
Submitted:
23 April 2025
Posted:
24 April 2025
You are already at the latest version
Abstract
Cross-reactivity between house dust mites and scabies mites is a clinically significant but often underestimated phenomenon that can complicate both the management of respiratory allergies and the diagnosis of parasitic infections such as scabies. The antigenic properties shared between these species are supported by a bioinformatic analysis we conducted, as well as by proteomic and immunological studies reported in the literature, which highlight structurally similar proteins capable of inducing cross-immune responses. This article provides a comprehensive analysis of the main allergens involved, focusing on the molecular mechanisms underlying cross-reactivity, its clinical implications, and potential diagnostic and therapeutic applications.
Keywords:
cross-reactivity
; Sarcoptes scabiei
; dust mites
; bioinformatic analysis
1. Introduction
Dust mites, particularly Dermatophagoides pteronyssinus and Dermatophagoides farinae, are among the leading causes of respiratory allergies worldwide [1,2]. It is estimated that dust mite allergy affects 1–2% of the global population, equivalent to 65–130 million people worldwide [3].
Scabies mites, primarily Sarcoptes scabiei, are ectoparasites responsible for scabies, a condition characterized by intense itching and skin lesions [4,5]. The incidence of scabies is increasing in many regions of the world, although epidemiological data are often imprecise due to the lack of mandatory reporting in many countries [6]. According to the 2015 Global Burden of Disease study, approximately 204 million people worldwide are affected by scabies [7].
Sarcoptes scabiei is an obligate parasite that burrows into the stratum corneum of human skin to live and reproduce, causing localized inflammation and immune responses against its excretory products [4]. The disease is more common in overcrowded environments with poor hygiene conditions, typically found in economically disadvantaged areas. However, scabies is not limited to developing countries; in developed nations, it frequently presents as outbreaks in institutions such as nursing homes and hospitals. The prevalence is higher among children, highlighting a particular vulnerability in this age group. Transmission primarily occurs through direct skin-to-skin contact, which requires prolonged exposure, making household members or individuals with close physical contact particularly at risk. In cases of crusted scabies, transmission can also occur via fomites (e.g., clothing, bedding, or towels) due to the extremely high number of mites present. Sexual transmission is also possible, classifying scabies as a sexually transmitted disease in certain contexts [7].
Despite their differences in habitat and biological functions, these mites share numerous antigenic proteins that can induce cross-reactive IgE responses in sensitized individuals [8,9,10,11]. The immune response generated by Sarcoptes scabiei infestation includes the production of IgE antibodies directed against its allergens, which exhibit significant cross-reactivity with the major allergens of house dust mites. For this reason, positive immediate skin tests to house dust mite allergen extracts should be carefully evaluated to avoid misdiagnosis [12].
This article examines the molecular basis of cross-reactivity between dust mites and scabies mites, leveraging new bioinformatic approaches. It explores the diagnostic challenges posed by cross-reactivity and discusses therapeutic interventions, including allergen-specific immunotherapy and novel diagnostic tools based on recombinant allergens.
2. Global Distribution and Health Implications of Different Mite Species
Mites constitute a vast group of arthropods that have colonized a wide range of terrestrial habitats, adapting to diverse environmental conditions and developing equally varied feeding behaviors. Their presence has been documented in both natural and human-made environments, where they can directly interact with humans and contribute to public health concerns.
The allergens of house dust mites, such as Dermatophagoides pteronyssinus, Dermatophagoides farinae, and Euroglyphus maynei, have been extensively studied. However, the degree of cross-reactivity between their allergens and those of other mite species remains unclear. It is known, however, that E. maynei, D. pteronyssinus, and D. farinae are among the main species involved in allergic sensitization and share significant cross-allergenic reactivity, particularly through specific allergens like Der p 2 [13].
The medically relevant mite species can be categorized into three major groups:
- House dust mites
- Storage mites
- Parasitic or ectoparasitic mites
Each of these groups has a specific geographical distribution and distinct health implications [14]. These characteristics are summarized in Table 1.

Table 1.
Characteristics of Different Mite Species.
| Category | Characteristics | Species | Family | Environments | Geographical distribution | Related pathologies | References |
|---|---|---|---|---|---|---|---|
| House Dust Mite | Present in domestic environments, where they feed on human dandruff and other organic debris | Dermatophagoides pteronyssinus | Pyroglyphidae | Temperate and humid climates | Europe (Italy, France, United Kingdom), Asia, and Oceania | Allergic rhinitis, asthma, and atopic dermatitis | Fernández-Caldas E, Iraola Calvo V. Mite allergens. Curr Allergy Asthma Rep. 2005 Sep;5(5):402-10. doi: 10.1007/s11882-005-0014-z. PMID: 16091214.) |
| Dermatophagoides farinae | Dry climates | North America and Central Asia | |||||
| Euroglyphus maynei | Humid climates | Western Europe, tropical regions of Asia | |||||
| Storage Mite | Present in agricultural environments and warehouses, where they feed on stored food products such as grain, flour, and seeds | Blomia tropicalis | Pyroglyphidae | Domestic environments | Latin America, Southeast Asia, and the Caribbean | Asthma and allergic rhinitis | |
| Lepidoglyphus destructor | Glycyphagidae | Granaries and farms | Northern Europe | Occupational allergies | |||
| Tyrophagus putrescentiae | Acaridae | Warehouses, humid houses | United States, United Kingdom, and Japan | Asthma, allergic rhinitis, and occupational allergies | |||
| Parasitic and Ectoparasitic Mites | Parasites of humans or animals | Sarcoptes scabiei | Sarcoptidae | Low-income areas, poor hygienic conditions | Sub-Saharan Africa, Southeast Asia, and Latin America | Scabies | |
| Demodex folliculorum | Demodecidae | Hair follicles | Ubiquitous | Dermatitis | |||
| Varroa destructor | Varroidae | Parasite of bees | Allergic reactions in beekeepers | ||||
| Ornithonyssus spp | Dermanyssidae | Parasites of birds | |||||
| Agricultural and Environmental Mites | Found in agricultural contexts, either as crop pests or as biological control agents | Tetranychus urticae | Tetranychidae | Temperate and warm climates | Mediterranean Basin and southeastern United States | Sensitization in agricultural workers | |
| Panonychus citri | Subtropical areas | Asia and Florida |
Figure 1, created by merging images from the Global Biodiversity Information Facility (GBIF.org) portal, illustrates the regions of the world where different mite species have been effectively isolated, collected, and recorded in the GBIF.org database.
3. Discussion
3.1. Molecular Mechanisms of Cross-Reactivity
Cross-reactivity among mites is primarily attributed to the structural and functional homology of their allergenic proteins. Notable examples include tropomyosin, paramyosin, and cysteine proteases such as Der p 1 and Der f 1 [15,16].
Tropomyosin and paramyosin, despite having different functions, exhibit high conservation among mite species, with sequence identity exceeding 95% [17]. In particular, tropomyosin (Der p 10) shares 98% sequence identity with the homologous protein in S. scabiei, making it a key mediator of IgE cross-reactivity [8,17].
A key study found that 35% of scabies patients tested positive for Dermatophagoides pteronyssinus using the radioallergosorbent test (RAST), compared to only 2% in the control group. This finding, along with the high total IgE concentration in scabies patients (45% compared to 4% in controls), supports the hypothesis that scabies infection stimulates the production of IgE antibodies even in non-atopic individuals, suggesting cross-reactivity between the antigenic proteins of Sarcoptes scabiei and house dust mites [18].
Studies using SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis), immunoblotting, and inhibition assays have identified up to 15 cross-reactive proteins in S. scabiei, including muscle and cytoskeletal proteins such as paramyosin. Four of these proteins exhibit strong IgE reactivity, making them key antigenic determinants of cross-reactivity [8]
The serum of patients with positive skin tests for dust mites has been shown to bind multiple proteins from S. scabiei extracts, indicating that individuals sensitized to house dust mites may also react to scabies mite antigens. Furthermore, scabies patients, even after treatment and recovery, maintain elevated levels of specific IgE against dust mites [8].
Proteomic analyses further confirm significant similarities between the enzymatic profiles of dust mites and scabies mites, particularly in proteases and structural proteins [19]. Antigenic similarities have also been confirmed at the genomic and proteomic levels, with the identification of orthologs (genes present in different species derived from a common ancestor) among the major allergenic proteins of Dermatophagoides spp. and Sarcoptes scabiei [20].
A study highlights the presence of orthologs between the allergens of dust mites (Euroglyphus maynei and Dermatophagoides farinae) and the scabies mite (Sarcoptes scabiei), with over 5,000 shared proteins across these species. However, S. scabiei shows a greater expansion of gene families, particularly for inactive proteases (SMIPP-C), which are used to evade the host’s immune system—a strategy less pronounced in dust mites. In contrast, the shared allergens include proteases (cysteine and serine) and lipid-binding proteins with MD-2 domains, but they differ in function and evolutionary adaptations: S. scabiei is specialized for host interaction, whereas dust mites adapt to the domestic environment [20].
Scabies has two significant implications in clinical allergology: first, pruritus as a reason for medical consultation, and second, its influence on allergenicity and the diagnosis of house dust mite (HDM) allergy. Specifically, the allergenicity of Sarcoptes scabiei can induce immunological responses that complicate the diagnosis of house dust mite allergy.
In patients with scabies, IgE antibodies against Dermatophagoides pteronyssinus allergens have been detected. It has also been shown that these patients develop serum IgE against recombinant cysteine and serine proteases of S. scabiei, as well as against apolipoproteins. In contrast, naïve individuals (never exposed) exhibit significantly lower IgE reactivity. Positive immediate-type skin tests to house dust mite allergens have been observed in patients with active or past scabies infestations, likely due to cross-reactive antibodies directed against Der p 4 and Der p 20 [8,21,22].
These aspects underscore the need for careful clinical evaluation to distinguish between sensitization to house dust mite allergens and cross-reactivity induced by scabies, preventing misdiagnoses and inappropriate treatments.
3.2. Main Proteins Involved in Cross-Reactivity
3.2.1. Tropomyosin
Tropomyosin (Der p 10) is a highly conserved allergen present in arthropods, including dust mites, scabies mites, and even crustaceans. It plays a critical role in allergic responses, with significant cross-reactivity documented across different sources [16]. Tropomyosin extracted from S. scabiei demonstrates a high IgE-binding affinity in sera from patients sensitized to dust mites, underscoring its clinical relevance [17].
3.2.2. Paramyosin
Another crucial allergen, paramyosin (Der p 11 and Der f 11), is structurally conserved among invertebrates. Paramyosin is a protein found in the thick muscle filaments of invertebrates. Sequencing studies and biochemical analyses have shown that scabies mite paramyosin shares 97% sequence identity with Dermatophagoides pteronyssinus paramyosin and 95% with Dermatophagoides farina [17].
Its high sequence identity (>95%) between dust mites and scabies mites supports its role in cross-reactivity. Clinical studies show IgE binding to epitopes derived from S. scabiei paramyosin, explaining positive skin test results in individuals sensitized to dust mites without prior exposure to scabies [17]. This phenomenon may explain why patients with scabies infestations exhibit positive reactivity in allergy tests for dust mites, even in the absence of an allergic history.
3.2.3. Cysteine Proteases
Proteins such as Der p 1 and Der f 1 are potent cysteine proteases that contribute to allergenicity through extracellular matrix degradation and immune modulation. Ectoparasitic mites like Sarcoptes scabiei also possess significant enzymatic activity, but their proteolytic properties are less studied compared to dust mites [23].
Sarcoptes scabiei extracts exhibit a more limited enzyme repertoire than dust mites, as demonstrated by their reduced ability to digest protein substrates such as gelatin and casein. This suggests a lower proteolytic capacity in S. scabiei, which could explain its lower allergenic potential compared to dust mites, which possess a wide variety of proteolytic and glycolytic enzymes [23].
Nevertheless, homologous proteins in S. scabiei exhibit structural and functional similarities, supporting the observed immunological cross-reactivity [24]. Comparative protein sequence studies have highlighted significant homology between Dermatophagoides proteases and certain Sarcoptes scabiei proteins, suggesting cross-reactivity mediated by the similar three-dimensional structure of these molecules [20].
In particular, antigenic cross-reactivity has been demonstrated between Der p 10 and Sar s 14, two protein isoforms that trigger strong IgE responses in patients sensitized to either mite. Immunoblotting and ELISA analyses have confirmed that sera from patients allergic to dust mites react significantly to Sarcoptes scabiei antigens [9]. Additionally, Sar s 1, a scabies mite protein, has been shown to exhibit high homology with Der p 10 [9].
3.2.4. Glutathione-S-Transferase (GST)
GST is a ubiquitous enzyme involved in cellular detoxification processes. It plays a role in defense against oxidative stress and protein degradation and is expressed in both dust mites and scabies mites. It has been identified in various dust mites, including D. pteronyssinus and D. farinae, as well as in Sarcoptes scabiei. Strong cross-reactivity between GST from different species has been demonstrated through IgE-binding inhibition experiments, showing that Sarcoptes scabiei GST can induce an immune response similar to that observed for dust mites [16].
3.2.5. Lipid-Binding Protein Family (MD-2 Like)
Lipid-binding proteins belonging to the “MD-2 like” family are a group of molecules involved in various biological functions, ranging from innate immunity to the regulation of cellular interactions. These proteins share distinct structural characteristics, including a domain composed of β-sheets arranged in a way that forms a hydrophobic cavity, designed to accommodate different lipid molecules. This structural versatility makes them functionally adaptable but also susceptible to conformational variations.
The ability to bind lipids grants MD-2 like proteins a crucial role in immune responses. For example, the binding of lipopolysaccharide (LPS) to the MD-2 protein, which is part of the Toll-like receptor 4 (TLR4) complex, is essential for triggering immune responses against bacterial infections.
In the context of allergology, MD-2 like proteins can act as allergens. Their structure facilitates cross-reactivity, allowing them to stimulate immune responses in sensitized individuals. Once recognized as antigens, these proteins can bind to IgE antibodies, triggering allergic reactions. Their susceptibility to conformational changes plays a significant role in this process. Minor variations in the three-dimensional structure of the hydrophobic cavity or exposed regions of the protein can alter how they are recognized by antibodies. This mechanism may explain why some individuals develop cross-reactive responses between dust mites and other allergens, such as pollens or foods that contain proteins with similar domains [24].
3.2.6. SMIPPs (Scabies Mite Inactivated Protease Paralogs)
A key element in understanding cross-reactivity is the discovery of “Scabies Mite Inactivated Protease Paralogs” (SMIPPs). These proteins, found in scabies mites such as Sarcoptes scabiei, have a structure similar to that of proteases but lack catalytic activity.
Serine and cysteine proteases are active enzymes that play essential roles in cellular metabolism and immune responses, participating in protein degradation and the activation of inflammatory pathways. In contrast, SMIPPs, despite their structural resemblance to proteases, are inactive. Their inactivity results from mutations in critical residues of the catalytic site, which would normally enable them to degrade protein substrates.
Despite their lack of enzymatic activity, SMIPPs are believed to have an important biological function by acting as “antagonists” of active proteases. These inactive proteins may compete with active proteases for peptide substrates, thereby interfering with processes that lead to immune response activation [19].
A significant example is Sar s 3, a SMIPP identified in scabies mites. This protein exhibits remarkable structural similarity to Der p 3, an active serine protease from dust mites. This similarity suggests that individuals sensitized to dust mite allergens may also react to scabies mite allergens. It may explain why people sensitized to dust mites tend to experience more severe or prolonged reactions in the presence of scabies infestations [19].
The identification and study of SMIPPs, such as Sar s 3, not only enhance our understanding of the immunological mechanisms underlying these reactions but may also pave the way for novel therapeutic strategies aimed at modulating inflammatory responses.
3.3. Clinical Implications
This phenomenon has significant clinical implications. In patients with scabies infection, the presence of specific IgE against Dermatophagoides can lead to an overestimation of sensitization to dust mites, resulting in misdiagnosis or overdiagnosis of dust mite allergy [22]. Additionally, in some cases, allergic symptoms such as asthma and rhinitis may worsen during or after a scabies infection due to cross-stimulation of the immune system [8].
It has been observed that some patients with a history of scabies infestation, despite being non-atopic and having no symptoms of respiratory or cutaneous allergy, may show positive skin tests to house dust mite allergens [21]. This reactivity has been attributed to immunological cross-reactivity, mediated by IgE antibodies specific to scabies allergens that also recognize house dust mite allergens due to structural similarity. This phenomenon highlights the need for careful interpretation of allergy tests in individuals with prior exposure to mites, in order to avoid misdiagnosis of allergic conditions.
Panzer R. et al. [25] confirmed that patients with active scabies infestations show elevated levels of specific IgE against Dermatophagoides and other dust mite species, suggesting that scabies infestation may sensitize the immune system in a way that promotes cross-reactive responses.
In patients with active scabies infection, high levels of specific IgE against dust mites have been observed, suggesting that the infection may induce a temporary or permanent allergic state [22]. The reverse condition has also been documented: individuals sensitized to dust mites can develop IgE antibodies that also recognize S. scabiei proteins [15].
This phenomenon is particularly evident in dogs with atopic dermatitis, where sera from affected animals have shown positive reactions against both dust mite and scabies mite antigens [15,26]. Another study on canine sera revealed that the recognition of common proteins between these two mite categories is not limited to IgE but also involves IgG [27]. These studies suggest that mite cross-reactivity is relevant not only in humans but also in other animal species.
Additionally, scabies infestation may induce activation of cell-mediated immunity against dust mites. A study on scabies patients reported that 69% of subjects tested positive in the atopy patch test (APT) when exposed to dust mite allergens applied to the skin.²⁸ Changes in immunological parameters in scabies patients (positive SPT and/or specific IgE for D. pteronyssinus/D. farinae, as well as APT positivity to dust mites) suggest that scabies infestation could cause a transient state of “atopy” or an atopic dermatitis-like (AD-like) condition. In several studies, some patients who had previously suffered from scabies remained positive in the SPT or intradermal test (IDT) against dust mites for a period ranging from six weeks to several months [28].
3.4. Diagnostic Challenges
Cross-reactivity between these allergens can lead to false-positive or false-negative results in allergy tests, complicating diagnosis.
In patients with active scabies or a history of infestation, skin tests for dust mite allergens may yield positive results not due to true allergic sensitization but rather because of cross-reactivity to shared antigens. This situation can lead to misdiagnosis of dust mite allergy and inappropriate treatment, especially in patients with prior exposure to scabies infestations [9].
Scabies patients often exhibit elevated IgE levels reactive to dust mite allergens, leading to potential misdiagnoses of dust mite allergy [29]. Similarly, individuals sensitized to dust mites may react to scabies antigens, further complicating test interpretation [1].
The differential diagnosis between dust mite allergy and scabies mite sensitization is primarily based on skin prick tests (SPT) and measurements of specific IgE. However, due to the strong cross-reactivity between the allergenic proteins of the two mites, test results may show positive reactivity to both, making clear distinction difficult. Moreover, advanced techniques such as allergen protein microarrays have demonstrated that many of the IgE antibodies in patient sera cross-react with structurally similar proteins from both Dermatophagoides and Sarcoptes [1].
The use of specific immunoassays for allergens, such as ImmunoCAP and recombinant allergen microarrays, may help distinguish between true sensitization and cross-reactivity. New markers, such as Sar s 14 and Sar s 1, show potential in differential diagnosis [10].
Using purified allergens in in vitro tests could reduce false positives and improve diagnostic accuracy, preventing adverse reactions in already sensitized patients [20].
3.5. Therapeutic Considerations
Therapeutic management must consider potential cross-reactivity: treatments such as allergen-specific immunotherapy (AIT) for dust mites may influence reactions to scabies allergens. Some studies have shown that immunization with Dermatophagoides farinae extracts provided protection against scabies infestation, suggesting a possible protective role of cross-reactivity [8].
AIT for dust mites may inadvertently modulate immune responses to scabies antigens, requiring careful patient selection [8]. Additionally, corticosteroids and antihistamines provide symptomatic relief in cases of cross-sensitization, but targeted immunotherapy would offer greater potential for long-term control [4].
3.6. Bioinformatic Analysis
In light of the findings from the literature review, we conducted a bioinformatic analysis to verify the existence of a robust scientific basis supporting this cross-reactivity.
From the perspective of amino acid sequence homology between two proteins, the following considerations must be taken into account [30]:
- An identity greater than 70% is considered a strong indicator of clinical cross-reactivity.
- Values between 50% and 70% can still lead to cross-reactivity, but they depend more on the three-dimensional structure of the allergen and the similarity of IgE-binding sites. Indeed, it is not always possible to establish a simple and direct relationship between sequence homology and cross-reactivity, as structural and conformational factors of allergenic proteins also influence their ability to bind IgE.
- For identity values below 50%, cross-reactivity is less likely but not excluded, especially for allergens with conserved IgE-binding sites [30].
Cantillo et al. [31] demonstrated that proteins with high sequence homology (85% and 71%) exhibit cross-reactivity with known allergens, supporting the idea that high sequence identity is a good indicator of cross-reactivity. Conversely, proteins with lower sequence identity (51% and 60%) did not show cross-reactivity, suggesting that a homology threshold of 70–80% is commonly used to indicate potential cross-reactivity between allergens. Homology above this value suggests a likely cross-reactivity.
The data we present were obtained through the analysis of protein sequences in FASTA format, retrieved from UniProt and compared using the SDAP 2.0 (Structural Database of Allergenic Proteins). Sequence alignments were analyzed to identify identity percentages, similarity, and significant homology scores. The analysis focused on the most relevant allergens identified in the literature review presented in this article, particularly Sar s 14, Sar s 1, and Sar s 3 from Sarcoptes scabiei.
4. Results
For Better Understanding, We Summarize Below the Meaning of the Individual Entries in the Analysis [32]
- Sequence info: The unique identifier code for the protein in the UniProt database.
- Name: Common name of the protein.
- Length: The length of the protein in terms of the number of amino acids.
- Opt: Optimized alignment score, reflecting the overall similarity between the sequences.
- Bits Score: The “bit” score, which helps assess the quality of the alignment; a higher score indicates a better alignment.
- E-Value: The E-value represents the expected number of alignments that would occur by chance with a score equal to or higher than the observed one, given the database and query size. A lower E-value indicates a statistically significant alignment.
- Identities (%): The percentage of identity between the sequences; it shows how many amino acid positions are exactly the same in the aligned sequences.
- Positives (%): The percentage of similarity, including conservatively substituted amino acids with similar chemical properties.
- Gaps (%): The percentage of gaps in the alignment, indicating where the sequences have been extended or truncated to optimize alignment.
Table 2.
Sequence Homology of Sar s 14.
| Sequence Info | Allergen Name | Lenght | Opt | Bits Score | E Value |
|---|---|---|---|---|---|
| AAF14270 | Eur m 14 | 1668 | 6992 | 1599.7 | 0 |
| AAM21322 | Der p 14 | 1662 | 6936 | 1586.9 | 0 |
| BAA04558 | Der f 14 | 349 | 1558 | 361.4 | 15e-100 |
| CAA31942 | Gal d vitellogenin | 1850 | 163 | 43.1 | 0.00055 |
4.1. Results of Sar s 14 Protein Analysis
The analysis of the Sar s 14 protein revealed a high degree of homology with allergenic proteins from dust mites, indicating a potential ability of these proteins to induce immunological cross-reactivity, particularly:
-
Eur m 14 (Euroglyphus maynei):
- ○
- Identity: 59% (995/1662 residues), Overall similarity: 81%.
- ○
- Score: 1599.7 bits, E-value: 0.
- ○
- These data indicate a highly conserved sequence, with an extremely high level of homology and almost no significant differences. The strong positivity (81%) highlights the presence of conservative substitutions (amino acids with similar biochemical properties even if not identical).
-
Der p 14 (Dermatophagoides pteronyssinus):
- ○
- Identity: 59% (980/1657 residues), Overall similarity: 81%.
- ○
- Score: 1586.9 bits, E-value: 0.
- ○
- This result confirms a strong similarity with Sar s 14, suggesting a similar evolutionary role and a probable sharing of immunogenic regions relevant to the immune system.
-
Der f 14 (Dermatophagoides farinae):
- ○
- Identity: 65% (227/349 residues), Overall similarity: 83%.
- ○
- Score: 361.4 bits, E-value: 2e-100.
- ○
- In this case, the identity level is even higher (65%), with a very high positivity percentage. However, the lower bits score and E-value indicate a shorter sequence. This is because only a protein fragment was available in bioinformatics databases rather than the full amino acid sequence.
4.2. Comparison with Vitellogenins (Chicken Egg, Gallus domesticus)
A significant difference was observed when comparing Sar s 14 with vitellogenins:
-
Gal d vitellogenin:
- ○
- Score: 43.1 bits / 40.6 bits.
- ○
- Identity: 22%.
- ○
- Positivity: 46%.
- ○
- Gaps: ~15% (134/868 residues).
- ○
- The similarity is low, with only 22% sequence identity and multiple gaps. This suggests that while vitellogenins have a similar structural function, they share low homology with Sar s 14. The evolutionary distance between phyla explains these differences.
In contrast, Sar s 1 (cysteine protease) and Sar s 3 proteins also exhibit homology but with lower bit scores and identity percentages compared to Sar s 14. The specific values and percentages are provided in the following tables:
Table 3.
Sequence Homology of Sar s 1.
| Sequence Info | Allergen Name | Lenght | Opt | Bits Score | E Value |
|---|---|---|---|---|---|
| BAC53948 | Der f 1 | 321 | 793 | 170.6 | 8.1e-44 |
| P25780 | Eur m 1 | 321 | 785 | 168.9 | 2.5e-43 |
| CAQ68250 | Der p 1 | 302 | 763 | 164.5 | 5.1e-42 |
| AAQ24541 | Blo t 1 | 333 | 481 | 107.1 | 1e-24 |
Table 4.
Sequence Homology of Sar s 3.
| Sequence Info | Allergen Name | Lenght | Opt | Bits Score | E Value |
|---|---|---|---|---|---|
| O97370 | Eur m 3 | 261 | 657 | 156.4 | 9.1e-40 |
| P39675 | Der p 3 | 261 | 652 | 155.3 | 2e-39 |
| P49275 | Der f 3 | 259 | 642 | 153.0 | 9.7e-39 |
| AAM10779 | Blo t 3 | 266 | 583 | 139.5 | 1.1e-34 |
| AAP57077 | Der p 9 | 273 | 487 | 117.6 | 4.5e-28 |
The findings indicate that the allergenic proteins of Sarcoptes scabiei share significant similarity with those of other dust mites, particularly within the “14” protein family. This provides a plausible molecular explanation for allergological cross-reactivity. Future studies should further investigate the role of shared epitopes in immune responses and assess the clinical impact of these interactions. The inclusion of allergens derived from Sarcoptes scabiei in diagnostic panels could represent a step forward in the personalized management of allergies.
Additionally, we generated a phylogenetic tree of the analyzed proteins using the Phylogeny.fr server by uploading the corresponding FASTA sequences.
As shown in Figure 2, Sar s 14 appears on a separate branch from the other allergens, as indicated by the observed phylogenetic distance (0.31). This suggests that, while it retains evolutionary similarities with the other sequences in the tree, its divergence is significant.
In the tree, the isolated position of Sar s 14 compared to other allergens indicates that, while it belongs to the same general evolutionary lineage, it has undergone modifications that make it less closely related to other nearby proteins.
The value of 0.31 above the branch of Sar s 14 represents the evolutionary distance from its divergence point with the common node. A higher value suggests a greater reliability of the evolutionary relationship³². This value indicates that Sar s 14 exhibits significant differences, despite sharing a common origin with other proteins in the tree.
The line below the phylogenetic tree, labeled 0.1, is a scale bar that indicates phylogenetic distance. This represents the number of observed changes (or differences) in the sequences, which translates, in evolutionary terms, into the degree of divergence between species or proteins.
A 0.1 scale bar means that a branch length equivalent to that bar represents a 10% difference between the aligned sequences [32].
In summary, the image demonstrates that Sar s 14 is evolutionarily related to the other allergens but is sufficiently divergent to form a distinct phylogenetic branch, separate from the other analyzed allergens.
We then conducted a study to predict the potential B-cell epitopes from the protein structures of the studied allergens, focusing on regions capable of inducing an IgE-mediated immune response. This analysis was performed using BepiPRED 2.0, a Machine Learning algorithm specifically adapted for predicting antibody-recognizable epitopes [32].
Epitopes—small peptides composed of eight to fifteen amino acids—are the allergenic determinants responsible for the specific binding with IgE. They play a crucial role in cross-reactivity among different allergens, which occurs when structurally similar but distinct proteins share epitopes capable of binding IgE [14].
Below are the results for Sar s 14, Eur m 14, Der p 14, and Der f 14, shown in Figures 3–6.
Figures 3–6.
Epitope Prediction Analysis.
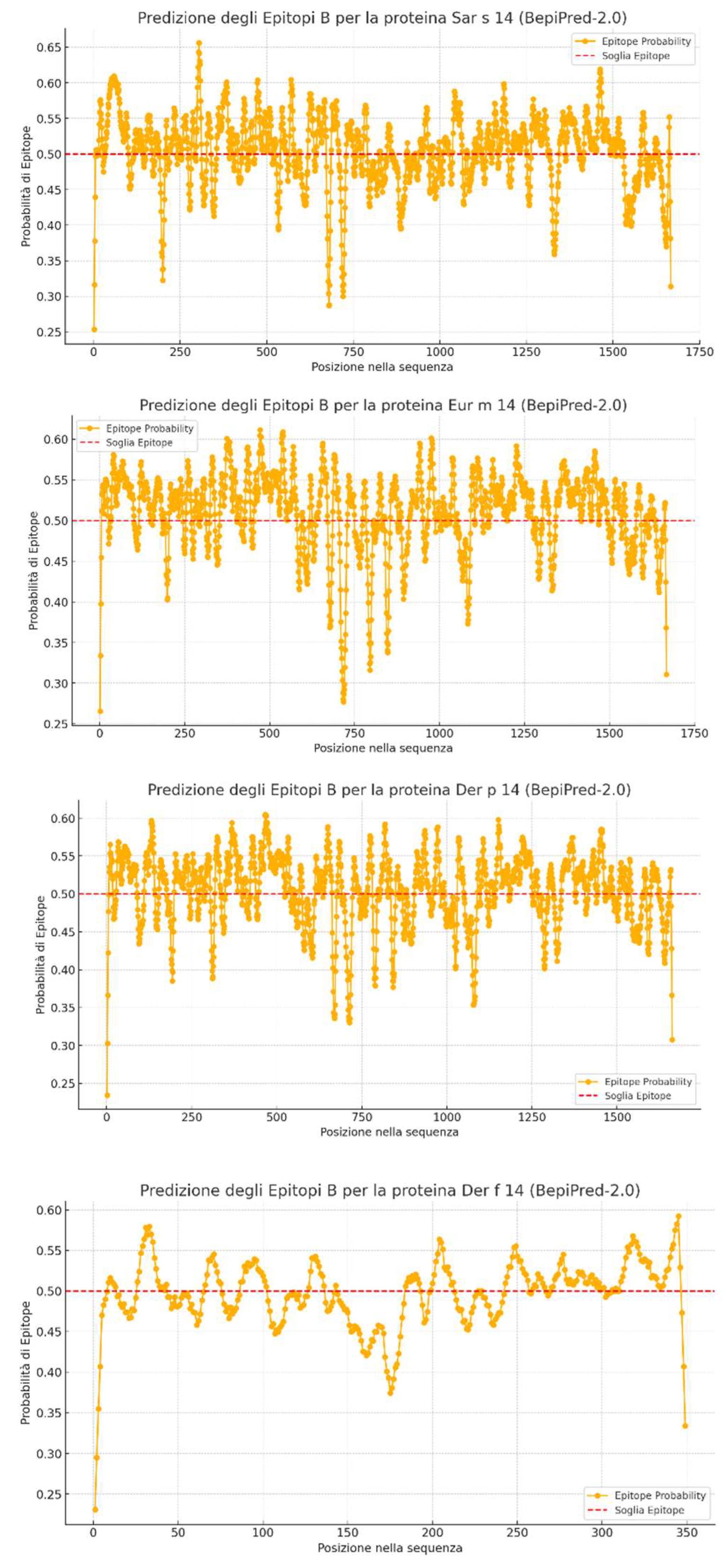
- The X-axis represents the position of amino acids within the protein sequence.
- The Y-axis shows the epitope prediction score.
- The dashed red line is the cutoff threshold set at 0.5: values above this threshold are considered potential epitopes.
BepiPred generated a CSV (Comma separated values) file containing all the computed data, from which we conducted a detailed comparison of the three analyzed proteins (Sar s 14, Der p 14, Eur m 14). Der f 14 was excluded from this analysis as its full sequence was unavailable in bioinformatics databases, and only a fragment was analyzed (as observed from the low number of residues on the X-axis in previous graphs).
1. Comparison of Epitope Probability
- The highest average epitope probability was observed for Eur m 14 (0.5107), followed by Der p 14 (0.5053) and Sar s 14 (0.5047).
- This suggests that Eur m 14 has a higher tendency to contain antigenic epitopes than the other two proteins.
-
When considering residues with a probability above the 0.5 threshold, the distribution was:
- ○
- Eur m 14: 1132 antigenic residues (67.9%)
- ○
- Der p 14: 1007 antigenic residues (60.6%)
- ○
- Sar s 14: 979 antigenic residues (58.7%)
- ○
- This indicates that Eur m 14 has a wider distribution of epitopes and a greater likelihood of immune recognition.
2. Residue Exposure
-
The protein with the highest number of exposed residues was Sar s 14 (240 residues, 14.4%), followed by:
- ○
- Eur m 14 (237 residues, 14.2%)
- ○
- Der p 14 (231 residues, 13.9%)
- Although the differences are minimal, Sar s 14 shows higher surface exposure, potentially making it more accessible for antibody interaction.
3. Surface Accessibility
-
Relative Surface Accessibility analysis revealed:
- ○
- Eur m 14: 0.1831
- ○
- Der p 14: 0.1783
- ○
- Sar s 14: 0.1757
- This means that Eur m 14 has a structural profile that makes it more accessible, potentially explaining its higher epitope density.
4. Secondary Structure Analysis
-
Helix Probability (α-helices formation tendency):
- ○
- Sar s 14: 0.2060
- ○
- Der p 14: 0.2046
- ○
- Eur m 14: 0.2013
-
Sheet Probability (β-sheet formation tendency):
- ○
- Eur m 14: 0.0934
- ○
- Sar s 14: 0.0920
- ○
- Der p 14: 0.0917
-
Coil Probability (flexible/disordered structure tendency):
- ○
- Eur m 14: 0.7052
- ○
- Der p 14: 0.7036
- ○
- Sar s 14: 0.7019
- ○
- Coil regions are often implicated in antigenic epitopes, which may further explain the higher antigenicity of Eur m 14.
5. Which Protein is the Most Antigenic?
- Based on:
- The highest average epitope probability
- The largest number of residues with epitope probability > 0.5
- The highest surface accessibility
- The strongest tendency to form flexible regions (coil structure)
Eur m 14 appears to be the most antigenic protein among the three. However, Sar s 14 has the highest surface exposure, potentially making it a more accessible target for antibodies.
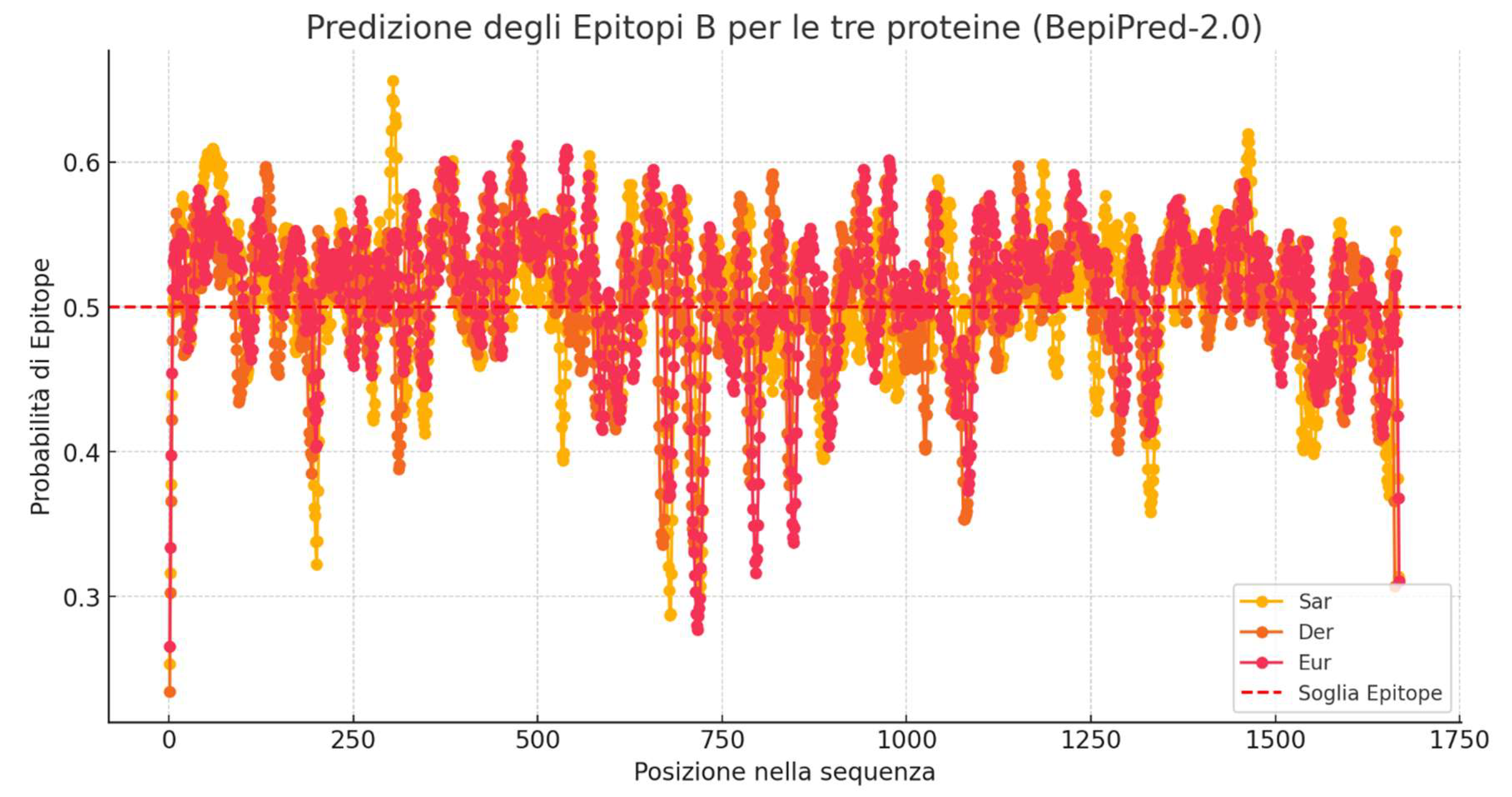
In the following graph (Figure 7), epitope predictions for the analyzed proteins have been overlaid to provide a comparative visualization of their antigenic potential.
- Positions with at least two proteins above the 0.5 threshold: Multiple regions along the sequence show high epitope probability in at least two of the three analyzed proteins. This suggests that certain sequence regions are likely antigenic in more than one protein, indicating conserved epitopes or structurally favorable antigenic sites.
- Positions where all three proteins exceed the 0.5 threshold: Several regions exhibit a probability above the critical threshold for all proteins. These areas are particularly significant as they represent highly conserved epitopes, which could serve as ideal targets for immunogenicity strategies or vaccine development.
5. Conclusions
The conducted bioinformatic analysis revealed significant sequence homology between the allergenic proteins of Sarcoptes scabiei and those of various dust mites, particularly within the Sar s 14 family. This suggests a plausible molecular basis for allergological cross-reactivity.
The obtained results indicate that the percentage identity between these allergens falls within the predictive range for potential cross-reactivity, with values often exceeding the critical 50-70% threshold. Additionally, the overlapping antigenic epitopes identified through BepiPred 2.0 further support the hypothesis that an immune response triggered by dust mites could potentially extend to Sarcoptes scabiei allergens.
Including these allergens in diagnostic panels could enhance the identification of individuals with cross-sensitization and contribute to a more personalized approach to allergy management. However, while our analysis suggests a strong probability of cross-reactivity, future studies are needed to confirm these findings through in vitro IgE-binding assays and in vivo experimental models.
Understanding the relationship between these allergens could open new perspectives in the diagnosis and treatment of allergic reactions, emphasizing the need for further structural and immunological research.
Cross-reactivity between dust mites and scabies mites represents a complex interaction of immunological and molecular mechanisms with significant clinical implications. Despite progress in allergen characterization, further research is essential to clarify the full spectrum of cross-reactive antigens and refine diagnostic and therapeutic approaches.
Future research should focus on high-resolution mapping of allergenic epitopes, development of highly specific diagnostic markers, and evaluation of the effectiveness of AIT in patients with documented cross-reactivity.
A promising strategy for improving diagnosis is the use of S. scabiei-specific antigens that do not share homology with dust mite allergens. However, significant challenges remain, such as the precise identification of exclusive antigens and the optimization of serological tests.
A deeper understanding of this phenomenon will not only improve diagnostic accuracy but also enhance therapeutic outcomes for individuals affected by allergies and mite-related infestations.
Fine modulo
References
- Arlian LG. Dust mites: update on their allergens and control. Curr Allergy Asthma Rep. 2001 Nov;1(6):581-6. [CrossRef]
- Sidenius, K.E., Hallas, T.E., Poulsen, L.K. and Mosbech, H. (2001), Allergen cross-reactivity between house-dust mites andother invertebrates. Allergy, 56: 723-733. [CrossRef]
- Huey-Jy Huang, Eszter Sarzsinszky, Susanne Vrtala, House dust mite allergy: The importance of house dust mite allergens for diagnosis and immunotherapy, Molecular Immunology, Volume 158, 2023, Pages 54-67, ISSN 0161-5890. [CrossRef]
- Hengge UR, Currie BJ, Jäger G, Lupi O, Schwartz RA. Scabies: a ubiquitous neglected skin disease. Lancet Infect Dis. 2006 Dec;6(12):769-79. [CrossRef]
- Govindarajan R, Rajamannar V, Kumar A, Samuel PP. Current status of mites and mite-borne diseases in India. J Vector Borne Dis. 2023 Jan-Mar;60(1):1-10. [CrossRef]
- Sunderkötter C, Wohlrab J, Hamm H. Scabies: Epidemiology, Diagnosis, and Treatment. Dtsch Arztebl Int. 2021 Oct 15;118(41):695-704. [CrossRef]
- Arora P, Rudnicka L, Sar-Pomian M, Wollina U, Jafferany M, Lotti T, Sadoughifar R, Sitkowska Z, Goldust M. Scabies: A comprehensive review and current perspectives. Dermatol Ther. 2020 Jul;33(4):e13746. [CrossRef]
- Arlian, L. G., Vyszenski-Moher, D. L., & Gilmore, A. M. (1988). Cross-antigenicity between sarcoptes scabiei and the house dust mite, dermatophagoides farinae (acari: sarcoptidae and pyroglyphidae). Journal of Medical Entomology, 25(4), 240-247. [CrossRef]
- Larry G Arlian, Diann L. Vyszenski-Moher, Salva G. Ahmed, Stephen A. Estes, Cross-Antigenicity Between the Scabies Mite, Sarcoptes scabiei, and the House Dust Mite, Dermatophagoides pteronyssinus, Journal of Investigative Dermatology, Volume 96, Issue 3, 1991, Pages 349-361, ISSN 0022-202X, . [CrossRef]
- Arlian LG, Feldmeier H, Morgan MS (2015) The Potential for a Blood Test for Scabies. PLoS Negl Trop Dis 9(10): e0004188. [CrossRef]
- Falk ES, Dale S, Bolle R, Haneberg B. Antigens Common to Scabies and House dust Mites. Allergy. 1981 May;36(4):233-8. [CrossRef]
- Sánchez-Borges M, et al. Scabies, crusted (Norwegian) scabies and the diagnosis of mite sensitisation. Allergol Immunopathol (Madr). 2017. [CrossRef]
- Smith AM, Benjamin D, Hozic N, et al.: The molecular basis of antigenic cross-reactivity between the group 2 mite allergens. J Allergy Clin Immunol 2001, 107:977–984.
- Fernández-Caldas E, Iraola Calvo V. Mite allergens. Curr Allergy Asthma Rep. 2005 Sep;5(5):402-10. [CrossRef]
- Schumann RJ, Morgan MS, Glass R, Arlian LG. Characterization of house dust mite and scabies mite allergens by use of canine serum antibodies. Am J Vet Res. 2001 Sep;62(9):1344-8. [CrossRef]
- Aalberse, R.C. (1998), Allergens from mites: implications of cross-reactivity between invertebrate antigens. Allergy, 53: 47-48. [CrossRef]
- Nisbet AJ, MacKellar A, Wright HW, Brennan GP, Chua KY, Cheong N, Thomas JE, Huntley JF. Molecular characterization, expression and localization of tropomyosin and paramyosin immunodominant allergens from sheep scab mites (Psoroptes ovis). Parasitology. 2006 Oct;133(Pt 4):515-23. [CrossRef]
- Falk ES, Bolle R. IgE antibodies to house dust mite in patients with scabies. Br J Dermatol. 1980 Sep;103(3):283-8. [CrossRef]
- K. Fischer, D.C. Holt, B.J. Currie, S.F. Walton, D.J. Kemp, Scabies mite inactivated protease paralogues, International Congress Series, Volume 1289, 2006, Pages 85-88, ISSN 0531-5131, ISBN 9780444522054, . [CrossRef]
- Rider SD, Jr., Morgan MS, Arlian LG (2017) Allergen homologs in the Euroglyphus maynei draft genome. PLoS ONE 12(8): e0183535. [CrossRef]
- Bajoghli AA, Bajoghli M, Adler S. Positive house dust mite skin test in a nonatopic patient with scabies. Ann Allergy Asthma Immunol. 2014 Dec;113(6):667. [CrossRef]
- Walton SF, Slender A, Pizutto S, Mounsey KE, Opresecu F, Thomas WR, Hales BJ, Currie BJ. Analysis of IgE binding patterns to house dust mite allergens in scabies-endemic communities: insights for both diseases. Clin Exp Allergy. 2015 Dec;45(12):1868-72; Erratum in: Clin Exp Allergy. 2016 Mar;46(3):508. doi: 10.1111/cea.12713. [CrossRef]
- Marjorie S. Morgan, Larry G. Arlian, Enzymatic Activity in Extracts of Allergy-Causing Astigmatid Mites, Journal of Medical Entomology, Volume 43, Issue 6, 1 November 2006, Pages 1200–1207, . [CrossRef]
- Wayne R. Thomas, Hierarchy and molecular properties of house dust mite allergens, Allergology International, Volume 64, Issue 4, 2015, Pages 304-311, ISSN 1323-8930, . [CrossRef]
- Panzer R, Krebs S. Mites, caterpillars and moths. J Dtsch Dermatol Ges. 2020 Aug;18(8):867-880. [CrossRef]
- Arlian LG, Morgan MS. Serum antibody to Sarcoptes scabiei and house dust mite prior to and during infestation with S. scabiei. Vet Parasitol. 2000 Jul 4;90(4):315-26. [CrossRef]
- Bigler, B. and Virchow, F. (2004), P-43. IgG antibodies against sarcoptic mite antigens in dogs cross-reacting with house dust and storage mite antigens. Veterinary Dermatology, 15: 54-54. [CrossRef]
- Taşkapan O, Harmanyeri Y. Atopy patch test reactions to house dust mites in patients with scabies. Acta Derm Venereol. 2005;85(2):123-5. [CrossRef]
- Aalberse RC. Clinically Significant Cross-Reactivities among Allergens. Int Arch Allergy Immunol. 1992;99(2-4):261-264. [CrossRef]
- Chapman MD, Pomés A, Breiteneder H, Ferreira F. Nomenclature and structural biology of allergens. J Allergy Clin Immunol. 2007 Feb;119(2):414-20. [CrossRef]
- Cantillo, Puerta, L.; Fernandez-Caldas, E.; et al. Tropomyosins in mosquito and house dust mite cross-react at the humoral and cellular level. Clin Exp Allergy 2018, 48, 1354-1363.
- ChatGPT as a bioinformatic partner. Gianluca Mondillo, Alessandra Perrotta, Simone Colosimo, Vittoria Frattolillo- medRxiv 2024.08.20.24312291; [CrossRef]
Figure 1.
Isolation of Different Mite Species. Legend: Yellow – Sarcoptes scabiei; Green – D. farinae; Red – D. pteronyssinus; Blue – Euroglyphus maynei. Global Biodiversity Information Facility (GBIF.org).
Figure 1.
Isolation of Different Mite Species. Legend: Yellow – Sarcoptes scabiei; Green – D. farinae; Red – D. pteronyssinus; Blue – Euroglyphus maynei. Global Biodiversity Information Facility (GBIF.org).
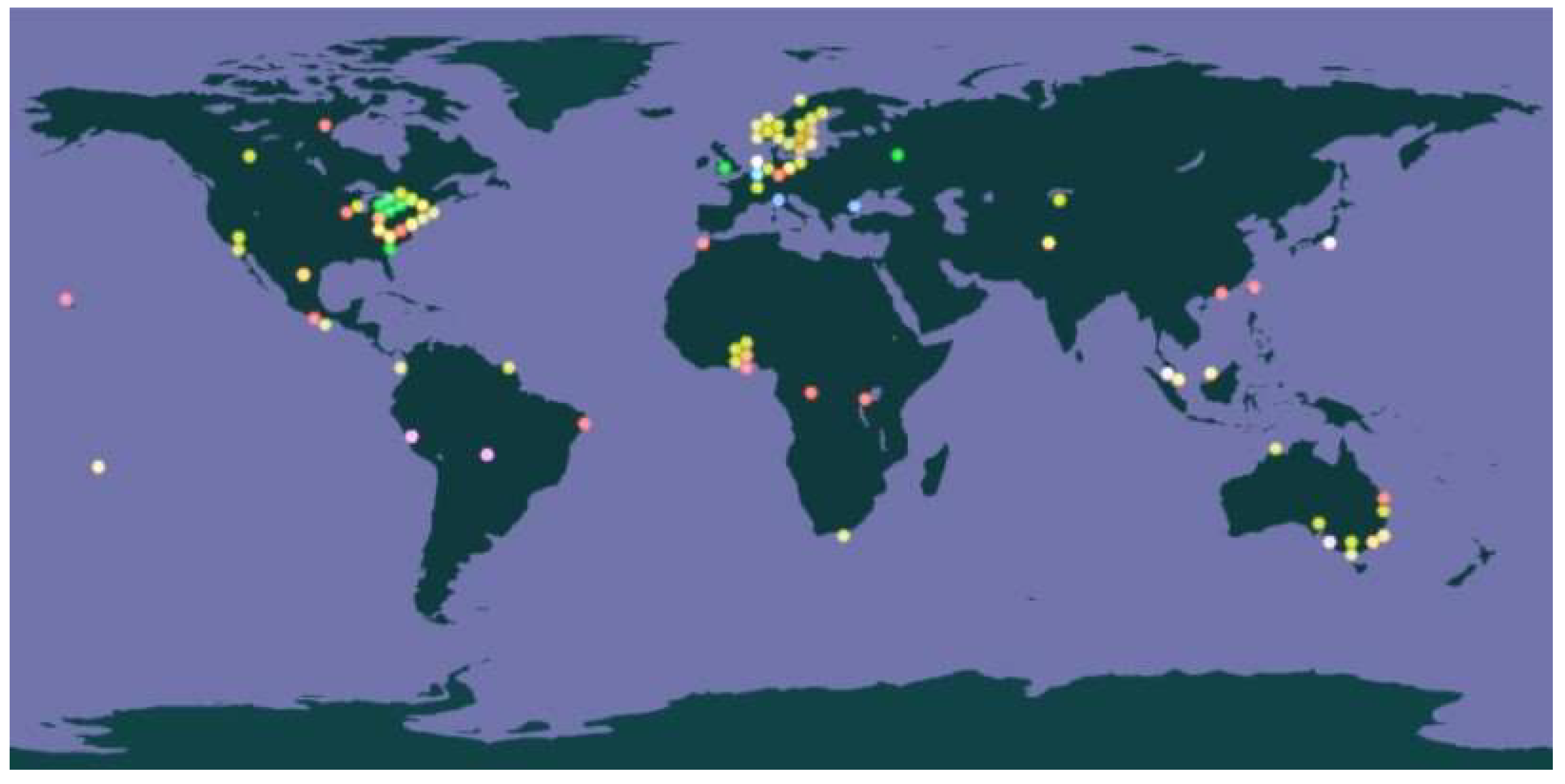
Figure 2.
Phylogenetic Tree.

Figure 7.
Overlapping Epitope Predictions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



