Submitted:
12 November 2020
Posted:
13 November 2020
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
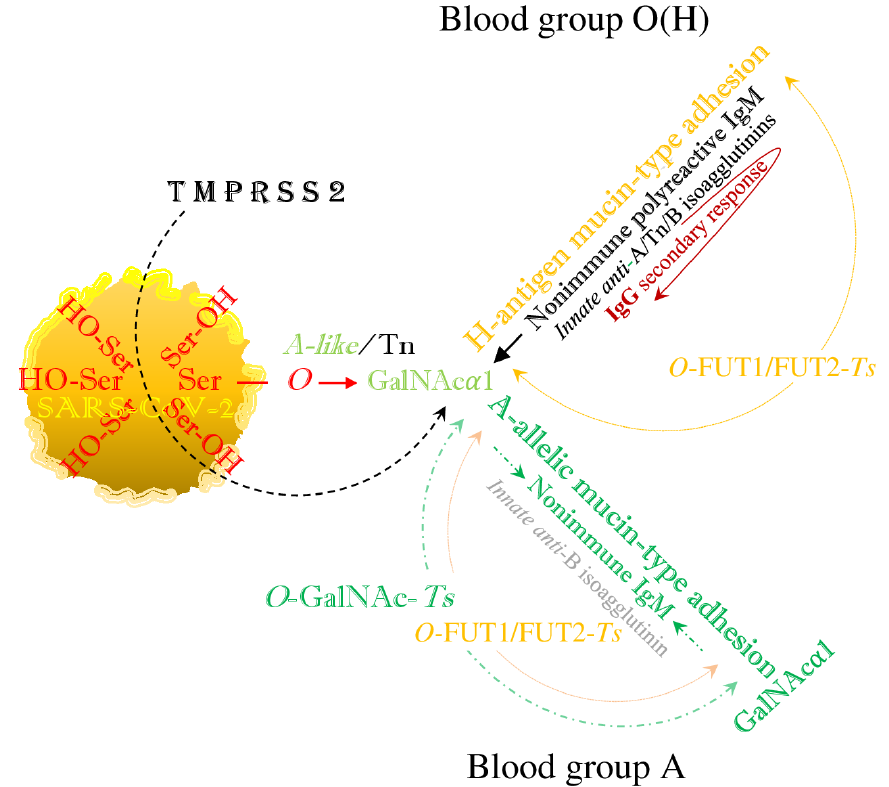
While the angiotensin converting enzyme 2 (ACE2) protein is defined as the primary severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) receptor, the viral serine molecule might be mobilized by the host's transmembrane protease serine subtype 2 (TMPRSS2) enzyme from the viral spike (S) protein and hijack the host’s N-acetyl-D-galactosamine (GalNAc) metabolism. The resulting hybrid, serologically A-like/Tn (T-nouvelle) structure potentially acts as a host–pathogen functional molecular bridge. In humans, this intermediate structure will hypothetically be replaced by ABO(H) blood group-specific, mucin-type structures, in the case of infection hybrid epitopes, implicating the phenotypically glycosidic accommodation of plasma proteins. The virus may, by mimicking the synthetic pathways of the ABO(H) blood groups, bind to the cell surfaces of the blood group O(H) by formation of a hybrid H-type antigen as the potential precursor of hybrid non-O blood groups, which does not affect the highly anti-glycan aggressive anti-A and anti-B isoagglutinin activities, exerted by the germline-encoded nonimmune immunoglobulin M (IgM). In the non-O blood groups, which have developed from the H-type antigen, these IgM activities are downregulated by phenotypic glycosylation, while adaptive immunoglobulins might arise in response to the hybrid A and B blood group structures, bonds between autologous carbohydrates and foreign peptides, suggesting the exertion of autoreactivity. The non-O blood groups thus become a preferred target for the virus, whereas blood group O(H) individuals, lacking the A/B phenotype-determining enzymes and binding the virus alone by hybrid H-type antigen formation, have the least molecular contact with the virus and maintain the critical anti-A and anti-B isoagglutinin activities, exerted by the ancestral IgM, which is considered the humoral spearhead of innate immunity.
Keywords:
COVID-19
; SARS-CoV-2–human carbohydrate interaction
; trans-species glycosylation
; A-like/Tn structure
; trans-species glycan bridge
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



