Submitted:
02 May 2023
Posted:
04 May 2023
You are already at the latest version
Abstract
Fire regimes in mountain landscapes of southern Europe have been shifting from their baselines due to rural abandonment and fire exclusion policies. Understanding the effects of fire on biodiversity is paramount to implement adequate management. Herein, we evaluated the relative role of burn severity and heterogeneity on bird abundance in an abandoned mountain range located in the biogeographic transition between the Eurosiberian and Mediterranean region (the Natural Park ‘Baixa Limia–Serra do Xurés’). We surveyed the bird community in 206 census plots distributed across the Natural Park, both inside and outside areas affected by wildfires over the last 11 years (from 2010 to 2020). We used satellite images of Sentinel 2 and Landsat missions to quantify the burn severity and heterogeneity of each fire within each surveyed plot. We also accounted for the past land use (forestry or agropastoral use) by using a land cover information for year 2010 derived from satellite image classification. We recorded 1,735 contacts from 28 bird species. Our models, fitted by using GLMs with Poisson error distribution (pseudo-R2-average of 0.22 ± 0.13), showed that up to 71% of the modelled species were linearly correlated with at least one attribute of the fire regime. The spatiotemporal variation in burnt area and severity were relevant factors for explaining the local abundance of our target species (39% of the species; Akaike weights > 0.75). We also found a quadratic effect of at least one fire regime attribute on bird abundance for 60% of the modeled species. The past land use, and its legacy after 10 years, was critical to understand the role of fire (Akaike weights > 0.75). Our findings confirm the importance of incorporating remotely sensed indicators of burn severity into the toolkit of decision makers to accurately anticipate the response of birds to fire management.
Keywords:
Bird response
; burn severity indices
; land-use legacy
; Poisson GLMs
; time since fire
; time series of satellite images
1. Introduction
Fire is a major ecological and evolutionary driver of biodiversity in Mediterranean-type ecosystems (He et al., 2019). The effects of fire on biodiversity have been studied across different taxa (plants, vertebrates and invertebrates; see e.g., Clavero et al., 2011; Mateos et al., 2011; Pausas and Ribeiro, 2017; Santos et al., 2022), considering different species traits (McLauchlan et al., 2020; Pausas, 2015), pre-fire conditions (Jones et al., 2016; Taillie et al., 2018), and biogeographic contexts (Farnsworth et al., 2014; Martínez et al., 2022; Parr and Andersen, 2006; Tingley et al., 2016). However, land managers are facing new challenges due to novel fire regimes, land-use and climate change (Kelly et al., 2020; Regos et al., 2016a). In southern Europe, rural landscapes are complex socioecological systems, shaped by traditional agricultural and pastoral activities since millennia. Fire regimes are shifting from their baselines due to cross-scale interactions between global climate change, regional land-use and fire suppression policies (Batllori et al., 2013; Moreira et al., 2020). Over the last decades, these extensive agroforestry activities have been gradually lost, affecting biodiversity patterns and fire (Lehikoinen et al., 2019; Wretenberg et al., 2007; Zakkak et al., 2014). In addition, the traditional use of fire by rural communities – commonly used to open spaces for pastures – has being eradicated as tool to manage landscapes, which has led to profound changes in fire ignition and fuel load patterns (Loepfe et al., 2010; Uyttewaal et al., 2023). The lack of a deep knowledge of the possible and complex responses of biodiversity to altered fire regimes undermines the capacity of managers to decide when, where and how to implement fire management. Besides, the impacts of fire on biodiversity is often evaluated through proxies for patch-mosaic configurations and post-fire vegetation regeneration (e.g., through ‘burnt area’ or ‘time since fire’) (Chalmandrier et al., 2013; Taillie et al., 2018; Watson et al., 2012). Despite being a critical descriptor of fire regime, burn severity is an aspect not easily accessible for ecologists or land managers, being an under-studied issue of the fire–biodiversity relationships (see meta-analysis in Fontaine and Kennedy, 2012).
Biodiversity patterns are largely influenced by past land uses (Foley et al., 2005; Herrando et al., 2016). For instance, forest birds are benefited from vegetation encroachment and forest spread (Gil-Tena et al., 2009; Regos et al., 2016a, 2016b), being the species more dependent on open habitats (such as grasslands and farmlands) the most negatively affected by agricultural abandonment (Scozzafava and De Sanctis, 2006; Wretenberg et al., 2007; Zakkak et al., 2014). In these abandoned landscapes, wildfires offer a window opportunity for early successional habitat species (Brotons et al., 2005; Campos et al., 2021; Regos et al., 2015a) —an adaptative response that strongly depends on burn severity and post-fire vegetation recovery (Lindenmayer et al., 2014; Puig-Gironès et al., 2022; Taillie et al., 2018). In areas with large-scale land-use changes, legacies from previous land-use can persist and affect post-fire vegetation dynamics (Puerta-Piñero et al., 2012). In fact, past land use was found to be even more important than propagule source distance or pine tree density in explaining levels of native forest regeneration in Mediterranean forests (Navarro-González et al., 2013). Historical land-use change and fire suppression were predicted to drive regional bird distributions in directions opposite to those expected from climatic trends (De Cáceres et al., 2013). Thereby, understanding the role of fire on biodiversity in these complex socioecological systems (driven by altered fire regimes and land abandonment processes) is paramount for implementing adequate management —especially for endangered species in protected areas (see e.g., Maillard et al., 2022; Pérez-Granados et al., 2013; Regos et al., 2016).
This study assesses the effect of wildfires on bird abundance in the Natural Park ‘Baixa Limia–Serra do Xurés’ (NW Iberia), a mountain range located in the biogeographic transition between the Eurosiberian and Mediterranean region, strongly affected by rural abandonment and human-caused fires (Regos et al., 2016b). Considering the high fire frequency and rapid post-fire vegetation recoveries in the region, we expect that wildfires had largely influenced local bird abundance. In particular, we evaluated the relative role of burn severity and heterogeneity on bird abundance while accounting for legacy factors such as previous land uses and past fires [see e.g., (Puig-Gironès et al., 2022; Watson et al., 2012; Zozaya et al., 2010)]. To assess the role of these factors on bird abundance, we surveyed local bird community in 206 census plots distributed across the Natural Park, both inside and outside areas affected by wildfires over the last 11 years (from 2010 to 2020). We used satellite images of Sentinel 2 and Landsat missions (freely available from European Space Agency and NASA, respectively) to measure the burn severity and heterogeneity of each fire that took place within each surveyed plot. We also accounted for the past land use (forestry or agropastoral use) by using land cover information for year 2010 derived from image classification of Landsat satellite.
2. Materials and Methods
2.1. Study Area
This study was carried out in the Natural Park ‘Baixa Limia-Serra do Xurés’ (29,345 ha), a mountain rural area located in the south-west of Galicia (NW Spain, 42°–8°; Figure 1), included in the Transboundary Biosphere Reserve ‘Gerês-Xurés’ (Macedo et al., 2009). The elevation ranges from 323 to 1,529 m a.s.l. with an average slope of 13°. The region is located in the transition between the Mediterranean and Eurosiberian biogeographic zones, in the proximity of the Atlantic coast. The climate regime is temperate oceanic sub-Mediterranean, with an average maximum temperature during the hottest month (June) of 22.8 °C and an average minimum temperature during the coldest month (January) of 0.29 °C. Average annual rainfall is 1,223 mm (Martínez-Cortizas and Pérez-Alberti, 1999). The landscape in the study area is dominated by shrubs (broom, gorse and heath, c.a. 32% of the study area) and sparsely vegetated areas (rocky areas with poor soils and little vegetation, 25%), maintained by fire and extensive agropastoral activities; followed by a variety of fragmented forests, such as deciduous woodlands (mostly represented by Quercus robur and Q. pyrenaica; 18%) and coniferous plantations (dominated by Pinus sylvestris and P. pinaster; 11%) (Regos et al., 2015b). The study area has been subjected to land abandonment processes (i.e., vegetation encroachment and forest spread) since the second part of the last century due to rural exodus (Regos et al., 2015b).
The Natural park ‘Baixa Limia-Serra do Xurés’ is included in the Natura 2000 network, since it holds high levels of biodiversity and serves as refuge for several emblematic species of conservation concern such as Golden eagle (Aquila chrysaetos), Eagle owl (Bubo bubo), Red-backed Shrike (Lanius collurio), Skylark (Alauda arvensis) or Dartford warbler (Curruca undata), among others; being designated as Spatial Protection Area (Directive 2009/147/CE) (Domínguez et al., 2012, 2005).
The study area is classified within the intermediate-cool-small pyrome (characterized by intermediate fire return but fairly small fires, see Archibald et al., 2013). The fire regime in the study area is therefore characterized by a large number of small- and medium-sized fires (Pais et al., 2020). Most of these fires are caused by human activity and classified as arson (87%). The land covers most affected by fire in the Natural park between 2000 and 2010 were sparsely vegetated areas and closed shrubland (49 and 15 % respectively), followed by pine plantations and oak woodlands (8.8 and 8.7 %) (Regos et al., 2015b). The speed of initial post-fire recovery is related to differences in fire–response traits of vegetation and to climatic conditions immediately following fire. The mid-term recovery is mainly influenced by fire traits and post-fire climatic conditions. However, the long-term recovery is more influenced by burn severity than by vegetation type and structure or by post-fire climatic conditions (Torres et al., 2018).
2.2. Bird sampling Design
The bird community was surveyed by means of 5-min point counts (206 sampling units, Figure 1) with unlimited distance. The censuses were undertaken during the breeding season (from mid-May to mid-July) of 2021. The censuses were carried out during the 4 h after sunrise (peak vocal activity) and under uniform weather conditions (days without marked rainfall or wind) to avoid possible detection biases caused by the time of survey, wind speed or cloud cover (Gregory et al., 2004). Each plot was considered to cover an area of about 3 hectares (ha), as most individuals were recorded within a radius of 100 m from the point count. This census method is designed to provide information on abundance of diurnal songbirds (Gregory et al., 2004). Raptors and crepuscular species were not considered in the analysis since this census method is not suitable for estimating the abundance of those species (Bibby et al., 1992). Migrants and rare species were also excluded.
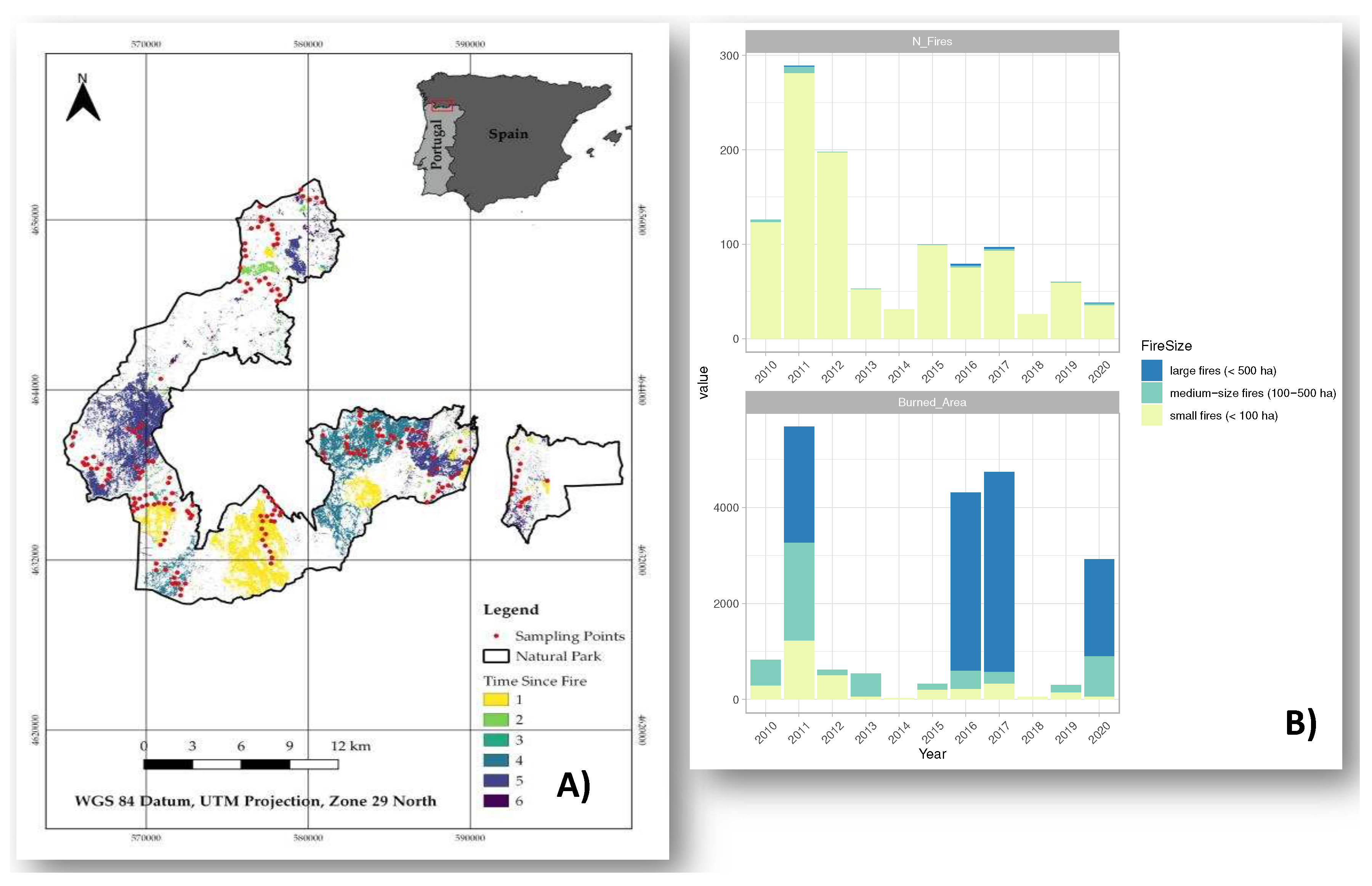
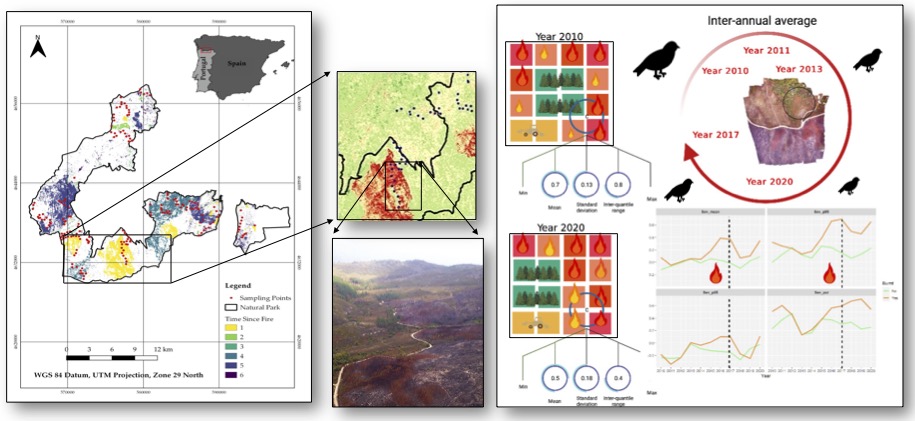
Figure 1.
Location of the Natural Park ‘Baixa Limia-Serra do Xurés’ and ‘Time Since Fire’ for the period 2010-2020. Red dots represent the point counts used to estimate bird density at species level (A). Number of fires and burned area classified by fire size (B).
Figure 1.
Location of the Natural Park ‘Baixa Limia-Serra do Xurés’ and ‘Time Since Fire’ for the period 2010-2020. Red dots represent the point counts used to estimate bird density at species level (A). Number of fires and burned area classified by fire size (B).
The census plots were spatially selected to conduct standardized intensive surveys in a stratified fashion to cover all land-cover types present within each of 10-km squares in which the study area was divided (Figure 1). This survey design allows to capture a large range of environmental conditions, in term of burn severity and time after fire (Figure 1). The census plots were separated by a minimum distance of 250 m to avoid overlapping census plots and possible cases of pseudo-replication. In this respect, the European Green Woodpecker (Picus viridis) was also excluded from the analysis because its conspicuous song is easily detected over long distances (more than 250 m).
2.3. Fire Regime
We created a set of fire-related variables to characterize the fire regime. These fire-related variables were based on burn severity indices derived from satellite imagery and the official database of fire perimeters of the Galician Government (Xunta de Galicia). For each year between 2010 and 2020, we used a pair of satellite images, one before (April - July) and one after (September - November) the fire season. To do so, we used the best available information for each year from the USGS (United States Geological Survey, available at https://earth.explorer.usgs.gov/): Landsat 5 imagery for years 2010 and 2011, Landsat 7 for year 2012, since Landsat 5 imagery was not useful due to high cloudiness. Since its launch in 2013, we shifted to Landsat 8 data, and finally to Sentinel 2 data from 2015 onwards (Table 1). All downloaded images were L1T (a processing level that includes a geometric correction performed with ground control points (GCP) and the use of a digital elevation model) and were re-projected to the UTM coordinate system (WGS 84 datum, UTM projection, Zone 29 North). We applied a radiometric correction method to all images, following the ‘Dark Object Subtraction’ methodology (available in the R-package ‘RStoolbox’), which assumes that the radiometric minimum for the image is located in pixels completely covered by shadow or underwater (Chavez, 1996). The method assumes that the reflectance in those pixels is exclusively due to the atmospheric effects and uses these values to correct the whole image, converting the values from TOA reflectance to surface reflectance.
For each of these years, we calculated the Normalized Burn Ratio (NBR), which is the normalized ratio between near-infrared (NIR) and short-wave infrared (SWIR) radiation (Equation (1)). NIR and SWIR bands of satellite sensors respond in opposite ways to burned vegetation, allowing to identify burned areas (Key and Benson, 2006).
To obtain a quantitative measure of change, we calculated the dNBR for each year by subtracting the NBR of the post-fire season image from the NBR of the pre-fire season image (Equation (2). Finally, dNBR values were used as an estimate for fire severity of each fire season (see Figure 2).
We computed several metrics within the buffer of 100 meter around each point count to characterize the burn severity at census-plot level for each year (i.e., spatial component): the average, minimum, and maximum values within the buffer. The inter-quartile range (i.e., the difference between the 5% and 95% quartile values) and the standard deviation were calculated to measure the heterogeneity in burn severity within the census plot (i.e., spatial variation; sensu Kelly et al., 2016) (Table S1). In addition, we estimated the percentage of burnt area in each census plot from the fire scars, available in vectorial format from official data of the Galicia Government (Xunta de Galicia) for the 2010-2020 period. Vectorial data was rasterized at the same spatial resolution as satellite images (30 meters). From the raster maps of annual burnt area and burn severity (Figure S1), we estimated the average value along the 11-year period in order to account for post-fire vegetation recovery and subsequent fire events (i.e., temporal component) (Table S1). To characterize other important attributes of fire regime over that period, we calculated fire recurrence (i.e., times each plot was affected by a fire event) and time since a fire event took place at each plot (hereafter, TSF) (Table S2).
2.4. Land-Use legacy
The inclusion of LULC covariates into the models allows assessing bird abundance variation across sites in relation to fire metrics, thus avoiding possible spurious correlations with other habitat characteristics. Satellite images were captured by Landsat 5 TM (Thematic Mapper) for year 2010 (July 30 and October 18). To generate the LULC map from each image, we applied different supervised classification methods. Six LULC classes were identified in our study site: (i) water bodies, including rivers, lakes and dams; (ii) deciduous forests, mainly oak woodlands; (iii) evergreen forests, mostly pine plantations; (iv) shrublands, mostly covered by health and gorse species; (v) croplands and grasslands; and (vi) sparsely vegetated areas, mostly shrub and rocky areas located at high elevation. We used two images for each year to enhance the spectral separability among LULC classes due to differences in vegetation phenology, thus increasing our ability to classify them (see details in Cánibe et al., 2022).
We performed image supervised classification using 4 different algorithms (‘Random Forest’, ‘Least Squared Support Vector Machines with Radial Basis Function Kernel’ from the ‘Support Vector Machines’ family of classifiers, ‘Monotone Multi-Layer Perceptron Neural Network’ from the artificial neural network family, and ‘adaptive boosting’ from the gradient boosting models family) accessible from the ‘superClass’ function of the ‘RStoolbox’ R package (Leutner and Horning, 2017). Map accuracy for each algorithm was measured using confusion matrices from which sensitivity and positive predictive power were calculated. We finally used the LULC map derived from ‘Random forest’ classifier since it was the most accurate (overall accuracy of 0.95; Kappa coefficient of 0.94) (see details in Cánibe et al., 2022). Finally, we reclassified the six LULC classes into three main land cover/use types of the study area (forest, cropland and grassland, and shrubland), and estimated the percentage of occupancy of each class to account for past land-use effects on bird abundance.
2.5. Effects of Fire on Bird Abundance
To assess the effects of fire regime on the abundance of each target species (N = 28; Table 2), we fitted generalized linear models (hereafter: GLMs; Burnham and Anderson, 2002). Since the response variable (i.e., relative bird abundance index) represents counts of individuals, our models were fitted by using GLMs with Poisson error distribution. The good-of-fitness of each model was measured by the pseudo-R2 (R2 Nagelkerke). We also measured the dispersion parameter (deviance/degrees of freedom), which should approximate to 1.0, to ensure no overdispersion within the models. The analyses were performed with the ‘MASS’ R package. We calculated the variance inflation factor (VIF) with the ‘usdm’ R package (Naimi, 2017) to estimate how much a regression coefficient’s variance is inflated by multicollinearity. The variables with VIF value greater than 3 were excluded from further analyses (Dormann et al., 2013). We applied a multimodel inference approach to run the GLMs for all (valid) combinations of explanatory variables (Burnham and Anderson, 2002); that is, (1) percentage of forest, (2) cropland and grassland, and (3) burned area; (4) TSF, and (5) mean and (6) standard deviation of burn severity (with the dredge function available in the ‘MuMIn’ R package; Barton, 2016). Models were built assuming linear and non-linear (i.e., second-order polynomial) relationships between the response and exploratory variables. For each model, we calculated the Akaike information criterion (AIC) and ∆I, where ∆I = AICi − AICminimum. All the models with ∆I < 4 were considered to have support. The importance of each variable was obtained by adding the Akaike weights (Wi) to the models in which such variable was present (Burnham and Anderson, 2002).
3. Results
The bird dataset included 2,041 records of 61 species, from which only a third had enough records to be modelled (i.e., species with presence in more than 5% of the 206 census plots) (number of species = 28; total contacts = 1,735; see Table 1, and Table S3 in Supp. Mat.). The bird abundance models yielded moderate explanatory capacity (i.e., the proportion of the variation in the dependent variable that is predictable from the independent variables) for most of our target species (pseudo-R2-average of 0.22 ± 0.13, Figure 2). The abundance models fitted assuming linear relationships showed similar explanatory capacity than those built upon non-linear relationships (hereafter: second-order polynomial GLMs; pseudo-R2-average of 0.20 ± 0.12), although depending on the target species (Figure 2; Table S4).
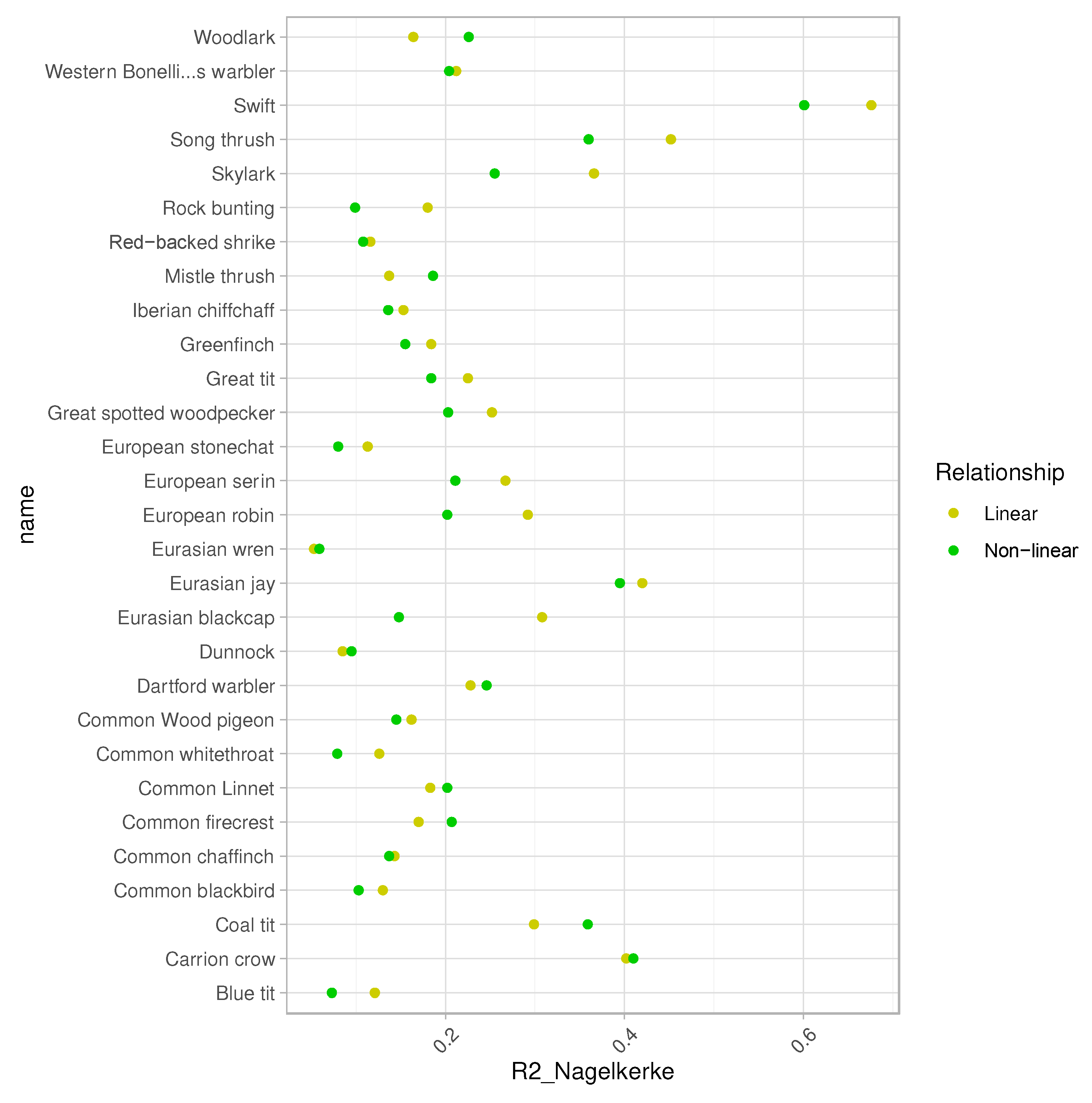

Table 1.
List of modelled species and total number of counts across the census plot carried out in the study area in 2021.
Table 1.
List of modelled species and total number of counts across the census plot carried out in the study area in 2021.
| Common Name | Scientific Name | Acronyms | Counts |
|---|---|---|---|
| Common Wood pigeon | Columba palumbus | C_palum | 20 |
| Common swift | Apus apus | Apus | 61 |
| Great spotted woodpecker | Dendrocopos major | D_major | 12 |
| woodlark | Lullula arborea | L_arbor | 24 |
| Skylark | Alauda arvensis | A_rvnss | 65 |
| Eurasian wren | Troglodytes troglodytes | Trogl | 153 |
| European robin | Erithacus rubecula | E_rubec | 72 |
| European stonechat | Saxicola rubicola | S_torq | 132 |
| Common blackbird | Turdus merula | T_merul | 130 |
| Song thrush | Turdus philomelos | T_philm | 27 |
| Mistle thrush | Turdus viscivorus | T_viscv | 49 |
| Dartford warbler | Curruca undata | S_undat | 186 |
| Common whitethroat | Sylvia communis | S_comm | 26 |
| Eurasian blackcap | Sylvia atricapilla | S_atrcp | 62 |
| Western Bonelli’s warbler | Phylloscopus bonelli | P_bonll | 36 |
| Iberian chiffchaff | Phylloscopus ibericus | P_ibrcs | 28 |
| Common firecrest | Regulus ignicapilla | R_igncp | 56 |
| Coal tit | Periparus ater | P_ater | 104 |
| Blue tit | Cyanistes caeruleus | P_caerl | 17 |
| Great tit | Parus major | P_major | 11 |
| Red-backed shrike | Lanus collurio | L_coll | 18 |
| Eurasian jay | Garrulus glandarius | G_gland | 22 |
| Carrion crow | Corvus corone | C_coron | 34 |
| Common chaffinch | Fringilla coelebs | F_colbs | 157 |
| European serin | Serinus serinus | Serinus | 49 |
| Greenfinch | Carduelis chloris | C_chlrs | 15 |
| Common Linnet | Carduelis cannabina | C_cannb | 121 |
| Rock bunting | Emberiza cia | E_cia | 48 |
Figure 2.
Pseudo-R2 (R2 Nagelkerke) for linear and second-order polynomial (non-linear) GLMs fitted with Poisson error distribution. Scientific species names available in Table 1.
Figure 2.
Pseudo-R2 (R2 Nagelkerke) for linear and second-order polynomial (non-linear) GLMs fitted with Poisson error distribution. Scientific species names available in Table 1.
3.1. Linear Relationships between Bird Abundance and Fire regime
According to the GLMs, the abundance of 71% of the target species (20/28) were significatively explained by at least one attribute of the fire regime (hereafter: ‘fire-sensitive’ species) (Figure 3; Table S5). The abundance of 46% of species (13/28) was found to be affected by at least one of the two burn severity metrics (Figure 3; Table S5). The percentage of forest was the main factor driving bird abundance (19/28) (see WAkaike > 0.5 in Figure 4). Regarding the fire-related variables, the percentage of burnt area was the most important factor affecting the local abundance for 39% of the modeled species (11/28), followed by the spatiotemporal descriptors of burn severity (namely, mean and standard deviation of dNBR) that significatively affected 28% of species (8/28) (see Figure 3; and WAkaike > 0.5 in Figure 4). The models showed positive correlations between the standard deviation of burn severity and bird abundance for half of the species (6/13) (Figure 3), while a third of the species did so with the burned area (4/11) and TSF (4 species; Figure 3).
3.2. Non-Linear Relationships between Bird Abundance and Fire Regime
For several birds, the relationship between their abundance and fire regime attributes was also nonlinear, even though the explanatory power of second-order polynomial GLMs was slightly lower than assuming linear relationships (Figure 2). The abundance of 60% of the modeled species were non-linearly influenced by at least one attribute of the fire regime (17/28) (Figure 3; Table S5). The percentage of forest was also the main factor driving bird abundance (16/28) (see WAkaike > 0.5 in Figure 4). However, in the second-order polynomial GLMs, the standard deviation of burn severity was the most important factor affecting the abundance patterns for 32% of the modeled species (9/28), followed by the burnt area and TSF that significatively affected 7 and 6 species, respectively (Figs. 3 and 4; Table S5). The models showed that the standard deviation of burn severity correlated positively with the abundance of 7 of the 17 species sensitive to fire (Figure 3). The abundance of a fourth of the species increased with the burnt area (4/17) (Figure 3). TSF was found to be positively correlated with the abundance of 3 species, while other 3 species did so negatively with this factor (Table 3; Table S5).
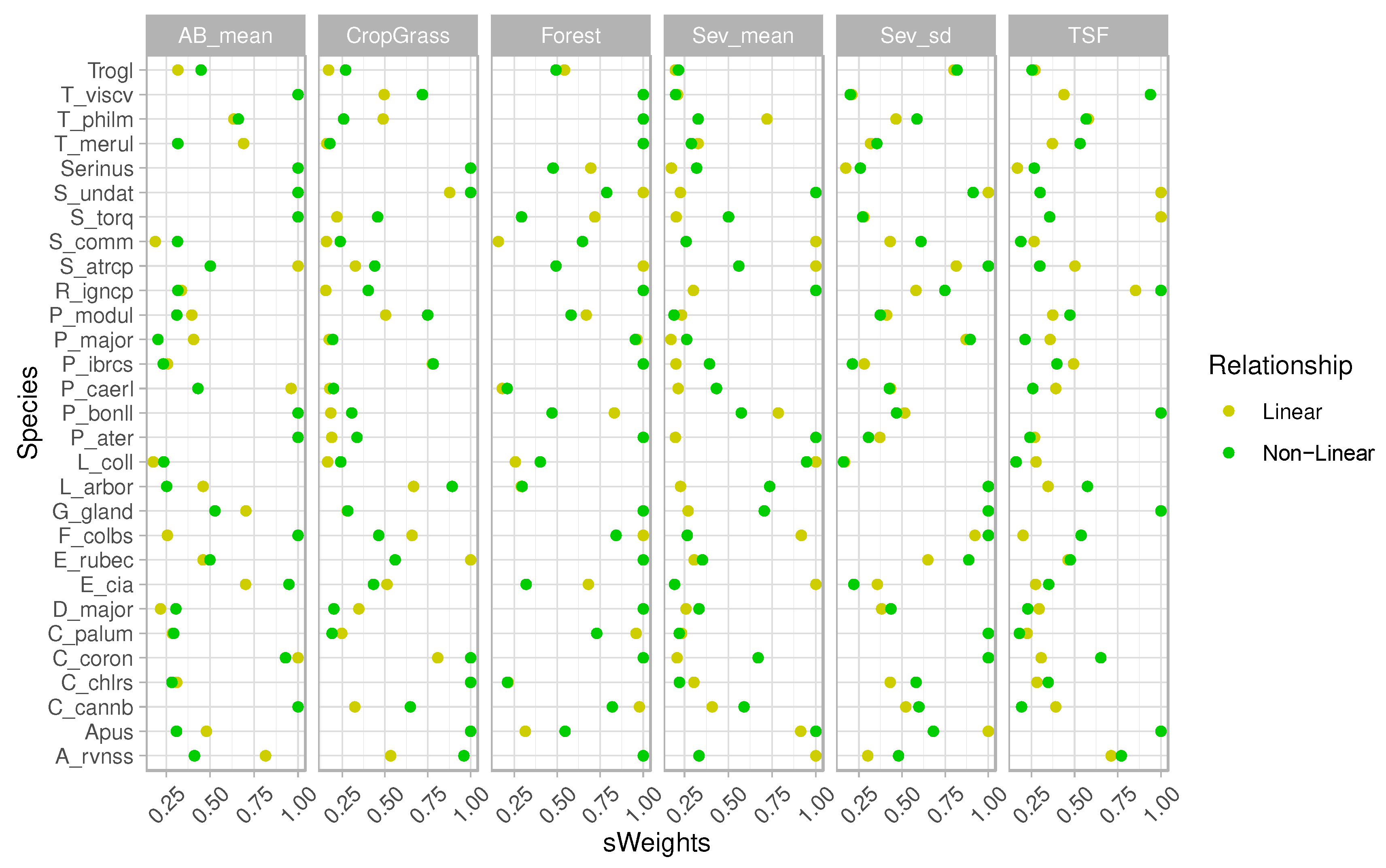
Figure 4.
Relative variable importance obtained by adding the Akaike weights (Wi) of the consequential models (∆I < 4) in which such variable was present, assuming linear or second-order polynomial (non-linear) relationships. Common and scientific names of the modelled species available in Table 1.
Figure 4.
Relative variable importance obtained by adding the Akaike weights (Wi) of the consequential models (∆I < 4) in which such variable was present, assuming linear or second-order polynomial (non-linear) relationships. Common and scientific names of the modelled species available in Table 1.
4. Discussion
Our models showed that the majority of our target species (up to 71%) are correlated with at least one attribute of the fire regime in the ‘Baixa Limia – Serra do Xurés’ Natural Park (see Figs. 3 and 4). The spatiotemporal variation in the burnt area and its severity were relevant factors for explaining the local abundance patterns of our target species (up to 39% of the 28 modeled species, see Figure 3). These results confirm the importance of burn severity and heterogeneity to explain bird abundance (see e.g., Puig-Gironès et al., 2022; Stephens et al., 2015), which has clear implications for fire management (see meta-analysis in Fontaine and Kennedy, 2012). Overall, the spatiotemporal variation of burn severity was positively correlated with the abundance patterns for 6 species (Figs. 3 and 4), which can be explained by an increase in the habitat heterogeneity created by fire (Nimmo et al., 2013; Parr and Andersen, 2006; Zozaya et al., 2010). The amount of burnt area was correlated with the abundance of 39% of species, with both positive and negative effects on bird abundance (see Figure 3). Overall, open habitat and early successional species were found to be positively affected by fire (i.e., the higher the burnt area, the higher the bird abundance), even though some of them were negatively influenced by its severity (see e.g., Dartford warbler and Skylark in Table S5). These results are in line with previous studies that suggest that early successional species, such as Dartford warbler, have peaks of abundance around 4-8 years after fire in Mediterranean ecosystems (Birdlife International, 2014; Pons et al., 2012). Fire creates habitat conditions to allow these species to occupy areas previously lost due to vegetation encroachment or afforestation processes driven by the joint effect of rural abandonment and fire suppression (see e.g., Regos et al., 2015a). In fact, recent studies suggested the use of fire as a tool to enhance bird conservation in this region, since fire could offset the negative effects of land abandonment on ecotone and open-habitat species (Campos et al., 2021; Regos et al., 2016b). On the contrary, forest-dwelling species (see e.g., Eurasian blackcap or Western Bonelli’s warbler in Table S5) were negatively affected by fire, being favored by an increasing heterogeneity in burn severity (i.e., the presence of un-burnt patches) (Berry et al., 2014).
Despite the number of studies supporting ‘time since fire’ as a good predictor of bird abundance (Chalmandrier et al., 2013; Moghli et al., 2021; Pons and Clavero, 2010), this variable only correlated significantly with the abundance of 6 species in our study area (see Figure 3 and Figure 4). The variable ‘time since fire’ is often used to delineate successional vegetation states, being a good proxy for post-fire recovery, fuel age or vegetation structure (Moritz et al., 2004; Parks et al., 2014). However, our study region is characterized by fast recovery rates (Torres et al., 2018) due to its bioclimatic position, which could explain the less important role of ‘time since fire’ on bird abundance (see Figure 4). Nevertheless, future studies in the region should include longer time series for burn severity and fire occurrence in their analyses. Short- and mid-term bird responses to fire can significatively differ from longer-term post-fire conditions and management (Nappi et al., 2010; Rost et al., 2013; Santos et al., 2016; White et al., 2015) —a critical issue that is context-specific.
The relationship between bird abundance and fire regime attributes is not always linear. Some studies have found more complex relationships between fire and bird abundance (Taillie et al., 2018; Watson et al., 2012). It is therefore important to consider non-linear effects of fire severity and ‘time since fire’ to fully understand post-wildfire responses for a majority of bird species (see e.g., Taillie et al., 2018). In our case, we found a quadratic effect of at least one of the three fire regime attributes on bird abundance for 60% of the modeled species (see Figure 3). Yet, the explanatory power of these models was slightly lower than those fitted assuming linear relationships (see Figure 2; Table S4), which suggests that adding non-linear relationships in our model would not provide additional information. On the contrary, the inclusion of variables on the past land use, and its legacy after 10 years (mainly percentage of ‘forest’ and, in less extent, ‘cropland and grassland’ for year 2010), was critical to understand the role of fire (Figs 3 and 4). Previous studies showed a significant increase for 13 shrubland and forest bird species (including species of conservation concern such as Turtle Dove, Dartford Warbler and Western Bonelli’s Warbler), while only 4 ecotone and open-habitat species (e.g., Red-backed Shrike) showed a significant negative trend in the Natural park ‘Baixa Limia – Serra do Xurés’ between 2000 and 2010, due to vegetation encroachment and forest spread in the region (Corbelle-Rico et al., 2022; Regos et al., 2016b; Salaverri et al., 2019). In this sense, this study confirms the added value of accounting for fire and land-use legacy factors when modelling bird abundance to enhance their predictive capacity (De Cáceres et al., 2013; Vallecillo et al., 2009). The complex relationships found between each fire regime descriptor and the relative bird abundance highlights the need of tailor-made management at species level, and the role that fire can play in such management. The creation of small burnt patches through fire (e.g., prescribed burning) could be a cost-effective management option to maintain open habitats in rural abandoned landscape of NW Iberia, while reducing wildfire hazard (Campos et al., 2021; Regos et al., 2016b).
5. Conclusions
Burn severity and heterogeneity – measured through the spatiotemporal variation of dNBR at plot level for the last 11 years – significatively correlated with bird abundance, being relevant factors affecting bird abundance in the Natural park ‘Baixa Limia – Serra do Xurés’. The majority of the 28 modeled species (71%) was found to be sensitive to fire, whose abundance were correlated with at least one fire regime attribute. These results confirm the role that fire can play in the conservation of these species in mountain regions largely affected by rural abandonment. Our burn severity metrics, computed from the inter-annual variation of satellite images, were useful to explain the abundance of several target species, due to the capacity of such indices to inform about habitat conditions after fire across space and time. These results confirm the usefulness of burn severity indicators to explain bird abundance patterns, which clearly complements the information provided by well-known fire regime descriptors such as the amount of burnt area and ‘time since fire’. However, the relative importance of land-use legacy factors to explain the abundance of our target species highlights the need of longer-term analysis to better inform decision making. The particular biogeographic position of our study area – characterized by fast post-fire vegetation recovery rates – calls for caution when extrapolating our results to other socioecological contexts. Despite the context-specific response of birds, our findings confirm the importance of incorporating remotely sensed indicators of burn severity into the toolkit of decision makers to accurately anticipate the response of birds to fire management.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Video S1: Areas affected by those wildfires at census-plot scale at the Natural Park taken by J Gonçalves and A.R. from an Unmanaged Aerial Vehicle; https://youtu.be/NnR_5l6pxbM.
Author Contributions
Conceptualization, A.R..; methodology, A.R..; formal analysis, C.G-R and A.R.; bird surveys, L.T and A.G-C.; remote sensing data, P.F-M and M.C.; writing—original draft preparation, A.R and C.G-R.; writing—review and editing, all authors.; visualization, P.F-M. and M.G-R.; supervision, A.R. and M.D-R.; project administration, A.R.; funding acquisition, A.R. All authors have read and agreed to the published version of the manuscript.
Funding
Fieldwork for collecting bird data was funded by the Xunta de Galicia (contract-number 2021-CP083, co-funded by FEADER program), and the ‘FirESmart’ project (Grant number - PCIF/MOG/0083/2017), funded by the Portuguese Foundation for Science and Technology (FCT). Burn severity products were computed in the frame of the SeverusPT, funded by the Portuguese Foundation for Science and Technology (FCT), grant number PCIF/RPG/0170/2019. AR is supported by the ‘Juan de la Cierva’ fellowship program funded by the Spanish Ministry of Science and Innovation (IJC2019-041033-I). M.C. was financially supported by the ‘FirESmart’ project.
Institutional Review Board Statement
“Not applicable”.
Informed Consent Statement
“Not applicable”.
Data Availability Statement
Bird and fire perimeter datasets are available from the Xunta de Galicia upon request at https://sede.xunta.gal/. Burn severity products in raster format are freely available at https://doi.org/10.5281/zenodo.7262062.
Acknowledgements
We would like to thank Antonio Rodríguez Fernández (Consellería de Medio Ambiente - Xunta de Galicia) for supporting our bird monitoring protocol in the Natural Park ‘Baixa Limia - Serra do Xurés’. We also thank Xunta de Galicia for giving access to the perimeters of the areas burnt in the municipalities of the Natural Park.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Archibald, S., Lehmann, C.E.R., Gomez-Dans, J.L., Bradstock, R.A., 2013. Defining pyromes and global syndromes of fire regimes. Proc. Natl. Acad. Sci. 110, 6442–6447. [CrossRef]
- Barton, K., 2016. MuMIn: multi-model inference. R package version 1.15.6. https:// CRAN.R-project.org/package=MuMIn.
- Batllori, E., Parisien, M.-A., Krawchuk, M. a., Moritz, M. a., 2013. Climate change-induced shifts in fire for Mediterranean ecosystems. Glob. Ecol. Biogeogr. 22, 1118–1129. [CrossRef]
- Berry, L.E., Lindenmayer, D.B., Driscoll, D. a., 2014. Large unburnt areas, not small unburnt patches are needed to conserve avian diversity in fire-prone landscapes. J. Appl. Ecol. n/a-n/a. [CrossRef]
- Bibby, C.J., Burgess, N.D., Hill, D.A., 1992. Bird census techniques. Cambridge University Press, Cambridge. Cambridge University Press, Cambridge, UK.
- Birdlife International, 2014. Species factsheet: Sylvia undata. [WWW Document].
- Brotons, L., Pons, P., Herrando, S., 2005. Colonization of dynamic Mediterranean landscapes: where do birds come from after fire? J. Biogeogr. 32, 789–798. [CrossRef]
- Burnham, K.P., Anderson, D.R., 2002. Model Selection and Multimodel Inference: a Practical Information-theoretic Approach. Springer, New York.
- Campos, J.C., Bernhardt, J., Aquilué, N., Brotons, L., Domínguez, J., Lomba, Â., Marcos, B., Martínez-Freiría, F., Moreira, F., Pais, S., Honrado, J.P., Regos, A., 2021. Using fire to enhance rewilding when agricultural policies fail. Sci. Total Environ. 755, 142897. [CrossRef]
- Cánibe, M., Titeux, N., Domínguez, J., Regos, A., 2022. Assessing the uncertainty arising from standard land-cover mapping procedures when modelling species distributions. Divers. Distrib. 28, 636–648. [CrossRef]
- Chalmandrier, L., Midgley, G.F., Barnard, P., Sirami, C., 2013. Effects of time since fire on birds in a plant diversity hotspot. Acta Oecologica 49, 99–106. [CrossRef]
- Chavez, J., 1996. Image-Based Atmospheric Corrections - Revisited and Improved. Photogramm. Eng. Remote Sens. 62, 1025–1036. [CrossRef]
- Clavero, M., Brotons, L., Herrando, S., 2011. Bird community specialization, bird conservation and disturbance: the role of wildfires. J. Anim. Ecol. 80, 128–36. [CrossRef]
- Corbelle-Rico, E., Sánchez-Fernández, P., López-Iglesias, E., Lago-Peñas, S., Da-Rocha, J.M., 2022. Putting land to work: An evaluation of the economic effects of recultivating abandoned farmland. Land use policy 112. [CrossRef]
- De Cáceres, M., Brotons, L., Aquilué, N., Fortin, M.-J., 2013. The combined effects of land-use legacies and novel fire regimes on bird distributions in the Mediterranean. J. Biogeogr. 40, 1535–1547. [CrossRef]
- Domínguez, J., Arenas, M., Tapia, L., 2005. Guía de las Aves del Parque Natural Baixa Limia-Serra do Xurés y Lic Baixa Limia. Consellería de Medio Ambiente, Xunta de Galicia. Santiago de Compostela.
- Domínguez, J., Lamosa, A., Pardavila, X., Martínez-Freiría, F., Regos, A., Gil, A., Vidal, M., 2012. Atlas de los vertebrados terrestres reproductores en el Parque Natural Baixa Limia-Serra do Xurés y ZEPVN-LIC Baixa Limia. Xunta de Galicia.
- Dormann, C.F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., Marquéz, J.R.G., Gruber, B., Lafourcade, B., Leitão, P.J., Münkemüller, T., McClean, C., Osborne, P.E., Reineking, B., Schröder, B., Skidmore, A.K., Zurell, D., Lautenbach, S., 2013. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography (Cop.). 36, 027–046. [CrossRef]
- Farnsworth, L.M., Nimmo, D.G., Kelly, L.T., Bennett, A.F., Clarke, M.F., 2014. Does pyrodiversity beget alpha, beta or gamma diversity? A case study using reptiles from semi-arid Australia. Divers. Distrib. 20, 663–673. [CrossRef]
- Foley, J.A., Defries, R., Asner, G.P., Barford, C., Bonan, G., Carpenter, S.R., Chapin, F.S., Coe, M.T., Daily, G.C., Gibbs, H.K., Helkowski, J.H., Holloway, T., Howard, E.A., Kucharik, C.J., Monfreda, C., Patz, J.A., Prentice, I.C., Ramankutty, N., Snyder, P.K., 2005. Global consequences of land use. Science (80-. ). 309, 570–574. [CrossRef]
- Fontaine, J.B., Kennedy, P.L., 2012. Meta-analysis of avian and small-mammal response to fire severity and fire surrogate treatments in U.S. Fire-prone forests. Ecol. Appl. 22, 1547–1561. [CrossRef]
- Gil-Tena, A., Brotons, L., Saura, S., 2009. Mediterranean forest dynamics and forest bird distribution changes in the late 20th century. Glob. Chang. Biol. 15, 474–485. [CrossRef]
- Gregory, R.D., Gibbons, D.W., Donald, P.F., 2004. Bird census and survey techniques. Bird Ecol. Conserv. 17–56. [CrossRef]
- He, T., Lamont, B.B., Pausas, J.G., 2019. Fire as a key driver of Earth’s biodiversity. Biol. Rev. 3, 1983–2010. [CrossRef]
- Herrando, S., Brotons, L., Anton, M., Páramo, F., Villero, D., Titeaux, N., Quesada, J., Stefanescu, C., 2016. Assessing impacts of land abandonment on Mediterranean biodiversity using indicators based on bird and butterfly monitoring data. Environ. Conserv. 43, 69–78. [CrossRef]
- Jones, G.M., Gutiérrez, R.J., Tempel, D.J., Whitmore, S.A., Berigan, W.J., Peery, M.Z., 2016. Megafires: an emerging threat to old-forest species. Front. Ecol. Environ. 14, 300–306. [CrossRef]
- Kelly, L.T., Brotons, L., McCarthy, M.A., 2016. Putting pyrodiversity to work for animal conservation. Conserv. Biol. [CrossRef]
- Kelly, L.T., Giljohann, K.M., Duane, A., Aquilué, N., Archibald, S., Batllori, E., Bennett, A.F., Buckland, S.T., Canelles, Q., Clarke, M.F., Fortin, M., Hermoso, V., Herrando, S., Keane, R.E., Lake, F.K., Mccarthy, M.A., Morán-ordóñez, A., Parr, C.L., Pausas, J.G., Penman, T.D., Regos, A., Rumpff, L., Santos, J.L., Smith, A.L., Syphard, A.D., Tingley, M.W., Brotons, L., 2020. Fire and biodiversity in the Anthropocene. Science (80-. ). 370, eabb0355. [CrossRef]
- Key, C.H., Benson, N.C., 2006. Landscape Assessment: Ground Measure of Severity, the Composite Burn Index; and Remote Sensing of Severity, the Normalized Burn Ratio., in: D. C. Lutes, R. E. Keane, J. F. Caratti, C. H. Key, N. C. Benson, S. Sutherland, L.J.G. (Ed.), FIREMON: Fire Effects Monitoring and Inventory System. Ogden, UT: USDA Forest Service, Rocky Mountain Research Station, Gen. Tech. Rep.
- Lehikoinen, A., Brotons, L., Calladine, J., Campedelli, T., Escandell, V., Flousek, J., Grueneberg, C., Haas, F., Harris, S., Herrando, S., Husby, M., Jiguet, F., Kålås, J.A., Lindström, Å., Lorrillière, R., Molina, B., Pladevall, C., Calvi, G., Sattler, T., Schmid, H., Sirkiä, P.M., Teufelbauer, N., Trautmann, S., 2019. Declining population trends of European mountain birds. Glob. Chang. Biol. 25, 577–588. [CrossRef]
- Leutner, B., Horning, N., 2017. RStoolbox: Tools for Remote Sensing Data Analysis. R package version 0.1.8. https://CRAN.R-project.org/package=RStoolbox.
- Lindenmayer, D.B., Blanchard, W., McBurney, L., Blair, D., Banks, S.C., Driscoll, D. a., Smith, A.L., Gill, a. M., 2014. Complex responses of birds to landscape-level fire extent, fire severity and environmental drivers. Divers. Distrib. n/a-n/a. [CrossRef]
- Loepfe, L., Martinez-Vilalta, J., Oliveres, J., Piñol, J., Lloret, F., 2010. Feedbacks between fuel reduction and landscape homogenisation determine fire regimes in three Mediterranean areas. For. Ecol. Manage. 259, 2366–2374. [CrossRef]
- Macedo, A., Tavares, A., Fontes, A., Pinto, C., Rodrigues, C., Machado, C., Figueiredo, D., Silva, D., Carvalho, H., Regalo, H., Osório, M., Santarém, M., Gonzalez, R., Formoso, J.C., Gonzalez, F.J., Fernández, M.A., Gil, A., Veloso, N., 2009. Propuesta para la creación de la Reserva de la Biosfera Transfronteriza Gerês/Xurés. Xunta de Galicia, Comissão de Coordenação e Desenvolvimento Regional do Norte, Instituto de Conservação da Natureza e da Biodiversidade. Xunta de Galicia, Comissão de Coordenação e Desenvolvimento Regional do Norte, Instituto de Conservação da Natureza e da Biodiversidade.
- Maillard, O., Herzog, S.K., Soria-Auza, R.W., Vides-Almonacid, R., 2022. Impact of Fires on Key Biodiversity Areas (KBAs) and Priority Bird Species for Conservation in Bolivia. Fire 5, 1–20. [CrossRef]
- Martínez-Cortizas, A., Pérez-Alberti, A., 1999. Atlas Bioclimático de Galicia. Xunta de Galicia, Santiago de Compostela.
- Martínez, F.J., Cheli, G.H., Grismado, C.J., Bisigato, A.J., 2022. Ground-Dwelling Arachnids and Fire Disturbance: A Case Study in Northeastern Patagonia (Argentina). Fire 5. [CrossRef]
- Mateos, E., Santos, X., Pujade-Villar, J., 2011. Taxonomic and functional responses to fire and post-fire management of a Mediterranean Hymenoptera community. Environ. Manage. 48, 1000–1012. [CrossRef]
- McLauchlan, K.K., Higuera, P.E., Miesel, J., Rogers, B.M., Schweitzer, J., Shuman, J.K., Tepley, A.J., Varner, J.M., Veblen, T.T., Adalsteinsson, S.A., Balch, J.K., Baker, P., Batllori, E., Bigio, E., Brando, P., Cattau, M., Chipman, M.L., Coen, J., Crandall, R., Daniels, L., Enright, N., Gross, W.S., Harvey, B.J., Hatten, J.A., Hermann, S., Hewitt, R.E., Kobziar, L.N., Landesmann, J.B., Loranty, M.M., Maezumi, S.Y., Mearns, L., Moritz, M., Myers, J.A., Pausas, J.G., Pellegrini, A.F.A., Platt, W.J., Roozeboom, J., Safford, H., Santos, F., Scheller, R.M., Sherriff, R.L., Smith, K.G., Smith, M.D., Watts, A.C., 2020. Fire as a fundamental ecological process: Research advances and frontiers. J. Ecol. 108, 2047–2069. [CrossRef]
- Moghli, A., Santana, V.M., Baeza, M.J., Pastor, E., Soliveres, S., 2021. Fire Recurrence and Time Since Last Fire Interact to Determine the Supply of Multiple Ecosystem Services by Mediterranean Forests. Ecosystems 25, 1358–1370. [CrossRef]
- Moreira, F., Ascoli, D., Safford, H., Adams, M.A., Moreno, J.M., Pereira, J.M.C., Catry, F.X., Armesto, J., Bond, W., González, M.E., Curt, T., Koutsias, N., McCaw, L., Price, O., Pausas, J.G., Rigolot, E., Stephens, S., Tavsanoglu, C., Vallejo, V.R., Van Wilgen, B.W., Xanthopoulos, G., Fernandes, P.M., 2020. Wildfire management in Mediterranean-type regions: Paradigm change needed. Environ. Res. Lett. 15. [CrossRef]
- Moritz, M.A., Keeley, J.E., Johnson, E.A., Schaffner, A.A., 2004. Testing a basic assumption of shrubland fire management: how important is fuel age? Front. Ecol. Environ. 2, 67–72. [CrossRef]
- Naimi, B., 2017. usdm: Uncertainty Analysis for Species Distribution Models. R Package, Version 1.1–18. [WWW Document].
- Nappi, A., Drapeau, P., Saint-Germain, M., Angers, V.A., 2010. Effect of fire severity on long-term occupancy of burned boreal conifer forests by saproxylic insects and wood-foraging birds. Int. J. Wildl. Fire 19, 500–511. [CrossRef]
- Navarro-González, I., Pérez-Luque, A.J., Bonet, F.J., Zamora, R., 2013. The weight of the past: land-use legacies and recolonization of pine plantations by oak trees. Ecol. Appl. 23, 1267–76. [CrossRef]
- Nimmo, D.G., Kelly, L.T., Spence-Bailey, L.M., Watson, S.J., Taylor, R.S., Clarke, M.F., Bennett, a F., 2013. Fire mosaics and reptile conservation in a fire-prone region. Conserv. Biol. 27, 345–53. [CrossRef]
- Pais, S., Aquilué, N., Campos, J., Sil, Â., Marcos, B., Martínez-freiría, F., Domínguez, J., Brotons, L., Honrado, J.P., Regos, A., 2020. Mountain farmland protection and fire-smart management jointly reduce fire hazard and enhance biodiversity and carbon sequestration. Ecosyst. Serv. 44, 101143. [CrossRef]
- Parks, S. a., Miller, C., Nelson, C.R., Holden, Z. a., 2014. Previous Fires Moderate Burn Severity of Subsequent Wildland Fires in Two Large Western US Wilderness Areas. Ecosystems 17, 29–42. [CrossRef]
- Parr, C.L., Andersen, A.N., 2006. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv. Biol. 20, 1610–1619. [CrossRef]
- Pausas, J.G., 2015. Evolutionary fire ecology: lessons learned from pines. Trends Ecol. Evol. 20, 318–323. [CrossRef]
- Pausas, J.G., Ribeiro, E., 2017. Fire and plant diversity at the global scale. Glob. Ecol. Biogeogr. 5, 1–17. [CrossRef]
- Pérez-Granados, C., López-Iborra, G.M., Serrano-Davies, E., Noguerales, V., Garza, V., Justribó, J.H., Suárez, F., 2013. Short-Term Effects of a Wildfire on the Endangered Dupont’s Lark Chersophilus duponti in an Arid Shrub-Steppe of Central Spain. Acta Ornithol. 48, 201–210. [CrossRef]
- Pons, P., Clavero, M., 2010. Bird responses to fire severity and time since fire in managed mountain rangelands. Anim. Conserv. 13, 294–305. [CrossRef]
- Pons, P., Clavero, M., Bas, J.M., Prodon, R., 2012. Time-window of occurrence and vegetation cover preferences of Dartford and Sardinian Warblers after fire. J. Ornithol. 153, 921–930. [CrossRef]
- Puerta-Piñero, C., Espelta, J.M., Sánchez-Humanes, B., Rodrigo, A., Coll, L., Brotons, L., 2012. History matters: Previous land use changes determine post-fire vegetation recovery in forested Mediterranean landscapes. For. Ecol. Manage. 279, 121–127. [CrossRef]
- Puig-Gironès, R., Brotons, L., Pons, P., 2022. Aridity, fire severity and proximity of populations affect the temporal responses of open-habitat birds to wildfires. Biol. Conserv. 272. [CrossRef]
- Regos, A., D’Amen, M., Herrando, S., Guisan, A., Brotons, L., 2015a. Fire management, climate change and their interacting effects on birds in complex Mediterranean landscapes: dynamic distribution modelling of an early-successional species—the near-threatened Dartford Warbler (Sylvia undata). J. Ornithol. 156, 275–286. [CrossRef]
- Regos, A., D’Amen, M., Titeux, N., Herrando, S., Guisan, A., Brotons, L., 2016a. Predicting the future effectiveness of protected areas for bird conservation in Mediterranean ecosystems under climate change and novel fire regime scenarios. Divers. Distrib. 22, 83–96. [CrossRef]
- Regos, A., Domínguez, J., Gil-Tena, A., Brotons, L., Ninyerola, M., Pons, X., 2016b. Rural abandoned landscapes and bird assemblages: winners and losers in the rewilding of a marginal mountain area (NW Spain). Reg. Environ. Chang. 16, 199–211. [CrossRef]
- Regos, A., Ninyerola, M., Moré, G., Pons, X., 2015b. Linking land cover dynamics with driving forces in mountain landscape of the Northwestern Iberian Peninsula. Int. J. Appl. Earth Obs. Geoinf. 38, 1–14. [CrossRef]
- Rost, J., Hutto, R.L., Brotons, L., Pons, P., 2013. Comparing the effect of salvage logging on birds in the Mediterranean Basin and the Rocky Mountains: Common patterns, different conservation implications. Biol. Conserv. 158, 7–13. [CrossRef]
- Salaverri, L., Guitián, J., Munilla, I., Sobral, M., 2019. Bird richness decreases with the abandonment of agriculture in a rural region of SW Europe. Reg. Environ. Chang. 19, 245–250. [CrossRef]
- Santos, X., Badiane, A., Matos, C., 2016. Contrasts in short- and long-term responses of Mediterranean reptile species to fire and habitat structure. Oecologia 180, 205–216. [CrossRef]
- Santos, X., Belliure, J., Gonçalves, J.F., Pausas, J.G., 2022. Resilience of reptiles to megafires. Ecol. Appl. 32, 1–14. [CrossRef]
- Scozzafava, S., De Sanctis, A., 2006. Exploring the effects of land abandonment on habitat structures and on habitat suitability for three passerine species in a highland area of Central Italy. Landsc. Urban Plan. 75, 23–33. [CrossRef]
- Stephens, J.L., Ausprey, I.J., Seavy, N.E., Alexander, J.D., 2015. Fire severity affects mixed broadleaf–conifer forest bird communities: Results for 9 years following fire. Condor. [CrossRef]
- Taillie, P.J., Burnett, R.D., Roberts, L.J., Campos, B.R., Peterson, M.N., Moorman, C.E., 2018. Interacting and non-linear avian responses to mixed-severity wildfire and time since fire. Ecosphere 9. [CrossRef]
- Tingley, M.W., Ruiz-Gutiérrez, V., Wilkerson, R.L., Howell, C.A., Siegel, R.B., 2016. Pyrodiversity promotes avian diversity over the decade following forest fire. Proc. R. Soc. B Biol. Sci. [CrossRef]
- Torres, J., Gonçalves, J., Marcos, B., Honrado, J., 2018. Indicator-based assessment of post-fire recovery dynamics using satellite NDVI time-series. Ecol. Indic. 89, 199–212. [CrossRef]
- Uyttewaal, K., Prat-Guitart, N., Ludwig, F., Kroeze, C., Langer, E.R. (Lisa., 2023. Territories in Transition: how social contexts influence wildland fire adaptive capacity in rural Northwestern European Mediterranean areas. Fire Ecol. 19. [CrossRef]
- Vallecillo, S., Brotons, L., Thuiller, W., 2009. Dangers of predicting bird species distributions in response to land-cover changes. Ecol. Appl. 19, 538–549. [CrossRef]
- Watson, S.J., Taylor, R.S., Nimmo, D.G., Kelly, L.T., Haslem, A., Clarke, M.F., Bennett, A.F., 2012. Effects of time since fire on birds: how informative are generalized fire response curves for conservation management? Ecol. Appl. 22, 685–696. [CrossRef]
- White, A.M., Manley, P.N., Tarbill, G.L., Richardson, T.W., Russell, R.E., Safford, H.D., Dobrowski, S.Z., 2015. Avian community responses to post-fire forest structure: implications for fire management in mixed conifer forests. Anim. Conserv. 19, 256–264. [CrossRef]
- Wretenberg, J., Lindstrom, A., Svensson, S., Part, T., 2007. Linking agricultural policies to population trends of Swedish farmland birds in different agricultural regions. J. Appl. Ecol. 44, 933–941. [CrossRef]
- Zakkak, S., Kakalis, E., Radovic, A., Halley, J.M., Kati, V., 2014. The impact of forest encroachment after agricultural land abandonment on passerine bird communities: The case of Greece. J. Nat. Conserv. 22, 157–165. [CrossRef]
- Zozaya, E.L., Brotons, L., Herrando, S., Pons, P., Rost, J., Clavero, M., 2010. Monitoring Spatial and temporal dynamics of bird communities in Mediterranean Landscapes affected by large wildfires. Ardeola 57, 33–50.
Figure 2.
Inter-annual variability (‘temporal component’) of burnt area (‘AB_mean’), mean (‘Sev_mean’) and standard deviation of burn severity (‘Sev_sd’) at plot level. Boxplots show the variability (‘spatial component’) across the 206 census plots affected (‘Burnt) and not affected (‘Un-burnt’) by fire over the time period (2010-2020). For all boxplots, lower and upper whiskers encompass the 95% interval, lower and upper hinges indicate the first and third quartiles, and the central black line indicates the median value.
Figure 2.
Inter-annual variability (‘temporal component’) of burnt area (‘AB_mean’), mean (‘Sev_mean’) and standard deviation of burn severity (‘Sev_sd’) at plot level. Boxplots show the variability (‘spatial component’) across the 206 census plots affected (‘Burnt) and not affected (‘Un-burnt’) by fire over the time period (2010-2020). For all boxplots, lower and upper whiskers encompass the 95% interval, lower and upper hinges indicate the first and third quartiles, and the central black line indicates the median value.
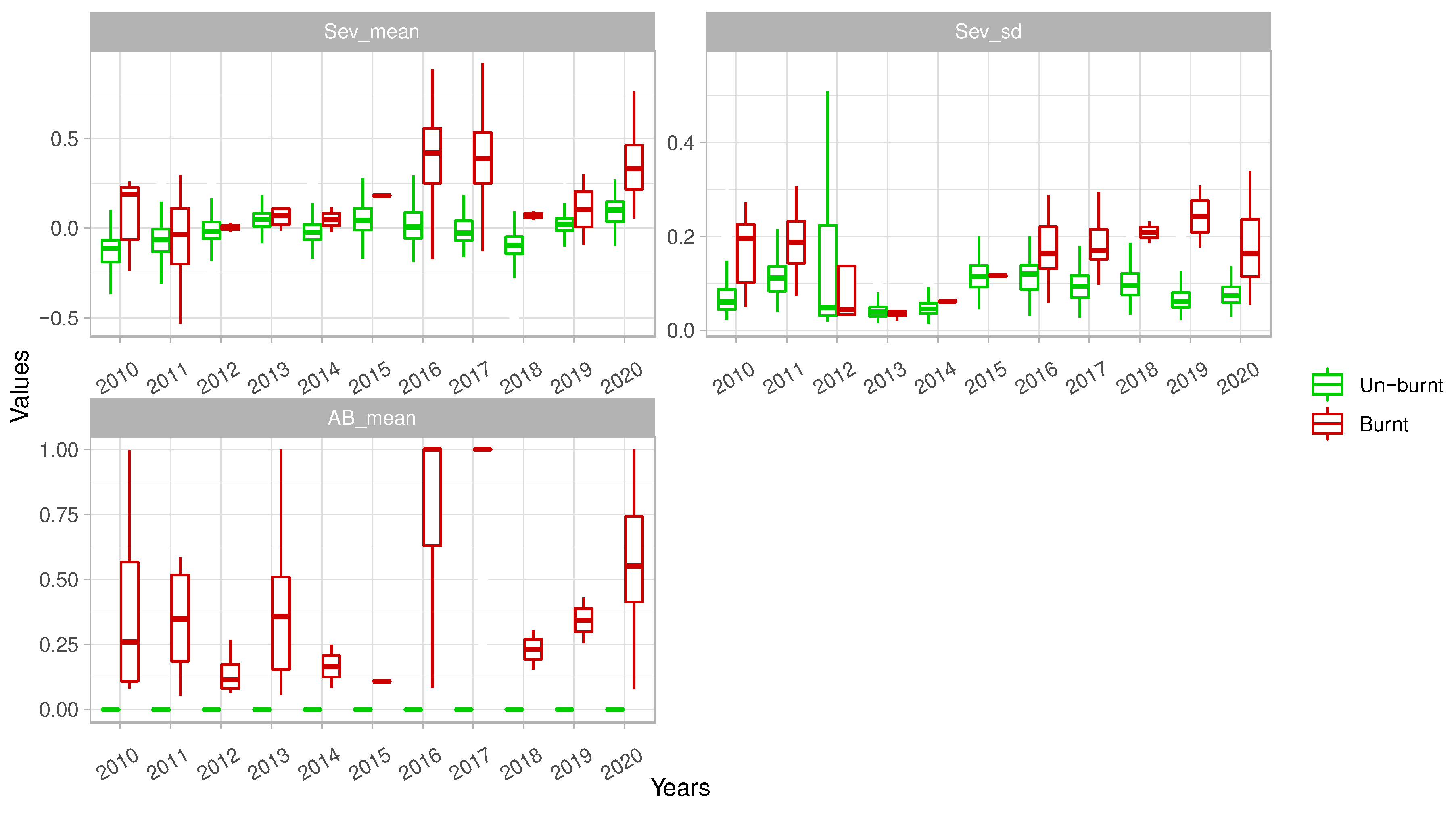
Figure 3.
Number of species significantly correlated (p-value < 0.05) with each exploratory variable: ‘AB_mean’ (average value of annual burn area), ‘CropGrass’ (% of cropland and grassland), ‘Forest’ (% of forest), ‘Sev_mean’ (average value of annual burn severity), ‘Sev_sd’ (standard deviation of burn severity), ‘TSF’ (time since last fire). ‘Positive’ and ‘Negative’ indicate the number of species with positive or negative relationships between the response and exploratory variables (determined by the ß parameter).
Figure 3.
Number of species significantly correlated (p-value < 0.05) with each exploratory variable: ‘AB_mean’ (average value of annual burn area), ‘CropGrass’ (% of cropland and grassland), ‘Forest’ (% of forest), ‘Sev_mean’ (average value of annual burn severity), ‘Sev_sd’ (standard deviation of burn severity), ‘TSF’ (time since last fire). ‘Positive’ and ‘Negative’ indicate the number of species with positive or negative relationships between the response and exploratory variables (determined by the ß parameter).
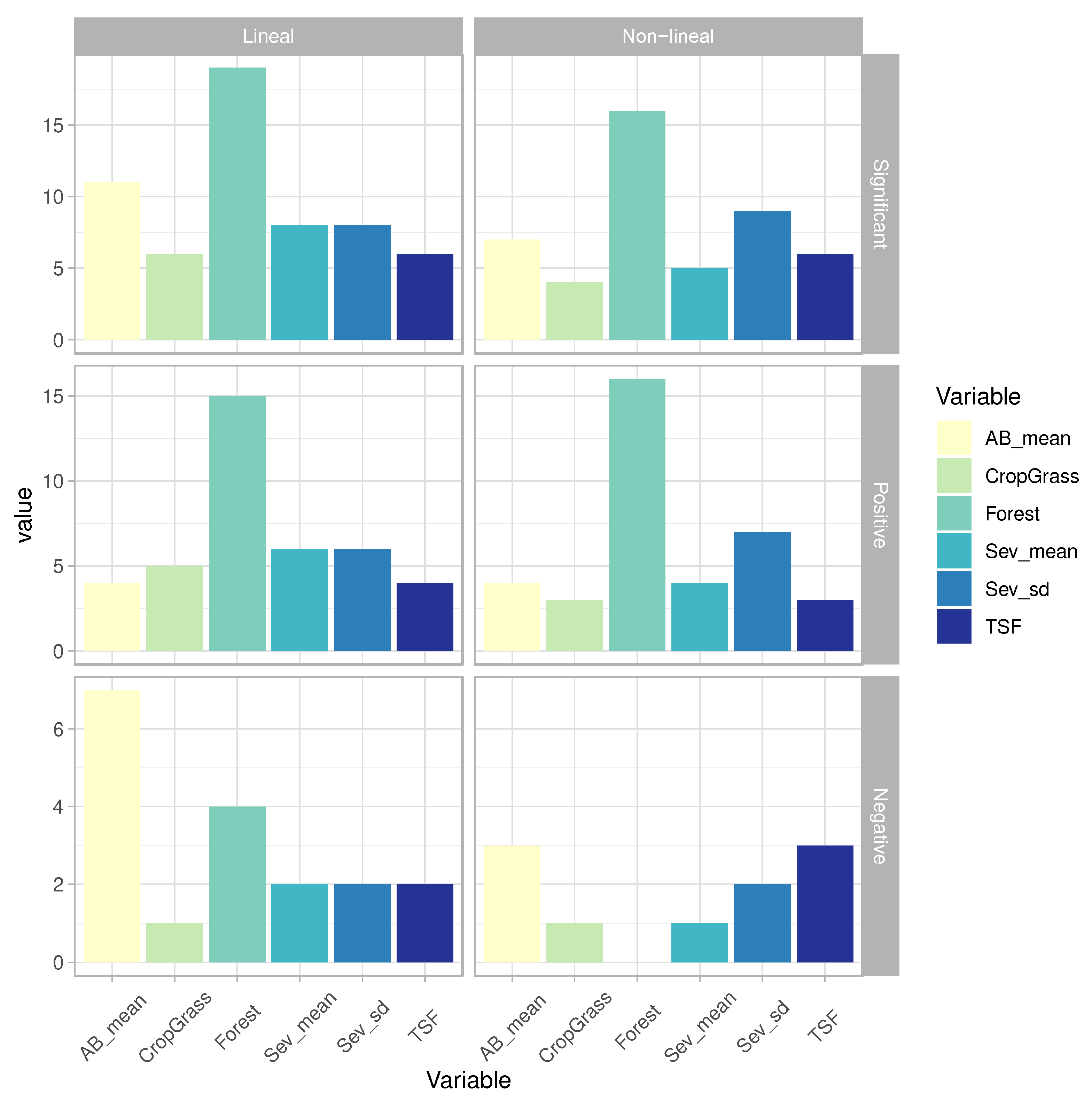
Table 1.
Satellite sensors used for each year, spatial resolution, and the corresponding NIR and SWIR bands used to calculate the Normalized Burn Ratio (NBR).
Table 1.
Satellite sensors used for each year, spatial resolution, and the corresponding NIR and SWIR bands used to calculate the Normalized Burn Ratio (NBR).
| Satellite | Resolution | Years | NIR band | SWIR band |
|---|---|---|---|---|
| Landsat-5 TM | 30 m | 2010 - 2011 | B4 | B7 |
| Landsat-7 ETM+ | 30 m | 2012 | B4 | B7 |
| Landsat-8 OLI/TIRS | 30 m | 2013 - 2014 | B5 | B7 |
| Sentinel-2 | 10 m | 2015 - 2021 | B8 | B12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



