
Submitted:
10 April 2025
Posted:
11 April 2025
You are already at the latest version
Abstract
Background/Objectives: Water and alcohol extracts of Epilobium have gained attention due to the high concentration of its bioactive compounds and associated health benefits. This review aimed to evaluate the effects of Epilobium parviflorum Shreb. (Onagraceae) preparations on vasculature in the light of its medical application on human health over the last five years. Materials and Methods: A literature search of databases such as PubMed/Medline, Scopus, and Google Scholar for original articles published between March 2000 and March 2025 was undertaken. The keywords used were: “aortic rings”, “ellagitannins”, “Epilobium”, “hypotension”, “oenothein B”, “Onagraceae”, “systolic blood pressure”, “vasorelaxation”, “willow herb”. Only one study was found, and this was discussed in this review. Results: Herb of E. parviflorum Shreb. is used as a remedy in folk medicine with a variety of therapeutic properties, including its preventive effects and ability to relieve symptoms in patients with benign prostate hyperplasia, prostatitis, and cancer. Other properties include kidney and urinary tract diseases, skin infections and antibacterial properties. E. parviflorum contains bioactive compounds such as oenothein B, quercetin-3-O-glucuronide and myricetin-3-O-rhamnoside. In a low dose they contribute to the reduction of oxidative stress due to their antioxidant and immunostimulant effects, positively reducing inflammation that causes certain conditions. At higher concentrations Epilobium generates reactive oxygen species that stimulate the body’s defense mechanisms against a variety of cancers. Conclusions: Although the potential application of E. parviflorum in metabolic disorders was not extensively studied before, its antioxidant and anti-inflammatory properties are well documented, which suggests potential pathways for future research and therapeutic development of preparations to benefit vascular health.
Keywords:
aortic rings
; ellagitannins
; Epilobium
; hypotension
; oenothein B
; Onagraceae
; polyphenols
; systolic blood pressure
; vasorelaxation
; Willow herb
1. Introduction
Epilobium parviflorum Schreb. (willow herb – UK or fireweed – US), is a perennial herbaceous plant from the Onagraceae family, that grows throughout Europe, Northern Africa, and Western Asia. Tea brewed from the E. parviflorum herb offers numerous health benefits, including alleviating disorders of the prostate gland, bladder and kidney, due to the anti-inflammatory, diuretic, and antioxidant activities of E. parviflorum. The infusion of 1–2 g in a cup of water is taken orally three to four times daily [1]. Infusion of Epilobium species is prepared in traditional medicine mainly to treat the early stages of benign prostate hyperplasia (BPH), urethritis, and micturition disorders [2], as well as in alleviating menstrual disorders [3], rectal bleeding [4], and the treatment of prostate adenoma [5]. It is also used in treating gastrointestinal and mucous membrane damage and promoting wound healing [4], Figure 1.
Epilobium species are rich in various bioactive compounds such as polyphenols represented by flavonoids (myricetin, quercetin, kaempferol, catechin), phenolic acids (gallic acid, chlorogenic acid, (Z)-p-coumaric acid, caffeoylquinic acid, p-coumaroylquinic acid, feruloylquinic acid), and hydrolysable tannins (ellagitannins) [6]. Other constituents such as lignans, steroids, triterpenoids, fatty acids, and essential oils, have also been identified [5,6]. Phytochemical analysis of the E. parviflorum extracts has shown that ellagitannin, oenothein B is the predominant compound, ranging from 20% to 35% of the total content of extracts and varies from 2 to 4.5% in the raw plant [6,7].
Oenothein B stands out as a distinct type of ellagitannin characterized by a macrocyclic structure that limits the flexibility of rotational bonds. This compound is particularly abundant in various medicinal plants such as Epilobium, Oenothera, and Eucalyptus species. Extensive research has demonstrated its diverse biological effects, including antioxidant and anti-inflammatory properties, potential anticancer activity, immunomodulatory, and antibacterial effects [8]. Phenolic acids are also important contributors to the antioxidant and therapeutic potential against BPH of this species [9].
2. Materials and Methods
A literature search of databases such as PubMed/Medline, Scopus, and Google Scholar for original articles published between March 2000 and March 2025 was undertaken. The keywords used were “aortic rings”, “ellagitannins”, “Epilobium”, “hypotension”, “oenothein B”, “Onagraceae”, “systolic blood pressure”, “vasorelaxation”, “willow herb”. Only one study was found, and this was included in this review, see Table 1.
3. Results
3.1. Direct Impact of Epilobium on the Vasculature
A study conducted by Isla et al. (2024) demonstrated the vasorelaxant effects of ellagitannins isolated from Cuphea carthagenensis (Jacq.) J. F. Macbr (Lythraceae) [10], see Table 1. C. carthagenensis is a popular plant in Brazilian folk medicine known for its hypotensive and depressant effects on the central nervous system. The presented results demonstrated the hypotensive effect of C. carthagenensis aqueous extract after one week of oral treatment of normotensive rats. In the same experiment, the rat aortic rings were pre-contracted with noradrenaline, and directly treated with three hydrolyzable tannins; oenothein B, woodfordin C, and eucalbanin B (20 – 180 µM), which induced a concentration-related vasorelaxation. Ellagitannins results found in studies induced vasorelaxation in vitro by activating nitric oxide (NO) synthesis/NO release from endothelial cells, without altering the vascular smooth muscles [10]. Authors concluded that the determined in vitro actions of these three compounds are unlikely to account for the hypotensive effect of alcoholic extract in vivo considering the low oral bioavailability of ellagitannins. It was found that other compounds and/or mechanisms are involved. Since oenothein B is the main constituent of E. parviflorum, it can be concluded that Epilobium herbal extract may also possess the hypotensive properties in vivo and regulate NO synthesis and/or release in vitro in a dose-dependent manner. Probably inhibition and/or release of NO involving E. parviflorum mechanism likely depends on the tissue’s oxygenation status, oxidative stress, inflammation, and reactive oxygen species (ROS), requiring further studies to determine.
Nitric oxide is a key signalling molecule playing a role in various biological activities. It regulates vascular tone and blood flow by activating soluble guanylate cyclase (sGC) in vascular smooth muscle, controls mitochondrial oxygen consumption by inhibiting cytochrome c oxidase, and activates immune cells, particularly macrophages, to elicit a protective response. Disturbances in NO production and transport within the vascular area led to cardiovascular diseases, including hypertension, atherosclerosis, and angiogenesis-related disorders [11].
Moreover, in another study Kim et al. [12] described how ethanol extract of Oenothera odorata (Onagraceae) seeds induced vasorelaxation via endothelium-dependent NO-cGMP signaling cascade through activation of Akt-eNOS-sGC pathway.
3.2. The Possible Impact of Epilobium on the Vasculature Based on its Properties
3.2.1. The Antioxidant and the Anti-Inflammatory Activity of Epilobium
The antioxidants [1,6,13,14], and the anti-inflammatory [8] activity of Epilobium extracts are a fundamental biological property in a plethora of Epilobium species [1]. Myricetin-3-O-rhamnoside (E. parviflorum) and quercetin-3-O-glucuronide (E. angustifolium) were shown to inhibit cyclooxygenases-1 and -2 (COX-1 and COX-2) [15]. In another study the methanol-aqueous extract from E. angustifolium aerial parts scavenged free radicals, and exerted a distinct impact on α-glucosidase, α-amylase and lipase. The studied extract also inhibited acetyl- and butyrylcholinesterase and had a prominent anti-tyrosinase effect [6]. Epilobium extracts may be considered a source of agents for treating disorders related to oxidative stress and inflammation.
Antioxidant and anti-inflammatory effects of E. parviflorum, and other two plant extracts Melilotus officinalis and Cardiospermum halicacabum showed a beneficial effect in macrophage and microglial cells. Merighi et al. [16] showed that 40% ethanol plant extracts from these herbs reduced NO production in macrophage and microglial cells with a conclusion that these extracts may be considered as a source of agents for treating disorders related to oxidative stress and inflammation. Ethanol extracts from the dried aerial part of E. parviflorum are characterized by the presence of several polyphenols, particularly flavonoids and condensed tannins, and can be considered a potential source of agents for treating disorders related to oxidative stress and inflammation.
In other studies, oenothein B was shown to be capable of inhibiting myeloperoxidase, hyaluronidase, and lipoxygenase-5 (LOX-5) thereby inhibiting the formation of reactive oxygen species (ROS) [17]. An imbalance in the body’s antioxidant system is associated with the pathogenesis of diseases such as cardiovascular diseases, inflammation, and cancers. This occurs through an increase in ROS [18].
Hiermann et al. [8] noticed that the aqueous but not methanolic extracts of E. angustifolii herb reduced the release of prostaglandins I2, E2 and D2 (in the perfused rabbit ear) approximately 5 times more effectively than did similar extracts of E. parviflori herb. Hiermann et al. isolated a minor component, myricetin 3-O-glucuronide, from E. angustifolium and identified it as a potent inhibitor of COX-1, COX-2, and 5-LOX activity [8]. Unfortunately, this discovery did not fully explain the inhibition of COX-1 and COX-2 activities by extracts of E. parviflorum [1,15]. Ethanol extracts of E. parviflorum also exhibit inhibitory effects on both COX-1 and COX-2-catalyzed prostaglandin biosynthesis and exert antioxidant activity [1]. COX-1 functioning is typically associated with maintaining physiological homeostasis and tissue protection, whereas COX-2 is the primary enzyme responsible for prostaglandin production in response to inflammation [19]. However, high activity of COX enzymes, particularly COX-2 (and in some cases COX-1), can impact the development of arterial hypertension. Both COX isoforms are involved in prostaglandin synthesis, which affects the regulation of blood vessel tension. Prostaglandins can act on vascular receptors, leading to impaired vasoconstriction or vasodilation. Additionally, they may influence sodium and water retention in the body, thereby increasing circulating blood volume and blood pressure. Chronic inflammatory conditions can lead to sustained increases in blood pressure by affecting blood vessels and the renal system. Elevated levels of ROS induced by COX activity are also associated with processes that can contribute to arterial hypertension through vascular damage and alterations in renal functioning. Hydroxyl radical (HO•) scavengers, including E. parviflorum, as noted by Kumagai et al. [19] and Feng et al. [20] inhibit COX regulation and subsequently reduce inflammation.
Early research by Okuda et al. [21] on the antioxidant activity of the new classes of tannins showed that the antioxidant properties of ellagitannins were generally higher than those of α-tocopherol and ascorbic acid. Ascorbic acid acts as a strong antioxidant, neutralizing free radicals responsible for the damage to the body’s cells and tissues, thereby providing protection against oxidative stress-related damage. Additionally, it helps regenerate the oxidized form of vitamin E into its active form, allowing its antioxidant properties to persist longer in the body. Ellagitannins have strong free radical scavenging abilities, which can inhibit the free radical chain reaction of other compounds, through their self-oxidation, thereby preventing the oxidation of lipids, proteins or DNA, which can significantly impact the development of hypertension [22,23,24,25].
Free radicals can damage the endothelium, leading to vascular dysfunction by reducing NO production and increasing peroxynitrite (ONOO–) formation disrupting the regulation of vascular tone. Reduced NO levels can lead to increased vascular resistance, decreased vasodilation and elevated blood pressure. Furthermore, tissue damage by free radicals can lead to chronic inflammation in the vascular wall, which in turn increases the production of pro-inflammatory cytokines and chemokines that can affect vascular contraction and increase blood pressure. Damage to DNA and proteins in the endothelium can lead to endothelial dysfunction. Lipid peroxidation leads to the formation of toxic metabolites and damage to cell membranes [19].
Oenothein B inhibits the release of hyaluronidase and myeloperoxidase from stimulated neutrophils, like the anti-inflammatory drug indomethacin. Myeloperoxidase, in the presence of iron ions and hydrogen peroxide (H2O2) generates oxygen radicals.
Scholar Kiss and others reported that oenothein B (from Oenothera paradoxa extract, Onagraceae) also can inhibit lipoxygenase, hyaluronidase, and ROS production from human neutrophils, demonstrating anti-inflammatory activity [26]. On the other hand, high concentrations of oenothein B, according to Sakagami et al., can have a pro-oxidative effect by inducing elevated levels of ROS against human oral tumour cell lines (Figure 2) [18]. LOX is an enzyme responsible for the oxidation of fatty acids, leading to the production of leukotrienes. Leukotrienes are lipid mediators of inflammation that can affect vascular tone by inducing smooth muscle contraction and reducing blood flow, contributing to increased blood pressure. Hyaluronidase is an enzyme responsible for the breakdown of hyaluronic acid, which is an important component of the extracellular matrix. On one hand, it inhibits the release of myeloperoxidase from neutrophils, however, excessive levels may lead to reduced hyaluronic acid levels and affect the permeability and function of the vascular wall as well as the elasticity of blood vessels. Therefore, the balance between the synthesis and secretion of this acid is crucial and regulation depends on the concentration of oenothein B [26,27]. In another study, authors evaluated the immunomodulatory effects induced by tannin treatment in human dendritic cells (DCs), by measuring the changes in cytokine production, cell differentiation, and cell viability. Significant changes were noticed in down-regulation of the expression of cell surface molecules, CD1a and CD83, the induction of apoptosis without the activation of caspase-3/7, 8, and 9, and suppressed production of inflammatory cytokines, such as interleukins (IL) especially: IL-1β and IL-6, in a dose-dependent manner. These may in part explain the traditional use of oenothein B-containing medicinal herbs for the treatment of a variety of inflammatory diseases, including inflammatory bowel disease, celiac disease, and rheumatoid arthritis. Moreover, studies conducted by Yoshimura and Okuyama demonstrated that oenothein B could reduce neuroinflammation in the brain during systemic inflammation when oenothein B was administered orally by enhancing neuronal signalling pathways as it suppressed (i) LPS-induced abnormal behavior in open field; (ii) LPS-induced microglial activation in the hippocampus and striatum; and (iii) LPS-induced COX-2 production in the hippocampus and striatum of mice [28,29,30].
3.2.2. The Impact of Epilobium on the Lipid Profile and Atherosclerotic Plaque Synthesis
High levels of cholesterol in the blood, especially low-density lipoprotein (LDL), play a crucial role in the development of atherosclerotic plaques. Elevated LDL cholesterol damages the endothelium of blood vessels, which is the first step of atherosclerosis development [31,32]. Damaged endothelium allows LDL to penetrate the arterial wall, where it undergoes oxidation, triggering an inflammatory response. Oxidized LDL (oxLDL) is recognized by macrophages, which migrate into the arterial wall and ingest oxLDL. These macrophages transform into foam cells filled with lipids, leading to the formation of initial atherosclerotic lesions. Thus, high LDL cholesterol is a major risk factor for atherosclerosis and related cardiovascular diseases [33].
One study demonstrated that strawberry extract (rich in ellagitannins) added to a high-fat diet attenuated disturbances in the redox and lipid status as well as in the inflammatory response in rats. This manifested as decreased triglycerides and total cholesterol. Moreover, body weight gain, hepatic fat, oxidized glutathione, and thiobarbituric acid-reactive substance concentrations also decreased [34]. In another study conducted by the same research team, ellagitannin-rich extract obtained from strawberries induced beneficial changes in the hepatic fat content, reduced and oxidized glutathione (GSH and GSSG) concentrations and GSH/GSSG ratio and modified blood plasma antioxidant status (FRAP, ACL), HDL-cholesterol, and atherogenic coefficient values in rats fed with a high-fat diet [24].
Oenothein B treatment has protective effects and supports liver function. Oenothein B ameliorates hepatic injury in alcoholic liver disease mice by improving oxidative stress and inflammation and modulating the gut microbiota. Xu et al. demonstrated that oenothein B treatment reduced alcohol-induced hepatic injury, as evidenced by decreased levels of aminotransferases, and inflammatory biomarkers and restored gut microbiota dysbiosis [35]. Oenothein B mitigated pathological damage by reducing lipid droplet accumulation, lipid vacuolation, inflammation, and fibrosis. The authors concluded that supplementation with oenothein B is beneficial in maintaining hepatocyte integrity and regulating lipid metabolism in hepatocytes. Oenothein B increased the expression of Nuclear factor erythroid 2-related factor 2 (Nrf2), Hemoxygenase-1 (HO-1), NQO1 genes and Kelch-like ECH-associated protein 1 (Keap1) levels. The Keap1/Nrf2 signalling pathway is a crucial component of the liver’s antioxidative system with protective effects. Under normal conditions, Keap1 binds to Nrf2, promoting its ubiquitination and degradation in the cytoplasm, maintaining low Nrf2 levels. During inflammation and oxidative stress, Nrf2 dissociates from Keap1 and translocates into the nucleus, promoting the expression of target genes such as HO-1, NQO1, SOD, CAT, and GSH, which reduce oxidative stress-induced damage [35]. Oenothein B increases the levels of antioxidant enzymes (SOD, CAT, GSH) and decreases oxidative stress markers (MDA, CYP2E1) [35]. This indicates that oenothein B effectively reduces oxidative stress in the liver. In vivo studies demonstrated that in HepG2 cells, levels of SOD, CAT, and GSH were lower, while MDA and ROS levels were higher, confirming oenothein B impact on reducing oxidative stress. Oenothein B plays an important role in reducing atherosclerotic plaque formation through several mechanisms related to regulating inflammation, oxidative stress, and gut microbiota. Oxidative stress and inflammation play a key role in the pathogenesis of alcoholic liver disease (ALD). Acute alcohol consumption increases gut permeability, allowing lipopolysaccharides (LPS) to enter the liver, where they activate toll-like receptor 4 (TLR4) on Kupffer cells. TLR4 activation leads to increased expression of inflammatory genes such as NF-κB, Myd88, and CD14, resulting in the production of inflammatory cytokines (IL-1β, IL-6, TNF-α), which cause liver inflammation and damage. Oenothein B significantly reduces TLR4 expression and downstream inflammatory mediators, leading to decreased levels of inflammatory cytokines in the liver. This anti-inflammatory effect has also been confirmed in vitro, where oenothein B reduced ethanol-induced NO levels in HepG2 cells. Oenothein B limits the nuclear translocation of p65, reducing NF-κB binding activity and inhibiting TLR/NF-κB pathway-dependent NO synthesis [35]. Thus, oenothein B alleviates oxidative stress and inflammation, which are crucial factors in atherosclerotic plaque development. Alcohol abuse alters gut microbiota composition, leading to disturbances of the gut-liver axis. These changes can influence the development of inflammation and oxidative stress in the body. Supplementation with oenothein B restores gut microbiota balance by increasing the abundance of short-chain fatty acid (SCFA) producers such as Muribaculaceae and Erysipelotrichaceae. SCFAs are gut microbiota metabolites that activate the Nrf2 signalling pathway in the liver, enhancing the antioxidative system. Oenothein B reduces the abundance of Gram-negative bacteria (Akkermansia), whose lipopolysaccharide (LPS) activates the TLR4/NF-κB pathway in the liver, leading to inflammation. Reducing the number of these bacteria decreases overall inflammation in the body [35].
In summary, oenothein B plays a multifaceted role in reducing atherosclerotic plaque by regulating oxidative stress, reducing inflammation and restoring gut microbiota balance. Through modulation of these processes, oenothein B may contribute to improving vascular health and reducing the risk of atherosclerosis.
3.2.3. Anti Colorectal Cancer Activity
Extracts from Epilobium species induce apoptosis in human hormone-dependent prostate cancer cells through activation of the mitochondrial pathway and caspase-3 activation, Figure 1 [5]. In this process, disruption of the mitochondrial membrane potential occurs, triggering the release of cytochrome c from the mitochondria into the cytosol. In the cytoplasm, cytochrome c binds with Apaf-1 (apoptosis protease-activating factor-1) and pro-caspase-9, forming a complex called the apoptosome. Upon formation of the apoptosome, pro-caspase-9 is activated and converted into caspase-9, which in turn activates other caspases, such as caspases-3, -6, and -7, through proteolytic conversion of proenzymes into their active forms. Activation of caspases leads to fragmentation of cellular proteins, DNA degradation [18], changes in cytoskeletal structure, and formation of apoptotic vesicles, resulting in controlled cell breakdown without inducing inflammation [36].
Extracts of various species of Epilobium inhibit the proliferation of human prostatic epithelial cells in vitro by affecting the progression of the cell cycle [37]. The active compounds – ellagitannins reduce the growth of cancer cells by inhibiting proliferation and inducing apoptotic cell death [18]. They exhibit cytotoxicity against cancer cell lines by disrupting the balance between proliferation and apoptosis. In this process, induction of apoptotic cell death occurs through DNA fragmentation and cleavage of cytokeratin 18 by activated caspases [18]. Furthermore, oenothein B is a potent inhibitor of 5α-reductase and aromatase, enzymes involved in the etiology of benign prostatic hyperplasia [38]. This prevents the development of inflammation and disturbance of cell division balance within the prostate gland. Oenothein B induces neutral endopeptidase in PC-3 prostate cancer cells, which inactivates growth-stimulating neuropeptides. Oenothein B is responsible for inhibiting poly(ADP-ribose) polymerase on specific chromosomal proteins in eukaryotic cells, mainly through poly(ADP-ribose) glycohydrolase. This process is a significant factor in regulating gene activation, DNA replication, transcription, and cell death. Ellagitannins exert an antimutagenic activity toward the direct mutagen medication mitomycin C. [39]. In another study, antigenotoxic and anticytotoxic effects against DNA damage induced by cyclophosphamide were studied [40]. The authors concluded that oenothein B protects DNA from CPA-induced damaging factors and induces DNA repair after damage [40]. Additionally, Miyamoto et al. concluded that ellagitannins with a marked antitumor activity should possess a dimeric structure with several galloyl groups on the glucose core [41]. Later authors demonstrated that oenothein B has anticancer activity through host-mediated induction of cytotoxic peritoneal exudate cells, via activation of macrophages and secreting interleukin 1β [42]. Agrimoniin, an antitumor tannin of Agrimonia pilosa Ledeb. (Rosaceae) like oenothein B induces interleukin-1 [43,44]. Evening primrose (Oenothera biennis L, Onagraceae) root extract and its active compound, oenothein B targeting the PD-1/PD-L1 blockade in the in vivo experiments utilizing a colorectal cancer mouse model [45].
Stolarczyk et al. [5] demonstrated that macrocyclic ellagitannins, including oenothein B, may exhibit prooxidant activity in prostate cancer cells. Oenothein B induces elevated levels of ROS [5,18], which in turn may contribute to the activation of intrinsic apoptosis through mitogen-activated protein kinase and/or phosphatidylinositol 3-kinase pathways. In response to increased ROS levels, cells can initiate defensive and signalling mechanisms that may beneficially affect their ability to adapt to changing environmental conditions. However, excessive ROS and its chronic maintenance can lead to oxidative stress, where the balance between ROS production and the body’s antioxidant mechanisms is disrupted in favor of ROS. Oxidative stress is associated with accelerated aging processes and the development of many chronic diseases, including hypertension. Therefore, understanding the impact of concentration and duration of supplementation with the extracts from Epilobium herbs requires further research.
Oenothein B participates in stimulating peripheral blood mononuclear cells (PBMCs) and releases IL-1β, IL-2, and TNF-α depending on the dose [46]. It also activates T lymphocytes through IL-1β, which are responsible for eliciting a host-mediated anticancer response rather than direct cytotoxic effects on cancer cells [44]. Oenothein B increases the expression of BAX, BAK, and BAD, p53, cytochrome c (cytoplasmic), and PARP, activates caspase-3 and 9, and decreases the expression of BCL-2, FADD, TRADD, TNFRSF10A, and TNFRSF10C [46]. In another study 5 Epilobium species (E. hirsutum L., E. parviflorum Schreb., E. palustre L. E. dodonaei Vill., and E. angustifolium L.) extracts were studied and all of them demonstrated good antioxidant activity, evaluated by DPPH and TEAC assays [47]. From the evaluated optimized extracts, E. angustifolium aerial parts had the highest selectivity toward killing cancerous cells (prostate carcinoma cell lines), followed by E. hirsutum (commonly known as hairy willowherb) aerial parts extract. For the antioxidant effect, E. hirsutum leaves and E. hirstum aerial parts extracts displayed the highest potency, decreasing ROS. The highest anti-inflammatory potential, based on the IL-6 and IL-8 levels, was displayed by E. dodonaei aerial parts and E. angustifolium leaf extracts [47]. E. hirsutum L. extract also exhibited anti-inflammatory and antitumor effects in the studied animal models [48].
3.2.4. Other than Colorectal Cancer Activity
Epilobium possesses cytotoxic properties, impacting the reduction of malignant melanoma (an aggressive type of skin cancer) [49], sarcoma [17], breast cancer (MCF-7) [50], prostate cancer (PC-3 and LNCaP) [38], lung cancer (A549), colon cancer (HT-29), as well as glioblastoma (1231N1) and neuroblastoma (SK-N-SH), human oral cavity epidermis (KB), cervical cancer (HeLa), and liver cancer cell lines (Hep-3B), with lower cytotoxicity towards normal cell lines (WISH) [49], see Figure 3.
3.2.5. Nuclear factor (NF-κB)
Oenothein B (from E. angustifolium) is responsible for the induction of intracellular Ca2+ influx, activation of NF-κB, and production of ROS and pro-inflammatory cytokines [46]. A low level of ROS is essential for maintaining redox balance and cell proliferation. High levels of ROS can activate PI3K/Akt signalling mainly by inhibiting phosphatases such as PTEN or directly activating oncogenes, including AKT. The PI3K/Akt signalling pathway is a mediator of NF-κB, which plays a significant role in cell proliferation, cell cycle progression, and cell viability in cancer. By increasing ROS levels, oenothein B may prevent cell proliferation and stimulation of cell death of lung cancer, likely through the PI3K/Akt/NF-κB signalling pathway. This is opposite to ROS inhibitors like N-acetyl-L-cysteine and the PI3K agonist (insulin-like growth factor 1, IGF-1) [51]. It has also been noted that oenothein B can induce keratinocyte growth and stimulate neutrophil influx.
It has been proven that oenothein B dose-dependently reduces NO production, inducible nitric oxide synthase (iNOS), and iNOS protein levels without inhibiting iNOS enzymatic activity using murine macrophages [16,52]. This action is NF-κB dependent but independent of the interferon (IFN)-γ/JAK-STAT pathway. As is known, improper or excessive NO production by iNOS is closely linked to numerous inflammatory diseases, making oenothein B a promising lead in developing therapeutic agents as effective inhibitors of NO production.
3.2.6. Lymphocytes Cells and Interferons (IFNs)
Ramstead et al. reported that oenothein B stimulated innate γδ T cells, αβ T cells, and NK cells, leading to increased expression of CD25 and/or CD69. This pathway increases IFN-γ production by NK cells and T lymphocytes [53]. In another study, aging influences the response of T cells to stimulation by the oenothein B [54]. Studies by Caillon et al. determined that a small percentage of γδ T lymphocytes in circulation contribute to hypertension mediated by angiotensin II [55]. Gene expression microarray analysis of whole blood in patients with or without coronary artery disease revealed a positive correlation between the frequency of γδ T cells in peripheral blood and systolic blood pressure. Yoshimura et al. noted that oenothein B dose-dependently reduces IFN-γ levels, suggesting its involvement in regulating systolic pressure through changes in IFN-γ levels [28].
3.2.7. Dendritic Cells
Moreover, Yoshimura et al. reported the immunomodulatory effect of oenothein B on human dendritic cells (DCs), present in various tissues in contact with the external environment, such as the skin, nose, lungs, stomach, and intestines [28]. These cells play critical roles in the initial immune response as antigen-presenting cells. Immature DCs have strong phagocytic abilities, and after antigen uptake and maturation, mature DCs migrate to lymph nodes and present antigens to naive T cells. DCs, macrophages, and B cells are involved in antigen presentation with MHC class II molecules, thus called professional antigen-presenting cells. Another important function of cytokine production inherently associated with DCs is its constitutive role in initiating inflammation associated with certain autoimmune diseases, including inflammatory bowel diseases, celiac disease, and rheumatoid arthritis. Oenothein B down-regulates CD1a and CD83 cell surface molecules and cytokines IL- 1β, IL-6, IL-12, IL-17, IFN-γ, and MIP-1β, dose-dependently inhibiting inflammatory processes. Although oenothein B has been noted to accelerate inflammatory cytokine release from monocytes, significant suppression of IL-6 production below detectable levels and downregulation of IL-1β may thus induce the anti-inflammatory effects of tannins via DCs. This study demonstrated that tannins exert peripheral anti-inflammatory effects by reducing cytokines and inducing dysfunction and apoptosis in DCs. Oenothein B suppresses cell surface molecules, delaying antigen presentation, lowering cytokine production, and inducing their apoptosis. Activation of caspases-3/7, 8, and 9 suggests a caspase-dependent mechanism of this apoptosis [10,20].
3.2.8. Brain Inflammation
Okuyama et al. suggested that oenothein B can reduce nerve inflammation in the brain during systemic inflammation [29,30]. Oenothein B itself barely crosses the blood-brain barrier (BBB), but metabolites produced by gut microflora can cross the BBB and act directly in the brain as anti-inflammatory agents. Ellagitannins are converted in the intestine to ellagic acid (2,3,7,8- tetrahydroxy-benzopyrano [5,4,3-cde] benzopyran-5-10-dione), which is then converted into metabolites such as urolithins (i.e., urolithin A, 3,8- dihydroxyurolithin, and urolithin B, 3-hydroxyurolithin) by gut bacteria [56]. According to recent studies, ellagic acid has neuroprotective functions in the brain, preventing cognitive deficits, hippocampal deficits, and amnesia. Urolithin A performs neuroprotective functions, protecting against ischemic neuronal damage by enhancing autophagy.
3.2.9. cAMP-Responsive Transcription Factor (CREB)
Oenothein B activates CREB by phosphorylating the serine residue via various kinases, such as protein kinase A, Akt/protein kinase B, Ca2+/calmodulin-dependent protein kinase, and glycogen synthase kinase-3 [29,30,55]. Garat et al. reported that CREB depletion in smooth muscle cells contributes to medial thickening, adventitial fibrosis, and pulmonary hypertension. CREB deficiency led to structural and hemodynamic changes characteristic of pulmonary hypertension (PH) in vivo. Rats with reduced CREB levels showed serum-independent proliferation and hypertrophy in vitro, and they secreted soluble factors that stimulated proliferation and extracellular matrix protein expression by adventitial fibroblasts. This indicates that CREB plays a crucial role in regulating the pathological transformation of arterial smooth muscle cells (SMCs) from a homeostatic, resting form to a proliferative, synthetic form, driving arterial remodelling and contributing to PH development [57].
3.2.10. Antimicrobial Activity
Epilobium extracts possess anti-inflammatory and antiseptic properties effective against the fungi: Candida albicans [58], Microsporum canis, Trichophyton tonsurans, dermatophytes Arthroderma spp. [59], and is beneficial within skin ulcers, swelling [60], and wound healing [4].
Moreover, several reports characterized the beneficial effects of Epilobium active compounds, ellagitannins towards variety of bacteria: Escherichia coli [1,59], Staphylococcus aureus [59,61], Pseudomonas aeruginosa [59,60], Bacillus cereus, Micrococcus luteus, Klebsiella pneumoniae and Acinetobacter baumannii [59], protozoa Leishmania donovani [62], and fungi Paracoccidioides brasiliensis [63]. Particularly noteworthy is the synergistic action of polyphenols with antibiotics against antibiotic-resistant bacteria. Methicillin-resistant Staphylococcus aureus (MRSA) has shown sensitivity to β-lactam antibiotics combined with oenothein B [61].
4. Conclusions
E. parviflorum offers a wide range of health benefits, primarily due to its rich content of bioactive compounds like flavonoids, phenolic acids and hydrolysable tannins (oenothein B). Its applications in traditional and modern medicine highlight its potential in treating prostate disorders, reducing inflammation, combating oxidative stress, and offering anticancer and antimicrobial effects. E. parviflorum influences the production and storage of NO, which may increase nitrosyl hemoglobin (Hb) synthesis (since NO can react directly with the heme of Hb), facilitating the transport of NO to areas with low oxygen levels. This, in turn, promotes vasodilation and blood pressure regulation. Moreover, extracts from Epilobium alleviate the effects of excess ROS and inflammation, thereby protecting against oxidative stress and related vascular diseases. The limited number of experimental studies regarding the effects of E. parviflorum on the vascular system justifies the need for further scientific research in this field. Therefore, it seems reasonable to pursue additional studies on E. parviflorum to fully understand its therapeutic potential and optimize its use in clinical settings.
Author Contributions
Conceptualization, K.L. and M.M.; methodology, K.L. and M.M.; software, K.L. and M.M.; validation, K.L. and M.M.; formal analysis, K.L. and M.M.; investigation, K.L..; resources, M.M.; data curation, M.M.; writing—original draft preparation, K.L.; writing—review and editing, K.L. and M.M.; visualization, K.L.; supervision, M.M.; project administration, K.L. and M.M.; funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Minister of Science under “the Regional Initiative of Excellence Program”.
Conflicts of Interest
Non-declared.
References
- Steenkamp, V.; Gouws, M.C.; Gulumian, M.; Elgorashi, E.E.; van Staden, J. Studies on antibacterial, anti-inflammatory and antioxidant activity of herbal remedies used in the treatment of benign prostatic hyperplasia and prostatitis. J Ethnopharmacol 2006, 103, 71–75. [Google Scholar] [CrossRef]
- Barbarossa, A.; Rosato, A.; Carocci, A.; Arpini, S.; Bosisio, S.; Pagni, L.; Piatti, D.; Spinozzi, E.; Angeloni, S.; Sagratini, G.; Zengin, G.; Cespi, M.; Maggi, F.; Caprioli, G. Efficacy of Willow Herb (Epilobium angustifolium L. and E. parviflorum Schreb.) Crude and Purified Extracts and Oenothein B Against Prostatic Pathogens. Antibiotics (Basel) 2025, 14, 117. [Google Scholar] [CrossRef]
- Deng, L.; Zong, W.; Tao, X.; Liu, S.; Feng, Z.; Lin, Y.; Liao, Z.; Chen, M. Evaluation of the therapeutic effect against benign prostatic hyperplasia and the active constituents from Epilobium angustifolium L. J Ethnopharmacol 2019, 232, 1–10. [Google Scholar] [CrossRef]
- Karakaya, S.; Süntar, I.; Yakinci, O.F.; Sytar, O.; Ceribasi, S.; Dursunoglu, B.; Ozbek, H.; Guvenalp, Z. In vivo bioactivity assessment on Epilobium species: A particular focus on Epilobium angustifolium and its components on enzymes connected with the healing process. J Ethnopharmacol 2020, 262, 113207. [Google Scholar] [CrossRef]
- Stolarczyk, M.; Naruszewicz, M.; Kiss, A.K. Extracts from Epilobium sp. herbs induce apoptosis in human hormone-dependent prostate cancer cells by activating the mitochondrial pathway. J Pharm Pharmacol 2013, 65, 1044–1054. [Google Scholar] [CrossRef]
- Gevrenova, R.; Zengin, G.; Ozturk, G.; Zheleva-Dimitrova, D. Exploring the Phytochemical Profile and Biological Insights of Epilobium angustifolium L. Herb. Plants (Basel) 2025, 14, 415. [Google Scholar] [CrossRef]
- Kiss, A.K.; Bazylko, A.; Filipek, A.; Granica, S.; Jaszewska, E.; Kiarszys, U.; Kośmider, A.; Piwowarski, J. Oenothein’s contribution to the anti-inflammatory and antioxidant activity of Epilobium sp. Phytomedicine 2011, 18, 557–560. [Google Scholar] [CrossRef]
- Hiermann, A.; Juan, H.; Sametz, W. Influence of Epilobium extracts on prostaglandin biosynthesis and carrageenin induced oedema of the rat paw. J Ethnopharmacol 1986, 17, 161–169. [Google Scholar] [CrossRef]
- Yoshida, T.; Yoshimura, M.; Amakura, Y. Chemical and Biological Significance of Oenothein B and Related Ellagitannin Oligomers with Macrocyclic Structure. Molecules 2018, 23, 552. [Google Scholar] [CrossRef]
- Isla, K.K.Y.; Tanae, M.M.; de Lima-Landman, M.T.R.; de Magalhães, P.M.; Lapa, A.J.; Souccar, C. Vasorelaxant effects of ellagitannins isolated from Cuphea carthagenensis. Planta Med 2024, 90, 276–285. [Google Scholar] [CrossRef]
- Cosby, K.; Partovi, K.S.; Crawford, J.H.; Patel, R.P.; Reiter, C.D.; Martyr, S.; Yang, B.K.; Waclawiw, M.A.; Zalos, G.; Xu, X.; Huang, K.T.; Shields, H.; Kim-Shapiro, D.B.; Schechter, A.N.; Cannon, R.O., 3rd; Gladwin, M.T. , 3rd; Gladwin, M.T. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med 2003, 9, 1498–1505. [Google Scholar] [CrossRef]
- Kim, H.Y.; Oh, H.; Li, X.; Cho, K.W.; Kang, D.G.; Lee, H.S. Ethanol extract of seeds of Oenothera odorata induces vasorelaxation via endothelium-dependent NO-cGMP signaling through activation of Akt-eNOS-sGC pathway. J Ethnopharmacol 2011, 133, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Shikov, A.N.; Poltanov, E.A.; Dorman, H.J.; Makarov, V.G.; Tikhonov, V.P.; Hiltunen, R. Chemical composition and in vitro antioxidant evaluation of commercial water-soluble willow herb (Epilobium angustifolium L.) extracts. J Agric Food Chem 2006, 54, 3617–3624. [Google Scholar] [CrossRef]
- Stajner, D. ; Popović; B M., Boza, P. Evaluation of willow herb's (Epilobium angustofolium L.) antioxidant and radical scavenging capacities. Phytother Res 2007, 21, 1242–1245. [Google Scholar] [CrossRef]
- Hevesi, B.T.; Houghton, P.J.; Habtemariam, S.; Kéry, A. Antioxidant and antiinflammatory effect of Epilobium parviflorum Schreb. Phytother Res 2009, 23, 719–724. [Google Scholar] [CrossRef]
- Merighi, S.; Travagli, A.; Tedeschi, P.; Marchetti, N.; Gessi, S. Antioxidant and Antiinflammatory Effects of Epilobium parviflorum, Melilotus officinalis and Cardiospermum halicacabum Plant Extracts in Macrophage and Microglial Cells. Cells 2021, 10, 2691. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.L.; Wang, C.C.; Yen, K.Y.; Yoshida, T.; Hatano, T.; Okuda, T. Antitumor activities of ellagitannins on tumor cell lines. Basic Life Sci 1999, 66, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, H.; Jiang, Y.; Kusama, K.; Atsumi, T.; Ueha, T.; Toguchi, M.; Iwakura, I.; Satoh, K.; Ito, H.; Hatano, T.; Yoshida, T. Cytotoxic activity of hydrolyzable tannins against human oral tumor cell lines--a possible mechanism. Phytomedicine 2000, 7, 39–47. [Google Scholar] [CrossRef]
- Kumagai, T.; Kawamoto, Y.; Nakamura, Y.; Hatayama, I.; Satoh, K.; Osawa, T.; Uchida, K. 4-hydroxy-2-nonenal, the end product of lipid peroxidation, is a specific inducer of cyclooxygenase-2 gene expression. Biochem Biophys Res Commun 2000, 273, 437–441. [Google Scholar] [CrossRef]
- Feng, L.; Xia, Y.; Garcia, G. E. , Hwang, D., Wilson, C. B. Involvement of reactive oxygen intermediates in cyclooxygenase-2 expression induced by interleukin-1, tumor necrosis factor-alpha, and lipopolysaccharide. J Clin Invest 1995, 95, 1669–1675. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T. Pharmacologically active tannins isolated from medicinal plants. Basic Life Sci 1992, 59, 539–569. [Google Scholar] [CrossRef]
- Rolnik, A.; Olas, B.; Szablińska-Piernik, J.; Lahuta, L.B.; Gromadziński, L.; Majewski, M.S. Antioxidant and anticoagulant properties of myo-inositol determined in an ex vivo studies and gas chromatography analysis. Sci Rep 2024, 14, 25633. [Google Scholar] [CrossRef]
- Wojtacha, P.; Bogdańska-Chomczyk, E.; Majewski, M.K.; Obremski, K.; Majewski, M.S.; Kozłowska, A. Renal Inflammation, Oxidative Stress, and Metabolic Abnormalities During the Initial Stages of Hypertension in Spontaneously Hypertensive Rats. Cells 2024, 13, 1771. [Google Scholar] [CrossRef]
- Żary-Sikorska, E.; Fotschki, B.; Kołodziejczyk, K.; Jurgoński, A.; Kosmala, M.; Milala, J.; Majewski, M.; Ognik, K.; Juśkiewicz, J. Strawberry phenolic extracts effectively mitigated metabolic disturbances associated with high-fat ingestion in rats depending on the ellagitannin polymerization degree. Food Funct 2021, 12, 5779–5792. [Google Scholar] [CrossRef]
- Rolnik, A.; Olas, B.; Szablińska-Piernik, J.; Lahuta, L.B.; Rynkiewicz, A.; Cygański, P.; Socha, K.; Gromadziński, L.; Thoene, M.; Majewski, M. Beneficial In Vitro Effects of a Low Myo-Inositol Dose in the Regulation of Vascular Resistance and Protein Peroxidation under Inflammatory Conditions. Nutrients 2022, 14, 1118. [Google Scholar] [CrossRef]
- Kiss, A.K.; Kapłon-Cieślicka, A.; Filipiak, K.J.; Opolski, G.; Naruszewicz, M. Ex vivo effects of an Oenothera paradoxa extract on the reactive oxygen species generation and neutral endopeptidase activity in neutrophils from patients after acute myocardial infarction. Phytother Res 2012, 26, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Granica, S.; Czerwińska, M.E.; Piwowarski, J.P.; Ziaja, M.; Kiss, A.K. Chemical composition, antioxidative and anti-inflammatory activity of extracts prepared from aerial parts of Oenothera biennis L. and Oenothera paradoxa Hudziok obtained after seeds cultivation. J Agric Food Chem 2013, 61, 801–810. [Google Scholar] [CrossRef]
- Yoshimura, M.; Akiyama, H.; Kondo, K.; Sakata, K.; Matsuoka, H.; Amakura, Y.; Teshima, R.; Yoshida, T. Immunological effects of Oenothein B, an ellagitannin dimer, on dendritic cells. Int J Mol Sci 2012, 14, 46–56. [Google Scholar] [CrossRef]
- Okuyama, S.; Furukawa, Y.; Yoshimura, M.; Amakura, Y.; Nakajima, M.; Yoshida, T. Oenothein B, a Bioactive Ellagitannin, Activates the Extracellular Signal-Regulated Kinase 2 Signaling Pathway in the Mouse Brain. Plants (Basel) 2021, 10, 1030. [Google Scholar] [CrossRef]
- Okuyama, S.; Makihata, N.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Nakajima, M.; Furukawa, Y. Oenothein B suppresses lipopolysaccharide (LPS)-induced inflammation in the mouse brain. Int J Mol Sci 2013, 14, 9767–9778. [Google Scholar] [CrossRef]
- Majewski, M.; Gromadziński, L.; Cholewińska, E.; Ognik, K.; Fotschki, B.; Juśkiewicz, J. Dietary Effects of Chromium Picolinate and Chromium Nanoparticles in Wistar Rats Fed with a High-Fat, Low-Fiber Diet: The Role of Fat Normalization. Nutrients 2022, 14, 5138. [Google Scholar] [CrossRef]
- Majewski, M.; Lis, B.; Juśkiewicz, J.; Ognik, K.; Jedrejek, D.; Stochmal, A.; Olas, B. The composition and vascular/antioxidant properties of Taraxacum officinale flower water syrup in a normal-fat diet using an obese rat model. J Ethnopharmacol 2021, 265, 113393. [Google Scholar] [CrossRef]
- Mortensen, M.B.; Dzaye, O.; Bøtker, H.E.; Jensen, J.M.; Maeng, M.; Bentzon, J.F.; Kanstrup, H.; Sørensen, H.T.; Leipsic, J.; Blankstein, R.; Nasir, K.; Blaha, M.J.; Nørgaard, B.L. Low-Density Lipoprotein Cholesterol Is Predominantly Associated With Atherosclerotic Cardiovascular Disease Events in Patients With Evidence of Coronary Atherosclerosis: The Western Denmark Heart Registry. Circulation 2023, 147, 1053–1063. [Google Scholar] [CrossRef]
- Żary-Sikorska, E.; Fotschki, B.; Jurgoński, A.; Kosmala, M.; Milala, J.; Kołodziejczyk, K.; Majewski, M.; Ognik, K.; Juśkiewicz, J. Protective Effects of a Strawberry Ellagitannin-Rich Extract against Pro-Oxidative and Pro-Inflammatory Dysfunctions Induced by a High- Fat Diet in a Rat Model. Molecules 2020, 25, 5874. [Google Scholar] [CrossRef]
- Xu, L.; Li, W.; Chen, S.Y.; Deng, X.W.; Deng, W.F.; Liu, G.; Chen, Y.J.; Cao, Y. Oenothein B ameliorates hepatic injury in alcoholic liver disease mice by improving oxidative stress and inflammation and modulating the gut microbiota. Front Nutr 2022, 9, 1053718. [Google Scholar] [CrossRef]
- Gozzelino, R.; Jeney, V.; Soares, M.P. Mechanisms of cell protection by heme oxygenase-1. Annu Rev Pharmacol Toxicol 2010, 50, 323–354. [Google Scholar] [CrossRef]
- Vitalone, A.; Guizzetti, M.; Costa, L.G.; Tita, B. Extracts of various species of Epilobium inhibit proliferation of human prostate cells. J Pharm Pharmacol 2003, 55, 683–690. [Google Scholar] [CrossRef]
- Lesuisse, D.; Berjonneau, J.; Ciot, C.; Devaux, P.; Doucet, B.; Gourvest, J.F.; Khemis, B.; Lang, C.; Legrand, R.; Lowinski, M.; Maquin, P.; Parent, A.; Schoot, B.; Teutsch, G. Determination of oenothein B as the active 5-alpha-reductase-inhibiting principle of the folk medicine Epilobium parviflorum. J Nat Prod 1996, 59, 490–492. [Google Scholar] [CrossRef]
- Silva, C.A.; Silva, C.R.; Véras, J.H.; Chen-Chen, L.; Ferri, P.H.; Santos, S.d.C. Genotoxicity and cytotoxicity evaluation of oenothein B and its protective effect against mitomycin C-induced mutagenic action. Mutat Res Genet Toxicol Environ Mutagen 2014, 767, 8–12. [Google Scholar] [CrossRef]
- Silva, C.A.; Véras, J.H.; Ventura, J.A.; de Melo Bisneto, A.V.; de Oliveira, M.G.; Cardoso Bailão, E.F.L.; ESilva, C.R.; Cardoso, C.G.; da Costa Santos, S.; Chen-Chen, L. Chemopreventive effect and induction of DNA repair by oenothein B ellagitannin isolated from leaves of Eugenia uniflora in Swiss Webster treated mice. J Toxicol Environ Health A 2023, 86, 929–941. [Google Scholar] [CrossRef]
- Miyamoto, K.; Kishi, N.; Koshiura, R.; Yoshida, T.; Hatano, T.; Okuda, T. Relationship between the structures and the antitumor activities of tannins. Chem Pharm Bull (Tokyo) 1987, 35, 814–822. [Google Scholar] [CrossRef]
- Miyamoto, K.; Nomura, M.; Sasakura, M.; Matsui, E.; Koshiura, R.; Murayama, T.; Furukawa, T.; Hatano, T.; Yoshida, T.; Okuda, T. Antitumor activity of oenothein B, a unique macrocyclic ellagitannin. Jpn J Cancer Res 1993, 84, 99–103. [Google Scholar] [CrossRef]
- Murayama, T.; Kishi, N.; Koshiura, R.; Takagi, K.; Furukawa, T.; Miyamoto, K. Agrimoniin, an antitumor tannin of Agrimonia pilosa Ledeb., induces interleukin-1. Anticancer Res 1992, 12, 1471–1474. [Google Scholar]
- Miyamoto, K.; Murayama, T.; Nomura, M.; Hatano, T.; Yoshida, T.; Furukawa, T.; Koshiura, R.; Okuda, T. Antitumor activity and interleukin-1 induction by tannins. Anticancer Res 1993, 13, 37–42. [Google Scholar]
- Lee, E.J.; Kim, Y.S.; Kim, J.H.; Woo, K.W.; Park, Y.H.; Ha, J.H.; Li, W.; Kim, T.I.; An, B.K.; Cho, H.W.; Han, J.H.; Choi, J.G.; Chung, H.S. Uncovering the colorectal cancer immunotherapeutic potential: Evening primrose (Oenothera biennis) root extract and its active compound oenothein B targeting the PD-1/PD-L1 blockade. Phytomedicine 2024, 125, 155370. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Kirpotina, L.N.; Jakiw, L.; Khlebnikov, A.I.; Blaskovich, C.L.; Jutila, M.A.; Quinn, M.T. Immunomodulatory activity of oenothein B isolated from Epilobium angustifolium. J Immunol 2009, 183, 6754–6766. [Google Scholar] [CrossRef]
- Vlase, A.M.; Toiu, A. ; Tomuță; I; Vlase, L.; Muntean, D.; Casian, T.; Fizeșan, I.; Nadăș; GC; Novac, C.Ș.; Tămaș; M; Crișan, G. Epilobium Species: From Optimization of the Extraction Process to Evaluation of Biological Properties. Antioxidants (Basel) 2022, 12, 91. [Google Scholar] [CrossRef]
- Vlase, A.M.; Toiu, A.; Gligor, O.; Muntean, D.; Casian, T.; Vlase, L.; Filip, A.; Bȃldea, I.; Clichici, S.; Decea, N.; Moldovan, R.; Toma, V.A.; Virag, P.; Crișan, G. Investigation of Epilobium hirsutum L. Optimized Extract's Anti-Inflammatory and Antitumor Potential. Plants (Basel) 2025, 13, 198. [Google Scholar] [CrossRef]
- Kyriakou, S.; Tragkola, V.; Paraskevaidis, I.; Plioukas, M.; Trafalis, D.T.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Chemical Characterization and Biological Evaluation of Epilobium parviflorum Extracts in an In Vitro Model of Human Malignant Melanoma. Plants (Basel) 2023, 12, 1590. [Google Scholar] [CrossRef]
- Hatefi Kia, B.; Kazemi Noureini, S.; Vaezi Kakhki, M.R. The Extracts of Epilobium Parviflorum Inhibit MCF-7 Breast Cancer Cells. Iran J Toxicol 2021, 15, 65–72. [Google Scholar] [CrossRef]
- Pei, X.; Xiao, J.; Wei, G.; Zhang, Y.; Lin, F.; Xiong, Z.; Lu, L.; Wang, X.; Pang, G.; Jiang, Y.; Jiang, L. Oenothein B inhibits human non-small cell lung cancer A549 cell proliferation by ROS-mediated PI3K/Akt/NF-κB signaling pathway. Chem Biol Interact 2019, 298, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Schmid, D.; Gruber, M.; Piskaty, C.; Woehs, F.; Renner, A.; Nagy, Z.; Kaltenboeck, A.; Wasserscheid, T.; Bazylko, A.; Kiss, A.K.; Moeslinger, T. Inhibition of NF-κB-dependent cytokine and inducible nitric oxide synthesis by the macrocyclic ellagitannin oenothein B in TLR-stimulated RAW 264.7 macrophages. J Nat Prod 2012, 75, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Ramstead, A.G.; Schepetkin, I.A.; Quinn, M.T.; Jutila, M.A. Oenothein B, a cyclic dimeric ellagitannin isolated from Epilobium angustifolium, enhances IFNγ production by lymphocytes. PLoS One 2012, 7, e50546. [Google Scholar] [CrossRef] [PubMed]
- Ramstead, A.G.; Schepetkin, I.A.; Todd, K.; Loeffelholz, J.; Berardinelli, J.G.; Quinn, M.T.; Jutila, M.A. Aging influences the response of T cells to stimulation by the ellagitannin, oenothein B. Int Immunopharmacol 2015, 26, 367–377. [Google Scholar] [CrossRef]
- Caillon, A.; Mian, M.O.R.; Fraulob-Aquino, J.C.; Huo, K.G.; Barhoumi, T.; Ouerd, S.; Sinnaeve, P.R.; Paradis, P.; Schiffrin, E.L. γδ T Cells Mediate Angiotensin II-Induced Hypertension and Vascular Injury. Circulation 2017, 135, 2155–2162. [Google Scholar] [CrossRef]
- Iglesias-Aguirre, C.E.; García-Villalba, R.; Beltrán, D.; Frutos-Lisón, M.D.; Espín, J.C.; Tomás-Barberán, F.A.; Selma, M.V. Gut Bacteria Involved in Ellagic Acid Metabolism To Yield Human Urolithin Metabotypes Revealed. Agric Food Chem 2023, 71, 4029–4035. [Google Scholar] [CrossRef]
- Garat, C.V.; Majka, S.M.; Sullivan, T.M.; Crossno, J.T.; Jr Reusch, J.E.B.; Klemm, D.J. CREB depletion in smooth muscle cells promotes medial thickening, adventitial fibrosis and elicits pulmonary hypertension. Pulm Circ 2020, 10, 2045894019898374. [Google Scholar] [CrossRef]
- Hiermann, A.; Reidlinger, M.; Juan, H.; Sametz, W. Isolierung des antiphlogistischen Wirkprinzips von Epilobium angustifolium [Isolation of the antiphlogistic principle from Epilobium angustifolium]. Planta Med 1991, 57, 357–360. [Google Scholar] [CrossRef]
- Dreger, M.; Adamczak, A.; Foksowicz-Flaczyk, J. Antibacterial and Antimycotic Activity of Epilobium angustifolium L. Extracts: A Review. Pharmaceuticals (Basel) 2023, 16, 1419. [Google Scholar] [CrossRef]
- Battinelli, L.; Tita, B.; Evandri, M.G.; Mazzanti, G. Antimicrobial activity of Epilobium spp. extracts. Farmaco 2001, 56, 345–348. [Google Scholar] [CrossRef]
- Zhao, L.; Zhou, A.; Liu, Z.; Xiao, J.; Wang, Y.; Cao, Y.; Wang, L. Inhibitory mechanism of lactoferrin on antibacterial activity of oenothein B: isothermal titration calorimetry and computational docking simulation. J Sci Food Agric 2020, 100, 2494–2501. [Google Scholar] [CrossRef] [PubMed]
- Kolodziej, H.; Kayser, O.; Kiderlen, A.F.; Ito, H.; Hatano, T.; Yoshida, T.; Foo, L.Y. Antileishmanial activity of hydrolyzable tannins and their modulatory effects on nitric oxide and tumour necrosis factor-alpha release in macrophages in vitro. Planta Med 2001, 67, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.D.; Ferri, P.H.; Santos, S.C.; Bao, S.N.; Soares, C.M.; Pereira, M. Oenothein B inhibits the expression of PbFKS1 transcript and induces morphological changes in Paracoccidioides brasiliensis. Med Mycol 2007, 45, 609–618. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Illustration of the E. parviflorum Schreb. (Onagraceae) therapeutic properties (edited using BioRender.com) ↓: decrease, ↑: increase.
Figure 1.
Illustration of the E. parviflorum Schreb. (Onagraceae) therapeutic properties (edited using BioRender.com) ↓: decrease, ↑: increase.
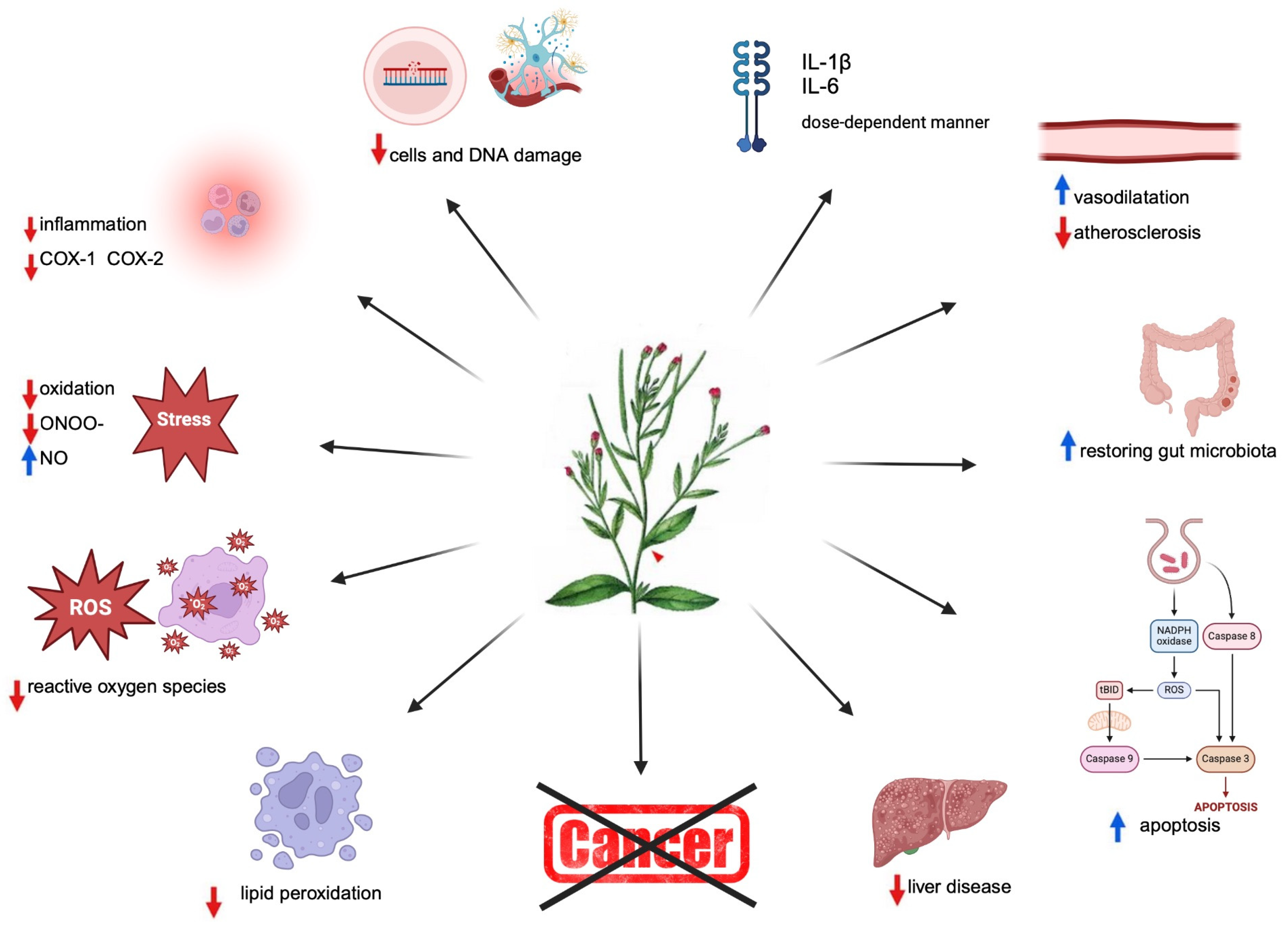
Figure 2.
Schematic of oxidative stress and reactive oxygen species (ROS) formation. An imbalance between ROS production and the body’s ability to neutralize them (via antioxidant enzymes like superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase) leads to oxidative stress. Cellular Signaling: Small amounts of ROS are used as signaling molecules in cells. Immune Defense: ROS are used by immune cells to kill pathogens. Cell Damage: Excessive ROS can lead to damage to lipids, proteins, and DNA, which is associated with various diseases, including cancers, neurodegenerative diseases, and cardiovascular diseases.
Figure 2.
Schematic of oxidative stress and reactive oxygen species (ROS) formation. An imbalance between ROS production and the body’s ability to neutralize them (via antioxidant enzymes like superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase) leads to oxidative stress. Cellular Signaling: Small amounts of ROS are used as signaling molecules in cells. Immune Defense: ROS are used by immune cells to kill pathogens. Cell Damage: Excessive ROS can lead to damage to lipids, proteins, and DNA, which is associated with various diseases, including cancers, neurodegenerative diseases, and cardiovascular diseases.

Figure 3.
Illustration of the E. parviflorum Schreb. (Onagraceae) effect on the inhibition of tumor progression (edited using BioRender.com).
Figure 3.
Illustration of the E. parviflorum Schreb. (Onagraceae) effect on the inhibition of tumor progression (edited using BioRender.com).
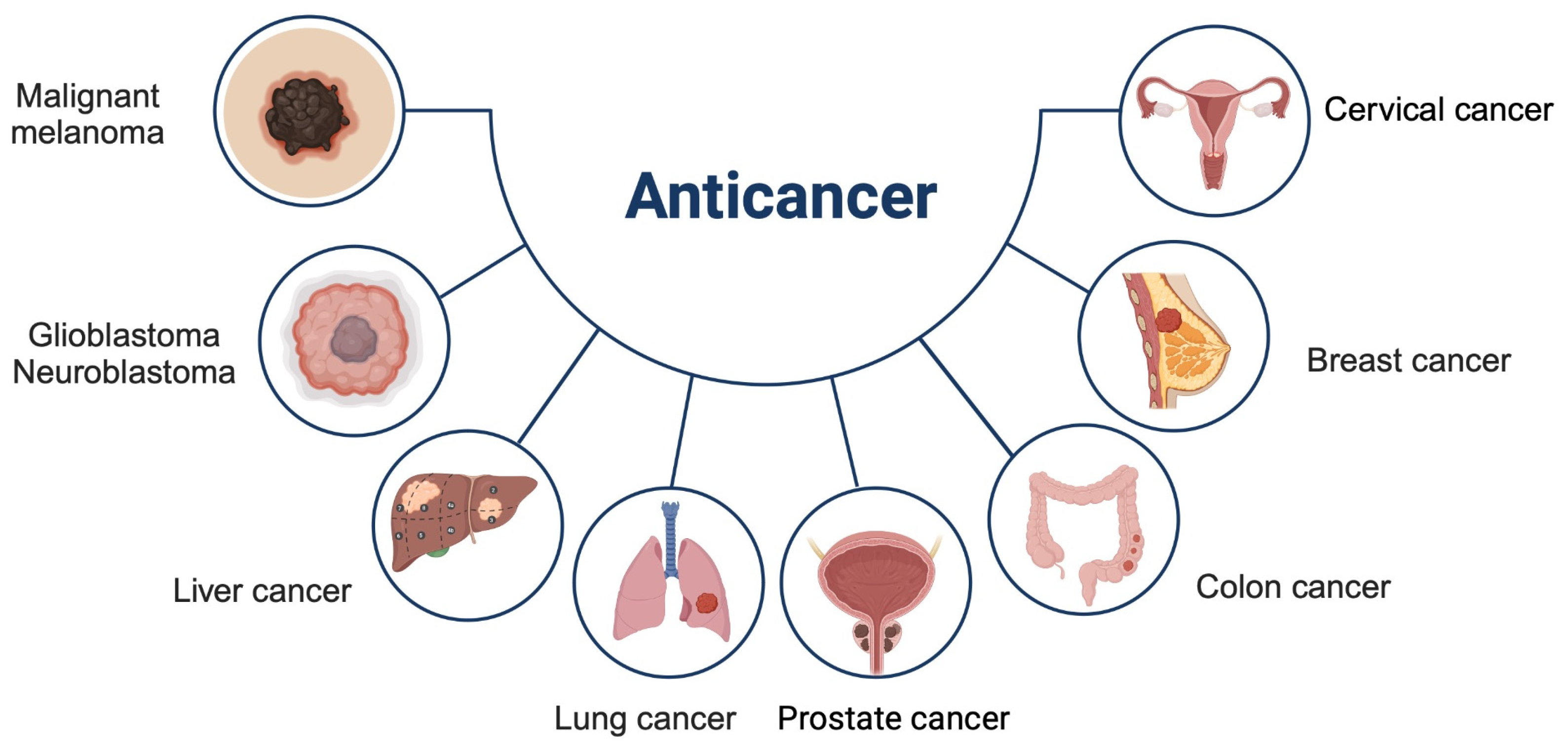

Table 1.
Vascular research results.
| Study | Plant | Material | Model (ex vivo/in vitro) |
Intervention | Duration | Parameters measured | Effect | Mechanism |
|---|---|---|---|---|---|---|---|---|
| Isla et al. 2024 [10] |
Cuphea carthagenensis (Jacq.) J. F. Macbr (Lythraceae) |
aqueous extract (AE) | Supplementation to rats | 0.5 and 1.0 g/kg/day | 1-week | systolic blood pressure (the non-invasive tail-cuff method) |
hypotensive effect | unknown, ellagitannin independent due to the low oral bioavailability of ellagitannins |
| oenothein B woodfordin C eucalbanin B isolated from AE | Ex vivo – rat aortic rings pre-contracted with vasoconstrictor noradrenaline | 20 – 180 µM | vasorelaxation | concentration-related vasorelaxation | Endothelium dependent, via activating NO synthesis/NO release from endothelial cells without alteration of Ca2+ influx in vascular smooth muscle preparations |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



