
Submitted:
15 January 2021
Posted:
18 January 2021
You are already at the latest version
Abstract
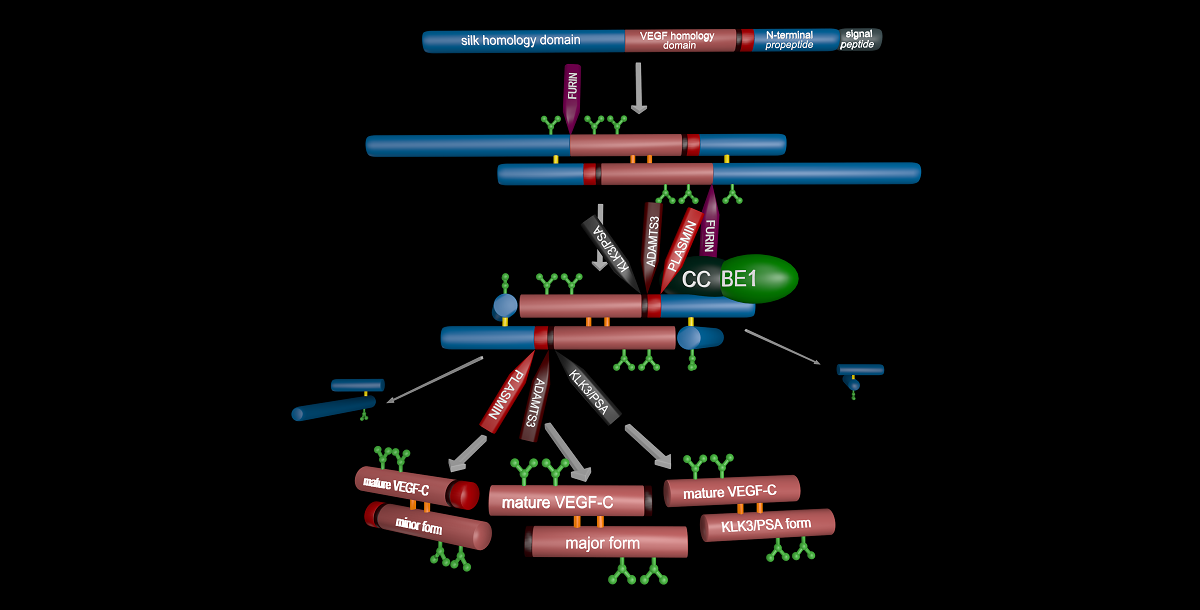
Specific proteolytic cleavages turn on, modify, or turn off the activity of vascular endothelial growth factors (VEGFs). Proteolysis is most prominent among the lymphangiogenic VEGF-C and VEGF-D, which are synthesized as precursors that need to undergo enzymatic removal of their C- and N-terminal propeptides before they can activate their receptors. The activating cleavage of VEGF-C is mediated by at least five different proteases: plasmin, ADAMTS3, prostate-specific antigen, cathepsin D, and thrombin. All of these proteases except for ADAMTS3 can also activate VEGF-D. Processing by different proteases results in distinct forms of the "mature" growth factors, which differ in affinity and receptor activation potential. The “default” VEGF-C-activating enzyme ADAMTS3 does not activate VEGF-D and therefore, VEGF-C and VEGF-D do function in different contexts. VEGF-C itself is also regulated in different contexts by different proteases. During embryonic development, ADAMTS3 activates VEGF-C. In contrast, thrombin and plasmin likely activate VEGF-C/-D during tissue injury-induced lymphangiogenesis, and PSA and cathepsin D perhaps during tumor-associated pathological lymphangiogenesis. Additionally, cathepsin D from saliva might activate latent VEGF-C/-D upon wound licking, thereby accelerating healing. Similar to tyrosine kinase receptors and VEGFs themselves, these activating proteases could be targeted to modulate angiogenesis and lymphangiogenesis in relevant diseases.
Keywords:
Vascular Endothelial Growth Factors (VEGFs)
; VEGF-A
; PlGF
; VEGF-B
; VEGF-C
; VEGF-D
; angiogenesis
; lymphangiogenesis
; CCBE1
; proteases
; ADAMTS3
; plasmin
; cathepsin D
; KLK3
; prostate-specific antigen (PSA)
; thrombin
; wound healing
; metastasis
; proteolytic activation
; vascular biology
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



