depression and non-psychiatric disorders associated with aberrant DNAm patterns. Further, it may be refined to target other genes in a simpler and safer way that can circumvent some of the limitations posed by the CRIPR technology.
Methods
Probe Design and Amplification
Methylated DNA probes targeting the conserved glucocorticoid response element (GRE) in intron 5 of the human (chr6:35,601,961 – 35,602,194; GRCh38/hg38, 256 bp) and mouse (chr17: 28,639,321 – 28,639,560; GRCm39/mm39, 239 bp) of FKBP5 were designed. A separate DNA probe designed against an intronic, regulatory region of the human MAOA gene (chrX:43,656,383 – 43,656,553; GRCh38/hg38, 170 bp) was also tested [31]. Additional tests for non-specific effects of the human FKBP5 probe investigated another adjacent region in intron 5 (chr6:35,610,962 – 35,611,313), intron 1 (chr6:35,687,767-35,688,045), and intron 7 (chr6:35,590,524-35,591,014). For both human and mouse probes, PCR primers targeted smaller regions than those analyzed by bisulfite sequencing to preclude the amplification of probe DNA during methylation analysis. Primers used for generating the probes are shown in Table 1. The genomic organization of the human FKBP5 locus is shown in Figure 1.
In Vitro Methylation of DNA Probes
Purified FKBP5 probes were subjected to in vitro methylation using the bacterial CpG methyltransferase (M.SssI, New England Biolabs) [32]. One µg of the probe was incubated with 4 units of M.SssI, 160 µM S-adenosylmethionine (SAM), and 1X NEBuffer 2 in a total reaction volume of 100 µL at 37°C for two 1-hour cycles followed by 20 minutes at 65°C for enzyme inactivation. A negative control reaction (FKBP5 SssI-) was performed in parallel, where the M.SssI enzyme was replaced with an equal volume of water. Following in vitro methylation, probes were purified again and eluted in 20 µL of EB buffer. The concentrations of the methylated (FK SssI+) and unmethylated (FK SssI-) probes were measured using Qubit 4 Thermo Fisher Scientific, Waltham, MA). Similar reactions were performed for the mouse Fkbp5 and human MAOA probes.
Cell Culture and Transfection
To induce demethylation at the GRE of the endogenous FKBP5 gene, we chose cell lines that we have previously shown to undergo DEX or glucocorticoid-induced loss of methylation [33,34]. Human embryonic kidney 293 (293HEK) and mouse pituitary AtT-20 cells were purchased from Atcc.org and were treated with 1 µM dexamethasone (DEX) for 5 days and cultured for an additional 5 days without any DEX [27]. This would allow CpGs to undergo persistent loss of DNA methylation which would then be restored by introducing methylated probes. Separate wells of cells were left untreated as negative controls or transfection reagent only controls. Prior to transfection, cells were seeded in 24-well plates at a density of 2 × 105 cells per well and allowed to adhere overnight in DMEM free of antibiotics. The transfection of the methylated and unmethylated probes was performed in triplicate (500 ng of probe per well) using X-tremeGENE 360 (MilliporeSigma, Burlington, MA) according to the manufacturer’s instructions. Cells were fed fresh DMEM media one day after transfection and harvested on the second day for collecting gDNA. For the assessment of glucocorticoid-induced gene expression, a subset of the transfected 293HEK cells was treated with 1 µM DEX for 4 hours prior to harvesting. Cells were harvested for total RNA extraction using RNeasy Mini Kit (Qiagen) according to the manufacturer’s instructions.
DNA Extraction and Methylation Analysis by Bisulfite Pyrosequencing
Genomic DNA was extracted from 293HEK and AtT-20 cells using the DNeasy Blood & Tissue Kit (Qiagen) according to the manufacturer's instructions. The extracted DNA was bisulfite-converted using the EZ DNAm-Gold Kit (Zymo Research, Irvine CA) according to the manufacturer’s instructions. Conditions for bisulfite pyrosequencing have been published elsewhere [35]. Primers used for assessing DNAm levels of FKBP5 and MAOA genes are shown in Table 1.
Gene Expression Analysis
Reverse Transcription Quantitative Real-Time PCR (RT-qPCR) was performed to assess the impact of targeted DNAm on gene expression. Complementary DNA (cDNA) was synthesized from 293HEK or AtT-20 RNA samples using QuantiTect Reverse Transcription Kit (Qiagen) according to the manufacturer’s instructions. Procedures for gene expression analysis using the Taqman probe and QuantStudio5 platform (Thermo Fisher Scientific) have been published elsewhere [36].
Results
The Use of Methylated, Single-Stranded Probe to Induce Target-Specific DNAm
To test whether a simple DNA probe can target-specifically alter DNAm, we generated a probe against specific CpG sites within the glucocorticoid response element (GRE) of the human FKBP5 gene. In vitro-methylated, single- or double-stranded DNA probes were generated and transfected into human HEK293 cells. Probes were designed with sufficiently short lengths to avoid amplification by subsequent epigenetic assays (Figure 1). Two days post-transfection, cells were analyzed for DNAm levels of the FKBP5 GRE by bisulfite pyrosequencing. Pyrosequencing analysis showed an increase of 20.5% increase in DNAm at CpG-1 (P=7.0x10-4) and an increase of 15.0% at CpG-2 (P= P=4.9x10-4) in 500 ng of methylated, single-stranded probe compared to untransfected samples (Figure 2A). We also observed a dose-response when we performed transfections with only 250 ng of probe, with CpG-1 showing a more modest increase of 8.6% (P=0.0066) and CpG-2 showing an increase of 7.6% (P=1.1x10-4) compared to untransfected samples. Samples transfected with unmethylated, single-stranded or methylated, double-stranded probes did not lead to an appreciable increase in DNAm (P>0.05). Representative pyrosequencing tracing is shown in Figure 2B.
The Effect of Probe-Induced DNAm on Gene Expression
We also investigated whether the increase in DNAm was associated with differential gene expression. Previous studies have shown that while DNAm changes may not immediately result in changes in gene expression, they can modulate the level to which a gene can respond to a stimulus. In this case, the GREs of FKBP5 are responsive to glucocorticoids in a methylation-sensitive way [34]. Treatment of the 293HEK cells with dexamethasone (DEX) caused a significant increase in FKBP5 expression (2.5-fold, P=0.036). While transfection with unmethylated, single-stranded or methylated, double-stranded probes did not lead to an appreciable attenuation of DEX-induced increase in expression, methylated, single-stranded probes significantly reduced the DEX-induced increase in FKBP5. Specifically, cells treated with methylated, single-stranded 500 ng of probe DNA showed 76.2% reduction in expression compared to that of unmethylated, single-stranded 500 ng of probe DNA treated with DEX. Furthermore, we observed a dose-response with methylated, single-stranded 250 ng of probe DNA, where 29.2% reduction in expression was observed compared to unmethylated, single-stranded 500 ng of probe DNA treated with DEX (Figure 2C).
Persistence of Probe-Induced DNAm Patterns and Accumulation of DNAm Following Multiple Probe Transfections
Previously, we demonstrated that glucocorticoid-induced DNA methylation persisted throughout development [26]. This persistence was replicated in a simpler mouse neuronal cell line treated with DEX [34]. Therefore, we tested whether DNA probe-induced increase in DNA methylation can persist through several weeks. Cells transfected once with 500 ng of methylated DNA probe showed appreciable increase in DNAm across all 5 CpGs compared to unmethylated DNA probe (P<0.038) at Week 1 (W1). Further culturing the transfected cells for four weeks showed that those transfected once with methylated DNA probe retained their DNA methylation patterns at CpG-1, CpG-3, and CpG-5 (P<0.0054) compared to cells transfected with unmethylated DNA probe at Week 1 or Week 4 (Figure 3A). We observed no significant increase in DNAm at CpG-2 at Week 1 and hence no significant increase at Week 4. At CpG-4, we observed significant increase in DNAm by 10.4% but this probe-induced increase in DNAm showed a decay of 5.2% by Week 4 (P=0.003). We also tested whether repeated transfections can have a cumulative effect on DNA methylation. 293HEK cells were transfected once, twice, three-times, and four-times with the same amount of DNA probes, each transfection taking place three days following the previous transfection. Results showed a cumulative effect on CpGs 3,4, and 5, with a more profound increase in DNAm at CpGs 1 and 2 following third and fourth transfections (Figure 3B).
Effect of DNA Probes in Non-Targeted Regions
We also tested the effect of our targeted probe on non-targeted regions. Chromosome conformation capture experiments have demonstrated physical interactions between GREs and the promoter [5,34], and it has been speculated that GREs that are scattered across several intronic regions of a glucocorticoid-responsive gene interact in the 3-D space of the nucleus to coordinate glucocorticoid-induced gene expression. It may be possible that other genomic regions in proximity with the region targeted by the probe may be exposed to the DNA methylation machinery. Therefore, we assayed the DNAm levels of a GRE immediately adjacent to the probe-targeted GRE in intron 5, a GRE in intron 7, and a GRE immediately downstream of the first exon in intron 1 (Figure 4A,B). DNAm analysis showed that most of the CpGs tested in introns 5 and 7 were too hypermethylated to undergo probe-induced DNA methylation. CpG-6 in intron 7, which was strangely hypomethylated compared to other CpGs (<2% for all treatment groups), did not show any increase in DNAm. Further, hypomethylated intron 1 CpGs showed no differences among untransfected cells, cells treated with the unmethylated probe, and cells treated with the methylated probe. Only exception was at CpG-1, where cells transfected with the methylated probe showed a small 1.3% increase in DNAm compared to the other groups (P=0.004, Figure 4C).
Epigenetic and Transcriptional Effects of Methylated, Single-Stranded Probe in a Mouse Pituitary Cell Line
We also tested mouse pituitary cells to determine whether the findings in 293HEK can be recapitulated in a different cell-type and species. Mouse AtT-20 cells were transfected with the mouse version of the probe against the conserved Fkbp5 intron 5 GRE. Results showed that the use of methylated, single-stranded probes increased DNAm at three of the four CpGs when compared to unmethylated single-stranded probes: CpG-2 (9.5%, P=0.003), CpG-3 (6.1%, P=0.003), and CpG-4 (7.2%, P=0.03). There were no appreciable differences in DNAm between unmethylated, single-stranded probes and methylated, double-stranded probes (Figure 5A). We then tested whether transfected DNA probes can modulate DEX-inducibility as the human probes had in the 293HEK cells. Fkbp5 expression analysis showed that samples treated with no DNA probe, methylated, double-stranded probe, or unmethylated, single-stranded probe had similar levels of gene induction by DEX treatment (7.8-, 6.3-, 6.7-fold induction, respectively, and P<1.3x10-4). However, samples transfected with methylated, single-stranded probe showed a significant reduction in DEX-induced expression compared to samples transfected with unmethylated, single-stranded probe (42.5% reduction, P=6.4x10-5, Figure 5B).
Additional Genomic Target of DNAm Probe: MAOA.
To demonstrate the broader applicability of our methylated probe approach beyond the FKBP5 gene, we tested the regulatory intronic region of Monoamine Oxidase A (MAOA) [31], a gene whose encoded protein metabolizes monoamine neurotransmitters such as dopamine, serotonin, and norepinephrine. Following the transfection of methylated single-stranded probes targeting the CpG-dense MAOA intronic region into 293HEK cells, we observed significant increases in DNAm across multiple CpG sites compared to cells transfected with unmethylated probes: CpG-1 (10.1%, P=0.01), CpG-5 (12.7%, P=0.04), CpG-6 (11.2%, P=0.02), CpG-8 (9.3%, P=0.006), CpG-10 (11.8%, P=0.006), and CpG-12 (14.3%, P=0.01) (Figure 6A). A one-way ANOVA revealed a significant difference between the two treatments (F(1, 26) = 6.25, P = 0.019), where methylation levels were higher in the methylated probe-treated group (M = 58.68, SD = 7.52) compared to the unmethylated probe-treated group (M = 52.31, SD = 10.25). Consistent with the increased methylation, we observed a corresponding decrease in MAOA gene expression. qRT-PCR analysis revealed that cells transfected with methylated probes showed a 27.3% reduction in MAOA expression compared to cells transfected with unmethylated probes (P=0.041) (Figure 6B).
Discussion
In this study, we introduce a simple approach for inducing targeted DNAm using methylated DNA probes. Our approach consists of the PCR amplification against a region of interest, which in this case was a methylation-sensitive, glucocorticoid response element in the intronic region of the FKBP5 gene [27]. The PCR product was first in vitro-methylated using the bacterial methyltransferase M.SssI and denatured to yield single-stranded DNA probes. Using the DNA probes against the FKBP5 GRE, we observed appreciable increases in DNAm across at least two of the five CpGs across the GRE in the human 293HEK cells. The observed increase in DNAm was specific to the single-stranded, methylated probe, as double-stranded, methylated probe and single-stranded unmethylated probe did not induce an increase in DNAm. Importantly, increase in DNAm at these CpGs was able to attenuate DEX-induced increase in FKBP5 levels. Further, gene regulation by the DNA probe was dose-dependent, as the introduction of half the amount of DNA probe resulted in a reduced addition of DNAm and reduced attenuation of FKBP5 induction by DEX compared to the full amount of DNA probe.
Compared to the other two epigenetic mechanisms, namely histone modification and RNA-mediated gene silencing, DNA methylation tends to be more stable. As such, a single administration of the DNA probe lasted through at least a four-week period, suggesting robustness of the probe-induced DNA methylation. This phenomenon is reminiscent of glucocorticoid loss of DNAm, which persisted for at least four weeks in the mouse brain and in a DEX-treated neuronal cell line [26,34]. It should be noted that the reversal of DNAm induction observed at CpG-4 in Figure 3A is not new, as some of the CpGs that lost DNAm by glucocorticoid treatment also showed a similar reversion to baseline by the fourth week [34]. In addition, repeated introduction of the probe had a cumulative effect on DNA methylation, although the increase in DNAm with successive transfections was not at the same magnitude as following the first transfection.
Investigation of other GREs in intronic regions, which have been thought to physically interact in 3D-space with the intronic GRE (intron 5) targeted by the probe, did not show any significant changes in DNAm, emphasizing the specificity of probe targeting. One exception to the negative results at other GREs is the increase in DNAm at one CpG in intron 1 (Figure 4C). However, the magnitude of the difference was less than 2%, which is unlikely to be functionally relevant.
We then tested whether probe-induced DNAm was unique to human cells by testing the mouse version of the Fkbp5 probe in the AtT-20 mouse pituitary cell line. Although more subtle than those observed in the human cell lines, increase in DNAm was significant. Similarly, this increase in DNAm was able to thwart DEX-induction of Fkbp5. There are a few factors that can potentially explain the difference in the magnitude of probe-induced increase in DNAm, including cell-type and species-specific differences, growth rate differences of cells, and different levels of DNAm-modifying enzymes. Growth rate may play an important role, as some methylation-altering events, such as glucocorticoid-induced loss of methylation, depends on cell proliferation [27].
An additional genomic region was tested to further generalize the probe-induced increase in DNAm. This time we designed a probe against an important regulatory region of MAOA, whose methylation levels correlated with enzymes levels determined by PET brain imaging [31]. In addition, the MAOA enzyme metabolizes the neurotransmitter serotonin and is thus targeted by a class of antidepressants called monoamine inhibitors (MAOI). Although significant increases in DNAm were observed across many CpGs, many CpGs were impervious to the probe. The sub-optimal increase in DNAm at these CpGs may be due to the sequence of the regulatory region in MAOA. It is also possible that CpG-dense regions may be harder to modify, as the MAOA probe covers approximately the same base pairs of DNA as that for FKBP5 but contains almost three times more CpGs.
Our current study highlights the potential of this method to modulate stress response pathways, as demonstrated by the targeted methylation of the FKBP5 gene. FKBP5 is known to be influenced by glucocorticoid exposure and methylation status, with demethylation of the glucocorticoid response element leading to increased gene expression upon subsequent glucocorticoid exposure [27]. By inducing targeted methylation of this region, we were able to attenuate the glucocorticoid-induced upregulation of FKBP5 expression, suggesting that our approach could be used to fine-tune stress response pathways and potentially mitigate the detrimental effects of chronic stress or glucocorticoid exposure.
The success of our method in both human and mouse cell lines indicates its potential for translation to in vivo models and eventual clinical applications. However, several challenges need to be addressed before this approach can be fully realized. First, the efficiency of probe delivery and cellular uptake may vary depending on the cell type and target tissue, requiring optimization of transfection methods and probe design. Second, the long-term stability and persistence of the induced methylation changes need to be evaluated in vivo to determine the durability of the therapeutic effects. The safety and potential off-target effects of the methylated probes must also be thoroughly assessed in animal models before considering human applications. Finally, we will need a suitable delivery device that can tissue-specifically deliver DNA probes for in vivo studies. Small extracellular vesicles (EVs) such as exosomes hold immense therapeutic value as delivery devices, as their vesicle cargo space can easily accommodate a ~200 bp DNA fragment and their surface proteins profiles can be modified to alter tissue targeting [37].
Despite these challenges, our study provides a proof-of-concept for the use of methylated DNA probes as a targeted epigenetic therapy. The ability to induce site-specific DNAm changes can open new avenues for the treatment of a wide range of diseases characterized by aberrant methylation patterns. As our understanding of the epigenetic basis of disease continues to grow, the development of targeted epigenetic therapies, such as the one presented here, will become increasingly important. In conclusion, we have developed a novel method for targeted induction of DNAm using methylated DNA probes. This approach offers a promising tool for modulating gene expression and function, with potential therapeutic applications in various diseases characterized by aberrant methylation patterns.
Author Contributions
Nishtaa Modi – conceptualization, investigation, funding acquisition, and original draft preparation; Jeffrey Guo – investigation and validation; Ryan Lee – investigation and validation; Alisha Greenstein – investigation and validation; and Richard Lee - conceptualization, investigation, funding acquisition, and original draft preparation.
Funding
This work was supported by the Woodrow Wilson Fellowship (Johns Hopkins University) granted to N.M. and George Browne Genetics Cell Laboratory Fund granted to R.S.L.
References
- Jones PL, Veenstra GJ, Wade PA, Vermaak D, Kass SU, Landsberger N, Strouboulis J, Wolffe AP. Methylated DNA and MeCP2 recruit histone deacetylase to repress transcription. Nat Genet 1998; 19:187-91.
- Baylin SB. Tying it all together: epigenetics, genetics, cell cycle, and cancer. Science 1997; 277:1948-9. [CrossRef]
- Jones PA, Baylin SB. The fundamental role of epigenetic events in cancer. Nat Rev Genet 2002; 3:415-28.
- Landgrave-Gomez J, Mercado-Gomez O, Guevara-Guzman R. Epigenetic mechanisms in neurological and neurodegenerative diseases. Front Cell Neurosci 2015; 9:58. PMC4343006.
- Klengel T, Mehta D, Anacker C, Rex-Haffner M, Pruessner JC, Pariante CM, Pace TW, Mercer KB, Mayberg HS, Bradley B, et al. Allele-specific FKBP5 DNA demethylation mediates gene-childhood trauma interactions. Nat Neurosci 2013; 16:33-41. PMC4136922.
- Uddin M, Aiello AE, Wildman DE, Koenen KC, Pawelec G, de Los Santos R, Goldmann E, Galea S. Epigenetic and immune function profiles associated with posttraumatic stress disorder. Proc Natl Acad Sci U S A 2010; 107:9470-5. PMC2889041.
- Murphy TM, O'Donovan A, Mullins N, O'Farrelly C, McCann A, Malone K. Anxiety is associated with higher levels of global DNA methylation and altered expression of epigenetic and interleukin-6 genes. Psychiatr Genet 2015; 25:71-8. [CrossRef]
- Tsankova NM, Berton O, Renthal W, Kumar A, Neve RL, Nestler EJ. Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action. Nat Neurosci 2006; 9:519-25. [CrossRef]
- Weaver IC, Cervoni N, Champagne FA, D'Alessio AC, Sharma S, Seckl JR, Dymov S, Szyf M, Meaney MJ. Epigenetic programming by maternal behavior. Nat Neurosci 2004; 7:847-54.
- Erdmann RM, Picard CL. RNA-directed DNA Methylation. PLoS Genet 2020; 16:e1009034. PMC7544125.
- Yang X, Lay F, Han H, Jones PA. Targeting DNA methylation for epigenetic therapy. Trends Pharmacol Sci 2010; 31:536-46. PMC2967479. [CrossRef]
- Liu XS, Wu H, Ji X, Stelzer Y, Wu X, Czauderna S, Shu J, Dadon D, Young RA, Jaenisch R. Editing DNA Methylation in the Mammalian Genome. Cell 2016; 167:233-47 e17. PMC5062609.
- Vojta A, Dobrinic P, Tadic V, Bockor L, Korac P, Julg B, Klasic M, Zoldos V. Repurposing the CRISPR-Cas9 system for targeted DNA methylation. Nucleic Acids Res 2016; 44:5615-28. PMC4937303.
- Xu X, Tao Y, Gao X, Zhang L, Li X, Zou W, Ruan K, Wang F, Xu GL, Hu R. A CRISPR-based approach for targeted DNA demethylation. Cell Discov 2016; 2:16009. PMC4853773.
- Kim S, Koo T, Jee HG, Cho HY, Lee G, Lim DG, Shin HS, Kim JS. CRISPR RNAs trigger innate immune responses in human cells. Genome Res 2018; 28:367-73. PMC5848615.
- Ewaisha R, Anderson KS. Immunogenicity of CRISPR therapeutics-Critical considerations for clinical translation. Front Bioeng Biotechnol 2023; 11:1138596. PMC9978118.
- Hakim CH, Kumar SRP, Perez-Lopez DO, Wasala NB, Zhang D, Yue Y, Teixeira J, Pan X, Zhang K, Million ED, et al. Cas9-specific immune responses compromise local and systemic AAV CRISPR therapy in multiple dystrophic canine models. Nat Commun 2021; 12:6769. PMC8613397.
- Zhang XH, Tee LY, Wang XG, Huang QS, Yang SH. Off-target Effects in CRISPR/Cas9-mediated Genome Engineering. Mol Ther Nucleic Acids 2015; 4:e264. PMC4877446.
- Hoijer I, Emmanouilidou A, Ostlund R, van Schendel R, Bozorgpana S, Tijsterman M, Feuk L, Gyllensten U, den Hoed M, Ameur A. CRISPR-Cas9 induces large structural variants at on-target and off-target sites in vivo that segregate across generations. Nat Commun 2022; 13:627. PMC8810904.
- Frangoul H, Altshuler D, Cappellini MD, Chen YS, Domm J, Eustace BK, Foell J, de la Fuente J, Grupp S, Handgretinger R, et al. CRISPR-Cas9 Gene Editing for Sickle Cell Disease and beta-Thalassemia. N Engl J Med 2021; 384:252-60.
- Stadtmauer EA, Fraietta JA, Davis MM, Cohen AD, Weber KL, Lancaster E, Mangan PA, Kulikovskaya I, Gupta M, Chen F, et al. CRISPR-engineered T cells in patients with refractory cancer. Science 2020; 367. PMC11249135.
- Scammell JG, Denny WB, Valentine DL, Smith DF. Overexpression of the FK506-binding immunophilin FKBP51 is the common cause of glucocorticoid resistance in three New World primates. Gen Comp Endocrinol 2001; 124:152-65.
- Nelson JC, Davis JM. DST studies in psychotic depression: a meta-analysis. Am J Psychiatry 1997; 154:1497-503.
- Binder EB, Salyakina D, Lichtner P, Wochnik GM, Ising M, Putz B, Papiol S, Seaman S, Lucae S, Kohli MA, et al. Polymorphisms in FKBP5 are associated with increased recurrence of depressive episodes and rapid response to antidepressant treatment. Nat Genet 2004; 36:1319-25.
- Tozzi L, Farrell C, Booij L, Doolin K, Nemoda Z, Szyf M, Pomares FB, Chiarella J, O'Keane V, Frodl T. Epigenetic Changes of FKBP5 as a Link Connecting Genetic and Environmental Risk Factors with Structural and Functional Brain Changes in Major Depression. Neuropsychopharmacology 2018; 43:1138-45. PMC5854813.
- Lee RS, Tamashiro KL, Yang X, Purcell RH, Harvey A, Willour VL, Huo Y, Rongione M, Wand GS, Potash JB. Chronic corticosterone exposure increases expression and decreases deoxyribonucleic acid methylation of Fkbp5 in mice. Endocrinology 2010; 151:4332-43. PMC2940504. [CrossRef]
- Cox OH, Song HY, Garrison-Desany HM, Gadiwalla N, Carey JL, Menzies J, Lee RS. Characterization of glucocorticoid-induced loss of DNA methylation of the stress-response gene Fkbp5 in neuronal cells. Epigenetics 2021; 16:1377-97. PMC8813076.
- Wochnik GM, Ruegg J, Abel GA, Schmidt U, Holsboer F, Rein T. FK506-binding proteins 51 and 52 differentially regulate dynein interaction and nuclear translocation of the glucocorticoid receptor in mammalian cells. J Biol Chem 2005; 280:4609-16.
- Hawn SE, Sheerin CM, Lind MJ, Hicks TA, Marraccini ME, Bountress K, Bacanu SA, Nugent NR, Amstadter AB. GxE effects of FKBP5 and traumatic life events on PTSD: A meta-analysis. J Affect Disord 2019; 243:455-62. PMC6487483. [CrossRef]
- Qiu B, Luczak SE, Wall TL, Kirchhoff AM, Xu Y, Eng MY, Stewart RB, Shou W, Boehm SL, Chester JA, et al. The FKBP5 Gene Affects Alcohol Drinking in Knockout Mice and Is Implicated in Alcohol Drinking in Humans. Int J Mol Sci 2016; 17. PMC5000669.
- Shumay E, Logan J, Volkow ND, Fowler JS. Evidence that the methylation state of the monoamine oxidase A (MAOA) gene predicts brain activity of MAO A enzyme in healthy men. Epigenetics 2012; 7:1151-60. PMC3469457.
- Renbaum P, Abrahamove D, Fainsod A, Wilson GG, Rottem S, Razin A. Cloning, characterization, and expression in Escherichia coli of the gene coding for the CpG DNA methylase from Spiroplasma sp. strain MQ1(M.SssI). Nucleic Acids Res 1990; 18:1145-52. PMC330428.
- Duis J, Cox OH, Ji Y, Seifuddin F, Lee RS, Wang X. Effect of Genotype and Maternal Affective Disorder on Intronic Methylation of FK506 Binding Protein 5 in Cord Blood DNA. Front Genet 2018; 9:648. PMC6305129.
- Cox OH, Song HY, Garrison-Desany HM, Gadiwalla N, Carey JL, Menzies J, Lee RS. Characterization of glucocorticoid-induced loss of DNA methylation of the stress-response gene Fkbp5 in neuronal cells. Epigenetics 2021:1-21. [CrossRef]
- Seifuddin F, Wand G, Cox O, Pirooznia M, Moody L, Yang X, Tai J, Boersma G, Tamashiro K, Zandi P, et al. Genome-wide Methyl-Seq analysis of blood-brain targets of glucocorticoid exposure. Epigenetics 2017; 12:637-52. PMC5687336. [CrossRef]
- Cox OH, Seifuddin F, Guo J, Pirooznia M, Boersma GJ, Wang J, Tamashiro KLK, Lee RS. Implementation of the Methyl-Seq platform to identify tissue- and sex-specific DNA methylation differences in the rat epigenome. Epigenetics 2024; 19:2393945. PMC11418217. [CrossRef]
- Murphy DE, de Jong OG, Brouwer M, Wood MJ, Lavieu G, Schiffelers RM, Vader P. Extracellular vesicle-based therapeutics: natural versus engineered targeting and trafficking. Exp Mol Med 2019; 51:1-12. PMC6418170.
Figure 1.
Genomic organization of FKBP5. The human FKBP5 locus is located on the negative strand of chromosome 6. For this project, three intronic regions have been tested, and these are indicated by thin horizontal gray lines. Two sets of large black arrows represent the outside and nested PCR primers used for bisulfite pyrosequencing. These black arrows flank the primers used for generating the probe (red arrows) which cannot be amplified by the pyrosequencing primers. The intron 5 GREs are composed of the main GRE formed by GRE1 and GRE2 for which the DNA probe was designed and an adjacent GRE that was tested as a negative control region.
Figure 1.
Genomic organization of FKBP5. The human FKBP5 locus is located on the negative strand of chromosome 6. For this project, three intronic regions have been tested, and these are indicated by thin horizontal gray lines. Two sets of large black arrows represent the outside and nested PCR primers used for bisulfite pyrosequencing. These black arrows flank the primers used for generating the probe (red arrows) which cannot be amplified by the pyrosequencing primers. The intron 5 GREs are composed of the main GRE formed by GRE1 and GRE2 for which the DNA probe was designed and an adjacent GRE that was tested as a negative control region.
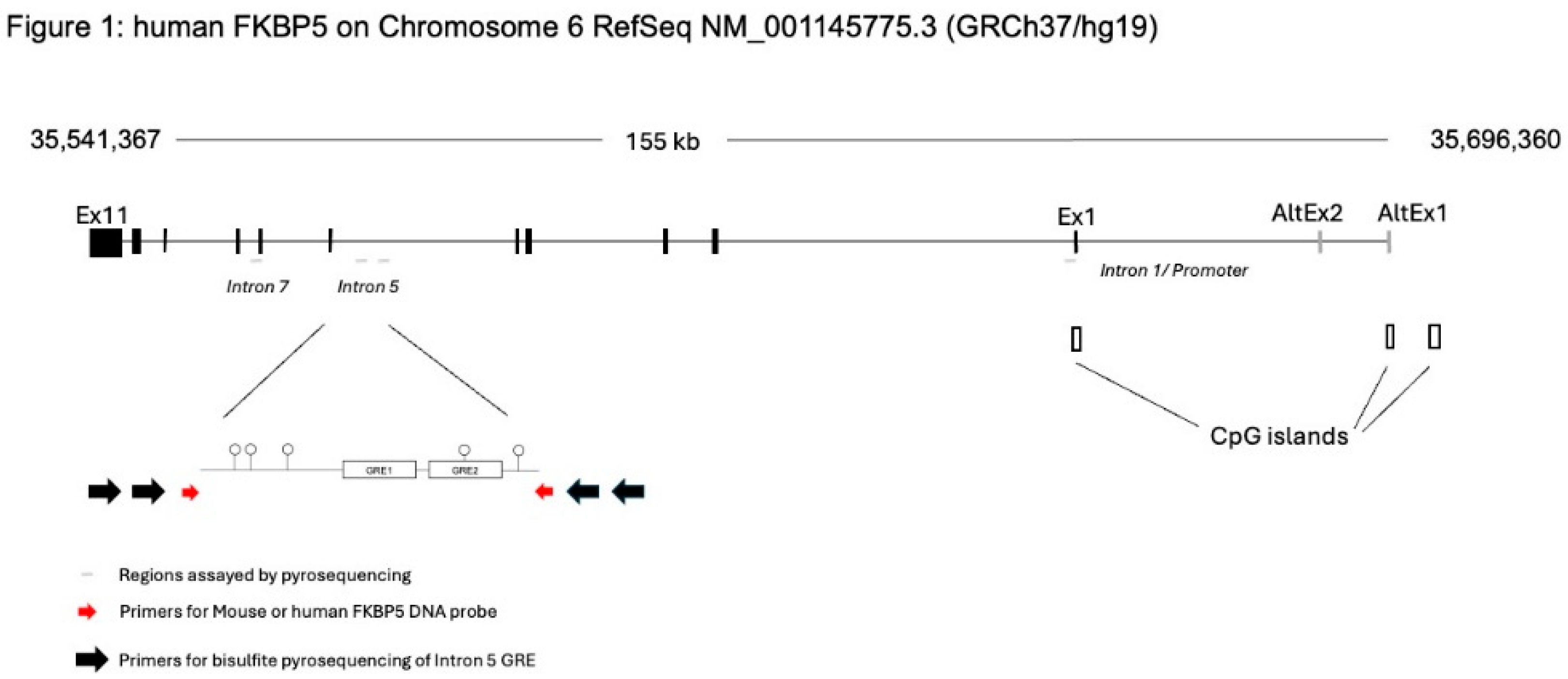
Figure 2.
Dose-dependent DNAm and gene expression changes following transfection of methylated DNA probes against FKBP5. (A) 293HEK cells were transfected with single-stranded unmethylated probe (500 ng -SssI), single-stranded methylated probe at two different concentrations (250 ng and 500 ng +SssI), or double-stranded methylated probe (500 ng +SssI +DS). Untransfected cells served as controls (Control). DNAm levels of five CpG sites at the conserved glucocorticoid response element (GRE) of human FKBP5 intron 5 were analyzed by bisulfite pyrosequencing. Data for the first two CpGs are shown. (B) Typical pyrograms obtained from bisulfite pyrosequencing is shown for each group at CpG-1. The percent DNAm determination occurs when R (or A/G) is dispensed, and it corresponds to the reverse complement of T/C (T for unmethylated and C for methylation CpG, respectively). (C) FKBP5 expression was measured by qRT-PCR in the same groups of 293HEK cells as in (A) treated with 1 μM dexamethasone (DEX) for four hours prior to collection. Bar graphs represent mean ± SEM from samples processed in triplicate. ***P<0.001, **P<0.01, and *P<0.05.
Figure 2.
Dose-dependent DNAm and gene expression changes following transfection of methylated DNA probes against FKBP5. (A) 293HEK cells were transfected with single-stranded unmethylated probe (500 ng -SssI), single-stranded methylated probe at two different concentrations (250 ng and 500 ng +SssI), or double-stranded methylated probe (500 ng +SssI +DS). Untransfected cells served as controls (Control). DNAm levels of five CpG sites at the conserved glucocorticoid response element (GRE) of human FKBP5 intron 5 were analyzed by bisulfite pyrosequencing. Data for the first two CpGs are shown. (B) Typical pyrograms obtained from bisulfite pyrosequencing is shown for each group at CpG-1. The percent DNAm determination occurs when R (or A/G) is dispensed, and it corresponds to the reverse complement of T/C (T for unmethylated and C for methylation CpG, respectively). (C) FKBP5 expression was measured by qRT-PCR in the same groups of 293HEK cells as in (A) treated with 1 μM dexamethasone (DEX) for four hours prior to collection. Bar graphs represent mean ± SEM from samples processed in triplicate. ***P<0.001, **P<0.01, and *P<0.05.
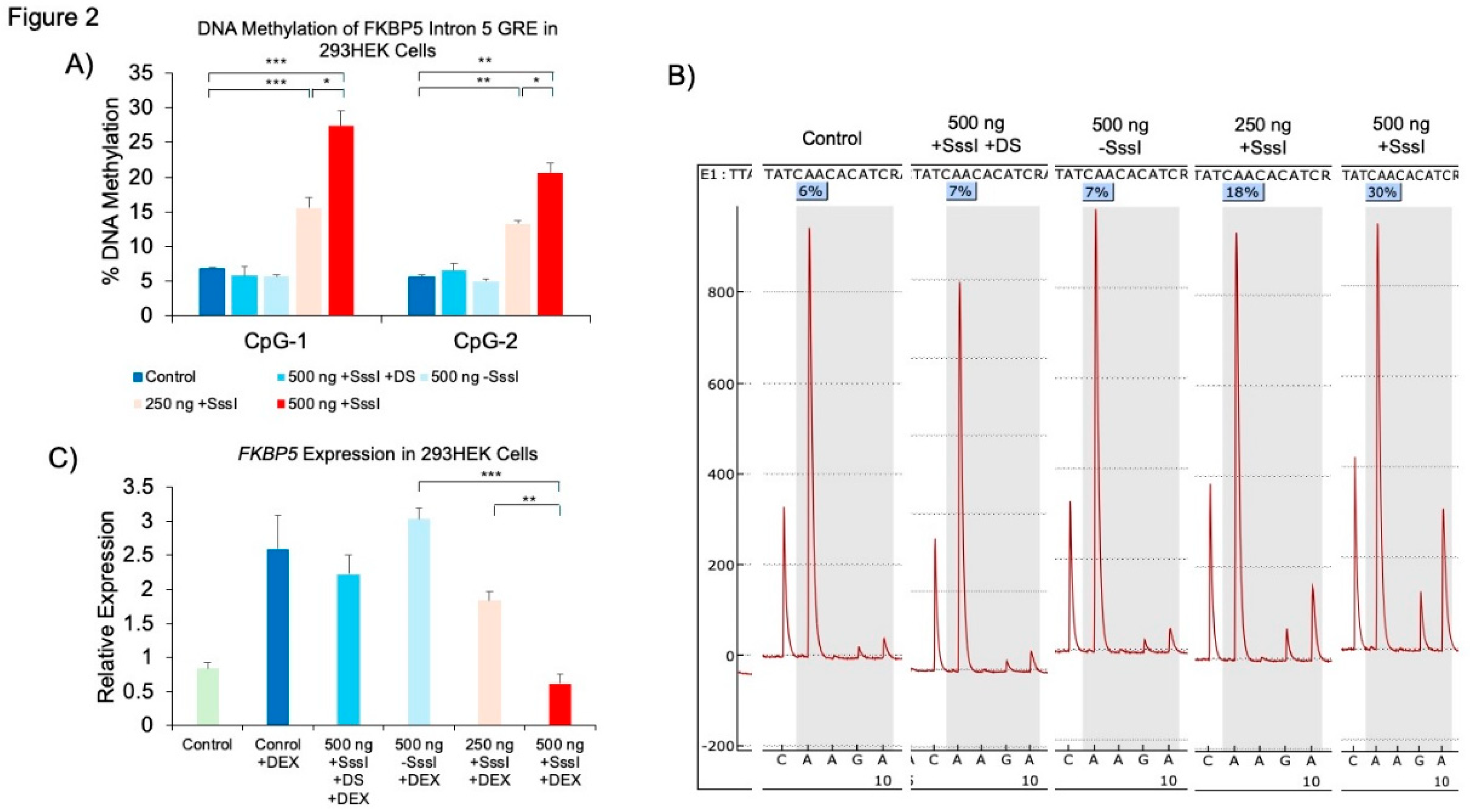
Figure 3.
Persistence and accumulation of DNA methylation. (A) 293HEK cells were transfected with the unmethylated (-SssI) and methylated (+SssI) FKBP5 DNA probe at Week 1 (W1), after which the cells were cultured for an additional four weeks (W4) before analysis by bisulfite pyrosequencing. (B) 293HEK cells were transfected with unmethylated (-SssI) and methylated (+SssI) FKBP5 DNA probe consecutively every three days and expanded, while 50% of the cells were collected for analysis. ***P<0.001, **P<0.01, and *P<0.05.
Figure 3.
Persistence and accumulation of DNA methylation. (A) 293HEK cells were transfected with the unmethylated (-SssI) and methylated (+SssI) FKBP5 DNA probe at Week 1 (W1), after which the cells were cultured for an additional four weeks (W4) before analysis by bisulfite pyrosequencing. (B) 293HEK cells were transfected with unmethylated (-SssI) and methylated (+SssI) FKBP5 DNA probe consecutively every three days and expanded, while 50% of the cells were collected for analysis. ***P<0.001, **P<0.01, and *P<0.05.
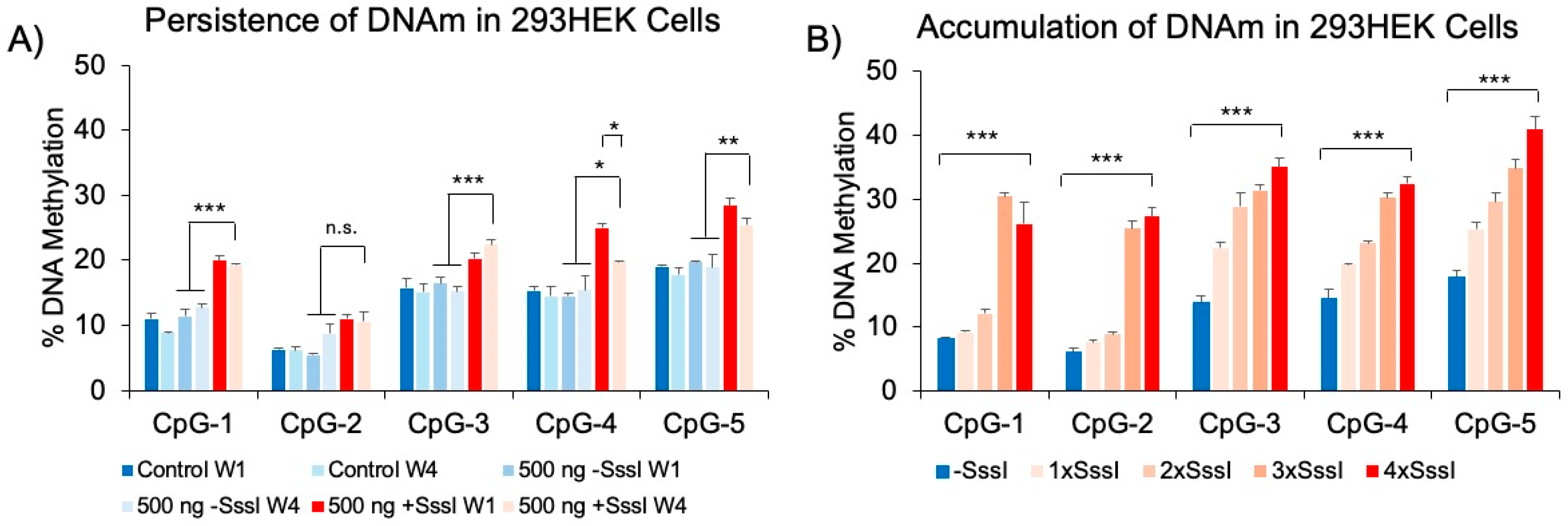
Figure 4.
DNAm analysis of human FKBP5 GREs at additional intronic regions. Experimentally verified glucocorticoid response elements (GREs) at introns 1, 5, and 7 were evaluated in probe-transfected 293 HEK samples for non-specific epigenetic effects. There were no significant differences in cells transfected with unmethylated vs. methylated DNA probes. **P<0.01.
Figure 4.
DNAm analysis of human FKBP5 GREs at additional intronic regions. Experimentally verified glucocorticoid response elements (GREs) at introns 1, 5, and 7 were evaluated in probe-transfected 293 HEK samples for non-specific epigenetic effects. There were no significant differences in cells transfected with unmethylated vs. methylated DNA probes. **P<0.01.
Figure 5.
DNAm and gene expression analysis following transfection of methylated DNA probes against mouse Fkbp5. (A) AtT-20 cells were transfected with single-stranded unmethylated probe (500 ng -SssI), single-stranded methylated probe (500 ng +SssI), or double-stranded methylated probe (500 ng +SssI +DS). Untransfected cells served as controls (Control). DNAm levels of four CpG sites at the conserved glucocorticoid response element (GRE) of human FKBP5 intron 5 were analyzed by bisulfite pyrosequencing. Data for all four GRE CpGs are shown. (B) Fkbp5 expression was measured by qRT-PCR in the same groups of AtT-20 cells as in (A) treated with 1 μM dexamethasone (DEX) for four hours prior to collection. Bar graphs represent mean ± SEM from samples processed in triplicate. ***P<0.001, **P<0.01, and *P<0.05.
Figure 5.
DNAm and gene expression analysis following transfection of methylated DNA probes against mouse Fkbp5. (A) AtT-20 cells were transfected with single-stranded unmethylated probe (500 ng -SssI), single-stranded methylated probe (500 ng +SssI), or double-stranded methylated probe (500 ng +SssI +DS). Untransfected cells served as controls (Control). DNAm levels of four CpG sites at the conserved glucocorticoid response element (GRE) of human FKBP5 intron 5 were analyzed by bisulfite pyrosequencing. Data for all four GRE CpGs are shown. (B) Fkbp5 expression was measured by qRT-PCR in the same groups of AtT-20 cells as in (A) treated with 1 μM dexamethasone (DEX) for four hours prior to collection. Bar graphs represent mean ± SEM from samples processed in triplicate. ***P<0.001, **P<0.01, and *P<0.05.
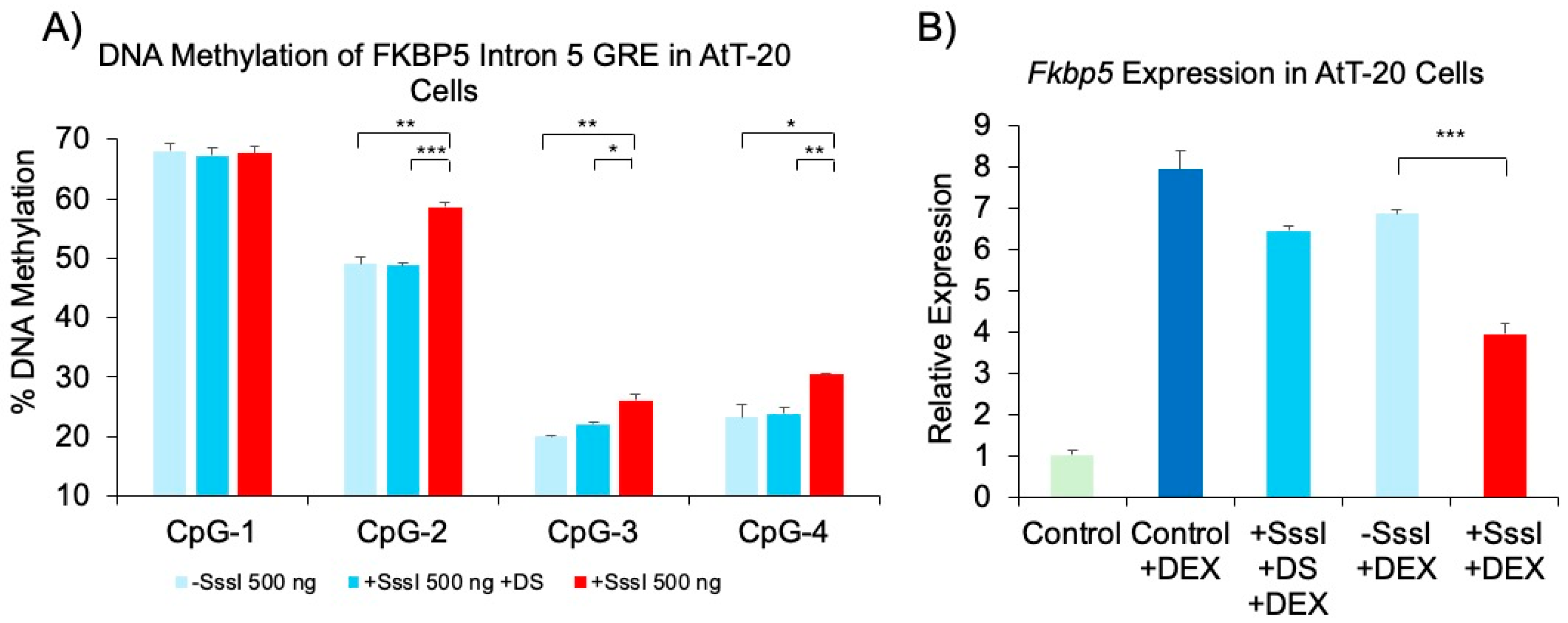
Figure 6.
DNAm and gene expression analysis following transfection of methylated DNA probes against MAOA. (A) 293HEK cells were transfected with single-stranded unmethylated probe (-SssI) or single-stranded methylated probe (+SssI). DNAm levels of 14 CpG sites at a regulatory region of human MAOA were analyzed by bisulfite pyrosequencing. (B) MAOA expression was measured by qRT-PCR in the same groups of 293HEK cells. Bar graphs represent mean ± SEM from samples processed in triplicate. *P<0.05 and **P<0.01.
Figure 6.
DNAm and gene expression analysis following transfection of methylated DNA probes against MAOA. (A) 293HEK cells were transfected with single-stranded unmethylated probe (-SssI) or single-stranded methylated probe (+SssI). DNAm levels of 14 CpG sites at a regulatory region of human MAOA were analyzed by bisulfite pyrosequencing. (B) MAOA expression was measured by qRT-PCR in the same groups of 293HEK cells. Bar graphs represent mean ± SEM from samples processed in triplicate. *P<0.05 and **P<0.01.
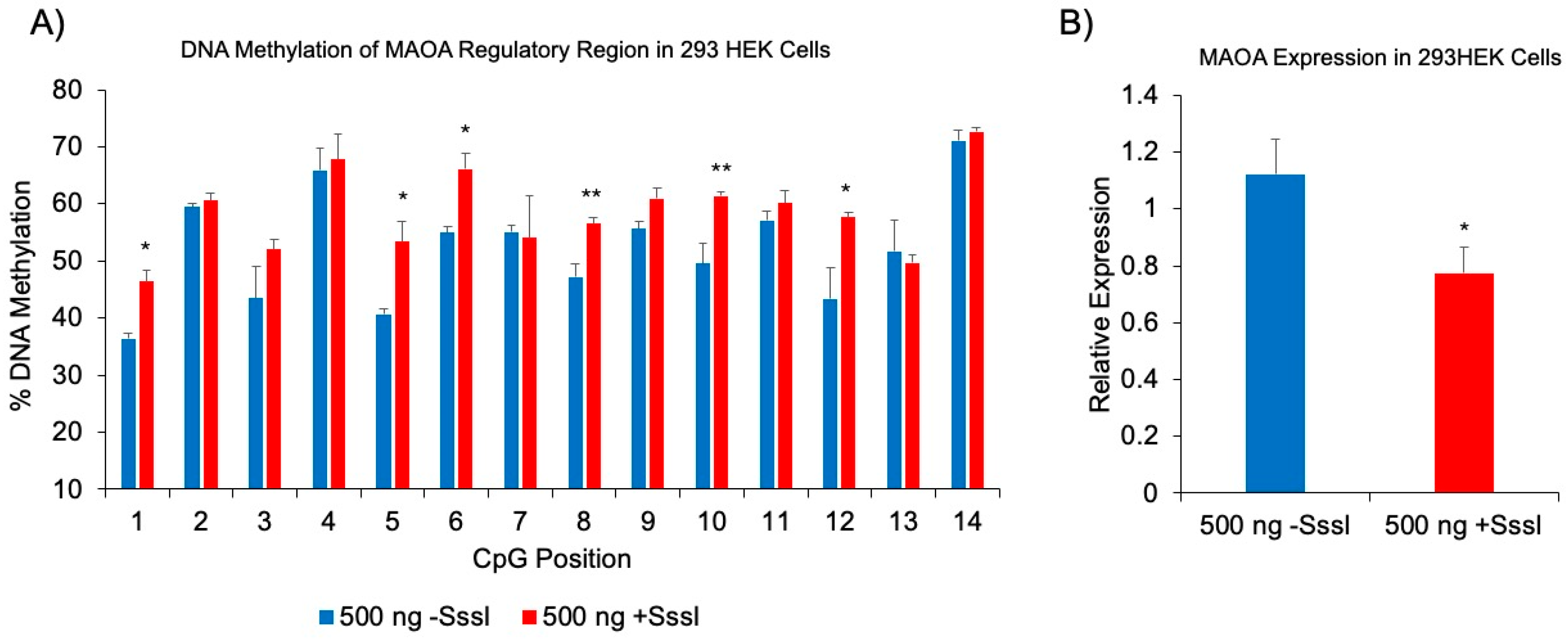

Table 1.
Sequence of primers for Probe Design and Pyrosequencing.
| Probe Primers | Sequence | Size |
| Human FKBP5 Probe – Forward | 5’- AAAGTCAAACCAAACCAAATTACC -3’ | 256 bp |
| Human FKBP5 Probe – Reverse | 5’- TTTGTTACTGCTGTGCACTCTCT -3’ | |
| Human MAOA Probe – Forward | 5’- TCGACGTAGTCGTGATCGG -3’ | 170 bp |
| Human MAOA Probe – Reverse | 5’- GCAGGATATGGGGCCAAG -3’ | |
| Mouse Fkbp5 Probe – Forward | 5’- CAGACACCAGCTACTATAATTAG -3’ | 239 bp |
| Mouse Fkbp5 Probe – Reverse | 5’- GCACATGAACTCGATGTGCTGACA -3’ | |
| Pyrosequencing Primers | Sequence | Size |
| Human FKBP5 Intron 5 Outside – A | GGTAGAGAAAGAAATAAATAAGTTA |
286 bp |
| Human FKBP5 Intron 5 Outside – B | TTCTTACATTTCATTTTTATTACTACTA | |
| Human FKBP5 Intron 5 Inside – A* | AAGATTATGTAATTTAAAGGGGGAGGG | |
| Human FKBP5 Intron 5 Inside – B | CTCTCTTTCCTTTTTTCCCCCCTAT | |
| Human FKBP5 Intron 5 Pyro 1 | TCTTTCCTTTTTTCCCCCCTATT | |
| Human FKBP5 Intron 5 Pyro 2 | CAATTTAAATAATATTTTACAACT | |
| Human FKBP5 Intron 5_2 Out – A | ATTTAATTGGTTTGGGTGTTAGAA |
406 bp |
| Human FKBP5 Intron 5_2 Out – B | CCTCTCAATACTTTCAACCACA | |
| Human FKBP5 Intron 5_2 In – A* | GAGAATTATTGTATTGGAGGTT | |
| Human FKBP5 Intron 5_2 In – B | ATTCTACAAATTCCAATTATTAAC | |
| Human FKBP5 Intron 5_2 Pyro 1 | GTATTGGAGGTTTATTGGTT | |
| Human FKBP5 Intron 5_2 Pyro 2 | TAGATGATTATGAGTTTGGAGTT | |
| Human FKBP5 Intron 5_2 Pyro 3 | GTTTAAGTTTTTTTTATATTTGTT | |
| Human FKBP5 Intron 5_2 Pyro 4 | GATTTGGAGAGGGAAAGGAGGT | |
| Human FKBP5 Intron 1 Out – A | AGTTTAAATTGTTTTATGTAGAATTTATTGA |
350 bp |
| Human FKBP5 Intron 1 Out – B | TCACTCCCAAACCATACC | |
| Human FKBP5 Intron 1 Inside – A | GTTTTGAATTATATTGAAGGGTATTT | |
| Human FKBP5 Intron 1 Inside – B* | CAAAACTCCTTATACTCTTCTATTCTAA | |
| Human FKBP5 Intron 1 Pyro 1 | GTAGAATTYGATTTTAGAGA | |
| Human FKBP5 Intron 7 Outside – A | AGAGTGAAATTGAGATGGAAATATGT |
503 bp |
| Human FKBP5 Intron 7 Outside – B | AATTTCTTCTCCATCCACTTCCTATA | |
| Human FKBP5 Intron 7 Inside – A | AGGAGGTATGTTGTTTTTGGAATTTAAG | |
| Human FKBP5 Intron 7 Inside – B* | AATTTATCTCTTACCTCCAACACT | |
| Human FKBP5 Intron 7 Pyro 1 | GGAGAAGTATAAAAAAAAAATGG | |
| Human FKBP5 Intron 7 Pyro 2 | GTTATAGAGTTTAGTGGTTT | |
| Human FKBP5 Intron 7 Pyro 3 | GGAGTTATAGTGTAGGTTTT | |
| Human FKBP5 Intron 7 Pyro 4 | TTAAGGAGTTATTTGGTAGA | |
| Human FKBP5 Intron 7 Pyro 5 | TGATATATAGGAATAAAATAAGAAT | |
| Human MAOA Outside – A | GATTTAGGAGYGTGTTAGTTAAAGT |
278 bp |
| Human MAOA Outside – B | TTATTATATCTACCTCCCCCAATC | |
| Human MAOA Inside – A | AGTTAAAGTATGGAGAATTAAG | |
| Human MAOA Inside – B* | ATCTACCTCCCCCAATCACACCACCAAC | |
| Human MAOA Pyro 1 | AAAGTATGGAGAATTAAGAGAAGG | |
| Human MAOA Pyro 2 | GAGTATYGYGGGTTATATG | |
| Human MAOA Pyro 3 | AGGTGGTATTTTAGGTTAGTGTGGA | |
| Mouse Fkbp5 Intron 5 Outside – A | GATGATTAGTTTTTTTTAGTAGTGATGT |
308 bp |
| Mouse Fkbp5 Intron 5 Outside – B | CTTATTATTCTCTTACTACCCTAA | |
| Mouse Fkbp5 Intron 5 Inside – A | TAGTTTTTGGGGAAGAGTGTAGAGTTAT | |
| Mouse Fkbp5 Intron 5 Inside – B* | ATTTTAAAAAACACAAAACACCCTATT | |
| Mouse Fkbp5 Intron 5 Pyro 1 | AGAAAAGGGAAAGTAGG | |
| Mouse Fkbp5 Intron 5 Pyro 2 | TAGTTTTTGTTATTGTTGTATG |
*These primers have been biotinylated and HPLC-purified for pyrosequencing.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



