
Submitted:
25 March 2023
Posted:
27 March 2023
You are already at the latest version
Abstract
Alfalfa (Medicago sativa L.) is the most cultivated forage legume around the world. Under a variety of growing conditions, forage yield in alfalfa is stymied by biotic and abiotic stresses including heat, salt, drought, and disease. Given the sessile nature of plants, they use strategies such as differential gene expression to respond to environmental cues. Transcription factors control the expression of genes that contribute to or enable tolerance and survival during periods of stress. Basic-leucine zipper (bZIP) transcription factors have been demonstrated to play a critical role in regulating plant growth and development as well as mediate the responses to abiotic stress in several species, including Arabidopsis thaliana, Oryza sativa, Lotus japonicus, and Medicago truncatula. However, there is little information about bZIP transcription factors in cultivated alfalfa. In the present study, 237 bZIP genes were identified in alfalfa from publicly available sequencing data. Multiple sequence alignments showed the presence of intact bZIP motifs in the identified sequences. Based on previous phylogenetic analyses in Arabidopsis thaliana, alfalfa bZIPs were similarly divided and fell into 10 groups. The physicochemical properties, motif analysis, and phylogenetic study of the alfalfa bZIPs revealed high specificity within groups. The differential expression of alfalfa bZIPs in a suite of tissues indicates that particular bZIP genes are specifically expressed at different developmental stages in alfalfa. Similarly, expression analysis in response to ABA, cold, drought, and salt stresses, indicates that a subset of bZIP genes are also differentially expressed and likely play a role in abiotic stress signaling and/or tolerance. These expression patterns were further verified by qRT-PCR. However, further functional characterization of bZIP transcription factors in alfalfa will help illuminate the role they play in stress tolerance mechanisms in legumes and facilitate the molecular breeding of stress tolerance in alfalfa.
Keywords:
Alfalfa
; bZIP transcription factor
; phylogenetic analysis
; expression pattern
; abiotic stress
1. Introduction
Alfalfa (M. sativa L.) is a highly outcrossing forage legume, widely cultivated in the United States with approximately 16 million hectares (1). It is well suited for animal and livestock feed due to its high nutritional content. It also improves soil fertility through its symbiotic association with the soil bacterium Sinorhizobium meliloti for biological nitrogen fixation, which augments the nitrogen content in the soil for future crops (2–4). This deep-rooted perennial crop also helps to prevent soil erosion. However, genetic improvement in terms of forage yield has been relatively stagnant in alfalfa (5). Major hinderances are genomic complexity, severe inbreeding depression upon selfing, and self-incompatibility which complicate alfalfa breeding. Although the multi-purpose use of alfalfa increases its demand, adverse environmental conditions result in abiotic stresses, and ultimately hamper production. Breeding for stress resistance improves production to some extent; however, lack of completely annotated genome and expression profile data, eventually creates knowledge gap in fully understanding genotype and phenotype associations for stress-related traits.
The sessile nature of plants inevitably exposes them to adverse environmental conditions such as abiotic stress. However, plants have developed diverse mechanisms to cope with these abiotic stresses. One of them is the synthesis of proteins, metabolites, and other compounds to aid in survival through abiotic stress, which is often controlled by transcription factors (TFs). Transcription factors play a critical role in responses to environmental stresses via binding to cis-regulatory elements in promoters to regulate downstream gene expression. In plants, approximately 7% of the genome codes for transcriptional regulators, which bind promoter elements of downstream genes through their conserved sequence-specific DNA-binding domain (6). Among the 64 families (7) of transcription factors identified in the plant kingdom, the bZIP (basic leucine zipper) family is one of the largest and most diverse (7–9).
The basic leucine zipper (bZIP) family is distinguished by its highly conserved bZIP domain composed of 60 – 80 amino acids (10). Structurally, the bZIP domain is divided into two functionally distinct regions: a basic region and a leucine zipper motif (10). The basic region is composed of an invariant motif (N-x7-R/K-x9) of 18 amino acids residues that facilitates sequence-specific DNA binding, while the leucine zipper contains several heptad repeats of leucine or other bulky hydrophobic amino acids such as isoleucine, valine, phenylalanine, or methionine, for dimerization specificity (8,11). Molecular studies of bZIP genes in A. thaliana show that they are involved in the regulation of diverse biological processes including pathogen defense, light and stress signaling, seed maturation, and flower development (11). Additional information on the bZIP transcription factor family has provided evidence of their role in response to biotic and abiotic stresses in a diversity of plant species (11,12).
The availability of whole-genome sequences for plants allows the identification or prediction of bZIP TF family members at the genome-wide level. The number of bZIP TFs identified in different plant and crop species varies from 78 (AtbZIPs) in A. thaliana (9), 89 (OsbZIPs) in O. sativa subs. japonica (8), 125 (ZmbZIPs) in Zea mays (12), 131 (GmbZIPs) in Glycine max (13), 92 (SbbZIPs) in Sorghum bicolor (14), 55 (VvbZIPs) in Vitis vinifera (15), 64 (CsbZIPs) in Cucumis sativus (16) and 247 (BnbZIPs) in Brassica napus (17). The bZIP transcription factors play crucial roles in developmental processes and environmental tolerance in response to multiple stresses. They are involved in the regulation of the seed development (Izawa et al., 1994; Toh et al., 2012), cell elongation (20,21), vascular development (21), flower development (22–25), somatic embryogenesis (26), as well as in nitrogen/carbon and energy metabolism (27–29).
In addition to functions in plant growth and development, bZIPs also play an important role in responses to abiotic and biotic stresses. Several bZIPs from A. thaliana (AtbZIP17, AtbZIP24, AtbZIP12), rice (OsbZIP12, OsbZIP72, OsABF1), and soybean (GmbZIP44, GmbZIP62, GmbZIP78) were found to positively regulate salt stress adaptation in plants either directly or indirectly (Liao et al., 2008; Liu et al., 2008; Lu et al., 2009; Yang et al., 2009; Amir Hossain et al., 2010; Hossain et al., 2010; Ji et al., 2013). Several bZIPs from rice (OsbZIP52/RISBZ5, OsbZIP16, OsbZIP23, OsbZIP45, AREB1, AREB2, ABF3) were also found to be involved in the drought tolerance (Yoshida et al., 2010; Chen et al., 2012; Liu et al., 2012; Park et al., 2015). OsbZIP52/RISBZ5 negatively regulates cold stress responses (Liu et al., 2012) while OsbZIP72 was a positive regulator of ABA responses (34). Similarly, overexpression of GmbZIP44, GmbZIP62, and GmbZIP78 reduced ABA sensitivity (13). Interestingly, group D or so-called TGA bZIPs plant a role in systemic acquired resistance (SAR) and pathogen resistance (Fu et al., 2013; Gatz, 2013). However, there is little published information about the bZIP transcription factor family in cultivated alfalfa and its role in stress resistance.
With the availability of a chromosome-level genome assembly in alfalfa (42), we conducted a genome-wide search to identify and characterize alfalfa bZIP transcription factors. Since bZIP transcription factors were identified to play significant roles in the regulation of the abiotic stress tolerance (11,12), we speculated various bZIP transcription factors would be differentially expressed throughout distinct developmental stages and in response to abiotic stresses in alfalfa as well. The present study identifies several bZIPs from a protein database in tetraploid alfalfa (M. sativa). We also analyzed differential gene expression from transcriptomics during ABA, drought, salt, and cold stress conditions and verified a subset of differentially expressed bZIPs via qRT-PCR. This study will facilitate functional analysis of the bZIP transcription factor family in alfalfa. The identification of functions of alfalfa bZIP transcription factors during abiotic stress conditions will further help breeding efforts for improved stress tolerance.
2. Materials and Methods
Identification of bZIP transcription factor gene family in alfalfa
For comprehensive identification and analysis of the bZIP transcription factor (TF) gene family in alfalfa, the sequences of bZIP transcription factors from model and known species were downloaded from the Plant Transcription factor database (http://planttfdb.cbi.pku.edu.cn/), which included 127 sequences from Arabidopsis thaliana, 93 from Lotus japonicus, 124 from Medicago truncatula and 140 from Oryza sativa. The number of bZIPs used were more than that is mentioned in Table 1 as it included spliced variants as well. A local protein database was created using Basic Local Alignment Search Tool (58) with protein sequences from chromosome level assembly of alfalfa (42). A BLASTp search was conducted in the local database created using the protein sequences from alfalfa, taking the bZIP sequences from model organisms as a query with an E-value cut-off of 1E-05 (0.00001). The bZIP sequences obtained from the search were further confirmed based on the presence of the bZIP domain (N-x(7)-R/K-x(9)-L-x(6)-L-x(6)-L) using the Pfam web program (https://pfam.xfam.org/) with an E-value of 1.0. Further, the bZIP domain was used to search against the database of the identified bZIP sequences using the Prosite program of the ExPASy bioinformatics resource (http://protsite.expasy.org). The identified sequences with intact bZIP domains were predicted to be bonafide bZIP sequences.
Evolutionary analysis, protein properties and detection of conserved motifs in the bZIPs
To analyze the sequence features of bZIP transcription factors, multiple sequence alignment of 237 bZIP proteins were performed using multiple sequence comparison by log-expectation (MUSCLE) (59) command using default parameters. The output of the multiple sequence alignment was visualized using Unipro UGENE v.33. (60) For evolutionary analysis, 549 sequences were used which included sequences from M. sativa (237), A. thaliana (78), L. japonicus (70), M. truncatula (75) and O. sativa (89). Multiple sequence alignment was carried out by CLUSTALW with default parameters. Subsequently, the phylogenetic tree was constructed by the Neighbor Joining method using 1000 bootstraps replicates. Phylogenetic analyses were conducted using MEGA version X (61).
Other physical properties of the identified sequences like the molecular weight and theoretical isoelectric point (pI) were determined using Compute pI/Mw tools (http://web.expasy.org/compute_pi/) of ExPASy bioinformatics resource. The MEME (45) program was used to identify the conserved motifs within the full-length Alfalfa. The parameters used were maximum number of motifs to be 25, distribution of motifs = any number of repetitions, optimum motif width = 6 to 50 residues.
In silico functional analysis of bZIP genes
For predicting the MsbZIP protein function (gene ontology) GO annotation was performed using the web-accessible Blast2GO v4.1 annotation system (https://www.blast2go.com/) (62). Briefly, the MsbZIP protein sequences were used to search for similar sequences against the NCBI non-redundant (Nr) database using the Blast tool in the Blast2GO software, with an E-value of 10−3 (1e-03). Next, mapping and annotation were performed on Blast2GO using default parameters. Finally, functional classification was also performed by Blast2GO.
Expression analysis during plant development
The raw RNA sequence data was downloaded from the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA), SRP055547 (46). The data was generated from six tissues at different growth stages of Alfalfa namely, root, nodule, elonged stem, pre-elonged stem, leaf, and flower. The tissue sample for RNA-Seq was collected at the respective stage of alfalfa plants. Fastqc version 0.11.7 was used for quality check of the raw sequences. The reads passing the minimum Phred quality score of 30 were selected. The RNA-Seq analysis was carried out following the method described by (63), in which the filtered reads were aligned with the reference genome using HISAT2 version 2.1.0 (64) and sorted by Samtools ver 1.9. Transcript assembly and quantification was carried out using Stringtie version 2.1.1 (65). A python script was used to extract read count information directly from the files generated from Stringtie and edgeR package (66) in R was used for differential gene expression analysis. TBtools version 1.0692 (67) was used to generate heatmaps for the differentially expressed genes.
Transcriptome analysis of bZIP genes in response to abiotic stresses
The raw RNA sequence data from previous studies were downloaded from the National Center for Biotechnological Information (NCBI) Sequence Read Archive (SRA). The transcriptome data consist of cold treatment (SRR7091780-SRR7091794, (68,69), and ABA, drought, and salt treatments (SRR7160313-SRR7160357, (68,70). All these samples were collected from 12 days old alfalfa seedlings for RNA-Seq. Fastqc version 0.11.7 was used for quality check of the raw sequences. The reads passing the minimum Phred quality score of 40 were selected. The RNA-Seq analysis was carried out following the method described by (63), in which the filtered reads were aligned with the reference genome using HISAT2 ver2.1.0 and sorted by Samtools ver1.9. Transcript assembly and quantification was carried out using Stringtie version 2.1.1. A python script was used to extract read count information directly from the files generated from Stringtie and edgeR package in R was used for differential gene expression analysis. TBtools version 1.0692 was used to generate heatmaps for the differentially expressed genes.
Analysis of cis-regulatory elements
For this analysis, the bZIP genes with changed expression during abiotic stress were visualized using Integrated Genome Browser 9.1.4 (71) to locate the promoter sequences. Samtools (ver. 1.9) was used to extract the 2000 base pair sequence from the promoter of these changed bZIP genes to investigate the potential cis-regulatory elements by querying them through the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/). In total six cis-regulatory elements responsive to stress were analyzed. These elements included abscisic acid responsive (ABRE), methyl jasmonate responsive (CGTCA-motif), light inducible G-box motif, low-temperature responsive (LTR), drought responsive (MBS binding site) and defense and stress responsive (TC-rich repeats).
Plant materials and RNA isolation:
An alfalfa (Medicago sativa) variety Vernal was collected from Washington State University, Pullman, WA. Seeds were sterilized using 70% ethanol inside the laminar hood and kept in 4 °C dark for 3 days. After that, seeds were transplanted in half strength Murashige and Skoog’s medium (PhtyoTech Labs, KS) containing 3% sucrose and 0.7% agar inside a growth chamber with light cycle of 16:8-h (light/dark) and temperature of 22 °C. We simulated abiotic stress with abscisic acid (ABA, 10 μM), NaCl (250 mM), mannitol (300 mM) and 4 °C cold stress treatment at 2 weeks after germination (WAG). Leaf tissue samples were collected at the following time points: 0 (control, CT), 1, 3, and 24 h for selected MsbZIP gene expression analysis. For selected MsbZIP H-group gene expression analysis, we collected hypocotyl samples at 5 days after germination (DAG) and 2 WAG . The collected samples were quickly placed in liquid nitrogen and stored in a − 80 °C freezer for subsequent RNA extraction.
Total RNA was extracted using the SpectrumTM plant total RNA isolation kit (Sigma Aldrich, USA) and treated with DNaseI to eliminate genomic DNA contamination. Total RNA yield (ng/µL) and purity (260:280 wavelength ratios) was measured by using Nanodrop (Eppendorf, USA) instrument. 2 µg RNA from each of the samples were used for the synthesis of single stranded cDNA. We used Bio-Rad iScript™ (Hercules, CA, USA) for cDNA synthesis.
Reverse Transcription Quantitative Real-Time PCR analysis:
Reverse Transcription Quantitative real-time PCR was performed with volumes of 10 µL per well with Bio-Rad™ SYBR green Supermix (Hercules, CA, USA). The amount of cDNA was normalized to the level of Medicago sativa housekeeping gene elongation factor 1α (MsElf1α) used as an internal control. The amplification was conducted in Biorad 96 Real-Time PCR System. A standard thermal profile for SYBR green mix was as followed: cDNA synthesis at 37 °C for 15 min and enzyme inactivation at 85 °C for 5 s. qPCR conditions were: initial denaturation 96 °C for 30s, denaturation 96 °C for 5 s, annealing and extension 62 °C for 30s. Transcripts expression levels were calculated with the 2−ΔΔCt method, as previously mentioned in (72). Three biological and three technical replicates were used for gene expression analysis. Primers used for this analysis are mentioned in Supplementary Table 3.
RT-qPCR Data Analysis
One-way ANOVA followed by multiple comparisons test was performed (73)using GraphPad Prism version 8.0 Statistics for all selected MsbZIP candidate gene expression analysis used in RT-qPCR validation.
3. Results
Identification of the alfalfa bZIP gene family
We identified 237 bZIP sequences with the intact bZIP domain in alfalfa (M. sativa). These sequences were named MsbZIP1 to MsbZIP237 based on the order identified in the protein sequence database (42). We compared the genome size and number of bZIPs in different models and crop species (Table 1). The comparison shows that alfalfa has the highest number of bZIP sequences. Since the diploid model legume M. truncatula with a genome size of 390 Mega Base (Mb) has 75 bZIP sequences, tetraploid alfalfa is expected to have double the number of bZIP sequences. Not surprisingly, the number of bZIP TFs identified in alfalfa was 237, which likely represents the complete number of bZIP for tetraploid alfalfa
Chromosomal distribution of bZIP genes
Among the identified 237 bZIP genes, 233 were annotated among 32 chromosomes and the remaining 4 genes were annotated in 6 different Contigs (Contig 37287, Contig 43349, Contig 51828, Contig 57601, Contig 57602, Contig 57603). The largest gene was in Group A with a length of 688 amino acids while the smallest genes were in Group S with 119 amino acids in length (Supplementary Table 2). The distribution of genes on all the 32 chromosomes was also different (Supplementary Figure 2). In most of the chromosomes, they were distributed throughout, while, in a few of the chromosomes, these genes are concentrated towards telomeric regions of the chromosomes. The chromosomal distribution of bZIP genes and their chromosome related information is provided with supplementary information.
Phylogenetic Analysis and Multiple Sequence Alignment
The identified 237 bZIP proteins were divided into 10 groups (A, C, D, E, F, G, H, I, M, and S) based on the topology of the tree developed in Arabidopsis thaliana (9,11) and were used to generate a phylogenetic tree along with protein sequences from A. thaliana, L. japonicus, M. truncatula and O. sativa (Figure 1). Alfalfa bZIP proteins fell into 10 different groups and numbers ranged from four (H) to forty-three (A); however, no members were identified for groups B, J and K. To identify common conserved domains amongst the sequences, we carried out multiple sequence alignment. The alignment of 237 bZIP protein sequences showed the presence of intact and highly conserved bZIP domains (N-x(7)-R/K-x(9)-L-x(6)-L-x(6)-L) (Figure 2, Supplementary Figure 1). The domain is divided into the basic region with ~18 amino acids residues containing nuclear localization signal followed by an intact N-x(7)-R/K motif while the leucine zipper region contains heptad repeats of leucines or other bulky hydrophobic amino acids with nine amino acids towards the C-terminus (11). The presence of the intact bZIP domain further validates the identified sequences as bZIP proteins.
Conserved protein domain analysis
Identification of conserved protein motifs helps to elucidate protein functions and bZIPs usually possess additional conserved motifs that could provide sites for activation (44). Using the “MEME” (Multiple Em for Motif Elicitation) program (45), 25 conserved motifs were identified in the 237 bZIPs (Supplementary Table 2). Among the identified motifs, the basic region of the bZIP, containing an invariant motif (N-x7-R/K-x9) with 18 amino acid residues was found (Figure 3A), while the leucine zipper region that contains the heptad repeat of leucine or other bulky hydrophobic amino acids was also identified (Figure 3B). The basic region facilitates sequence specific DNA binding whereas the leucine zipper region is important for dimerization specificity. However, the function of the 23 motifs that were also identified in the bZIP sequences are unknown and require further study.
In silico functional classification of MsbZIP transcription factors
Among the 237 MsbZIP, 21 GO (Gene Ontology) categories were assigned to 203 of the MsbZIPs identified (Figure 4). The major molecular functions of these bZIPs were DNA-binding transcription factor activity, which is consistent with their demonstrated role as transcription factors in other species. In the biological process category, most of bZIPs were assigned to the regulation of transcription category and almost all these proteins were predicted to localize to the nucleus in the cellular component category. Transcription factors provide binding sites through which they can regulate gene expression. They may act as either positive or negative regulators of downstream genes depending upon the environmental condition. The current functional classification (GO terms) of these bZIP proteins further supports their regulatory nature.
Tissue-specific expression profile analysis of alfalfa bZIPs
After analysis of publicly available RNA-Seq data (46), we found differential expression of 177 bZIP genes. These genes were selected for having expression values in at least one of the tissues: stem, flowers, leaves, root nodules, roots, and pre-elongated stems (PES). They were then displayed in a heatmap to visualize the expression profile in different tissues and organs (Figure 5). Differential gene expression was observed for different developmental stages. Most of the genes were highly expressed in nodules and roots. Apart from nodules and roots, genes that were upregulated in one developmental stage were downregulated in other developmental stages which can be observed in the heatmap. Even within a group, the genes were differentially expressed across all developmental stages suggesting different bZIP genes are required for growth and development at different stages.
Alfalfa bZIP genes are differentially expressed in response to abiotic stresses
Analysis from the publicly available RNA-seq datasets showed differential expression of 146 genes during ABA, drought, and salt stress as well as 152 bZIP proteins during cold stress at 0, 2, 6, 24, and 48h, respectively. The expression pattern of MsbZIP genes during different abiotic stress conditions of cold, ABA, drought and salt showed differential expression. Across different time points of abiotic stress, the expression was different for different genes and even within a group the genes were expressed differently for different abiotic stress. Among 4 different time points of cold treatment (2h, 6h, 24h and 48 h), different genes were upregulated at different time points (Supplementary Figure 3). Even within a group, at different time points, different genes were upregulated and downregulated at different intervals of cold treatment. Like the cold treatment, abiotic stress of ABA, drought and salt treatment also showed multiple genes upregulated at different time points of stress treatment (Figure 6). However, no genes were actively expressed during different time points of the same treatment condition among ABA, drought, and salt, which indicates different transcription factors are active during different abiotic stress as well as different time points of stress.
RT-qPCR validation of gene expression analysis
For RNA-Seq result verification, five differentially expressed genes, two from group A (MsbZIP80 and MsbZIP88) and three from group S (MsbZIP31, MsbZIP 109 and MsbZIP117) were selected for RT-qPCR analysis. The expression pattern for most of the genes were consistent with the RNA-Seq analysis (Figure 7). In addition, the genes also showed high group specificity, as genes from Group A (MsbZIP80 and MsbZIP88) and Group S (MsbZIP31, MsbZIP 109 and MsbZIP117) were consistent in expression to their specific groups. Significant upregulation of all five genes were found at the 1-hour timepoint for salt stress. MsbZIP31 and MsbZIP117 expression then decreased at the 3 and 24-hour timepoints compared to the 1-hour timepoint, whereas the genes from the A- MsbZIP80 and MsbZIP88 (A Group) were the most upregulated at the 3-hour timepoint.
Similarly, for drought treatments, all the genes except MsbZIP117, were highly upregulated upon 1-hour of exposure of mannitol and remained upregulated relative to the control. However, MsbZIP117 was downregulated at 3-hours and then returned to normal expression levels at 24 hours. Similar to drought treatment and as expected, the same trend of upregulation and downregulation for all five genes continued during their treatment of ABA across the timepoints.
For the cold treatments, MsbZIP80 and MsbZIP109 were continuously upregulated when exposed to 4 °C at all the timepoints (from 1-h to 24-h). For the remaining three genes (MsbZIP31, MsbZIP88 and MsbZIP117), they followed completely different trend across the samples and treatments. While MsbZIP31 was significantly upregulated at 1-hour of exposure of cold, MsbZIP117 was downregulated and MsbZIP88 did not change. However, MsbZIP88 was significantly upregulated from 1-hour to 3-hour treatment while the two genes downregulated at the same treatment. MsbZIP31 and MsbZIP117 were then upregulated from 3-hour to 24-hour of treatment. MsbZIP88 showed slight but significant upregulation in response to cold at the 3-hour timepoint. The level of significance during upregulation and downregulation of genes is presented in Figure 7.
To validate the tissue specific expression of bZIPs, genes from Group H (MsbZIP79 and MsbZIP222) were selected. In comparison to the 5-day old hypocotyl, 2-week-old hypocotyl of both MsbZIP79 and MsbZIP222 were significantly downregulated (p < 0.01) (Figure 8). However, for both the genes, they were upregulated in leaf tissue in comparison to the hypocotyl tissues extracted from 2-week-old seedlings. For MsbZIP79, the upregulation was highly significant (p < 0.001); however, it was not the same for MsbZIP222. MsbZIP79 was significantly upregulated in 2-week-old leaves compared to 2-week-old hypocotyls. The expression pattern of these genes was also in consistent with the RNA-Seq analysis.
Cis-regulatory elements in bZIP gene promoter
The expression pattern of stress responsive-genes are often controlled by cis-regulatory elements. These elements are typically located 5’ upstream of the gene coding sequences. These elements provide a binding site for the transcription factors to switch on or off the gene based on the environmental condition. In this study, we analyzed 135 stress-responsive bZIP promoters, we identified 875 cis-regulatory elements distributed along these 135 bZIP promoter. The detailed distribution of these cis-elements along the bZIP promoters was performed (Supplementary Figure 4). We focused on cis-elements implicated in abiotic stress responses and found an abundance of the following cis-regulatory elements: abscisic acid responsive element (ABRE), methyl jasmonate responsive motif (CGTCA-motif), light inducible G-box motif, low-temperature responsive (LTR), drought responsive (MBS binding site) and defense and stress responsive (TC-rich repeats).Among the 875 cis-elements, light inducing G-box motif was the highest with 274 followed by abscisic stress responsive element (ABRE) with 234 while low temperature responsive (LTR) with 50 was the lowest.
4. Discussion
In the present study, we identified 237 bZIP sequences from tetraploid alfalfa that contained both a highly conserved basic region and the heptad repeat leucine zipper region, suggesting they are functional bZIPs. As predicted, the number of bZIP in tetraploid alfalfa (237) is more than double to that of diploid model legume M. truncatula (75). Not surprisingly, the number of bZIP genes varied amongst plant species with A. thaliana (78), L. japonicus (70), M. truncatula (75) and O. sativa (89) (7,11,43,47,48). Similarly, the allotetraploid B. napus genome contained 247 bZIP genes, which is roughly double that of the number found in the related diploid A. thaliana.
Based on phylogenetic analysis and previous analyses from A. thaliana, M. truncatula, L. japonicus and O. sativa, we classified the alfalfa bZIP genes into 10 groups (A, C, D, E, F, G, H, I, M, and S). The most recent classification of bZIPs from A. thaliana (9) sorted AtbZIPs into 13 groups. Notably, groups B, J and K are missing in our analysis of alfalfa. In A.thaliana there are three members of group B (bZIP17, bZIP28, and bZIP49) and one group K member (bZIP60), which are implicated in endoplasmic reticulum stress responses (49), but both these groups are missing in alfalfa which begs the question of which groups perform this function in alfalfa. Group J in A. thaliana is made up of a single copy gene, bZIP62, which is related to Group G bZIP GBF1– a negative regulator of blue-light responsive hypocotyl growth that acts antagonistically to HY5 and HYH, two group H bZIPs important in photomorphogenic growth (50). Another remarkable difference between groups is the group M bZIP72, which is single copy in A. thaliana but contains 13 members in alfalfa. It will be interesting to determine the role M group bZIPs play in alfalfa and it is intriguing to postulate why this group has increased in number.
It is well established that bZIP transcription factors have a myriad of roles in plant development such as seed maturation and germination (19), floral induction and development (22,25). Not surprisingly, tissue-specific expression of 177 bZIP genes in nodules, flowers, roots, leaves, and stems was found in alfalfa as well (Figure 5). Interestingly, group E members were most specifically expressed in stems, roots, and flowers, whereas several group F members were expressed in pre-elongated stems. In A. thaliana the group E member bZIP34 has been linked to pollen germination and pollen tube growth (24). In contrast, group F members regulate zinc (Zn) transporters and salt stress responses (35,51). Group C and S bZIPs are known to heterodimerize in the so-called C/S1 bZIP network involved in nutrient and energy metabolism (29,52). Likewise, group C and S bZIPs are co-expressed in some tissues such as roots and nodules in alfalfa.
In addition to regulating development, bZIPs play a wide array of roles in biotic and abiotic stress responses (11,12) in different crop species. (53) identified the OsABI5 bZIP TF that was involved in rice fertility and stress tolerance. (Nijhawan et al., (2008) related bZIP genes in rice to drought tolerance through genomic survey and gene expression analysis. Similarly, a root-specific bZIP transcription factor was isolated in tepary beans and found to be responsive to water-stress conditions (Rodriguez-Uribe et al., 2006). (13) isolated three bZIP genes (GmbZIP44, GmbZIP62, GmbZIP78) and found a negative regulator of ABA and tolerance to salt and freezing stress by overexpression in A. thaliana. As several studies have shown the role of bZIP transcription factors in the response to plant stress, (Liu et al., 2012) further added to it by cloning a bZIP gene and measuring physiological changes mediated by it in alfalfa under different stress conditions. Additionally, the over-expressed cloned Alfalfa bZIP genes in tobacco plants resulted in transgenic tobacco plants conveying salt and drought tolerance. These results indicate that the over-expression of certain bZIP genes increases the tolerance of plants to different abiotic stresses.
Furthermore, RT-qPCR analysis was carried out to corroborate the expression trends from RNA-Seq analyses. The genes selected for abiotic stress were from Group A (MsbZIP80,MsbZIP88) and Group S (MsbZIP31, MsbZIP109,MsbZIP117), while Group H (MsbZIP79,MsbZIP222) genes were selected for expression during developmental stages. In A. thaliana, Group A genes encode abscisic acid-responsive element binding factors (ABF1) that act at the core of the ABA signaling pathway (55). During water deficit conditions like cold, salt and drought, these factors are induced for the adaptive response to overcome water deficit conditions (55). Similarly, expression analysis of Medicago truncatula revealed bZIP genes that were responsive to drought and salt stress conditions were concentrated in Group A and S (48). Furthermore, bZIPs from these groups were found to be involved in sugar signaling process (56), resulting in physiological and developmental changes, which integrates with other signaling pathways in plants for stress response (56). Similar to these studies, we also found the MsbZIPs from Group A and Group S were highly induced with significant expression during differential treatment of salt, cold, drought and ABA.
In A. thaliana, group H bZIPs contain elongated hypocotyl (HY5) and the HY5 homologue (HYH), which have been found to play important roles in developmental process (9). HY5 regulates developmental process through cell elongation, cell proliferation, chloroplast development, pigment accumulation and nutrient assimilation (57). These genes inhibit hypocotyl elongation in light and promote plant growth by inducing nutrient uptake and through expression of enzymes associated with nitrogen, sulfur and copper required for overall growth (50). The findings of the current study revealed that the bZIPs in Group H (MSbZIP79,MsbZIP222) are significantly downregulated in 2 weeks old hypocotyl tissue in comparison to 5 day-hypocotyl tissue. However, these genes were more highly expressed in 2-week-old leaf samples, which further establishes their role in the developmental processes in leaves as has been proposed in A. thaliana.
5. Conclusion
Here we report the first comprehensive in silico analysis of the bZIP transcription factor family in alfalfa (M. sativa). We identified 237 bZIP genes and named them MsbZIP1 to MsbZIP237. Phylogenetic analysis of these bZIP genes using A. thaliana as a reference divided the sequences into 10 groups, with B, J, and K missing in alfalfa. The physicochemical analysis and motif analysis showed high specificity within each group. The expression profile of bZIPs suggests bZIPs are expressed in a tissue-specific manner. Finally, the expression profiles of bZIP genes during different abiotic stress conditions (cold, ABA, drought, and salt) showed the specific response of a few bZIP at specific time points during the stress response making them good candidates for stress-responsive transcription factors and further functional characterization. Taken together, this work provides a framework for the future study of bZIPs in alfalfa and presents candidate bZIPs involved in stress-response signaling.
References
- USDA. Crop Production Historical Track Records. Natl Agric Stat Serv. 2018;(April).
- Carelli, M.; Gnocchi, S.; Fancelli, S.; Mengoni, A.; Paffetti, D.; Scotti, C.; Bazzicalupo, M. Genetic Diversity and Dynamics of Sinorhizobium meliloti Populations Nodulating Different Alfalfa Cultivars in Italian Soils. Appl. Environ. Microbiol. 2000, 66, 4785–4789. [Google Scholar] [CrossRef]
- Pellock, B.J.; Cheng, H.-P.; Walker, G.C. Alfalfa Root Nodule Invasion Efficiency Is Dependent on Sinorhizobium meliloti Polysaccharides. J. Bacteriol. 2000, 182, 4310–4318. [Google Scholar] [CrossRef]
- Donnarumma, F.; Bazzicalupo, M.; Blažinkov, M.; Mengoni, A.; Sikora, S.; Babić, K.H. Biogeography of Sinorhizobium meliloti nodulating alfalfa in different Croatian regions. Res. Microbiol. 2014, 165, 508–516. [Google Scholar] [CrossRef]
- Statistics USD of ANA. United States Department of Agriculture National Agricultural Statistics Services. 2018.
- Udvardi, M.K.; Kakar, K.; Wandrey, M.; Montanari, O.; Murray, J.; Andriankaja, A.; Zhang, J.-Y.; Benedito, V.; Hofer, J.M.; Chueng, F.; et al. Legume Transcription Factors: Global Regulators of Plant Development and Response to the Environment. Plant Physiol. 2007, 144, 538–549. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, P.; Riaño-Pachón, D.M.; Corrêa, L.G.G.; Rensing, S.A.; Kersten, B.; Mueller-Roeber, B. PlnTFDB: updated content and new features of the plant transcription factor database. Nucleic Acids Res. 2009, 38, D822–D827. [Google Scholar] [CrossRef] [PubMed]
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic Survey and Gene Expression Analysis of the Basic Leucine Zipper Transcription Factor Family in Rice. Plant Physiol. 2007, 146, 323–324. [Google Scholar] [CrossRef]
- Dröge-Laser, W.; Snoek, B.L.; Snel, B.; Weiste, C. The Arabidopsis bZIP transcription factor family — an update. Curr. Opin. Plant Biol. 2018, 45, 36–49. [Google Scholar] [CrossRef]
- Hurst, HC. Transcription factors. 1: bZIP proteins. Protein Profile [Internet]. 1994;1(2):123–68. Available from: http://www.ncbi.nlm.nih. 8528. [Google Scholar]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Chen, J.; Wang, Y.; Chen, Y.; Chen, S.; Lin, Y.; Pan, S.; Zhong, X.; Xie, D. Genome-Wide Analysis of bZIP-Encoding Genes in Maize. DNA Res. 2012, 19, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Zou, H.-F.; Wei, W.; Hao, Y.-J.; Tian, A.-G.; Huang, J.; Liu, Y.-F.; Zhang, J.-S.; Chen, S.-Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, J.; Zhang, B.; Vanitha, J.; Ramachandran, S.; Jiang, S. Genome-wide Expansion and Expression Divergence of the Basic Leucine Zipper Transcription Factors in Higher Plants with an Emphasis on SorghumF. J. Integr. Plant Biol. 2011, 53, 212–231. [Google Scholar] [CrossRef]
- Liu, J.; Chen, N.; Chen, F.; Cai, B.; Santo, S.D.; Tornielli, G.B.; Pezzotti, M.; Cheng, Z.-M.M. Genome-wide analysis and expression profile of the bZIP transcription factor gene family in grapevine (Vitis vinifera). BMC Genom. 2014, 15, 281–281. [Google Scholar] [CrossRef] [PubMed]
- Baloglu, M.C.; Eldem, V.; Hajyzadeh, M.; Unver, T. Genome-Wide Analysis of the bZIP Transcription Factors in Cucumber. PLOS ONE 2014, 9, e96014. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, D.; Jia, L.; Huang, X.; Ma, G.; Wang, S.; Zhu, M.; Zhang, A.; Guan, M.; Lu, K.; et al. Genome-Wide Identification and Structural Analysis of bZIP Transcription Factor Genes in Brassica napus. Genes 2017, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Izawa T, Foster R, Nakajima M, Shimamoto K, Chua NH. The rice bZIP transcriptional activator RITA-1 is highly expressed during seed development 1994 Sep;6(9):1277–87. Available from: https://academic.oup. Plant Cell 1994, 6, 1277–1287. [CrossRef]
- Toh, S.; McCourt, P.; Tsuchiya, Y. HY5is involved in strigolactone-dependent seed germination in Arabidopsis. Plant Signal. Behav. 2012, 7, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa J, Sakai T, Ishida S, Yamaguchi I, Kamiya Y, Takahashi Y. REPRESSION OF SHOOT GROWTH, a bZIP Transcriptional Activator, Regulates Cell Elongation by Controlling the Level of Gibberellins. Available from: https://academic.oup. Plant Cell, 2000; 12, 901–915. [CrossRef]
- Yin, Y.; Zhu, Q.; Dai, S.; Lamb, C.; Beachy, R.N. RF2a, a bZIP transcriptional activator of the phloem-specific rice tungro bacilliform virus promoter, functions in vascular development. EMBO J. 1997, 16, 5247–5259. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Kobayashi, Y.; Yamamoto, S.; Daimon, Y.; Yamaguchi, A.; Ikeda, Y.; Ichinoki, H.; Notaguchi, M.; Goto, K.; Araki, T. FD, a bZIP Protein Mediating Signals from the Floral Pathway Integrator FT at the Shoot Apex. Science 2005, 309, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.-F.; Running, M.P.; Williams, R.W.; Meyerowitz, E.M. The PERIANTHIA gene encodes a bZIP protein involved in the determination of floral organ number in Arabidopsis thaliana. Genes Dev. 1999, 13, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Gibalová, A.; Reňák, D.; Matczuk, K.; Dupl’áková, N.; Cháb, D.; Twell, D.; Honys, D. AtbZIP34 is required for Arabidopsis pollen wall patterning and the control of several metabolic pathways in developing pollen. Plant Mol. Biol. 2009, 70, 581–601. [Google Scholar] [CrossRef]
- Iven, T.; Strathmann, A.; Böttner, S.; Zwafink, T.; Heinekamp, T.; Guivarc’h, A.; Roitsch, T.; Dröge-Laser, W. Homo- and heterodimers of tobacco bZIP proteins counteract as positive or negative regulators of transcription during pollen development. Plant J. 2010, 63. [Google Scholar] [CrossRef]
- Guan, Y.; Ren, H.; Xie, H.; Ma, Z.; Chen, F. Identification and characterization of bZIP-type transcription factors involved in carrot (Daucus carota L.) somatic embryogenesis. Plant J. 2009, 60, 207–217. [Google Scholar] [CrossRef]
- Baena-González, E.; Rolland, F.; Thevelein, J.M.; Sheen, J. A central integrator of transcription networks in plant stress and energy signalling. Nature 2007, 448, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Ciceri, P.; Locatelli, F.; Genga, A.; Viotti, A.; Schmidt, R.J. The Activity of the Maize Opaque2 Transcriptional Activator Is Regulated Diurnally. Plant Physiol. 1999, 121, 1321–1327. [Google Scholar] [CrossRef]
- Ehlert, A.; Weltmeier, F.; Wang, X.; Mayer, C.S.; Smeekens, S.; Vicente-Carbajosa, J.; Dröge-Laser, W. Two-hybrid protein–protein interaction analysis in Arabidopsis protoplasts: establishment of a heterodimerization map of group C and group S bZIP transcription factors. Plant J. 2006, 46, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Cho, J.-I.; Han, M.; Ahn, C.-H.; Jeon, J.-S.; An, G.; Park, P.B. The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. J. Plant Physiol. 2010, 167, 1512–1520. [Google Scholar] [CrossRef]
- Hossain, A.; Lee, Y.; Cho, J.-I.; Ahn, C.-H.; Lee, S.-K.; Jeon, J.-S.; Kang, H.; Lee, C.-H.; An, G.; Park, P.B. The bZIP transcription factor OsABF1 is an ABA responsive element binding factor that enhances abiotic stress signaling in rice. Plant Mol. Biol. 2009, 72, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Liu, G.; Liu, Y.; Zheng, L.; Nie, X.; Wang, Y. The bZIP protein from Tamarix hispida, ThbZIP1, is ACGT elements binding factor that enhances abiotic stress signaling in transgenic Arabidopsis. BMC Plant Biol. 2013, 13, 151–151. [Google Scholar] [CrossRef]
- Liu, J.; Srivastava, R.; Howell, S.H. Stress-induced expression of an activated form of AtbZIP17 provides protection from salt stress in Arabidopsis. Plant, Cell Environ. 2008, 31, 1735–1743. [Google Scholar] [CrossRef]
- Lu, G.; Gao, C.; Zheng, X.; Han, B. Identification of OsbZIP72 as a positive regulator of ABA response and drought tolerance in rice. Planta 2008, 229, 605–615. [Google Scholar] [CrossRef]
- Yang, O.; Popova, O.V.; Süthoff, U.; Lüking, I.; Dietz, K.-J.; Golldack, D. The Arabidopsis basic leucine zipper transcription factor AtbZIP24 regulates complex transcriptional networks involved in abiotic stress resistance. Gene 2009, 436, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, W.; Zhou, J.; He, H.; Chen, L.; Chen, H.; Deng, X.W. Basic leucine zipper transcription factor OsbZIP16 positively regulates drought resistance in rice. Plant Sci. 2012, 193-194, 8–17. [Google Scholar] [CrossRef]
- Liu, C.; Wu, Y.; Wang, X. bZIP transcription factor OsbZIP52/RISBZ5: a potential negative regulator of cold and drought stress response in rice. Planta 2011, 235, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Jeong, J.S.; Lee, K.H.; Kim, Y.S.; Choi, Y.D.; Kim, J.-K. OsbZIP23 and OsbZIP45, members of the rice basic leucine zipper transcription factor family, are involved in drought tolerance. Plant Biotechnol. Rep. 2015, 9, 89–96. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Gatz, C. From Pioneers to Team Players: TGA Transcription Factors Provide a Molecular Link Between Different Stress Pathways. Mol. Plant-Microbe Interactions® 2013, 26, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zeng, Y.; Yang, Y.; Huang, L.; Tang, B.; Zhang, H.; Hao, F.; Liu, W.; Li, Y.; Liu, Y.; et al. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, W.; Qi, X.; Liu, Z.; Xie, W.; Wang, Y. Genome-wide identification, expression profiling, and SSR marker development of the bZIP transcription factor family in Medicago truncatula. Biochem. Syst. Ecol. 2015, 61, 218–228. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Li, H.; Yang, Y.; Guang, Y.; Zhou, Y. The bZIP gene family in watermelon: genome-wide identification and expression analysis under cold stress and root-knot nematode infection. PeerJ 2019, 7, e7878. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, w202–w208. [Google Scholar] [CrossRef]
- O’rourke, J.A.; Fu, F.; Bucciarelli, B.; Yang, S.S.; Samac, D.A.; Lamb, J.F.S.; Monteros, M.J.; Graham, M.A.; Gronwald, J.W.; Krom, N.; et al. The Medicago sativa gene index 1.2: a web-accessible gene expression atlas for investigating expression differences between Medicago sativa subspecies. BMC Genom. 2015, 16, 1–17. [Google Scholar] [CrossRef]
- E ZG, Zhang YP, Zhou JH, Wang L. Mini Review Roles of the bZIP gene family in rice. Genet. Mol. Res. 2014, 13, 3025–3036. [CrossRef]
- Wang, Z.; Cheng, K.; Wan, L.; Yan, L.; Jiang, H.; Liu, S.; Lei, Y.; Liao, B. Genome-wide analysis of the basic leucine zipper (bZIP) transcription factor gene family in six legume genomes. BMC Genom. 2015, 16, 1053. [Google Scholar] [CrossRef]
- Howell, S.H. Endoplasmic Reticulum Stress Responses in Plants. Annu. Rev. Plant Biol. 2013, 64, 477–499. [Google Scholar] [CrossRef]
- Gangappa, S.N.; Srivastava, A.K.; Maurya, J.P.; Ram, H.; Chattopadhyay, S. Z-Box Binding Transcription Factors (ZBFs): A New Class of Transcription Factors in Arabidopsis Seedling Development. Mol. Plant 2013, 6, 1758–1768. [Google Scholar] [CrossRef]
- Assunção, A.G.L.; Herrero, E.; Lin, Y.-F.; Huettel, B.; Talukdar, S.; Smaczniak, C.; Immink, R.G.H.; Van Eldik, M.; Fiers, M.; Schat, H.; et al. Arabidopsis thaliana transcription factors bZIP19 and bZIP23 regulate the adaptation to zinc deficiency. Proc. Natl. Acad. Sci. USA 2010, 107, 10296–10301. [Google Scholar] [CrossRef]
- Weltmeier, F.; Ehlert, A.; Mayer, C.S.; Dietrich, K.; Wang, X.; Schütze, K.; Alonso, R.; Harter, K.; Vicente-Carbajosa, J.; Dröge-Laser, W. Combinatorial control of Arabidopsis proline dehydrogenase transcription by specific heterodimerisation of bZIP transcription factors. EMBO J. 2006, 25, 3133–3143. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.; Guan, Y.; Ren, H.; Zhang, F.; Chen, F. A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol. Biol. 2008, 66, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Uribe, L.; O'Connell, M.A. A root-specific bZIP transcription factor is responsive to water deficit stress in tepary bean (Phaseolus acutifolius) and common bean (P. vulgaris). J. Exp. Bot. 2006, 57, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Roychoudhury, A. Abscisic-acid-dependent basic leucine zipper (bZIP) transcription factors in plant abiotic stress. Protoplasma 2015, 254, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Hanson J, Smeekens S. Sugar perception and signaling—an update. Available from: https://linkinghub.elsevier. Curr. Opin. Plant. Biol. 2009; 12, 562–567. [CrossRef]
- Jing, Y.; Zhang, D.; Wang, X.; Tang, W.; Wang, W.; Huai, J.; Xu, G.; Chen, D.; Li, Y.; Lin, R. ArabidopsisChromatin Remodeling Factor PICKLE Interacts with Transcription Factor HY5 to Regulate Hypocotyl Cell Elongation. Plant Cell 2013, 25, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; The UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S. Blast2GO: A Comprehensive Suite for Functional Analysis in Plant Genomics. Int. J. Plant Genom. 2008, 2008, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Luo, D.; Wu, Y.; Liu, J.; Zhou, Q.; Liu, W.; Wang, Y.; Yang, Q.; Wang, Z.; Liu, Z. Comparative Transcriptomic and Physiological Analyses of Medicago sativa L. Indicates that Multiple Regulatory Networks Are Activated during Continuous ABA Treatment. Int. J. Mol. Sci. 2018, 20, 47. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Luo, D.; Chai, X.; Wu, Y.; Wang, Y.; Nan, Z.; Yang, Q.; Liu, W.; Liu, Z. Multiple Regulatory Networks Are Activated during Cold Stress in Medicago sativa L. Int. J. Mol. Sci. 2018, 19, 3169. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Zhou, Q.; Wu, Y.; Chai, X.; Liu, W.; Wang, Y.; Yang, Q.; Wang, Z.; Liu, Z. Full-length transcript sequencing and comparative transcriptomic analysis to evaluate the contribution of osmotic and ionic stress components towards salinity tolerance in the roots of cultivated alfalfa (Medicago sativa L.). BMC Plant Biol. 2019, 19, 1–20. [Google Scholar] [CrossRef]
- Freese, N.H.; Norris, D.C.; Loraine, A.E. Integrated genome browser: visual analytics platform for genomics. Bioinformatics 2016, 32, 2089–2095. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Brown MB, Forsythe AB. Robust Tests for the Equality of Variances. J. Am. Stat. Assoc. 1974; 69, 364. [CrossRef]
Figure 1.
Phylogenetic analysis and group classification of bZIP proteins from alfalfa. 237 bZIP proteins and 312 proteins from A. thaliana (78), L. japonicus (70), M. truncatula (75) and O. sativa (89) were used to create a neighbor joining tree with 1000 bootstraps. The bZIPs are grouped into 11 groups (A - K, M and S) based on tree topology results from A. thaliana and M. truncatula. A detailed information of the tree including the genes from all the species mentioned above is presented in Genes expressed during stress conditions are distributed throughout the groups. Group D contained genes highly expressed during drought stress conditions and ABA, while Group I and S contained genes that are upregulated during salt stress conditions. Although genes highly expressed during cold stress conditions were distributed over different groups, most of them were from group A and S.
Figure 1.
Phylogenetic analysis and group classification of bZIP proteins from alfalfa. 237 bZIP proteins and 312 proteins from A. thaliana (78), L. japonicus (70), M. truncatula (75) and O. sativa (89) were used to create a neighbor joining tree with 1000 bootstraps. The bZIPs are grouped into 11 groups (A - K, M and S) based on tree topology results from A. thaliana and M. truncatula. A detailed information of the tree including the genes from all the species mentioned above is presented in Genes expressed during stress conditions are distributed throughout the groups. Group D contained genes highly expressed during drought stress conditions and ABA, while Group I and S contained genes that are upregulated during salt stress conditions. Although genes highly expressed during cold stress conditions were distributed over different groups, most of them were from group A and S.
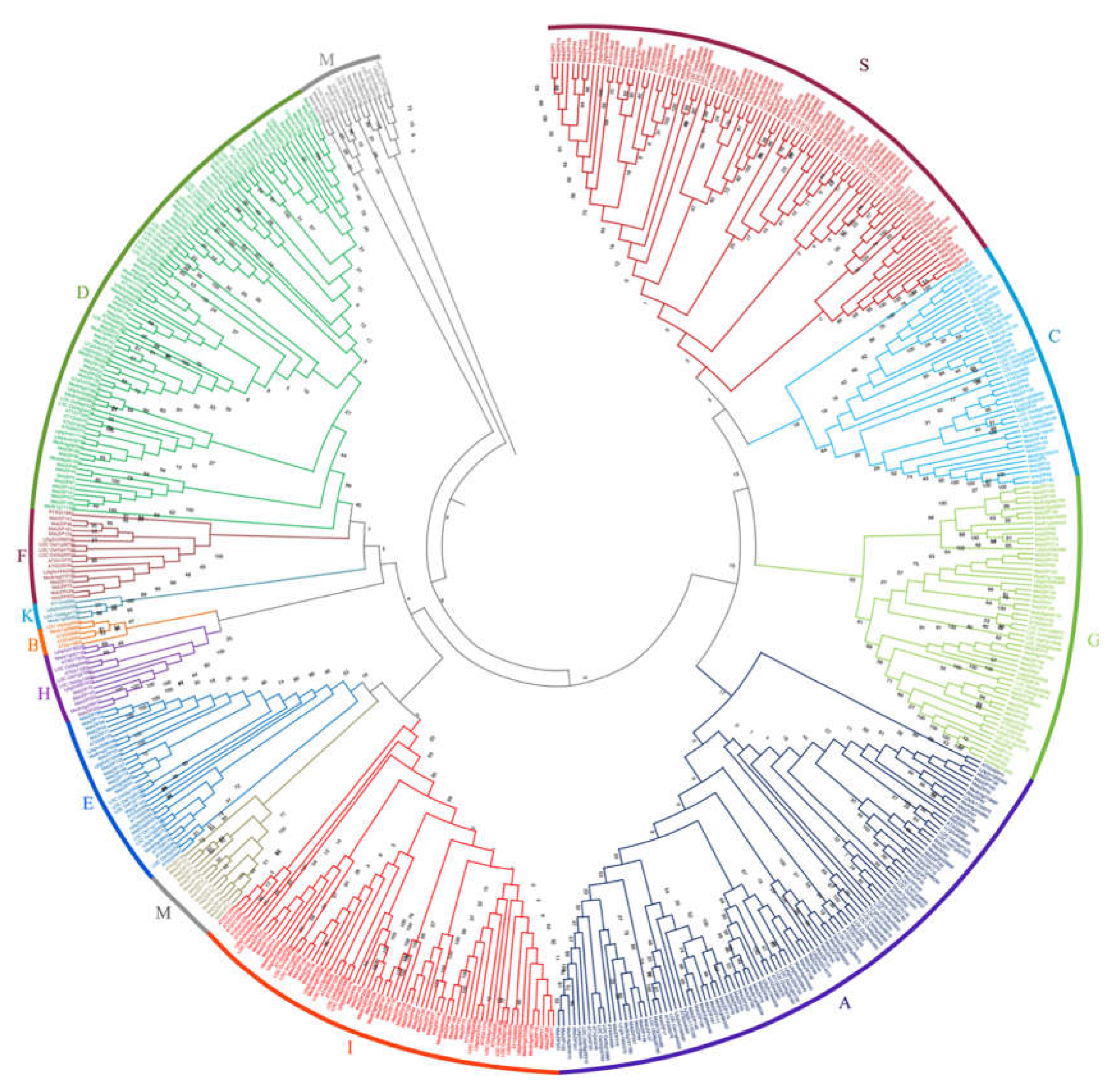
Figure 2.
Multiple sequence alignment of alfalfa bZIP proteins of group A. The alignment was performed using MUSCLE 3.8.31 and visualized using Unipro UGENE v.33. The A group contains 43 bZIPs in alfalfa, which are highly expressed during abiotic stress. A consensus sequence is provided at the top of the figure (in bold). The bars above the consensus sequence present the percentage of consensus amino acid base between the aligned sequences. The ruler below the consensus sequence provides the position of the amino acid base on the aligned sequences. The color changes from light to dark with dark color indicating highly conserved amino acid bases. The consensus sequence at the top indicates the level of conservation with a capital letter indicating high conservation and a small letter with low conservation. The highly conserved bZIP domain (N-x(7)-[RK]-x(9)-L-x(6)-L-x(6)-L), between 429 to 461, is presented in dark color.
Figure 2.
Multiple sequence alignment of alfalfa bZIP proteins of group A. The alignment was performed using MUSCLE 3.8.31 and visualized using Unipro UGENE v.33. The A group contains 43 bZIPs in alfalfa, which are highly expressed during abiotic stress. A consensus sequence is provided at the top of the figure (in bold). The bars above the consensus sequence present the percentage of consensus amino acid base between the aligned sequences. The ruler below the consensus sequence provides the position of the amino acid base on the aligned sequences. The color changes from light to dark with dark color indicating highly conserved amino acid bases. The consensus sequence at the top indicates the level of conservation with a capital letter indicating high conservation and a small letter with low conservation. The highly conserved bZIP domain (N-x(7)-[RK]-x(9)-L-x(6)-L-x(6)-L), between 429 to 461, is presented in dark color.

Figure 3.
Conserved bZIP domain. (A) the conserved basic region of the bZIP motif (motif 12-29). The basic region is composed of an invariant motif (N-x7-R/K-x9) with 18 amino acid residues which can be observed. The basic region facilitates the sequence specific DNA binding. (B) The leucine zipper region that contains the heptad repeat of leucine (motif 5-19) or other bulky hydrophobic amino acids is represented by the figure. This region is important as it facilitates the dimerization specificity. This consensus sequence was generated using MEME suite 5.3.3. MEME (Multiple Em for Motif Elicitation) allows discovery of novel motifs in collection of nucleotides or protein sequences. The height of the character corresponds to how frequently the character occurs at that position in the motif.
Figure 3.
Conserved bZIP domain. (A) the conserved basic region of the bZIP motif (motif 12-29). The basic region is composed of an invariant motif (N-x7-R/K-x9) with 18 amino acid residues which can be observed. The basic region facilitates the sequence specific DNA binding. (B) The leucine zipper region that contains the heptad repeat of leucine (motif 5-19) or other bulky hydrophobic amino acids is represented by the figure. This region is important as it facilitates the dimerization specificity. This consensus sequence was generated using MEME suite 5.3.3. MEME (Multiple Em for Motif Elicitation) allows discovery of novel motifs in collection of nucleotides or protein sequences. The height of the character corresponds to how frequently the character occurs at that position in the motif.
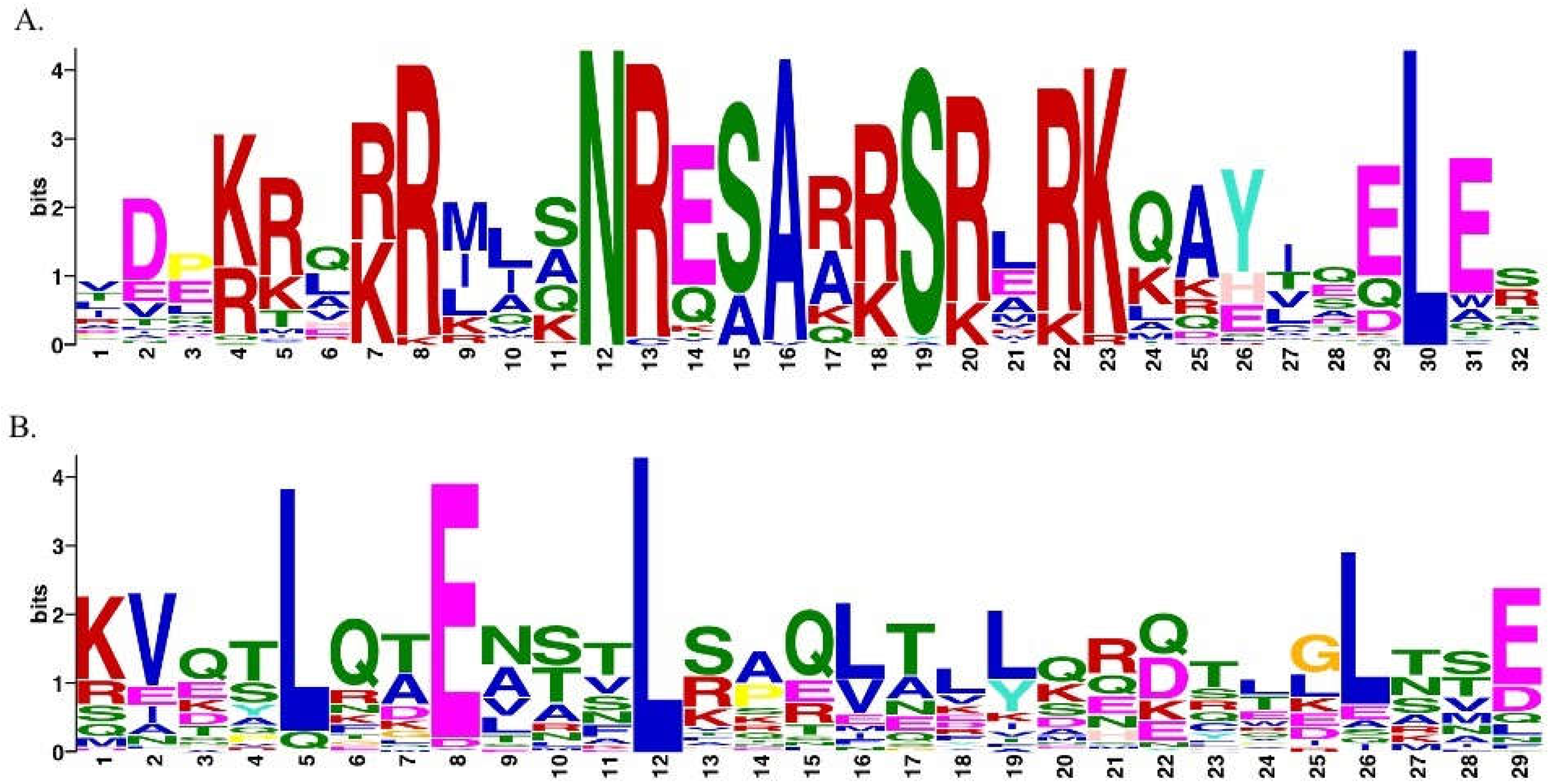
Figure 4.
Functional annotation of bZIP proteins in alfalfa. Distribution of genes in different GO categories for Biological Process (pink), Cellular Component (green) and Molecular Function (blue). In the molecular function category, most of the genes were assigned to DNA binding transcription factor activity followed by sequence-specific DNA binding. Similarly, most of the biological process of these genes involves regulation of transcription and these genes are mostly located inside the nucleus as presented in the cellular component category.
Figure 4.
Functional annotation of bZIP proteins in alfalfa. Distribution of genes in different GO categories for Biological Process (pink), Cellular Component (green) and Molecular Function (blue). In the molecular function category, most of the genes were assigned to DNA binding transcription factor activity followed by sequence-specific DNA binding. Similarly, most of the biological process of these genes involves regulation of transcription and these genes are mostly located inside the nucleus as presented in the cellular component category.
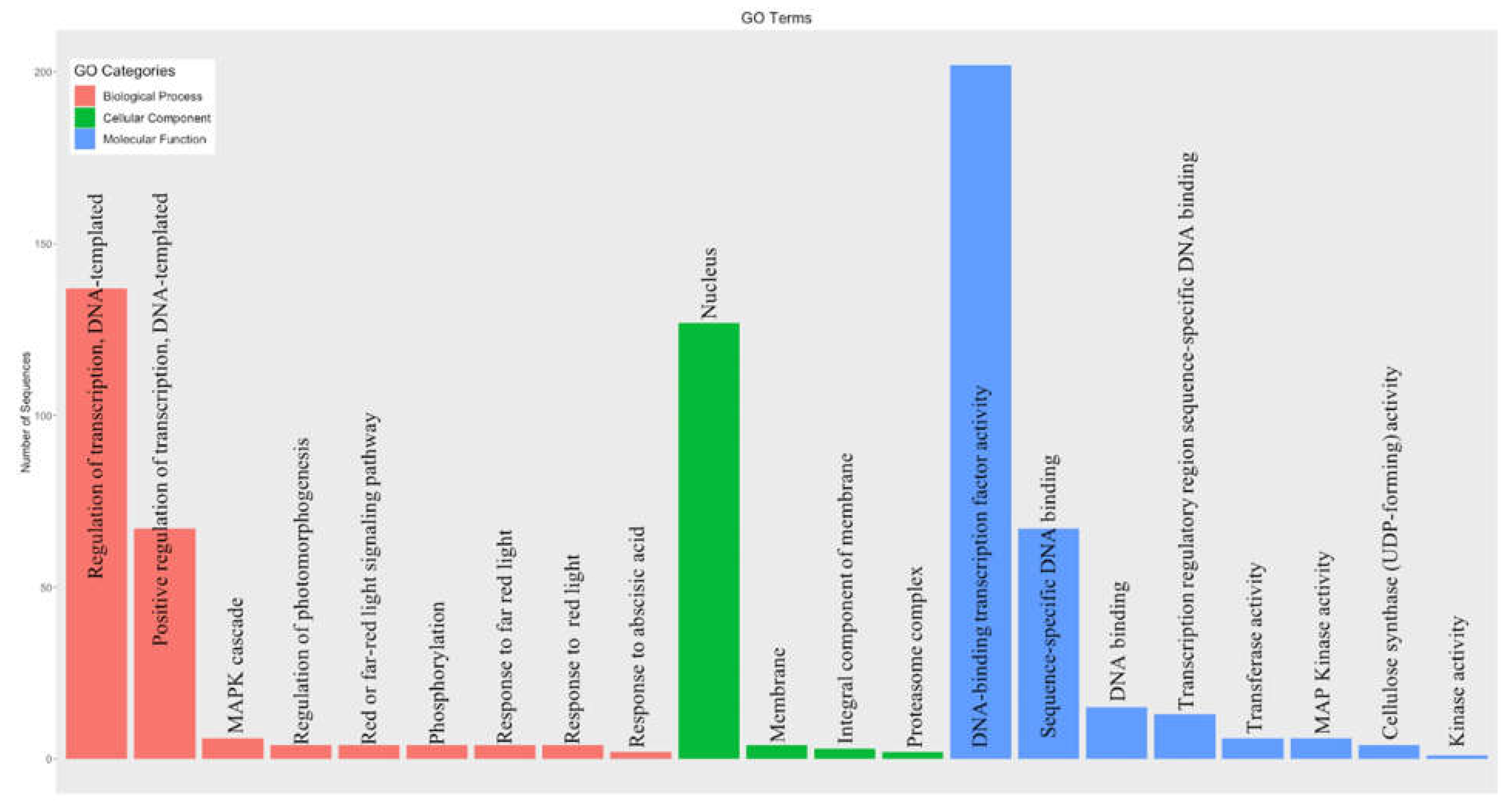
Figure 5.
Expression profile of alfalfa bZIP genes among different tissues and organs. In the figure, PES is pre-elongated stem. Most of the genes were highly expressed in Stem, Flower, Leaf, Nodules, Pre-elongated stem (PES) and Root. The genes that were expressed in one tissue were not expressed in the other tissue indicating different genes may be required during different growth stages of alfalfa. Different groups are represented to the side of the genes by the respective group name along with the color as represented in the phylogenetic tree.
Figure 5.
Expression profile of alfalfa bZIP genes among different tissues and organs. In the figure, PES is pre-elongated stem. Most of the genes were highly expressed in Stem, Flower, Leaf, Nodules, Pre-elongated stem (PES) and Root. The genes that were expressed in one tissue were not expressed in the other tissue indicating different genes may be required during different growth stages of alfalfa. Different groups are represented to the side of the genes by the respective group name along with the color as represented in the phylogenetic tree.
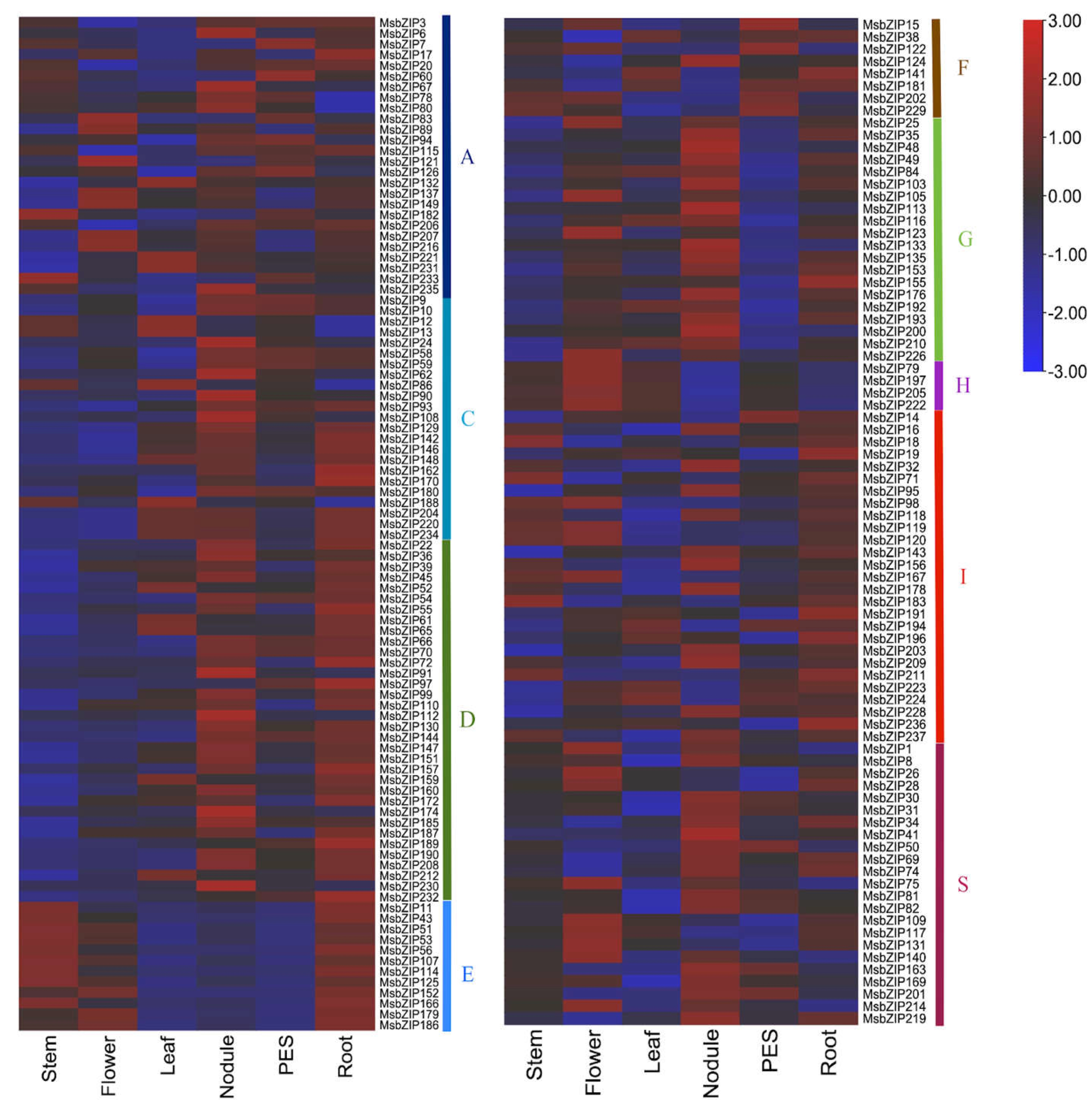
Figure 6.
Expression profile of 146 alfalfa bZIP genes in response to ABA, drought, and salt stress. Most of the genes were highly expressed during initial treatment of salt stress from 0.5 to 3 hours. For salt stress, the genes that were highly expressed during the 0.5 hour of treatment were also actively expressed during 3 hours of treatment. For drought stress, gene expression levels were increased as the duration of drought stress was increased from 1 hour to 3 hours to 6 hour and more genes were expressed during 6 hour of drought stress. ABA treatment showed only a few genes that were expressed as the stress duration was increased to 3 hours and 12 hours. Different groups are represented to the side of the genes by the respective group name along with the color as represented in the phylogenetic tree.
Figure 6.
Expression profile of 146 alfalfa bZIP genes in response to ABA, drought, and salt stress. Most of the genes were highly expressed during initial treatment of salt stress from 0.5 to 3 hours. For salt stress, the genes that were highly expressed during the 0.5 hour of treatment were also actively expressed during 3 hours of treatment. For drought stress, gene expression levels were increased as the duration of drought stress was increased from 1 hour to 3 hours to 6 hour and more genes were expressed during 6 hour of drought stress. ABA treatment showed only a few genes that were expressed as the stress duration was increased to 3 hours and 12 hours. Different groups are represented to the side of the genes by the respective group name along with the color as represented in the phylogenetic tree.

Figure 7.
Expression analysis of five genes during abiotic stresses based on RT-qPCR. Columns represents individual genes, while rows represent four stress conditions (cold, drought, salt, and ABA). The different treatment time point of 0-hour, 1 hour, 3 hour and 24 hour is presented in xaxis, while y-axis presents the expression value. A line is used to show the significant expression at different level of significance (0.05, 0.01, 0.001).
Figure 7.
Expression analysis of five genes during abiotic stresses based on RT-qPCR. Columns represents individual genes, while rows represent four stress conditions (cold, drought, salt, and ABA). The different treatment time point of 0-hour, 1 hour, 3 hour and 24 hour is presented in xaxis, while y-axis presents the expression value. A line is used to show the significant expression at different level of significance (0.05, 0.01, 0.001).
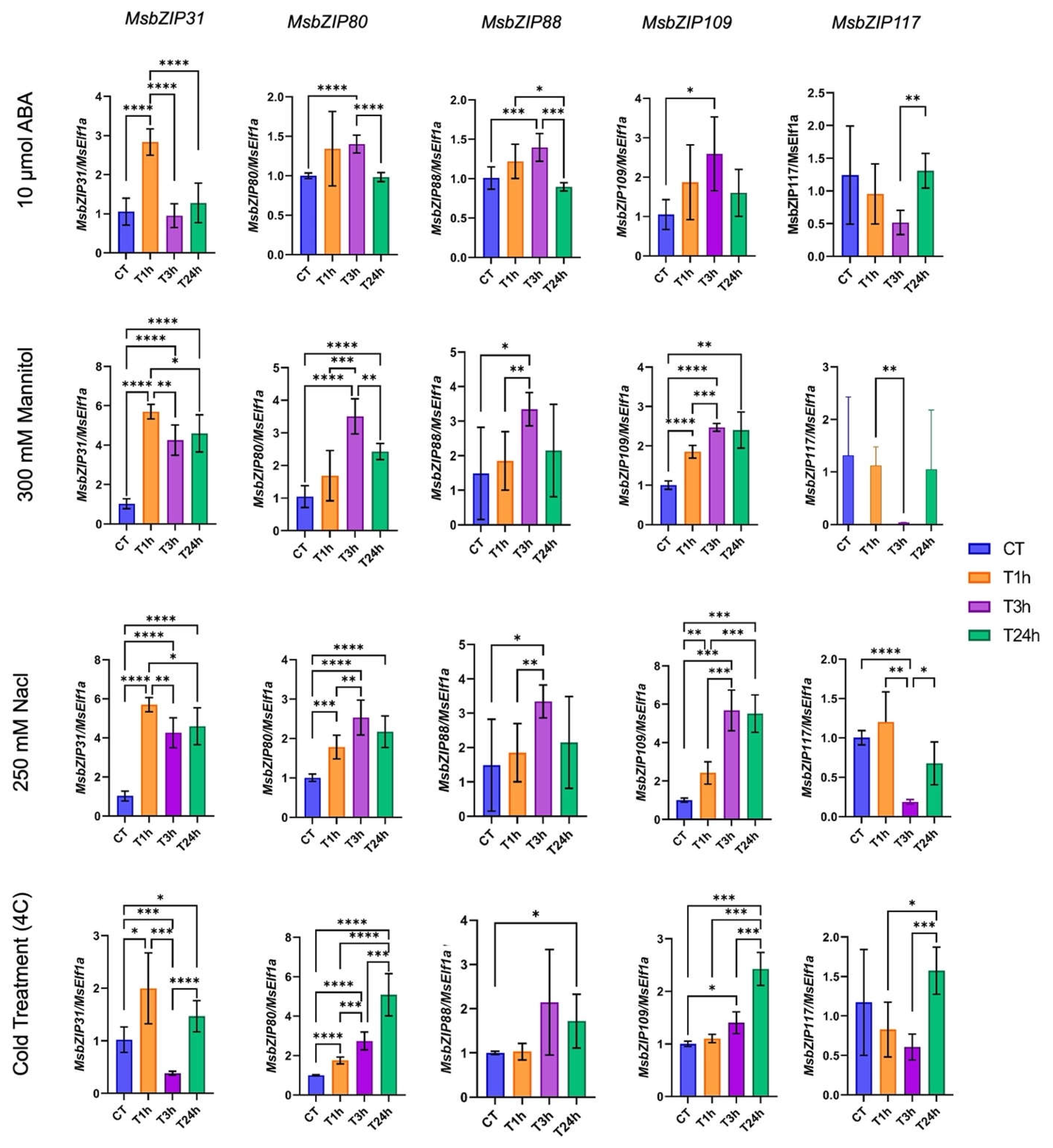
Figure 8.
Expression analysis of genes for developmental stages. (a.) represents tissues extracted from 5 days after germination (DAG) and 2 weeks after germination (WAG), while (b.) represents tissues extracted from 2 weeks seedlings from hypocotyl and leaf. A horizontal line showing significant expression at different level of significance (0.05, 0.01, 0.001) is also presented.
Figure 8.
Expression analysis of genes for developmental stages. (a.) represents tissues extracted from 5 days after germination (DAG) and 2 weeks after germination (WAG), while (b.) represents tissues extracted from 2 weeks seedlings from hypocotyl and leaf. A horizontal line showing significant expression at different level of significance (0.05, 0.01, 0.001) is also presented.


Table 1.
Comparative genome size and number of bZIP proteins in different model crops used in the study.
Table 1.
Comparative genome size and number of bZIP proteins in different model crops used in the study.
| Species | Chromosome | Genome Size | Number of bZIPs |
|---|---|---|---|
|
A. thaliana (9,11) |
2n = 2x =10 | 135 Mb | 78 |
|
B. napus (17) |
2n=2x1+2x2=38 | 1.16 Gb | 247 |
|
L. japonicus (7) |
2n = 2x = 12 | 470 Mb | 70 |
|
M. truncatula (43) |
2n = 2x =16 | 390 Mb | 75 |
|
O. sativa (8) |
2n = 2x = 20 | 430 Mb | 89 |
| M. sativa (Current Study) | 2n = 4x = 32 | 3,150 Mb | 237 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



