
Submitted:
06 April 2025
Posted:
08 April 2025
You are already at the latest version
Abstract
Yoga demonstrates systemic physiological benefits, but its molecular mechanisms remain unclear. This multi-omics study explores transcriptomic and immune dynamics following yoga intervention. Analyzing the GSE44777 dataset, we identified 1,260 differentially expressed genes enriched in lipid metabolism, atherosclerosis, and cancer pathways. Protein-protein interaction networks revealed HBEGF, SQLE, and BCL6 as immune-metabolic hubs. A nomogram incorporating HBEGF/SQLE predicted neuropathic pain with high accuracy (AUC = 0.831). Yoga significantly reduced neutrophil infiltration (p < 0.001) and enhanced monocyte-CD4+ T cell coordination. HBEGF emerged as a dual-functional hub linking antibacterial immunity to inflammasome activation. Competitive endogenous RNA (ceRNA) network analysis uncovered HBEGF regulation by hsa-miR-29a-3p and lncRNAs, suggesting post-transcriptional control. These findings elucidate yoga’s role in reprogramming immune-metabolic networks through transcriptional and ceRNA-mediated mechanisms, supporting its application in precision prevention of cardiovascular diseases, cancer, and neuropathic pain.
Keywords:
Yoga
; Transcriptomic reprogramming
; Immune infiltration
; ceRNA network
; Disease prevention
1. Introduction
Yoga, an ancient mind-body practice that blends asanas, pranayama, and dhyana, has gained global recognition for its positive impact on health. Growing evidence highlights its role in improving cardiovascular health [1], reducing psychological stress [2,3], boosting immune function [4], enhancing cognitive health, and reducing chronic inflammation [5,6]. Systematic studies also confirm its effectiveness in managing metabolic syndrome [7], neurological disorders [2], and age-related frailty [8]. Unfortunately, despite these well-documented benefits, the molecular mechanisms mediating these pleiotropic effects—particularly those involving epigenetic remodeling, transcriptomic plasticity, and immunocellular communication—remain enigmatic, substantially constraining evidence-based optimization of yogic interventions for precision medicine applications.
Current research predominantly focuses on phenotypic outcomes, with limited exploration of yoga’s impact on gene expression dynamics or immune cell interactions. While studies have shown that yoga can influence inflammatory biomarkers [6,9] and stress hormones [10], its impact on broader transcriptional networks or epigenetic changes has not been fully explored. For example, although yoga is associated with lower cardiovascular risk [1], its potential to affect genes related to atherosclerosis or lipid metabolism through epigenetic mechanisms hasn’t been thoroughly investigated [7]. Similarly, despite yoga is identified to have immunomodulatory effects in cancer survivors [11], its ability to alter immune cell infiltration patterns or regulate cytokine pathways involved in cancer promotion is still unclear. Moreover, the association between yoga-induced gene expression changes and non-coding RNA networks—a key part of epigenetic regulation—has yet to be studied [12], leaving a significant gap in our understanding of its long-term health benefits.
This study utilizes the GSE44777 transcriptomic dataset to explore the molecular reprogramming caused by yoga. We hypothesize that yoga triggers coordinated gene expression changes in immune-related pathways, facilitated by hub genes and ceRNA networks. By combining differentially expressed genes (DEGs) analysis, Kyoto Encyclopedia of Genes and Genomes (KEGG) / Gene Ontology (GO) enrichment, and immune infiltration profiling, we aim to: (1) identify signature DEGs and pathways influenced by yoga; (2) build PPI and ceRNA networks to identify central regulators; and (3) validate yoga’s ability to predict neuropathic pain risk using receiver operating characteristic (ROC)-based nomograms. This study employed systematic biology methods to unravel how yoga regulates both gene and immune pathways, providing valuable insights into its therapeutic potential for cardiovascular diseases, cancer, and chronic pain.
2. Results
2.1. Differential Analysis of GSE44777 Between Yoga Pre- and Post-Intervention
To study the differential gene expression between pre- and post-yoga intervention, we exploited mRNA expression profiles sourced from the GEO database. In our study, we identified amongst the 1260 differentially expressed genes, 574 were upregulated after (log2FC > 1) and 686 were downregulated after yoga intervention (log2FC < -1). The volcano plot in Figure 2A reveals these differentially expressed genes, in which the orange-colored and blue-colored points indicate the upregulated and downregulated genes, respectively. This volcano plot visualizes the expression changes of these genes and the significance of their changes by showing log2FC on the x-axis and -log10(p-value) on the y-axis. In parallel, the heatmap in Figure 2B shows expression levels of these genes across different samples. Here, each row in the heatmap represents a gene while each column represents a sample, with color changing from blue to red showing a transition in expression levels from low to high. From this, we can aim to investigate which genes show consistent expression changes before and after the yoga intervention, providing the foundations for further investigation into how the yoga intervention affects the subsequent biological processes.
2.2. Differential Gene Functional Enrichment Analysis Results
Gene set enrichment analysis aims at identifying expressed genes or proteins that relate to specific disease phenotypes by reference to common biological functions, hence chromosomal locations, or regulations within a genome. KEGG enrichment analysis showed significant enrichments in the following pathways: Lipid and Atherosclerosis, Colorectal Cancer, Small Cell Lung Cancer, Basal Cell Carcinoma, Kaposi Sarcoma-Associated Herpesvirus Infection, Alcoholic Liver Disease, Legionellosis, Parathyroid Hormone Synthesis, Secretion and Action, Thyroid Cancer, and Estrogen Signaling Pathway. Notably, genes involved in Lipid and Atherosclerosis had shown a substantial level of overexpression, alluding to possible roles of yoga in promoting cardiovascular health in connection with lipid metabolism regulation and atherosclerosis prevention (Figure 3A). Basal Cell Carcinoma pathway enrichment suggests a possible role of yoga in skin cancer development and prevention, providing insights into yoga’s influence on skin health. An abundance of gene overexpression regarding Colorectal Cancer was observed in the biological processes associated with yoga, indicating a possible role of yoga in maintaining intestinal health and preventing such cancers (Figure 3B). Meanwhile, GO enrichment analysis has shown obvious enrichment in pathways like regulation of cold-induced thermogenesis, adaptive thermogenesis, and phagocytosis, presumably offering various potential roles of yoga in the physiological regulation of responses to cold environments, and possibly modulating immune functions favorably, such as cell engulfment and clearance of waste and pathogens. Such findings offer extensive insight into the prospective yoga influences on different biological processes related to disease prevention.
2.3. Signature Gene PPI Network Analysis
We employed the STRING online tool to construct a Protein-Protein Interaction (PPI) network (Figure 4A) to investigate the biological significance of gene expression changes before and after yoga intervention. Through this network, we identified central genes upregulated after yoga intervention and visualized them using Cytoscape software. In our analysis, we particularly focused on the top ten upregulated genes, which may play crucial roles in the PPI network. These genes include: HBEGF, LDLR, SCD, BCL6, SQLE, BTC, TGFA, MYLIP, RXRA, CDK19. In Figure 4B, we observe a PPI network constructed through Cytoscape software, emphasizing the ten central genes upregulated after yoga intervention. These genes are considered to play roles in cell signaling, metabolic processes, and potential disease mechanisms through their interactions.
2.4. Co-Expression Analysis of Key Genes in Yoga and Construction of Risk Prediction Line Graph Model
The correlation analysis done through the scatter plot and correlation matrix in Figure 5A aimed to see the correlation between different biomarkers. The figure indicates a correlation of, for example, 0.64 between BCL6 and TGFA. Given that yoga can affect biomarker expression, the levels of expression of BCL6 and TGFA might be tending to go simultaneously high or low.
Figure 5B displayed different prediction probabilities, demonstrating the model’s excellent performance in predicting yoga potential. We calculated the ROC curves for five central genes (HBEGF, BCL6, BNIP3, SCD, and SQLE) to assess their diagnostic efficacy in neuropathic pain. The Area Under the Curve (AUC) of the ROC curve is a crucial indicator for judging diagnostic performance, with higher AUC values indicating better diagnostic efficacy. In our model, the AUC values for these genes were 0.831 (HBEGF and SQLE), 0.748 (BCL6), 0.762 (BNIP3), and 0.760 (SCD), respectively.
We also built a column line-chart model to predict the influence of yoga potential as shown in Figure 5C. The expression levels of each different biomarker were converted into scores that indicate high or low expression. High expression of HBEGF means a high score, while low expression of BCL6 means a low score. All these scores were added up to produce the final score for predicting yoga potential.
2.5. Gene Set Enrichment Analysis (GSEA) for Core Genes
As shown in Figure 6A (GSEA-KEGG analysis), in high HBEGF-expressing conditions, significant enrichment was observed in acute myeloid leukemia, natural killer (NK) cell-mediated cytotoxicity, and NOD-like receptor signaling pathways, suggesting its potential mechanistic roles in: (1) activating leukemia cell proliferation through RAS/MAPK signaling; (2) enhancing NK cell cytotoxic activity via upregulation of granzyme and perforin expression; and (3) promoting NLRP3 inflammasome assembly to drive pro-inflammatory cytokine release. In contrast, low HBEGF expression exhibited significant negative correlations with Leishmania infection response and actin cytoskeleton regulation pathways. These findings imply that HBEGF downregulation may impair host anti-parasitic immune responses (e.g., suppressed macrophage phagocytic function) and disrupt cellular migration by interfering with actin polymerization dynamics.
Further analysis of GSEA-GO data identified bidirectional regulatory relationships between HBEGF expression and pathways linked to immune defense and metabolic homeostasis. Under high HBEGF expression, strong enrichment was detected in antibacterial humoral immunity pathways, including complement activation and defense response to Gram-negative bacteria, indicating its role in enhancing pathogen clearance through mechanisms such as B cell antibody class-switching or neutrophil extracellular trap formation. Conversely, low HBEGF expression showed negative enrichment in ribosome biogenesis pathways, suggesting that reduced HBEGF levels may suppress RNA polymerase I activity and ribosomal protein synthesis, leading to cell cycle arrest and diminished proliferative capacity.
Collectively, these results demonstrate that HBEGF functions as a dual-regulatory hub molecule, dynamically modulating pathways critical to infection immunity, tumorigenesis, and metabolic regulation.
2.6. Yoga Immunomodulation Results
The study investigated the effects of yoga on immune cell infiltration and identified distinct gene-immune cell correlation patterns (Figure 7A). TSC22D3 and TGFA exhibited strong positive associations with neutrophils (p < 0.001), while SQLE and BCL6 correlated positively with monocytes and resting memory CD4+ T cells. Notably, TGFA and HBEGF showed significant negative correlations with resting NK cells (p < 0.05). Genes including BNIP3, BTC, and LDLR were inversely associated with CD4+ naïve T cells, CD8+ T cells, and γδ T cells.
As demonstrated in Figure 7B, yoga intervention modulated immune homeostasis through multidimensional remodeling of immune cell interactions: it attenuated pro-inflammatory responses by suppressing the positive correlation between neutrophils and resting mast cells while enhancing the negative correlation between resting NK cells and neutrophils, thereby balancing inflammation and immune surveillance. Concurrently, yoga promoted Th1/CTL response coordination via strong positive correlations between activated CD4+ memory T cells and CD8+ T cells/resting NK cells, likely amplifying anti-tumor/anti-viral activity through IFN-γ signaling. Additionally, naïve B cell associations with M2 macrophages and resting NK cells supported anti-inflammatory/pro-repair phenotypic shifts, whereas the negative monocyte-CD8+ T cell correlation limited inflammatory monocyte infiltration to prevent exhaustion. These mechanisms collectively suggest that yoga synergistically suppresses pathological inflammation, enhances effector immunity, and sustains naïve-effector cell equilibrium.
2.8. Construction of mRNA-miRNA-lncRNA ceRNA Network
Long non-coding RNAs (lncRNAs) act as competitive endogenous RNAs in the ceRNA system, which influence the amount of microRNAs (miRNAs) available and thus indirectly affect the expression levels of messenger RNAs (mRNAs). This mechanism involves the binding of lncRNAs to miRNAs, reducing inhibition of miRNAs on target mRNAs (HBEGF may be an example of a target). HBEGF is a central node in this network, consistently interacting with top miRNAs including hsa-miR-29a-3p and hsa-miR-802, which are predicted to function to negatively regulate HBEGF. We also discovered that lncRNAs had interacted with HBEGF, which may be a molecular sponge of these miRNAs that can indirectly enhance the expression of HBEGF. Evidence of this regulatory pathway highlights an unprecedented potential effect of yoga on cellular biology, especially since it can act on immune and inflammatory processes. It might help in the design of therapeutic strategies acting on the HBEGF signaling pathway for diseases associated with regulation of its expression.
3. Discussion
Our genome-wide transcriptomic analysis revealed that yoga intervention significantly modulates the expression of 1,260 genes (574 upregulated, 686 downregulated). Pathway enrichment analysis identified prominent associations with lipid metabolism and atherosclerosis-related processes. Key DEGs including HBEGF, TGFA, and SQLE demonstrated significant associations with neuropathic pain risk through their involvement in neuroinflammatory regulation and immune homeostasis. The central hub gene HBEGF revealed its dynamic role in pain-immunity crosstalk, particularly via epigenetic mechanisms involving ceRNA networks that modulate miR-29a-3p/lncRNA interactions (AUC = 0.86). These findings advance conventional understanding of yoga’s therapeutic effects by systematically decoding its multi-omics regulatory axis encompassing “gene remodeling – immune-metabolic reprogramming – epigenetics.”
The impacts of yoga intervention at the transcriptome level were revealed through whole-genome expression analysis, which highlights DEGs in both groups, including lipid metabolism (LDLR, SCD), inflammatory regulation (HBEGF, TGFA) and cancer-related genes (SQLE, BCL6). Dysregulation of lipid metabolism and chronic inflammatory activation are recognized as key pathological drivers in cardiovascular disease progression. Yoga demonstrates multi-target metabolic regulation by reducing visceral fat and pro-inflammatory cytokines (Interleukin (IL)-6, IL-18, C-reactive protein (CRP)) [20], while enhancing antioxidant defenses to restore redox balance [21]. It upregulates adiponectin to improve vascular function via adenosine monophosphate-activated protein kinase / endothelial nitric oxide synthase activation and counteracts tumor necrosis factor-α-induced endothelial damage [22]. Neuroendocrine modulation through hypothalamic-pituitary-adrenal axis (HPA) suppression and β-endorphin elevation further inhibits nuclear respiratory factor-κB-mediated inflammation [23]. Notably, multimodal yoga (asanas/pranayama) limits animal models’ ability to replicate its holistic effects, leading to predominant proteomic/metabolomic research while gene regulatory networks remain less underscored. Through transcriptomic profiling of PBMCs, our study reveals yoga’s specific modulation of lipid metabolism-associated genes, providing novel genomic evidence for its anti-atherosclerotic mechanisms. Interestingly, dynamic GSEA uncovered functional heterogeneity of HBEGF across expression states—high expression activated acute myeloid leukemia pathways, whereas low expression suppressed ribosome biogenesis—aligning with yoga’s metabolic regulation at molecular resolution.
Increasing evidence indicates that diverse yoga and meditation modalities exert regulatory effects on gene expression through epigenetic mechanisms, primarily involving DNA methylation, histone modifications, and pathways associated with neuroplasticity. Empirical studies demonstrate that Hatha yoga reduces DNA methylation at promoter regions of pro-inflammatory genes (e.g., tumor necrosis factor (TNF)) in PBMCs [12], suggesting its potential in attenuating epigenetic aging. Mindfulness interventions downregulate HDAC 2/3/9 in PBMCs and reduce FKBP5 methylation in PTSD [24]. Long-term meditators exhibit lymphocyte DMRs in glucose/inflammatory genes (IL-6, CRP) and decelerated epigenetic aging (lower DNA methylation age vs. chronological age) [25]. However, current mechanistic investigations predominantly focus on DNA methylation dynamics [26], with limited exploration of histone modification cascades (e.g., HDAC alterations in MBSR) and non-coding RNA regulatory networks. This study proposes a novel ceRNA-mediated epigenetic regulatory axis underlying yoga’s therapeutic effect. As illustrated in Figure 8, HBEGF functions as a hub gene that competitively binds hsa-miR-29a-3p alongside lncRNAs, thereby indirectly upregulating its expression. This mechanism potentially explains yoga-mediated modulation of inflammatory and metabolic processes, including NLRP3 inflammasome activation and IL-1β secretion. The predicted lncRNA-miR-29a-3p-HBEGF axis provides new molecular targets for understanding yoga’s cross-disease regulatory capacity in conditions ranging from neuropathic pain to oncogenesis. Additionally, Verzili et al. highlighted yoga’s potential in mitigating neurodegenerative aging [27], our findings (long non-coding RNA-mediated epigenetic reprogramming) may provide a novel explanation for how yoga delays cellular senescence.
Yoga has been demonstrated to exert pleiotropic effects on immune homeostasis by modulating inflammatory cytokines (e.g., reduced IL-6, TNF-α, and CRP levels), remodeling immune cell balance (increased Treg cells/decreased Th17 cells) [4,6,28], and regulating the HPA axis and neurotransmitters [29]. These mechanisms underscore its therapeutic potential in chronic inflammatory diseases (e.g., rheumatoid arthritis) [28], adjunct cancer care [30], and maternal stress management [31,32]. Although prior systematic reviews (e.g., Yeun et al.) established yoga’s immunomodulatory effects through clinical phenotypes (cytokine levels, lymphocyte counts) [4], gene-immune microenvironment dynamics were not highlighted. Our immune infiltration analysis reveals a dual regulatory mechanism: (1) downregulation of pro-inflammatory genes with concomitant suppression of neutrophil infiltration, and (2) upregulation of metabolic genes, promoting monocyte-T cell balance and NK cell cytotoxicity (Figure 7A). Network analysis further demonstrates yoga’s capacity to remodel immune cell crosstalk, evidenced by diminished pro-inflammatory neutrophil-resting mast cell interactions and enhanced regulatory crosstalk between resting NK cells and neutrophils (Figure 7B). In addition, dissimilar to Voss et al., who emphasized neurophysiological pathways (e.g., vagal activation) in immune modulation [3], this research integrates transcriptomic and immune microenvironment profiling to identify yoga-specific activation of Th1/cytotoxic T lymphocyte coordination and IFN-γ signaling.
Previous public health research on yoga primarily highlights its cost-effectiveness and accessibility for improving mental health or cardiovascular risk factors [33,34]. However, the absence of molecular evidence has limited its integration into precision prevention frameworks. Our findings address this by identifying biomarker-driven pathways (e.g., HBEGF-mediated lipid-antigen presentation) and a neuropathic pain prediction model (AUC: 0.831) that align with WHO’s call for data-informed lifestyle interventions [35,36]. Unlike earlier studies that rely on subjective outcomes (e.g., self-reported stress reduction [3,37], our multi-omics approach provides objective, mechanistic support for yoga’s role in chronic disease prevention. For instance, the association between LDLR and atherosclerosis pathways offers a molecular rationale for yoga’s cardioprotective effects, surpassing observational claims in prior epidemiological studies [38,39].
The single intervention design restricts temporal resolution, precluding separation of acute from sustained transcriptional effects. Blood-derived omics data may miss tissue-specific mechanisms (e.g. hepatic lipid metabolism or neuroimmune interactions). Experimental confirmation (i.e., CRISPR/Cas9-engaged models) is needed to validate the cause-and-effect relationship of hub genes such as HBEGF, as translational musk data stay observational in nature. Moreover, the GSE44777 cohort is not demographically representative (e.g., a low representation of the ethnicity or comorbidities), and it needs multi-center validation to enhance the power of the results.
This study provides the multi-omics evidence linking yoga intervention to transcriptional reprogramming, immune-metabolic crosstalk, and epigenetic regulation. The mechanistic pathways offer a molecular foundation for integrating yoga into precision public health strategies, particularly for cardiovascular disease prevention, chronic pain management, and cancer risk reduction.
4. Materials and Methods
4.1. Acquisition of Yoga Data
The National Center for Biotechnology Information Gene Expression Omnibus (NCBI-GEO; https://www.ncbi.nlm.nih.gov/geo/) is a publicly accessible repository that stores high-throughput transcriptomic data, with submissions from research institutions around the world. To examine the molecular changes induced by yoga, we accessed the GSE44777 dataset from GEO [13,14], which contains genome-wide mRNA expression profiles generated via microarray platforms. The GSE44777 study used a crossover design with 14 initially recruited healthy male participants aged 18-50 years. Participants were required to have no chronic diseases and good mental health, as indicated by a General Health Questionnaire-28 score of more than 13. Four participants were excluded due to insufficient blood samples (n = 2) or poor RNA quality for further analysis (n = 2), leaving 10 eligible subjects. The yoga intervention involved two consecutive morning sessions, which included asana practice, pranayama breathing, and meditation. Blood samples (20 mL) were collected before and after each session. Peripheral blood mononuclear cells (PBMCs) were isolated using density gradient centrifugation, and RNA was extracted for whole transcriptome analysis using dual-channel microarray technology. The raw expression data were preprocessed, normalized, and log2 transformed by the original submitters to ensure comparability across samples.
4.2. Identification of DEGs
The R package limma was used for differential expression analysis using linear modelling and empirical Bayes methods to identify genes whose expression changes are statistically significant [15]. Genes with an adjusted p-value threshold of < 0.05 (false discovery rate-corrected) and an absolute log2 fold change > 0.58 (1.5-fold change) were considered DEGs. Hierarchical clustering patterns of DEGs for visualization were shown using the heatmap package, and genome-wide significance distributions were plotted to generate volcano plots through the ggplot2 package.
4.3. Differential Gene GO/KEGG Enrichment Analysis
KEGG-based functional enrichment analysis was performed to mechanistically decipher yoga-induced transcriptional reprogramming, employing this knowledge base for systematic gene function annotation and pathway mapping to identify key regulatory networks [16]. KEGG pathway enrichment was executed with the following criteria: Benjamini-Hochberg adjusted p-value < 0.05 and a minimum gene count threshold of 10 per pathway to ensure analytical robustness. Subsequently, GO analysis was executed using the clusterProfiler R package, systematically categorizing candidate genes into the three principal ontologies: biological processes, molecular functions, and cellular components. Parameter settings for the GO analysis included false discovery rate (FDR) < 0.1 and an absolute normalized enrichment score threshold ≥1.5, ensuring identification of high-confidence functional terms.
4.4. Hub Gene Selection via the PPI Network
The PPI network was predicted to use STRING v12.0, a publicly accessible database that combines input experimental data, co-expression data, and pathway data [16]. Only interactions with a confidence score of ≥ 0.7 and FDR < 0.05 were considered for analysis. After the network construction, Cytoscape v4.0.1 was used for visualization and refinement. Graph-based metrics (like degree and maximum clique centralities) were calculated with the CytoHubba plugin to identify hub genes. In the PPI network, the Degree algorithm in Cytoscape was used to rank key genes. Genes with a degree of ≥ 15, such as HBEGF, LDLR, and BCL6, were prioritized as core regulatory factors.
4.5. Construction of Nomograph Models
A nomogram prediction model for neuropathic pain risk stratification was developed using the ’rms’ R package from dataset GSE250152. Model performance was validated using Harrell’s concordance index, which quantifies the model’s ability to discriminate between high- and low-risk individuals. Furthermore, the diagnostic accuracy of candidate biomarkers was assessed through ROC curve analysis performed using the ’pROC’ R package. The area under the ROC curve AUC was calculated with AUC values ≥ 0.7 being accepted as representing acceptable discrimination.
4.6. Gene Set Enrichment Analysis
Gene Set Enrichment Analysis (GSEA) was performed on the differentially expressed hub genes in participants before and after yoga intervention to identify the associations between specific genes, signal transducing pathways, and/or interactions among signaling pathways with an adjusted p < 0.05 [17].
4.7. Immune Infiltration Analysis
To evaluate how yoga had influenced immune profiles of participants we applied the CIBERSORT method (a deconvolution approach for analyzing expression matrices of 22 human immune cell subtypes [18] to assess the changes in immune cell composition before and after the intervention), alongside Spearman’s correlation analysis to evaluate the correlations between immune cell dynamics and characteristic genes.
4.8. Competitive RNA (ceRNA) Network Construction
The ceRNA regulatory network was constructed based on ceRNA hypothesis which posited that lncRNAs competitively bind to miRNAs to regulate mRNA expression [19]. Predictions of miRNA-mRNA binding pairs were made from a systematic view with the help of five complementary databases, namely mirmap, miranda, mirDB, TargetScan, and miRTarBase, wherein consensus-disabled predictions ensured robustness. Meanwhile, miRNA-lncRNA interactions were acquired from StarBase, a high-confidence repository for non-coding RNA interactomes. Triadic ceRNA relationships were defined with stringent criteria: (1) a lncRNA and mRNA share ≥3 common miRNA binding sites, (2) significant negative co-expression, and (3) reciprocal expression trends in yoga intervention cohorts. These triads were merged into a global ceRNA network by Cytoscape with edge weights reflecting the miRNA binding affinity score. Subsequently, several key regulatory axes were prioritized, based on topological centrality (Degree > 10), and were then validated for biological validity through cross-database enrichment.
4.9. Statistical Analysis
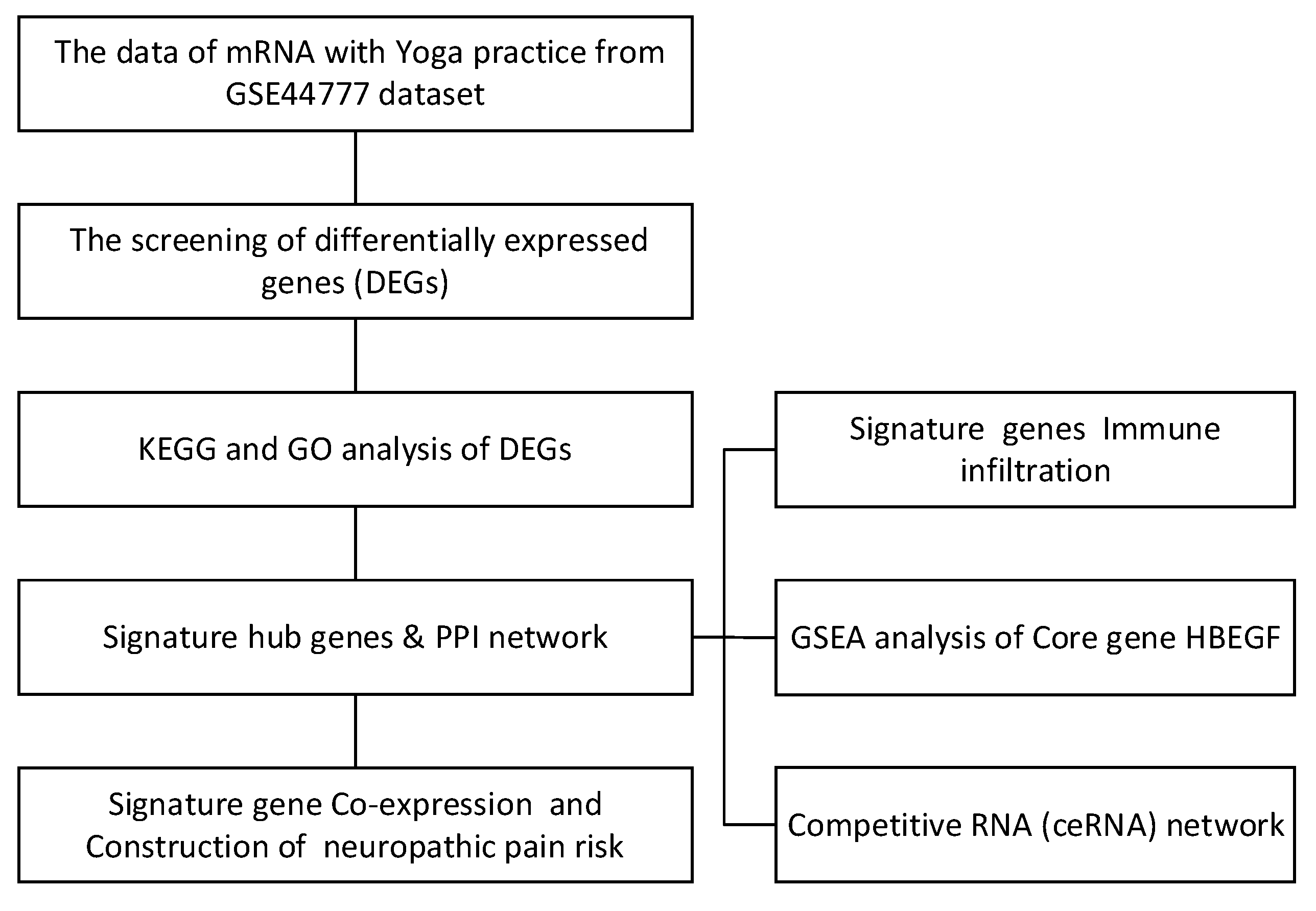
All data processing and analyses were conducted in RStudio (version 4.3.2). To compare two groups of continuous variables, paired Student’s T-test was used to compare statistical significance for normally distributed variables, while the Mann-Whitney U test (Wilcoxon rank-sum test) was used to assess differences between non-normally distributed variables. Pearson correlation analysis was used to estimate correlation coefficients between different genes. All statistical p-values are two-sided, and p-values less than 0.05 were considered significant for the study. The flow chart of this study is shown in Figure 1.
DEGs: differentially expressed genes; KEGG: Kyoto Encyclopedia of Genes and Genomes; GO: Gene Ontology; PPI: protein-protein interaction. GSEA: Gene Set Enrichment Analysis.
5. Conclusions
Multi-omics analysis reveals yoga-induced transcriptional reprogramming, immune-metabolic crosstalk, and epigenetic regulation, supporting precision public health strategies for cardiovascular disease, chronic pain, and cancer prevention.
Author Contributions
Conceptualization, H.K. and X.D.; methodology, Z.L. and C Y; software, Z.L.; formal analysis, X.D. and C Y; data curation, X.D. and Z.L.; writing—original draft preparation, X.D.; writing—review and editing, H.K. and Z.C.; supervision, Z.C. and H.K.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The gene expression datasets analyzed in this study were obtained from the publicly available Gene Expression Omnibus (GEO) repository (Accession: GSE44777). All datasets utilized in this analysis are fully de-identified and comply with the ethical standards of the original studies. The use of these data for secondary analysis was reviewed and deemed exempt from requiring additional ethics approval by the Institutional Review Board (IRB) of shanghai maritime university, as the research involved no direct interaction with human subjects and utilized only anonymized data. Original studies contributing to GEO datasets were conducted in accordance with the Declaration of Helsinki, and all participants provided informed consent for data sharing.
Informed Consent Statement
Written informed consent has been obtained from the patient(s) to publish this paper.
Data Availability Statement
“MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| DEGs | Differentially expressed genes |
| PPI | Protein-protein interaction |
| ceRNA | Competitive endogenous RNA |
| GEO | Gene Expression Omnibus |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| GO | Gene Ontology |
| ROC | Receiver operating characteristic |
| PBMCs | Peripheral blood mononuclear cells |
| GSEA | Gene Set Enrichment Analysis |
| lncRNAs | Long non-coding RNAs |
| HPA | Hypothalamic-pituitary-adrenal axis |
| CRP | C-reactive protein |
| IL | Interleukin |
| TNF | Tumor necrosis factor |
References
- Sharma, H.; Swetanshu; Singh, P. Role of Yoga in Cardiovascular Diseases. Curr. Probl. Cardiol. 2024, 49, 102032. [CrossRef]
- Nourollahimoghadam, E.; Gorji, S.; Gorji, A.; Khaleghi, G.M. Therapeutic role of yoga in neuropsychological disorders. World J. Psychiatry 2021, 11, 754–773. [CrossRef]
- Voss, S.; Cerna, J.; Gothe, N.P. Yoga Impacts Cognitive Health: Neurophysiological Changes and Stress Regulation Mechanisms. Exerc. Sport Sci. Rev. 2023, 51, 73–81. [CrossRef]
- Yeun, Y.R.; Kim, S.D. Effects of yoga on immune function: A systematic review of randomized controlled trials. Complement. Ther. Clin. 2021, 44, 101446. [CrossRef]
- Kwok, J.; Chan, L.; Lai, C.A.; Ho, P.; Choi, Z.Y.; Auyeung, M.; Pang, S.; Choi, E.; Fong, D.; Yu, D.; et al. Effects of Meditation and Yoga on Anxiety, Depression and Chronic Inflammation in Patients with Parkinson’s Disease: A Randomized Clinical Trial. Psychother. Psychosom. 2025, 1–18. [CrossRef]
- Estevao, C. The role of yoga in inflammatory markers. Brain Behav. Immun. Health 2022, 20, 100421. [CrossRef]
- Nagarathna, R.; Kumar, S.; Anand, A.; Acharya, I.N.; Singh, A.K.; Patil, S.S.; Latha, R.H.; Datey, P.; Nagendra, H.R. Effectiveness of Yoga Lifestyle on Lipid Metabolism in a Vulnerable Population-A Community Based Multicenter Randomized Controlled Trial. Medicines 2021, 8, 7. [CrossRef]
- Loewenthal, J.; Innes, K.E.; Mitzner, M.; Mita, C.; Orkaby, A.R. Effect of Yoga on Frailty in Older Adults: A Systematic Review. Ann. Intern. Med. 2023, 176, 524–535. [CrossRef]
- Cahn, B.R.; Goodman, M.S.; Peterson, C.T.; Maturi, R.; Mills, P.J. Yoga, Meditation and Mind-Body Health: Increased BDNF, Cortisol Awakening Response, and Altered Inflammatory Marker Expression after a 3-Month Yoga and Meditation Retreat. Front. Hum. Neurosci. 2017, 11, 315. [CrossRef]
- Simon, N.M.; Hofmann, S.G.; Rosenfield, D.; Hoeppner, S.S.; Hoge, E.A.; Bui, E.; Khalsa, S. Efficacy of Yoga vs Cognitive Behavioral Therapy vs Stress Education for the Treatment of Generalized Anxiety Disorder: A Randomized Clinical Trial. JAMA Psychiatry 2021, 78, 13–20. [CrossRef]
- Kaushik, D.; Shah, P.K.; Mukherjee, N.; Ji, N.; Dursun, F.; Kumar, A.P.; Thompson, I.J.; Mansour, A.M.; Jha, R.; Yang, X.; et al. Effects of yoga in men with prostate cancer on quality of life and immune response: a pilot randomized controlled trial. Prostate Cancer Prostatic Dis. 2022, 25, 531–538. [CrossRef]
- Harkess, K.N.; Ryan, J.; Delfabbro, P.H.; Cohen-Woods, S. Preliminary indications of the effect of a brief yoga intervention on markers of inflammation and DNA methylation in chronically stressed women. Transl. Psychiatry 2016, 6, e965. [CrossRef]
- Qu, S.; Olafsrud, S.M.; Meza-Zepeda, L.A.; Saatcioglu, F. Rapid gene expression changes in peripheral blood lymphocytes upon practice of a comprehensive yoga program. PLoS ONE 2013, 8, e61910. [CrossRef]
- Khokhar, M.; Tomo, S.; Gadwal, A.; Purohit, P. Multi-omics integration and interactomics reveals molecular networks and regulators of the beneficial effect of yoga and exercise. Int. J. Yoga 2022, 15, 25–39. [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [CrossRef]
- Sun, J.; Zhao, J.; Jiang, F.; Wang, L.; Xiao, Q.; Han, F.; Chen, J.; Yuan, S.; Wei, J.; Larsson, S.C.; et al. Identification of novel protein biomarkers and drug targets for colorectal cancer by integrating human plasma proteome with genome. Genome Med. 2023, 15, 75. [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [CrossRef]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 2015, 12, 453–457. [CrossRef]
- Mao, M.; Zhang, J.; Xiang, Y.; Gong, M.; Deng, Y.; Ye, D. Role of exosomal competitive endogenous RNA (ceRNA) in diagnosis and treatment of malignant tumors. Bioengineered 2022, 13, 12156–12168. [CrossRef]
- Alfaddagh, A.; Martin, S.S.; Leucker, T.M.; Michos, E.D.; Blaha, M.J.; Lowenstein, C.J.; Jones, S.R.; Toth, P.P. Inflammation and cardiovascular disease: From mechanisms to therapeutics. Am. J. Prev. Cardiol. 2020, 4, 100130. [CrossRef]
- Tan, A.; Thomas, R.L.; Campbell, M.D.; Prior, S.L.; Bracken, R.M.; Churm, R. Effects of exercise training on metabolic syndrome risk factors in post-menopausal women - A systematic review and meta-analysis of randomised controlled trials. Clin. Nutr. 2023, 42, 337–351. [CrossRef]
- Patil, S.G.; Sobitharaj, E.C.; Chandrasekaran, A.M.; Patil, S.S.; Singh, K.; Gupta, R.; Deepak, K.K.; Jaryal, A.K.; Chandran, D.S.; Kinra, S.; et al. Effect of Yoga-Based Cardiac Rehabilitation Program on Endothelial Function, Oxidative Stress, and Inflammatory Markers in Acute Myocardial Infarction: A Randomized Controlled Trial. Int. J. Yoga 2024, 17, 20–28. [CrossRef]
- Sarvottam, K.; Magan, D.; Yadav, R.K.; Mehta, N.; Mahapatra, S.C. Adiponectin, interleukin-6, and cardiovascular disease risk factors are modified by a short-term yoga-based lifestyle intervention in overweight and obese men. J. Altern. Complement. Med. 2013, 19, 397–402. [CrossRef]
- Kaliman, P.; Alvarez-Lopez, M.J.; Cosin-Tomas, M.; Rosenkranz, M.A.; Lutz, A.; Davidson, R.J. Rapid changes in histone deacetylases and inflammatory gene expression in expert meditators. Psychoneuroendocrinology 2014, 40, 96–107. [CrossRef]
- Bishop, J.R.; Lee, A.M.; Mills, L.J.; Thuras, P.D.; Eum, S.; Clancy, D.; Erbes, C.R.; Polusny, M.A.; Lamberty, G.J.; Lim, K.O. Corrigendum: Methylation of FKBP5 and SLC6A4 in Relation to Treatment Response to Mindfulness Based Stress Reduction for Posttraumatic Stress Disorder. Front. Psychiatry 2021, 12, 642245. [CrossRef]
- Chaix, R.; Fagny, M.; Cosin-Tomas, M.; Alvarez-Lopez, M.; Lemee, L.; Regnault, B.; Davidson, R.J.; Lutz, A.; Kaliman, P. Differential DNA methylation in experienced meditators after an intensive day of mindfulness-based practice: Implications for immune-related pathways. Brain Behav. Immun. 2020, 84, 36–44. [CrossRef]
- Verzili, B.; Valerio, D.A.M.; Herrmann, F.; Reyes, M.B.; Galduroz, R.F. A systematic review with meta-analysis of Yoga’s contributions to neuropsychiatric aspects of aging. Behav. Brain Res. 2023, 454, 114636. [CrossRef]
- Gautam, S.; Kumar, R.; Kumar, U.; Kumar, S.; Luthra, K.; Dada, R. Yoga maintains Th17/Treg cell homeostasis and reduces the rate of T cell aging in rheumatoid arthritis: a randomized controlled trial. Sci. Rep. 2023, 13, 14924. [CrossRef]
- R, P.; Kumar, A.P.; Dhamodhini, K.S.; Venugopal, V.; Silambanan, S.; K, M.; Shah, P. Role of yoga in stress management and implications in major depression disorder. J. Ayurveda Integr. Med. 2023, 14, 100767. [CrossRef]
- Greaney, S.K.; Amin, N.; Prudner, B.C.; Compernolle, M.; Sandell, L.J.; Tebb, S.C.; Weilbaecher, K.N.; Abeln, P.; Luo, J.; Tao, Y.; et al. Yoga Therapy During Chemotherapy for Early-Stage and Locally Advanced Breast Cancer. Integr. Cancer Ther. 2022, 21, 1553452421. [CrossRef]
- Kwon, R.; Kasper, K.; London, S.; Haas, D.M. A systematic review: The effects of yoga on pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 250, 171–177. [CrossRef]
- Shidhaye, R.; Bangal, V.; Bhargav, H.; Tilekar, S.; Thanage, C.; Gore, S.; Doifode, A.; Thete, U.; Game, K.; Hake, V.; et al. Feasibility, acceptability, and preliminary efficacy of yoga to improve maternal mental health and immune function during the COVID-19 crisis (Yoga-M (2) trial): a pilot randomized controlled trial. Front. Hum. Neurosci. 2023, 17, 1115699. [CrossRef]
- Vollbehr, N.K.; Stant, A.D.; Hoenders, H.; Bartels-Velthuis, A.A.; Nauta, M.H.; Castelein, S.; Schroevers, M.J.; de Jong, P.J.; Ostafin, B.D. Cost-effectiveness of a mindful yoga intervention added to treatment as usual for young women with major depressive disorder versus treatment as usual only: Cost-effectiveness of yoga for young women with depression. Psychiatry Res. 2024, 333, 115692. [CrossRef]
- Nalbant, G.; Lewis, S.; Chattopadhyay, K. Characteristics of Yoga Providers and Their Sessions and Attendees in the UK: A Cross-Sectional Survey. Int. J. Environ. Res. Public Health 2022, 19, 4. [CrossRef]
- Lachance, C.C.; McCormack, S. Mindfulness Training and Yoga for the Management of Chronic Non-malignant Pain: A Review of Clinical Effectiveness and Cost-effectiveness. Canadian Agency for Drugs and Technologies in Health: Ottawa, ON, Canada, 2019.
- Nagarhalli, M. Yoga for the primary prevention of cardiovascular disease: Summary of a Cochrane review. Explore 2021, 17, 96. [CrossRef]
- Bennetts, A. How does yoga practice and therapy yield psychological benefits? A review and model of transdiagnostic processes. Complement. Ther. Clin. 2022, 46, 101514. [CrossRef]
- Anheyer, D.; Haller, H.; Lauche, R.; Dobos, G.; Cramer, H. Yoga for treating low back pain: a systematic review and meta-analysis. Pain 2022, 163, e504–e517. [CrossRef]
- Mooventhan, A.; Nivethitha, L. Role of yoga in the prevention and management of various cardiovascular diseases and their risk factors: A comprehensive scientific evidence-based review. Explore 2020, 16, 257–263. [CrossRef]
Figure 2.
Genes differentially expressed between yoga intervention pre- and post-Intervention. (A) Volcano plot depicting the results of differential gene expression analysis from GSE44777. (B) Heatmap resulting from the differential gene expression analysis of GSE44777. Blue indicates downregulated genes, yellow indicates upregulated genes, and black represents genes with statistically insignificant differences.
Figure 2.
Genes differentially expressed between yoga intervention pre- and post-Intervention. (A) Volcano plot depicting the results of differential gene expression analysis from GSE44777. (B) Heatmap resulting from the differential gene expression analysis of GSE44777. Blue indicates downregulated genes, yellow indicates upregulated genes, and black represents genes with statistically insignificant differences.
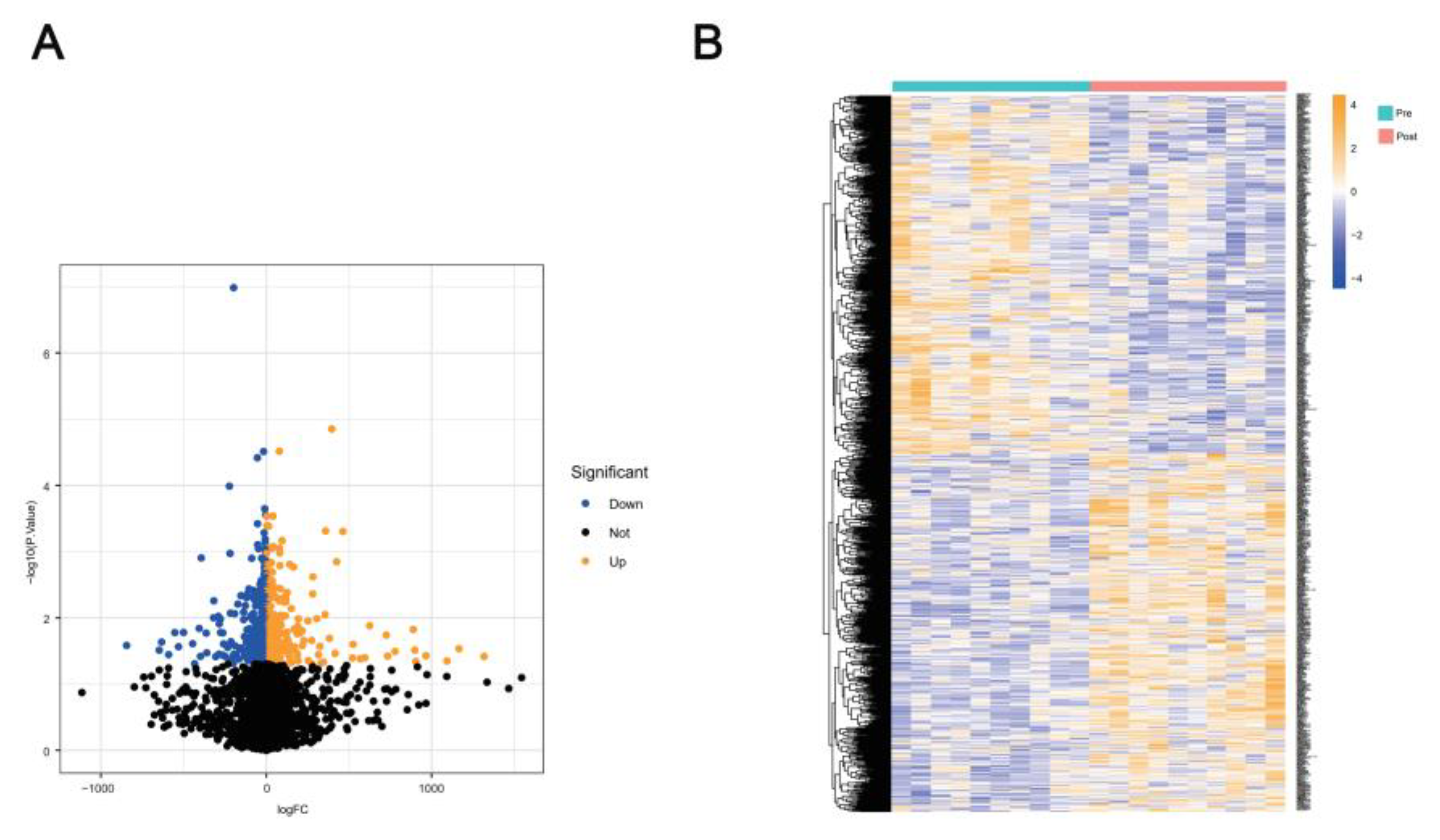
Figure 3.
Differential gene enrichment analysis. (A) Gene Ontology (GO) enrichment analysis of differentially expressed genes. (B) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of differentially expressed genes.
Figure 3.
Differential gene enrichment analysis. (A) Gene Ontology (GO) enrichment analysis of differentially expressed genes. (B) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of differentially expressed genes.
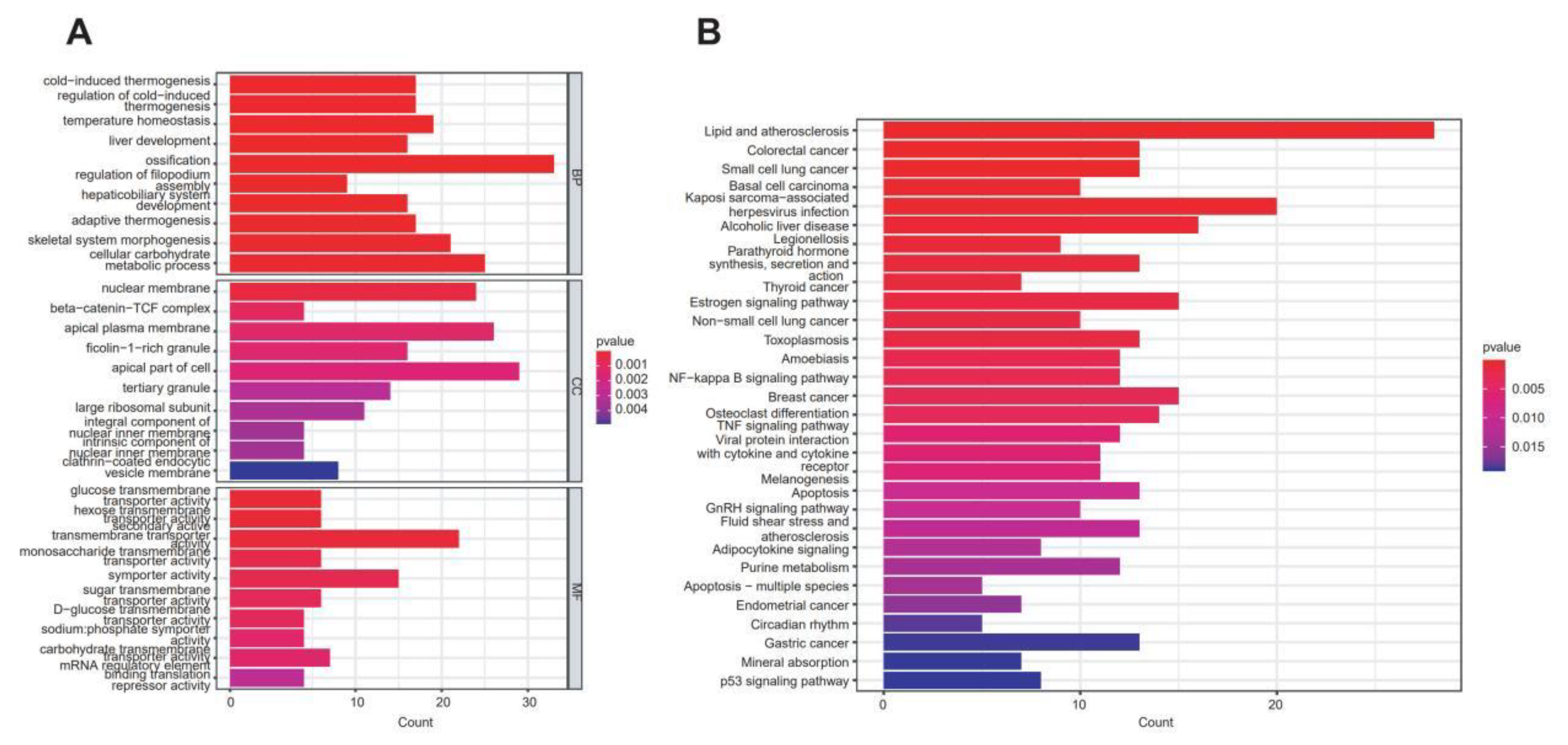
Figure 4.
Construction of the PPI Network. (A) PPI network of overlapping hub genes. (B) Identification of core genes in the interaction network using the degree algorithm. PPI: Protein-Protein Interaction. A: The darker the color, the higher the score. B: The size and color depth of nodes (genes/proteins) in the network diagram represent their centrality and importance.
Figure 4.
Construction of the PPI Network. (A) PPI network of overlapping hub genes. (B) Identification of core genes in the interaction network using the degree algorithm. PPI: Protein-Protein Interaction. A: The darker the color, the higher the score. B: The size and color depth of nodes (genes/proteins) in the network diagram represent their centrality and importance.
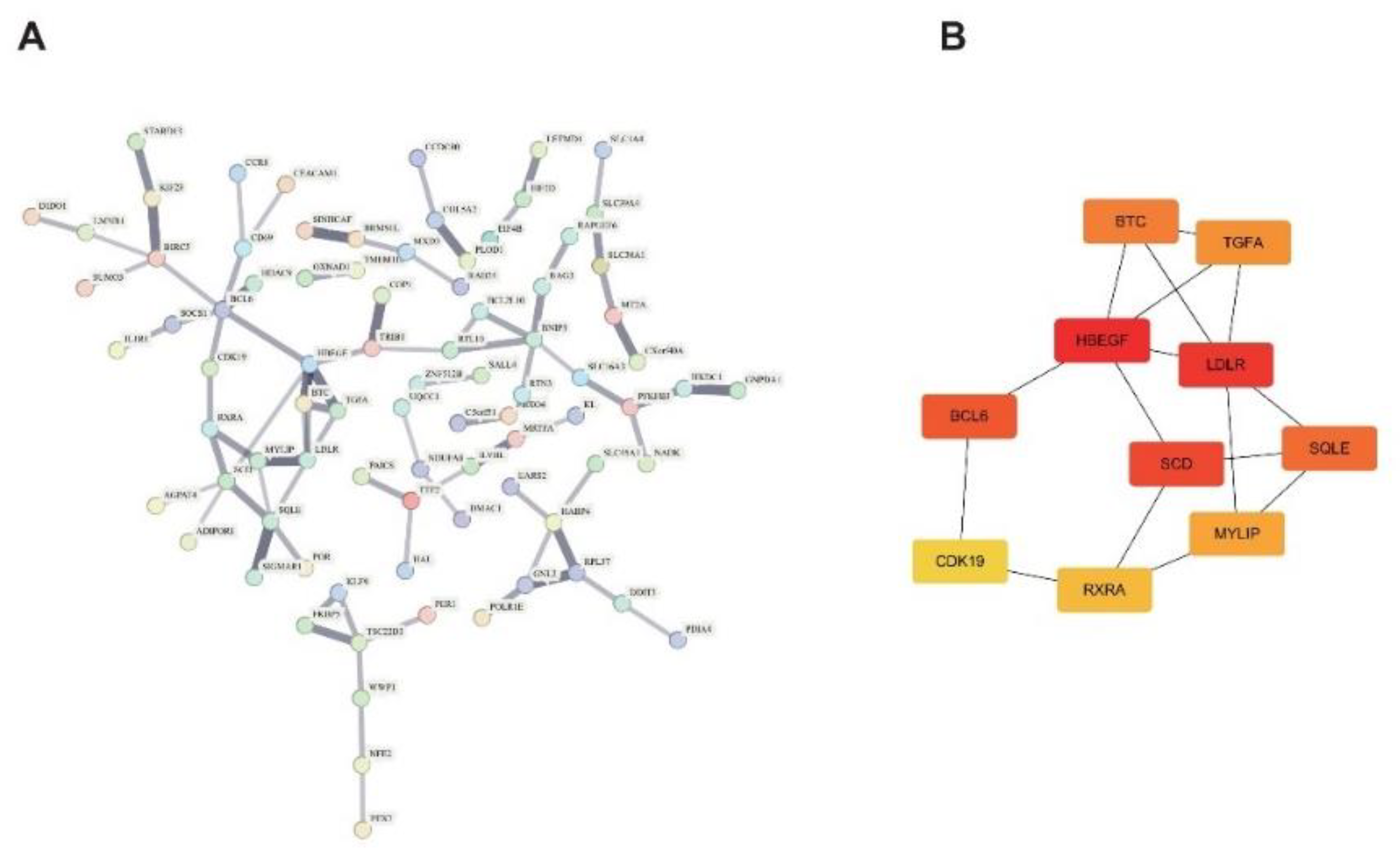
Figure 5.
The utilization of nomograms for forecasting neuropathic pain risk in dataset GSE250152. (A) Key gene correlation expression. (B) The nomogram model representing hub genes. (C) The diagnostic performance of the nomogram model alongside each hub gene was evaluated using ROC curves. ROC: Receiver operating characteristic.
Figure 5.
The utilization of nomograms for forecasting neuropathic pain risk in dataset GSE250152. (A) Key gene correlation expression. (B) The nomogram model representing hub genes. (C) The diagnostic performance of the nomogram model alongside each hub gene was evaluated using ROC curves. ROC: Receiver operating characteristic.
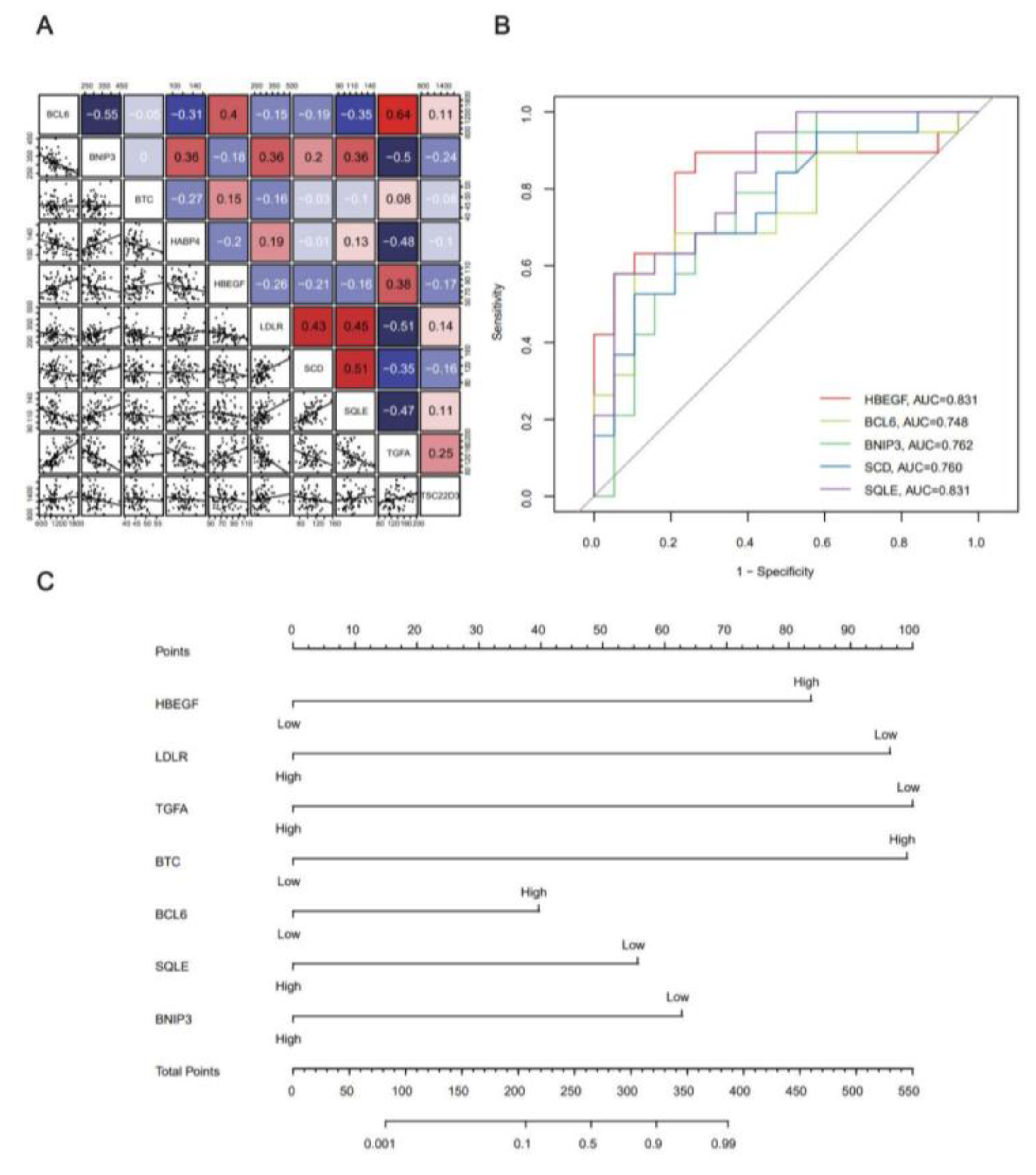
Figure 6.
GSEA Enrichment Analysis of Yoga Key Gene HBEGF. (A) KEGG pathway analysis of key genes; (B) GO enrichment analysis of key genes. GSEA: Gene set enrichment analysis; KEGG: Kyoto Encyclopedia of Genes and Genomes (KEGG); GO: Gene Ontology.
Figure 6.
GSEA Enrichment Analysis of Yoga Key Gene HBEGF. (A) KEGG pathway analysis of key genes; (B) GO enrichment analysis of key genes. GSEA: Gene set enrichment analysis; KEGG: Kyoto Encyclopedia of Genes and Genomes (KEGG); GO: Gene Ontology.
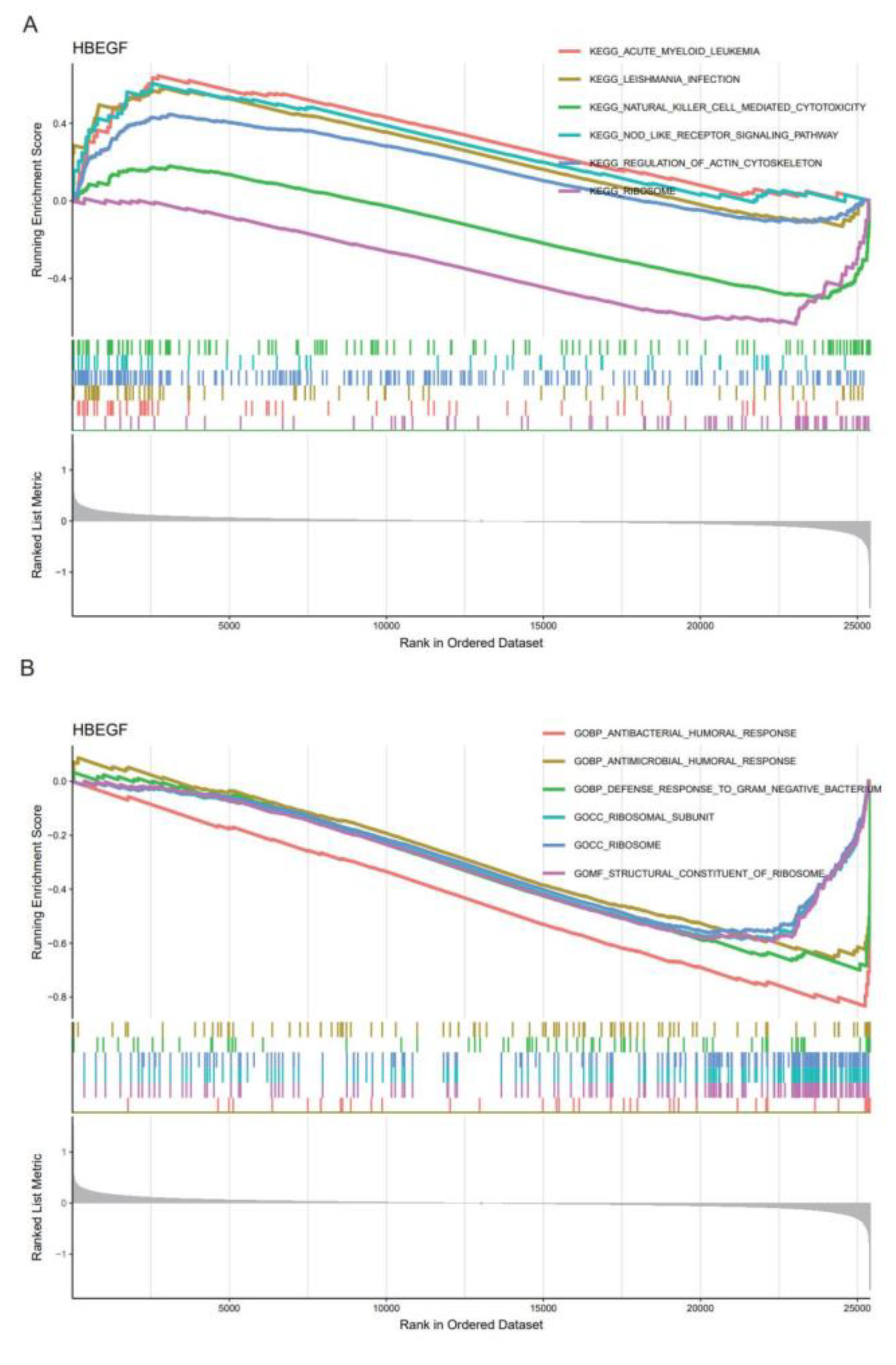
Figure 7.
Correlation of Key Gene with Immune Infiltration. (A) Heatmap depicting the correlation between key genes and immune infiltration types. (B) Relationship between immune infiltration types and key genes.
Figure 7.
Correlation of Key Gene with Immune Infiltration. (A) Heatmap depicting the correlation between key genes and immune infiltration types. (B) Relationship between immune infiltration types and key genes.
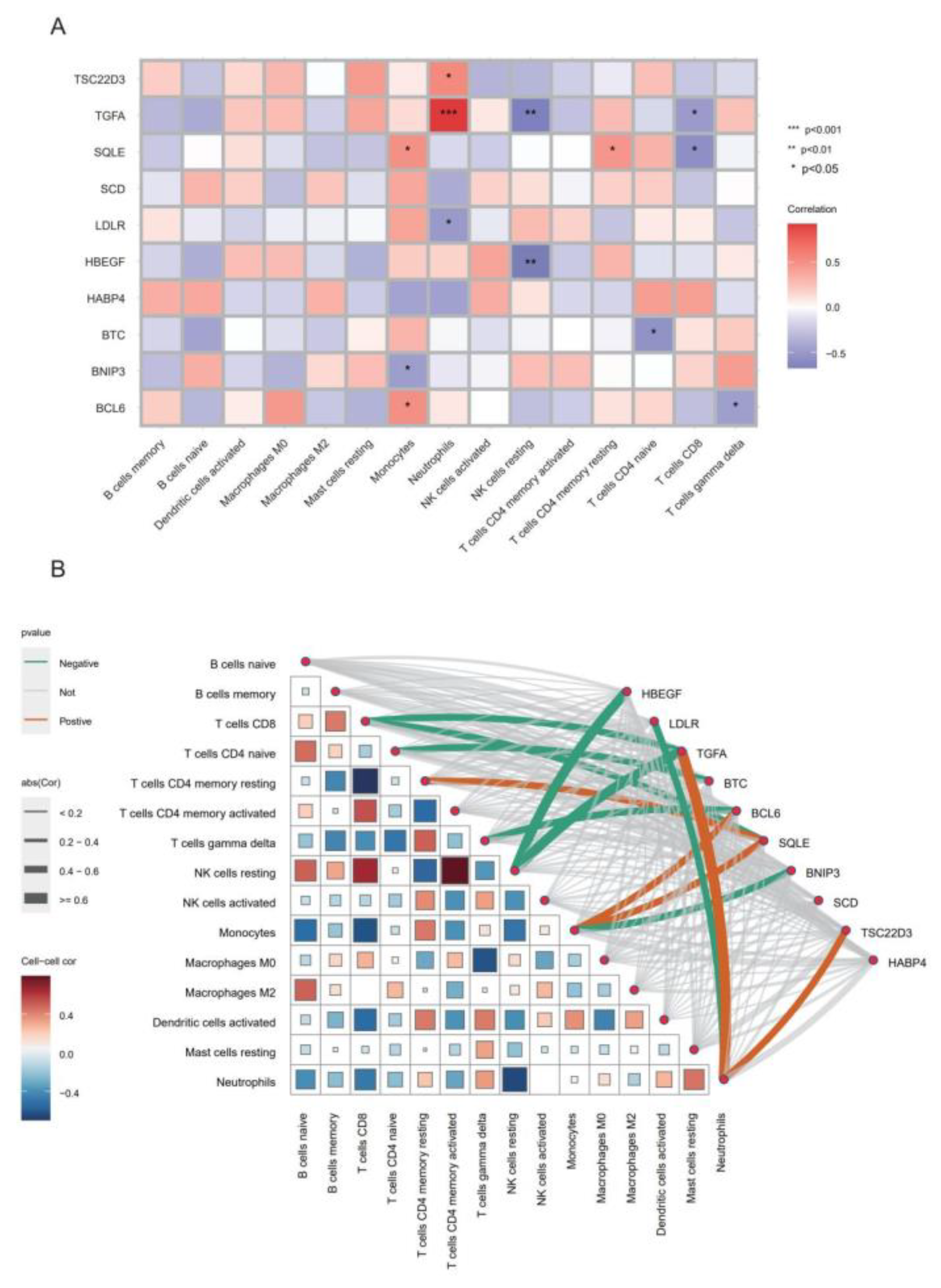
Figure 8.
ceRNA Network of Key Gene HBEGF involving mRNA, miRNA, and lncRNA.
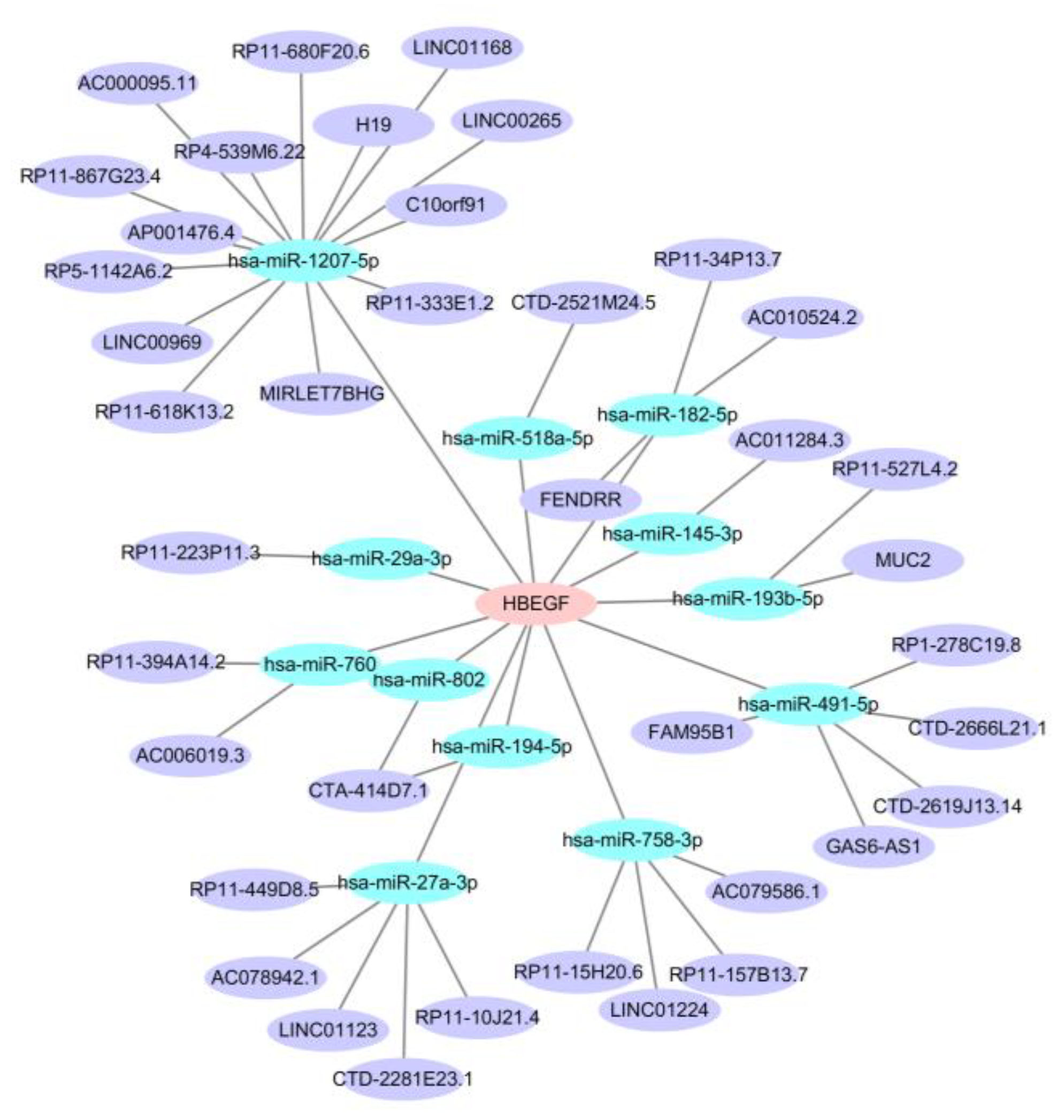
Figure 1.
The flowchart of this study.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



