Submitted:
04 April 2024
Posted:
08 April 2024
Read the latest preprint version here
Abstract
Lipofuscin is indigestible garbage that accumulates in the autophagic vesicles and cytosol of post-mitotic cells with age. Drs. Brunk and Terman postulated that lipofuscin accumulation is the main or at least a major driving factor in aging. They even posited that the evolution of memory is the reason why we get lipofuscin at all, as stable synaptic connections must be maintained over time, meaning that the somas of neurons must also remain in the same locale. In other words, they cannot dilute out their garbage over time through cell division. Mechanistically, their position certainly makes sense given that rendering a large percentage of a post-mitotic cell’s lysosomes useless must almost certainly negatively affect that cell and the surrounding microenvironment. Here, I explore the possibility that the accumulation of lipofuscin may to some extent exacerbate every other kind of age-related damage. I do not think that lipofuscin removal will reverse/prevent all forms of aging, just the major component facing us currently. In this piece, I will review what is known about lipofuscin accumulation from evolutionary and mechanistic standpoints and discuss a method of removing it from single cells in vitro.
Keywords:
Anti-aging
; lipofuscin
; Hydra vulgaris
; Alzheimer's disease
; TFEB
; FluidFM
Introduction
Biological aging is a complex molecular process that takes place over time in all organisms. However, organisms have evolved mechanisms to repair various forms of age-related damage. For example, DNA repair enzymes exist that can fix damage in nuclear DNA. Mitophagy enables the degradation of damaged mitochondria. The immune system, when one is young at least, eliminates at least some proportion of one’s senescent cells.
There are many theories about why we age, but one stands out to me as being the most plausible based on the evolutionary and mechanistic evidence. That is the “garbage catastrophe theory of aging.” Drs. Brunk and Terman postulated years ago that the problem of aging can essentially be summed up as a “garbage disposal issue [1].” The main idea is that basically old molecules are sometimes damaged in ways that prevent the lysosomes from breaking them down properly, and over time these damaged, old molecules accumulate inside the lysosomes. Eventually, the lysosomes become full of this indigestible garbage, i.e., “lipofuscin”, and cannot perform their normal function - then there is a garbage back-up and the cell starts to decline health-wise.
There are two arguments for lipofuscin removal being the most important goal of anti-aging science currently. One is evolutionary and one is mechanistic.
Evolutionary Argument
In nature, there are only a handful of organisms that can be said to be essentially biological immortal. Hydra vulgaris (i.e., magnipapillata) is one of such organisms, and the reason for this might be that its indigestible garbage is essentially released from its body over time. It has three cell lineages - ectodermal epithelial, interstitial, and endodermal epithelial. All the epithelial cells in its body column are stem cells that continuously divide - displacing cells toward its extremities. The cells at the extremities slough off eventually [2,3]. In terms of the interstitial lineage, the differentiated cells that its stem cells produce are closely associated with epithelial cells and so are continuously displaced as well. This is a convenient way to dispose of lipofuscin - i.e., through dilution and cell shedding. However, continuous replacement of neurons may not allow for the stable inter-neuronal interactions required for long-term memory [4]. Lobsters continually grow throughout life; their fully differentiated cells express telomerase, allowing them to keep dividing as needed [5]. This includes the cells of their central nervous system, which allows for adult neurogenesis [6]. They also shed their shells periodically. Thus, the same logic appears to apply to them. However, their growth process does not appear to be fast enough to prevent lipofuscin from accumulating over time [7]. Notably, lipofuscin accumulation in eyestalk ganglia [8] is used as a gauge of biological age in lobsters (and myocardial lipofuscin accumulation can be used a marker of chronological age in humans [9]). Lobsters can retain memories, but only for a short time span [10]. Naked mole rats are also very long lived, and it has been shown that they have unusually active autophagic systems [11,12] (in addition to better anti-cancer defenses [13]). Even with better autophagy, naked mole rats still do accumulate lipofuscin in their post-mitotic tissues [14].
It appears as though all animals that age, e.g., flies [15], worms [16,17], lobsters, naked mole rats, mice [18], non-human primates [19], and humans [20] accumulate lipofuscin in their post-mitotic tissues. None of the aforementioned organisms seem to possess any evolutionarily “built-in” ways of exporting the lipofuscin that accumulates in their post-mitotic cells from their bodies, presumably because that would be an unnecessary expenditure of energy in light of procreation.
Export from the post-mitotic cells themselves is possible through exocytosis [21], extracellular vesicle secretion, or secretory autophagy. That part is not too energetically costly. However, from in vitro studies, it does not seem as though lipofuscin is exported from post-mitotic cells very often [22]. It also has only rarely been observed in vivo [23].
More importantly, when exported, there would ultimately be nowhere for the garbage to go except to be picked up by tissue-resident or circulating phagocytes, which themselves become bloated with lipofuscin. (Transfer of lipofuscin to tissue-resident phagocytes through tunneling nanotubes [TNTs] or partial cell fusion is also theoretically possible.)
Lipofuscin within aged tissue-resident macrophages is perhaps mostly derived from damaged molecules generated by their own, internal metabolic processes - rather than the phagocytosis of extracellular, lipid-saturated debris or efferocytosis in the context of aged, lipofuscin-laden cells [24].
Crucially, I have seen no evidence in the literature that tissue-resident or circulating phagocytes efficiently leave the body through migration to the gastrointestinal tract, urogenital tract, skin, or lungs, except possibly when there are infections in those areas.
Mechanistic Argument
Lipofuscin is broadly a complex amalgam of highly oxidized cross-linked macromolecules, including proteins, lipids, sugars, and metal cations. It varies in composition between species, individuals, cell types, and plausibly even cells of the same type [20,25,26,27].
While it was originally widely believed that lipofuscin is inert, it may in fact permeabilize or otherwise destabilize lysosomes and promote apoptosis or necrosis [20,28,29,30,31] Even if the damaged molecules are mostly sequestered within lysosomes and are not actively harmful to the cell, the fact that many lysosomes become full of garbage and therefore are almost surely unable to perform their normal functions nearly as well just logically seems as though it must be a major problem for the cell. If a critical threshold is reached in enough cells in a tissue, e.g., the brain, it clearly would be problematic. The cells may try to produce more lysosomes - but will eventually reach capacity.
Along these lines, lipofuscin accumulation decreases the ability of cells to adapt to amino acid starvation [32] and increases their susceptibility to oxidative stress [33]. Increases in the dietary intake of metal cations such as Fe2+, which plays a key role in the formation of lipofuscin, augments lipofuscin accumulation [34,35,36] and speeds up aging [37]. Manganese acts similarly [38]. Furthermore, artificial lipofuscin loading into human cells results in a significant loss of cellular viability [39,40] - although it is unclear how similar artificial lipofuscin is to real, age-related lipofuscin. Another study showed that the dietary intake of artificial lipofuscin shortens the lifespan of Drosophila melanogaster [41].
With regard to short-lived species, like mice and rats, lipofuscin may not have enough time to accumulate to pathological levels before they die of cancer. It is estimated that 50-90% of aged mice die of cancer [42]. Even still, it was shown that in the cerebral cortex neurons of lamina Vb in 630-700-day old rats, lipofuscin occupied 23% of the soma volume [43,44]. This could still certainly have a negative physiological effect. Unsurprisingly, we do see a cognitive decline in mice with age [45]. However, even the oldest mice do not develop age-related neurodegenerative disorders such as Alzheimer’s disease, Parkinson’s disease, or amyotrophic lateral sclerosis [46]. Ostensibly, they simply do not live long enough for sufficient build-up of lipofuscin in their neurons. Other conditions like age-related macular degeneration and sarcopenia do occur in mice, but whether their most severe cases are as bad as the most severe human cases is unclear to me. It is also possible that some mouse tissues accumulate lipofuscin more rapidly than others due to cell type differences in metabolic rates, etc. Some types of human neurons, for example, have not accumulated much lipofuscin by the time others are nearly full [47].
With regard to humans, it has been demonstrated that multiple neuronal cell subtypes become densely packed with lipofuscin granules with age [48,49,50]. In large motor neurons of centenarians, lipofuscin constitutes up to 75% of total cytoplasmic volume [51]. Other post-mitotic cell types also accumulate substantial amounts of lipofuscin [9,52,53]. Lipofuscin-laden lysosomes are often much larger than typical lysosomes. The typical size of a lysosome in a fed, unaged cell is ~100 nm-500 nm in diameter [54]. In contrast, lipofuscin granules are generally 1-5 microns in diameter [54].
Finally, lipofuscin accumulation also explains the downward spiral of functionality that is seen in aging - i.e., the rapid acceleration in decline starting around 60-70 years of age [55]. That is because the decline in autophagy probably increases the rate at which lipofuscin is formed by allowing aggregates to stay around longer and develop further oxidative damage. Also, lipofuscin accumulation may lead to more free radical production [56], thus accelerating its accumulation as lipofuscin is heavily comprised of oxidatively damaged biomolecules. Furthermore, eventually, when autophagy levels have decreased substantially in many cells in a tissue, the rate of accumulation of other forms of age-related damage probably accelerates as well, which could further accelerate lipofuscin accumulation.
If whole-body lipofuscin removal were started at 30 years of age and repeated periodically every decade or so, the extracellular matrix (ECM) may be kept in a good condition. With regard the elderly, however, the extracellular matrix may be an issue. That is because cells throughout their bodies would have been disabled for so long - and would not have been able to properly take care of their surrounding ECM during that time period. It is likely that lipofuscin removal will still be substantially rejuvenating, however, as damaged ECM components can be excised - although they may also be indigestible or become lipofuscin once inside the lysosomes of nearby cells. Induced ECM turnover is also a possibility [57].
The hydra may also shed its ECM when it sheds cells, so perhaps lipofuscin shedding is not the only consideration for that organism. The ECM surrounding its stem cells may not be shed or turn over that frequently - but still remain youthful.
Testing the Importance of Lipofuscin Accumulation in Aging In Vitro Now
TFEB is the master regulator of autophagy (and lysosomal biogenesis) [58]. TFEB overexpression increases lysosomal acidity and boosts lysosomal hydrolase levels in lysosomes. It was shown that TFEB overexpression can shrink lipofuscin deposits - but this work was done in the context of a transgenic mouse model with overproduction of an aggregating protein, which I would define as “false” lipofuscin [59]. Namely, if the protein ceased to be produced and the cells weren’t already completely overwhelmed, it could be degraded over a reasonable amount of time by endogenous lysosomal hydrolases.
TFEB could still be of use for true lipofuscin, however. It also induces the exocytosis of lysosomal contents through fusion of the lysosomal and plasma membranes. One problem with boosting endogenous exocytosis processes is that they may lose potency with age; oxidatively warped lipids from lipofuscin may insert themselves into the membranes of the lysosomes that contain them and prevent efficient fusion with the plasma membrane. The main problem, however, is that the two most well-known inducers of lysosomal exocytosis, Ca2+ and TFEB, do not induce a substantial amount of exocytosis - even in youthful cells [60,61].
Notably, a small molecule was shown to induce some lipofuscin exocytosis in monkey retinal pigment epithelial cells, which are important in macular degeneration. However, this small molecule, remofuscin, is a potent and reversible inhibitor of the H+/K+ ATPase [62]. Proton pump inhibitors increase lysosomal pH and activate TFEB - so remofuscin’s effects may not be additive with TFEB overexpression.
In order to test the theory that lipofuscin is currently the main issue with regard to age-related disease in a way that may be currently feasible, we could utilize a recently developed organelle sampling FluidFM approach [63] to remove lipofuscin-laden lysosomes from single skeletal muscle cells taken from biopsies of elderly humans [64,65,66]. By doing so, we could observe whether lipofuscin removal reverses the age-related phenotype of these cells. We could also determine how long they live in culture compared to untreated single skeletal muscle cells.
Along these lines, in one study, it was shown that at ~day 25, rat hippocampal neurons in vitro have an aged phenotype, including dysfunctional mitochondria, higher reactive oxygen species production, and at least one feature of senescence [67]. It was also shown that defective autophagy was linked to this senescence-like phenotype - and that aged neurons in culture accumulate lipofuscin [68]. Thus, if the post-mitotic cells’ lipofuscin were removed from the cells and new, functional lysosomes were introduced exogenously or generated through TFEB, their aged phenotype may reverse.
Cells from elderly patients may require a more extensive initial session of lipofuscin removal. That is because once the garbage back-up is cleared at the end-point, i.e., the lysosomal level, oxidatively damaged mitochondria would then be mitophagocytosed and cytoplasmic lipofuscin granules would possibly be engulfed by autophagosomes and delivered to new lysosomes. Thus, new lysosomes might be quickly overwhelmed again - and they should be exported as well. Perhaps three rounds of lysosomal removal in quick succession would be appropriate for the first treatment session. The elderly cell may also have autophagosomes that have been present for too long in the cytoplasm. Their outer membranes may have become corrupted by oxidatively warped lipids; thus, they may not even be able to fuse with new lysosomes. They might have to be exported as well.
It is important to keep in mind that while primary cell cultures often die after a few days-weeks [69], the cause of death may not be related to normal aging. It may instead be related to infection or hyperosmolality due to evaporation of the media [70]. These factors should be mitigated or negated if possible. This is perhaps especially tricky when it comes to single-cell manipulations.
The FluidFM-based approach involves a small needle that penetrates the cell membrane and can inject content into or aspirate content from a single cell [63]. Single skeletal muscle cells from elderly patients could be injected with magnetic nanoparticles (MNPs) conjugated with a nanobodies that target LAMP-1. Then, a magnetic field could be applied to cluster the lysosomes to one small area at the plasma membrane. They would then be removed by the needle. New lysosomes could then be injected to replenish the cell’s stores - or perhaps generated via transient, induced TFEB overexpression.
To prevent cell death, one may need to co-inject some number of new lysosomes with the nanobody-conjugated MNPs. The new lysosomes would first be pre-incubated with nanobodies that are not conjugated to MNPs, however - to sterically block the relevant LAMP-1 site.
Figure 1 FluidFM for Lipofuscin Removal.
Proof-of-Concept In Vivo
Proof-of-concept for lipofuscin removal in vivo could potentially be undertaken in an animal model with various transgenes installed in a genomic safe harbor locus [71], once secretory autophagy [72] of lysosomes can be reliably induced in a wide variety of cell types - ideally with a small number of proteins overall.
Tissue-resident macrophages (TRMs) would secrete a peptide that activates a synthetic receptor in post-mitotic cells [73], triggering a substantial duration of TFEB overexpression, as well as a burst of secretory autophagy of old/mixed lysosomes.
For microglia, cycles of random migration and peptide secretion could potentially be effected via inducible LRRK2 inhibition [74], and for peripheral TRMs, inducible Bmal1 inhibition may suffice [75]. For the latter, LRRK2 inhibition seems to have the opposite effect on as on microglia [76]. However, in vivo migration in an extensive and random enough way may require a tactic involving a self-generated chemokine gradient and a synthetic gene circuit like a noisy repressilator [77,78].
TFEB overexpression would generate new lysosomes in the cell to compensate for some of the remaining unladen lysosomes being exported via secretory autophagy. TFEB-mediated lysosomal exocytosis should be inhibited, however. That is because secretory autophagy would allow the lipofuscin to stay encapsulated in the extracellular space, which would facilitate pick-up by bioengineered macrophages in vivo - and perhaps reduce the chances of lipofuscin pieces to be taken up by neighboring cells or even taken back up by the cells that ejected the granule initially.
Notably, mitochondria secreted via secretory autophagy seem to lack an encapsulating membrane after export [79]; thus, in the case of lysosomes, a lysosomal membrane protein, e.g., LAMP-1, could be targeted by a TRM chimeric antigen receptor for phagocytosis (CAR-P) [80].
Secretory autophagy could possibly be enacted in one or more cell types via a particular strategy. The cells could express a nanobody targeting LAMP-1 with a fused domain that interacts with Sec22b - as well as an LC3-interaction region (LIR) [81]. They would also express Sec22b, Stx3/4, and SNAP23/29 [72]. Furthermore, autophagosome-lysosome fusion could be inhibited via a dominant-negative STX17 variant [82]. Targeting a plus-end directed motor protein to the “differentiated” cell-containing autophagosome would also be of use to move it to the periphery.
TRMs could employ a synthetic gene circuit to count the number of lysosomes they phagocytose [83,84]. Also, the phagosomes containing said lysosomes should be unable to fuse with TRM lysosomes. Once an experimentally determined number of lysosomes have been phagocytosed - a given TRM would asymmetrically divide, wherein one progeny cell inherits all of the aforementioned phagosomes. They would also irreversibly express a receptor that would mediate their chemoattraction [85,86] to the gastrointestinal lumen [87] for expulsion from the body. Alternatively, if necessary, a device could be placed in the peritoneal space that slowly releases a chemoattractant for the TRMs - allowing for non-invasive, albeit manually effected egress [88].
As whole lysosomes are being extracted from the body, true lipofuscin may not be required to test the efficacy of the approach. Mice with a Cre-inducible, central nervous system (CNS)-specific lysosomal storage disorder could be employed [89]. Alternatively, an adult version of the P301S mouse model of tauopathy could perhaps be employed [59]. An immunocompromised mouse model could be utilized for testing purposes. Also, extraction from the peritoneal space would perhaps be easiest initially, as deschloroclozapine [85,86] can be slowly released from biodegradable beads placed there laparoscopically [88].
Periodic whole-body lipofuscin removal:
In the future, the systemic depletion of tissue-resident macrophages (TRMs) could be effected via the CSF1R inhibitors, PLX3397 [90] and PLX5622 [91] - perhaps combined with an anti-CD45 immunotoxin [92]. PLX3397 has been tested in clinical trials [93], and PLX5622, an inhibitor that crosses the blood-brain barrier more efficiently than PLX3397, has been extensively tested in a preclinical context [94]. BLZ945 is another BBB-permeable CSF1R inhibitor, which has been tested in clinical trials [95]. “Lytic" anti-CD45 monoclonal antibodies have also been tested in human patients [96]. Microglial depletion in non-human primates appears to be safe, and microglial depletion can be effected multiple times in mice - as long as sessions are spaced out enough [94].
Notably, induced pluripotent stem cell (iPSC)-derived macrophages (iMacs) can be generated in large quantities in bioreactors [97], and edited autologous iMacs could be intravenously infused to repopulate the patient’s TRMs. iMacs take on the signature of the tissue they inhabit, to some degree [98]. It is also possible to culture iPSC-derived monocytes in large quantities [99]; these may be more suitable for systemic replacement, as they may be more able to take on the transcriptional signature of multiple TRM-subtypes. Perhaps circulating monocytes can even be harvested and edited en masse outside the body, and then reinfused.
One of the edits would be to have a CSF1R variant that resists PLX3397 and PLX5622, to enable continuous selection of the iMacs or iPSC-derived monocytes over endogenous TRMs, which would normally replenish themselves quite effectively - or be reseeded by circulating monocytes produced by the patient’s hematopoietic stem cells (HSCS) [100]. The edited TRMs could also at least transiently secret a CD11b or CD64 immunotoxin when they reach their niches, to locally eliminate the patient’s original TRMs.
Notably, a CD45 immunotoxin would also deplete the patient’s hematopoietic stem cells (HSCs) [101]. However, clinicians could first edit HSCs with liposomally encapsulated mRNA [102] to generate a CD45 variant receptor that can evade the immunotoxin [103]. However, it was recently shown that TRMs have multiple subpopulations, two of which are replenished by patient HSCs [100]. One may need to edit the HSCs ex vivo to have the same (extensive) edits as the iMacs or iPSC-derived monocytes.
Imbuing patient HSCs ex vivo with the same edits as the infused myeloid cells prior would also allow for central tolerance of constitutively expressed gene products after lymphoid cell ablation via an intravenously administered CD45 immunotoxin.
The CSF1R variant would also be helpful for the HSCs - as the aforementioned inhibitors may affect HSCs as well as TRMs.
The edited TRMs could secrete proteins, RNA [104], and ribonucleoproteins [105] to surrounding cells.
COURIER, a form of RNA export [104], could be used to initiate secretory autophagy of lysosomes in surrounding cells, possibly using small RNAs to activate endogenous genes. To enhance the potency of the export process in this context, the RNA could be made self- or trans-replicating [106,107]. The packaging capacity of COURIER is at least 9.8 kb - different cargos could potentially be expressed and packaged into different nanocage copies, though - and alternative protein nanocage architectures could theoretically allow for larger cargo capacities if necessary [104]. A benefit of using nanocages rather than viral capsids is that they may be less immunogenic [104].
As TRMs only make up around 10% of many tissues, the motility aspect of approach would be crucial. Microglial random motility can be increased via LRRK2 inhibition, although this may have the opposite effect for other TRMs. Bmal1 inhibition is a possibility for TRMs as well. Perhaps a self-driven chemotaxis gradient combined with a gene circuit like the represillator could be utilized to enact migration in random directions for defined periods of time. However, other methods may be required instead or in addition. A noisy lysis gene circuit could help; repopulation by the remaining TRMs or by HSCs could increase the range of the treatment within tissues, as migration would be inherent in repopulation since the TRMs are spaced out within tissues.
Lysis could also be a side effect of adenovirus, vaccinia virus [108], or HSV-1 vector replication - as a means of transferring large DNA packets to surrounding cells. In the future, cytoplasmic plasmids that replicate up to a controlled copy number set point could be donated to surrounding cells as well. The same is true for a coronavirus, vaccinia virus, or an HSV-1 virus, all of which can bud from the plasma membrane. However, for the coronavirus, vaccinia virus or HSV-1, if a copy number control mechanism is utilized, continuous donation may be possible.
TRMs may wander outside of their host tissues, but perhaps they would simply assume the identity of whatever tissue they end up in - at least given time.
Even with enhanced motility - TRMs in some tissues like the eye only make up 1% of the resident cells. Induced replication of the TRMs in some tissues may be necessary - it will be important to ensure that this does not cause excessive inflammation. M1 macrophages are typically considered inflammatory, while M2 macrophages are anti-inflammatory. This binary classification may not represent the entire picture with regard to macrophage phenotypic states, but may be sufficient in this context.
Random motility should only occur - inducibly - when the periodic treatment is required - i.e., perhaps once a decade. Otherwise, it may impair the ability of the TRMs to fight infection.
One final mechanism of both motility and gene delivery is possible. Bacteria can take care of both motility and gene delivery. If the TRMs contain bacteria that replicate up to a tolerable copy number via quorum sensing - they could continuously donate them via secretory autophagy to the surrounding cells. HSC-derived TRMs could secrete a chemoattractant molecule and entry receptor for the bacteria - perhaps combined with a noisy gene circuit - so that some of the exported bacteria enter them and reach quorum sensing levels as well — for subsequent donation. Or, the HSC-derived TRMs could be lysed by the self-renewing TRMs potentially - if depletion allows them to take over the niches of the HSC-derived TRMs. This may put a lot of stress on the patient’s HSCs, however. Perhaps more hTERT expression would help with that potential issue. Alternatively, perhaps we could shut down monocyte production from the HSCs - temporarily at least.
The TRMs can migrate randomly and secrete the bacteria - which could secrete a peptide small molecule that causes the TRMs to “chase” them [109]. Also, the bacteria could enter surrounding cells and lyse to release a replicating RNA component [110] - or a overproduce an engineered T4 phage and lyse - to release DNA that can reach the nucleus of target cells [111].
Eventually, in an opposite strategy - the HSCs could be used to reseed all three niches potentially - as artificial TRMs may not be healthy long-term [112].
The bacteria can express CD47 minimal peptides [113] or recombinant CD47 on their surfaces and. A noisy gene circuit could allow them to shed the capsule when they need to enter target cells - through a K5 lyase [114] and a few rounds of division. Also, a noisy gene circuit could cause lysis if the bacteria are extracellular for too long - if they cannot enter a target cell for some reason. A lack of ActA promoter activity after cell egress would trigger a countdown mechanism for this.
Also, asymmetric bacterial division inside the surrounding cells - perhaps in the context of a Salmonella Typhimurium vector [115,116] - could lead to one bacterium lysing the vacuole and overproducing the T4 phage - the remaining bacterium could detect whether gene transfection has occurred properly.
During the process of lipofuscin removal, dexamethasone may be sufficient - along with the capsule element - for attenuating immunogenicity.
Gene silencing is an issue for both surrounding cells and the TRMs/HSCs themselves. That is why an RNA delivery method may be more ideal. Also, the TRMs/HSCs can be imbued with a kill switch. Each time the treatment is required, some HSCs can be harvested and all the TRMs and HSCs in the patient can be eliminated, followed by reintroducing the HSCs to their niches. New TRMs would be infused, and hTERT overexpression in the reintroduced HSCs could help with repopulation.
Whole-body lipofuscin removal should probably be effected once every decade - ideally starting at ~30 years of age.
The innate immune response to dsRNA could possibly be attenuated by the expression of the MERS-CoV ORFa protein [117].
As periodic induction would also be required in this context, the adaptive immune response to the RNA-dependent RNA polymerase may need to be attenuated as well. First, the vector could encode endogenous or deimmunized protein components [118]. Second, it may help if the vector were to incorporate multiple, tandem miR-142-3p binding sites in the protein mRNA 3’UTRs [119]. Third, the vector could express the SARS-CoV-2 ORF6 protein, which inhibits the MHC class I pathway [120].
As mentioned before, mice do not seem to accumulate as much lipofuscin as humans, so testing this approach in mice would not necessarily be too informative. Non-human primates would be the best model organisms in this context. However, perhaps pigs, cats, or dogs would be possible model organisms that could be useful in this context, as well.
Periodic, whole-body lipofuscin removal should be tested in a preventative as well as a rejuvenative and maintenance capacity.
There are two considerations for elderly patients - i.e., the rejuvenative and maintenance capacity. Initially eliminating their TRMs may release a lot of lipofuscin into tissues. Perhaps that will not be much of an issue, but if so - the therapy can be started immediately with the infused, edited TRMs/edited HSCs, employing TRM TFEB overexpression and, round of symmetrical division, and migration of one of the progeny cells to an extraction point.
TRM TFEB overexpression and asymmetric division could also be employed, but with lysosomes being inherited by one progeny cell rather than phagosomes, as the much of the lipofuscin released from the initial TRMs may have resided in their cytoplasms rather than lysosomes.
Alternatively or in addition to the two previous mechanisms, a chaperone-mediated autophagy (CMA) mechanism could potentially be employed. The TRMs could be bioengineered so that they can detect when they are full of lipofuscin. One potential way of doing this would be to express a TEVp variant of with a KFERQ-motif, which targets protein substrates for CMA [121]. When enough lipofuscin has accumulated, TEVp would no longer be degraded. It would thus be free to liberate cyclic or endoplasmic reticulum-tethered transcription factors [122,123]. Using a synthetic gene circuit, it would be necessary to reach a threshold of TEV protease activity before expression of the chemotaxis receptor is permanently activated via recombinase-mediated gene cassette inversion [124].
Secondly, elderly patients will likely need an extensive first lipofuscin removal session for reasons described in the in vitro section. Also, if extracellular matrix (ECM) turnover efficiency is restored to youthful levels, damaged extracellular molecules could potentially be excised from the surrounding ECM and taken up; at least some of them may be indigestible by lysosomal enzymes [125].
In a previous piece, I called this strategy “CURE”, i.e., Clearance of Undigested Rubbish by Encapsulation, but I think “ via Extraction” is better. (The version of whole-body lipofuscin clearance that involves elimination in situ can also be encompassed by this acronym, of course.)
Again, ECM issues may be a problem - at least for the elderly. TRMs and the surrounding cells could inducibly secrete enzymes to turn over the ECM more frequently than normal, at least if DNA delivery is enacted to the surrounding cells.
Periodic, Systemic Senescent Cell Destruction
Periodic, systemic senescent cell elimination in the INK-ATTAC mouse model was shown in 2016 to substantially increase healthspan and lifespan [126]. The protein p16Ink4a has been shown to be expressed at higher levels by senescent cells; thus, in the INK-ATTAC mouse model, a minimal p16Ink4a promoter was designed to drive the expression of a small molecule-inducible Casp8.
Currently available small molecule-based senolytic drugs seem to be fairly weak, at least in human patients [127]. Recently, researchers have employed CAR T-cells to destroy senescent cells, although I’m not sure it is known how efficiently they can target senescent cells in the CNS after intravenous infusion. Also, I don’t believe they’ve shown as impressive results as the genetic INK-ATTAC model [128,129]. (CAR NK-cells are also being considered as senolytic agents [130].)
TRM replacement combined with COURIER could potentially be used for periodic senescent cell destruction - using a tuned ADAR approach targeting the p16Ink4a transcript [131,132] involving self- or trans-amplifying RNA, wherein the ADAR output is Casp8.
The amplification aspect could be limited via superinfection exclusion, copy number restriction, and gene dosage compensation.
Additional Longevity Therapies THAT Involve “Tweaking Metabolism”
Dr. Aubrey de Grey has stated that therapies seeking to treat aging by altering metabolic pathways, e.g., over or underexpressing various proteins, may be dangerous. Gene pathways have evolved over billions of years, and there is much still to learn about how they interact with each other. However, some of such interventions appear to be relatively safe in preclinical and clinical models.
For example, hTERT overexpression via adeno-associated viral (AAV) vector or cytomegaloviral (CMV) vector [133,134] seem to be safe approaches for increasing the lifespan of mice. In fact, a single human patient, Liz Parrish of BioViva, was intravenously injected with an AAV vector encoding hTERT [135].
Partial reprogramming is potentially an option as well [136].
Periodic TFEB overexpression may be a safe way to increase the amount of time between lipofuscin removal sessions [137].
FOXN1 expression in thymic epithelial cells may help to rejuvenate the aged thymus and counter involution [138].
Gene therapy could be crucial to help stave off cancer. Tumor suppressor protein overexpression has been shown to decrease the risk of cancer without causing side effects [139,140,141,142]. In fact, PTEN overexpression even seems to improve metabolism in mice [142]. Finally, DREAM complex inhibition could enable as effective DNA repair in somatic cells as in germline cells [143].
These proteins could be continuously provided to target cells by the TRMs, but continuously enhanced motility could be an issue for normal TRM functionality, and levels would likely be lower than with continuous expression in the target cells. DNA construct delivery to the nucleus of target cells would be better. As mentioned before, though, gene silencing could be problematic. Delivery of endosymbiont bacteria to the cytoplasm of target cells and maintained expression would be more ideal.
*Sequencing Our Mitochondrial and Nuclear DNA as Soon as Possible
Ideally, we would sequence our mitochondrial and nuclear genomes at as young an age as possible, to make sure our “reference sequences” are as close to the original as possible. Perhaps harvesting a small number of germline stem cells (GSCs) and sequencing the DNA from them would be most appropriate. They appear to have the best nuclear DNA repair mechanisms of all male cells [144] - and possibly the best mitochondrial DNA repair mechanisms. Evidence is starting to accrue that female GSCs exist as well [145]. (Oocytes can be harvested up until menopause - they have very efficient DNA repair mechanisms as well [146]; there are strict selection processes for oocyte mitochondria [147].)
If there a few mutations scattered throughout our mitochondrial and nuclear DNA reference genomes, however, it may not be much of an issue - or an issue at all.
Many, if not most individuals, may have mitochondrial heteroplasmy, wherein more than one genome variant is present in their tissues. Homoplasmy may be more rare, but it has been shown in mice that homoplasmy with a healthy genome variant allows for normal functionality [148]. However, it was also shown it is important for there to be a concordance between mitochondrial genomes and an organism’s nuclear genome [149] - although this is in only in the context of aging. Periodic lipofuscin removal may negate this issue. This is all to say that heteroplasmy should be taken into account when seeking to define an individual’s mitochondrial “reference” genome or genomes. We must be able to determine which of the patient’s mitochondrial genomes to utilize for their treatment - whether that be one or possibly multiple.
*Possibly Saving Cells with Pristine DNA as Soon as Possible
While we may eventually be able to “print” entire eukaryotic genomes into patient-derived or even synthetic cells, that may be a long ways off. We could save cells with pristine genomes for each patient now, so that we can convert them to iMacs or iPSC-derived monocytes to effect the adult stem cell replacement strategy described in the next section.
Eventually Fixing Nuclear and Mitochondrial Mutations
Dr. de Grey has suggested allotopic expression as a means of addressing mitochondrial DNA mutations with age [150]. Another approach would be to utilize the bacterial “launch pad” here. Bacteria can conjugate with mitochondria [151]. They could utilize a T4SS to inject pristine mtDNA into patient mitochondria and proteins that degrade the old mtDNA (exploiting synonymous mutations in the new mtDNA) [152].
Said proteins could also potentially be secreted by the bacteria then imported into the mitochondria, rather than being injected via T4SS [153].
After conjugation, the bacteria could also lyse and release a replicating RNA molecule that is subjected to copy number restriction. Proteins that degrade the old mtDNA could be continuously expressed by the construct and imported into the mitochondria to select for the new mtDNA over a longer period of time.
Fixing nuclear DNA mutations and damage in stem cells and long-lived post-mitotic cells on a fundamental level may be the most difficult challenge we face with aging. Notably, it has become clear that nuclear DNA mutations damage accumulate with age in post-mitotic cells as well as stem cells [154].
There may only be only feasible strategy for addressing this on fundamental level in the relatively near future - “whole-body induced cell turnover” [155]. Edited TRMs could eliminate adult stem cells via immunotoxins. They could also asymmetrically divide, wherein one progeny cell dedifferentiates into an iPSC via Yamanaka factor expression. The empty niche could potentially then guide the iPSC into engrafting and differentiating into the appropriate adult stem cell type. Then, the new, edited adult stem cell could be induced to kill tissue-resident cells and divide to repopulate the tissue. This could be a viable strategy even in the brain, if done slowly over time - and it may be necessary in the short-term. The telomeres of the edited, adult stem cells could be elongated via hTERT overexpression to enable more rapid repopulation of tissues than is typical.
However, iPSCs can form teratomas in vivo, so this may not be appropriate. The progeny cell of the TRMs that is for reseeding adult stem cells may need to more directly convert to the given stem cell type instead.
WILT
Blood cancers can be effectively treated with immunotherapy, although perhaps CAR T-cells with multiple, orthogonal receptors targeting a number of ubiquitous cell surface ligands expressed by the targeted white blood cell type could be employed in the future to avoid escape variants [156].
I have proposed a solid tumor treatment called “Oncolytic Vector Efficient Replication Contingent on Omnipresent Mutation Engagement” (OVERCOME). It involves using an intracellular, oncolytic vector that can only replicate once it has detected patient clonal mutations via molecular switches [157,158]. However, cancer can theoretically evolve to resist essentially any treatment. The ideal cancer therapy biologically speaking might be to pre-empt it entirely.
Dr. Aubrey de Grey proposed such a therapy, and called it “Whole-body Interdiction of Lengthening of Telomeres” (WILT) [159]. This TRM replacement-based delivery system could also help with WILT potentially. COURIER can mediate dual mRNA cargo delivery [104]. In this case, it could be two mRNA molecules that together encode a heterodimer of evolved designer-recombinases that excises hTERT from the genome of cells [160].
Patient stem cells would need to be periodically replaced for this to work. However, as mentioned before, the ability to do so may be crucial for correcting nuclear DNA mutations anyway - at least initially.
There is a possible version of WILT that can be implemented using “steward”, endosymbiont bacteria. This might be helpful because cancer can lengthen their telomeres using a mechanism called alternative lengthening of telomeres (ALT) as well as hTERT. However, ALT proteins are required for cellular processes other than telomere elongation [161]. A bacterial steward could theoretically “dole out” the ALT proteins as needed.
In the distant future, we almost certainly will be able to develop computerized microrobots and/or nanorobots that can enter our cells, scan their DNA, and eliminate mutated, oncogenic variants.
Conclusion
It would be extremely interesting to put the “garbage catastrophe theory of aging” to the test with organelle-sampling FluidFM - using single skeletal muscle cells (in large replicate) from elderly humans. If in vitro testing reveals lipofuscin accumulation to be a critical factor in the aging of such cells, we will have to eliminate it in situ or remove it from the body somehow. Through synthetic biology, I believe we can clear lipofuscin from our tissues.
Funding
Funding not received for the study.
Authors’ Contributions
M.R. wrote the paper.
Ethics Approval and Consent to Participate
N/A.
Consent for Publication
N/A.
Availability of Data and Material
N/A.
Acknowledgements
Thank you to my family and friends. The figures in this piece were created with BioRender.com.
Competing Interests
The author declares that he has no competing interests.
References
- Terman, A.; Brunk, U.T. Lipofuscin. Int. J. Biochem. Cell Biol. 2004, 36, 1400–1404. [Google Scholar] [CrossRef]
- Siebert, S.; Farrell, J.A.; Cazet, J.F.; Abeykoon, Y.; Primack, A.S.; Schnitzler, C.E.; Juliano, C.E. Stem Cell Differentiation Trajectories in Hydra Resolved at Single-Cell Resolution. Science 2019, 365, eaav9314. [Google Scholar] [CrossRef] [PubMed]
- Murad, R.; Macias-Muñoz, A.; Wong, A.; Ma, X.; Mortazavi, A. Coordinated Gene Expression and Chromatin Regulation during Hydra Head Regeneration. Genome Biol. Evol. 2021, 13, evab221. [Google Scholar] [CrossRef] [PubMed]
- Terman, A.; Brunk, U.T. Is aging the price for memory? Biogerontology 2005, 6, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Klapper, W.; Kühne, K.; Singh, K.K.; Heidorn, K.; Parwaresch, R.; Krupp, G. Longevity of lobsters is linked to ubiquitous telomerase expression. FEBS Lett. 1998, 439, 143–146. [Google Scholar] [CrossRef]
- Beltz, B.S.; Sandeman, D.C. Regulation of life-long neurogenesis in the decapod crustacean brain. Arthropod Struct. Dev. 2003, 32, 39–60. [Google Scholar] [CrossRef] [PubMed]
- Peregrim, I. Why we age—A new evolutionary view. Biologia 2017, 72, 475–485. [Google Scholar] [CrossRef]
- Sheehy, M.; Shelton, P.; Wickins, J.; Belchier, M.; Gaten, E. Ageing the European lobster Homarus gammarus by the lipofuscin in its eyestalk ganglia. Mar. Ecol. Prog. Ser. 1996, 143, 99–111. [Google Scholar] [CrossRef]
- Kakimoto, Y.; Okada, C.; Kawabe, N.; Sasaki, A.; Tsukamoto, H.; Nagao, R.; Osawa, M. Myocardial lipofuscin accumulation in ageing and sudden cardiac death. Sci. Rep. 2019, 9, 3304. [Google Scholar] [CrossRef]
- Karavanich, C.; Atema, J. Individual recognition and memory in lobster dominance. Anim. Behav. 1998, 56, 1553–1560. [Google Scholar] [CrossRef]
- Zhao, S.; Lin, L.; Kan, G.; Xu, C.; Tang, Q.; Yu, C.; Sun, W.; Cai, L.; Xu, C.; Cui, S. High autophagy in the naked mole rat may play a significant role in maintaining good health. Cell. Physiol. Biochem. 2014, 33, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Triplett, J.C.; Tramutola, A.; Swomley, A.; Kirk, J.; Grimes, K.; Lewis, K.; Orr, M.; Rodriguez, K.; Cai, J.; Klein, J.B.; et al. Age-related changes in the proteostasis network in the brain of the naked mole-rat: Implications promoting healthy longevity. Biochim. et Biophys. Acta (BBA) - Mol. Basis Dis. 2015, 1852, 2213–2224. [Google Scholar] [CrossRef] [PubMed]
- Hadi, F.; Kulaberoglu, Y.; Lazarus, K.A.; Bach, K.; Ugur, R.; Beattie, P.; Smith, E.S.J.; Khaled, W.T. Transformation of naked mole-rat cells. Nature 2020, 583, E1–E7. [Google Scholar] [CrossRef] [PubMed]
- Edrey, Y.H.; Hanes, M.; Pinto, M.; Mele, J.; Buffenstein, R. Successful aging and sustained good health in the naked mole rat: a long-lived mammalian model for biogerontology and biomedical. ILAR J. 2011, 52, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Panno, J.; Nair, K. Effects of increased lifespan on chromatin condensation in the adult male housefly. Mech. Ageing Dev. 1986, 35, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Clokey, G.V.; Jacobson, L.A. The autofluorescent “lipofuscin granules” in the intestinal cells of Caenorhabditis elegans are secondary lysosomes. Mech. Ageing Dev. 1986, 35, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Houthoofd, K.; Braeckman, B.P.; Lenaerts, I.; Brys, K.; De Vreese, A.; Van Eygen, S.; Vanfleteren, J.R. Ageing is reversed, and metabolism is reset to young levels in recovering dauer larvae of C. elegans. Exp. Gerontol. 2002, 37, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Goyal, V.K. Lipofuscin pigment accumulation in the central nervous system of the mouse during aging. Exp. Gerontol. 1982, 17, 89–94. [Google Scholar] [CrossRef]
- Gilissen, E.P.; Staneva-Dobrovski, L. Distinct Types of Lipofuscin Pigment in the Hippocampus and Cerebellum of Aged Cheirogaleid Primates. Anat. Rec. 2013, 296, 1895–1906. [Google Scholar] [CrossRef]
- Moreno-García, A.; Kun, A.; Calero, O.; Medina, M.; Calero, M. An Overview of the Role of Lipofuscin in Age-Related Neurodegeneration. Front. Neurosci. 2018, 12, 464. Available online: https://www.frontiersin.org/article/10.3389/fnins.2018.00464 (accessed on 31 March 2022). [CrossRef]
- Gray, D.A.; Woulfe, J. Lipofuscin and Aging: A Matter of Toxic Waste. Sci. Aging Knowl. Environ. 2005, 2005, re1. [Google Scholar] [CrossRef]
- Terman, A.; Brunk, U.T. Is Lipofuscin Eliminated from Cells? Investigative Ophthalmology & Visual Science 1999, 40, 2463–2464. [Google Scholar]
- Wang, L.; Xiao, C.-Y.; Li, J.-H.; Tang, G.-C.; Xiao, S.-S. Transport and Possible Outcome of Lipofuscin in Mouse Myocardium. Adv. Gerontol. 2022, 12, 247–263. [Google Scholar] [CrossRef]
- Burns, J.C.; Cotleur, B.; Walther, D.M.; Bajrami, B.; Rubino, S.J.; Wei, R.; Franchimont, N.; Cotman, S.L.; Ransohoff, R.M.; Mingueneau, M. Differential accumulation of storage bodies with aging defines discrete subsets of microglia in the healthy brain. eLife 2020, 9, e57495. [Google Scholar] [CrossRef] [PubMed]
- Boellaard, J.W.; Schlote, W. Ultrastructural Heterogeneity of Neuronal Lipofuscin in the Normal Human Cerebral Cortex. Acta Neuropathol. 1986, 71, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Sohal, R.S.; Wolfe, L.S. Chapter 11 Lipofuscin: characteristics and significance. In Progress in Brain Research; Swaab, D.F., Fliers, E., Mirmiran, M., Van Gool, W.A., Van Haaren, F., Eds.; Elsevier: Amsterdam, The Netherlands, 1986; Volume 70, pp. 171–183. [Google Scholar] [CrossRef]
- Sheehy, M.R.J. Individual variation in, and the effect of rearing temperature and body size on, the concentration of fluorescent morphological lipofuscin in the brains of freshwater crayfish, Cherax cuspidatus (Crustacea: Parastacidae). Comp. Biochem. Physiol. Part A: Physiol. 1990, 96, 281–286. [Google Scholar] [CrossRef]
- Brunk, U.T.; Terman, A. Lipofuscin: mechanisms of age-related accumulation and influence on cell function. Free Radic. Biol. Med. 2002, 33, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Kurz, T.; Terman, A.; Gustafsson, B.; Brunk, U.T. Lysosomes and oxidative stress in aging and apoptosis. Biochim. et Biophys. Acta (BBA) - Gen. Subj. 2008, 1780, 1291–1303. [Google Scholar] [CrossRef]
- Gabandé-Rodríguez, E.; Keane, L.; Capasso, M. Microglial phagocytosis in aging and Alzheimer's disease. J. Neurosci. Res. 2020, 98, 284–298. [Google Scholar] [CrossRef]
- Pan, C.; Banerjee, K.; Lehmann, G.L.; Almeida, D.; Hajjar, K.A.; Benedicto, I.; Jiang, Z.; Radu, R.A.; Thompson, D.H.; Rodriguez-Boulan, E.; et al. Lipofuscin causes atypical necroptosis through lysosomal membrane permeabilization. Proc. Natl. Acad. Sci. 2021, 118, e2100122118. [Google Scholar] [CrossRef]
- Terman, A.; Dalen, H.; Brunk, U.T. Ceroid/lipofuscin-loaded human fibroblasts show decreased survival time and diminished autophagocytosis during amino acid starvation☆. Exp. Gerontol. 1999, 34, 943–957. [Google Scholar] [CrossRef] [PubMed]
- Terman, A.; Abrahamsson, N.; Brunk, U.T. Ceroid/lipofuscin-loaded human fibroblasts show increased susceptibility to oxidative stress. Exp. Gerontol. 1999, 34, 755–770. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Jiang, H.; Xu, H.; Song, N.; Xie, J. Increased iron levels correlate with the selective nigral dopaminergic neuron degeneration in Parkinson’s disease. J. Neural Transm. 2011, 118, 361–369. [Google Scholar] [CrossRef]
- Maccarinelli, F.; Pagani, A.; Cozzi, A.; Codazzi, F.; Di Giacomo, G.; Capoccia, S.; Rapino, S.; Finazzi, D.; Politi, L.S.; Cirulli, F.; et al. A novel neuroferritinopathy mouse model (FTL 498InsTC) shows progressive brain iron dysregulation, morphological signs of early neurodegeneration and motor coordination deficits. Neurobiol. Dis. 2015, 81, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Bhoiwala, D.; Song, Y.; Cwanger, A.; et al. High iron diet causes elevation of retinal iron levels and RPE autofluorescence. Investigative Ophthalmology & Visual Science 2015, 56, 4203. [Google Scholar]
- Mangan, D. Iron: an underrated factor in aging. Aging 2021, 13, 23407–23415. [Google Scholar] [CrossRef] [PubMed]
- Ohgami, N.; Yajima, I.; Iida, M.; Li, X.; Oshino, R.; Kumasaka, M.Y.; Kato, M. Manganese-mediated acceleration of age-related hearing loss in mice. Sci. Rep. 2016, 6, 36306. [Google Scholar] [CrossRef] [PubMed]
- Höhn, A.; Grune, T. Lipofuscin: formation, effects and role of macroautophagy. Redox Biol. 2013, 1, 140–144. [Google Scholar] [CrossRef] [PubMed]
- von Zglinicki, T.; Nilsson, E.; Döcke, W.D.; Brunk, U.T. LIPOFUSCIN Lipofuscin accumulation and ageing of fibroblasts. Gerontology 1995, 41 (Suppl. 2), 95–108. [Google Scholar] [CrossRef]
- Tsakiri, E.N.; Iliaki, K.K.; Höhn, A.; Grimm, S.; Papassideri, I.S.; Grune, T.; Trougakos, I.P. Diet-derived advanced glycation end products or lipofuscin disrupts proteostasis and reduces life span in Drosophila melanogaster. Free. Radic. Biol. Med. 2013, 65, 1155–1163. [Google Scholar] [CrossRef]
- Seluanov, A.; Gladyshev, V.N.; Vijg, J.; Gorbunova, V. Mechanisms of cancer resistance in long-lived mammals. Nat. Rev. Cancer 2018, 18, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Samorajski, T.; Ordy, J.M.; Rady-Reimer, P. Lipofuscin pigment accumulationin the nervous system of aging mice. Anat. Rec. 1968, 160, 555–573. [Google Scholar] [CrossRef] [PubMed]
- Brizzee, K.R.; Johnson, F.A. Depth distribution of lipofuscin pigment in cerebral cortex of albino rat. Acta Neuropathol. 1970, 16, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Yanai, S.; Endo, S. Functional Aging in Male C57BL/6J Mice Across the Life-Span: A Systematic Behavioral Analysis of Motor, Emotional, and Memory Function to Define an Aging Phenotype. Front. Aging Neurosci. 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Lutz, C.M.; Osborne, M.A.; Anisimov, S.V.; Donnelly, C.J.; Grima, J.C.; Sattler, R.; Monani, U.R.; De Vivo, D.C.; Sturrock, A.; Leavitt, B.R.; et al. Optimizing mouse models of neurodegenerative disorders: are therapeutics in sight? Futur. Neurol. 2014, 9, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Double, K.L.; Dedov, V.N.; Fedorow, H.; Kettle, E.; Halliday, G.M.; Garner, B.; Brunk, U.T. The comparative biology of neuromelanin and lipofuscin in the human brain. Cell. Mol. Life Sci. 2008, 65, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.M.; Yates, P.O.; Stamp, J.E. The relationship between lipofuscin pigment and ageing in the human nervous system. J. Neurol. Sci. 1978, 37, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Goyal, V.K. Lipofuscin pigment accumulation in human brain during aging. Exp. Gerontol. 1982, 17, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Benavides, S.H.; Monserrat, A.J.; Fariña, S.; et al. Sequential histochemical studies of neuronal lipofuscin in human cerebral cortex from the first to the ninth decade of life. Arch. Gerontol. Geriatr. 2002, 34, 219–231. [Google Scholar] [CrossRef]
- Yin, D. Biochemical basis of lipofuscin, ceroid, and age pigment-like fluorophores. Free. Radic. Biol. Med. 1996, 21, 871–888. [Google Scholar] [CrossRef]
- Wing, G.L.; Blanchard, G.C.; Weiter, J.J. The topography and age relationship of lipofuscin concentration in the retinal pigment epithelium. Investigative Ophthalmology & Visual Science 1978, 17, 601–607. [Google Scholar]
- Dayan, D.; Abrahami, I.; Buchner, A.; Gorsky, M.; Chimovitz, N. Lipid pigment (lipofuscin) in human perioral muscles with aging. Exp. Gerontol. 1988, 23, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ren, D. Lysosomal Physiology. Annu. Rev. Physiol. 2015, 77, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-K.; Min, B.; Eom, J.; Park, J.S. Different phases of aging in mouse old skeletal muscle. Aging 2022, 14, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Vida, C.; de Toda, I.M.; Cruces, J.; Garrido, A.; Gonzalez-Sanchez, M.; De la Fuente, M. Role of macrophages in age-related oxidative stress and lipofuscin accumulation in mice. Redox Biol. 2017, 12, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Furber, J.D. Extracellular glycation crosslinks: prospects for removal. Rejuvenation Res. 2006, 9, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Settembre, C.; Di Malta, C.; Polito, V.A.; Garcia Arencibia, M.; Vetrini, F.; Erdin, S.; Erdin, S.U.; Huynh, T.; Medina, D.; Colella, P.; et al. TFEB Links Autophagy to Lysosomal Biogenesis. Science 2011, 332, 1429–1433. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, R.; Carrera, I.; Xu, S.; Lakshmana, M.K. TFEB Overexpression in the P301S Model of Tauopathy Mitigates Increased PHF1 Levels and Lipofuscin Puncta and Rescues Memory Deficits. eNeuro 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, J.K.; Andrews, N.W.; Simon, S.M. Membrane proximal lysosomes are the major vesicles responsible for calcium-dependent exocytosis in nonsecretory cells. J. Cell Biol. 2002, 159, 625–635. [Google Scholar] [CrossRef]
- Medina, D.L.; Fraldi, A.; Bouche, V.; Annunziata, F.; Mansueto, G.; Spampanato, C.; Puri, C.; Pignata, A.; Martina, J.A.; Sardiello, M.; et al. Transcriptional Activation of Lysosomal Exocytosis Promotes Cellular Clearance. Dev. Cell 2011, 21, 421–430. [Google Scholar] [CrossRef]
- Julien, S.; Schraermeyer, U. Lipofuscin can be eliminated from the retinal pigment epithelium of monkeys. Neurobiol. Aging 2012, 33, 2390–2397. [Google Scholar] [CrossRef]
- Gäbelein, C.G.; Feng, Q.; Sarajlic, E.; Zambelli, T.; Guillaume-Gentil, O.; Kornmann, B.; Vorholt, J.A. Mitochondria transplantation between living cells. PLOS Biol. 2022, 20, e3001576. [Google Scholar] [CrossRef]
- Beregi, E.; Regius, O.; Hüttl, T.; Göbl, Z. Age-related changes in the skeletal muscle cells. Z Gerontol 1988, 21, 83–6. [Google Scholar]
- Hütter, E.; Skovbro, M.; Lener, B.; Prats, C.; Rabøl, R.; Dela, F.; Jansen-Dürr, P. Oxidative stress and mitochondrial impairment can be separated from lipofuscin accumulation in aged human skeletal muscle. Aging Cell 2007, 6, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Triolo, M.; Hood, D.A. Manifestations of Age on Autophagy, Mitophagy and Lysosomes in Skeletal Muscle. Cells 2021, 10, 1054. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Cheng, S.; Huang, F.; Fan, W.; Chen, Y.; Shi, H.; He, H. Mitochondrial dysfunction in long-term neuronal cultures mimics changes with aging. Med Sci. Monit. 2011, 17, BR91–BR96. [Google Scholar] [CrossRef]
- Moreno-Blas, D.; Gorostieta-Salas, E.; Pommer-Alba, A.; Muciño-Hernández, G.; Gerónimo-Olvera, C.; Maciel-Barón, L.A.; Konigsberg, M.; Massieu, L.; Castro-Obregón, S. Cortical neurons develop a senescence-like phenotype promoted by dysfunctional autophagy. Aging 2019, 11, 6175–6198. [Google Scholar] [CrossRef] [PubMed]
- Li, Z. 5.43—In Vitro Micro-Tissue and -Organ Models for Toxicity Testing. In Comprehensive Biotechnology (Second Edition); Moo-Young, M., Ed.; Academic Press: Burlington, 2011; pp. 551–563. [Google Scholar] [CrossRef]
- Potter, S.M.; DeMarse, T.B. A new approach to neural cell culture for long-term studies. J. Neurosci. Methods 2001, 110, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.; Kraner-Scheiber, S.; Petersen, B.; Rieblinger, B.; Buermann, A.; Flisikowska, T.; Flisikowski, K.; Christan, S.; Edlinger, M.; Baars, W.; et al. Efficient production of multi-modified pigs for xenotransplantation by ‘combineering’, gene stacking and gene editing. Sci. Rep. 2016, 6, 29081. [Google Scholar] [CrossRef]
- Kimura, T.; Jia, J.; Kumar, S.; Choi, S.W.; Gu, Y.; Mudd, M.; Dupont, N.; Jiang, S.; Peters, R.; Farzam, F.; et al. Dedicated SNARE s and specialized TRIM cargo receptors mediate secretory autophagy. EMBO J. 2017, 36, 42–60. [Google Scholar] [CrossRef]
- Scheller, L.; Strittmatter, T.; Fuchs, D.; Bojar, D.; Fussenegger, M. Generalized extracellular molecule sensor platform for programming cellular behavior. Nat. Chem. Biol. 2018, 14, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Kim, B.; Byun, J.-W.; Baik, S.H.; Huh, Y.H.; Kim, J.-H.; Mook-Jung, I.; Song, W.K.; Shin, J.-H.; Seo, H.; et al. LRRK2 G2019S mutation attenuates microglial motility by inhibiting focal adhesion kinase. Nat. Commun. 2015, 6, 8255. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, G.B.; Cunningham, P.S.; Poolman, T.M.; Iqbal, M.; Maidstone, R.; Baxter, M.; Bagnall, J.; Begley, N.; Saer, B.; Hussell, T.; et al. The clock gene Bmal1 inhibits macrophage motility, phagocytosis, and impairs defense against pneumonia. Proc. Natl. Acad. Sci. USA 2020, 117, 1543–1551. [Google Scholar] [CrossRef] [PubMed]
- Russo, I.; Bubacco, L.; Greggio, E. LRRK2 as a target for modulating immune system responses. Neurobiol. Dis. 2022, 169, 105724. [Google Scholar] [CrossRef] [PubMed]
- Insall, R.H.; Paschke, P.; Tweedy, L. Steering yourself by the bootstraps: how cells create their own gradients for chemotaxis. Trends Cell Biol. 2022, 32, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Tigges, M.; Marquez-Lago, T.T.; Stelling, J.; Fussenegger, M. A tunable synthetic mammalian oscillator. Nature 2009, 457, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.W.S.; Lu, G.; Dong, H.; Cho, Y.-L.; Natalia, A.; Wang, L.; Chan, C.; Kappei, D.; Taneja, R.; Ling, S.-C.; et al. A degradative to secretory autophagy switch mediates mitochondria clearance in the absence of the mATG8-conjugation machinery. Nat. Commun. 2022, 13, 3720. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, M.A.; Williamson, A.P.; Steinbach, A.M.; Roberts, E.W.; Kern, N.; Headley, M.B.; Vale, R.D. Chimeric antigen receptors that trigger phagocytosis. eLife 2018, 7, e36688. [Google Scholar] [CrossRef]
- Birgisdottir, Å.B.; Lamark, T.; Johansen, T. The LIR motif – crucial for selective autophagy. J. Cell Sci. 2013, 126 Pt 15, 3237–3247. [Google Scholar] [CrossRef]
- Uematsu, M.; Nishimura, T.; Sakamaki, Y.; Yamamoto, H.; Mizushima, N. Accumulation of undegraded autophagosomes by expression of dominant-negative STX17 (syntaxin 17) mutants. Autophagy 2017, 13, 1452–1464. [Google Scholar] [CrossRef]
- Friedland, A.E.; Lu, T.K.; Wang, X.; Shi, D.; Church, G.; Collins, J.J. Synthetic Gene Networks that Count. Science 2009, 324, 1199–1202. [Google Scholar] [CrossRef] [PubMed]
- VanHook, A.M. Macrophages don’t take more than they can eat. Sci. Signal. 2017, 10, eaao1183. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Rhau, B.; Hermann, A.; McNally, K.A.; Zhou, C.; Gong, D.; Weiner, O.D.; Conklin, B.R.; Onuffer, J.; Lim, W.A. Synthetic control of mammalian-cell motility by engineering chemotaxis to an orthogonal bioinert chemical signal. Proc. Natl. Acad. Sci. USA 2014, 111, 5896–5901. [Google Scholar] [CrossRef] [PubMed]
- Nagai, Y.; Miyakawa, N.; Takuwa, H.; Hori, Y.; Oyama, K.; Ji, B.; Takahashi, M.; Huang, X.-P.; Slocum, S.T.; DiBerto, J.F.; et al. Deschloroclozapine, a potent and selective chemogenetic actuator enables rapid neuronal and behavioral modulations in mice and monkeys. Nat. Neurosci. 2020, 23, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Ronda, C.; Chen, S.P.; Cabral, V.; Yaung, S.J.; Wang, H.H. Metagenomic engineering of the mammalian gut microbiome in situ. Nat. Methods 2019, 16, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Maxwell, K.G.; Wang, K.; Bowers, D.T.; Flanders, J.A.; Liu, W.; Wang, L.-H.; Liu, Q.; Liu, C.; Naji, A.; et al. A nanofibrous encapsulation device for safe delivery of insulin-producing cells to treat type 1 diabetes. Sci. Transl. Med. 2021, 13, eabb4601. [Google Scholar] [CrossRef] [PubMed]
- Spampanato, C.; Feeney, E.; Li, L.; Cardone, M.; Lim, J.-A.; Annunziata, F.; Zare, H.; Polishchuk, R.; Puertollano, R.; Parenti, G.; et al. Transcription factor EB (TFEB) is a new therapeutic target for Pompe disease. EMBO Mol. Med. 2013, 5, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Sung, C.Y.W.; Hayase, N.; Yuen, P.S.T.; Lee, J.; Fernandez, K.; Hu, X.; et al. Macrophage Depletion Protects Against Cisplatin-Induced Ototoxicity and Nephrotoxicity. bioRxiv 2023. [Google Scholar] [CrossRef]
- Lund, H.; Pieber, M.; Parsa, R.; Han, J.; Grommisch, D.; Ewing, E.; Kular, L.; Needhamsen, M.; Espinosa, A.; Nilsson, E.; et al. Competitive repopulation of an empty microglial niche yields functionally distinct subsets of microglia-like cells. Nat. Commun. 2018, 9, 4845. [Google Scholar] [CrossRef]
- Gustafsson, K.; Rhee, C.; Frodermann, V.; Scadden, E.W.; Li, D.; Iwamoto, Y.; Palchaudhuri, R.; Hyzy, S.L.; Boitano, A.E.; Nahrendorf, M.; et al. Clearing and replacing tissue-resident myeloid cells with an anti-CD45 antibody–drug conjugate. Blood Adv. 2023, 7, 6964–6973. [Google Scholar] [CrossRef]
- Butowski, N.; Colman, H.; De Groot, J.F.; Omuro, A.M.; Nayak, L.; Wen, P.Y.; Cloughesy, T.F.; Marimuthu, A.; Haidar, S.; Perry, A.; et al. Orally administered colony stimulating factor 1 receptor inhibitor PLX3397 in recurrent glioblastoma: an Ivy Foundation Early Phase Clinical Trials Consortium phase II study. Neuro-Oncology 2016, 18, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Green, K.N.; Crapser, J.D.; Hohsfield, L.A. To Kill Microglia: A Case for CSF1R Inhibitors. Trends Immunol. 2020, 41, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-C.; Gil-Martin, M.; Bauer, T.M.; Naing, A.; Lim, D.W.-T.; Sarantopoulos, J.; Geva, R.; Ando, Y.; Fan, L.; Choudhury, S.; et al. Abstract CT171: Phase I study of BLZ945 alone and with spartalizumab (PDR001) in patients (pts) with advanced solid tumors. Cancer Res 2020, 80, CT171. [Google Scholar] [CrossRef]
- Krance, R.A.; Kuehnle, I.; Rill, D.R.; Mei, Z.; Pinetta, C.; Evans, W.; Brown, M.P.; Pulé, M.; Heslop, H.E.; Brenner, M.K. Hematopoietic and immunomodulatory effects of lytic CD45 monoclonal antibodies in patients with hematologic malignancy. Biol. Blood Marrow Transplant. 2003, 9, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Rafiei Hashtchin, A.; Manstein, F.; Carvalho Oliveira, M.; Kempf, H.; Zweigerdt, R.; et al. Continuous human iPSC-macrophage production by suspension culture in stirred tank bioreactors. Nat. Protoc. 2022, 17, 513–539. [Google Scholar] [CrossRef] [PubMed]
- Fattorelli, N.; Martinez-Muriana, A.; Wolfs, L.; Geric, I.; De Strooper, B.; Mancuso, R. Stem-cell-derived human microglia transplanted into mouse brain to study human disease. Nat. Protoc. 2021, 16, 1013–1033. [Google Scholar] [CrossRef]
- Cao, X.; Yakala, G.K.; van den Hil, F.E.; Cochrane, A.; Mummery, C.L.; Orlova, V.V. Differentiation and Functional Comparison of Monocytes and Macrophages from hiPSCs with Peripheral Blood Derivatives. Stem Cell Rep. 2019, 12, 1282–1297. [Google Scholar] [CrossRef] [PubMed]
- Dick, S.A.; Wong, A.; Hamidzada, H.; Nejat, S.; Nechanitzky, R.; Vohra, S.; Mueller, B.; Zaman, R.; Kantores, C.; Aronoff, L.; et al. Three tissue resident macrophage subsets coexist across organs with conserved origins and life cycles. Sci. Immunol. 2022, 7, eabf7777. [Google Scholar] [CrossRef] [PubMed]
- Palchaudhuri, R.; Saez, B.; Hoggatt, J.; Schajnovitz, A.; Sykes, D.B.; Tate, T.A.; Czechowicz, A.; Kfoury, Y.; Ruchika, F.; Rossi, D.J.; et al. Non-genotoxic conditioning for hematopoietic stem cell transplantation using a hematopoietic-cell-specific internalizing immunotoxin. Nat. Biotechnol. 2016, 34, 738–745. [Google Scholar] [CrossRef]
- Breda, L.; Papp, T.E.; Triebwasser, M.P.; Yadegari, A.; Fedorky, M.T.; Tanaka, N.; Abdulmalik, O.; Pavani, G.; Wang, Y.; Grupp, S.A.; et al. In vivo hematopoietic stem cell modification by mRNA delivery. Science 2023, 381, 436–443. [Google Scholar] [CrossRef]
- Wellhausen, N.; O’Connell, R.P.; Lesch, S.; Engel, N.W.; Rennels, A.K.; Gonzales, D.; Herbst, F.; Young, R.M.; Garcia, K.C.; Weiner, D.; et al. Epitope base editing CD45 in hematopoietic cells enables universal blood cancer immune therapy. Sci. Transl. Med. 2023, 15, eadi1145. [Google Scholar] [CrossRef] [PubMed]
- Horns, F.; Martinez, J.A.; Fan, C.; Haque, M.; Linton, J.M.; Tobin, V.; Santat, L.; Maggiolo, A.O.; Bjorkman, P.J.; Lois, C.; et al. Engineering RNA export for measurement and manipulation of living cells. Cell 2023, 186, 3642–3658.e32. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, C.T.; Homma, S.; Suchy, F.; Wang, S.; Bhadhury, J.; Amaya, A.K.; et al. Secreted Particle Information Transfer (SPIT)—A Cellular Platform for In Vivo Genetic Engineering. bioRxiv 2024. [Google Scholar] [CrossRef]
- Mc Cafferty, S.; De Temmerman, J.; Kitada, T.; Becraft, J.R.; Weiss, R.; Irvine, D.J.; Devreese, M.; De Baere, S.; Combes, F.; Sanders, N.N. In Vivo Validation of a Reversible Small Molecule-Based Switch for Synthetic Self-Amplifying mRNA Regulation. Mol. Ther. 2021, 29, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Perkovic, M.; Gawletta, S.; Hempel, T.; Brill, S.; Nett, E.; Sahin, U.; Beissert, T. A trans-amplifying RNA simplified to essential elements is highly replicative and robustly immunogenic in mice. Mol. Ther. 2023, 31, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.L.; Moss, B. Infectious poxvirus vectors have capacity for at least 25,000 base pairs of foreign DNA. Gene 1983, 25, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.E.; Oggier, A.; Füglistaler, A.; Camviel, N.; Hijazi, M.; Villarreal, A.R.; Arber, C.; Barth, P. Computational design of dynamic receptor—peptide signaling complexes applied to chemotaxis. Nat. Commun. 2023, 14, 2875. [Google Scholar] [CrossRef]
- Saberi, A.; Gulyaeva, A.A.; Brubacher, J.L.; Newmark, P.A.; Gorbalenya, A.E. A planarian nidovirus expands the limits of RNA genome size. PLOS Pathog. 2018, 14, e1007314. [Google Scholar] [CrossRef]
- Zhu, J.; Batra, H.; Ananthaswamy, N.; Mahalingam, M.; Tao, P.; Wu, X.; Guo, W.; Fokine, A.; Rao, V.B. Design of bacteriophage T4-based artificial viral vectors for human genome remodeling. Nat. Commun. 2023, 14, 2928. [Google Scholar] [CrossRef]
- Hohsfield, L.A.; Najafi, A.R.; Ghorbanian, Y.; Soni, N.; Hingco, E.E.; Kim, S.J.; Jue, A.D.; Swarup, V.; Inlay, M.A.; Green, K.N. Effects of long-term and brain-wide colonization of peripheral bone marrow-derived myeloid cells in the CNS. J. Neuroinflammation 2020, 17, 279. [Google Scholar] [CrossRef]
- Rodriguez, P.L.; Harada, T.; Christian, D.A.; Pantano, D.A.; Tsai, R.K.; Discher, D.E. Minimal “Self” peptides that inhibit phagocytic clearance and enhance delivery of nanoparticles. Science 2013, 339, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Hickey, A.M.; Bhaskar, U.; Linhardt, R.J.; Dordick, J.S. Effect of eliminase gene (elmA) deletion on heparosan production and shedding in Escherichia coli K5. J. Biotechnol. 2013, 165, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.-W.; Liu, Y.; Lee, Y.-Q.; Yang, P.-J.; Ho, C.-T.; Hong, J.-C.; Hsiao, J.-C.; Liao, D.-C.; Liang, A.-J.; Hung, T.-C.; et al. Construction of intracellular asymmetry and asymmetric division in Escherichia coli. Nat. Commun. 2021, 12, 888. [Google Scholar] [CrossRef]
- Szczesna, M.; Huang, Y.; Lacoursiere, R.E.; Bonini, F.; Pol, V.; Koc, F.; et al. Dedicated bacterial esterases reverse lipopolysaccharide ubiquitylation to block immune sensing. Res Sq 2023, rs.3.rs-2986327. [Google Scholar] [CrossRef]
- Blakney, A.K.; McKay, P.F.; Bouton, C.R.; Hu, K.; Samnuan, K.; Shattock, R.J. Innate Inhibiting Proteins Enhance Expression and Immunogenicity of Self-Amplifying RNA. Mol. Ther. 2021, 29, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- Ferdosi, S.R.; Ewaisha, R.; Moghadam, F.; Krishna, S.; Park, J.G.; Ebrahimkhani, M.R.; Kiani, S.; Anderson, K.S. Multifunctional CRISPR-Cas9 with engineered immunosilenced human T cell epitopes. Nat. Commun. 2019, 10, 1842. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Muhuri, M.; Li, S.; Qin, W.; Xu, G.; Luo, L.; Li, J.; Letizia, A.J.; Wang, S.K.; Chan, Y.K.; et al. Circumventing cellular immunity by miR142-mediated regulation sufficiently supports rAAV-delivered OVA expression without activating humoral immunity. JCI Insight 4, e99052. [CrossRef] [PubMed]
- Yoo, J.-S.; Sasaki, M.; Cho, S.X.; Kasuga, Y.; Zhu, B.; Ouda, R.; Orba, Y.; de Figueiredo, P.; Sawa, H.; Kobayashi, K.S. SARS-CoV-2 inhibits induction of the MHC class I pathway by targeting the STAT1-IRF1-NLRC5 axis. Nat. Commun. 2021, 12, 6602. [Google Scholar] [CrossRef] [PubMed]
- Koga, H.; Martinez-Vicente, M.; Macian, F.; Verkhusha, V.V.; Cuervo, A.M. A photoconvertible fluorescent reporter to track chaperone-mediated autophagy. Nat. Commun. 2011, 2, 386. [Google Scholar] [CrossRef]
- Lonzarić, J.; Lebar, T.; Majerle, A.; Manček-Keber, M.; Jerala, R. Locked and proteolysis-based transcription activator-like effector (TALE) regulation. Nucleic Acids Res. 2016, 44, 1471–1481. [Google Scholar] [CrossRef]
- Slabaugh, E.; Brandizzi, F. Membrane-tethered transcription factors provide a connection between stress response and developmental pathways. Plant Signal. Behav. 2011, 6, 1210–1211. [Google Scholar] [CrossRef] [PubMed]
- Rubens, J.R.; Selvaggio, G.; Lu, T.K. Synthetic mixed-signal computation in living cells. Nat. Commun. 2016, 7, 11658. [Google Scholar] [CrossRef] [PubMed]
- Streeter, M.D.; Rowan, S.; Ray, J.; McDonald, D.M.; Volkin, J.; Clark, J.; Taylor, A.; Spiegel, D.A. Generation and Characterization of Anti-Glucosepane Antibodies Enabling Direct Detection of Glucosepane in Retinal Tissue. ACS Chem. Biol. 2020, 15, 2655–2661. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally occurring p16Ink4a-positive cells shorten healthy lifespan. Nature 2016, 530, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. Send in the senolytics. Nat. Biotechnol. 2020, 38, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Amor, C.; Feucht, J.; Leibold, J.; Ho, Y.-J.; Zhu, C.; Alonso-Curbelo, D.; Mansilla-Soto, J.; Boyer, J.A.; Li, X.; Giavridis, T.; et al. Senolytic CAR T cells reverse senescence-associated pathologies. Nature 2020, 583, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Yang, D.; Sun, B.; Sun, B.; Li, S.; Li, S.; Wei, W.; Wei, W.; Liu, X.; Liu, X.; et al. NKG2D-CAR T cells eliminate senescent cells in aged mice and nonhuman primates. Sci. Transl. Med. 2023, 15, eadd1951. [Google Scholar] [CrossRef]
- VitaDAO - ApoptoSENS - Senolytic CAR-NK Cells. Available online: https://www.vitadao.com/projects/apoptosens-senolytic-car-nk-cells (accessed on 3 April 2024).
- Kaseniit, K.E.; Katz, N.; Kolber, N.S.; Call, C.C.; Wengier, D.L.; Cody, W.B.; Sattely, E.S.; Gao, X.J. Modular, programmable RNA sensing using ADAR editing in living cells. Nat. Biotechnol. 2023, 41, 482–487. [Google Scholar] [CrossRef]
- Gayet, R.V.; Ilia, K.; Razavi, S.; Tippens, N.D.; Lalwani, M.A.; Zhang, K.; Chen, J.X.; Chen, J.C.; Vargas-Asencio, J.; Collins, J.J. Autocatalytic base editing for RNA-responsive translational control. Nat. Commun. 2023, 14, 1339. [Google Scholar] [CrossRef]
- Bernardes de Jesus, B.; Vera, E.; Schneeberger, K.; Tejera, A.M.; Ayuso, E.; Bosch, F.; et al. Telomerase gene therapy in adult and old mice delays aging and increases longevity without increasing cancer. EMBO Mol. Med. 2012, 4, 691–704. [Google Scholar] [CrossRef]
- Jaijyan, D.K.; Selariu, A.; Cruz-Cosme, R.; Tong, M.; Yang, S.; Stefa, A.; Kekich, D.; Sadoshima, J.; Herbig, U.; Tang, Q.; et al. New intranasal and injectable gene therapy for healthy life extension. Proc. Natl. Acad. Sci. USA 2022, 119, e2121499119. [Google Scholar] [CrossRef] [PubMed]
- Sewell, P.E. Systemic Human Htert Aav Gene Transfer Therapy And The Effect On Telomere Length And Biological Age, A Case Report. J. Regen. Biol. Med. 2022. [Google Scholar] [CrossRef]
- Ocampo, A.; Reddy, P.; Martinez-Redondo, P.; Platero-Luengo, A.; Hatanaka, F.; Hishida, T.; Li, M.; Lam, D.; Kurita, M.; Beyret, E.; et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell 2016, 167, 1719–1733. [Google Scholar] [CrossRef] [PubMed]
- Decressac, M.; Mattsson, B.; Weikop, P.; Lundblad, M.; Jakobsson, J.; Björklund, A. TFEB-mediated autophagy rescues midbrain dopamine neurons from α-synuclein toxicity. Proc. Natl. Acad. Sci. USA 2013, 110, E1817–E1826. [Google Scholar] [CrossRef] [PubMed]
- Bredenkamp, N.; Nowell, C.S.; Blackburn, C.C. Regeneration of the aged thymus by a single transcription factor. Development 2014, 141, 1627–1637. [Google Scholar] [CrossRef] [PubMed]
- García-Cao, I.; García-Cao, M.; Martín-Caballero, J.; Criado, L.M.; Klatt, P.; Flores, J.M.; Weill, J.-C.; Blasco, M.A.; Serrano, M. 'Super p53' mice exhibit enhanced DNA damage response, are tumor resistant and age normally. EMBO J. 2002, 21, 6225–6235. [Google Scholar] [CrossRef] [PubMed]
- Matheu, A.; Maraver, A.; Klatt, P.; Flores, I.; Garcia-Cao, I.; Borras, C.; Flores, J.M.; Viña, J.; Blasco, M.A.; Serrano, M. Delayed ageing through damage protection by the Arf/p53 pathway. Nature 2007, 448, 375–379. [Google Scholar] [CrossRef]
- Zhao, Y.; Burikhanov, R.; Qiu, S.; Lele, S.M.; Jennings, C.D.; Bondada, S.; Spear, B.; Rangnekar, V.M. Cancer resistance in transgenic mice expressing the SAC module of Par-4. Cancer Res 2007, 67, 9276–9285. [Google Scholar] [CrossRef]
- Garcia-Cao, I.; Song, M.S.; Hobbs, R.M.; Laurent, G.; Giorgi, C.; de Boer, V.C.; Anastasiou, D.; Ito, K.; Sasaki, A.T.; Rameh, L.; et al. Systemic Elevation of PTEN Induces a Tumor-Suppressive Metabolic State. Cell 2012, 149, 49–62. [Google Scholar] [CrossRef]
- Bujarrabal-Dueso, A.; Sendtner, G.; Meyer, D.H.; Chatzinikolaou, G.; Stratigi, K.; Garinis, G.A.; Schumacher, B. The DREAM complex functions as conserved master regulator of somatic DNA-repair capacities. Nat. Struct. Mol. Biol. 2023, 30, 475–488. [Google Scholar] [CrossRef]
- Barroca, V.; Fouchet, P. Germline Stem Cells: The First Guards of Heredity. Cell Stem Cell 2008, 2, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Wang, B.; Zhu, Y.; Wu, J.; Qiu, L.; Ling, S.; Zhou, Z.; Dai, Y.; Zhong, Z.; Zheng, Y. Female germline stem cells: aging and anti-aging. J. Ovarian Res. 2022, 15, 79. [Google Scholar] [CrossRef] [PubMed]
- Stringer, J.M.; Winship, A.; Zerafa, N.; Wakefield, M.; Hutt, K. Oocytes can efficiently repair DNA double-strand breaks to restore genetic integrity and protect offspring health. Proc. Natl. Acad. Sci. 2020, 117, 11513–11522. [Google Scholar] [CrossRef]
- St. John, J.C. Mitochondria and Female Germline Stem Cells—A Mitochondrial DNA Perspective. Cells 2019, 8, 852. [Google Scholar] [CrossRef] [PubMed]
- Lechuga-Vieco, A.V.; Latorre-Pellicer, A.; Calvo, E.; Torroja, C.; Pellico, J.; Acín-Pérez, R.; García-Gil, M.L.; Santos, A.; Bagwan, N.; Bonzon-Kulichenko, E.; et al. Heteroplasmy of Wild-Type Mitochondrial DNA Variants in Mice Causes Metabolic Heart Disease With Pulmonary Hypertension and Frailty. Circ. 2022, 145, 1084–1101. [Google Scholar] [CrossRef] [PubMed]
- Latorre-Pellicer, A.; Moreno-Loshuertos, R.; Lechuga-Vieco, A.V.; Sánchez-Cabo, F.; Torroja, C.; Acín-Pérez, R.; Calvo, E.; Aix, E.; González-Guerra, A.; Logan, A.; et al. Mitochondrial and nuclear DNA matching shapes metabolism and healthy ageing. Nature 2016, 535, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Zealley, B.; de Grey, A.D.N.J. Strategies for Engineered Negligible Senescence. Gerontology 2013, 59, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.G.; Koob, M.D. Transformation of isolated mammalian mitochondria by bacterial conjugation. Nucleic Acids Res. 2005, 33, e139. [Google Scholar] [CrossRef]
- Zekonyte, U.; Bacman, S.R.; Smith, J.; Shoop, W.; Pereira, C.V.; Tomberlin, G.; Stewart, J.; Jantz, D.; Moraes, C.T. Mitochondrial targeted meganuclease as a platform to eliminate mutant mtDNA in vivo. Nat. Commun. 2021, 12, 3210. [Google Scholar] [CrossRef]
- Sato, T.K.; Kawano, S.; Endo, T. Role of the membrane potential in mitochondrial protein unfolding and import. Sci. Rep. 2019, 9, 7637. [Google Scholar] [CrossRef]
- Abascal, F.; Harvey, L.M.R.; Mitchell, E.; Lawson, A.R.J.; Lensing, S.V.; Ellis, P.; Russell, A.J.C.; Alcantara, R.E.; Baez-Ortega, A.; Wang, Y.; et al. Somatic mutation landscapes at single-molecule resolution. Nature 2021, 593, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Cortese, F.A.B.; Santostasi, G. Whole-Body Induced Cell Turnover: A Proposed Intervention for Age-Related Damage and Associated Pathology. Rejuvenation Res. 2016, 19, 322–336. [Google Scholar] [CrossRef] [PubMed]
- Scheller, L.; Strittmatter, T.; Fuchs, D.; Bojar, D.; Fussenegger, M. Generalized extracellular molecule sensor platform for programming cellular behavior. Nat. Chem. Biol. 2018, 14, 723–729. [Google Scholar] [CrossRef]
- Renteln, M.A. Promoting Oncolytic Vector Replication with Switches that Detect Ubiquitous Mutations. Curr. Cancer Ther. Rev. 2024, 20, 40–52. [Google Scholar] [CrossRef]
- Renteln, M. A New Paradigm in Cancer Treatment: Identifying and Targeting Clonal Mutations 2024. [CrossRef]
- de Grey, A.D. Whole-body interdiction of lengthening of telomeres: a proposal for cancer prevention. Front. Biosci. 2005, 10, 2420–9. [Google Scholar] [CrossRef] [PubMed]
- Lansing, F.; Paszkowski-Rogacz, M.; Schmitt, L.T.; Schneider, P.M.; Romanos, T.R.; Sonntag, J.; Buchholz, F. A heterodimer of evolved designer-recombinases precisely excises a human genomic DNA locus. Nucleic Acids Res. 2020, 48, 472–485. [Google Scholar] [CrossRef]
- Sommer, A.; Royle, N.J. ALT: A Multi-Faceted Phenomenon. Genes 2020, 11, 133. [Google Scholar] [CrossRef]
Figure 1.
(A) Single aged skeletal muscle cells are injected via the FluidFM needle with MNPs conjugated to nanobodies against LAMP-1 and perhaps new lysosomes with sterically-blocked LAMP-1. (B) The nanobodies conjugated to MNPs find their LAMP-1 targets on the surface of pre-existing lysosomes. (C) Magnetism is used to cluster at least the majority of the pre-existing lysosomes in one area of the cell (i.e., near the needle tip - opposite the region where new lysosomes were injected). (D) The needle is used to aspirate the indigestible garbage-filled lysosomes. (E) Another set of new lysosomes are injected into the cell where the pre-existing lysosomes were extracted.
Figure 1.
(A) Single aged skeletal muscle cells are injected via the FluidFM needle with MNPs conjugated to nanobodies against LAMP-1 and perhaps new lysosomes with sterically-blocked LAMP-1. (B) The nanobodies conjugated to MNPs find their LAMP-1 targets on the surface of pre-existing lysosomes. (C) Magnetism is used to cluster at least the majority of the pre-existing lysosomes in one area of the cell (i.e., near the needle tip - opposite the region where new lysosomes were injected). (D) The needle is used to aspirate the indigestible garbage-filled lysosomes. (E) Another set of new lysosomes are injected into the cell where the pre-existing lysosomes were extracted.
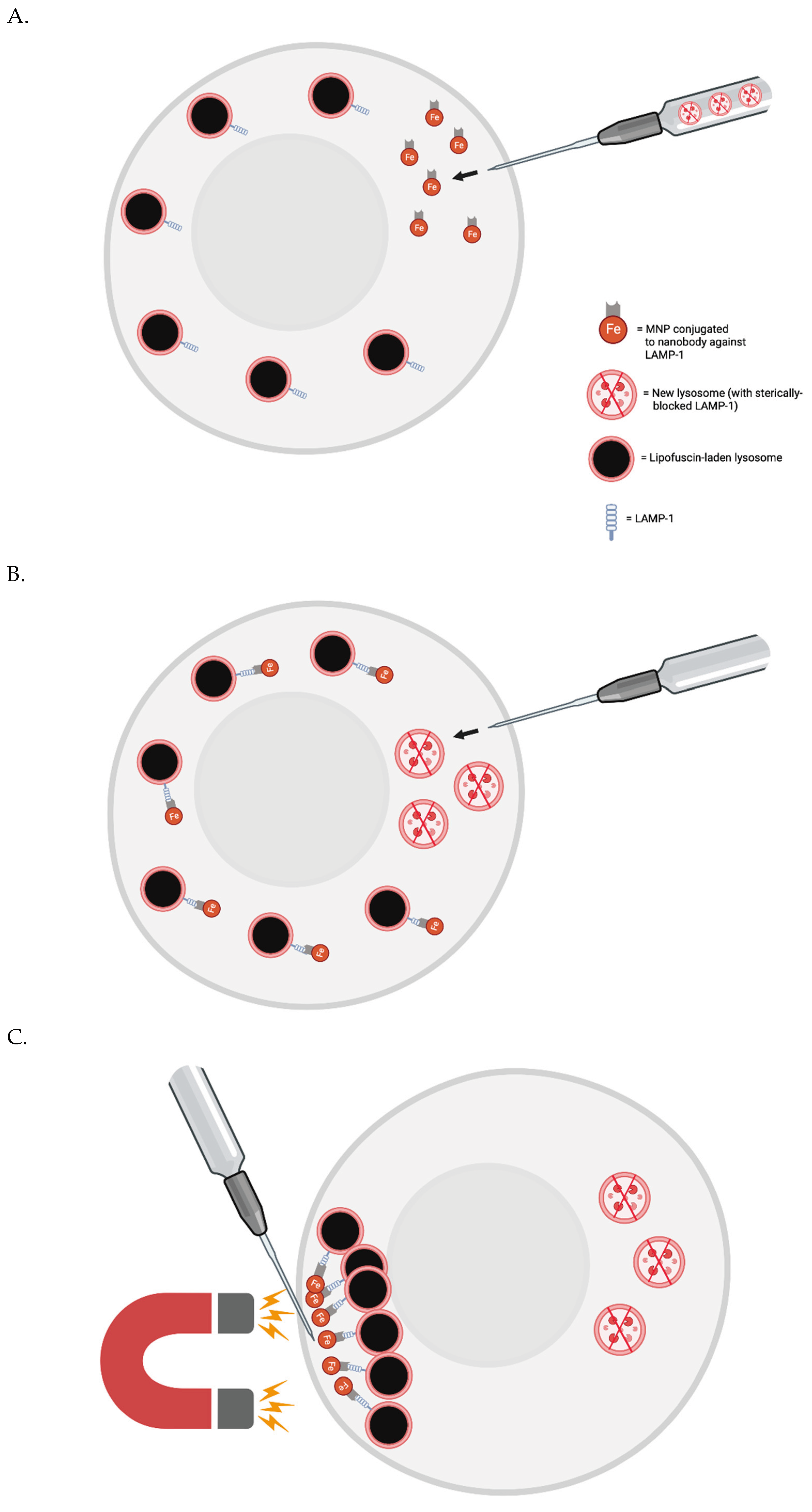
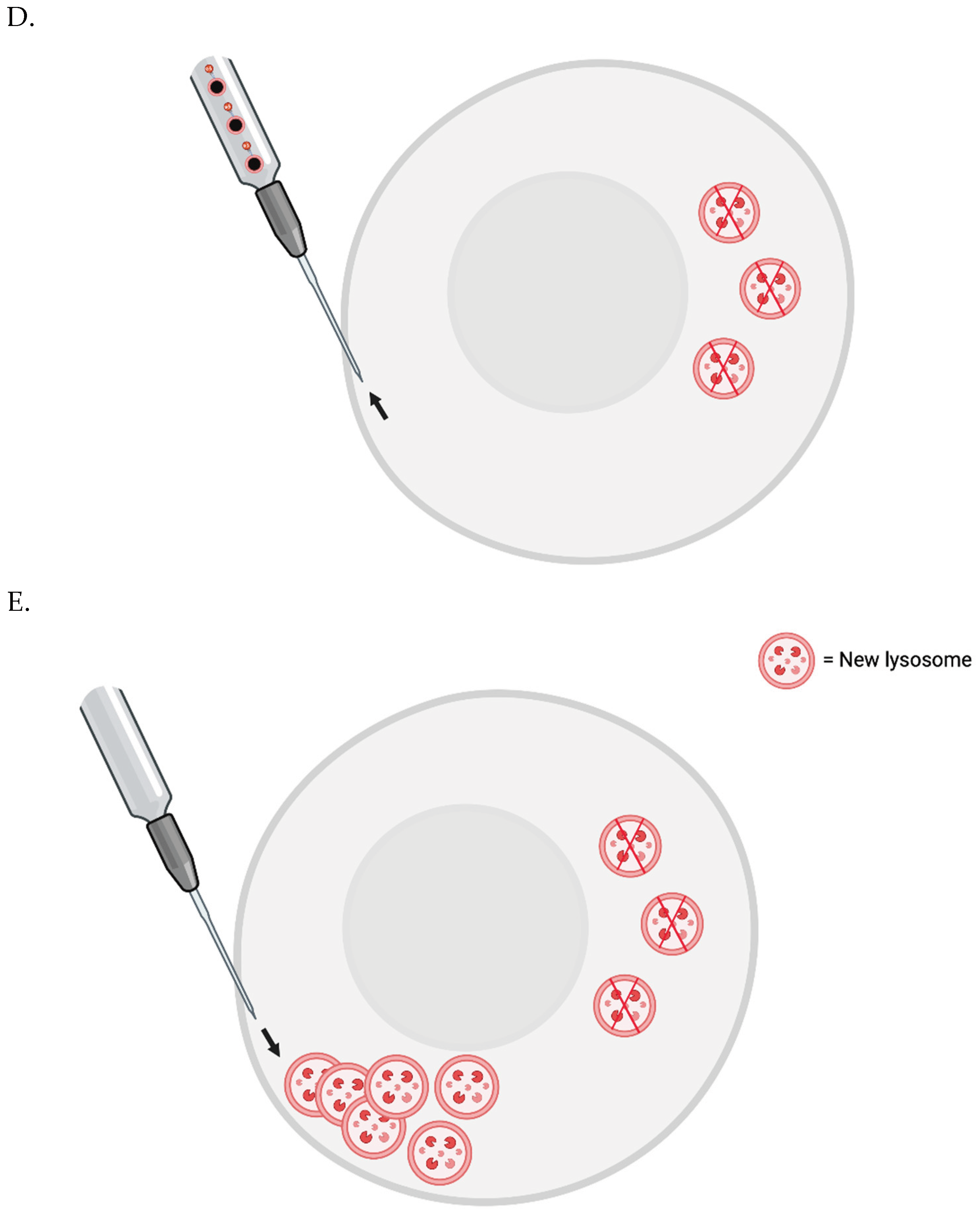
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



