Submitted:
22 February 2023
Posted:
23 February 2023
You are already at the latest version
Abstract
A synoptic review of plant disease epidemics and outbreaks was made using two complementary approaches. The first approach involved reviewing scientific literature published in 2021, in which quantitative data related to new plant disease epidemics or outbreaks were obtained via surveys or similar methodologies. The second approach involved retrieving new records added in 2021 to the CABI Distribution Database, which contains over a million global geographic records of organisms from over 50,000 species. The literature review retrieved 186 articles, describing studies in 62 categories (pathogen species/species complexes) across >40 host species on 6 continents. Pathogen species with >5 articles were: Bursaphelenchus xylophilus, Candidatus Liberibacter asiaticus, cassava mosaic viruses, citrus tristeza virus, Erwinia amylovora, Fusarium spp. complexes, Fusarium oxysporum f. sp. cubense, Magnaporthe oryzae, maize lethal necrosis co-infecting viruses, Meloidogyne spp. complexes, Pseudomonas syringae pvs, Puccinia striiformis f. sp. tritici, Xylella fastidiosa, and Zymoseptoria tritici. Automated searches of the CABI Distribution Database identified 617 distribution records new in 2021 of 283 plant pathogens. A further manual review of these records confirmed 15 pathogens reported in new locations: apple hammerhead viroid, apple rubbery wood viruses, Aphelenchoides besseyi, Biscogniauxia mediterranea, Ca. Liberibacter asiaticus, citrus tristeza virus, Colletotrichum siamense, cucurbit chlorotic yellows virus, Erwinia rhapontici, Erysiphe corylacearum, Fusarium oxysporum f. sp. cubense Tropical Race 4, Globodera rostochiensis, Nothophoma quercina, potato spindle tuber viroid, and tomato brown rugose fruit virus. Of these, 4 pathogens had at least 25% of all records reported in 2021. We assessed two of these pathogens – tomato brown rugose fruit virus and cucurbit chlorotic yellows virus – to be actively emerging in/spreading to new locations. Although three important pathogens – Ca. Liberibacter asiaticus, citrus tristeza virus and Fusarium oxysporum f. sp. cubense – were represented in the results of both our literature review and our interrogation of the CABI Distribution Database, in general our dual approaches revealed distinct sets of plant disease outbreaks and new records, with little overlap.
Keywords:
Plant disease epidemics
INTRODUCTION
Plant diseases pose a continuing threat to food security, plant health, and ecosystem health globally (Jeger et al. 2021; Ristaino et al. 2021; Rizzo et al. 2021). Historically, plant disease epidemics and outbreaks have been associated with increased interconnectedness of the landscape, climate change, human movement, and increased intensification of crop production practices such as irrigation. Plant pathogens are responsible for an estimated 20-40% of crop losses, although net effects are hard to quantify due to a lack of evidence related to precise distribution and impact (Savary et al. 2019, 2022). Even so, even less is known about the prevalence of pathogens of minor crops, of those of regional importance only, of non-food crops or concerning the provision of ecosystem services from native wild plant populations and communities (Gougherty and Davies 2021, Jeger 2021). It is reasonable to assume that epidemics or major disease outbreaks in major crops are at least somewhat well documented, but what is often missing is comprehensive information on the extent and frequency of new outbreaks across all plant pathosystems. Here we test a potential approach to fill that knowledge gap by exploring approaches to retrieve information related to new plant outbreaks and epidemics from literature and database searches.
This review has two main objectives. The first objective is to provide a synoptic view of plant disease outbreaks across all hosts as reported in peer-reviewed scientific articles published in 2021. The second objective is to summarise information from a comprehensive database on new reports of pathogens spreading or emerging in new geographical regions – or on new host species – in 2021. Our working hypothesis was that there would be little overlap in the results obtained from meeting the two objectives of our study, mainly because it is reasonable to assume that new reports of a pathogen species will usually precede epidemics or major outbreaks of that species that might subsequently occur. Nevertheless, by accessing information from two complementary approaches, we anticipate that this will identify any limitations in identifying emerging and re-emerging plant diseases by automated literature reviews and/or database searches.
Objective 1. A literature search was made to identify articles in which the objectives of the study and the underlying methodology, particularly the use of surveys and other surveillance methods, are well-described. As we describe below, the key importance of such studies lies in providing information that goes beyond that obtained in reports of the presence or absence of a given disease. We accept that restricting the review to publications from just a single year, 2021, may be considered a limitation, but the periods referred to often span multiple years. However, difficulties in obtaining up-to-date information on epidemics/outbreaks may affect decision-making in disease management (Thompson et al., 2018; Severns & Mundt, 2021). A wider problem is how to correct for inevitable time lags between data collection, analysis, and reporting (Bastos et al., 2019).
The breadth of our ambition makes it impractical to use expert-based assessment methods (Savary et al. 2019), important though these may be for estimating global crop losses of major crops. We do not limit the review to major food crops, or indeed any plant host category, or to pathogens that experts consider to be important. Instead, we use methods accounting for disease outbreaks reported in the scientific literature irrespective of geographical regions, hosts, or causal organisms. In the latter case, comprehensive reviews have been published for most pathogen taxonomic groupings, for example, viruses and viroids (Jones, 2021), fungi and oomycetes (Fones et al., 2020), bacteria and mollicutes (Agrios 2005), and nematodes (Bernard et al., 2017). The methods we use are based on principles of a systematic review, and so recognise the need to: (i) avoid as much bias as possible in the choice of pathogens or hosts; (ii) identify pathogen species in the search and not specify them a priori because of their perceived importance; and (iii) avoid giving undue emphasis to pathogens or plants which are used as model systems or for presenting novel scientific interest.
We note that, in searching the scientific literature, it is important to recognise the different terminologies used. For example, ‘harmful organisms’ (a term used in plant health and pest risk analysis, Devorshak, 2012) is a generic term, including pests and weeds. Pests could also be used to refer to both invertebrate organisms that cause damage due to, for example, herbivory, and pathogens causing disease. The term parasite can be used to describe a wide range of symbiotic associations, from mutualistic to pathogenic (Jiang et al., 2020). The term pathogen is used to describe a parasite that causes disease, although there can be some fluidity depending on the biotic and abiotic environment (Méthot and Alizon, 2014). Disease itself often requires both a pathogen and a vector for it to manifest. A vector may itself be a pest in the absence of the pathogen. For this reason, it is important that the results of automated searches are scrutinised to confirm their relevance to the search strings used. Plant pathology is also still influenced – and arguably over-influenced – by the ‘one pathogen-one host’ paradigm (Lamichhane and Venturi, 2015). However, disease complexes, mixed infections, the co-occurrence of closely related species (a major problem with plant viruses), sub-species, and other intraspecific designations of variety, biovar, pathovar and forma specialis, are all known to occur. These factors complicate disease assessments, risk assessments and indeed systematic reviews. For this reason, we have been careful to reflect these complexities in our methodology and interpretation of results.
Objective 2. The second objective of the review reflects the distinction between outbreaks of long-established and well-characterised pathogens, and new reports of pathogens, sometimes poorly or newly characterised, spreading or emerging in new geographical regions or hosts (Bebber et al. 2013, 2014). Such reports often arise from the requirements of national and regional plant health regulations for official surveys to determine the presence or absence of a named harmful organism, usually at the species level. For new reports across all hosts, pathogens, and geographical regions, the use of a comprehensive database is essential to assess patterns that emerge from records made in any given year, in our case 2021.
The comprehensive database we use here is the CABI Distribution Database (DDB). This database holds over 1.2 million geographic records of organisms and their locations, with a focus on species which are important to agriculture and the environment. Over 55,000 species are represented, across key areas such as crop pests, crop and animal diseases, invasive species, and natural enemies. Most of the data has been sourced from peer reviewed published literature and other authoritative third parties such as plant protection organisations. Whenever available, information is stored on the origin, extent of distribution and the invasive status. The DDB was used for this review as it has the most comprehensive global coverage of plant pests and diseases relevant to agriculture, and since it is continuously updated to reflect changing species distributions.
METHODS
Objective 1. Published Articles on Disease Outbreaks and Epidemics
A systematic review was made on Web of Science of journal articles published in 2021 (including online access of copy-edited but not necessarily typeset articles prior to full publication, aka ‘Early View’ or ‘First Look’ or other synonyms) following a systematic map protocol and search criteria based on thematic blocks and search strings (Vilvert et al., 2021). This was done in two stages (Figure 1). A scoping, proof-of-concept study used three thematic blocks and the preliminary search strings “Plant disease” AND “survey” AND (“epidemic” OR “outbreak”). The intention was to identify pathogen species for further study and to assess the extent of coverage in the set of recovered articles. This resulted in 45 articles which were checked manually for relevance. Of these, 8 articles were excluded, since they reported on food-borne diseases, plant-based vaccines, pharmacological and therapeutic activity of plants, mosquitoes/malaria, or animal/human diseases, leaving 37 articles remaining. This was considered an inadequate coverage and hence the search was extended to four thematic blocks with search strings “plant disease” AND (“survey” OR “surveillance” OR “monitoring”) AND (“pathogen” OR “vector”) AND (“epidemic” OR “outbreak” OR “impact” OR “damage”). We term this the ‘Stage 1’ search.
The extended search for Stage 1 resulted in a larger set of 141 articles which again were checked for relevance. Given the larger number of articles recovered, criteria were set for excluding articles, recognising that in some cases this is a matter of judgement rather than the result of any absolute criterion. In the main, excluded were articles on: inoculative weed control with pathogens; non-refereed literature reviews or routine listings of pathogens; soil surveys of microbial communities; development of detection sensors; and primer design and genomic approaches. Unless linked directly to new field observations of disease, also excluded were articles reporting analysis of seed samples; small scale experimental plots using artificial inoculation; analysis of historical culture collections; aerobiological studies; and mathematical models. However, articles that dealt with functional aspects of microbial communities associated with the natural occurrence of disease in the field were retained. Also retained were grower surveys and citizen science that referred to or were combined with field observations of disease. This selection process resulted in 90 articles that were retained for further analysis.
A separate search was done in Stage 1 for plant parasitic nematodes by substituting the term “plant disease” with “plant nematode” in the search string. This was done because nematodes are often referred to as parasites associated with pest damage rather than pathogens causing disease. Following the same exclusion procedures as above, 7 further articles were retained. Hereafter, we exclusively refer to plant parasitic nematodes as pathogens.
The 97 (= 90 + 7) articles recovered in Stage 1 were allocated to categories corresponding to: 1) a single named pathogen species, but also including a recognised disease complex with co-infecting often related species (65 categories); and 2) multiple pathogens in a crop, cropping/production system, forest trees, or in wild plant populations (6 categories). These 97 articles are termed the “Core articles” (labelled “Publications included in review” in Figure 1).
Each core article was flagged by host (crop, cropping/production system, or wild plant), pathogen group (fungus, oomycete, bacterium, phytoplasma, virus, nematode) and geographical origin (regions, country, continent, global), and one of 10 descriptor types for the main motivation of the study, recognising that many articles were motivated by more than one of these descriptors.
The descriptors were as follows.
- detection, discrimination, and analysis
- surveillance, including growers and citizen science
- emergence of new pathogens, strains, and vectors
- wild hosts and weeds as inoculum reservoirs
- wild relatives, land races, and genetic resources
- wild plant populations and communities
- disease management, including varietal resistance
- biological control
- climate change and variability
- yield loss estimation
The purpose of the Stage 1 search was to identify pathogen species for which disease epidemics or outbreaks had been identified through surveys or other systematic approaches. To provide additional context and information on the pathogen species identified, an augmented Stage 2 search was done with two thematic blocks and the search strings: “<pathogen name>” AND (“survey” OR “epidemic” OR “outbreak”), again restricted to 2021 publications. In Stage 2, the “survey” and the “epidemic or outbreak” thematic blocks were combined in a single thematic block. Hence, the search was intended to recover articles reporting on an epidemic or outbreak of a given pathogen but without reporting a survey, and vice-versa. The search strings used in Stage 2 for the pathogen species identified did not include “surveillance”, “monitoring”, “impact”, or “damage” as used in Stage 1. The 89 additional articles thus retrieved are termed the “Supplementary articles” (labelled “Publications retained and added to review” in Figure 1). A similar approach was followed for the multiple pathogen categories identified, but with an appropriate thematic block defined for the cropping/production system rather than the single pathogen name. Articles recovered with these modified search criteria were allocated to the defined categories from Stage 1, giving a total of 186 retained articles across all categories after the results of both Stages were included. The periods of time over which surveys were made were noted for each study.
Objective 2. Records in the CABI Distribution Database
Automated Identification of Potential New Pathogen Records
The CABI DDB attempts to maintain up-to-date presence/absence status records across 507 geographic areas. These can be national or sub-national (e.g., state, province), administrative areas and other regions, islands, or sea areas of biogeographic importance. To perform the following analysis, data were initially extracted from the MS SQL Server database and subsequently analysed in MS Excel and QGIS.
Firstly, all distribution records with a 'Present' status and a source reference citation year of 2021 were extracted from the database (Figure 2). If no citation year was published, we only considered records added to the database between 01/01/2021-31/12/2021. Records with a poor “Data Quality” flag were excluded.
The next step was to filter to consider only plant pathogens. To classify organisms, the GBIF taxonomic library (GBIF Secretariat, 2021) was used to exclude organisms which are not pathogenic, e.g., Plantae and the Animalia phyla: Chordata, Arthropoda, and Mollusca. The classification was also cross-referenced with the Crop Protection channel of the CABI Compendium (https://www.cabidigitallibrary.org/product/QC). If an organism had datasheet type ‘Pest’ it was considered a plant pathogen. Any remaining unclassified organisms were manually checked against EPPO Global Database (EPPO, 2021) and/or search of scientific literature for evidence that they cause plant disease. This resulted in a set of 25,809 distribution records from 931 organisms in 416 geographic areas identified as plant pathogens.
To establish which of the 2021 records represented a first record of a pathogen in a location, a check was made against the rest of the DDB for any pre-2021 records of the organism in the geographic location. Where no previous ‘Present’ status records were held in the location in question, the record was considered a ‘new’ occurrence record.
The data were then summarised for each pathogen to count the number of geographic locations with at least one new record. A list ordered by the number of locations was used to select pathogens for manual review. Pathogens which had new 2021 records for at least four locations were selected for further investigation. This threshold of four records was chosen because the number of pathogens returned with three records or fewer was much larger, to the extent a manual review would have been overwhelming.
Manual Review
To test the efficacy of the automated process, and to assess the proportion of records which represented pathogen spread to a new location or presence on a new host, for each of the pathogens with four or more new 2021 records, the individual distribution records and their sources were reviewed in detail. Records were categorised as either:
- A confirmed first report in a location or on a new host
- Not a confirmed first report
Records were considered first reports if the cited reference confirmed that a pathogen had been recorded in a new geographic area (of the 507 used in the DDB), where it had not previously been recorded. A record was also considered to be a first report if the cited reference confirmed that the pathogen had been identified on a host plant on which it had not previously been recorded. Records that failed to meet these criteria were considered not to be confirmed first reports.
The date of each confirmed first report was identified to be the year that symptoms were first observed or the year that samples of the pathogen or affected host plant/s were collected, not the year the report itself was published.
Details of the confirmed first reports for each pathogen were then described, summarising the recent changes in distribution of the pathogen, the host plants with which it was associated and any other information from the reports that was considered relevant.
RESULTS
Objective 1. Published Articles on Disease Outbreaks and Epidemics
Core articles from Stage 1 (single pathogen species or species complexes allocated to categories, hereafter ‘named species’) are tabulated in Electronic Supporting Information S1 for fungi, oomycetes, bacteria, phytoplasmas, viruses, and nematodes, respectively. The greatest number of named species was for fungal pathogens (29), followed by plant viruses (11) (Figure 3). Together these groups accounted for 60% of all named species. No core articles were identified for viroids. In many surveys of plant diseases, multiple rather than single pathogens are recorded for a given plant host, cropping system (mixed cropping, rotation, or within season sequential planting), or wild plant populations (both native and invasive). Production may vary from annuals to perennial, with different production systems from seed production to vegetative propagation in plant nurseries to the harvested crop. Core articles relevant for multiple pathogens affecting a crop or cropping system, including crop-associated weeds, production system (including aquaculture, tree/forest systems), or wild plants, are tabulated in Electronic Supporting Information S2.
Of 45 host species covered in the core articles, only pathogens of maize (6), wheat (4), rice (4), tomato (4), citrus (3) and potato (3) were reported at least twice (Figure 3). Geographically, the core articles reported studies in 42 countries. Most articles described studies done in the United States (20), followed by China (9). At the continental scale (Figure 4), the comparable numbers were Europe (25), North America (23, including Canada and Mexico), Asia (15 in total, with 9 from China and 6 across Central, South, East and South-east Asia), Africa (14), South America (9, including Central America and the Caribbean) and Australasia (5, only Australia). Accounting for articles which reported multiple pathogen-location pairs, as well as for two articles which took a worldwide focus and so were relevant to every continent, allowed us to project a graphical summary of the results of Objective 1 onto a global map showing species counts per taxonomic group and continent (Figure 4).
Five article types were dominant (Figure 3): disease management, including deployment of resistant cultivars (20), emergence of new pathogens, strains, and vectors (18); disease surveillance, either remote or ground-based (15); detection, discrimination, and analysis of pathogens (14); and across all articles for wild plant populations, weeds, and wild relatives (14).
Combining the results of Stage 1 and Stage 2 resulted in 14 categories for named species or species complexes with five or more articles (Table 1; Figure 5). The years over which surveys were made are shown in the final column of Table 1 (see also Electronic Supporting Information, S4). These included five fungal pathogens (or complexes): Puccinia striiformis f. sp. tritici, Zymoseptoria tritici, Magnaporthe oryzae, Fusarium spp. complexes and Fusarium oxysporum f. sp. cubense; four bacterial pathogens: Candidatus Liberibacter asiaticus (and vectors), Xylella fastidiosa (and vectors), Erwinia amylovora and Pseudomonas syringae pvs; three viral pathogens (or diseases caused by interactions between pathogens): citrus tristeza virus, cassava mosaic viruses and maize lethal necrosis co-infecting viruses; the nematode Bursaphelenchus xylophilus; and Meloidogyne spp. complexes. These named species are discussed individually in Electronic Supporting Information, S5. Where articles were grouped into multiple pathogen categories or by cropping/production practices, combining the Stage 1 and Stage 2 searches resulted in five categories with five or more articles - further discussed in Electronic Supporting Information, S6.
Objective 2. Records in the CABI Distribution Database
The automated process to identify new 2021 records from the DDB resulted in 617 distribution records from 283 pathogen designations (Electronic Supporting Information, S3: Figures S3-4 and S3-5). The vast majority of the first reports identified by Objective 2 were based on the results of field surveys. Surveys were initiated for several reasons but were predominantly due to the observation of novel symptoms on commercially grown crops. In many cases, sequencing was necessary to accurately identify the causal agent of the observed symptoms (e.g., Cho et al. 2021; Hofer et al. 2021; Noronha et al. 2021). This was particularly important in characterising mixed infections and revealing the presence of cryptic species. Studies like these are a starting point to understanding their relative contributions to crop yield loss and designing more effective management strategies (Hofer et al. 2021; Hu et al. 2021; Kavalappara et al. 2021).
Of the 617 records identified, 180 plant pathogens returned a single new 2021 record. A total of 255 geographic locations recorded at least one new 2021 record. The majority of new 2021 records were for occurrences of pathogens in Asia (226), and the fewest records were from Oceania (20), with a high proportion of records for fungi and viruses for all continents. Of these, 28 pathogens met the threshold for manual review, i.e., four or more new 2021 records (Figure 6 and Figure 7). The manual review determined whether the records were confirmed first reports in a new geographic location or on a new host plant. Fifty-nine records were classified as such, covering 15 plant pathogens (Table 2); again, a more detailed description of these pathogens is provided in the Electronic Supporting Information, S7. Records for the remaining manually reviewed 13 plant pathogens were not considered to be first reports for various reasons, including taxonomic revisions, external databases referenced by the date the information was accessed, and when it was determined surveys focused on a pathogen in a region it was already known to occur.
Which Pathogens Are Actively Spreading/Emerging?
Of the 15 plant pathogens identified (Table 2), 4 showed a signature of possible actively spreading and/or emerging in new region(s). These were identified by filtering the results after manual review to isolate pathogen species for which: i) there were at least two new records in 2021; ii) at least 25% of the total records for the pathogen in question were from 2021. These 4 pathogens were: Tomato brown rugose fruit virus, Cucurbit chlorotic yellows virus, Apple hammerhead viroid and Colletotrichum siamense.
Tomato brown rugose fruit virus (ToBRFV) is a serious threat to tomato (and pepper) cultivation. First identified in 2014, in Israel, it is now found in at least 35 countries across Asia, Europe, North America, and Africa (Zhang et al., 2022). The pathogen breaks the Tm-1, Tm-2 and Tm-22 resistance genes present in many varieties of tomato, as well as L1 and L2 alleles in pepper, leading to the potential for very rapid spread following first incursion into a new agricultural region. Despite many outstanding knowledge gaps around the pathogen’s spread, mechanical transmission via workers and/or farm/greenhouse machinery/irrigation is thought to be important at local scales, with very long distance spread possible because the virus can be seed borne (Oladokun et al., 2019). There are currently no resistant cultivars, and control is often based on phytosanitary measures. However, despite some limited success at local scales, the pathogen is still actively spreading to new countries, and presents significant threat to tomato cultivation worldwide (Zhang et al., 2022). The total of 15 records from 2021 in the DDB included reports from Iran, Saudi Arabia, Syria, and Uzbekistan in Asia; from Austria, Belgium, Czechia, Estonia, Hungary, Malta, Norway, Portugal, Slovenia, and Switzerland in Europe; and from Mexico, two states of the USA and Brazil in the Americas. As well as development of resistant varieties, which will be necessary for durable control, particularly important will be developing accurate, cost effective and reliable methodologies for early detection, since – in common with many viral diseases – symptoms can be rather difficult to distinguish unambiguously. Notably, the pathogen provides one of the few examples of tracking the spread of a plant pathogen in real time based on genetic information, as demonstrated for outbreaks in the Netherlands using the Nextstrain platform (van de Vossenberg et al., 2020).
Cucurbit chlorotic yellows virus was first described in 2004 in Japan following reports of leaf yellowing symptoms on greenhouse grown cucurbits (Gyoutoku et al. 2009). The pathogen has subsequently been detecting in several Mediterranean and Middle Eastern countries, including – but not limited to – Lebanon, Iran, Greece, Egypt, Turkey, Jordan, Spain, Cyprus, and Algeria (CABI, 2023). First reports in North America appeared in 2018, although subsequent examination of frozen plant materials revealed the pathogen had been present since at least 2014 (Wintermantel et al. 2019), and the virus is now reported on cucurbits in several US states (Hernandez et al., 2021). Local spread of the virus is vectored by multiple species within the Bemisia tabaci species complex, with a semi-persistent transmission mode (Li et al. 2016). Long distance spread is thought to occur via infected plant materials and the nursery trade. As well cucurbit crops, including melon, watermelon, cucumber, zucchini and pumpkin, the virus infects various wild hosts, including various weed species. The potential wild reservoir of inoculum this presents, together with rapidly evolving insecticide resistance in B. tabaci populations, combine to present significant difficulties in effective disease management. Management is also affected by the lack of easily recognisable disease symptoms, although genetic tests are available. The economic impacts are potentially severe – in Lebanon yield losses of up to 30% are reported, driven in part by coinfection with other viruses (Abrahamian et al, 2015) – and there are currently significant concerns around further impacts in cucurbit producing regions worldwide. The 11 records from 2021 in the DBB included new reports from China, India, Philippines, and South Korea in Asia, as well as from four separate US States (Alabama, Florida, Georgia, and Texas).
Apple hammerhead viroid was initially discovered by analysis of high throughput sequencing data (Zhang et al. 2014). It has subsequently been reported worldwide, including countries in Africa, Asia, Europe and North and South America (CABI, 2023). However, it is invariably found in samples from plants infected with other viruses, making the correlation between viroid infection and any specific symptoms impossible to determine (Várallyay et al., 2022). We therefore assess the new reports in 2021 – in Tunisia, India (two different states) and Brazil – as not necessarily reflecting new spread of a potentially damaging pathogen to a new location, but instead more likely reflecting the increasing availability of increasingly sophisticated bioinformatic methods to detect a long-standing and perhaps even benign association, and so do not discuss this species further.
Anthracnose diseases are caused worldwide by Colletotrichum spp. fungi and affect a very large number of different host species. This large monophyletic group of ascomycete fungi consists of more than 200 accepted species, classified into at least 14 species complexes (Damm et al. 2019). Major losses are routinely caused to economically important crops, especially fruits, vegetables and ornamentals, with increasingly severe impacts on stable food crops in the developing world (Dean et al. 2021). In 2021 our search of the DDB revealed new reports caused by the species C. siamense on Vernicia Montana (in China), rubber trees (China), Thai basil (in Malaysia), onion (Sri Lanka) and avocado (New Zealand). However, a recent EFSA pest categorisation reports a worldwide distribution of this species in particular, with records from Argentina, Australia, Bangladesh, Brazil, China, Colombia, Egypt, Ghana, India, Indonesia, Israel, Japan, Kenya, Laos, Malaysia, Malawi, Mexico, New Zealand, Nigeria, Pakistan, Philippines, Puerto Rico, Republic of Korea, South Africa, Sri Lanka, Taiwan, Thailand, Turkey, Uruguay, USA, Vietnam, Zimbabwe (EFSA PLH Panel, 2022). It is therefore likely that the driver of the apparent signature of new spread in DDB records – with no records from before 2021 – is due to confusion around classification of historic reports. C. siamense belongs to the C. gloeosporioides species complex, and until recently was not always reported separately. We therefore do not discuss this species further here.
Comparing the Results Obtained for Objective 1 and Objective 2
Six named pathogens were identified by both of our two approaches. In alphabetical order, these were: Candidatus Liberibacter asiaticus, Citrus tristeza virus, Colletotrichum siamense, Fusarium oxysporum f. sp. cubense Tropical race 4, Globodera rostochiensis and Tomato brown rugose fruit virus. Of these, three were reported in at least 5 publications in Objective 1 (Table 1). These were Ca. Liberibacter asiaticus, Citrus tristeza virus and Fusarium oxysporum f. sp. cubense. We note that these three named species are well-established in the results from the DDB, with many records from before 2021. This highlights the advantage of our dual approach, in which records in a database like the DDB are followed by epidemic spread and so publications in the scientific literature.
Thirteen additional pathogens identified by the automated review of DDB records overlap with pathogens identified by the literature review, but these were not reviewed manually, as they did not reach the threshold of four new 2021 records. In some cases, the literature review identified pathogens at the genus level or as species complexes or groups of pathovars. There is some overlap in DDB with these groupings, e.g., Colletotrichum spp. (13 species identified in DDB results), Pseudomonas syringae pathovars (2 pathovars identified in DDB results).
A map showing the combined counts of locations from both the literature review and the CABI Distribution Database is shown in Figure 8.
Combining data from both approaches (Objectives 1 and 2) revealed differences in the dominant pathogen types between continents (Figure 8). In Australasia and North America, there was a dominance of fungal pathogens. These occurred on a wide variety of host types, for example Diplodia corticola on oak (Fereira et al 2021), Fusarium graminearum on cereals (Breunig and Chilvers 2021), and Pyrenopeziza brassicae on vegetable crops (Claasen et al. 2021) in North America. In Australasia fungal outbreaks reported included Alternaria brassicola on ornamentals (Papaix et al. 2021) and Puccinia striiformis f.sp tritici on wheat (Sandhu et al. 2021). In contrast, in Asia, Europe and Africa, there was a dominance of viruses and viroids. These included 8 reports of apple rubbery wood phytoplasma across China, Cassava viruses in Kenya (Munguti et al. 2021) and Thailand (Soakham et al. 2021), and multiple reports of Tomato brown rugose fruit virus across Europe. Bacterial pathogens did not dominate in any one continent but were represented as the highest proportion of new reports in Europe due mainly to multiple new findings of Xylella fastidiosa. The greatest variability in pathogen types was in South America. This included new outbreaks of Candidatus liberibacter asiaticus (causal agent of citrus greening syn. HLB) (EPPO, 2021a), Fusarium oxysporum f.sp cubensis on Banana (Heck et al. 2021), and multiple reports of the nematode Aphelenchoides besseyi in Brazil (EPPO 2021a, Heck et al. 2021).
The descriptors assigned to each article in Objective 1 revealed the main motivation for each study retrieved, and this also varied strongly with continent (Figure 3C). In North America, the majority of studies were motivated by disease management factors, whereas in Asia both disease management and detection were primary motivations. In Europe, most studies were motivated by the emergence of new pathogens, strains and vectors, as well as detection, driven by the large number of new reports of Xylella fastidiosa. In Africa and South America disease management was a primary motivator, whereas in Australia most studies were motivated by wild plant populations and communities, with many new reports on ornamental hosts.
DISCUSSION
Although yield losses in major crops are increasingly well documented (Savary et al. 2019, 2022), the underlying disease dynamics driving losses are much less well studied. Diseases in minor crops, non-food crops and natural plant systems are particularly poorly characterised. Here we used two contrasting approaches to analyse articles published in 2021 on plant disease epidemics across the world. The first (Objective 1) was a systematic review of the 2021 literature reporting surveys of plant disease occurrences. The second (Objective 2) retrieved new plant pathogen presence records added to the CABI Distribution Database (DDB) in 2021.
The approaches are complementary, with both helping to build a holistic picture of the range of plant health threats that are impacting, or have the potential to impact, food security, as well as plant and environmental health. In turn, information such as this can inform appropriate surveillance measures or management strategies to either prevent pathogens from entering a novel location or limit the likelihood or impact of recurring outbreaks.
Geographically, both approaches revealed the highest numbers of outbreaks and new records in North America, Asia, and Europe, with lower numbers in Africa and South America. There were also similarities in the pathogen taxa identified, with fungi and viruses dominating. Interestingly, however, the two approaches led to a pair of relatively distinct lists of specific pathogen species. Exceptions to this were Candidatus Liberibacter asiaticus, Citrus tristeza virus, Colletotrichum siamense, Fusarium oxysporum f. sp. cubense Tropical race 4, Globodera rostochiensis and Tomato brown rugose fruit virus. These pathogens were retrieved from both approaches and of them Ca. Liberibacter asiaticus, Citrus tristeza virus and Fusarium oxysporum f. sp. cubense were reported in at least 5 publications in the 2021 literature (Objective 1, Table 1).
Which Limitations Might Affect Our Methodology?
In Objective 1, we concentrated on published literature reporting the results of surveys. This task would have been made easier if journals specified general guidance for good practice in planning, designing, carrying out, analysing, and writing papers reporting the results of this type of survey. Such guidance would include reporting the rationale for surveys, the purposes they serve, the questions being asked, and whether they are hypothesis-driven or exploratory: often this vital information was lacking from the literature returned by our searches. Ideally, this guidance would also encourage at least a partial move away from single pathogen surveys, given the increasing recognition of disease complexes and syndromes and the recent awareness of mixed or co-infections and cryptic species (Lamichhane and Venturi, 2015). The increased splitting into various sub-specific forms and other genomic characterisations arising from surveys presents additional major data analysis challenges and difficulties in biological interpretation.
In Objective 2, we initially extracted records automatically from the DDB, but manual intervention was required to identify confirmed first reports of pathogens in new geographic locations or on new hosts. This is unsurprising since confirmed new reports are not the only focus of the DDB. Records required manual review for various reasons, predominantly due to taxonomic revisions and/or differences in the date of addition to the DDB versus the date of the survey itself. On other occasions, the literature cited by records found in the automated stage discusses surveys carried out with the purpose of determining pathogen prevalence, disease severity or pathogen impacts in locations where a given pathogen is already well known to occur. This was not our focus here, and so such reports required manual removal.
Certainly, pathogen distributions present a constantly evolving picture, and databases as used in Objective 2 can only possibly ever provide a snapshot in time. For example, Tomato brown rugose fruit virus was first reported as present in 2021 in Estonia, but has subsequently been reported as absent, having been eradicated (EPPO, 2022). It is also notable that the first reports from Objective 2 contain little to no information about the scale of crop losses caused by the pathogens identified. There is a paucity of quantitative data on the impacts of pathogens on their host plants. However, the publication of first reports is a vital first step in monitoring the subsequent impacts that occur.
Which Factors Might Bias Our Results?
Both approaches adopted here were systematic, explicitly designed to attempt to avoid bias toward certain crops or pathosystems. Results, therefore, ought to reflect the balance of current outbreaks and new findings. However, there are nevertheless unavoidable biases that remain. Both the literature we searched in Objective 1, and most of the literature and records contributing to the DDB in Objective 2, were in English, introducing a “Tower of Babel” bias into our results (Grégorie et al., 1995). This has been acknowledged to affect the results of past studies using similar sources of data (Bebber et al., 2019). Certain pathogens are topical for scientific reasons since they constitute model systems for experiments, for example A. thaliana and P. syringae (e.g., Karasov et al. 2018). The pathogens that appear in the literature are also driven, in part, by the need to attract research funding, or indeed the availability of funding, and do not necessarily reflect the impact of a disease in economic, social, or environmental importance.
Additionally, our results will have been affected by the way in which regulatory surveys for quarantine pests in plant health are principally targeted at substantiating claims for pest freedom. Such surveys could even be argued to provide perverse incentives which, in fact, reward under-sampling of those pests. Though risk and statistically based survey approaches are increasingly being adopted (EFSA 2020, Mastin et al., 2020), detection surveys are often used to establish presence/absence rather than providing representative surveys of disease dynamics. Lack of resources for representative and widespread monitoring of disease has in many cases led to significant outbreaks being undiscovered for decades. For example, Xylella fastidiosa was first reported in France in 2015, but independent studies have estimated it was introduced in the 1980s (Soubeyrand et al., 2018; Dupas et al., 2022). In the UK, ash dieback disease was first reported in 2012 but may have been present and spread from ash stock planted as early as 1991 (Wylder et al., 2018).
Our results likely underestimate the importance of pathogens in the “Global South”, reflecting greater surveillance and research effort in richer, developed countries (Jones et al., 2008). Certain pathogens have significant regulatory implications following first discovery, for example, Tomato brown rugose fruit virus, and others receive increased attention because of the impacts they might cause. Conversely, pathogens sometimes do not receive very much attention because they are perceived as being unimportant. This can have unintended consequences. As an example, there is a paucity of records on Xylella fastidiosa on deciduous tree species in North America. This likely reflects underreporting of the disease in these areas due to perceived low impact but has led to considerable current uncertainty in Europe as to what the impact could be if X. fastidiosa spreads in forest tree species present in the EU (EFSA 2019).
The structure of the geographic regions in the DDB also introduced some possibility for bias; for example, Ca. Liberibacter asiaticus was reported for the first time in Bahia, a state in Brazil. This only showed up in our results as a new outbreak because Brazil is one of the countries for which sub-national geographical resolution is retained in the DDB. A similar issue underpinned our identification of apple rubbery wood viruses as a pathogen presenting the signature of emergence/spread: the pathogen is certainly spreading, but all records in the DDB in 2021 relate to different locations in (mainland) China.
Other issues that impact on reporting of disease include a potential over-representation of those diseases that have strong symptom expression and existing tools for diagnosis, as well as time lags between when samples are collected in the field and when laboratory diagnosis occurs. Moreover, our approach does not capture data from the any number of regulatory surveys that are not published nor reported via the DDB, which might particularly affect pathogens which are not notifiable.
Implications for Disease Management
Studies concerned with aspects of disease management accounted for many of the articles recovered under Objective 1, very often linked to disease surveillance in regulated zones (Lazaro et al. 2021) and detection/ discrimination of new pathogens or novel strains using molecular diagnostics. Examples include Sri Lankan cassava mosaic virus emerging in Southeast Asia (Charoenvilaisiri et al. 2021), Xylella subspp in the Balearic Islands (Olmo et al. 2021), citrus tristeza virus strains (Park et al. 2021, Llanes-Alvarez et al. 2021), Erwinia spp in the USA (Singh et al. 2021), antibiotic resistance in Pseudomonas syringae pv actinidiae (Psa) (Lee et al., 2021), fungicide resistance in Zymoseptoria tritici (Garnault et al., 2021) (see detailed summaries in Electronic Supporting Information, S5). Many studies were concerned with the field performance and deployment of resistant varieties: such as for pokkah boeng disease in sugarcane (Shan et al. 2021, Solic-Palacios et al. 2021), wheat yellow rust (Perronne et al. 2021, Esmail 2021). Fewer field studies were made on the use of biological control agents in disease management: such as for root knot nematode in vegetable crops (Sabri et al. 2021), Psa in kiwi fruit (Ares et al. 2021) (Electronic Supporting Information, S5).
From a broader perspective, disease management is increasingly seen as a multifunctional issue involving the securing of food supplies while safeguarding ecosystem health and reducing dependency on natural resources (He et al. 2021), including the extended use of within-crop diversification (Wang et al. 2021b) as shown for Zymoseptoria tritici in intraspecific mixed stands of wheat varieties (Orelana-Torrejon et al., 2021; Electronic Supporting Information, S5). Integrated crop management in both low- and high-input cropping systems is seen as an instrument for maintaining yields while reducing reliance on synthetic inputs such as pesticides and fertilizers (Richard et al. 2021). The use of even intermittent conventional insecticides to control Diaphorina citri, the vector of Ca. Liberibacter asiaticus (Clas) may affect the activity of natural enemies (Shrestha et al. 2021; Electronic Supporting Information, S5). Studies on Ca. Liberibacter asiaticus (Clas) in feral understorey citrus in forested areas have suggested that shading may have the potential to reduce disease transmission in commercial citrus production (Vincent et al. 2021: Electronic Supporting Information, S5). Disease management may differ between optimal and sub-optimal areas: such as for Fusarium oxysporum f. sp. cubense (Foc) in banana cultivation (Teixeira et al., 2021), maize lethal necrosis (MLN) and maize seed productions systems in different agro-ecological zones (Eunice et al., 2021) (Electronic Supporting Information, S5).
Where containment is practised as a disease management objective, the importance of involving a broad spectrum of participants – farmers, extension workers, practitioners, and plant health regulators – is now well recognised (Ladisa et al. 2021). Such involvement is necessary to improve cooperation and coordination amongst participants (Sherman et al. 2019) and should form part of any risk-based approach to disease management (Hyatt-Twynam et al. 2017). These concerns were mirrored in studies on Clas in California (Garcia-Figuera et al., 2021), cassava mosaic virus disease in Zambia (Szyniszewska et al. 2021) (Electronic Supporting Information, S5). Other studies on disease management of Clas have given detailed accounts of control techniques such as precision trunk inoculation but have not considered the implications for all participants (Li and Nangong, 2021: Electronic Supporting Information, S5).
The complexities of disease management where there are multiple pathogens present in a mixed or rotational cropping system are illustrated for maize-soybean systems (Hampf et al. 2021, Meier et al. 2021), rice-wheat/maize systems (Shan et al., 2021b, Sekiya et al. 2022), and grapevine-cover crop systems (Léon et al., 2021) (Electronic Supporting Information, S6). However, in general, this important topic is arguably under-represented in the literature, given the real world context in which many growers are placed.
On detecting the first report of a pathogen, some publications identified under Objective 2 emphasised the requirement to monitor and control vector populations that may facilitate the spread of pathogens. This was a particular concern for disease management of Cucurbit chlorotic yellows virus in Texas, USA, where whiteflies in the Bemisia tabaci species complex are abundant (Hernandez et al. 2021). Publications also speculated on the possible sources of infection by novel pathogens, with many suggesting they originated from the importation of commercial seed (Sabra et al. 2021). It was also highlighted that for Tomato brown rugose fruit virus, the movement of unregulated seed and propagation by keen gardeners may be another pathway of introduction and a further threat to commercial crop production (Hamborg and Blystad 2022, cited by EPPO 2021). All these aspects need to be taken into consideration to develop robust management strategies to prevent introductions, to quickly identify them when they do occur and to limit their subsequent spread and potential crop losses.
Some first reports in Objective 2 were identified from samples of asymptomatic plants, often the result of official surveys carried out as part of a routine monitoring program. Such programs are more likely to be established for pathogens known to be high risk, such as Tomato brown rugose fruit virus in Europe and Candidatus Liberibacter asiaticus in Citrus growing regions of Brazil (EPPO 2021). They can also be particularly useful in detecting pathogens that might otherwise have spread undetected due to their symptoms being consistent with those of other, perhaps more common, pathogens (Chynoweth et al. 2021). Another category of first reports represented the outcome of pre-export examinations of both commercial and non-commercial seed (EPPO, 2021 for Tomato brown rugose fruit virus; Skelton et al. 2019, cited by EPPO 2021 for Potato spindle tuber viroid). These studies highlight the critical role border inspections of imports and exports play in preventing spread of novel pathogens.
Predicting Future Spread of Disease
Often linked to disease management, disease prediction assumes that being able to predict future disease risk and occurrence can lead to improved disease management or choice of disease control options. Predictions may be relevant at different temporal and spatial scales, and with different levels of statistical sophistication, from individual fields or production units, based on weather data (Kim et al. 2020, Shah et al. 2019) and/or phenological data (Hjelkrem et al. 2021) to the landscape level (Yuen and Mila, 2015). Prediction may relate to individual pathogen species or, more generally, to taxa with varying levels of vulnerability based on phylogenetic relationships (Robles-Fernández & Lira-Noriega 2017). Neural network models and standard daily meteorological data gave the best prediction of the population dynamics of D. citri, the vector of CLas, at the field scale in Pakistan (Bibi et al. 2021; Electronic Supporting Information, S5). Typical field-based prediction models linked to weather data and crop phenology/morphology were developed for B. tabaci, the vector of cassava mosaic viruses, in Uganda (Katono et al. 2021; Electronic Supporting Information, S5). A weather-based model was developed to predict when a threshold would be reached for use of a systemic fungicide to control Z. tritici (Beyer et al. 2022; Electronic Supporting Information, S5).
Risk-based prediction at the regional level has become more common in recent years and is most relevant to the topic of this paper. Risk-based surveillance methods have been developed for early detection of invasive plant pathogens. The perhaps counter-intuitive result is that basing surveillance only on areas predicted to be high risk is not always the optimal strategy (Mastin et al. 2020). A model was developed to predict the probability of pine wood nematode occurrence in Hubei province, China, based on vegetation condition, anthropogenic activity and topographic features obtained from remote sensing data and geographic information systems (Zhang et al. 2021; Electronic Supporting Information, S5). Distinct subpopulations of the nematode occur in China; based on genomic variation, it was predicted that the nematode survives better at lower temperatures than previously thought (Ding et al. 2021; Electronic Supporting Information, S5). Ecological niche modelling was used to predict the likely occurrence in regions of Mexico of Trioza erytreae, a vector of Ca. Liberibacter africanus (Claf) as well as Clas (Espinoza-Zaragoza et al., 2021; Electronic Supporting Information, S5). Species distribution models have also predicted that in future years climatic suitability may be less favourable for Philaenus spumarius, the main vector of X. fastidosa in Europe (Godefroide et al. 2022; Electronic Supporting Information, S5).
Climate Change, Yield & Ecosystem Services
Disease prediction may prove of limited value unless the impact of climate change on crops, pathogens, vectors, and the broader biotic environment are taken into consideration (Lops 2022, Sreenivas 2022). This broader view, however, often provides general opinion rather than specific detail. In some cases, detailed experimental information on the host-pathogen interaction in relation to temperature gives some indication of likely responses to a global climate (e.g., Amari et al. 2021). Historical data over extended periods have been used to predict trends in future years and countries for specific diseases or groups of pathogens of crops and wild plants (Bebber et al. 2019, Burdon & Zhan 2020, Jeger 2021, Chen et al. 2022, Tang et al. 2017). In a comprehensive review, Chaloner et al. (2021) tracked the likely global trends in crop production, yields, and disease pressure. These authors predicted that yield and infection risk will increase at high latitudes over the 21st century, but that in the tropics there will be limited productivity gain and a reduction in infection risk. The balance between yield gains and infection risk in relation to climate change and variability makes the consequences for food security uncertain. A process-based simulation model was used to predict the incidence and severity of rice blast disease under different time scenarios and climate change scenarios (Wang et al. 2021b; Electronic Supporting Information, S5). Little change was predicted by 2030, with a slight decline by 2070. A key factor in modulating trends in disease was the level of cultivar resistance. However, no predictions were made on the impacts on yields.
The avoidance of crop loss due to disease is a major motivation for making improvements in disease management and prediction and for securing global food supplies (Savary et al. 2012, Savary et al. 2019, Savary and Willoquet, 2020), especially for pathogens of the major staple food crops such as wheat and rice (Ficke et al. 2018, Savary et al. 2022). Household surveys in Kenya have shown very high losses in maize due to maize lethal necrosis (MLN), approaching 80% in the western highland tropical and moist transitional zones (de Groote et al. 2021; Electronic Supporting Information, S5). Fewer quantitative data are available for pathogens of minor or speciality food crops, including pome fruit and grapevine (Trad and Taoueb 2021, Frem et al. 2021; Electronic Supporting Information, S5) although, because of higher per unit horticultural value, estimates of economic loss have been made. Losses of forest trees in nurseries or young plantations have often been expressed in terms of % mortality (Thu et al. 2021; Electronic Supporting Information, S6).
Analogous to yield in wild plant populations and communities is the provision of ecosystem services (Paseka et al. 2020, Windsor et al. 2021, Cavender-Bares et al. 2022, Nowak-Olejnik et al. 2022) recognising that there may also be ecosystem disservices, i.e., negative effects (Guo et al. 2022). Losses in the provision of ecosystem services are then analogous to crop losses. However, this aspect has rarely been reported, and most studies have concentrated on the role of wild plants as reservoirs or alternate hosts of pathogens infecting crop or cultivated plants (Mugerwa et al., 2021, Doolotkeldieva et al. 2021, Pak et al., 2021, Regassa et al., 2021, Chen et al., 2021; Electronic Supporting Information, S5).
Outlook
Clearly, more investment is needed in plant health monitoring, not just to accurately characterise presence or absence at a country level, but also to monitor spatial distribution and temporal changes for improved epidemiological intelligence (Ristaino et al., 2021). This is sorely needed for rapid responses to new and emerging diseases, particularly in low- and middle-income countries where the impacts can be particularly severe (Carvajal-Yepes et al., 2019). Better sources of data on disease presence and particularly absence, would allow predictive mathematical models to be properly parameterised, which remains challenging in practice (Cunniffe et al., 2015). It would also allow novel outbreaks of re-emerging diseases which can no longer be controlled in traditional ways (Bhattacharya et al., 2017) to be targeted. New pathogen detection technologies, for example with the application of remote sensing (Zarco-Tejada et al., 2018) are being developed, and could potentially feed into more cost effective and widespread monitoring. There are also opportunities to include diffuse data sources into the information stream concerning the spread of pathogens, including, for example, citizen science (Brown et al., 2020), landowner reports (Defra, 2020) and results of horizon scanning activities by regulators (EFSA, 2022).
This study has confirmed the value of two complementary approaches to identifying current plant disease outbreaks and potential new risks through publications in the peer-reviewed scientific literature (Objective 1), and new reports in an extensive database (Objective 2). These two approaches give different perspectives on current and future plant disease threats. It is noteworthy that only three pathogens, Ca. Liberibacter asiaticus, citrus tristeza virus and Fusarium oxysporum f. sp. cubense, were identified in both approaches, reflecting their continuing impact in areas they have long been established and their spread to new areas where previously absent. Six of the 14 pathogens reported extensively in 2021 in the scientific literature are vectored by insects. The role of vectors in disease transmission, epidemiology and spread needs as much attention as the pathogen. A good example of this concern is the African citrus psyllid Trioza erytreae, first recognised as having spread to Europe in the last decade (Cocuzza et al. 2017), and which now poses the risk of allowing huanglongbing to establish. Although the pathogens identified are well-known and characterised, they (or their vectors) continue to spread. The question remains which of the pathogens identified as new records in databases will in future years cause major disease outbreaks and be reported extensively in the scientific literature.
Very few pathogens spread intercontinentally through natural dispersal, mostly rust fungi such as sugarcane rust and cereal rusts (Jeger 1999, Brown and Hovmøller 2002). In many cases the role of international trade in plants and plant products can seed subsequent establishment, spread and impact of pathogens (Brasier, et al. 2008, Jeger et al. 2021). A traded plant may carry a pathogen to a new location, or a naïve introduced plant may acquire a new pathogen in the new location. Similarly, a vector may move with the traded plant, or an invasive pathogen may acquire a new vector. Numerous examples of the introductions of exotic and emerging pathogens can be found in the listings of pathogens identified in Objective 2 of this review. Finally, problems remain of delays in reporting, publishing, and communicating plant disease outbreaks and new records for plant health regulation, disease management and farm strategic planning. We suggest that the approach taken in this review provides a mechanism for integrating information and a synoptic view on current plant disease outbreaks and potential future risks and might usefully be adopted regularly in the future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
ACKNOWLEDGEMENTS
We thank Alice Campain for assistance in preparing Figure 5, Bryony Taylor for useful discussions, Nian Wang for encouraging us to write this article, and the four anonymous reviewers who provided very helpful suggestions to help us sharpen the focus of our work.
OPEN ACCESS STATEMENT
For the purpose of open access, the authors have applied a Creative Commons Attribution (CC BY) licence to any Author Accepted Manuscript version arising from this submission.
References
- Abrahamian, P.E.; Sobh, H.; Seblani, R.; Abou-Jawdah, Y. Coinfection of two criniviruses and a begomovirus enhances the disease severity in cucumber. Eur. J. Plant Pathol. 2015, 142, 521–530. [Google Scholar] [CrossRef]
- Amari, K.; Huang, C.; Heinlein, M. Potential impact of global warming on virus propagation in infected plants and agricultural productivity. Front. Plant Sci. 2021, 12, 649768. [Google Scholar] [CrossRef]
- Agrios, G. Agrios, G. Plant diseases caused by procaryotes: bacteria and mollicutes. Chapter 12 in Plant Pathology (Fifth Edition) Elsevier Press inc. 2005, pp 615-703.
- Ares, A.; Pereira, J.; Garcia, E.; Costa, J.; Tiago, I. The leaf bacterial microbiota of female and male kiwifruit plants in distinct seasons: assessing the impact of Pseudomonas syringae pv. Actinidiae Phytobiomes J. 2021, 5, 275–287. [Google Scholar] [CrossRef]
- Bastos, L.S.; Economou, T.; Gomez, M.F.C.; Villela, D.A.M.; Coehlo, F.C.; Cruz, O.G.; et al. A modelling approach for correcting reporting delays in disease surveillance data. Stat. Med. 2019, 38, 4363–4377. [Google Scholar] [CrossRef] [PubMed]
- Bebber, D.P. Climate change effects on Black Sigatoka disease of banana. Philos. Trans. R. Soc. B 2019, 374, 20180269. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Change 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Bebber, D.P.; Holmes, T.; Gurr, S.J. The global spread of plant pests and pathogens. Glob. Ecol. Biogeogr. 2014, 23, 1398–1407. [Google Scholar] [CrossRef]
- Bebber, D.P.; Field, E.; Gui, H.; Mortimer, P.; Holmes, T.; Gurr, S.J. Many unreported crop pests and pathogens are probably already present. Glob. Change Biol. 2019, 25, 2703–2713. [Google Scholar] [CrossRef] [PubMed]
- Bernard, G.C.; Egnin, M.; Bonsi, C. ‘The Impact of Plant-Parasitic Nematodes on Agriculture and Methods of Control’, in M. M. Shah, M. Mahamood (eds.), Nematology – Concepts, Diagnosis and Control, IntechOpen, London. 2017.
- Beyer, M.; Marozsak, B.; Dam, D.; Parisot, O.; Pallez-Barthel, M.; Hoffmann, L. Enhancing Septoria leaf blotch forecasts in winter wheat II: model architecture and validation results. J. Plant Dis. Prot. 2022, 129, 45–51. [Google Scholar] [CrossRef]
- Bhattacharya, S. Deadly new wheat disease threatens Europe’s crops. Nature 2017, 542, 145–146. [Google Scholar] [CrossRef]
- Bibi, M.; Hanif, M.K.; Sarwar, M.U.; Khan, M.I.; Khan, S.Z.; Shivachi, C.S.; et al. Monitoring population phenology of Asian citrus psyllid using deep learning. Complexity: online 2021, 4644213.
- Brasier, C.M. The biosecurity threat to the UK and global environment from international trade in plants. Plant Pathol. 2008, 57, 792–808. [Google Scholar] [CrossRef]
- Brown, J.K.M. and Hovmøller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.; Perez-Sierra, A.; Crow, P.; Parnell, S. The role of passive surveillance and citizen science in plant health. CABI Agric. Bioscience. 2020, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Burdon, J.J.; Zhan, J. Climate change and disease in plant communities. PloS Biol. 2020, 18, e3000949. [Google Scholar] [CrossRef] [PubMed]
- CABI, 2023. Cucurbit chlorotic yellows virus [original text by Maliogka, V.; Orfanidou, C. and Katis, N.]. In: Invasive Species Compendium. Wallingford, UK: CAB International. www.cabi.org/isc.
- CABI, 2023. Apple hammerhead viroid [Distribution map]. In: Distribution Maps of Plant Diseases. Wallingford: UK: CAB International https://www.cabidigitallibrary.org/journal/dmpd.
- Carvajal-Yepes, M.; Cardwell, K.; Nelson, A.; Garrett, K.; Giovani, B.; Saunders, D.G.O. A global surveillance system for crop diseases. Science 2019, 364, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Cavender-Bares, J.M.; Nelson, E.; Meireles, J.E.; Lasky, J.R.; Miteva, D.A.; Nowak, D.J.; et al. The hidden value of trees: quantifying the ecosystem services of tree lineages and their major threats across the contiguous US. PloS Sustain. Transform. 2022, 1, e0000010. [Google Scholar] [CrossRef]
- Chaloner, T.M.; Gurr, S.J.; Bebber, T.P. Plant infection risk tracks global crop yields under climate change. Nat. Clim. Change 2021, 11, 710–715. [Google Scholar] [CrossRef]
- Charoenvilaisiri, S.; Seepiban, C.; Kumpoosiri, M.; Rukpratanporn, S.; Warin, N.; Phuangrat, B.; et al. Development of a triple antibody sandwich enzyme-linked immunosorbent assay for cassava mosaic disease detection using a monoclonal antibody to Sri Lankan cassava mosaic virus. Virol. J. 2021, 18, 100. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Cao, C.; Fang, Z.; Jiang, H.; Fang, X.; Bao, S. Potential occurrence risk prediction of sudden oak death under different future climate scenarios based on SVM model. IEEE International Geoscience and Remote Sensing Symposium. IGARSS 2018 Valencia, Spain, July 22-27, 2018. 5228-5231.
- Chen, W.; Zhang, Z.D.; Ma, X.; Zhang, G.; Yao, Q.; Kang, Z.; et al. Phenotyping and genotyping analyses reveal the spread of Puccinia striiformis f. sp tritici aeciospores from susceptible barberry to wheat in Qinghai of China. Front. Plant Sci. 2021, 12, 764304. [Google Scholar] [CrossRef]
- Cocuzza, G.E.M.; Alberto, U.; Hernández-Suárez, E.; Siverio, F.; Di Silvestro, S.; Tena, A. A review on Trioza erytreae (African citrus psyllid), now in mainland Europe, and its potential risk as vector of huanglongbing (HLB) in citrus. J. Pest Sci. 2017, 90, 1–17. [Google Scholar] [CrossRef]
- Cunniffe, N.J.; Koskella, B.; Metcalf, J.E.; Parnell, S.R.; Gottwald, T.R. and Gilligan, C.A. Thirteen challenges in modelling plant diseases. Epidemics 2015, 10, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. magnum and C. orchidearum species complexes. Studies in Mycology 2019, 92, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Dean, R. , van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kossack, K.; Pietro, A.D.; Spanu, P.D. et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathology. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Defra, 2020. Agricultural Transition Plan 2021 to 2024. https://www.gov.uk/government/publications/agricultural-transition-plan-2021-to-2024.
- De Groote, H.; Munyua, B.G.; Palmas, S.; Suresh, L.M.; Bruce, A.Y.; Kimenju, S. Using panel community surveys to track the impact of crop pests over time and space – the case of maize lethal necrosis (MLN) disease in Kenya from 2013-2018. Plant Disease 2021, 105, 1259–1271. [Google Scholar] [CrossRef] [PubMed]
- Devorshak, C. Terminology used in pest risk analysis. In Plant Pest Risk Analysis: Concepts and Application (ed. C. Devorshak) CAB International, Wallingford, UK. 2012. Pp 45-56.
- Ding, X.; Guo, Y.; Ye, J.; Wu, X.; Lin, S.; Chen, F. Population differentiation and epidemic tracking of Bursaphelenchus xylophilus in China based on chromosome-level assembly and whole-genome sequencing data. Pest Management Science: published online 17 December 2021.
- Doolotkeldieva, T.; Bobushova, S.; Carnal, S.; Rezzonico, F. Genetic characterization of Erwinia amylovora isolates detected in the wild walnut-fruit forests of South Kyrgyzstan. J. Plant Pathol. 2021, 103, 109–120. [Google Scholar] [CrossRef]
- Dupes, E.; Durand, K.; Rieux, A.; Briand, M.; Pruvost, O.; Cunty, A.; et al. 2022. Two bridgehead invasions of Xylella fastidiosa subsp. Multiplex in France. bioRxiv. [CrossRef]
- EFSA. Guidelines for statistically sound and risk-based surveys of Xylella fastidiosa. EFSA J. 2020, 7, 1873. [Google Scholar]
- EFSA, 2022. Plant Health Newsletter on Horizon Scanning. [CrossRef]
- EFSA PLH Panel. Scientific Opinion on the pest categorisation of Colletotrichum aenigma, C. alienum, C. perseae, C. siamense and C. theobromicola. EFSA J. 2022, 20, 7529. [Google Scholar] [CrossRef]
- Esmail, S.M.; Draz, I.S.; Ashmawy, M.A.; El-Oraby, W.M. Emergence of new aggressive races of Puccinia striiformis f. sp. Tritici causing yellow rust epiphytotic in Egypt. Physiol. Mol. Plant Pathol. 2021, 114, 101612. [Google Scholar] [CrossRef]
- Espinosa-Zaragoza, S.; Pérez-De la, O.; Aquirre-Medina, J.F.; López-Martínez, V. Does the African citrus psyllid, Trioza erytreae, represent a phytosanitary threat to the citrus industry in Mexico? Insects 2021, 12, 450. [Google Scholar] [CrossRef]
- Eunice, J.; Miano, D.; Muiru, W.M.; Mutitu, E.; Macharia, I. Status of maize lethal necrosis disease in seed production systems in Kenya. Cogent Food Agric. 2021, 7, 1918406. [Google Scholar] [CrossRef]
- Ficke, A.; Cowger, C.; Bergstrom, G.; Brodal, G. Understanding yield loss and pathogen biology to improve disease management: Septoria nodorum blotch – a case study in wheat. Plant Dis. 2018, 102, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Fones, H.N.; Bebber, D.P.; Chaloner, T.M.; Kay, W.T.; Steinberg, G.; Gurr, S.J. Threats to global food security from emerging fungal and oomycete crop pathogens. Nat. Food 2020, 1, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Frem, M.; Fucilli, V.; Nigro, F.; El Moujabber, M.; Abou Kubaa, R.; La Notte, P.; et al. The potential direct economic impact and private management costs of an invasive alien species: Xylella fastidiosa on Lebanese wine grapes. NeoBiota 2021, 70, 43–67. [Google Scholar] [CrossRef]
- Garcia-Figuera, S.; Deniston-Sheets, H.; Grafton-Cardwell, E.; Babcock, B.; McRoberts, N. Perceived vulnerability and propensity to adopt best management practices for Huanglongbing disease of citrus in California. Phytopathology 2021, 111, 1758–1773. [Google Scholar] [CrossRef] [PubMed]
- Garnault, M.; Duplaix, C.; Leroux, P.; Couleaud, G.; David, O.; Walker, A.-S.; et al. Large-scale study validates that regional fungicide applications are major determinants of resistance evolution in the wheat pathogen Zymoseptoria tritici in France. New Phytol. 2021, 229, 3508–3521. [Google Scholar] [CrossRef] [PubMed]
- GBIF Secretariat. 2021. GBIF Backbone Taxonomy. Checklist dataset https://doi.org/10.15468/39omei accessed via GBIF.org on 2022-05-23. [CrossRef]
- Godefroid, M.; Morente, M.; Schartel, T.; Cornara, D.; Purcell, A.; Gallego, D.; et al. Climate tolerances of Philaenus spumarious should be considered in risk assessments of disease outbreaks related to Xylella fastidiosa. J. Pest Sci. 2022, 95, 855–868. [Google Scholar] [CrossRef]
- Gougherty, A.V.; Davies, T.J. Towards a phylogenetic ecology of plant pests and pathogens. Philos. Trans. R. Soc. B 2021, 376, 20200359. [Google Scholar] [CrossRef] [PubMed]
- Grégorie, G.; Derderian, F.; Le Lorier, J. Selecting the language of the publications included in a meta-analysis: Is there a tower of Babel bias? J. Clin. Epidemiol. 1995, 48, 159–163. [Google Scholar] [CrossRef]
- Gyoutoku, Y.; Okazaki, S.; Furuta, A.; Etoh, T.; Mizobe, M.; Kuno, K.; Hayashida, S.; Okuda, M. Chlorotic yellows disease of melon caused by Cucurbit chlorotic yellows virus, a new crinivirus. Jpn. J. Phytopathology. 2009, 75, 109–111. [Google Scholar] [CrossRef]
- Guo, R.-Z.; Song, Y.-B.; Dong, M. Progress and prospects of ecosystem disservices: an updated literature review. Sustainability 2022, 14, 10396. [Google Scholar] [CrossRef]
- Hamborg, Z.; Blystad, D. First Report of Tomato Brown Rugose Fruit Virus in Tomato in Norway. Plant Disease 2022, 106, 1773–2006. [Google Scholar] [CrossRef] [PubMed]
- Hampf, A.C.; Nendel, C.; Strey, S.; Strey, R. Biotic yield losses in the southern Amazon, Brazil: making use of smartphone-assisted plant disease diagnosis data. Front. Plant Sci. 2021, 12, 621168. [Google Scholar] [CrossRef]
- Haynes, A.F. Presence of N-fixing neighbors increases leaf N and δ13C in Castilleja applegatei, a root hemiparasite. Plant Ecol. 2022, 223, 213–228. [Google Scholar] [CrossRef]
- He, D.; Zhan, J.; Xie, L. Problems, challenges and future of plant disease management: from an ecological point of view. J. Integr. Agric. 2016, 15, 705–715. [Google Scholar] [CrossRef]
- Heck, D.W.; Dita, M.; Del Ponte, E.M.; Mizubuti, E.S.G. Incidence, spatial pattern and temporal progress of Fusarium wilt of bananas. J. Fungi 2021, 7, 646. [Google Scholar] [CrossRef]
- Hernandez, R.N.; Isakeit, T.; Al Rwahnih, M.; Hernandez, R. and Alabi, O.J. First Report of Cucurbit chlorotic yellows virus Infecting Cantaloupe(Cucumis melo) in Texas. Plant Disease. 2021, 105, 3313. [Google Scholar] [CrossRef]
- Hjelkrem, A.-G.; R. , Ficke, A.; Abrahamsen, U.; Hofgaard, I.S.; Brodal, G. Prediction of leaf blotch disease risk in Norwegian spring wheat based on weather factors and host phenology. Eur. J. Plant Pathol. 2021, 160, 199–213. [Google Scholar] [CrossRef]
- Hyatt-Twynam, S.R.; Parnell, S.; Stutt, R.O.J.H.; Gottwald, T.R.; Gilligan, C.A.; Cunniffe, N.J. Risk-based management of invading plant disease. New Phytol. 2017, 214, 1317–1329. [Google Scholar] [CrossRef]
- Jeger, M.J. Improved understanding of dispersal in crop pest and disease management: current status and future directions. Agric. For. Meteorol. 1999, 97, 331–349. [Google Scholar] [CrossRef]
- Jeger, M.J. The impact of climate change on disease in wild plant populations and communities. Plant Pathology. 2021, 71, 111–130. [Google Scholar] [CrossRef]
- Jeger, M.; Beresford, R.; Bock, C.; Brown, N.; Fox, A.; Newton, A.; et al. Global challenges facing plant pathology: multidisciplinary approaches to meet the food security and environmental challenges in the mid-twenty-first century. CABI Agric. Biosci. 2021, 2, 20. [Google Scholar] [CrossRef]
- Jiang, J.; Abbott, K.C.; Baudena, M.; Eppinga, M.B.; Umbanhowar, J.A.; Bever, J.D. Pathogens and mutualists as joint drivers of host species coexistence and turnover: implications for plant competition and succession. Am. Nat. 2020, 195, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Jones, R. Plant virus disease pandemics and epidemics. Plants 2021, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Katono, K.; Macfadyen, S.; Omongo, C.A.; Odong, T.L.; Colvin, J.; Karungi, J.; et al. Influence of cassava morphological traits and environmental conditions on field populations of Bemisia tabaci. Insects 2021, 12, 604. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Do, K.S.; Park, J.H.; Kang, W.S.; Lee, W.H.; Park, E.W. Application of numerical weather prediction data to estimate infection risk of bacterial grain rot of rice in Korea. Plant Pathol. J. 2020, 36, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Ladisa, G.; Bogliotti, C.; Calabrese, G.; Kalaitzidis, C.; Livieratos, I.; Owen, C.; et al. Stakeholders’ perception of Xylella fastidiosa (Xf) disease risk assessment: First results from Puglia (IT), Chania (GR), Valencia and Andalucia (ES). Open Journal of Social Sciences 2021, 9, 188–223. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Venturi, V. Synergisms between microbial pathogens in plant disease complexes: a growing trend. Front. Plant Sci. 2015, 6, 385. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, E.; Sese, M.; Lopez-Quilez, A.; Conesa, D.; Dalmau, V.; Ferrer, A.; et al. Tracking the outbreak: an optimised sequential adaptive strategy for Xylella fastidiosa delimiting surveys. Biol. Invasions 2021, 23, 3243–3261. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, G.H.; Koh, Y.J.; Jung, J.S. Identification of strA – strB genes in streptomycin-resistant Pseudomonas syringae pv. Actinidiae biovar 2 strains isolated in Korea. Plant Pathol. J. 2021, 37, 489–493. [Google Scholar]
- León, M.; Berbegal, M.; Abad-Campos, P.; Ramón-Albalat, A.; Caffi, T.; Rossi, V.; et al. Evaluation of sown cover crops and spontaneous weed flora as a potential reservoir of black-foot pathogens in organic Viticulture. Biology 2021, 10, 498. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Nangong, Z. Precision trunk injection technology for treatment of huanglongbing (HLB)-affected citrus trees – a review. J. Plant Dis. Prot. 2021, 29, 15–34. [Google Scholar]
- Li, J.; Liang, X.; Wang, X.; Shi, Y.; Gu, Q.; Kuo, Y.-W.; Falk, B.W.; Yan, F. Direct evidence for the semipersistent transmission of Cucurbit chlorotic yellows virus by a whitefly vector. Sci. Rep. 2016, 6, 36604. [Google Scholar] [CrossRef] [PubMed]
- Lops, F. Mitigation and adaption to climate change of plant pathogens. J. Plant Sci. Phytopathol. 2022, 6, 101–102. [Google Scholar]
- Mannino, M.R.; Laurenaudie, M.; Ling, J.P.; Candresse, T.; Miret, J.A.J.; Jeger, M.J.; et al. 2021. Horizon scanning for plant health: report on 2017-2020 activities. EFSA Supporting Publication EN-2010, 113 pp.
- Mastin, A.J.; Gottwald, T.R. , van den Bosch, F.; Cunniffe, N.J.; Parnell, S.R. Optimising risk-based surveillance for early detection of invasive plant pathogens. PLOS Biol. 2020, 18, e3000863. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.A.; Lopez-Guerrero, M.G.; Guo, M.; Schmer, M.R.; Herr, J.R.; Schnable, J.C. Rhizosphere microbiomes in a historical maize-soybean rotation system respond to host species and nitrogen fertilization at the genus and subgenus levels. Appl. Environ. Microbiol. 2022, 87, e03132–20. [Google Scholar]
- Méthot, P.O.; Alizon, S. What is a pathogen? Toward a process view of host-parasite interactions. Virulence 2014, 5, 775–85. [Google Scholar] [CrossRef]
- Mugerwa, H.; Colvin, J.; Alicai, T.; Omongo, C.A.; Kabaalu, R.; Visendi, P.; et al. Genetic diversity of whitefly (Bemisia tabaci spp.) on crop and uncultivated plants in Uganda: implications for the control of this devastating pest species complex in Africa. J. Pest Sci. 2021, 94, 1307–1330. [Google Scholar] [CrossRef]
- Nowak-Olejnik, A.; Mocior, E. Provisioning ecosystem services of wild plants collected from seminatural habitats: a basis for sustainable livelihood and multifunctional landscape conservation. Mt. Res. Dev. 2021, 42, R11–R19. [Google Scholar] [CrossRef]
- Oladokun, J.O.; Halabi, M.H.; Barua, P. and Nath, P.D. Tomato brown rugose fruit disease: current distribution, knowledge and future prospects. Plant Pathology. 2019, 68, 1579–1586. [Google Scholar] [CrossRef]
- Olmo, D.; Nieto, A.; Borràs, D.; Montesinos, M.c.F.; Pascual, A.; et al. Landscape epidemiology of Xylella fastidiosa in the Balearic Islands. Agronomy 2021, 11, 473. [Google Scholar] [CrossRef]
- Orelana-Torrejon, C.; Vidal, T.; Boixel, A.-L.; Gélisse, S.; Saint-Jean, S.; Suffert, S. Annual dynamics of Zymoseptoria tritici in wheat cultivar mixtures: A compromise between the efficacy and durability of a recently broken-down resistance gene? Plant Pathol. 2022, 71, 289–303. [Google Scholar] [CrossRef]
- Pak, D.; You, M.P.; Lanoiselet, V.; Barbetti, M.J. Management of rice blast (Pyricularia oryzae): implications of alternative hosts. Eur. J. Plant Pathol. 2021, 161, 343–355. [Google Scholar] [CrossRef]
- Park, J.-W. , da Graça, J.V.; Sétamou, M.; Kunta, M. Diversity of Citrus tristeza virus strains in the upper gulf coast area of Texas. Plant Dis. 2021, 105, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Paseka, R.E.; White, L.E.; Van der Waal, D.B.; Strauss, A.T.; González, A.L.; Everett, R.A.; et al. Disease-mediated ecosystem services: pathogens, plants, and people. Trends Ecol. Evol. 2020, 35, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Perronne, R.; Dubs, F. , de Vallavieille-Pope, C.; Leconte, M., du Cheyron, P.; Cadot, V.; et al. Spatiotemporal changes in varietal resistance to wheat yellow rust in France reveal an increase in field resistance level during the period 1985-2018. Phytopathology 2021, 111, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Regassa, B.; Abraham, A.; Fininsa, C.; Wegary, D. Alternate hosts and seed transmission of maize lethal necrosis in Ethiopia. J. Phytopathol. 2021, 169, 303–315. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Anderson, P.K.; Bebber, D.P.; Braumann, K.A.; Cunniffe, N.J.; Federoff, N.V.; et al. The persistent threat of emerging plant disease pandemics to food security. PNAS 2021, 118, e2022239118. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, D.M.; Lichtveld, M.; Mazet, J.A.K.; Togami, E.; Miller, S.A. Plant Health and its effects on food safety and security in a One Health framework: four case studies. One Health Outlook 2021, 3, 6. [Google Scholar] [CrossRef]
- Robles-Fernández, Á.; L. ; Lira-Noriega, A. Combining phylogenetic and occurrence information for risk assessment of pest and pathogen interactions with host plants. Front. Appl. Math. Stat. 2017, 3, 17. [Google Scholar] [CrossRef]
- Sabri, K.; Mokabli, A.; Mokrini, F.; Khayi, S.; Laasli, S.-E.; Smaha, D.; et al. Diversity of nematophagous fungi associated with vegetable crops in Northern Algeria. Arch. Phytopathol. Plant Prot. 2022, 55, 405–419. [Google Scholar] [CrossRef]
- Savary, S.; Willoquet, L. Modeling the impact of crop diseases on global food security. Annu. Rev. Phytopathol. 2020, 58, 313–341. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Ficke, A.; Aubertot, J.-N.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Willocquet, L.; Castilla, N.; Nelson, A.; Singh, U.S.; Kumar, J.; et al. Whither rice health in the lowlands of Asia: Shifts in production situations, injury profiles, and yields. Plant Pathol. 2022, 71, 55–85. [Google Scholar] [CrossRef]
- Sekiya, N.; Nakajima, T.; Oizumi, N.; Kurosawa, C.; Tibanyendela, N.; Peter, M.A. Agronomic practices prevented local outbreaks of rice yellow mottle virus disease revealed by spatial autoregressive analysis. Agron. Sustain. Dev. 2022, 42, 15. [Google Scholar] [CrossRef]
- Severns, P.M.; Mundt, C.C. Delays in epidemic outbreak control cost disproportionately large treatment footprints to offset. Pathogens 2022, 11, 393. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.A.; Paul, P.A.; De Wolf, E.D.; Madden, L.V. Predicting plant disease epidemics from functionally represented weather series. Philos. Trans. R. Soc. B 2019, 374, 20180273. [Google Scholar] [CrossRef] [PubMed]
- Shan, H.L.; Cang, X.Y.; Qin, W.; Li, W.F.; Wang, X.Y.; Zhang, R.Y.; et al. Resistance to pokkah boeng disease in new and main sugarcane varieties cultivated in China. Euphytica 2021, 217, 194. [Google Scholar] [CrossRef]
- Shan, A. , Pan, A., Kang, K.J.; Pan, M., Wang, G., Wang, M.; et al. Effects of straw return with N fertilizer reduction on crop yield, plant diseases and pests and potential heavy metal risk in a Chinese rice paddy: A field study of 2 consecutive wheat-rice cycles. Environ. Pollut. 2021, 288, 117741. [Google Scholar] [CrossRef]
- Sherman, J.; Burke, J.M.; Gent, D.H. Cooperation and coordination in plant disease management. Phytopathology 2019, 109, 1720–1731. [Google Scholar] [CrossRef]
- Shrestha, B.; Martini, X.; Stelinski, L.L. Population fluctuations of Diaphorina citri and its natural enemies in response to various management practices in Florida. Fla. Entomol. 2022, 104, 178–185. [Google Scholar] [CrossRef]
- Skelton, A.; Buxton-Kirk, A.; Fowkes, A.; Harju, V.; Forde, S.; Ward, R. Potato spindle tuber viroid detected in seed of uncultivated Solanum anguivi, S. coagulans and S. dasyphyllum collected from Ghana, Kenya and Uganda. New Dis. Rep. 2019, 39, 23. [Google Scholar] [CrossRef]
- Singh, J.; Cobb-Smith, D.; Higgins, E.; Khan, A. Comparative evaluation of lateral flow immunoassays, LAMP, and quantitative PCR for diagnosis of fire blight in apple orchards. J. Plant Pathol. 2021, 103, S131–S142. [Google Scholar] [CrossRef]
- Soubeyrand, S. , de Jerphanion, P.; Martin, O.; Saussac, M.; Manceau, C.; Hendrikx, P. et al. Inferring pathogen dynamics from temporal count data: the emergence of Xylella fastidiosa in France is probably not recent. New Phytol. 2018, 219, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Sreenivas, S.S. Impact of climate change on plant pathogenic fungi, bacteria and viruses: a review. Asian J. Res. Rev. Agric. 2022, 4, 35–45. [Google Scholar]
- Szyniszewska, A.M.; Chikoti, P.C.; Tembo, M.; Mulenga, R.; Gilligan, C.A. , van den Bosch, F.; et al. Smallholder cassava planting material movement and grower behavior in Zambia: implications for the management of cassava virus diseases. Phytopathology 2021, 111, 1952–1962. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Cao, X.; Xu, X.; Jiang, Y.; Luo, Y.; Ma, Z.; et al. Effects of climate change on epidemics of powdery mildew in winter wheat in China. Plant Dis. 2017, 101, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Heck, D.; Nomura, E.; Viera, H.; Dita, M. Soil attributes, plant nutrition, and Fusarium wilt of banana in São Paulo, Brazil. Trop. Plant Pathol. 2021, 46, 443–454. [Google Scholar] [CrossRef]
- Thompson, R.N.; Gilligan, C.A.; Cunniffe, N.J. Control fast or control smart: When should invading pathogens be controlled? PloS Comput. Biol. 2018, 14, e1006014. [Google Scholar] [CrossRef]
- Thu, P.Q.; Quang, D.N.; Chi, N.M.; Hung, T.X.; Binh, L.V.; Dell, B. New and emerging insect pest and disease threats to forest plantations in Vietnam. Forests 2021, 12, 1301. [Google Scholar] [CrossRef]
- Trad, M. and Taoueb, S.M. A critical review of pear genetic resources in Tunisia after the fire blight outbreak: risk assessment and geographical limits. Euro-Mediterr. J. Environ. Integr. 2021, 6, 50. [Google Scholar] [CrossRef]
- Várallyay, E.; Přibylová, J.; Galbacs, Z.N.; Jahan, A.; Varga, T.; Špak, J.; et al. Detection of Apple Hammerhead Viroid, Apple Luteovirus 1 and Citrus Concave Gum-Associated Virus in Apple Propagation Materials and Orchards in the Czech Republic and Hungary. Viruses. 2022, 14, 2347. [Google Scholar] [CrossRef] [PubMed]
- van den Vossenberg, B.T.L.H.; Visser, M.; Bruinsma, M.; Koenraadt, H.M.S.; Westenberg, M. and Botermans, M. Real-time tracking of Tomato brown rugose fruit virus (ToBRFV) outbreaks in the Netherlands using Nextstrain. PLOS ONE. 2020, 15, e0234671. [Google Scholar]
- Vilvert, E.; Stridh, L.; Andersson, B.; Olson, A.; Aldén, L.; Berlin, A. Evidence based disease control methods in potato production: a systematic map protocol. Environ. Evid. 2022, 11, 6. [Google Scholar] [CrossRef]
- Vincent, C.; Guha, A.; Killiny, L.N.; Diepenbrock, L. Understorey environment promotes photosynthetic efficiency and mitigates severity and function of an introduced, vectored pathosystem: a study of a feral citrus population in central Florida. Tree 2021, 35, 1711–1725. [Google Scholar] [CrossRef]
- Wang, H.; Mongiano, G.; Fanchini, D.; Titone, P.; Tamborini, L.; Bregaglio, S. Varietal susceptibility overcomes climate change effects on future trends of rice blast disease in Northern Italy. Agric. Syst. 2021, 193, 103233. [Google Scholar] [CrossRef]
- Wang, Y.-P.; Pan, Z.-C.; Yang, L.-N.; Burdon, J.L.; Friberg, H.; Sui, J.; et al. Optimising plant disease management in agricultural ecosystems through rational in-crop diversification. Front. Plant Sci. 2021, 12, 767209. [Google Scholar] [CrossRef]
- Windsor, F.M.; Tavella, J.; Rother, D.C.; Raimundo, R.L.G.; Devoto, M.; Guimarães, P.R.; et al. Identifying plant mixes for multiple ecosystem service provision in agricultural systems using ecological networks. J. Appl. Ecol. 2021, 58, 2770–2782. [Google Scholar] [CrossRef]
- Wintermantel, W.M.; Jenkins Hladky, L.L.; Fashing, P.; Ando, K. and McCreight, J.D. First Report of Cucurbit Chlorotic Yellows Virus Infecting Melon in the New World. D. First Report of Cucurbit Chlorotic Yellows Virus Infecting Melon in the New World. Plant Disease. 2019, 103. [Google Scholar]
- Wylder, B.; Briddle, M.; King, K.; Baden, R.; Webber, J. Evidence from mortality dating of Fraxinus excelsior indicates ash dieback (Hymenoscyphus fraxineus) was active in England in 2004–2005. For. Int. J. For. Res. 2018, 91, 434–443. [Google Scholar] [CrossRef]
- Yuen, J.; Mila, A. Landscape-scale disease risk quantification and prediction. Annu. Rev. Phytopathol. 2015, 53, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Zarco-Tejada, P.J.; Camino, C.; Beck, P.S.A.; Calderon, R.; Honero, A.; Hernandez-Clemente, R. Previsual symptoms of Xylella fastidiosa infection revealed in spectral plant-trait alterations. Nat. Plants 2018, 4, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Griffiths, J.S.; Marchand, G.; Bernards, M.A.; Wang, A. Tomato brown rugose fruit virus: an emerging and rapidly spreading plant RNA virus that threatens tomato production worldwide. Mol. Plant Pathol. 2022, 23, 1262–1277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dian, Y.; Zhou, J.; Peng, S.; Hu, Y.; Hu, L.; et al. Characterizing spatial patterns of pine wood nematode outbreaks in subtropical zone in China. Remote Sens. 2021, 13, 4682. [Google Scholar] [CrossRef]
- Zhang, Z.; Qi, S.; Tang, N.; Zhang, X.; Chen, S.; Zhu, P.; et al. Discovery of replicating circular RNAs by RNA-Seq and Computational Algorithms. PLOS Pathog. 2014, 10, e1004553. [Google Scholar] [CrossRef]
Figure 1.
Methodology for the survey of published articles on disease outbreaks and epidemics. The workflow and number of results after each stage of filtering for the review of the scientific literature published in 2021. Numbers represent the number of identified articles at each stage: the final review after all exclusions were applied summarises n = 186 articles.
Figure 1.
Methodology for the survey of published articles on disease outbreaks and epidemics. The workflow and number of results after each stage of filtering for the review of the scientific literature published in 2021. Numbers represent the number of identified articles at each stage: the final review after all exclusions were applied summarises n = 186 articles.
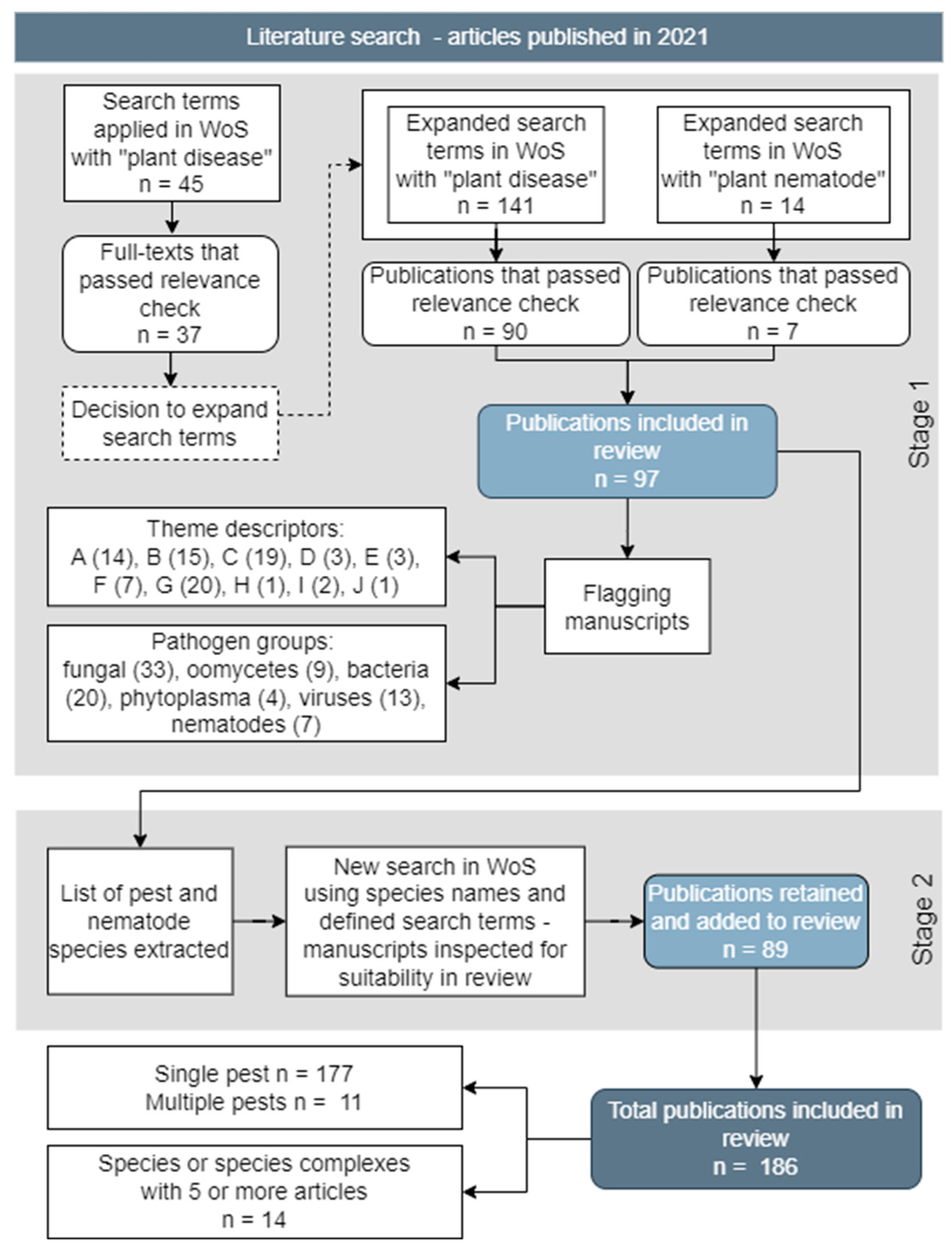
Figure 2.
Review of records in the CABI Distribution Database (DDB). The workflow and number of results from the exploration of the DDB for new species occurrence records added to the database in 2021. Numbers represent the number of identified records which consist of a concept and the related geographic area. Totals are also shown for the number of unique geographic areas and concepts at each stage of the process. A total of 28 pathogens had four or more new occurrence records in 2021 according to DDB. It was followed by manual review of all pathogens with more than 4 new records (n = 28), to identify confirmed first reports in a new location (n=15). “Concept”a = A generic name for an entity in the distribution database for which location and presence/absence data are stored. Most commonly an organism recorded at species level, but in some cases recording occurs at higher or lower taxonomic levels depending on data available. “Geographic area”b = One of 507 defined regions covering the globe of national or sub-national (e.g., state, province) administrative areas and other regions, islands and sea areas of biogeographic importance.
Figure 2.
Review of records in the CABI Distribution Database (DDB). The workflow and number of results from the exploration of the DDB for new species occurrence records added to the database in 2021. Numbers represent the number of identified records which consist of a concept and the related geographic area. Totals are also shown for the number of unique geographic areas and concepts at each stage of the process. A total of 28 pathogens had four or more new occurrence records in 2021 according to DDB. It was followed by manual review of all pathogens with more than 4 new records (n = 28), to identify confirmed first reports in a new location (n=15). “Concept”a = A generic name for an entity in the distribution database for which location and presence/absence data are stored. Most commonly an organism recorded at species level, but in some cases recording occurs at higher or lower taxonomic levels depending on data available. “Geographic area”b = One of 507 defined regions covering the globe of national or sub-national (e.g., state, province) administrative areas and other regions, islands and sea areas of biogeographic importance.
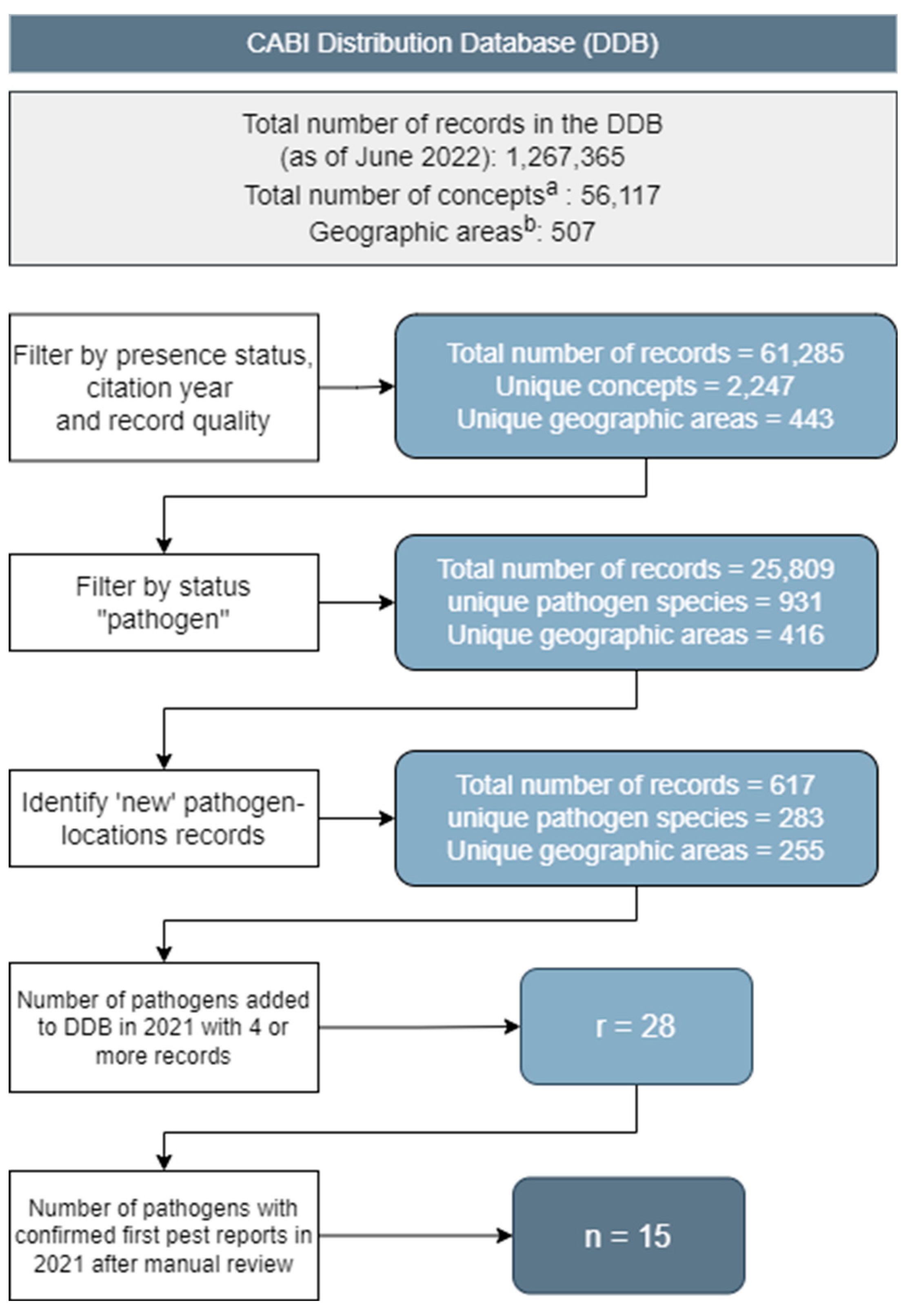
Figure 3.
Categorisation of core articles on single named species identified in Stage 1 of the review of published articles on disease outbreaks and epidemics. A – Number of host plants per continent where retrieved pathogen species were observed. B – Number of pathogen species per group of pathogens and host plants on which they were observed. C – Number of pathogen species per continent and study descriptor. D – Number of reported species grouped by study descriptors and pathogen group.
Figure 3.
Categorisation of core articles on single named species identified in Stage 1 of the review of published articles on disease outbreaks and epidemics. A – Number of host plants per continent where retrieved pathogen species were observed. B – Number of pathogen species per group of pathogens and host plants on which they were observed. C – Number of pathogen species per continent and study descriptor. D – Number of reported species grouped by study descriptors and pathogen group.
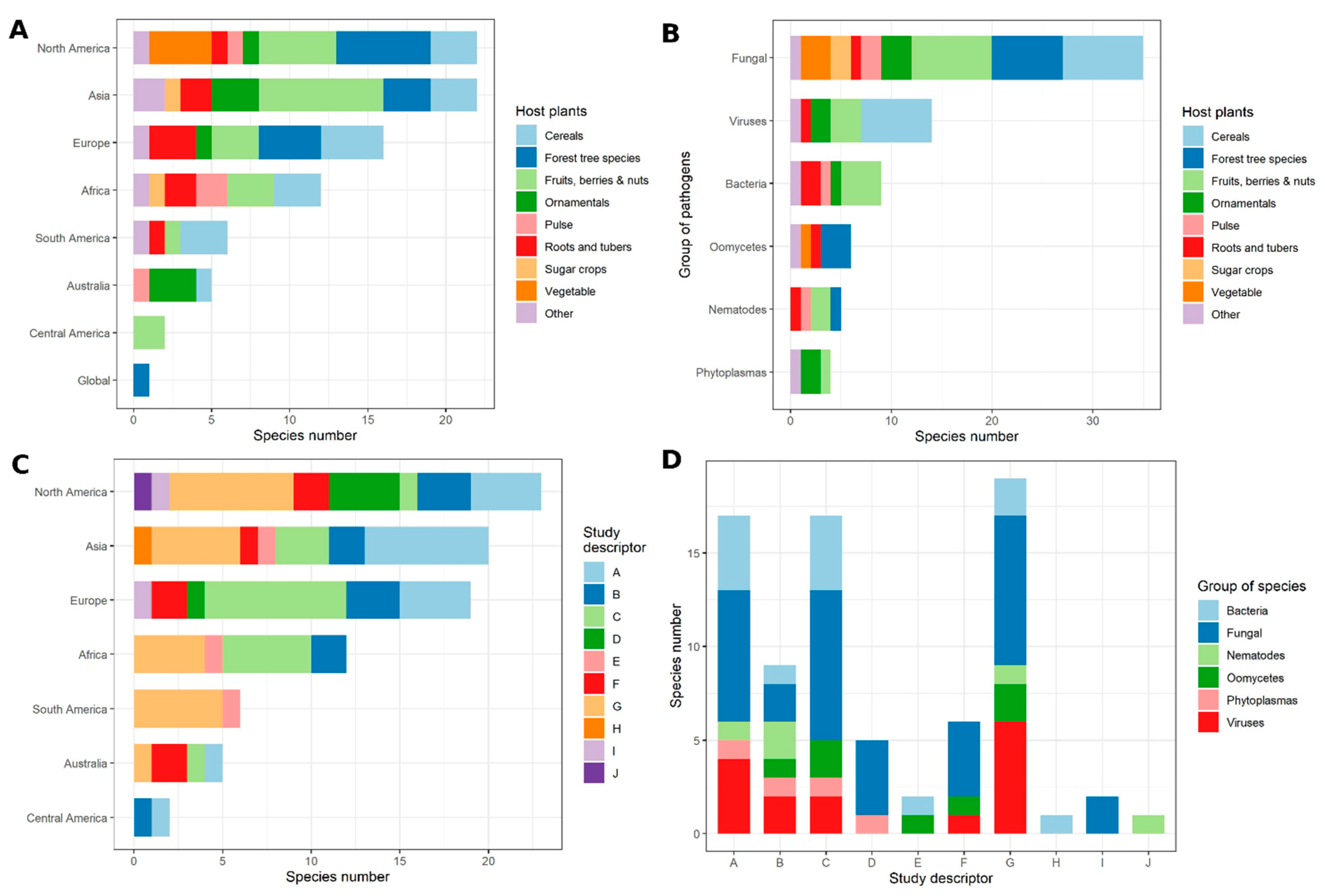
Figure 4.
Map showing number of pathogen-location pair records retrieved from core articles on single named species and multiple pathogens identified in Stage 1 of the review of published articles on disease outbreaks and epidemics. The number of records by continent and species type of pathogen. Two studies had global remit, therefore n=2 was added to each continent’s totals. A map showing country by country data is available in Electronic Supporting Information, S3 (Figure S3-1).
Figure 4.
Map showing number of pathogen-location pair records retrieved from core articles on single named species and multiple pathogens identified in Stage 1 of the review of published articles on disease outbreaks and epidemics. The number of records by continent and species type of pathogen. Two studies had global remit, therefore n=2 was added to each continent’s totals. A map showing country by country data is available in Electronic Supporting Information, S3 (Figure S3-1).
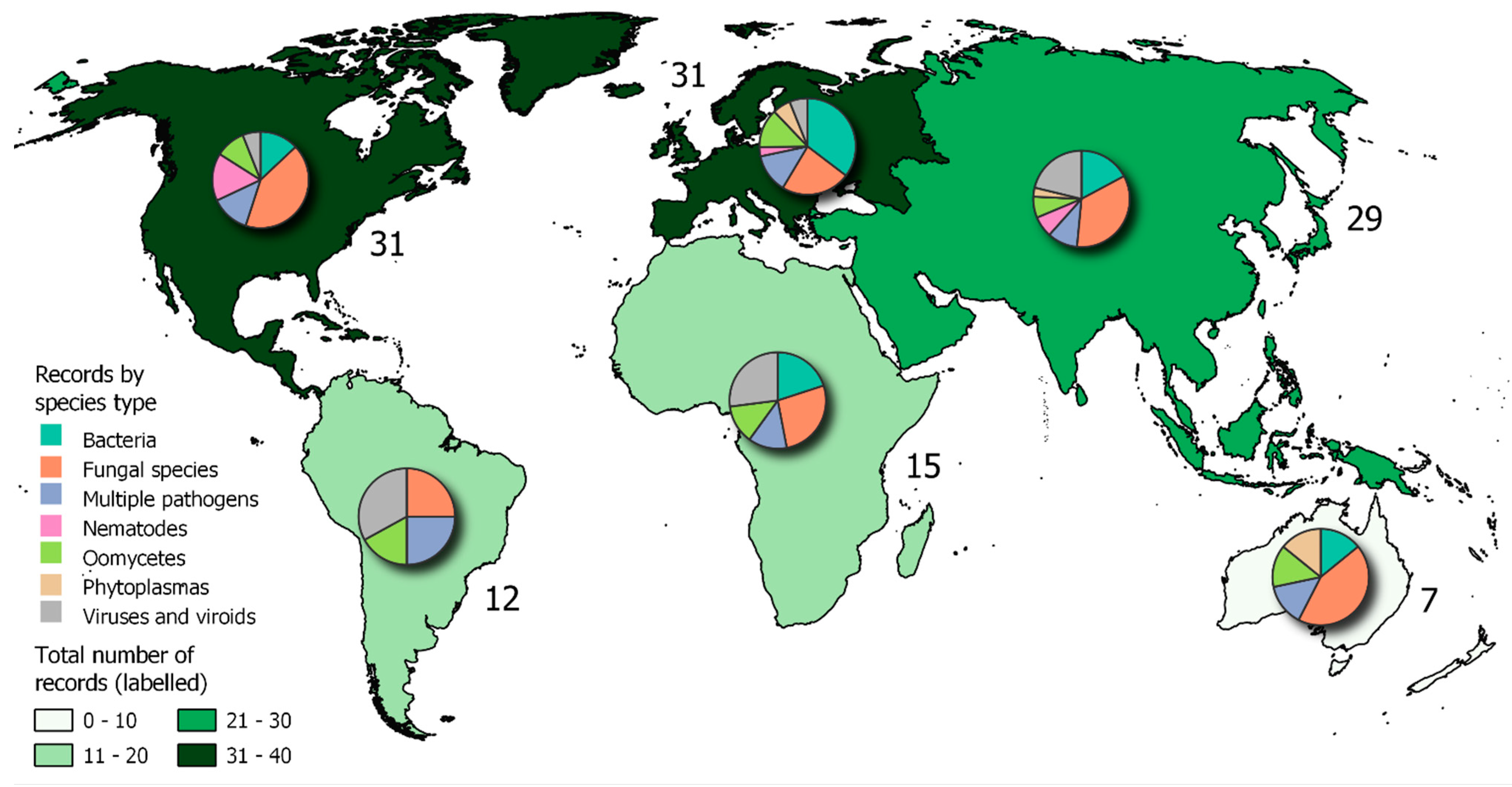
Figure 5.
Symptoms caused by the diseases identified in the review of published articles on disease outbreaks and epidemics. A) Fusarium oxysporum f. sp. cubense (Panama disease of banana); symptoms on a banana plant (Public Domain - Released by Scot Nelson/via flickr - CC0); B) Meloidogyne enterolobii (Pacara earpod tree root-knot nematode); rootknot symptoms on an ornamental Petunia species (©Jeffrey W. Lotz/Florida Department of Agriculture & Consumer Services/Bugwood.org - CC BY 3.0 US); C) Cassava mosaic disease (African cassava mosaic); symptoms on cassava leaf. (©Phil Taylor, CABI); D) Citrus tristeza virus (©Tim Gottwald); E) Erwinia amylovora (fireblight) on pear (©Phil Taylor, CABI); F) Candidatus Liberibacter asiaticus (Asian greening); HLB symptoms. April 2016 (©Nian Wang (Citrus Research and Education Center)); G) Pseudomonas syringae pv. syringae (bacterial canker) symptoms on peach (Prunus persica), bark peeled to show discoloration beneath (©University of Georgia Plant Pathology/University of Georgia/Bugwood.org - CC BY 3.0 US); H) Fusarium on wheat (©Phil Taylor, CABI); I) Magnaporthe oryzae (rice blast) in Myanmar (©Phil Taylor, CABI); J) Puccinia striiformis f. sp. tritici; symptoms of yellow rust on triticale leaf (©Rasbak/via Wikimedia Commons - CC BY-SA 4.0); K) Zymoseptoria tritici, septoria on wheat (©Phil Taylor, CABI); L) Symptoms of maize lethal necrosis disease (©Joseph Mulema, CABI); M) Bursaphelenchus xylophilus (pine wilt nematode); pine wilt symptoms, one year after death of tree (©USDA Forest Service/Region 2- Rocky Mountain Region/USDA Forest Service/Bugwood.org - CC BY 3.0 US); N) Symptoms of Xyella fastidiosa on olive in Apulia, Italy, 2015 (©Dr Stephen Parnell).
Figure 5.
Symptoms caused by the diseases identified in the review of published articles on disease outbreaks and epidemics. A) Fusarium oxysporum f. sp. cubense (Panama disease of banana); symptoms on a banana plant (Public Domain - Released by Scot Nelson/via flickr - CC0); B) Meloidogyne enterolobii (Pacara earpod tree root-knot nematode); rootknot symptoms on an ornamental Petunia species (©Jeffrey W. Lotz/Florida Department of Agriculture & Consumer Services/Bugwood.org - CC BY 3.0 US); C) Cassava mosaic disease (African cassava mosaic); symptoms on cassava leaf. (©Phil Taylor, CABI); D) Citrus tristeza virus (©Tim Gottwald); E) Erwinia amylovora (fireblight) on pear (©Phil Taylor, CABI); F) Candidatus Liberibacter asiaticus (Asian greening); HLB symptoms. April 2016 (©Nian Wang (Citrus Research and Education Center)); G) Pseudomonas syringae pv. syringae (bacterial canker) symptoms on peach (Prunus persica), bark peeled to show discoloration beneath (©University of Georgia Plant Pathology/University of Georgia/Bugwood.org - CC BY 3.0 US); H) Fusarium on wheat (©Phil Taylor, CABI); I) Magnaporthe oryzae (rice blast) in Myanmar (©Phil Taylor, CABI); J) Puccinia striiformis f. sp. tritici; symptoms of yellow rust on triticale leaf (©Rasbak/via Wikimedia Commons - CC BY-SA 4.0); K) Zymoseptoria tritici, septoria on wheat (©Phil Taylor, CABI); L) Symptoms of maize lethal necrosis disease (©Joseph Mulema, CABI); M) Bursaphelenchus xylophilus (pine wilt nematode); pine wilt symptoms, one year after death of tree (©USDA Forest Service/Region 2- Rocky Mountain Region/USDA Forest Service/Bugwood.org - CC BY 3.0 US); N) Symptoms of Xyella fastidiosa on olive in Apulia, Italy, 2015 (©Dr Stephen Parnell).

Figure 6.
Unique pathogen-host pairs in new pathogen records reported in 2021 in the CABI Distribution Database. Host species were identified in each location where new pathogen records were added to DDB in 2021 and were reported in four or more geolocations defined in DDB. One or more host species could be identified in each geolocation. Host species were classified in groups, and locations by continent.
Figure 6.
Unique pathogen-host pairs in new pathogen records reported in 2021 in the CABI Distribution Database. Host species were identified in each location where new pathogen records were added to DDB in 2021 and were reported in four or more geolocations defined in DDB. One or more host species could be identified in each geolocation. Host species were classified in groups, and locations by continent.
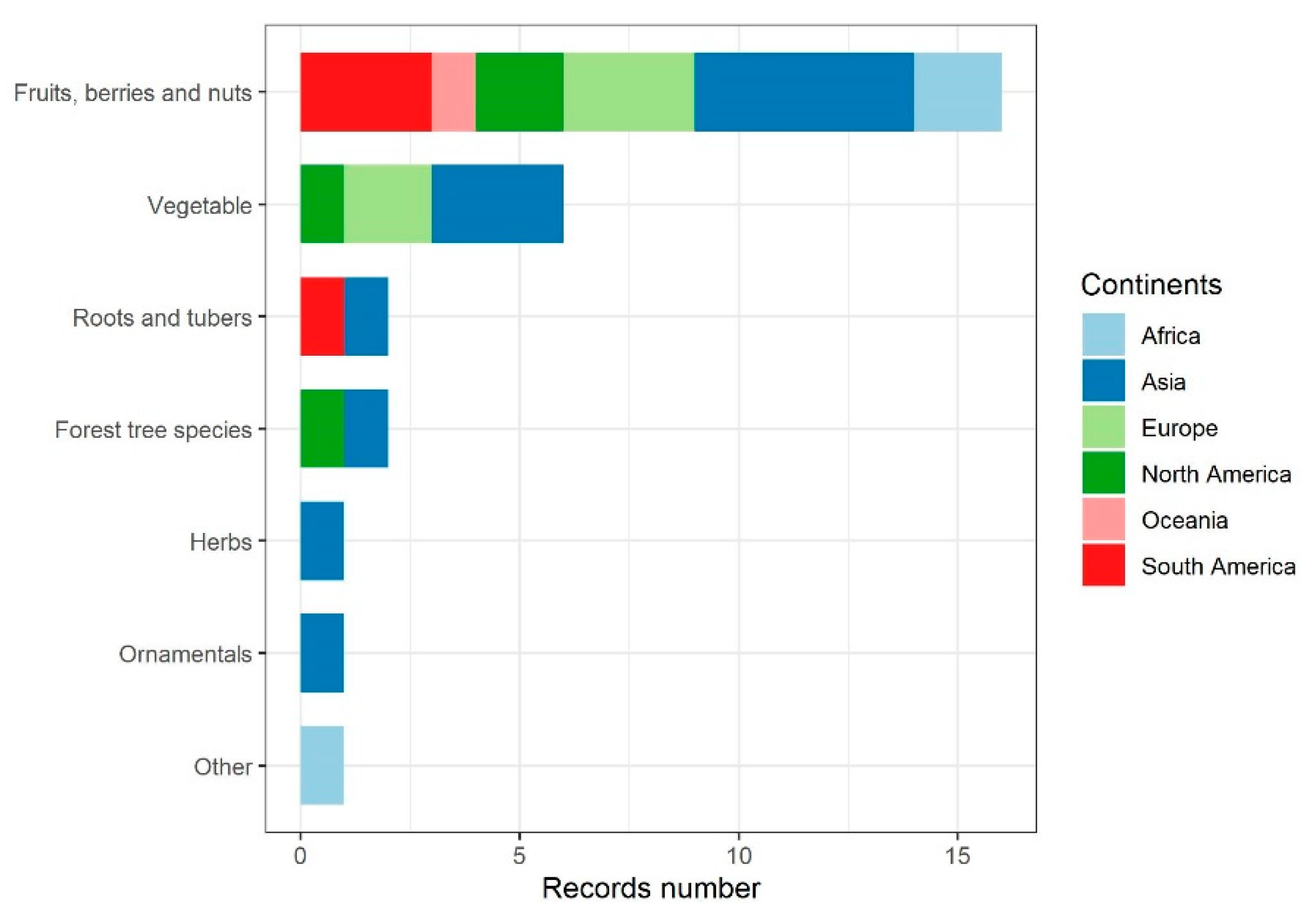
Figure 7.
Map showing locations of new pathogen records reported in 2021 in the CABI Distribution Database following manual review. Counts are summarised by continent, with a breakdown of pathogen type by kingdom. A map showing country by country data is available in Electronic Supporting Information, S3 (Figure S3-2).
Figure 7.
Map showing locations of new pathogen records reported in 2021 in the CABI Distribution Database following manual review. Counts are summarised by continent, with a breakdown of pathogen type by kingdom. A map showing country by country data is available in Electronic Supporting Information, S3 (Figure S3-2).
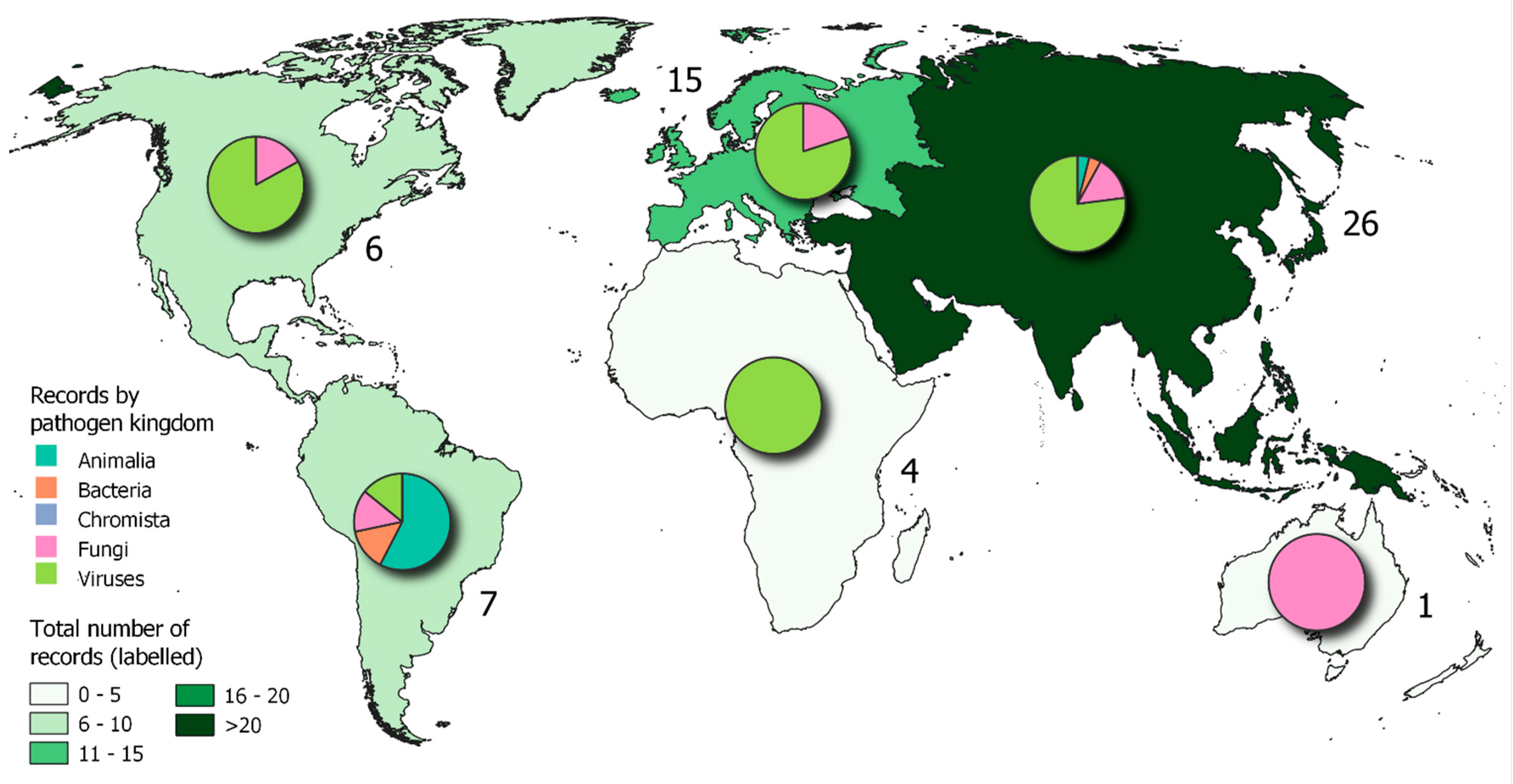
Figure 8.
Map showing combined locations of new pathogen records reported in 2021 from both the literature review and from the CABI Distribution Database (CDD) (following manual review). Counts at CDD geolocation level are summarised by continent, with a breakdown of pathogen type by kingdom. A map showing country by country data is available in Electronic Supporting Information, S3 (Figure S3-3).
Figure 8.
Map showing combined locations of new pathogen records reported in 2021 from both the literature review and from the CABI Distribution Database (CDD) (following manual review). Counts at CDD geolocation level are summarised by continent, with a breakdown of pathogen type by kingdom. A map showing country by country data is available in Electronic Supporting Information, S3 (Figure S3-3).
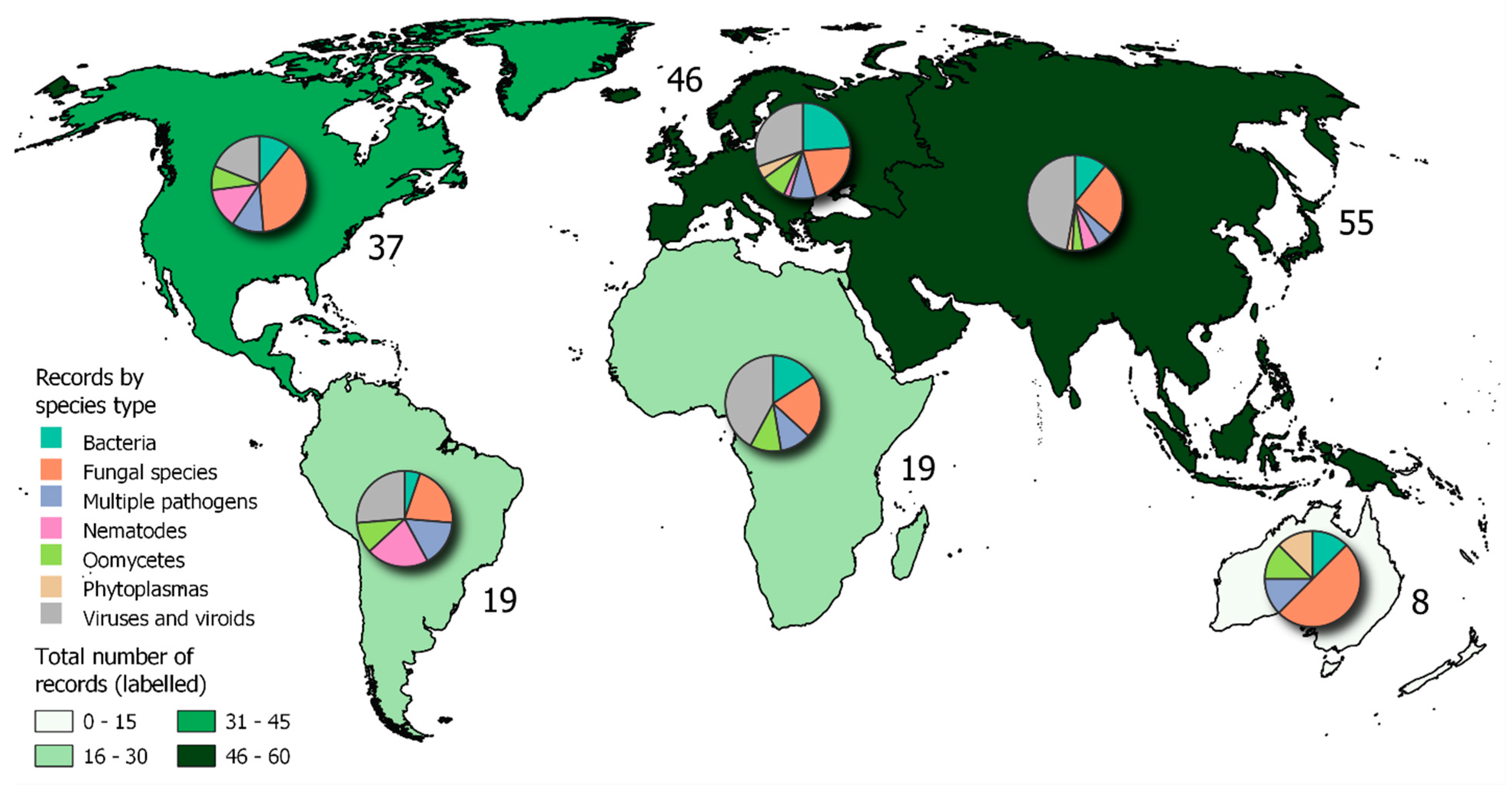

Table 1.
Combined results following Stage 1 and 2 from the review of published articles on disease outbreaks and epidemics. Summaries of the results for each of these categories are given in the main text; the range of survey dates for the named species/complexes is also indicated.
Table 1.
Combined results following Stage 1 and 2 from the review of published articles on disease outbreaks and epidemics. Summaries of the results for each of these categories are given in the main text; the range of survey dates for the named species/complexes is also indicated.
| NAMED SPECIES OR SPECIES COMPLEX | |||
| Number of articles | Year/s of survey | ||
| Candidatus Liberibacter asiaticus and vectors | 10 | 2009-2019 | |
| Cassava mosaic viruses | 10 | 2013-2020 | |
| Xylella fastidiosa and vectors | 10 | 2011-2020 | |
| Erwinia amylovora | 7 | 2015-2020 | |
| Puccinia striiformis f. sp. tritici | 7 | 1985-2020 | |
| Zymoseptoria tritici | 7 | 1999-2020 | |
| Bursaphelenchus xylophilus | 7 | 2000-2020 | |
| Meloidogyne spp. complexes | 6 | 2013-2019 | |
| Magnaporthe oryzae | 6 | 2003-2019 | |
| Citrus tristeza virus | 6 | 2012-2019 | |
| Fusarium spp. complexes | 6 | 2015-2019 | |
| Fusarium oxysporum f. sp. cubense | 5 | 2016-2017 | |
| Pseudomonas syringae pvs | 5 | 1982-2019 | |
| Maize lethal necrosis co-infecting viruses | 5 | 2013-2019 | |
| MULTIPLE PATHOGENS/CROPPING PRODUCTION PRACTICE | |||
| Number of articles | |||
| Aquaculture | 5 | ||
| Crop-associated weeds | 5 | ||
| Cropping systems | 5 | ||
| Forest tree pathogens | 5 | ||
| Wild plants | 5 | ||
Table 2.
New pathogen records reported in 2021 in the CABI Distribution Database. Of the 28 pathogens which met the threshold for manual review – i.e., had at least four new records – the 15 shown above were confirmed as constituting new reports of a named pathogen in a new location. The "Percentage of records from 2021” column was calculated by taking “New records in 2021 after manual review” as a percentage of “Pre 2021 records” + “New records in 2021 after manual review”. This allowed us to designate four pathogens as exhibiting a signature of actively emerging and/or spreading: pathogens for which the percentage of records from 2021 was >25% of the total and for which there were at least 2 records in 2021 after manual review. These pathogens are discussed in the main text.
Table 2.
New pathogen records reported in 2021 in the CABI Distribution Database. Of the 28 pathogens which met the threshold for manual review – i.e., had at least four new records – the 15 shown above were confirmed as constituting new reports of a named pathogen in a new location. The "Percentage of records from 2021” column was calculated by taking “New records in 2021 after manual review” as a percentage of “Pre 2021 records” + “New records in 2021 after manual review”. This allowed us to designate four pathogens as exhibiting a signature of actively emerging and/or spreading: pathogens for which the percentage of records from 2021 was >25% of the total and for which there were at least 2 records in 2021 after manual review. These pathogens are discussed in the main text.
| PATHOGEN | NUMBER OF DDB RECORDS | ||||
|---|---|---|---|---|---|
| Pre 2021 records | New 2021 records |
2021 records after manual review | Percentage of new records from 2021 | Emerging and/or spreading? | |
| Tomato brown rugose fruit virus | 18 | 16 | 15 | 45.45% | ✓ |
| Cucurbit chlorotic yellows virus | 24 | 12 | 11 | 31.43% | ✓ |
| apple rubbery wood viruses | 27 | 8 | 8 | 22.86% | |
| Aphelenchoides besseyi | 130 | 8 | 4 | 2.99% | |
| Apple hammerhead viroid | 1 | 5 | 4 | 80.00% | ✓ |
| Colletotrichum siamense | 0 | 4 | 4 | 100.00% | ✓ |
| Potato spindle tuber viroid | 56 | 5 | 3 | 5.08% | |
| Erysiphe corylacearum | 16 | 4 | 3 | 15.79% | |
| Erwinia rhapontici | 10 | 8 | 1 | 9.09% | |
| Citrus tristeza virus | 160 | 7 | 1 | 0.62% | |
| Globodera rostochiensis | 99 | 7 | 1 | 1.00% | |
| Biscogniauxia mediterranea | 3 | 5 | 1 | 25.00% | |
| Fusarium oxysporum f. sp. cubense Tropical race 4 | 31 | 5 | 1 | 3.13% | |
| Candidatus Liberibacter asiaticus | 105 | 4 | 1 | 0.94% | |
| Nothophoma quercina | 2 | 4 | 1 | 33.33% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



