
Submitted:
22 April 2025
Posted:
24 April 2025
You are already at the latest version
Abstract
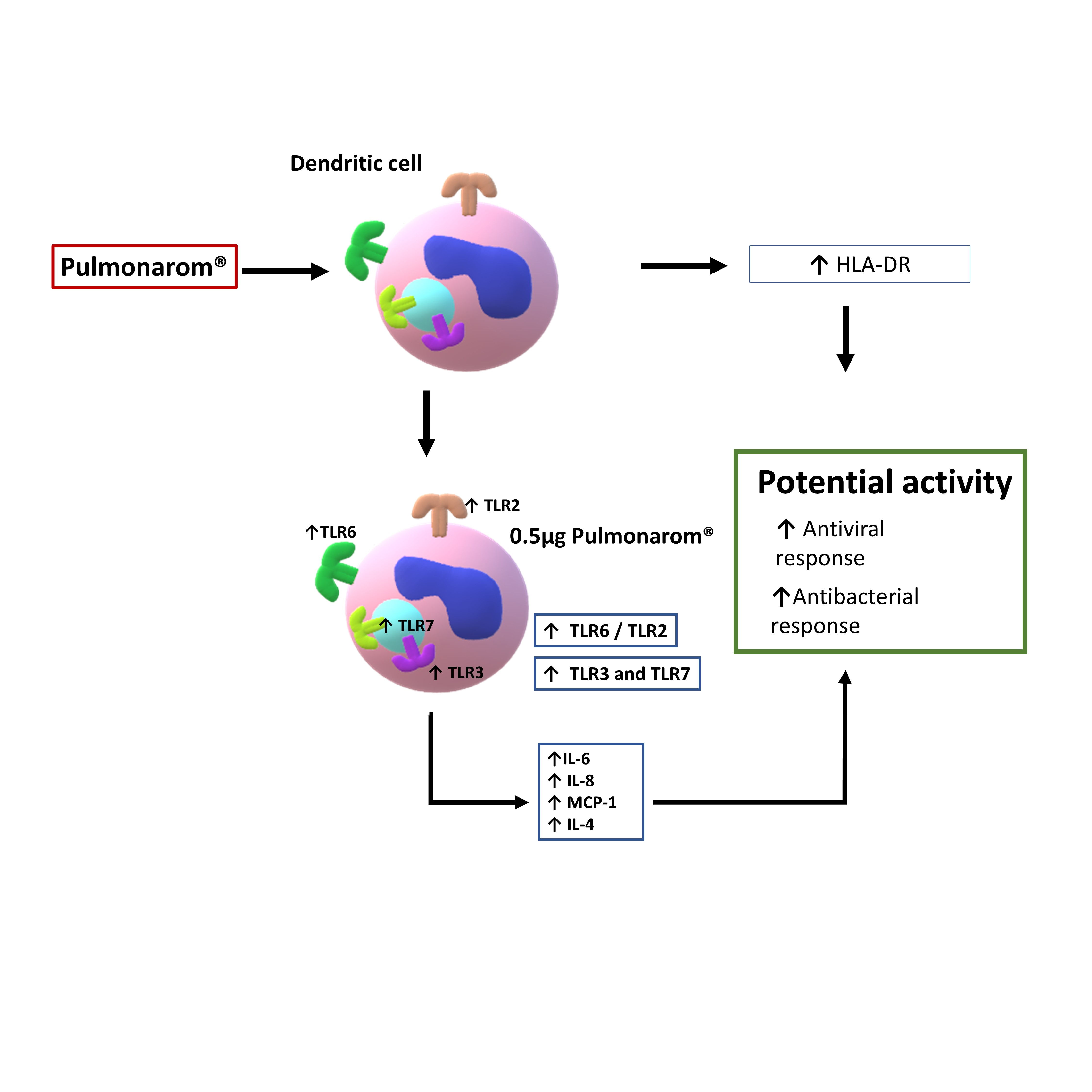
Background Evidence shows bacterial lysates may participate in the immune response against respiratory infections. However, few studies have been conducted on dendritic cells, and, to our knowledge, none on human dendritic cells, and few have analyzed certain TLRs and cytokines. Materials and Methods Monocytes were obtained from the buffy coat and subsequently differentiated into dendritic cells (moDCs). The protein of commercial bacterial lysate Pulmonarom® was lyophilized and quantified. Ultrastructural changes induced by Pulmonarom® were evaluated in moDC by electron microscopy, and the expression of TLRs and some cytokines in these cells was evaluated by flow cytometry. Results Pulmonarom®-stimulated moDCs showed increased numbers of dendrites and the presence of lysosomes; they also showed increased expression of MHC-II and TLRs 2, 3, 6, and 7, IL-4, IL-6, IL-8, and MCP-1. Major conclusions The effects of Pulmonarom® on moDC correlate with an increased capacity to mobilize to lymph nodes and present antigens, initiating an effective adaptive immune response. Lysosomal activity and maturation of moDC, increased expression of TLR2, TLR3, TLR6, and TLR7 receptors, and increased production of the cytokines IL-8, MCP-1, IL-6, and IL-4, indicate a training process in moDC after their activation with Pulmonarom® bacterial lysate. This suggests that Pulmonarom® could enhance responses against viruses and bacteria by activating moDC. To our knowledge, this is the first study to address the effect of Pulmonarom® on one of the most important cells orchestrating the innate and acquired immune responses.
Keywords:
Pulmonarom
; Dendritic cell
; monocyte derived drndritic cell
; moDC
; TLR
; Cytokine
; Immunomodulator
; respiratory infectious disease
1. Introduction
Acute respiratory tract infections represent a serious worldwide public health problem due to their high mortality and morbidity rates, complications, and sequelae, and also to economic repercussions in part due to the days of disability required by patients, representing a complex problem.
The Global Burden of Disease Study reports that in 2021, the global number of new episodes of upper respiratory tract infections was 12.8 billion for all ages in men and women. [1] Globally, the incidence rate of upper respiratory tract infections was from 162 to 484.8 per 100,000 population, with the highest incidence rate in children under 2 years old, while the number of episodes recorded in children aged 5 to 9. [1]
Regarding lower respiratory tract infections (LRI) during 2024, the Global Burden of Disease Study reports that in 2021, there were 344 million cases of LRI and 2.18 million deaths worldwide; 502,000 deaths occurred in children under 5 years of age, of which 254,000 occurred in countries with a low sociodemographic index. [2]
Considering this serious problem worldwide, preventive measures have been adopted to help reduce the impact of these infections; one of them, and probably the most important, is vaccination, which has had a great impact in several diseases caused by viral and bacterial agents. [3]Although, unfortunately, not all respiratory infections can be prevented through immunization.[4]
Another measure adopted to deal with respiratory tract infections is the use of bacterial lysate compounds for the therapy against these infections, which have been of utility for their prevention and treatment. [5]. Bacterial lysates fall under the classification of postbiotics proposed by ISAPP, which states that "A postbiotic is a preparation of inanimate microorganisms and/or their components that confer a health benefit on the host." This definition makes no distinction between the microorganism of origin.[6,7,8]
These bacterial lysates act as immunomodulators, and some reports point to their effects through the increase of specific antibodies. These properties are probably due to their capacity to stimulate the innate immune response. Considering that dendritic cells are one of the main relevant cells that orchestrate the immune response. [9,10] It is important to know which effects these bacterial lysates have on this antigen-presenting cell.
Dendritic cells (DCs) are a heterogeneous cell population whose members differ in ontogeny, anatomical location, migration, cytokine secretion pattern, and immunological functions. They are located in lymphoid and non-lymphoid tissues, and sense the presence of pathogens or danger molecules mainly through their pattern recognition receptors (PRRs): pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) [11]. DCs have several types of PRRs; the most well-known are the Toll-like receptors (TLRs). DCs can also phagocytose microorganisms, a process that favors the migration of DCs to lymphoid organs where they perform their functions as antigen presenters to T lymphocytes, thus initiating the adaptive immune response. [10] It has also been shown that DC participates in the modulation of the immune response towards an inflammatory response (Th1) or antibody-producing and antiparasitic response (Th2) [9] and in the regulation of cytotoxic T lymphocytes and immunological tolerance through the production of different cytokines. [12].
Intranasal immunization with Postbiotic induced in mice increased mRNA expression levels of CCL3, CCL4, CXCL1, CXCL2, CXCL9, and CXCL10 and moderate levels of TNF-α, as well as increased influx of activated DC (CD80Hi/CD86Hi) and neutrophils.[13] In a subsequent study, mice orally administered with Postbiotic and subsequently infected with sublethal doses of influenza virus were shown to have lower viral loads in lung tissue and increased cytotoxic CD8+ T cells, indicating heterologous protection. [14]. In addition, nonspecific DC maturation was observed with overexpression of antigen presentation markers, polyclonal B cell activation with a significant increase in IgG serum levels, and trends toward increased influenza-specific IgA and IgG in the airways. These mice also showed little protection against secondary bacterial infections. [14] This same finding of protection against secondary bacterial infection was reported in a later study with protection against Streptococcus pneumoniae. [15] Furthermore, in a recent study, it was reported that immunized mice with a Postbiotic showed protection against respiratory syncytial virus, mediated by increased levels of IFNβ. [16]
Although some research demonstrates the involvement of DC-TLRs in the interaction with bacterial lysates, most of these studies were performed on cell lines or with mouse bone marrow-derived DC. In some of these studies, no more than two TLRs were analyzed (TLR2/TLR4, TLR2/TLR6, or TLR4/7), and some molecules of the signaling pathway, including MyD88 (an adaptor intracellular signaling molecule) [17]. The above limits the understanding of the preventive and therapeutic effects of Postbiotic in human respiratory infections, and which different TLR receptor profiles are involved in the dendritic cell activation compared to mice.
Pulmonarom® is used for the treatment and prevention of viral and bacterial respiratory infections. It is available for sale to the public, making it an easily accessible alternative for people. However, it is important to define its effects on the immune system, with dendritic cells being of the greatest interest due to their implications in the innate and adaptive immune responses. Thus, this study aims to evaluate DC, activation markers, immunoinflammatory cytokines, and TLR overexpression pattern profile induced by Pulmonarom®.
2. Material and Methods
2.1. Monocyte-Derived Dendritic Cells Obtention
Monocyte-derived dendritic cells (moDC) were differentiated from CD14+ monocytes as described previously. [18,19] Peripheral blood monocytes were purified from buffy coats from healthy donors, which were kindly supplied by the blood bank of the National Institute of Cardiology “Ignacio Chávez”. The handling of blood samples was performed according to the Declaration of Helsinki, donors signed informed consent forms, and the local scientific and ethics committees approved the protocol. PBMC were separated by gradient centrifugation with Histopaque-1077 (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). For this, buffy coats were diluted 1:2 WITH D-PBS to form the gradient and centrifuged at 400× g at 24 °C for 45 min. Then, the layer containing mononuclear cells WAS recovered, and cells were washed three times with D-PBS at 300× g at 4 °C for 10 min. Afterward, mononuclear cells were blocked with BSA contained in a blocking buffer (D-PBS, 2 mM EDTA, and 0.5% BSA cell culture grade) and incubated at 4 °C for 15 min. Immediately, mononuclear cells were incubated with anti-CD14 magnetic microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany) for 20 min at 4 °C and washed with D-PBS. Cells marked with anti-CD14 were separated through an LS magnetic column coupled to a magnet (Miltenyi Biotec, Bergisch Gladbach, Germany). The column was washed three times with 3 mL of D-PBS to eliminate CD14—cells and then separated from the magnet, which permitted the recovery of CD14+ monocytes with the help of a plunger and D-PBS. Afterward, monocytes were counted and seeded in RPMI-1640 medium with stable glutamine (Biowest, Riverside, MO, USA) supplemented with 10% FBS, 100 U/mL penicillin, 100 µg/mL streptomycin, and 50 µM 2-mercaptoethanol at pH 7.2 (R-10 medium) at a density of 1 × 106 cells/mL on 24-well tissue culture-treated plates The following day, fresh R-10 medium enriched with 500 U/mL granulocyte-macrophage–colony-stimulating factor (GM-CSF) and 1000 U/mL IL-4 (BD Biosciences, Franklin Lakes, NJ, USA) was used to replace 50% of the medium of each well to promote the differentiation of CD14+ monocytes to immature moDC. This procedure was repeated during days 2 and 4 of culture. moDCs with an immature phenotype were harvested on days 5–6 of culture and used for all the assays.
2.2. Lyophilization and Protein Quantification
Commercial bacterial lysate Pulmonarom® was purchased at a pharmacy. Pulmonarom® is sold in a commercial presentation of 3 ml glass ampoules containing bacterial lysates. The 3 mL of Pulmonarom® was dried (-120°C, Maxi Dry Lyo). Protein concentration was quantified using the Bradford method (Bradford 1976). The lyophilized powder was stored, and the concentration was adjusted until used.
2.3. moDC Stimulus
The moDC were stimulated with 0.01, 0.1, and 0.5 µg of Pulmonarom® protein per well in 24 wells of the cell culture plate. They were then incubated for 24h at 37ºC with 5% CO2. Thereafter, supernatants were collected, and the cells were processed for flow cytometry.
2.4. Characterization of moDC
For moDC characterization by flow cytometry, markers evaluating the expression of the CD11c, CD1a, HLA, CD80, and CD86 in the flow cytometer (FACS Aria, Becton Dickinson, San Jose, CA, USA) and data were analyzed with the software (FlowJo BD Life Science, V10.8, 2023). The antibodies employed were anti-CD11c (eBioscience, San Diego, CA, USA), CD1a, CD80, HLA, and CD86 (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The DCs harvested on day 7 of culture presented a phenotype: CD11c+, CD1a+, HLAlo, CD80lo, and CD86lo, corresponding to the phenotype of an immature DC with a purity of≥ 80%.
2.5. Scanning Transmission Electron Microscopy.
Samples were fixed with Karnovsky (2.5% glutaraldehyde/2% formaldehyde) and washed with 0.1M cacodylates. They were postfixed with osmium tetroxide OsO4 for 2 h, contracted with 1% uranyl for 1 h, and dehydrated with increasing alcohol concentrations: 70, 80, 90, 100, 100, and 100. 2 changes of propylene oxide were added for 30 min each. They were pre-embedded with propylene oxide/Epon resin 2:1 TN, 1:1 TN, and pure TN resin. The inclusion was carried out at 60 degrees C. Semi-thin sections of 1 micron were made, which were stained with toluidine blue. Thin sections of 100 nm were made and observed in the Crossbeam 550 field emission electron microscope STEM (Scanning Transmission Electron Microscopy) mode.
2.6. Cytokine Production
Cytokines were quantified in cell supernatants after 24 hours of stimulation with Pulmonarom®, using the Kit Human Anti-Virus Response Panel 1 (Biolegend, Cat. 741270, San Diego, California) and the Human Essential Immune Response Panel (Biolegend, Cat. 740930). Following the supplier's instructions. Briefly, supernatants were incubated with capture beads for 2 hours; thereafter, they were washed with wash buffer (buffer provided in the kit), and then detection biotinylated antibodies were added and incubated for 1 hour at room temperature with constant shaking (330 rpm on an orbital shaker). Without washing, streptavidin was added and incubated for another 30 min. Finally, the beads were washed and prepared for the flow cytometer.
2.7. Identification and Expression of Toll-like Receptors (TLRs)
Cells were recovered by cooling the culture plates to 4°C for 30 min and then washing them with cold phosphate-buffered saline (PBS; NaCl: 137 mM., KCl: 2.7 mM. Na2HPO4: 10 mM. KH2PO4: 1.8 mM). The viability and number of the cells were assessed using trypan blue staining. Cells used for flow cytometry assays had a viability greater than 85%. Cells were resuspended in FACS buffer (PBS + 1% bovine serum albumin (BSA)). Cells were adjusted to a concentration of 50,000 cells per tube and stained for flow cytometry assessment. The assay started with viability staining using the Zombie Aqua Fixable Viability Kit (Biolgend Cat. 423102), washed with PBS, and then extracellular staining was performed by incubating the antibodies for 30 min with FACS buffer and then washed with PBS. The extracellular antibodies were anti-CD11c-PECy7 (Biolegend Cat. 337216), anti-HLA-DR AF700 (Biolegend Cat. 307626), anti-TLR2 AF 647 (Biolegend Cat. 309714), anti-TLR4 BV 412 (Biolegend Cat. 312811), and anti-TLR6 PE (Biolegend Cat. 334708). For isotype controls the following antibodies were used: PE Mouse IgG2a k isotype control (BioLegend Cat. 400211) and APC Mouse IgG1 k (Biolgend Cat. 400119).
Once the cells were stained extracellularly, they were fixed with a Fixation Buffer reagent (Biolgend Cat. 420801) for 20 minutes at room temperature. Thereafter, they were washed with wash buffer (Biolgend Cat. 421002), diluting the buffer to a 1x concentration. The cells were washed twice by centrifuging at 1500 rpm for 5 min at RT. The cells were incubated overnight (approximately 12 hours) with wash buffer and the intracellular antibodies used were: anti-TLR3 BV711 (Biolgend Cat. 309714), anti-TLR7 FITC (Biolegend Cat. 376908), anti-TLR8 PerCP CY5.5 (Biolegend Cat. 395512) and anti-TLR9 APC- Fire&TRADE 810 (Biolegend Cat. 394812). Finally, the cells were washed with 1x wash buffer and read by flow cytometer.
2.8. Flow Cytometry Analysis
Samples were read by the NxT Attune flow cytometer, capable of reading 13 colors at a time. Matrix compensation was performed with dendritic cells, and isotype controls were used in each experiment to ensure specific binding to cells. The files obtained were analyzed in FlowJo 10.8 software. Single-parameter analyses were performed using the analysis strategy presented in Supplementary Figure 1 and Figure 2.
2.9. Statistical Analysis
The data were first analyzed for distribution using a Shapiro-Wilks test, as appropriate. Data with a normal distribution were analyzed with parametric tests, while data with a non-normal distribution were analyzed with non-parametric tests. In the parametric analysis, ANOVA- Dunnett test was used to compare the groups. Mean ± standard deviation (SD) was plotted in each graph.
3. Results
3.1. Pulmonarom® Extract Induces Ultrastructural Morphological Changes in moDC.
Monocyte-derived dendritic cells (moDCs) were cultured with different concentrations of Pulmonaron® (from 0.01 to 0.5 µg/well). For the MET study, moDC were analyzed without stimulation and stimulated with 0.5 mg of Pulmonarom®. As shown in Figure 1, Pulmonaron® induced ultrastructural morphological changes of moDC as observed in Transmission electron microscopy (TEM)
3.2. Pulmonarom® Extract Increases Class II Histocompatibility Molecules in Dendritic Cells.
Considering the morphological changes induced by Pulmonaron® over moDC, we wondered if these changes corresponded to an activation of the cells. Figure 2 shows that Pulmonaron® at a dose of 0.5 µg/well induces a significant increase in class II histocompatibility molecules HLA-DR, which is related to their activation.
3.3. Pulmonarom® Extract Increases the Expression of TLRs 2, 3, 6, and 7
As it has been reported that activated dendritic cells can increase the expression of TLR, we studied if Pulmonarom® exerts these effects. As shown in Figure 3, moDC stimulated in vitro with Pulmonarom® increased the expression of TLR2, TLR3, TLR6, and TLR7. This increase was more evident with Pulmonarom® concentration 0.5 µg/well. For TLR6, a high increase was detected at 0.1 and 0.5 µg/well of Pulmonarom®. No significant effects were observed on the overexpression of TLR4, TLR8, and TLR9.
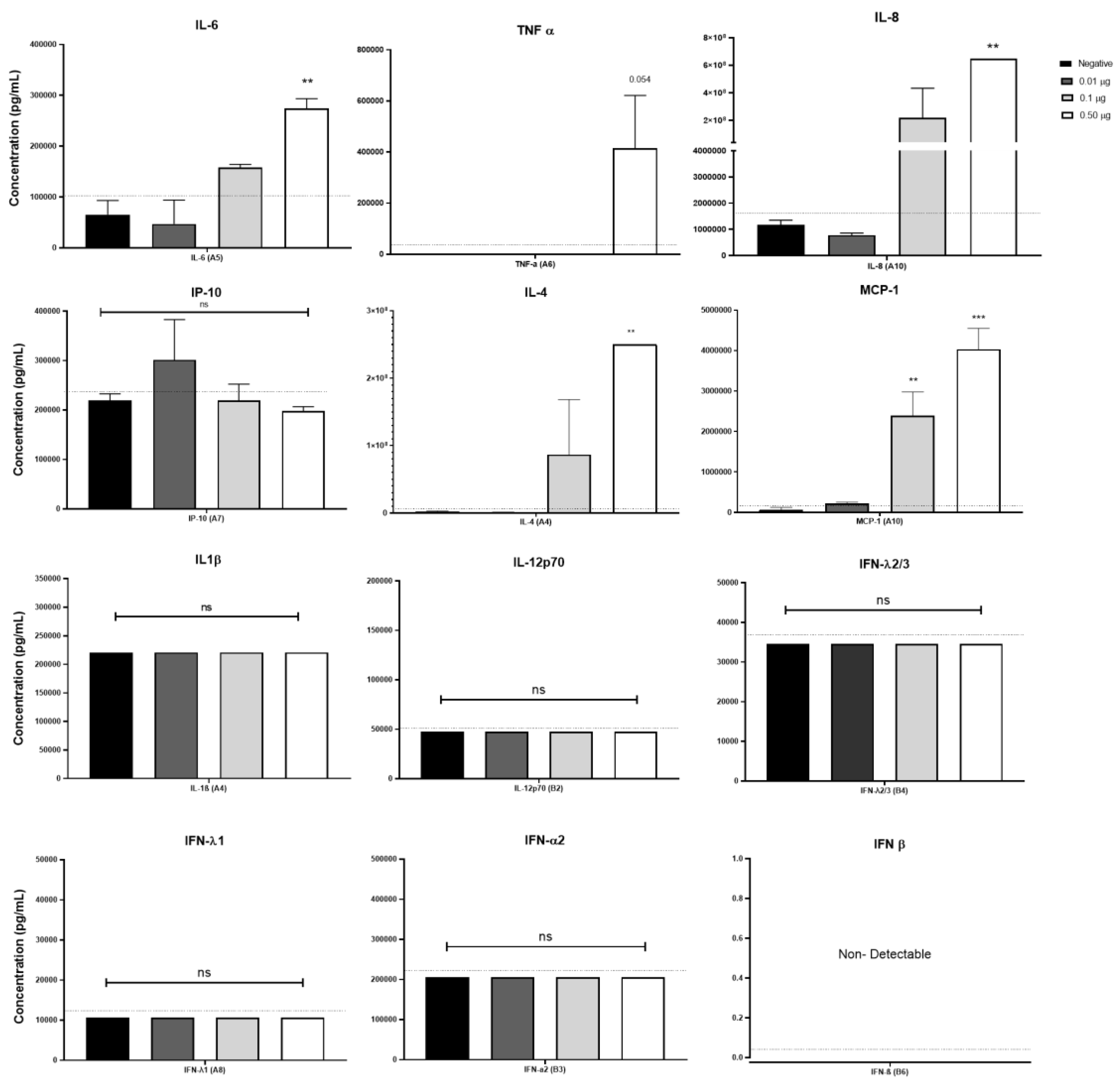
3.4. Pulmonarom® Increases the Production of IL-4, IL-6, IL-8 and MCP-1.
It is well known that some bacterial lysates can induce the expression of inflammatory cytokines. [20,21] Considering that Pulmonarom® was able to induce the overexpression of some TLRs and class II MHC molecules, this activation profile may be probably accompanied by the induction of inflammatory cytokines. As shown in Figure 4, Pulmonarom® was capable of increasing IL-6, IL-8, IL-4, and MCP-1 by moDC after 24 hours of stimuli. No increase in the most inflammatory cytokines (IL-1β and TNF-α) and of the type I IFNs (alfa, beta y lambda) was detected.
4. Discussion
Dendritic cells play a crucial role in linking innate and adaptive immune responses to deal with pathogens. During viral and bacterial infections, the proper functioning of dendritic cells is essential for the elimination of pathogens by recognizing pathogen-associated molecular patterns (PAMPs) [19] that interact with pattern recognition receptors (PRRs) on their membrane, such as Toll-like receptors (TLRs). DC activation, through various PAMPs, enhances the maturation of phagosomes and increases the phagocytosis of pathogens [22].
Therefore, DC activation is pivotal for an adequate activation of the immune system. Additionally, DC activation triggers the production of cytokines, which boost the defense against pathogens, including pro-inflammatory cytokines like TNFα. [23] or defensins such as β-defensin. [24] This activation also increases adhesion molecules in the endothelium, leading to greater cellular infiltration in the lesion area, thereby aiding in the elimination of various pathogens [25].
Several approaches to prevent and deal against respiratory infections have been applied worldwide, and since the 1950s, bacterial lysates have been used to reduce the incidence of recurrent respiratory tract infections. [26] Since then, commercially available bacterial lysates, including Ismigen, LW 50020, OM-85, Lantigen B and Pulmonarom®, have been prescribed in children and adults.
Pulmonarom® is a commercial bacterial lysate marketed in Mexico for 90 years with the registration number 13485 SSA IV. It is composed of 6 x 108UFC of each of the following bacteria: Haemophilus influenzae, Staphylococcus aureus, Moraxella catarrhalis, Klebsiella pneumoniae, Streptococcus pneumoniae, Streptococcus pyogenes, Streptococcus agalactiae, Streptococcus dysgalactiae, and Streptococcus anginosus.
Pulmonarom® is indicated for the prophylaxis of respiratory infections during endemic periods; and, since it reduces the duration of treatment for respiratory infections, it is also indicated as an adjuvant in these treatments. Pulmonarom® is also indicated for nonspecific respiratory tract conditions that present without fever but with local irritation, congestion, and inflammation of the airways related to environmental pollutants. Pulmonarom® is also indicated for the treatment of recurrent respiratory infections.
Despite the broadly employment of Pulmonarom® in Mexico, no studies have studied the mechanisms through which it can exert its therapeutic and preventive functions against respiratory tract infections. Thus, we deep inside the effects of this commercial Postbiotic on dendritic cells activation, considering the relevant role of these cells in modulating the innate and adaptive immune response.
In this study, we found that Pulmonarom® is capable of increasing antigen presentation molecules on the surface of moDC (Figure 2). These changes are accompanied by morphological alterations in moDC cultures, indicating the maturation and activation of dendritic cells induced by postbiotic [27,28] These effects on DC could imply an improved potential to mobilize lymph nodes and present antigens to initiate an effective adaptive immune response.
Transmission electron microscopy (TEM) analysis revealed ultrastructural changes that correlate with moDC maturation. Pulmonarom®-stimulated moDCs showed a greater number of dendrites, which were observed as a large number of semicircular bodies under TEM cross-section, as well as an increase in the number of lysosomes (Figure 1).
In immature DCs, internalized antigens are slowly degraded and inefficiently utilized for peptide loading by MHC-II; however, during maturation, the vacuolar proton pump is activated, which enhances lysosomal acidification and antigen proteolysis, facilitating the efficient formation of peptide-MHC class II complexes. Therefore, it is suggested that lysosomal function in DCs may play an important role in the processing of internalized antigens. [29] The importance of lysosomes in DC maturation, as well as their migration and chemotaxis capacity, is regulated by transient receptor potential melastatin 2 (TRPM2)-mediated lysosomal Ca2+ release. [30] When an appropriate DC activation occurred, the number of Lysosomes also increased, as observed in moDC stimulated with Pulmonarom®
While immature DC migration follows a random, homing pattern, mature DC migration exhibits a continuous, directional pattern toward lymph nodes for antigen presentation to Th cells. This pattern of mature DC migration is regulated by lysosomal calcium levels. Although much is unknown about intracellular signaling in DC maturation, lysosomal signaling is crucial for this process. When DC detect foreign particles, intracellular signaling is initiated, leading to lysosomal calcium being released by the ionic channel TRPML1 (transient receptor potential cation channel, mucolipin subfamily, member 1), which activates the actin-based motor protein myosin II, located at the posterior end of the cell, promoting rapid and directional migration. Lysosomal calcium also activates the transcription factor EB (TFEB), which translocates to the nucleus to maintain TRPML1 expression, thus sustaining DC migration. [31] The TRPML1-TFEB interaction negatively regulates macropinocytosis upon detection of microorganisms by DCs, thus promoting that the DC is not “distracted” by capturing other antigens during its migration. [31]
In summary, to meet the dynamic functional requirements during DC maturation, extensive remodeling of lysosomal compartments occurs. These changes are necessary for lysosomal calcium efflux and the contraction of Actomyosin for DC motility and migration. Furthermore, increased lysosomal membrane permeability participates in inflammasome activation. Lysosomal signaling collaborates with intracellular signaling for antigen presentation and other immune response mechanisms. [32]
Furthermore, when evaluating the TLRs, we found changes predominantly in the TLR2, TLR3, TLR6, and TLR7 receptors, which recognize bacterial and viral patterns. TLR2 recognizes peptidoglycan, which is present in higher concentrations in Gram-positive bacteria such as Staphylococcus spp and Streptococcus sp, both of which cause multiple respiratory infections [25]. Similarly, TLR6 recognizes diacyl peptides, which are also primarily expressed in Gram-positive bacteria. Consequently, the increased expression of these TLRs enhances the recognition of bacteria that mainly affect the respiratory tract.
Alyanakian et al. demonstrated that other commercial lysates stimulated mouse dendritic cells and induced TGF-beta production by splenocytes in a TLR-2, TLR-4, and MyD88-dependent manner. [33]. In this work, we also found an increase in receptors such as TLR2 and TLR6, which can recognize nonspecific components of the bacterial membrane (Figure 2), as well as TLR3 and TLR7, which could enhance the antiviral response. In 2017, Dang et al. demonstrated that other commercial lysates induced the production of interferon-β through MyD88 and TRIF (another TLR adapter molecule) in mouse bone marrow-derived DCs [34].
Although we did not observe changes in interferon molecules, two factors were different: first, the use of this commercial lysate was in cells from C57BL/6 mice characterized by inflammatory responses [35], and second, the concentration of this commercial lysate used was 40 times higher than Pulmonarom in this study (Figure 4).
In addition, it has been reported that the participation of TLR6, TLR2, and TLR7 actively participates against some viral infections, such as the SARS-CoV-2 virus, which is recognized by TLR2 and subsequently by TLR7 for an effective anti-viral response. [36]. Another example is the role of TLR6 and TLR2 in the antiviral response against respiratory viral disease (RSV), where infection increases TLR2 expression and the absence of TLR2 or TLR6 leads to a higher viral load and lower production of pro-inflammatory cytokines. [37]. In this work, we found an increase in TLR2, TLR6, and TLR7 (Figure 3) that could favor the recognition of some common viruses in the respiratory tract.
On the other hand, Pulmonarom ® increased the expression of TLR3, this receptor is a specialized receptor for the recognition of dsRNA, so it is of great importance in viral infections. It has been reported that its participation is critical for the response against RSV in epithelial cells of the respiratory tract. [38] In addition, TLR3 is involved in the recognition of the influenza virus. [39], Therefore, observing an increase in this receptor could increase the innate immune response and induce type I interferons in the presence of ligands to this receptor. [40], However, in cases of prolonged activation, it can cause dysfunction in the immune response. [41]. This is a very interesting result compared to other commercial lysates, which do not report increased TLR3 expression in their published models. Being such a ubiquitous receptor for virus recognition, it could be an advantage in functionality and coverage against various viruses that are recognized by TLR3.
The above indicates that the increase in TLRs by Pulmonarom® could impact a better response against viruses and bacteria, by increasing the activation of dendritic cells and their expression of TLRs, however, this has to be tested in vivo models. It should be noted that the safety of the extract, in the viability of the cells is an important point since it does not represent a risk for dendritic cells (Supplementary Figure 3).
Pulmonarom® induced the production of IL-6, IL-8 and MCP-1, indicating that it is activating NFKB in dendritic cells, a result similar to that obtained using OM-85 BV in vitro using a mouse macrophage cell line, they demonstrated that the production of IL-1β, IL-6 and TNF-α is induced through activation mediated by the TLR4 and TLR2 signaling pathway through ERK1/2 and NF-κB. [42]. In this work we did not explore the signal pathway, this is an important point to research in the future to know the molecular mechanism of Pulmonarom®
IL-6 and IL-8 are two cytokines that have been widely studied in the inflammatory response.
On the one hand, IL-6 is an interleukin with pleiotropic functions; in some contexts, it can increase the immune response by increasing the specificity of CD8 T lymphocytes. [43] and in others, promote infection and sometimes generate exacerbated inflammation, as occurs in the context of SARS-CoV-2 [44]. The participation of this cytokine at the beginning of an infection could be beneficial by increasing the phagocytic capacity of neutrophils and macrophages. [46,47] However, it is an important result that points out the importance of using in vivo models in future studies, to know the role that IL-6 may have in an infectious or inflammatory context in the presence of Pulmonarom®.
IL-8 is a chemotactic cytokine for neutrophils, cells of the immune system with great phagocytic, inflammatory and immunological capacity, being crucial to limit the replication of viruses in early stages. [45] and even, if antibodies exist previously, they can increase the elimination of viruses through antibody-mediated phagocytosis as occurs in antibodies against hemagglutinin of the influenza A virus. [46]. However, IL-8 also participates in the adaptive immune response, since the ability to allow the recruitment of effector memory T lymphocytes with greater cytotoxic activity has been reported. [47]. However, it is important to note the duality that IL-8 presents in inflammatory contexts, since it has also been seen associated in adults infected with the influenza virus, which is negatively correlated with the production of antibodies against the virus. [48] This is probably because IL-8 can recruit myeloid suppressor cells (MDSC), which are increased in adult stages. [49,50]. This highlights the importance of conducting in vivo studies to evaluate the effect on young and old mice, and thus understand the implications that the treatment could have on people susceptible to viral infections.
The immune response needs to have the ability to recognize and present antigen to develop an effective immune response.
In summary, the increased expression of HLA-DR molecules, as well as TLR2, TLR3, TLR6, and TLR7 receptors, accompanied by the increased production of cytokines IL-8, MCP-1, IL-6 and IL-4, could be indicative of a potential training process in moDCs following their activation with the bacterial lysate Pulmonarom® [51,52] This training may explain the protective effect of this bacterial lysate against certain respiratory infections. However, further studies are required to evaluate the presence of additional markers that are considered classic signs of immune training, signal pathways, and specific alterations in the energy metabolism of moDCs [53], as well as challenges in biological models.
5. Conclusion
Pulmonarom® is a safe product currently marketed in Mexico for the treatment of respiratory infections. In this study, we found that Pulmonarom® can modify dendritic cells, and these effects are likely related to the observed increase in the immune response in humans following its treatment.
Authors Contributions
Conceptualization: RVL, Data curation: RVL, JAH-A, SGS-G, AWR, JDD, AGL, CCG, Formal Analysis: RVL, JAH-A, SGS-G, Writing – original draft: RVL, JAH-A, SGS-G, Writing – review & editing: RVL, LG-K, GF.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
The authors declare that this study received funding from Opella, Sanofi Consumer Healthcare business unit.
Acknowledgments
The authors would like to thank Dr. Jesica Pozos Hernández for their comments and suggestions. We also thank Dr. Rebeca Illiana Arévalo Martínez, Dr. Salvador Bueno Valenzuela, and Dr. Ibarra for their institutional support in carrying out this project. JAH-a thanks SECIHTI for scholarship CVU: 967829.
Conflict of Interest
The authors have no conflicts of interest to declare.
References
- Sirota SB, Doxey MC, Dominguez RMV, Bender RG, Vongpradith A, Albertson SB, et al. Global, regional, and national burden of upper respiratory infections and otitis media, 1990–2021: a systematic analysis from the Global Burden of Disease Study 2021. Lancet Infect Dis [Internet]. 2024 Jan 1 [cited 2025 Jan 19];25[1]:36–51. Available from: http://www.thelancet.com/article/S1473309924004304/fulltext.
- Bender, R.G.; Sirota, S.B.; Swetschinski, L.R.; Dominguez, R.-M.V.; Novotney, A.; E Wool, E.; Ikuta, K.S.; Vongpradith, A.; Rogowski, E.L.B.; Doxey, M.; et al. Global, regional, and national incidence and mortality burden of non-COVID-19 lower respiratory infections and aetiologies, 1990–2021: a systematic analysis from the Global Burden of Disease Study 2021. Lancet Infect. Dis. 2024, 24, 974–1002. [Google Scholar] [CrossRef]
- Global immunization efforts have saved at least 154 million lives over the past 50 years [Internet]. [cited 2025 Mar 20]. Available from: https://www.who.int/news/item/24-04-2024-global-immunization-efforts-have-saved-at-least-154-million-lives-over-the-past-50-years.
- Communities vulnerable without immunization against infectious diseases [Internet]. [cited 2025 Mar 20]. Available from: https://www.who.int/malaysia/news/detail/23-04-2020-communities-vulnerable-without-immunization-against-infectious-diseases.
- Coviello, S.; Wimmenauer, V.; Polack, F.P.; Irusta, P.M. Bacterial lysates improve the protective antibody response against respiratory viruses through Toll-like receptor 4. Hum. Vaccines Immunother. 2014, 10, 2896–2902. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Cunningham, M.; Hill, C. Frequently asked questions about the ISAPP postbiotic definition. Front. Microbiol. 2024, 14, 1324565. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Kumar, A.; Green, K.M.; Rawat, M. A Comprehensive Overview of Postbiotics with a Special Focus on Discovery Techniques and Clinical Applications. Foods 2024, 13, 2937. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.; Murphy, K.M. Dendritic cell regulation of TH1-TH2 development. Nat. Immunol. 2000, 1, 199–205. [Google Scholar] [CrossRef]
- A Itano, A.; McSorley, S.J.; Reinhardt, R.; Ehst, B.D.; Ingulli, E.; Rudensky, A.Y.; Jenkins, M.K. Distinct Dendritic Cell Populations Sequentially Present Antigen to CD4 T Cells and Stimulate Different Aspects of Cell-Mediated Immunity. Immunity 2003, 19, 47–57. [Google Scholar] [CrossRef]
- Gallo PM, Gallucci S. The Dendritic Cell Response to Classic, Emerging, and Homeostatic Danger Signals. Implications for Autoimmunity. Front Immunol [Internet]. 2013 [cited 2025 Feb 8];4(JUN):138. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC3677085/.
- Solano-Gálvez, S.G.; Tovar-Torres, S.M.; Tron-Gómez, M.S.; Weiser-Smeke, A.E.; Álvarez-Hernández, D.A.; Franyuti-Kelly, G.A.; Tapia-Moreno, M.; Ibarra, A.; Gutiérrez-Kobeh, L.; Vázquez-López, R. Human Dendritic Cells: Ontogeny and Their Subsets in Health and Disease. Med Sci. 2018, 6, 88. [Google Scholar] [CrossRef] [PubMed]
- Rial, A.; Lens, D.; Betancor, L.; Benkiel, H.; Silva, J.S.; Chabalgoity, J.A. Intranasal Immunization with a Colloid-Formulated Bacterial Extract Induces an Acute Inflammatory Response in the Lungs and Elicits Specific Immune Responses. Infect. Immun. 2004, 72, 2679–2688. [Google Scholar] [CrossRef]
- Pasquali, C.; Salami, O.; Taneja, M.; Gollwitzer, E.S.; Trompette, A.; Pattaroni, C.; Yadava, K.; Bauer, J.; Marsland, B.J. Enhanced Mucosal Antibody Production and Protection against Respiratory Infections Following an Orally Administered Bacterial Extract. Front. Med. 2014, 1, 41–41. [Google Scholar] [CrossRef]
- Rossi, G.A.; Bessler, W.; Ballarini, S.; Pasquali, C. Evidence that a primary anti-viral stimulation of the immune response by OM-85 reduces susceptibility to a secondary respiratory bacterial infection in mice. Ital. J. Pediatr. 2018, 44, 112. [Google Scholar] [CrossRef] [PubMed]
- Antunes, K.H.; Cassão, G.; Santos, L.D.; Borges, S.G.; Poppe, J.; Gonçalves, J.B.; Nunes, E.d.S.; Recacho, G.F.; Sousa, V.B.; Da Silva, G.S.; et al. Airway Administration of Bacterial Lysate OM-85 Protects Mice Against Respiratory Syncytial Virus Infection. Front. Immunol. 2022, 13, 867022. [Google Scholar] [CrossRef]
- Duan, T.; Du, Y.; Xing, C.; Wang, H.Y.; Wang, R.-F. Toll-Like Receptor Signaling and Its Role in Cell-Mediated Immunity. Front. Immunol. 2022, 13, 812774. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Lanzavecchia, A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J. Exp. Med. 1994, 179, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Romani, N.; Gruner, S.; Brang, D.; Kämpgen, E.; Lenz, A.; Trockenbacher, B.; Konwalinka, G.; O Fritsch, P.; Steinman, R.M.; Schuler, G. Proliferating dendritic cell progenitors in human blood. J. Exp. Med. 1994, 180, 83–93. [Google Scholar] [CrossRef]
- Navarro, S.; Cossalter, G.; Chiavaroli, C.; Kanda, A.; Fleury, S.; Lazzari, A.; Cazareth, J.; Sparwasser, T.; Dombrowicz, D.; Glaichenhaus, N.; et al. The oral administration of bacterial extracts prevents asthma via the recruitment of regulatory T cells to the airways. Mucosal Immunol. 2011, 4, 53–65. [Google Scholar] [CrossRef]
- Huber M MHBW. Eur J Med Res. 2005 [cited 2022 Nov 29]. p. 209–17 Th1-orientated immunological properties of the bacterial extract OM-85-BV - PubMed. Available from: https://pubmed.ncbi.nlm.nih.gov/15946922/.
- McInturff, J.E.; Modlin, R.L.; Kim, J. The Role of Toll-like Receptors in the Pathogenesis and Treatment of Dermatological Disease. J. Investig. Dermatol. 2005, 125, 1–8. [Google Scholar] [CrossRef]
- Ruby, J.; Bluethmann, H.; Peschon, J.J. Antiviral Activity of Tumor Necrosis Factor (TNF) Is Mediated via p55 and p75 TNF Receptors. J. Exp. Med. 1997, 186, 1591–1596. [Google Scholar] [CrossRef]
- Hertz, C.J.; Wu, Q.; Porter, E.M.; Zhang, Y.J.; Weismüller, K.-H.; Godowski, P.J.; Ganz, T.; Randell, S.H.; Modlin, R.L. Activation of Toll-Like Receptor 2 on Human Tracheobronchial Epithelial Cells Induces the Antimicrobial Peptide Human β Defensin-2. J. Immunol. 2003, 171, 6820–6826. [Google Scholar] [CrossRef]
- Le, J.; Kulatheepan, Y.; Jeyaseelan, S. Role of toll-like receptors and nod-like receptors in acute lung infection. Front. Immunol. 2023, 14, 1249098. [Google Scholar] [CrossRef]
- Rahman, M.; Grice, I.D.; Ulett, G.C.; Wei, M.Q. Advances in Bacterial Lysate Immunotherapy for Infectious Diseases and Cancer. J. Immunol. Res. 2024, 2024, 4312908. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Kim, J. Properties of immature and mature dendritic cells: phenotype, morphology, phagocytosis, and migration. RSC Adv. 2019, 9, 11230–11238. [Google Scholar] [CrossRef]
- Landmann, S.; Mühlethaler-Mottet, A.; Bernasconi, L.; Suter, T.; Waldburger, J.-M.; Masternak, K.; Arrighi, J.-F.; Hauser, C.; Fontana, A.; Reith, W. Maturation of Dendritic Cells Is Accompanied by Rapid Transcriptional Silencing of Class II Transactivator (Ciita) Expression. J. Exp. Med. 2001, 194, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, E.S.; Ebersold, M.; Garrett, W.; Pypaert, M.; Mellman, I. Activation of Lysosomal Function During Dendritic Cell Maturation. Science 2003, 299, 1400–1403. [Google Scholar] [CrossRef]
- Sumoza-Toledo, A.; Lange, I.; Cortado, H.; Bhagat, H.; Mori, Y.; Fleig, A.; Penner, R.; Partida-Sánchez, S. Dendritic cell maturation and chemotaxis is regulated by TRPM2-mediated lysosomal Ca2+release. FASEB J. 2011, 25, 3529–3542. [Google Scholar] [CrossRef] [PubMed]
- Bretou, M.; Sáez, P.J.; Sanséau, D.; Maurin, M.; Lankar, D.; Chabaud, M.; Spampanato, C.; Malbec, O.; Barbier, L.; Muallem, S.; et al. Lysosome signaling controls the migration of dendritic cells. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef]
- Yang, J.; Rong, S.-J.; Zhou, H.-F.; Yang, C.; Sun, F.; Li, J.-Y. Lysosomal control of dendritic cell function. J. Leukoc. Biol. 2023, 114, 518–531. [Google Scholar] [CrossRef]
- Alyanakian MA, Grela F, Aumeunier A, Chiavaroli C, Gouarin C, Bardel E, et al. Transforming Growth Factor-β and Natural Killer T-Cells Are Involved in the Protective Effect of a Bacterial Extract on Type 1 Diabetes. Diabetes [Internet]. 2006 Jan 1 [cited 2022 Dec 4];55[1]:179–85. Available from: https://diabetesjournals.org/diabetes/article/55/1/179/12316/Transforming-Growth-Factor-and-Natural-Killer-T.
- Dang, A.T.; Pasquali, C.; Ludigs, K.; Guarda, G. OM-85 is an immunomodulator of interferon-β production and inflammasome activity. Sci. Rep. 2017, 7, srep43844. [Google Scholar] [CrossRef]
- Morokata T, Ishikawa J, Ida K, Yamada T. C57BL/6 mice are more susceptible to antigen-induced pulmonary eosinophilia than BALB/c mice, irrespective of systemic T helper 1/T helper 2 responses. Immunology [Internet]. 1999 [cited 2025 Mar 17];98[3]:345–51. Available from: https://pubmed.ncbi.nlm.nih.gov/10583592/.
- van der Sluis, R.M.; Cham, L.B.; Gris-Oliver, A.; Gammelgaard, K.R.; Pedersen, J.G.; Idorn, M.; Ahmadov, U.; Hernandez, S.S.; Cémalovic, E.; Godsk, S.H.; et al. TLR2 and TLR7 mediate distinct immunopathological and antiviral plasmacytoid dendritic cell responses to SARS-CoV-2 infection. EMBO J. 2022, 41, e109622. [Google Scholar] [CrossRef]
- Murawski, M.R.; Bowen, G.N.; Cerny, A.M.; Anderson, L.J.; Haynes, L.M.; Tripp, R.A.; Kurt-Jones, E.A.; Finberg, R.W. Respiratory Syncytial Virus Activates Innate Immunity through Toll-Like Receptor 2. J. Virol. 2009, 83, 1492–1500. [Google Scholar] [CrossRef]
- Groskreutz, D.J.; Monick, M.M.; Powers, L.S.; Yarovinsky, T.O.; Look, D.C.; Hunninghake, G.W. Respiratory Syncytial Virus Induces TLR3 Protein and Protein Kinase R, Leading to Increased Double-Stranded RNA Responsiveness in Airway Epithelial Cells. J. Immunol. 2006, 176, 1733–1740. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki A, Pillai PS. Innate immunity to influenza virus infection. Nature Reviews Immunology 2014 14:5 [Internet]. 2014 Apr 25 [cited 2025 Jan 26];14[5]:315–28. Available from: https://www.nature.com/articles/nri3665.
- Chen, Y.; Lin, J.; Zhao, Y.; Ma, X.; Yi, H. Toll-like receptor 3 (TLR3) regulation mechanisms and roles in antiviral innate immune responses. J. Zhejiang Univ. B 2021, 22, 609–632. [Google Scholar] [CrossRef] [PubMed]
- Stowell, N.C.; Seideman, J.; A Raymond, H.; A Smalley, K.; Lamb, R.J.; Egenolf, D.D.; Bugelski, P.J.; A Murray, L.; A Marsters, P.; A Bunting, R.; et al. Long-term activation of TLR3 by Poly(I:C) induces inflammation and impairs lung function in mice. Respir. Res. 2009, 10, 43–43. [Google Scholar] [CrossRef]
- Luan, H.; Zhang, Q.; Wang, L.; Wang, C.; Zhang, M.; Xu, X.; Zhou, H.; Li, X.; Xu, Q.; He, F.; et al. OM85-BV Induced the Productions of IL-1β, IL-6, and TNF-α via TLR4- and TLR2-Mediated ERK1/2/NF-κB Pathway in RAW264.7 Cells. J. Interf. Cytokine Res. 2014, 34, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, J.P.; Schanz, O.; Garbers, C.; Zaremba, A.; Hegenbarth, S.; Kurts, C.; Beyer, M.; Schultze, J.L.; Kastenmüller, W.; Rose-John, S.; et al. IL-6 trans-Signaling-Dependent Rapid Development of Cytotoxic CD8+ T Cell Function. Cell Rep. 2014, 8, 1318–1327. [Google Scholar] [CrossRef]
- Gubernatorova, E.; Gorshkova, E.; Polinova, A.; Drutskaya, M. IL-6: Relevance for immunopathology of SARS-CoV-2. Cytokine Growth Factor Rev. 2020, 53, 13–24. [Google Scholar] [CrossRef]
- George, S.T.; Lai, J.; Ma, J.; Stacey, H.D.; Miller, M.S.; Mullarkey, C.E. Neutrophils and Influenza: A Thin Line between Helpful and Harmful. Vaccines 2021, 9, 597. [Google Scholar] [CrossRef]
- Mullarkey, C.E.; Bailey, M.J.; Golubeva, D.A.; Tan, G.S.; Nachbagauer, R.; He, W.; Novakowski, K.E.; Bowdish, D.M.; Miller, M.S.; Palese, P. Broadly Neutralizing Hemagglutinin Stalk-Specific Antibodies Induce Potent Phagocytosis of Immune Complexes by Neutrophils in an Fc-Dependent Manner. mBio 2016, 7. [Google Scholar] [CrossRef]
- Hess, C.; Means, T.K.; Autissier, P.; Woodberry, T.; Altfeld, M.; Addo, M.M.; Frahm, N.; Brander, C.; Walker, B.D.; Luster, A.D. IL-8 responsiveness defines a subset of CD8 T cells poised to kill. Blood 2004, 104, 3463–3471. [Google Scholar] [CrossRef]
- Quach, H.Q.; Goergen, K.M.; E Grill, D.; Ovsyannikova, I.G.; A Poland, G.; Kennedy, R.B. IL-8 as a Causal Link Between Aging and Impaired Influenza Antibody Responses in Older Adults. J. Infect. Dis. 2025. [Google Scholar] [CrossRef]
- Bueno, V.; Sant’anna, O.A.; Lord, J.M. Ageing and myeloid-derived suppressor cells: possible involvement in immunosenescence and age-related disease. AGE 2014, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G.; Picard, E.; Bueno, V.; Verschoor, C.P.; Ostrand-Rosenberg, S. MDSCs, ageing and inflammageing. Cell. Immunol. 2021, 362, 104297. [Google Scholar] [CrossRef] [PubMed]
- Bekkering, S.; Domínguez-Andrés, J.; Joosten, L.A.; Riksen, N.P.; Netea, M.G. Trained Immunity: Reprogramming Innate Immunity in Health and Disease. Annu. Rev. Immunol. 2021, 39, 667–693. [Google Scholar] [CrossRef] [PubMed]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.B.; Ifrim, D.C.; Saeed, S.; Jacobs, C.; van Loenhout, J.; de Jong, D.; Stunnenberg, H.G.; et al. Bacille Calmette-Guérin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc. Natl. Acad. Sci. 2012, 109, 17537–17542. [Google Scholar] [CrossRef]
- Alcaraz-Serna, A.; Bustos-Morán, E.; Fernández-Delgado, I.; Calzada-Fraile, D.; Torralba, D.; Marina-Zárate, E.; Lorenzo-Vivas, E.; Vázquez, E.; de Alburquerque, J.B.; Ruef, N.; et al. Immune synapse instructs epigenomic and transcriptomic functional reprogramming in dendritic cells. Sci. Adv. 2021, 7, eabb9965. [Google Scholar] [CrossRef]
Figure 1.
Pulmonarom® simulates dendritic cells, increasing the vacuoles and vesicles. Transmission electron micrograph of human moDC, non-stimulated (A and C) and stimulated with 0.5 μg. Pulmonarom® (B and D). Scale bar 10mm (A and B) and 2 mm (C and D).
Figure 1.
Pulmonarom® simulates dendritic cells, increasing the vacuoles and vesicles. Transmission electron micrograph of human moDC, non-stimulated (A and C) and stimulated with 0.5 μg. Pulmonarom® (B and D). Scale bar 10mm (A and B) and 2 mm (C and D).
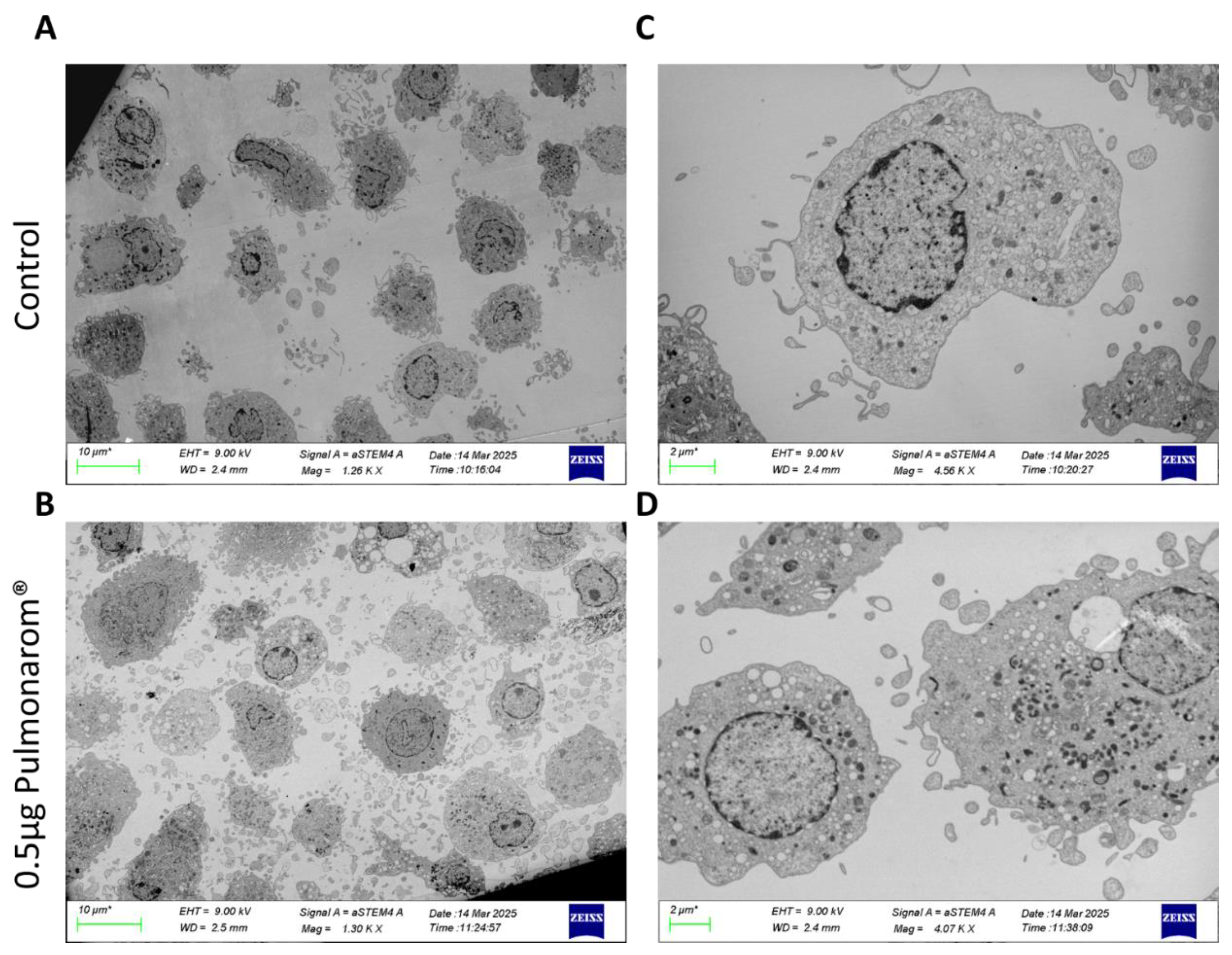
Figure 2.
Pulmonarom® increases the expression of MHC-II molecules in moDC. (A) Micrographs of cultured moDC depicting an increase in adherent cells, Scale bar 10 µm. (B) Representative Dot-plots from cell cultures, Left and right images correspond to non-stimulated and Pulmonarom® stimulated cells, respectively. An increase in HLA-DR expression is observed after stimulation with Pulmonarom®. (C) Representative histograms of HLA-DR expression. The negative control was treated only with culture medium, while the rest of the samples were stimulated with Pulmonarom® at different concentrations. An increase in the expression of the HLA-DR molecule is observed. (D) Quantification of HLA-DR expression using the geometric mean fluorescence intensity (gMFI). The normality of the data was assessed with a Shapiro-Wilks test. * p <0.05, ns: not statistical differences. ANOVA- Dunnett was used to compare the groups, finding only a significant difference in the 0.5 µg group. n=5 per group.
Figure 2.
Pulmonarom® increases the expression of MHC-II molecules in moDC. (A) Micrographs of cultured moDC depicting an increase in adherent cells, Scale bar 10 µm. (B) Representative Dot-plots from cell cultures, Left and right images correspond to non-stimulated and Pulmonarom® stimulated cells, respectively. An increase in HLA-DR expression is observed after stimulation with Pulmonarom®. (C) Representative histograms of HLA-DR expression. The negative control was treated only with culture medium, while the rest of the samples were stimulated with Pulmonarom® at different concentrations. An increase in the expression of the HLA-DR molecule is observed. (D) Quantification of HLA-DR expression using the geometric mean fluorescence intensity (gMFI). The normality of the data was assessed with a Shapiro-Wilks test. * p <0.05, ns: not statistical differences. ANOVA- Dunnett was used to compare the groups, finding only a significant difference in the 0.5 µg group. n=5 per group.
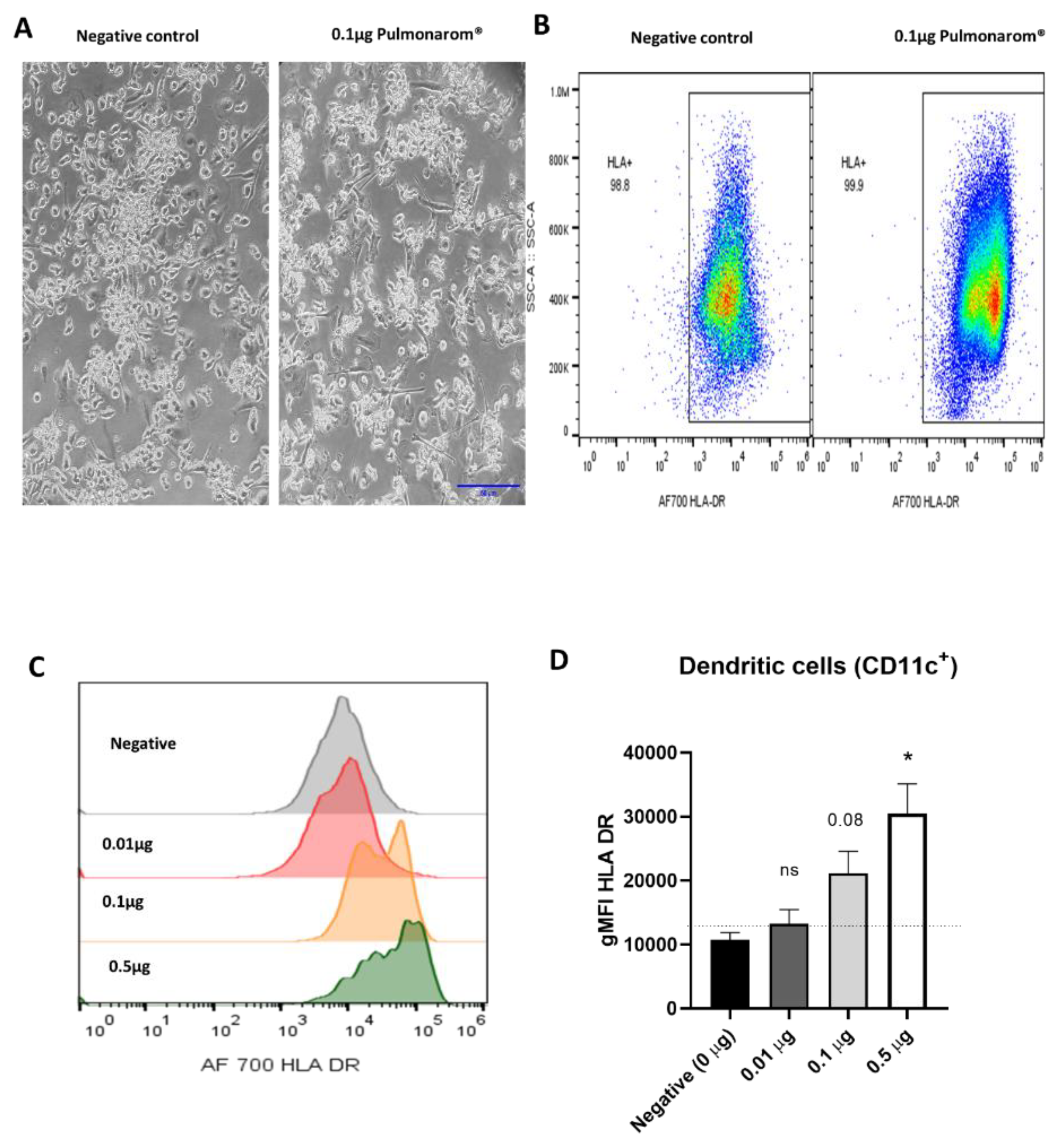
Figure 3.
Pulmonarom® increases the expression of TLRs 2, 6, 7, and 3 in dendritic cells. TLR expression in moDC phenotype as CD11c+ HLA DR+. Main changes are observed in TLR6, TLR7 and TLR3. The normality of the data was evaluated with a Shapiro-Wilks test. * p<0.05, **p<0.01 ns: no statistical differences. ANOVA- Dunnett was used to compare the groups. n=5 per group.
Figure 3.
Pulmonarom® increases the expression of TLRs 2, 6, 7, and 3 in dendritic cells. TLR expression in moDC phenotype as CD11c+ HLA DR+. Main changes are observed in TLR6, TLR7 and TLR3. The normality of the data was evaluated with a Shapiro-Wilks test. * p<0.05, **p<0.01 ns: no statistical differences. ANOVA- Dunnett was used to compare the groups. n=5 per group.
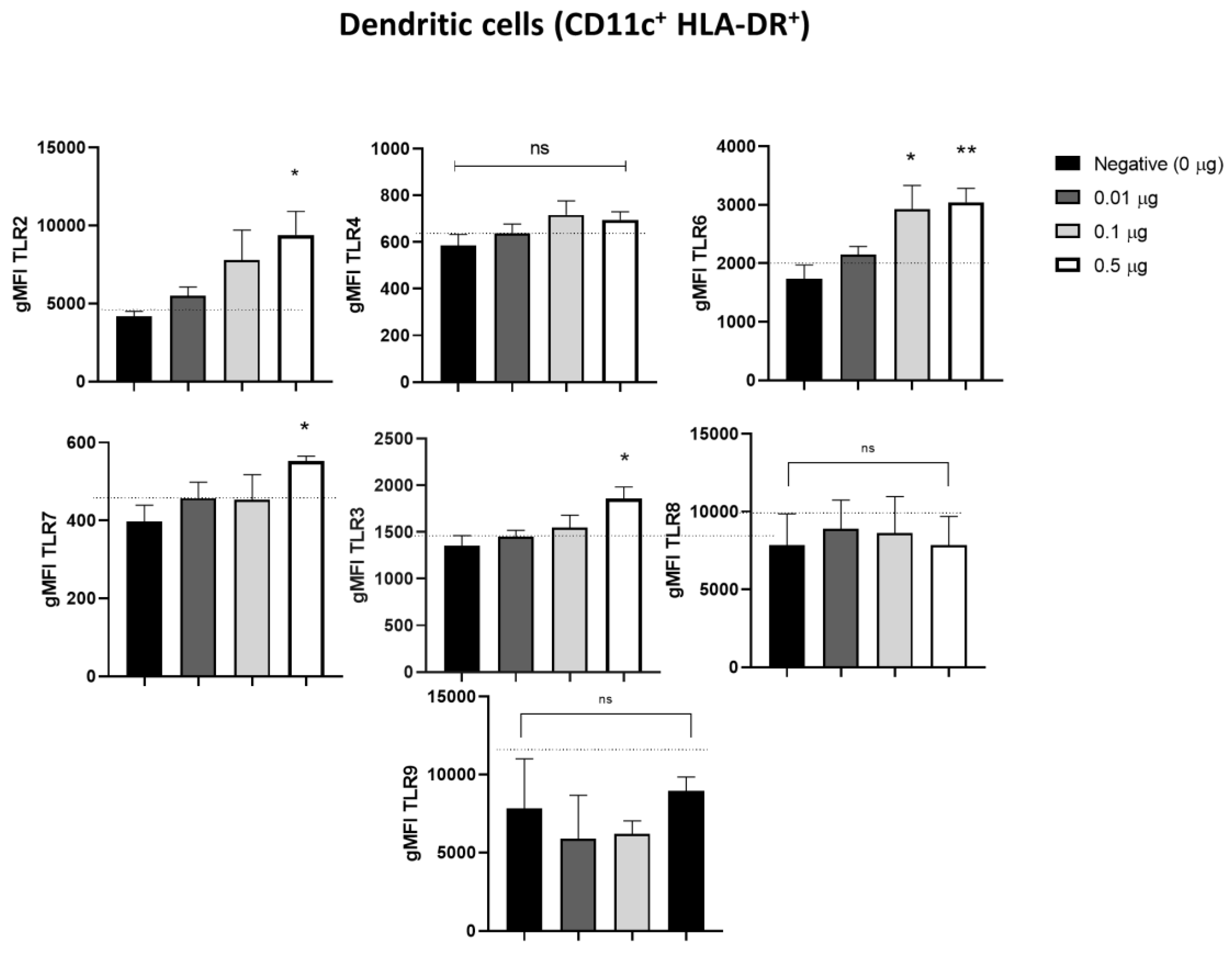
Figure 4.
Pulmonarom® increases the production of innate response cytokines. Monocyte-derived dendritic cells (moDCs) were cultured for 24 hours with different doses of Pulmonarom® (from 0.01 to 0.50 µg) to subsequently quantify the cytokines present in the culture supernatants. The results show a significant increase in the production of IL-6, IL-8, IL-4, and MCP-1. Mean ± standard deviation (SD) was plotted. Data normality was assessed with a Shapiro-Wilks test. ANOVA- Dunnett was used to compare the groups. * p<0.05, **p <0.01, ns: no statistical differences. n=5 per group.
Figure 4.
Pulmonarom® increases the production of innate response cytokines. Monocyte-derived dendritic cells (moDCs) were cultured for 24 hours with different doses of Pulmonarom® (from 0.01 to 0.50 µg) to subsequently quantify the cytokines present in the culture supernatants. The results show a significant increase in the production of IL-6, IL-8, IL-4, and MCP-1. Mean ± standard deviation (SD) was plotted. Data normality was assessed with a Shapiro-Wilks test. ANOVA- Dunnett was used to compare the groups. * p<0.05, **p <0.01, ns: no statistical differences. n=5 per group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



