
Submitted:
26 June 2024
Posted:
10 July 2024
Read the latest preprint version here
Preprints on COVID-19 and SARS-CoV-2
Abstract
The severe acquired respiratory coronavirus–2 (SARS–CoV-2) infection has initiated both acute and chronic COVID–19 disease between 2020 and 2023, currently evolving with other homologous prior coronavirus strains of the Nidoviridae order, which encompasses other prevalent alpha/ beta coronaviruses, but also the Middle East Respiratory Syndrome (MERS-CoV) and SARS-CoV-1, with recent SARS–CoV–2 variants, increasing demands for effective immunogens and therapeutic approaches that will reduce global disease burden and further infection from SARS–CoV-2 affected individuals that may experience post acute sequelae (PASC) or “Long COVID”. Following a worldwide programme of prophylactic vaccination, there is still a dilemma in the efforts to find prophylactic and early therapeutic approaches that would treat novel SARS-CoV-2 variants and prevent future epidemics or pandemics within host human and animal populations, where zoonotic or cross species transfer naturally occurs. Concerns about viral immune escape intersect at a specific point; a gained evolutionary ability of several viruses to co–infect and compete against previous scientific advances since 1796 that remain undetected or asymptomatic during the early stages of infection progressing to symptomatic and severe disease via the double methylation of the 5' end of eukaryotic DNA or RNA-based viral genomes, the 7-MeGpppA2’-O-Me cap, and its double methylation capping process is performed by the activated viral 2’ - O - Methyltransferase (MTase) enzyme, a complex of two viral non-structural proteins (NSPs) joined together through an activation process (NSP10/16) and by N7-Methyltransferase (N7-MTase/NSP14), respectively. Moreover, it was discovered that polymorphic viruses translate NSP1, which prevents the activation of various Pattern Recognition Receptors (PRRs), and consequently, detection of Pathogen-Associated Molecular Patterns (PAMPs) and Damage-Associated Molecular Patterns (DAMPs) alike. NSP1 also silences important interferon-encoding genes (INGs) and interferon-stimulated genes (ISGs), is signalled in a paracrine manner to neighbouring cells, and that induces the apoptosis of host cells, inducing an effect of “trace erase” effect and making the viral infection as immunologically “invisible” as possible during the initial, key stages of viral replication and distribution, all such mechanisms occurring independently of the viruses in cause. Another important viral NSP is NSP14, as it plays two functional roles that are independent of each other; to produce new viral genetic material for the purpose of maintaining the validity of the viral genome as well, and not just transfer a methyl group to the 5’ end of the viral genome. Other viral NSPs share a role with NSP1, 10, 14 and 16 in directly suppressing the activation of PRRs and ISGs, and all such viral proteins help the virus in its process of self-camouflaging against first- and second-line immunity, thereby often severely impacting the quality of the produced adaptive immune responses. The outcome of all such phenomena is the sharp decrease in the host Type I and Type III interferons' (IFNs) rate of synthesis by the host cells, that would usually occur and affect homeostatic cellular pathways, resulting in further viral replication and induced apoptosis. Nonetheless, effects of microbial immune evasion during the development of other viral or carcinogenic pathologies are not widely known. In short, polymorphic viruses developed a proportionate evolutionary response against developed adaptive immune responses, by currently relying on gaps mostly situated in the natural immune system in their process of molecular self-camouflaging. Scientists developed numerous approaches of early treatment that generally showed good success rates and fewer risks of adverse events, and the still early present stages of COVID-19 research should also be taken into consideration whilst filtering for the most appropriate solutions. For example, the administration of recombinant human interferons I and III into the nasal mucosa cellular layer, as key mediators of anti–viral activity, can simulate intracellular infection and stimulate cellular activity in a timely manner, training the innate and adaptive immune system cells to develop and appropriately stimulate an adequate immune response through B and T cells. Another example could involve the treatment of natural and adaptive lymphocytes with a low dose of IFNs I and possibly III, prior to their insertion into the host lymphatic system, possibly alongside additional recruitment of plasmacytoid dendritic cells (pDCs) as further interferon “factories”, all with the purpose of early infection management. It might be that focusing on directly offering the immune system the information about the genetics and protein structure of the pathogen, rather than training its first-line mechanisms to develop faster, excessively increases its specificity, making it reach a level that brings the virus the opportunity to evolve and escape previously-developed host immune mechanisms. It is until the scientific community realises this potentially crucial aspect that large proportions of the world population will probably continue to face serious epidemics and pandemics of respiratory diseases over the coming several decades, evidenced with dengue fever and more recently, monkeypox and possibly avian flu. Of note, it has been indicated that IFN I and / or III display significant immunising, early therapeutic and clinical disease onset-attenuating effects for many other microbial evoked diseases, as well as for a number of oncological diseases.
Keywords:
covid-19
; pandemic
; immune evasion
; first-line immunity
; viral evolution
; interferon
; dendritic cells
; cytokines
; chemokines
; natural lymphocytes
; adaptive lymphocytes
; innate immunity
; adaptive immunity
; vaccinology
; therapeutics.
Introduction
Interferons (IFN) are glycoproteins with antiviral activity that can be produced by dendritic cells (DCs), comparatively quicker upon infection that stimulate the immune system into its innate and adaptive compartments. WIthin these immune system compartments are IFN receptors located within not just DCs but also B-lymphocytes, monocytes, macrophages (Mϕ), and T-lymphocytes, as well as glial cells, neurons, and among others. These cellular receptors initiate both downstream and upstream effects that include initiating phagocytosing pathogenic viruses as soon as they infect such cells. There are three principal classes of IFN that are Type I, Type II and Type III with the first and the last that have more specific antiviral activities, whilst type II IFNs focus more upon the development of pro–inflammatory responses earlier in host infection. Interferons (IFNs) were originally characterised in 1957 by Isaacs and Lindemann (Stanifer M. L. et al., 2020), with comparatively recent differentiation of IFN subtypes further, as below. Recently, scientists further discovered that a number of plants also generate interferon–stimulating proteins, and we believe that by modifying dosages of such natural or synthetic proteins there could be a potential improvement of cellular effects of IFNs in various pathologies. The main focus in clinical research during viral pandemics like the recent COVID–19 is to balance the cellular immune responses precision and beneficial cellular effects, through ongoing IFN research, as timing of IFN release has been indicated to affect viral infection, propagation, replication and lysis in differential organ compartments and cell systems. The principal commonality between the majority of infectious diseases is that numerous forms of cancer and several proteinopathies is that all disease categories generally involve a late and exaggerated development of Type I and Type III IFN based immune responses, and this potentially means IFNs could play essential prophylactic, and sometimes also early therapeutic roles in oncogenic, retroviral and neurodegenerative forms of disease depending on disease aetiology. Therefore, we emphasised upon the search for safety data with regards to both clinical approaches and the physiology of each type of IFN signalling. The collected results and bibliographic references were then listed specifically, according to topic and degree of relevance to the title of the study. We present here projective analysis and clinical trials concerning the risks and efficacy of low-dose nasal recombinant human Type I and III IFN, with effects of subcutaneous dosing of Type I and III IFN that will require further research,.by means of gathering safety data and collateral evidence around the study’s principal scope in order to fully elucidate the nature of IFN cellular responses and pathways for therapeutic benefit.
i) Overview to Interferon Cellular Types
Type I IFNs are synthesised and secreted by various types of human cells, with timing of nuclear transcription resulting in differential effects indicated as key in host cellular responses to pathogens and within cancer cellular proliferation. For example, early synthesis and release from plasmacytoid dendritic cells (pDCs) that produce both Type I IFNs subtypes IFN–α and IFN–β can affect antiviral responses through innate and adaptive immune cell compartments initiating apoptosis in macrophages. Type I IFNs also include IFN–δ, IFN–ε, IFN–κ, IFN–τ, IFN–ω, and IFN–ζ subtypes, whilst Type III IFNs include IFN–λ and subtypes (Reizis B., 2019). There are more classifications of each subtype of such interferons that can vary between host species. To clarify further, human IFN–α presently consists of at least 18 subtypes, which include IFN–α1, IFN–α2, IFN–α5, IFN–α7 and IFN–α14, whilst IFN–λ consists of IFN–λ1, IFN-λ2, IFN-λ3 and IFN-λ4, and among the listed examples of IFN-α subtypes, IFN-α2 and IFN-α14 are glycosylated (George J and Mattapallil JJ, 2018). The first IFN subtype is glycosylated on its Nitrogen-terminus (N-terminus, left hand-side), whilst the latter is glycosylated on its Oxygen-terminus (O-terminus, right hand-side). Recombinant human IFN-α2 has often been used during clinical trials for the purpose of safety and efficacy testing, with numerous trials indicating safety and potential in disease treatment. In addition, all subtypes of IFN-β are N-glycosylated. Furthermore, another clinical study showed that the addition of a glycosyl group on IFN-λ4 raised its level of anti-inflammatory and antiviral efficacy (Chung J. H. et al., 2020). It is important to note that glycosylated interferons have a greater stability and thereby display more powerful antimicrobial effects (Li S. et al., 2018). Glycosylated IFNs perform a molecular interaction with the carbohydrate molecules present on the plasma membrane of cells and they may have better binding affinities to the IFNAR1/2 and IFNLR1/IL10R2, and an easier overall access into their cells. On the other hand, Type II IFNs include IFN−γ. Whilst IFN-β and IFN-λ are produced by all kinds of human cells, IFN-α are generally produced by immune cells, including plasmacytoid Dendritic Cells (pDCs) (Kim Y. M. et al., 2021). It is important to note that research involving Type III IFNs is still in early stages, and much of the specific data about early therapy and immunisation is unknown although comparatively well documented in bacterial infection, but indications are that potential use could be at least as effective for as naturally produced Type II IFNs.
ii) Pathological context
Immune system modulation and/or evasion could represent a serious immunological problem of the 21st century, and is the result of less knowledge with a comparative shift in the evolutionary development between the human immune system and prior pathogens, like smallpox, human immunodeficiency virus (HIV), and Streptococcus, that took place throughout history. Respiratory viruses represent the pathological category of viruses whose mechanisms of immune evasion are among the most prevalent., It is likely that regular mutations also lead to the development of ongoing seasonal variants of the Influenza A positive-sense single-stranded RNA virus (+ssRNA virus) and the Haemophilus Influenzae bacterium (with 198 potential sub–type combinations for the latter). The avian Influenza A variant (H5N1) has undergone mutations of concern, with a few recent events of zoonotic spillover into humans, and severe forms of Influenza A may favour severe morbidity following secondary bacterial infection. H. influenzae is composed of haemagglutinin (HA) and neuraminidase (NA) protein antigens that co–exist alongside SARS–CoV–2 viral antigens that include spike (S), membrane (M), nucleocapsid (N) and envelope (E) proteins, and an effective restriction of an increased viral and pathogenic protein load is produced by the initial, sensitised ability of the immune system to react and lyse infectious viruses effectively during replication. Generally speaking, it seems that powerful microbes evade their host cell’s immune mechanisms faster and develop trickier methods of direct and indirect molecular self-camouflaging than microbes causing routine infectious diseases. Four universal methods of microbial immune escape seem to comprise the phenomenon of pathogenic DNA or RNA 5’ capping, in which the microbe adds two methyl (CH3-) groups to the 5’ end of its genome to attempt significant evasion from Pattern Recognition Receptor (PRR)-mediated recognition of Pathogen-Associated Molecular Patterns (PAMPs), which are molecules specific of pathogenic agents, and Damage-Associated Molecular Patterns (DAMPs), which are molecules produced due to induced cellular damage and lysis, meaning that recognition of microbe-induced damage and lysis of cells is also impaired; the direct inhibition of the activation and expression of interferon-encoding genes (INGs) to facilitate viral replication and distribution of its increasing load to neighbouring cells and tissues, the method of mitochondrial disruption via the increased production rate of reactive oxygen species (ROS), and the method of viral usage of inter-cellular channelling nanotubes between host cells, which are developed by many cells to facilitate paracrine signalling of important proteins between each other, and viruses utilise the same method of paracrine signalling for the purpose of facilitating the exocytosis and paracrine signalling of non-structural proteins that will prevent the activation and expression of interferon-encoding genes (INGs) in uninfected cells as well, which tremendously aids the virus in escaping recognition by such PRRs, which include Toll-Like Receptors (TLRs) and RIG-I-Like Receptors (RLRs) (Amarante-Mendes D. P. et al., 2018). Such a method of molecular self-camouflaging facilitates an exponential increase of the viral load whilst preventing much of the development of immune responses that normally induce clinical signs and symptoms characteristic of infectious disease (Sherer N. M. et al., 2008). The final evolutionary objective seems to be a prevention of Type I and possibly of Type III Interferon production by the infected cells. To specify, the non-structural proteins 1 (NSP1) produced by Dengue and Zika viruses were discovered to be packaged into exosomes, exocytosed and transmitted to neighbouring cells as soon as they were translated by the viral genome to prevent signalling of IFN I and III in them also (Safadi D. E. et al., 2022), and this molecular event likely indicates that oftentimes, the NSP1 produced by the novel coronavirus is also transmitted to neighbouring cells often faster than secreted interferons are. Moreover, it was discovered that the NSP1 produced by the H5N1 strain of Influenza A Virus (IAV) induces the caspase-induced apoptosis of their host cells, thereby seemingly bringing an effect of “trace erasing” to “ensure” that the “intelligence” of the virus is ahead of the “intelligence” of the host immunity (Zhang C. et al., 2010), and such phenomena may also widely apply to SARS-CoV-2 infection. A relationship of proportionality between disease mortality rate and extent of NSP1-induced apoptosis may be projected. Likewise, the extent of viral evolution over human and animal natural immunity seems to be a proportionate reaction to the developed adaptive immune responses and memory over past viruses and viral variants, ultimately according to the pattern behind Newton's Third Law in Motion, given the foundational role that physics plays in material sciences. The gap currently used by novel polymorphic viruses on their evolutionary path to gain advantage over the immune methodology of responses is very likely situated mostly in the natural immune system.
Other viruses of major concern, including the novel Monkeypox Virus (MPV), Ebola virus (EBOV), Marburg virus, the Human Immunodeficiency Viruses 1 and 2 (HIV-1 and HIV-2), as well as the Sendai, Hendra and Nipah viruses, displayed significant molecular activity that antagonises the synthesis and exocytosis of Type I and often Type III Interferons. One important common method of the majority of such viruses is the translation of viral proteins that cleave the interferon-encoding mRNA. For example, the EBOV viral genome translates the VP24 and VP35 proteins to substantially inhibit interferon production (Ilinykh et al., 2015), although the latter protein was not found to suppress IFN production in plasmacytoid dendritic cells (Leung et al., 2011). Sensitised Type I Interferon-mediated immune responses were found to considerably weaken the EBOV infection (Kuroda et al., 2020). Furthermore, scientists who induced a loss-of-function mutation in the EBOV gene encoding VP35 discovered that such a variant of the filovirus is not virulent and mounts a long-term immune memory against the regular Ebola viral variant (Woolsey et al., 2019). Moreover, one study showed the ability of HIV-1 to suppress Type I Interferon synthesis and transmission by producing the Viral infectivity factor nonstructural protein (Vif), which targets the cGAS-STING-related signalling cascades by interacting with the cellular Tyrosine (Tyr/Y) Phosphatase enzyme known as SHP-1, leading ultimately to the dephosphorylation of STING at the Y162 amino acid position (Wang Yu et al., 2022). Another study highlighted the ability of HIV-1 to evade TLR-8 detection via the translation and signalling of the snapin viral protein to neighbouring cells (Khatamzas et al., 2017).

Table 2.
List of clinical trials involving the administration of Type I and Type III Interferons for both COVID-19 and non-COVID-19 purposes.
Table 2.
List of clinical trials involving the administration of Type I and Type III Interferons for both COVID-19 and non-COVID-19 purposes.
| NCT Order | Disease status | NCT & Phase - Type I IFNs | NCT & Phase - Type III IFNs |
| I | COVID-19 | NCT04469491, Phase 2 (Oral/Nasal) | NCT04354259, Phase 2 (Pegylated, subcutaneous) |
| II | COVID-19 | NCT04469491, Phase 2 (Oral/Nasal) | NCT04967430, Phase 3 (Pegylated, subcutaneous) |
| III | COVID-19 | NCT04732949, Completed (Oral/Nasal) | NCT04534673, Phase 2 (Pegylated, subcutaneous) |
| IV | COVID-19 | NCT04350281, Phase 2 (Subcutaneous) | NCT04343976, Phase 2 (Pegylated, subcutaneous) |
| V | COVID-19 | NCT05381363, Phase 1/2 (Oral/Nasal) | NCT04727424, Phase 3 (Pegylated, subcutaneous) |
| VI | IAV infection | NCT00895947, Completed (Oral/Nasal) | Not available yet |
| VII | Hepatitis B/C | NCT00917358, Completed | NCT01204762, Completed (Pegylated, subcutaneous) |
| VIII | HIV Infection | NCT01295515, Completed | NCT01866930, Completed (Pegylated, subcutaneous) - HCV patients co-infected with HIV |
| IX | Cancer | NCT01462773, Completed | NCT04469491, Unknown status |
| X | Cancer | NCT00278174, Completed | Not available yet |
Figure 1.
The cGAS-STING pathway inducing Type I Interferon, Interleukin-6 and Tumor Necrosis Factor expression and signalling (Created with BioRender.com under the IV2510F0GI License Number).
Figure 1.
The cGAS-STING pathway inducing Type I Interferon, Interleukin-6 and Tumor Necrosis Factor expression and signalling (Created with BioRender.com under the IV2510F0GI License Number).
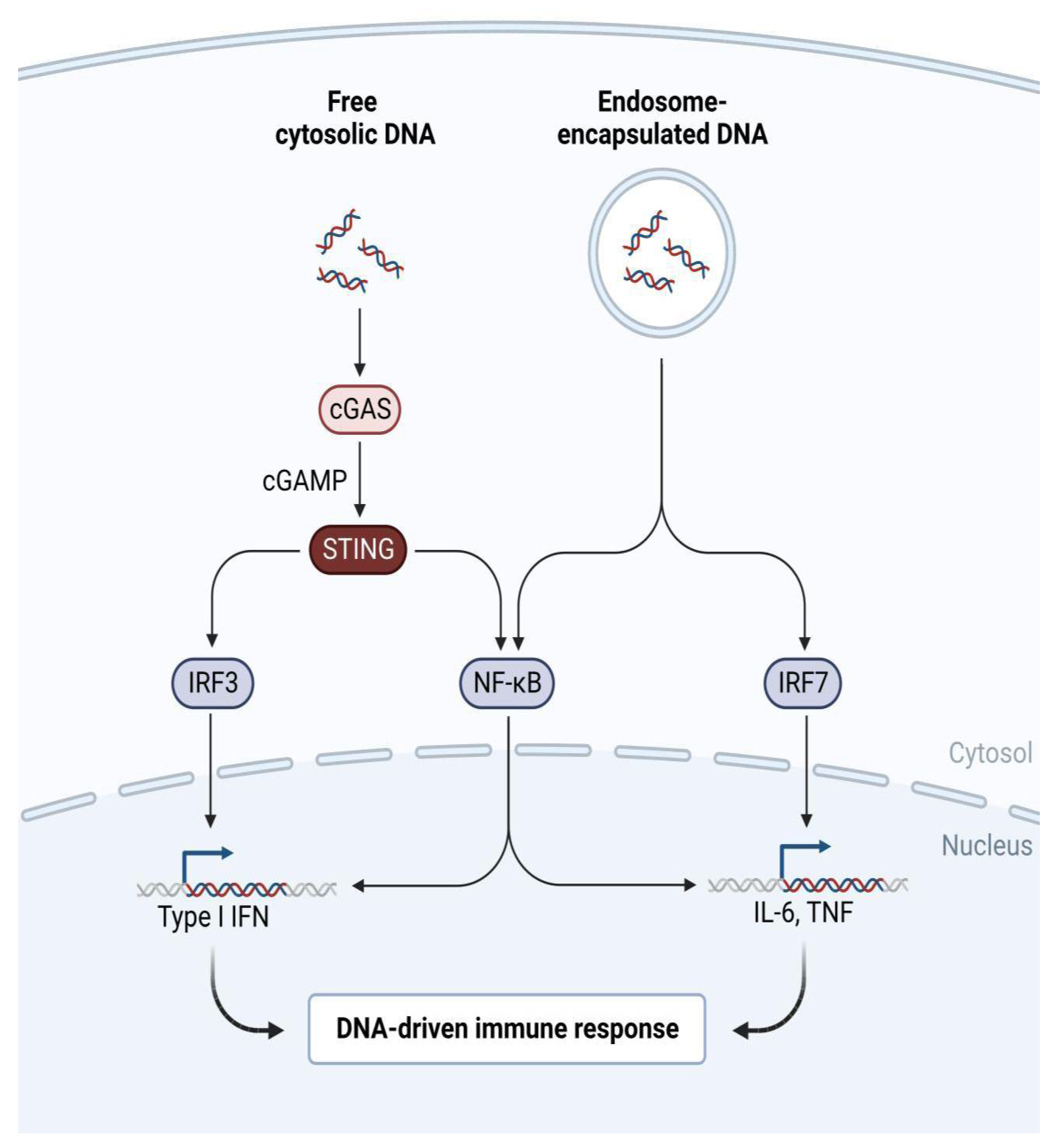
Overall, the evolutionary problem of viral antagonism against the depths of human immunity, the interferon system, may be more voluminous than previously believed. The increased ability of the novel coronavirus to evade recognition of cellular Toll-Like Receptors leads to a desensitised and delayed ability of the immune system to recognize, and let alone react against the virus, which explains the significant number of individuals that develop symptoms several days after an infection takes place, and the phenomenon is also known as a silent inflammation. Interestingly, a prolonged asymptomatic stage of the disease does not mark an immune preparation against an exponential increase of the viral load, but a lack of it. All infectious diseases begin with an asymptomatic stage, but the majority of such diseases include a far briefer stage that does not implicate the manifestation of clinical symptoms. The ability of microbes to evade first-line immunity could be proportional with the duration of asymptomatic disease.
iii) Limitations
It is important to note the difference of Type I and Type III IFNs with regards to the stage of research concerning their prophylactic and early therapeutic effects. Moreover, it is also essential to note a common problem that led to an interruption of some clinical trials; a lack of funding, which was often accompanied by insufficient participants. Likewise, in spite of the wide bioavailability of human recombinant interferons as such, as well as their significant impact upon the severity of multiple diseases of concern, there seems to be a slight impediment of funding and human resource availability in a number of cases, particularly in areas of the world affected by precarious welfare and sanitary conditions, given that wider research involving IFNs as such began just after the mid-twentieth century. Despite such occurrences, which could be a result of barriers met commonly during research efforts that later prove to be successful, the novel discoveries of natural immune effectiveness and of its existent “memory” strongly encourage researchers to bring a much wider inclusion of natural immunity in their process of therapeutics and vaccine-based research. Adverse reaction-related limitations generally include the utilisation of larger dosages of human recombinant Type I and Type III IFNs, in latter stages of infectious diseases that display more concerning magnitudes of morbidity. In such cases, complications involved the onset of autoimmune diseases and increased rates of infectious disease pathophysiology. Moreover, more severe adverse events included the onset of systemic inflammation, as well as neurological diseases, including depression and anxiety-related pathologies. The “double-edged sword”-like nature of interferons of such classes with regards to quantity, type of disorders and their stages may explain why there are existing contexts in which interferons cause more harm to the host organism than benefit. Likewise, a significant extent of caution requires to be practised during the projection and development of such novel approaches in clinical research covering human immunity, regardless of the extent of efficacy interferons would display if administered within the appropriate medical context.
Methodology
Using MESH searches and/or the clinicaltrials.gov site, we used the keywords below and detected IFN NCT trials listed globally, without date restriction, which were listed as follows with 209 “type I IFN”, 330 “type II IFN” but also 153 were classified as “ type III IFN”. Therefore, according to this a total of 692 NCTs were either entered or registered, awaiting, in progress, or completed prior to this analysis publication with or without results. However using keyword searches “IFN alpha” and/or “IFN beta” totalled 1845, but also “IFN gamma” totalled 2477 with 25 “IFN lambda”, 3 “IFN epsilon”, “1688 IFN delta”, 21 “IFN kappa” and 21 “IFN omega”. Therefore a total of 692 NCT are registered according to IFN type, however 6077 individual searches appear according to type of IFN. Only peer-reviewed scientific journals concerning the effectiveness of early therapies using low-dose Type I and III IFNs were analysed using the [MeSH] algorithm on PubMed. Data collected through numerous clinical trials (NCTs) implicating a low-dose IFN nasal spray and high-dose Type I, Type II or Type III IFN was collected, and the kinds of NCTs included preliminary, double-blind and meta-analyses, but also mainly in phase 2 and phase 3. We reviewed literature addressing similarities and differences between SARS-CoV-2, three other respiratory viruses, and a retrovirus, to determine other options open to clinical research or researchers for further investigation.
Discussion
1. Important highlights of innate immune evasion
Pathogenic microbes and the human organism have co–evolved since the natural selection of humans, and it is almost a matter of certainty that numerous microbes used the depths of first-line human immunity as a primary model for evolution. The novel SARS–CoV–2 impairs the IFN system in several methods. Initially, the virus aids the production of reactive oxygen species (ROS), which disrupts the activity of mitochondria and then, viral copies cross intracellular tunnelling nanotubes (TNTs), which were created for the purpose of cell-to-cell communication. Events as such prevent significant recognition by various pattern recognition receptors (PRRs), by translating open reading frames 6 and 8 (ORF 6 and 8), that downregulate the expression of major histocompatibility complex I (MHC–I) related genes and indirectly suppressing the immune stimulatory action of Type I and III IFNs by means of preventing the synthesis of ISGs proteins that have powerful anti–viral and anti–inflammatory molecular actions, and by translating 16 non-structural proteins (NSPs), that directly suppress the immune–stimulatory action of type I IFN and type III IFNs by preventing translation, as well as the autocrine and paracrine secretion of IFN proteins (Rubio-Casillas A. et al., 2022), and that effect lysis of ISG products that play a major role in antiviral defence (McKellar et al., 2021). As above, the SARS-CoV-2 ORF3a, ORF9b and ORF10 proteins also play a role in evading Type I and Type III IFN mediated immune responses by inhibiting nuclear transport of IRF3, NF-kB and STAT polymers, leading to an overall down-regulated expression of interferon-induced genes (IFI) (Rashid F. et al., 2022). Interestingly, prompt and robust translation and exocytosis of IFN–α (Glennon-Alty L. et al., 2021) and IFN–λ (Zanoni I. et al., 2017), as is historically though to occur by DCs, inhibits the production and degranulation of ROS quicker, thereby tending to secure further robust IFN–λ signalling from neighbouring cells, and both IFNs play a significant role in regulating the activation and recruitment of neutrophils. The SARS–CoV–2 genome produces a number of non-structural proteins (NSP) that significantly decrease the amount of cellular secreted IFN, either by viral self-camouflage or by cleaving IFN–producing mRNA and downregulating the activation of cellular transcription factors (TFs) that translate interferon inducible proteins (IFI). Similar events occur in the cases of respiratory syncytial virus (RSV) (Bossert et al., 2002, Hijano et al., 2019, H. influenza and subsequent variant infections (Ma D. Y et al., 2015), as both viruses synthesise NSP1 that inhibit the translation of type I and type III interferon-encoding genes leading to stimulation or inhibition of IFN receptor activity (e.g., IFNAR2), whilst RSV translates NSP2 to suppress responses as such (Munir S. et al., 2011). Respiratory syncytial virus (RSV) particularly activates Rab5a host cell proteins to downregulate the activation of type III IFN based signalling cascades (Mo S. et al., 2021). Furthermore, it was discovered that the NSP1 protein, translated by the avian H5N1 strain of the Influenza A virus, induces the caspase-induced apoptosis of the host cell, before it is independently exocytosed and signalled in a paracrine manner, undergoing receptor-induced endocytosis into neighbouring cells. In such a manner, polymorphic viruses like (A)H5N1 Influenza, and very likely SARS-CoV-2 as well, seem to also induce a “trace erasing” effect in order to prevent the buildup of natural and adaptive immune memory during the initial, most critical stages of the viral replication and distribution of the viral load to multiple, distinct types of host tissues (Zhang et al., 2010). This aspect seems to reflect a pattern of developed “counter-intelligence” of viruses as such, by means of imitating the “intelligence” of human natural and adaptive immunity. Despite the fact that scientists first generally observed a lack of specificity behind natural immune mechanisms, it was recently discovered that natural immunity develops “memory" against pathogens after all and likewise, the concept of “trained immunity” now covers the principal mechanisms of natural immune activation, with its quality first covering timing of action and then physical extent (Dagenais A. et al., 2023). Furthermore, there has recently been a collection of scientific data displaying unexpected results with regards to pathogen-derived vaccination of poultry against the H5N1 strain of IAV. Namely, it has been discovered that live attenuated H5N1 IAV-based poultry vaccination resulted in a faster evolutionary process of the viral haemagglutinin (HA) gene, involving thereby an accelerated process of viral zoonosis, with rates of transmission and intensity of virulence increasing as a result. Likewise, efforts of poultry vaccination against H5N1 IAV infection resulted in an accelerated viral evolutionary process, and such a case may further highlight the evolutionary stronghold polymorphic viruses as such have gained upon human first-line and second-line immune defences (Li B. et al., 2023).
With regards to SARS–CoV–2 induced COVID–19 disease, it is important to note methods used to escape animal host immune systems by means of replication and spread to host tissues. Namely, once the virus enters the host, modulation of the activities of cellular pattern recognition receptors occurs, such as Toll-like receptors (TLR) (e.g. TLR3, TLR7 and TLR8), but also others like pathogen-associated molecular patterns (PAMPs) found within host genomes generated through gene transcription and translation that occurs via diverse transcription factors (TFs). Therefore as IFNs generate anti-viral immunity through systemic, organ, tissue and cellular pathways, it is essential to consider the synthesis and expression rate of Type I and III Interferon-encoding genes and proteins (Kim You-Me and Eui-Cheol Shim, 2021). The cellular source and nature of IFN remains unclear. However, recent indications in 2021 indicate that type I IFNs are not produced or at a lesser rate by certain cells during SARS–CoV–2 infection that consist of either innate or adaptive immune cells and others, like macrophages (Mϕ), dendritic cells (DCs) or epithelial cells that could potentially affect immune systemic responses during pathological disease (Niles et al., 2021).
See Diagram 1 (IFN Receptors)
Figure 2.
The interferon proteins present on the surface of plasmacytoid Dendritic Cells (pDCs) as a gateway for the activation and secretion of Interferon-Stimulated Gene (ISG) products with major antiviral activities (Created with BioRender.com with the permission of publication under the XA2510ANAU licence code).
Figure 2.
The interferon proteins present on the surface of plasmacytoid Dendritic Cells (pDCs) as a gateway for the activation and secretion of Interferon-Stimulated Gene (ISG) products with major antiviral activities (Created with BioRender.com with the permission of publication under the XA2510ANAU licence code).
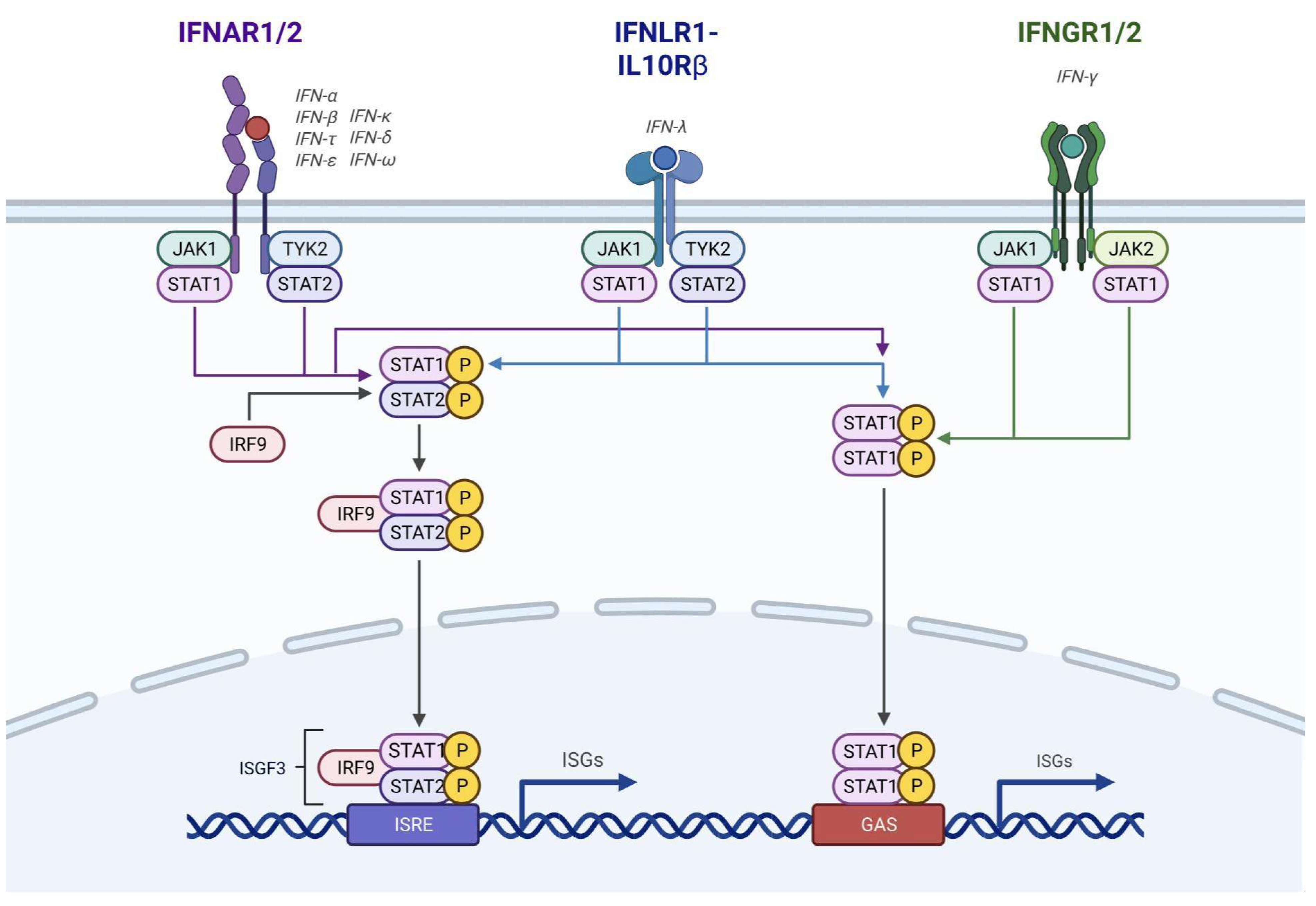
2. SARS-CoV-2 Mechanism of Cellular Action in Disease
Many of the NSPs that SARS-CoV-2 is composed of are defined as follows and include NSP1 responsible for cellular mRNA degradation, NSP2 - arresting cell cycle, NSP3 - 7 propagation, formation of replication complexes, and code for enzyme proteases but also others including NSP8/9/10/13/14/16 – that encode primase/helicases, polymerases, binding proteins and exonucleases and endonucleases. These end with NSP16 2ʹ-O-MTase, an enzyme that caps mRNA, conserved in a viral pocket known as the S-Adenosyl-L-Methionine pocket affecting the enzyme NSP16, which is ultimately activated by joining with NSP10 to form the 2’-O-Methyltransferase enzyme complex. NSP10 is known as an activator protein with NSP16 as an effector protein, and NSP16 transfers a methyl group (-CH3) to the 5’ end of the positive sense (+ss) RNA viral protein once activated. This viral genome 5’ methylation facilitates evasion and recognition of both extracellular and intracellular sensors TLR3, TLR7 but also an extracellular cellular receptor TLR4 expressed differentially and also cellular interferon-encoding genes that have differential and variable activation that facilitating compartmental cellular delay of Type I and III IFN signalling. Oral methioninase had been developed to degrade the S-Adenosyl-L-Methionine viral pocket and lower the ability of NSP13, NSP14 and NSP16 to be conserved (Hoffman et al., 2020), six natural drug-like compounds, including quercetin, oxopowelline and deacetylbowdensine (Saliu T. L. et al., 2021), were developed to prevent the interaction of NSP10 and NSP16, and ultimately, the formation of the NSP10/NSP16 methyltransferase complex. It is known that NSP14 and NSP16 are responsible for the final step of the methyl (-CH3) group transfer to the 5’ end of the viral genome. Whilst NSP14 does not require activation by NSP10 whilst acting as the C-terminal N7 Methyltransferase (N7-MTase) enzyme to place the 7Methyl (7Me) group to the 5’ 7MeGpppA2’-O-Me end of the viral genome, NSP16 does require molecular interaction with and activation by NSP10 to transfer the 2’-O-Methyl (2’-O-Me) group to the 5’ 7MeGpppA2’-O-Me end of the viral mRNA, in order to make it less recognisable as a particle foreign to the host cell (Park G. J. et al., 2022). Nonetheless, NSP14 also acts as an N-terminal 3’-5’ exonuclease (ExoN) enzyme, which requires activation by NSP10 through the formation of the NSP10/NSP14 enzyme complex to become an effector protein, playing a role in producing new viral RNA to maintain a threshold level of viral genomic validity. Even though the process of the viral 5’ end modification is not profoundly understood, it is presently known that a molecular self-camouflaging process of viruses implicates the modification of their 5’ end, which can fully occur if two methyl groups are added to it. One such group is added by the C-terminal N7-MTase enzyme, which does not require activation by NSP10, whilst the other methyl group is added by the NSP16 effector protein, which can only function if it is joined and activated by NSP10 beforehand, to form the 2’ - O-MTase complex. Likewise, NSP14 also likely represents a particularly important target for the development of antiviral drug-like molecules (Samarago M. et al., 2021). Furthermore, several years after the SARS outbreak in 2003, researchers developed a small drug-like peptide called TP29 to prevent an interaction between NSP10 and NSP16 (Wang Y. et al., 2015), and we also hypothesise that the peptide could show a similar extent of efficacy in COVID-19 cases. Furthermore, it is important to note that there is a particularly high binding affinity of sinefungin (SFG) to NSP16 effector methyltransferase component, and S-Adenosyl-L-Homocysteine (SAH) also displayed a lower, but still considerable affinity to NSP16, and both components have the potential of inhibiting NSP16, though more evidence needs to be collected to confirm such data (Mahalapbutr P. et al., 2020). Consequently, SARS–CoV–2 downregulates both autocrine/paracrine signalling and synthesis of Type I and Type III IFNs during critical early stages of infection and replication. Some viral NSPs also inhibit other products of inflammatory gene signatures (IGS) that lyse the viral genome. Therefore, SARS–CoV–2 has an ability to affect the innate and adaptive immune system that usually ensures quicker phagocytosis or stimulates viral cell apoptosis by activation of B cells, T cells and antigen-presenting cells (APCs) that could be considered antagonised by the mechanisms that Type I and Type III IFNs that usually robustly induces clearance of infected cells once released by the newly infected host cells to prevent further viral replication and spread. Moreover, by downregulating IFN synthesis, pathogens and viruses like SARS–CoV–2 gained an advantage over the ability of the immune system to create important antiviral Protein Kinase R (PKR) and anti-inflammatory signals, which are crucial for the development of effective immune responses. Protein Kinase R represents a molecule that plays a major role in the regulation of the ageing process and the mount of cellular antiviral defences, and it also has key functions in regulating intracellular transcription, mRNA translation, rates of apoptosis and cell proliferation (Gal-Ben-Ari et al., 2019). Moreover, the receptor binding domain (RBD) of the SARS–CoV–2 S glycoprotein forms a trimeric complex with a GRP78 chaperone, as well as with the ACE2 receptor, and it is the viral interaction with GRP78 that further enhances infectivity and ultimately, virulence.
GRP78 represents a protein that normally targets and degrades misfolded proteins, and it also plays an important role in preventing the onset of autoimmunity (Quinones Q. J. et al., 2008), and likewise, a viral interaction with the chaperone was shown to amplify its activity and favour the development of autoantibodies (Sabirli R. et al., 2021), ultimately resulting in a decrease of the quality of the overall antiviral immune response. Likewise, not only is it the novel SARS–CoV–2 that utilises numerous methods to delay or evade essential areas of human immunity, but it was also discovered that the virus detects and utilises other host proteins to increase its infective abilities (Carlos et al., 2021). It may be that a research focus that is primed upon the direct providence of the adaptive immune system with the genetic and proteic information of polymorphic viruses, like SARS–CoV–2, and ultimately not concentrated around the necessity of the training of first-line immune mechanisms to develop faster in front of the dilemma of first-line immune evasion, excessively increases the specificity of the host immune responses, which reaches a level that ultimately brings the pathogen the opportunity to evolve and escape previously developed mechanisms, as the developed immune memory does not cover mutated forms and novel antigens. Furthermore, given that early type I IFN and type III IFN based immune signalling pathways systems are associated with higher immuno-competency and in turn, to low extents of energy consumption due to lower viral loads and resulting induced disease, viral immune evasion could have been playing a major role in preventing a full increase of the human lifespan, particularly in world areas with higher incidences of hunger and poverty. Given that the “double-edged sword”-like immunological effects of Type I IFNs has shown visible signs of affecting cellular metabolism and even brain ageing, via the choroid plexus, there are considerable reasons to believe that stimulating early type I IFN and possibly type III IFN based immune signalling in large proportions of the world population by means of immunisation against infectious diseases and important forms of cancer will also play a major role in decreasing the average rates of oxidative stress, mutagenesis and metabolic acidification, leading overall to a better average quality and duration of life.
3. Cellular Mechanism of IFN Action in Disease
The molecular activities of Type I IFN have implications upon cellular mechanisms. These include inhibition of viral replication, activation of nitric oxide synthase (NOS), inactivation of PKR, activation of oligoadenylate synthetase (OAS), regulation of inflammatory responses, peptide presentation by the class I/II major histocompatibility complexes (MHC-/III), activation of adenosine deaminase 1 (ADAR1), maturation and development of leukocytes alongside apoptosis of infected cells. Activation of Protein Kinase R (PKR) leads to downregulated translation of viral RNA encoding pathogenic proteins, whilst the activation of OAS results in the degradation and lysing of RNA and the activation of ADAR1 enables RNA editing (Fatemeh S. et al, 2021). NSP1 activates the phosphatidylinositol 3-kinase (PI3K) pathway that inhibits type I IFN synthesis and activate cellular stress-response proteins, such as heat-shock proteins (HSPs), which inhibit apoptotic pathways of host cells at first, before they stimulate such inductions of host cell death (Ehrhardt et al., 2007). This selective inhibition of apoptosis stimulates a temporal increase of viral load in the host organism. Moreover, the virus further inhibits antigen presentation via the STAT1-IRF1-NLRC5 pathway and the MHC class I complexes present within the majority of nucleated cells but also MHC class II molecules expressed at higher levels on DCs, thereby affecting the specialisation of CD8+ T-lymphocytes (Yoo et al, 2021). Given that the antigen presentation process is affected by targeting of the same pathway as the one induced by Type I IFN activation, we can ascertain that inhibited type I IFN activation could be associated with down-regulated endogenous antigen presentation of peptides, via MHC class II molecules. The downregulation of pattern recognition reception affects signalling from TLR–3, TLR–4, TLR–7, RIG–1 and MDA5 receptors to the IFNA1, IFNA2 and IFNB1 genes. NSP1 lyses molecules of mRNA that encode Type IFNs, whilst NSP16 caps the 5’ end of viral RNA and makes the cell less capable of recognising the viral RNA as pathogenic.
NSP1 was also found to impair the synthesis of Type III IFN, potentially affecting the expression of IFNL1, IFNL2 and IFNL3 gene expression during infections with rotavirus and the porcine epidemic diarrhoea virus and, given that IFN–λ was only discovered in 2003, we have reasons to believe that the synthesis rate of this sub-type of IFN could also be affected during a SARS–CoV–2 infection. In other words, the production of NSP1 by SARS–CoV–2 and Influenza A viral genomes represents a pathogenic evolutionary trait of viruses that is essential for preservation in animal hosts that also includes the avian influenza virus (H5N1 IAV), as these comprise a reaction against the hidden abilities of host organisms to lyse them efficiently from comparative recent research discovery. Given that the rate of interferon-gamma, which is part of Type II IFNs, partially depends upon the synthesis rate of interferon-beta, the impact of the listed non-structural proteins touches the normal synthesis rate of interferon-gamma, which is responsible with a normal expression rate from the infected cells to neighbouring cells. And such signals in turn stimulate the neighbouring cells to produce and send antiviral signals to the immune system. SARS–CoV–2 NSP16 requires activation by NSP10 and hence, NSP10 is known as the activator protein, whilst NSP16 is known as the effector protein. These dimerize to form the 2’-O-Methyltransferase complex, and such nomenclature of the enzyme complex was established because NSP16 caps the 5’ end of the mRNA molecule by attaching a methyl group to it. One study showed that SARS-CoV-2 did not induce IFN production and distribution in pHAE cell cultures. Namely, there was no detectable interferon-alpha of any subtype, and a low rate of synthesis and transmission of interferon-beta1 and interferon-lambda1, with normalised read counts that were lower than the value of 10. Furthermore, several genes involved in the pattern recognition reception and signalling cascade leading to type I IFN synthesis, including RIG-I, MDA5, TBK1, TRAF6, IRF-3 and IRF-7, displayed little to no transcription activities in response to the viral infection, which further indicates the impact of non-structural proteins 1 and 16 upon the sensitivity of the host cell to the virus (Abigail V. et al, 2020).The methyl group is transported from the S-Adenosyl-L-Methionine pocket, which is formed after joining of the S-Adenosyl molecule with the L-Methionine amino acid. The methyl group is transported to NSP13 and/or NSP14, before the NSP16 effector protein binds it. NSP1 represents a problematic IFN antagonist because it significantly suppresses IFN–α and IFN–β synthesis, and was shown to suppress IFN–λ synthesis during rotavirus infection (Iaconis et al, 2021) and porcine epidemic diarrhoea virus (Zhang et al, 2018), thereby potentially amplifying the impairment of innate immune responses. An impairment of such responses can have significant implications upon adaptive immune responses, and likewise, can cause a higher incidence of moderate and severe disease. Given that Type III IFN was only discovered in 2003, has the scientific community investigated the relationship between the viral NSP1 and IFN-λ synthesis, and are there any odds that the SARS-CoV-2 NSP1 suppresses to any extent the synthesis of IFN-λ, since one experiment involving a dosage of IFN-λ2 in mice displayed positive results with regards to a mitigation of the disease (Chong et al, 2021)? Moreover, ORF6 was found to prevent the polymerization of STAT1-STAT2-IRF9 trimer with the ISG3 protein, thereby preventing the expression of important antiviral cytokines (Setaro A. C. et al., 2021). ORF9b was also found to impair Type I IFN and Type III IFN based responses, by targeting important elements of the RIG-I, MDA-5, MAVS, Toll-Like Receptor 3 and cGAS-STING pathways, which normally result in a robust expression of such interferons (Han L. et al., 2021), and such an event further highlights the importance of early Interferon III signals in the prevention of severe forms of COVID-19.
The extracellular matrix (ECM), cytoplasmic and nuclear proteins produced by ISGs, play diverse important roles in restricting IAV infection as well (McKellar et al., 2021), meaning that advanced methods of viral immune evasion may substantially impair diverse intracellular and intercellular mechanisms of antiviral first-line immunity, which include the restriction of viral replication, the inhibition of viral packaging and even the lysing of non-structural proteins, which restrict the expression of Type I and Type III IFNs. Likewise, viral co-infection and evolution with host cells may allow viruses to develop counter-mechanisms unknown to the host that directly inhibit the ability of viruses to replicate, translate their pathogenic proteins and spread to neighbouring cells.
With regards to the chronology of the extent of interferon signalling post-cellular infection, there are three principal categories of Type I and III IFN responses: early response, delayed response and an absent response. The first is followed by a firm restriction of viral load increase, enhanced regulation of pro-inflammatory responses and mild clinical forms of the disease. The second is followed by a dysregulated inflammatory monocyte-macrophage response, severe forms of pneumonia and lung tissue damage. A delayed and exaggerated Type I IFN response will generally overstimulate pro-inflammatory mechanisms and stimulate the development of more severe forms of the disease. The third is followed by a high viral load, longer intensive care unit visits, invasive ventilation and a poor prognosis. Likewise, we can deduce that administration of nasal sprays prophylactically might in turn be more important before or at the onset of the disease than after symptoms occur and our hypothesis is that individuals would receive the nasal spray either before they encounter any clinical forms of the disease, in the first stages of the clinical display, before the peak of the viral load has been reached, or in any other disease stages that do not involve serious symptoms and severe disease (Fatemeh S. et al, 2021). If the nasal spray was administered prophylactically, the sensitivity of the viral +ssRNA sensors will probably increase substantially, which will facilitate a larger extent of early type I IFN and type III IFN translation and exocytosis, alongside a more abundant secretion of the necessary anti-viral and anti-inflammatory ISG products. If the nasal spray is administered at a time that just precedes the moment of first-cell infection, there will already be an immune activity by the time the virus will have attempted evasion of first-line immune signals, and an event as such will make the virus likely much more detectable and vulnerable in the host cells. A delayed clearance of the viral load is likely a common consequence of Type I IFN and III IFN viral immune evasion. Different signalling ratios between Interferon-alpha and Tumour Necrosis Factor-beta usually lead to different outcomes of COVID-19. Mild and moderate forms of the disease involve a balanced ratio between produced interferon-alpha (IFN-alpha) and produced Tumour Growth Factor-beta (TGF-beta), whilst severe forms of the disease involve a far greater extent of Tumour Growth Factor-beta production and signalling. Moreover, severe COVID-19 also involves the inhibition of interferon-gamma (IFN-gamma) and Tumour-Growth Factor-alpha (TFG-alpha), and there is a significant inhibition of the expression of CD18 molecules on the surface of Natural Killer Cells (NK Cells) (Barros-Martins et al, 2022).
Dendritic cells constitute a type of immune cells that play a critical role in the development of responses to viral infectious diseases, such as COVID-19. Dendritic cells are able to recognize and capture viral antigens, process them, and present them to other immune cells, such as T- and B-lymphocytes. This process is known as antigen presentation and is essential for the activation of the adaptive immune response. There are several different subtypes of dendritic cells, each with different functions and surface markers. One important subtype of dendritic cells represents the plasmacytoid dendritic cells (pDCs), which are known for their ability to produce large amounts of interferon I (IFN-I) in response to viral infections. IFN-I is a key component of the innate immune response to viral infections and it is produced by a variety of cells, including dendritic cells, in response to viral infection. IFN-I has a range of antiviral effects, including the inhibition of viral replication, the activation of innate immune cells, and the enhancement of antigen presentation. Impaired first-line immune responses are visibly associated with the recruitment of immature dendritic cells and the development of weakened T-lymphocyte-mediated adaptive immune responses (Niles et al., 2021). CD123 represents a relevant marker for Dendritic Cell activation (Matic S. et al., 2020), meaning that a decreased CD123 expression on the surface of DCs could be an important marker of a delayed interferon-based expression and transmission that is predominantly anti-inflammatory in nature.
A low-dose Type I and Type III Interferon-based nasal spray administered to a patient with COVID-19 could activate the dendritic cells, leading to the subsequent activation of M1 and M2 macrophages. The activation of dendritic cells is mediated by the binding of IFN-I to the IFNAR1/2 receptor on the surface of the dendritic cell. Once activated, dendritic cells are able to produce a range of cytokines and chemokines that can recruit other immune cells to the site of infection, including M1 and M2 macrophages. The activation of M1 macrophages is mediated by the production of pro-inflammatory cytokines, such as Tumour Necrosis Factor (TNF)-α and interleukin (IL)-12. M1 macrophages are involved in the initial immune response to the virus and are able to kill infected cells via apoptosis, necroptosis and pyroptosis, through the production of reactive oxygen and nitrogen species, as well as of diverse caspase enzymes, including caspases 1 and 8. Severe inflammatory responses favour the onset of pyroptosis, which implicates the programmed death of macrophages (Shan C. et al., 2021). The activation of M2 macrophages is also important for the immune response to COVID-19. M2 macrophages are involved in tissue repair and the resolution of inflammation. They produce anti-inflammatory cytokines, such as Interleukin 10 (IL-10) and Transforming Growth Factor (TGF)-β, and help to limit tissue damage caused by the immune response. In COVID-19, M1 and M2 macrophage-mediated immune responses are unbalanced and often polarised, and this leads to an uncontrolled development of inflammation (Mortezaee K. et al., 2022). The activation of dendritic cells and M1 and M2 macrophages is mediated by a variety of surface markers, including CD markers. CD markers are cell surface antigens that are used to identify and characterise different cell types. Different subsets of dendritic cells express different CD markers, which can influence their function and activation. For example, pDCs express CD123 and CD303, which are used to identify and isolate these cells. CD40 is another important CD marker that is expressed on the surface of dendritic cells. CD40 is a co-stimulatory molecule that is involved in the activation of T-lymphocytes and the production of cytokines by dendritic cells. The use of a low-dose interferon I and III-based nasal spray may also influence the expression of CD markers on dendritic cells. For example, one study found that treatment with IFN-I increased the expression of CD80 and CD86 on the surface of dendritic cells, which are important co-stimulatory molecules that are involved in the activation of T-lymphocytes (Alculumbre S. et al., 2019).
Given the exponential nature of the increase of the viral load and the number of infected cells as SARS-CoV-2 infection progresses, early Type I and III IFN responses are less than delayed responses, given that early responses will only implicate the production and manufacture of better understood cytokines i infected cells. Delayed responses will involve the activation of interferon-encoding genes in many more cells, resulting in the production of an amount of IFN that will rather contribute to pathogenesis and aggravation of inflammatory disease, given that the products of ISGs include important pro-inflammatory cytokines, such as CXCL10, CCL2 and CCL5. An increased synthesis rate of pro-inflammatory cytokines is associated with a decreased quality of immune system performance. Likewise, it is important to acknowledge the "double-edged sword"-like nature of Type I (i.e. IFN-α2b, IFN-β1, IFN-δ, IFN-ε, IFN-κ, IFN-ω and IFN-τ) and Type III Interferons (i.e. IFN-λ1, IFN-λ2 and IFN-λ3) and hold fast to the criticality of robust first-line immune responses during SARS-CoV-2 infection. The most important sub-domains of IFNs with regards to building important antiviral and anti-inflammatory signals, alongside shaping important adaptive immunity pathways, represent IFN-α2b, IFN-ε, IFN-ω, IFN-λ1, IFN-λ2 and IFN-λ3, although IFN-β1 also represent glycoproteins with interesting potential immunomodulatory and boosting characteristics.
The timeline of SARS-CoV-2 infection and the onset of COVID-19 disease, using the viral load, symptomatic intensity and antibody count as the principal parameters, was studied in more detail. Generally speaking, COVID-19 symptoms begin when the viral load is approaching its peak and the peak of the intensity in symptomatology for patients experiencing moderate disease occurs when the viral load has already started decreasing. Severe disease generally occurs when the viral peak reaches the level where the disease has not started its clinical manifestations (Muge Cevik et al, 2020), and this could imply the increased abilities of the novel coronavirus to escape host immune sensing can often impair the immune system by resulting in the development of autoantibodies and/or the suppression of the quality and wellbeing of the immune system as a whole. Given that children and young adults generally have first and second-line immune defences that are more robust in nature than old adults, and that the levels of interferon epsilon and interferon omega were found to be significantly higher during SARS-CoV-2 infections in young people than in old adults, and that SARS-CoV-2 was found to affect older people pronouncedly more disproportionately, it is likely that the two interferon sub-domains also play an outstanding role in maintaining a balance between anti- and pro-inflammatory immune factors whilst strongly stimulating the recruitment of NK cells, dendritic cells, as well as of B- and T-lymphocytes (Pierangeli et al., 2022). Nevertheless, the Omicron variant was found to affect younger people much more than the previous major variants, which possibly means that the new variant escaped interferon epsilon and omega signals significantly more. Likewise, the debate on whether interferon alpha-2b plays more relevant immunising and immunisation-adjuvant roles than interferons epsilon and omega remains strong, and further research is needed upon this matter. Moreover, a recently-developed SARS-CoV-2 variant that was named XBB, underwent natural selection and was discovered to significantly outcompete vaccine-induced IgM and IgG immunoglobulins, sparking fresh concerns with regards to the ability of the virus to perform immune evasion even if herd immunity had previously been reached, either by means of mass vaccination or by means of a mass exposure to the virus.
RIG-I and MDA5, which are cytoplasmic pattern recognition receptors, detect pathogen-associated molecular patterns (PAMPs) on the viral genome and phosphorylate the IRF7 and IRF3 dimers, which in turn act as transcriptional factors of Type I IFN -encoding genes. Both MDA5 and RIG-I require activation by dsRNA, but RIG-I requires activation by a triphosphate group, before an interaction with MAVS, which will result in the phosphorylation of the TBK1 and IKK-epsilon dimer. In turn, the IRF7-IRF3 dimer is phosphorylated, which will then activate the type I IFN encoding genes and result in the translation, folding, secretion, as well as the autocrine and paracrine signalling of the immunological messengers (Brisse M. and Ly H., 2019). Once the +ssRNA of the novel coronavirus enters in the host cell, TLR3 and TLR7 become activated as a result of the detection of PAMPs, that are either found on the pathogen surface or synthesised by its genome or are generated during cellular infection. TLR4 activation also occurs, but not inside the host cell, whilst TLR8 is located on endosomes and becomes activated generally due to bacterial infection. Following TLR3/7 activation, MyD88 binds to the pattern recognition receptor and becomes phosphorylated. As a result, three relay proteins are phosphorylated and will act as transcription factors for the synthesis of Type I and Type III interferons; AP1, IRF7 and NF-kB. Following the expression of the interferon-encoding genes in cause, the newly produced interferon proteins will undergo exocytosis and transmission, which will be autocrine and paracrine in nature. Once reaching neighbouring cells, Type I Interferons bind to the IFNAR1/2 receptor, whilst Type III Interferons bind to the IFNLR1/IL10R2 receptor. Following this event, the JAK1 and STAT2 molecules will become phosphorylated, leading to the phosphorylation of STAT1 and STAT2 and their dimerisation. IRF9 then binds to the STAT1-STAT2 phosphorylated dimer to form the STAT1-STAT2-IRF9 trimer before Interferon-Stimulated Genes become activated. Following the signalling cascade, the ISGs will express anti-viral and anti-inflammatory signals that will be playing a critical role in shaping adaptive immune responses. Namely, the products of hundreds of activated ISGs seem to stimulate a desired level of antiviral immune responses by dendritic cells via antigen presentation, as well as helper CD4+ and cytotoxic CD8+ T-lymphocytes via supporting plasma cells in the production of qualitative antibodies and inducing the lysis of infected cells respectively.
Some of the ISG products, like IFITM3, play major flexible roles in linking first and second-line immune responses to the adaptive immune system. Likewise, a significant impairment of Type I and Type III Interferon production and signalling result in severe implications for the adaptive immune response. The viral non-structural protein 1 (NS1 or NSP1) represents an important example of a viral component that is a result of an evolutionary response to impair first-line immune responses. Namely, NSP1 has been shown to cleave and lyse the mRNA encoding Type I Interferons, alongside NSP2 (Xu et al., 2022). Interestingly, human host cells developed anti-viral evolutionary responses to include the ability of such viruses to inhibit first-line immune responses. The 2',5' oligoadenylate synthetase proteins 1,2 and 3, protein kinase R, nuclear factor 90 and interferon-stimulated gene product 15 represent proteins that restrict the ability of viruses like Influenza A and SARS-CoV-2 to replicate, and yet nsp1 was found to inhibit the activity of such proteins, alongside cleaving and lysing the mRNA encoding Type I Interferons. The binding capabilities of the viral RNA specifying NSP1 inhibits the activities of the 2',5' oligoadenylate synthetase proteins and prevents RNaseL activation, leading to an inhibited process of viral RNA degradation. The viral RNA-inhibiting activities of protein kinase R and nuclear factor 90 shows how human host cells and the virus have co-evolved, and NSP1 has been helping the virus escape such proteins. Nuclear factor 90 is possibly not produced by Interferon-Stimulated Genes, which indicates that the evolutionary conflict between first-line immune defences and respiratory viruses of such nature has been more generalised than previously thought. Furthermore, interferon-stimulated gene product 15, which is produced by one of the most expressed interferon-stimulated genes, has been shown to target non-structural protein 1, as Isg15-deficient mice were shown to be more susceptible to Influenza A infection and that the pathogenic protein was recently displayed as a target of ISGylation. Moreover, Influenza A viruses also developed PB1-F2 and PA-X proteins to bypass innate immune responses, by inhibiting the process of viral RNA sensing and significantly downregulating the Type I and Type III Interferon-induced signalling cascades and the apoptotic process of infected cells (McKellar et al., 2021).
TLR-3, TLR-7, TLR-8 and TLR-9, which are endosomal Pattern Recognition Receptors, as well as TLR-4, which is a cytoplasmic Pattern Recognition Receptor, detect Pathogen-Associated Molecular Patterns (PAMPs) on the viral genome and phosphorylate the IRF3 and IRF7 dimers. The activation of TLR-3 and TLR-4 results in the phosphorylation of TRIFF and then the IRF3 dimer, whilst the activation of TLR-7, TLR-8 and TLR-9 causes the phosphorylation of MyD88 and then the IRF7 dimer. Unlike the case of TLR-3, activating TLR-4 will also phosphorylate MyD88, TIRAP and TRAM before the activation of the IRF3 dimer. The final outcome is the same; the expression of Type I Interferon-encoding genes and the autocrine and paracrine signalling of the immunological messengers. (Cervantes J. L. and Weinerman B., 2012). Type I Interferons have recently been found not to recruit NK cells directly, but through the activation of inflammatory chemokines and monocytes (CCL2, CCL5, CXCL10 and IMMs) (Lee A. J. et al., 2019). Interferon-stimulated genes produce various inflammatory chemokines, such as CCL2 and CXCL10, which are responsible with the activation of inflammatory monocytes and dendritic cells, as well as with the recruitment of natural killer cells, which in turn activate macrophages and interferon-gamma, which belongs to the second domain of interferons, and induce the lysis of infected cells. Although interferon-gamma was found to have powerful preventive effects against important forms of respiratory, oncological and neurodegenerative diseases if produced, exocytosed and transmitted in moderation (Aiman et al., 2022) and it was also found to share a similar bi-directional nature with Type I and III Interferons with regards to their effect upon pathogen integrity and immune competency respectively, according to the extent of its activation, it is the first and the third interferon domains that play the most foundational roles in shaping and balancing much of the immune response. The activation of antigen-presenting cells and the recruitment of natural killer cells will ultimately shape the processes of CD4+ and CD8+ T-lymphocyte recruitment, as well as the quality of antibody production and specification via the process of V(D)J antibody gene rearrangement in maturing B-Lymphocytes. A dysregulated synthesis rate of Type I and III Interferons result in an increased CXCL10 signalling extent, which in turn will inhibit the proliferation of myeloid progenitor cells (Khalil et al., 2021) and increase the level of p38-mediated primary T-lymphocyte apoptosis (Sidahmed et al., 2012). As a result, the risks for the development of deficiencies in myeloid cell (i.e. neutrophil and dendritic cell) and lymphoid cell (helper CD4+ and cytotoxic CD8+ T-lymphocyte) counts, thereby increasing the probability of significant adaptive immune consequences.
CCL2 and CXCL2 were found to be capable of clearing tissues from the SARS-CoV viral load without the help of helper- and cytotoxic-T-lymphocytes, as well as of neutralising antibodies, twelve days after the moment of first-cell-infection, and this finding indicates the high importance of activating antiviral innate immune responses by recruiting neutrophils, monocytes and macrophages toward the infected tissues. It was also shown that hyper-activated interleukin-6 and interferon-gamma-related pathways were associated with a higher severity of COVID-19 (Lagunas-Rangel et al., 2020), potentially meaning that delayed Type I and III Interferon responses are associated with higher transmission rates of IFN gamma and IL-6, as a significantly higher number of infected cells would almost simultaneously produce Type I and III interferons and likewise, their number would be much higher than in the cases when interferons are produced and undergo transmission early. The fact that the levels of inflammatory chemokines like CCL3, CCL5, CCL20 and CXCL10 were considerably higher than the levels of inflammatory chemokines secreted by CD14+CD16+ inflammatory chemokines in COVID-19 patients with developed acute respiratory distress syndrome (ARDS), unlike in the case of non-COVID-19 related viral and bacterial infections that resulted in the development of ARDS, when the chemokine levels were similar, represents a significant sign that the principle immunological problem caused by the novel coronavirus is not only related to, but based upon a disrupted timing and extent of Type I and III Interferon system activation. Moreover, in the case of the SARS epidemic, the virus is also capable of inhibiting Type I and III Interferon expression and once interferon-stimulated genes are finally expressed, inflammatory chemokines, including CCL7 and CCL8 as well, are released and further contribute to the onset of the disease, which further suggests how several respiratory viruses have co-evolved with the interferon system. Although SARS-CoV and MERS-CoV display similar chemokine profiles, performed comparative studies showed that MERS-CoV infection results in higher activation rates of the CXCL10 inflammatory chemokine, and this may be an important reason why the systemic inflammatory extent and death rate of MERS are higher. CXCL10, CXCL8 and CCL2 represent potentially important markers of SARS, MERS and SARS-CoV-2 infection and onset of infectious disease, and the activation rate of CXCL10 is particularly analysed in COVID-19 patients. Such a chemokine binds to the CXCR3 receptor to become activated and stimulate the recruitment of natural killer cells, T-helper cells 1, cytotoxic CD8+ T-cells, as well as Th1-related immune responses, and its concentration is directly proportional with the severity of the infectious disease. It was found to be positively-regulated during early stages of the SARS-CoV-2 infection, which further indicates that its extent of synthesis is dependent on the timing of Type I and III Interferon synthesis, as well as autocrine and paracrine signalling.
Interferon I and III production and signalling rates were much lower in pHAE cells infected with SARS-CoV-2 than in pHAE cells infected with the H1N1 variant of Influenza A (Abigail V. et al, 2021), which suggests that the novel coronavirus is significantly more capable of escaping detection by PRRs, of capping the 5’ end of the +ssRNA viral genome and of lysing interferon and ISG proteins than the Influenza A +ssRNA virus. The option of using a UV-attenuated SARS-CoV-2 specimen with a deletion in the genes encoding the non-structural proteins 1 and 16 could have been the best vaccination candidate, had the spike protein not acted as a superantigen, entered the lymphatic system and caused damage to the endothelium, crossing the endothelial barrier and entering the bloodstream. We believe that the main problem is the great level of toxicity the spike protein has been displaying through severe cases of COVID-19, often resulting in the formation of sub-optimal antibodies and significant autoimmune implications. Furthermore, during the SARS epidemic, researchers developed the TP29 small peptide to separate the activator nsp10 from the effector nsp16 to prevent a large extent of 5' viral +ssRNA capping, as well as the oral methioninase enzyme to digest the S-Adenosyl-L-Methionine pocket of the virus in order to expose the concerned non-structural proteins to lytic factors and prevent the process of 5' viral mRNA capping. Almost two decades later, it was discovered that the novel coronavirus produces the same non-structural proteins and S-Adenosyl-L-Methionine pocket to camouflage itself and prevent the activation of the host cell's Pattern Recognition Receptors. Likewise, the two early approaches could show significant efficacy and bring insignificant financial demands in the pharmaceutical market, and researchers showed that oral methoninase displays efficacy against COVID-19 as well (Hoffman et al., 2020). Moreover, there seems to be another method to evolutionarily combat the pathogen, by manually stimulating immunisation through the development of IgM super-antibodies to directly remove the viral camouflage, by tackling the non-structural proteins 1 and 16 inside the infected cells.
The immune system could be trained in this way as well to build a better interferon-based defence against viruses that gained an evolutionary advantage of suppressing it. The problems with such an approach are the massive financial demands and a precision of the intervention that might be too elevated, which means it could overall bring an increased risk of adverse reactions. Boosting the mucosal immunity, on the other hand, represents an approach that has been tested numerous times, and many of the performed tests indicated outstanding positive results, despite a number of concerns of inefficacy and high financial demands from a number of critics. Concerns include a possible relatively weak connection between the development of qualitative IgA antibodies in the mucosal immune system and the development of qualitative IgG antibodies in the systemic immune system due to a high complexity of the local immunity. However, tests implicating the stimulation of IgA synthesis have shown outstanding prophylactic efficacy, with very few clinical trial participants experiencing infection or re-infection in the future. The COVID-19 pandemic was not exempt in this case, as attempts of intranasal prophylaxis and immunisation were associated with the development of long-term immune memory against the virus and the spike protein. Results have strongly indicated the importance of developing IgA-mediated mucosal immunity in the prevention of moderate and severe disease. It is suggested that the IgG1 to IgA1 antibody secretion ratio and the extent of neutrophil activation are proportional with disease severity (LaSalle T. J., 2021), meaning that a delayed, systemic Type I and III Interferon signalling and a widespread secretion of IgA antibodies may likely become markers of severe COVID-19 disease. The duration of the pre-clinical stage of COVID-19 is very likely also proportional with the probability that the COVID-19 disease will be severe. Moreover, the fact that oral methioninase was shown to have significant efficacy in prophylaxis and early treatment further indicates the high potential of mucosal immunity in preventing the onset of severe infectious disease. One early therapeutic approach implicated the administration of inhalable IgA immunoglobulins that had previously been exposed to the spike protein of the Omicron variant, into the nasal cavity of K18-ACE2 transgenic members of the Mus. musculus species that were infected with the Omicron variant. The approach was shown to be more efficacious than an IgG Fc-based prophylaxis and treatment, and it used IgA antibodies that had been synthesised and secreted in Pichia pastoris for cost-effectiveness purposes (Qi Li et al., 2022).
Another method to stimulate Interferon I and III-based immune responses could involve the gathering of proteins from diverse plants that stimulate a more robust production and exocytosis of Type I and possibly of Type III Interferons, find the right quantity of each of them in order to bring robust effects, since plant medicine often brings slow effects and dosages usually require to be consistent over a longer period of time, and such methods represent applications of Translational Medicine. The ingredients of a natural compound based on plant medicine that stimulates interferon responses are as follow: Silybum marianum – a hydroalcoholic extract with 80% silymarin from fruit (200 mg); Astragalus membranaceus – 10: a hydroalcoholic extract from the roots (150 mg); Schisandra chinensis – a hydroalcoholic extract with 2% schizandra from fruit (150 mg); Agaricus blazei – a hydroalcoholic extract with 20% polysaccharides (100 mg); Ganoderma lucidum – a hydroalcoholic extract with 20% polysaccharides (100 mg); Morinda citrifolia – a hydroalcoholic extract with 40% polysaccharides from fruit (75 mg); Aloe vera – 20: a hydroalcoholic extract from the aerial parts (50 mg); Foeniculum vulgare (fennel essential oil) (0.11 mg); filler: microcrystalline cellulose; anti-caking agents: colloidal silicon dioxide and vegetable magnesium stearate (Dacia Plant, 2021). We are aware some ingredients are preservatives that probably have controversial traits, and there is no claim that this particular compound is certainly perfect. Perhaps, such preservatives could be replaced with alternative compounds if the situation imposes it. If required, we could create a medical drug containing a mixture of at least some of these plant-derived interferon-stimulating proteins and interferon-alpha, beta, delta, epsilon, kappa, omega and lambda, and we believe that such an approach would more likely be needed if more severe SARS-CoV-2 and Influenza A variants emerged and spread around the world. Perhaps, such an approach could be combined with approaches of immune system-based prophylactic and early therapeutic vaccines involving human recombinant plasmacytoid dendritic cells as factories of interferon glycoproteins and/or natural and adaptive lymphocytes treated themselves with a fairly low dose of interferon glycoproteins beforehand, particularly in the context of preventing or attenuating the effects of disease in the early stages of HIV-induced AIDS (Carp T., 2024).
With regards to the evolutionary advantage of the virus to suppress key mechanisms of natural immunity, a traditional vaccination approach using a pathogenic fragment might not help significantly on a long-term basis because the pathogen is highly polymorphic and, most importantly, because it will not directly support the development of a counter-evolutionary response to outcompete the ability of the virus to suppress such natural immunity mechanisms. This might represent a challenge even for dendritic cell-based vaccines, not because such antigen-presenting cells will bring accurate forms of antigen to lymphocytes and train them to target the most relevant variants, but because of the same gap created by the virus in key parts of the innate immunity (Saadeldin et al, 2021). As a result, the virus will use such gaps to adapt and become more capable of suppressing natural immunity. A sign that such events are already happening is the higher capability of the BA.1 variant of SARS-CoV-2 to suppress Interferon I responses, which explains why more children develop complications and become hospitalised (Lyudmila Shalamova et al, 2022). Moreover, the BA.2 variant of SARS-CoV-2 has recently been found to have an even higher ability to suppress such interferon-based responses, particularly in children and teenagers. A full and long-lasting immunisation against SARS-CoV-2 could only apply if the entire immune system is continuously covered. The dosage could involve a puff of the spray per day in each nostril, and the duration of the prophylactic or early therapeutic session could last from two weeks to a month. Likewise, we would consider this vaccine candidate of a long-term kind. Given that the classical vaccine, which involves sharpening immunity purely from an adaptive immunity perspective, does not seem to address the problem of viral camouflaging using its non-structural proteins, whose impact affects the innate immune system, we believe that an interferon I and III-based approach will make a greater difference and, possibly, even evolutionarily outcompete the virus and its skills of self-camouflage. This clinical approach could also include the administration of a TP29 small peptide, which separates the non-structural protein 16 from the non-structural protein 10 to deactivate it (Wang et al, 2015), and oral recombinant methioninase to disintegrate the S-Adenosyl-L-Methionine pocket of the virus and decrease the activity of RNA-dependent RNA Polymerase, which catalyses the replication of the virus (Hoffman et al, 2020).
A more specific timeline of Interferon-alpha and Interferon-beta synthesis in the early stages of Herpes Simplex Virus 2 (HSV-2) infection has been studied. There is a considerable probability that interferon-beta will first be produced to signal to the immune system that the cell has just been infected, around 12 hours after the first HSV-2 virus entered its host cell. Interferon-alpha will be produced around 40 hours after the synthesis of interferon-beta and just over 48 hours after the first host cell becomes infected. Interferon-beta can be produced again at the time when the production of interferon-alpha commences (Amanda J. Lee and Ali A. Ashkar, 2018). This Type I Interferon synthesis timeline may be similar in the case of a SARS-CoV-2 infection. Given that it usually takes significantly more time than 40 hours for the coronavirus to infect more cells and cause symptoms, a sole interferon-alpha-based prophylaxis and therapy will probably not be substantially different from a therapy consisting of a combination with interferon-beta. Nevertheless, the synthesis of interferon-gamma depends on the extent of the Natural Killer (NK) Cell recruitment, which itself depends, to a certain degree, on the extent of interferon-beta synthesis. Importantly, NK Cells also form and develop an immune memory against pathogens and differentiate into memory cells, despite being part of the innate immune system (Lena Müller et al, 2017). One clinical study involving the administration of a nasal spray containing interferon-beta and serine residues to check the prophylactic effects against rhinovirus showed a higher degree of effectiveness than interferon-alpha-2b based nasal sprays, and likewise, trialling a nasal spray using a combination of interferon-alpha and interferon-beta could be worthwhile (Sperber et al, 1988).
The results of a pharmacokinetic in-vitro study displayed a high effectiveness of recombinant interferon-alpha-2b therapy for the respiratory syncytial virus (RSV), parainfluenza virus (HPIV) and coronavirus strains causing common cold (i.e. HCoV-OC43 and HCoV-229E). The therapy had particularly immune-sharpening effects for the parainfluenza virus and the common cold coronavirus infections, and results were better than the drug control section for certain Influenza A variants as well. Overall, the in-vitro experiment involving interferon-alpha residues showed broad antiviral effects, a high therapeutic index and a low toxicity (Hui Qiang Wang et al, 2014). Moreover, a preliminary in-vivo study was performed in Beijing, China, in order to verify whether a low-dose interferon-alpha-2b-based nasal spray is effective in preventing the rubella and measles infectious diseases, and the results showed that such a nasal spray can be used for preventing these diseases as well. In this context, a low dosage is in nanograms (ng) and a high dosage is in milligrams (mg), and only a low dosage of interferon-alpha is generally required to prevent and treat infectious diseases ranging from the flu and COVID-19, to the Acquired Immunodeficiency Syndrome (Joseph Cummings, 2020). An in-vitro research study of human cell cultures, performed at the University of Texas in 2020 as part of the Journal of Antiviral Resistance, displayed clear evidence that SARS-CoV-2 is highly influenced by the presence of interferon-alpha and interferon-beta, and has indicated a considerably possible link between the proportion of 80% of infected people who develop mild symptoms and the fact that in many cases, infected cells manage to robustly send Interferon I signals and stimulate immune responses rapidly and proportionally (E. Mantlo et al, 2020).
In June 2020, the University of West Australia published an article stating that Type I Interferon nasal and oral sprays, as well as lozenges, may play an important role in preventing COVID-19. They referenced a clinical trial that included 2,944 participants from the infectious diseases’ hospital departments in China involved the prophylactic administration of a low-dose of recombinant interferon-alpha-2b as a way of protecting frontline healthcare workers in areas of COVID-19 outbreaks, and the results were promising. No participant developed pneumonia afterward (M. Zhongji et al, 2021). Moreover, about 750 army soldiers in China participated in the experimental group of a clinical trial in 2005 involving the administration of a low dose of interferon-alpha, and the results were successful on a similar scale. There was no significant side effect reported in neither of the trials, and I believe this was so because the dosage was accurate and used within the right context; the quantity of the administered interferon-alpha matched the quantity of the deficient Type I Interferons (Gao et al, 2010). Furthermore, during the Hong Kong influenza outbreak in January and February 1969, there was a clinical trial conducted by Dr. V. Soloviev in the former USSR, who tested the efficacy of a nasal spray based on a small concentration of human leukocyte-produced interferons on a treatment group of 14,000 participants, and the calculated efficacy rate was between 56.3% and 69.2%, with variations according to the age group. Around 128 units of interferons were used in the nasal sprays, the treatment session lasted for five days and it was determined that the interferon-based treatment significantly reduced the number of influenza cases (p < 0.01). Furthermore, it had been suggested that the Hong Kong strains of influenza virus could have been divided into interferon-positive and interferon-negative variants. It was reported that the approach was harmless and that it should be applied in cases of threats of immediate infections, which means it should be regarded as an emergency prophylaxis (V D Solov’ev, 1969).
With regards to the exact mechanisms that a recombinant interferon alpha-2b-based therapy resulted in, a clinical trial indicated the ability of the human interferon alpha to decrease the duration of viral transmission and consequently, to reduce the markers of acute inflammation, such as Interleukin-6 (IL-6) and C-Reactive Protein (CRP). Namely, by testing human interferon alpha in COVID-19 patients, it was determined that a longer viral transmission from the host can be associated with stronger markers of acute inflammation and likewise, that an impaired interferon-alpha response causes the viral infection to become a more common factor of pathogenesis and severe illness. The results encouraged scientists to recommend interferon-alpha-2b therapy as a method to combat the pandemic (Q. Zhong et al, 2020). Also, interferon alpha-2b therapy also resulted in a shorter duration of time the viral load was detected in the upper respiratory tract (Qiong Zhou et al, 2020). Another clinical trial in Iran implicated an injectable dose of interferon-alpha of 12 million IU, and the study had encouraging results, despite the exceptionally higher dosage used. Specifically, the mortality rate from COVID-19 in the cohort was reduced by more than 50%, and early interventions sharply reduced the death rate (E. D. Monfared et al, 2020). We share the thoughts of Dr. Joseph Cummings that a low-dose interferon-alpha therapy would likely have further decreased, not only the mortality rate of the disease, but also the ability of the virus to increase its load and the probability of morbidity following infection in that cohort. One important aspect to add is that clarithromycin-based early therapy against COVID-19, which showed a high rate of efficacy and very low risks of adverse events, was associated with a decreased circulation of the C-reactive protein, tumour necrosis factor-alpha and interleukin-6, as well as an increased rate of Th1 and Th2 mononuclear responses and a clear restriction against the increase of the viral load. The effects of normal Interferon I signalling rates and early therapy result in an almost identical outcome as clarithromycin-based early therapy (Konstantinos Tsiakos et al, 2021).
The majority of the immune cells produce Type I Interferons, meaning that the glycoprotein cytokine is widely bioavailable (De Maeyer, E. et al., 1998). Plasmacytoid dendritic cells (pDCs) produce Type I Interferons most commonly, and they are also known as antigen presenting cells. Given that SARS-CoV-2 is known to decrease the ability of such cells to capture and present pathogenic antigens, that means the viral inhibition of Type I Interferon production automatically decreases the ability of pDCs to present captured viral antigens. As a result of the interferon inhibition, natural killer cells are also relatively absent. The absence of natural killer cells and interferon-dependent killer dendritic cells (IKDCs) leads to a poor production of Type I Interferons (mainly by plasmacytoid dendritic cells, or pDCs). Plasmacytoid dendritic cells are also known as conventional dendritic cells (cDCs) (David Vremec et al., 2007). In other words, a robust production of Type I Interferons during viral infection plays a central role in stimulating NK cell recruitment and antigenic presentation by pDCs, which produce the interferons in the first place. Viral apoptotic inhibition facilitates viral replication and spread to more kinds of tissues, particularly during the first few days of infection. Overall, this results in a lower recruitment rate of B and T-Lymphocytes, which in turn results in the development of poorer natural and adaptive antibodies. Furthermore, this facilitation of viral replication and spread, on top of the viral inhibition of first-line immune responses, further contributes to the problem of a compromised collective immune privilege against novel infectious viruses. Also, it is rather possible that this impairment of the immune system to effectively reduce the replication rate of the virus and tackle its pathogenicity in the end is an important factor of the dilemma of induced autoimmune diseases (i.e. long COVID post SARS-CoV-2 infection) as well. Such an induction of autoimmune disease is voluminously favoured by a disrupted anti-inflammatory to pro-inflammatory mediator ratio, as the probability of induced autoimmune disease is proportional to the intensity of the antiviral and pro-inflammatory immune responses.
With regards to the relationship between immunocompetency and the severity of COVID-19; it is essential to note that immunosuppression does not always lead to a severe outcome of the disease due to the complexity of the nature of the immune responses that are resulted from the viral infection. Namely, the clear innate immune suppression is the number one cause for a disrupted set of adaptive immune responses, which in turn can lead to the onset of severe symptoms and post-COVID autoimmune diseases. Immunosuppressed patients in this case often develop milder symptoms because the intensity of their immune responses is considerably lower. As a result, the risk of immune-derived tissue damage is actually lower, and this also explains the heated debate about whether certain immunosuppressants are actually effective in reducing the intensity of COVID-19 symptoms. Therefore, the outstanding level of the initial induced first-line immune suppression by SARS-CoV-2 brings higher risks of the onset of disruptive pro-inflammatory immune responses, meaning that the extent of viral self-camouflage, suppression of interferon production, inhibition of antigenic presentation and temporary suspension of induced host cell apoptosis is directly or even exponentially proportional with the intensity of the induced symptomatologic immune responses afterward.
An Interferon I and III-based response should be early and prompt, otherwise infected people can develop serious forms of the disease. The ability of the virus to camouflage itself and limit the amount of synthesised Type I and Type III Interferons likely has caused millions of unnecessary deaths and put further tens of millions of lives at risk all around the world since the first outbreak took place. There are three scenarios with regards to the timing and extent of Type I Interferon synthesis. The first scenario implicates the prompt synthesis and secretion of the right amount of Interferon I, and this leads to the development of a strong and intelligent response, to the development of mild or no symptoms and a quick recovery. The second situation implicates a delayed Interferon I response, which often can become exaggerated as well because of the increased viral load, and this scenario is associated with moderate to severe clinical symptoms, the development of pneumonia, as well as hospital admission, which sometimes can lead to a further admission into the Intensive Care Unit. The third situation implicates a completely absent Type I and Type III Interferon response, which is followed by a severe form of the disease, admission into the Intensive Care Unit, intubation, a systemic inflammation caused by a high viral load and the spread of the virus from the lungs to other important organs through the blood and often death. A delayed administration of prepared interferons not only does not improve symptoms or help the patient recover more quickly, but can worsen the disease, since because this delay allows a sharper increase of the viral load, this intervention would only contribute to an over-sensitised and exaggerated interferon expression. In turn, there could also be a number of instances when a proper interferon-gamma production and signalling rate will be delayed, and when the rate of response will finally increase, it could become exaggerated as well due to the higher viral load, which will lead to a higher immunological demand. As a result, a delayed increase of interferon-gamma response could also contribute to a worse disease form. In other words, timing is critical for a virus with a high replicative rate. Furthermore, approximately 10% of the cases when there is a delayed and exaggerated Interferon I-based response, patients develop autoantibodies and consequently, diverse forms of autoimmunity.
An Interferon-based therapeutic approach generally requires to be taken even before an infected person requires hospitalisation. The fact that the approach did help save lives brings hope that the approach is useful even in slightly more advanced stages of the disease. One important aspect to mention is that severe symptoms would likely represent a marker for a delayed and exaggerated interferon response, which means that only infected people with milder symptoms should be eligible for interferon-based therapies. In the Cuban study of interferon efficacy for COVID-19 hospital patients, 95.1% of the patients in the cohort who received the interferon-based therapy were discharged, compared to 21% of those who did not receive this therapy. The case-fatality ratio of the patients in the placebo group was 32.1%, compared to the 0.9% case-fatality ratio of the patients in the treatment group, and the case-fatality ratio for the patients with severe disease who did not the therapy (48.6%) was more than double the number of patients with severe disease who did (21.9%) (Ricardo Pereda et al, 2020). Likewise, it is possible that, the earlier and less severe the stage of the disease is, the more effective interferon-based therapy is, and it is likely that a prophylactic approach will almost always prevent the onset of the disease. Such events show the power of specific innate immune mechanisms in front of a virus that can cause significant morbidity and an unprecedented number of deaths.
Several clinical studies suggested that lozenges and nasal sprays containing a low dose of Type I Interferons show prophylactic and even early therapeutic efficacy against HIV-induced AIDS as well. For example, one interim clinical trial involved the recruitment of forty adults tested positive for the HIV-1 infection and they were offered daily lozenges containing an amount of IFN-alpha ranging from 75 to 600 IU. Eighteen other patients infected with HIV-1 were recruited but were not offered the interferon lozenges. From the total number of patients, eighteen patients received long-term treatment and five of them died. Among those patients, two of them had suffered from Kaposi’s Sarcoma and two others died because of HIV-1 related factors (Babiuch et al., 1993). Another clinical study involved the administration of Pegylated Interferon-alpha-based treatment (Peg-Intron A), in combination with antiretroviral treatment that involved the administration of Highly-Active Antiretroviral Therapeutics (HAART), Nelfinavir, Stavudine and Didanosine. From the total number of 168 injections with the Pegylated Interferons, only six were not administered at a maximum dose due to safety concerns. The IFN therapy was found to be tolerated well, to significantly restrict HIV replication , protein assembly, packaging and distribution, as well as to ultimately induce a robust reduction of the viral road during the early stages of infection, and to visibly prolong the lifespan of the infected CD4+ T-Lymphocytes (Emilie et al., 2001).
Furthermore, a recent study published in January 2022 indicates that the Omicron variant of SARS-CoV-2 is more capable of silencing pathways involved in interferon-alpha and interferon-beta expression and, as a result, there may be increased risks of delayed or impaired antiviral and anti-inflammatory responses, leading to a higher susceptibility to the development of chaotic adaptive immune responses. This phenomenon is tricky because the Omicron variant has been proven to generally be ten-fold less morbid and deadly than the Delta variant (Lyudmila et al, 2022). Given that children generally have a stronger innate immune system and likewise, their interferon I signalling is generally robust and in greater amounts, the higher ability of the Omicron variant to delay and impair Interferon I-based responses is the reason why more children are affected by the Omicron variant-induced disease than the Delta variant-induced disease. Likewise, we believe that there should be a greater awareness of the need for sharpening interferon I-based immunity, given that increases in the Tumour Necrosis Factor and IL-6 production rates, as well as NF-kB-based inflammatory processes were not enough to significantly restrict the increase of the viral load and the inflammatory responses (Lee et al, 2020). We hypothesise that around 125-150 IU of interferon-beta may be combined with around 175-200 IU of interferon-alpha residues and form a combined prophylaxis and therapy, which could be even more effective against COVID-19 and flu diseases induced by Influenza A variants of concern.
Due to the fact that interferon-gamma also plays an important role in sharpening antiviral immunity and restricting unnecessary inflammation, we believe that combining interferon-alpha with interferon-beta and interferon-gamma and forming a triple-interferon-based prophylaxis and therapy could also be a suitable candidate, although we believe that a combination of interferon-alpha, interferon-beta and interferon-lambda would be enough to reach the maximum potential of the interferon candidate. The candidate would have a maximum total interferon amount of 500 IU (i.e. 200 IU of IFN-alpha and 300 IU of IFN-beta). Given that the peak of the viral load is usually reached around two days after the symptomatic onset, a nasal recombinant interferon alpha-2b-based spray could be most suitable for administration up to two days after the first symptoms of COVID-19 have occurred for the desired therapeutic outcome. In other words, scientists could bring an emphasis upon this potential vaccine candidate in stages of local outbreaks and in people who have mild or no symptoms and have just received a positive polymerase chain reaction or rapid antigen test result. The product could be administered either just via the nasopharynx or both via the nasopharynx and the oropharynx to bring about a more extended mucosal immunity activation. The candidate should not be recommended to anyone with COVID-19 that has received a positive antibody test result, since the viral peak had most likely already been reached by then. The right kind of Interferon I therapy involves a low-dose administration before the viral peak has been reached, and according to the timeline presented above, the viral peak is reached around 48-55 hours after the onset of the first symptoms. Likewise, we believe that the dosage should take place only for prophylactic and early therapeutic purposes, and this includes people who do not display symptoms, are tested negative and happen to be in an area of outbreak and infected people who have just received a positive real-time polymerase chain reaction or rapid antigen test, and developed the first symptoms. Infected people who receive a positive antibody test should not receive any dosage of interferons, since the stage where antibodies are formed and more widely present in their system indicates that the viral peak has already been reached, and the patients would only risk aggravating their disease. A higher quantity of administered Interferon I can raise the risks of developing unwanted adverse reactions and sometimes autoimmune responses as well.
Type I Interferon synthesis and exocytosis from tumour cells represent an essential step in a correct signalling of tumour cells to immune components implicated in tackling cancerous formations (Ernest C. Burden, 2019). A disrupted balance between Interferon-alpha, which is an anti-inflammatory cytokine, and Tumour Necrosis Factor-beta, which is a proinflammatory cytokine, is highly associated with severe COVID-19, as in the case where there is a significantly higher number of TNF-beta residues than IFN-alpha residues, there will be much weaker restrictions against inflammation. Moreover, a lower amount of IFN-alpha will lead to weaker restrictions against viral replication and spread. These two issues combined can make a huge negative difference with regards to disease outcome. Moreover, an excess of Tumour-Necrosis Factor-beta had a dysregulatory effect upon many Natural Killer Cells, which as a result failed to attach and induce the apoptosis of cells infected with the virus. With the impairment of interferon-alpha synthesis, there will already be a disrupted interferon-alpha to tumour necrosis factor-beta ratio, which will make the immune system more prone to excessive inflammatory responses and impeded antiviral responses. Other clinical studies implicated an injectable amount of recombinant interferons of 3 to 6 million International Units for severe diseases, like malignant tumours and polycythaemia and, although the risks there were higher, the majority of the participants experienced recovery from their disorders and likewise, the clinical trials were deemed successful. People were often put into two main categories in order to differentiate dosage based on the underlying health condition, since a much higher quantity of interferon-alpha poses a higher risk of moderate and disruptive adverse reactions. Given that the majority of the participants did not experience visible adverse events and experienced a significant improvement of the clinical display of their disorder, we believe that the administration of recombinant interferon-alpha-2b represents a likely pivotal approach in immunology, microbiology and pathology restriction, and will potentially result in a scientific progress that would not have a precedent.
There is a circulating theory that the Fukushima disaster in 2011 has already started to cause forms of Acute Radiation Syndrome (ARS) around the world. The theory is based on the evidence that the Fukushima disaster is worse than the Chernobyl one, that it will be affecting the Earth for potentially 100 years to have significant negative social impacts upon animals and the fauna. It was mentioned numerous times that certain effects of the disaster will be irreversible. Likewise, it is very possible that some kind of radiation-induced disease or set of diseases will be circulating around the world, maybe even for decades. Symptoms of Acute Radiation Syndrome include inflammation of upper and lower respiratory tracts, shortness of breath, fever, loss of smell and taste happening regardless of whether the nose becomes blocked or not, headaches and dizziness. More severe cases involve the onset of pneumonia, cytokine storm and multi-systemic inflammation. In other words, the symptomatology is often very similar to the one of COVID-19, and we are wondering whether cases of Acute Radiation Syndrome perhaps infiltrated the SARS-CoV-2 pandemic (Rios et al, 2021), given that a number of patients who developed COVID-19 around the world experienced a more significant loss of hair as well, for example. The trickiest commonality between COVID-19 and Acute Radiation Syndrome is that the lung injury morbidity caused by both disorders can be significantly mitigated using inhibitors of the Angiotensin Converting Enzyme (ACE) (L. Cerezo and M. M. i Garau, 2012). Likewise, because of the visible commonalities between COVID-19 and ARS, we believe that certain methods of treatment developed for ARS could apply for COVID-19, and vice versa. Given the evidence that the future does not look bright with regards to respiratory illnesses and the projected general state of health of the planet, we are urging scientists and clinical researchers to create a nasal spray consisting of either interferon-alpha and interferon-beta, or probably even a combination of interferon-alpha, interferon-beta and interferon-lambda, and determine whether the immune-stimulating agents can be regarded as a vaccine for not only the current pandemic, but also for other serious respiratory illnesses that are significantly probably to emerge over the next decades. With regards to the lost sense of smell and taste as a result of radiation poisoning, we could perhaps apply important principles by Nikola Tesla into electricity-based therapies to repair the damaged ophthalmological nerves, and if SARS-CoV-2 also damages such nerves, then this therapeutic approach could be applied for COVID-19 as well. Furthermore, all pathogenic agents seem to have their own sound frequency, and perhaps there could be the development of a frequency-based therapy to target and neutralise the virus during diverse stages of infection and tissular spread. An interesting intervention for intubated COVID-19 could be the transport of Ultraviolet A (Ali Rezaie et al, 2021) and Ultraviolet C rays through the tracheal tube by means of reduction of the viral load in the trachea and the bronchi, and a general mitigation of the hyper-inflammatory processes (Stanislaw P Stawicki, 2020).
SARS-CoV-2 represents a pathogenic agent with a high degree of complexity with regards to developed evolution against human immunity. Both viral infection (Zhang et al., 2020) and the administration of the spike protein-encoding mRNA vaccines (Aldén M. et al., 2022) led to the activation of a number of LINE-1 Reverse Transcriptases and the reverse transcription and integration of around 1% of the viral genome and of the spike protein-encoding mRNA into the DNA of diverse host cells, such as hepatocytes. This phenomenon is rather novel for non-retroviral RNA particle-like pathogens. Furthermore, the novel coronavirus produces diverse non-structural proteins to inhibit first-line immune signals, it inhibits the activity of specific chaperones and consequently, the development of some important antibodies, and it produces the spike glycoprotein to further stimulate an already exaggerated, late interferon-based signalling and activation of Interferon-Stimulated Genes. The spike glycoprotein often showed problematic mechanisms even when it was used for immuno-stimulatory purposes, which displays the degree of severity in the evolutionary and immunological matter as such. This further indicates the super-antigenic nature of the spike protein, and likewise, this also highlights the ability of SARS-CoV-2 to eventually mutate into variant forms that escape the majority of vaccine-induced antibodies. We are possibly situated at a dead-end with regards to the current inability of immunology to solve the puzzle of preventing the onset of diseases caused by pathogens with mechanisms of immune evasion, upon which viral mutation and polymorphism is usually dependent upon. A continuation of viral mutations based upon interferon I and III-suppressive models could eventually result in the natural selection of pathogenic super-powers. Overall, the novel coronavirus has acquired evolutionary abilities to suppress all essential areas of human immunity. Likewise, the scientific community is being under an increasing pressure to shift the overall approach in order to include the direct stimulation of first-line immune responses whilst developing prophylactic, therapeutic and immunising agents to tackle epidemics of concern. The matter represents direct evidence of a degree of error and failure in past immunological research to tackle infectious pathogens of concern and treat significant forms and stages of induced diseases.
Given the significantly damaging effects of the spike protein upon endothelial cells, it was proposed that COVID-19 is mainly a blood-borne disease, although infection first occurs in the cells of the pharyngeal wall (Salk Institute, 2021). Likewise, it is more likely that important blood vessels (i.e. the venae cavae and the aorta) will be affected in people with more severe forms of COVID-19, as the number of tissues infected by the virus is generally proportional with the intensity of the symptoms. It is important to note that the spike protein is often capable of damaging the endothelial cell barrier and entering the bloodstream (Raghavan et al., 2021), and that the way the novel SARS-CoV-2 vaccines have been designed can often allow the pathogenic protein to reach the endothelial cells and then important blood vessels. For example, the mRNA from the BNT162b2 vaccine is nucleoside-modified, as it contains nucleosides with pseudouridine, rather than wild-type uridine, and is protected by an outer layer of lipid nanoparticles, which allow the molecule to last from a few days to a few weeks, thereby entering numerous cells and producing more copies of the spike protein. Given the peculiar nature and origins of the spike protein, there is a real likelihood that there will be a spread of the spike protein and induced collateral damages in a significant number of cases worldwide. Furthermore, not only the spike protein was demonstrated via 2D static and 3D microfluidic in vitro models to be capable of crossing the blood-brain barrier and damaging neurons, but also excessive inflammatory responses induced by the spike protein can prolong the activity of monocytes, which in turn will create leaks in the blood-brain barrier and make it easier for the pathogenic protein to cross it. Namely, whilst the spike protein upregulates genes expressing a number of metalloproteinases, including MMP3, MMP9 and MMP12, which then cause leaks in the blood-brain barrier, the S1 subunit of the spike protein interacts with the Toll-Like Receptor 4 (TLR-4) on macrophages in the alveoli and monocytes in mainstream blood vessels, and induces pro-inflammatory signalling, which in turn can cause further leaks in the blood-brain barrier. In addition to this, the receptor binding domain of the same pathogenic protein was shown to at least sometimes be capable of interacting with developing central nervous system proteins involved in neuronal protection and maintenance, and likewise, causing risks of protein misfolding and the onset of neurodegenerative disease.
It is important to note that the spike glycoprotein (S protein), which represents the principal viral protein of pathophysiological induction, is deemed as cytotoxic, not only for several tissues in the human body, but also for the marine environment (Ives Charlie-Silva et al., 2021). The S protein has been shown to function as a superantigen, reaching a magnitude of an immunological impact similar to the staphylococcal enterotoxin B super-antigen when COVID-19 patients developed complications, such as a multi-systemic inflammation (Mary H. Cheng et al, 2020). Moreover, the S1 subunit and the active trimer of the viral glycoprotein are also capable of disrupting the activity of mitochondria in brain endothelial cells and likewise, have implications upon metabolism (Eun Seon Kim et al, 2021) and the 674-685 amino acid fragment of the protein was shown to prevent mitochondria from synthesising cytochrome c, which is responsible for inducing apoptosis of infected cells (Olena Kalashnik, 2021). The S protein likely causes a significant number of host cells not to undergo apoptosis if the virus and the spike protein spread to tissues in various sites of the host organism. Likewise, the S protein displays a level of toxicity significantly greater than previously thought and, although we fully support the concept of immunisation, we believe the scientific community should reconsider the spike protein-based mass vaccination approach. The vaccinal full length spike is very similar to the viral full length spike. The S protein produced by the vaccinal mRNA is very likely not less toxic, but only more immunogenic, meaning that there is a higher immune sensitivity against it than against the viral spike protein, and researchers induced two Proline amino acid substitutions in the S2 subunit of the spike protein so that it became more immunogenic. The amino acid substitutions were induced to keep the S protein in a pre-fusion conformation, but it is not fully certain that the stabilised S protein will keep its initial conformation (Timothy P. Riley et al, 2021). Likewise, there is currently still no evidence determining that the vaccinal S glycoprotein is not at all capable of entering host cells after synthesis and exocytosis, given the emergency state of the COVID-19 vaccine clinical trials in 2020 and thereby, of the limited production of conclusive data. It may be necessary to consider that the scientific community probably still has not fully apprehended the complex nature, origins and pathological flexibility of the virus and, implicitly, of the spike glycoprotein. Importantly, the translation of mRNA into a protein is only possible within the intracellular matrix, in the cytoplasm, meaning that the spike glycoprotein can only be produced, fold and be manufactured inside of a host cell. Hence, risks of harmful intracellular interference may sadly not be significantly decreased in the case of the mass administration of spike protein-based vaccines, and an increased immune sensitivity could mean a higher susceptibility for hyper-inflammatory immune responses, especially in people with one or more comorbidities.
The receptor-binding domain of the vaccinal spike protein is intact, meaning that it is as capable of binding to the ACE2 receptors of host cells as the viral spike protein, intracellular signaling cascades can become visibly disrupted and likewise, the spike protein can still cause significant intracellular damage. Likewise, risks may not be substantially decreased, and an increased immune sensitivity could mean a higher susceptibility for hyper-inflammatory immune responses, especially in people with one or more comorbidities. The cell types that were among the most affected following SARS-CoV-2 infection were inducible pluripotent STEM cells (iPSCs) located in the myocardium and the alveoli (Zhang L. et al., 2020). Likewise, it is possible that the spike protein affects those cells the most, and it is known that children and young people generally have a higher number of STEM cells in their organism. From this information and the fact that children and young people generally have a very low risk of developing morbidity and dying from COVID-19, we deduced the likelihood that children are at a significantly higher risk of developing disease and dying as a result of vaccine adverse reactions than in the case of natural infection, given that their natural immune system is also generally more prepared to build their first and second-line defences than the one of adults. Likewise, with regards to the administration of spike protein-based vaccines in children and teenagers; the following affirmation ought to be addressed; just because health authorities will be recommending only a single dose of the mRNA vaccines, which will be around a third of the dose used in adults, it does not mean risks of significant adverse reactions for this age group will visibly drop. There likely will still be a significant amount of spike protein produced, since the mRNA is nucleoside-modified and protected by lipid nanoparticles, making it potentially immune against RNA-degrading factors for one to two weeks. To exemplify, instead of 27,000 produced spike protein copies overall, there will be around 3,000 copies (~27,000 / 3 / 3) in the case of the administration of a single dose, or 6,000 copies (~27,000 / 3 / 1.5) in the case of the administration of two doses, both of which still being significant numbers for a pathogenic protein with cytotoxic effects for multiple kinds of tissues, including the endothelial ones, and with toxic effects for the aquatic environment as well. Moreover, children have a significantly lower body mass index than adults and likewise, a third of the dose for children is often equivalent to a whole dose for adults (3,000 spike proteins in a 20 kg-weight organism ~ 9,000 spike proteins in a 60 kg-weight organism). Hence, risks likely still remain outstandingly high and are probably even higher than in the case of young adults aged 18-25. It is likely much safer for children and teenagers to receive low-dose Interferon I-based nasal sprays instead.
As time went by and the disease became visibly endemic, the mass vaccination campaign displayed poorer long-term results than it was desired and initially projected by the scientific community, despite the unprecedented intellectual and clinical efforts to create life-saving vaccines against COVID-19, and despite the successful induction of variant-specific IgM and IgG immunoglobulin production, as new viral variants often evaded the recognition of immunoglobulins developed after the reception of the spike glycoprotein-based vaccine(s). In the process, an unprecedented number of people also developed serious adverse reactions, and the frequency and severity of both short- and long-term reactions as such were particularly higher in people who either received the adenoviral vector 5/26-based, the mRNA-1273 mRNA-based or at least two or three doses of the BNT162b2 mRNA-based vaccine, depending also on other variables, like health status and age. Numerous people who received even three or four doses of the BNT162b2 mRNA vaccine eventually developed COVID-19 again, many people even repeatedly, and the extent of symptoms was numerous times nearly the same as the symptoms developed by unvaccinated people who contracted the illness from the time it started becoming endemic. During the recent couple of years, vaccinated people became even more susceptible to infection than unvaccinated people, likewise pointing toward the significantly important role trained immunity played in turning the pandemic disease into an endemic one, despite the paradox that the aid for reaching a threshold level of timing and quality with regards to adaptive immune activation plays a critical role in combating complex epidemics and pandemics. The overall events that occurred during the past few years may in fact expose an unintentional negligence of the critical role that natural immunity plays in protecting the human body against polymorphic viruses like SARS-CoV-2, and such a negligence occurred whilst an unprecedented extent of financial investment occurred in the effort to find effective approaches of clinical response against the disease. Would it not be much more efficient, less time consuming and financially demanding to create such sprays, support the mucosal immune system more directly and eventually outcompete the virus evolutionarily by filling in the gaps in the cellular immune system created mainly by the viral non-structural proteins 1, 10, 14 and 16?
Could interferons also contribute as a potential cancer therapeutic? Given that early Interferon I and III-based immune responses are associated with significant levels of prophylactic anti-cancer cellular mechanisms (Yu et al., 2022), it is probable that low dose interferon I and III-based nasal sprays will also upregulate the activity of important proto-oncogenes and tumour suppressor genes, thereby partially restoring the ability of cells to perform the necessary DNA repair mechanisms and potentially preventing numerous incidences of mutagenesis and genome toxicity over the next several decades. Furthermore, performing thorough studies on the links between a virally hijacked host immune system and the onset of cancer is likely the next important step in understanding the relationship between cellular immunology and cancer aetiology. Nevertheless, the highest impact of the antiviral proteins will be upon the ability of the virus to self-camouflage, to significantly downregulate interferon synthesis and signalling, and to inhibit antigen presentation. Furthermore, it has recently been discovered that fungi often grow in and around cancerous cells, and this finding further indicates that cancer is an immunological problem as well (Dohlman et al., 2022). Likewise, interferons may prevent the formation of tumours because they often sharpen first-line immune responses to the point of stimulating a rapid induction of apoptosis of the affected cells, as well as the spread of toxins that may be developing inside and around them.
Around the time the AIDS pandemic started, the TIME magazine came with two important suggestions: that interferon-alpha drops can significantly impact cancer and that it can treat the common cold. Moreover, Dr. Anthony Fauci acknowledged once the growing evidence that low dose Interferon I-based drops are effective against AIDS and yet, during the current pandemic, the same governmental scientist first insisted there was no cure or significant method of treatment against COVID-19, and now that the only way to pull civilisation out of the pandemic is to introduce another significant amount of spike proteins intramuscularly, according to classical methods of immunisation, which we have well-founded reasons to believe they do not quite apply in this context, especially for groups of lower risk. Withholding information from or refusing to testify to the public about critical past research represents one of the greatest bioethical issues that the world has been facing, and financial interests and institutional corruption may represent two important factors of this issue, and the publication of secret contracts between Pfizer and world governments that occurred at the beginning of 2022, containing information about serious adverse reactions that are probable to occur after the administration of BNT162b2 vaccines in a significant number of people represents a relevant example of such corruption, and this example also marks a failure of reaching informed consent in a significant number of cases, which is an unprecedented situation. Overall, we believe that trialling the prophylactic and therapeutic effects of Type I and Type III Interferons, and testing their position in relation to vaccinology are two steps of prime importance and may contribute to the greatest clinical research in modern history. The foundational factors of malignant tumours and neurodegenerative disease are immunological in nature, and the problem of immune escape represents an important stronghold of the worldwide epidemic of cancer and neurodegeneration. Immune escape not only facilitates pathogens to infect kinds of tissues that are vulnerable to mutagenesis and genome toxicity, but also prevents the apoptosis of cells that have already undergone tumour-related mutagenesis. Understanding the key mechanisms of cellular signalling resulting in the phenomenon known as "wise immune sharpening" represents the number one objective of cancer research. Weakened first and second-line immune responses are not only caused by active pathogens, but by a series of genetic factors that likely emerged as a result of a repeated exposure of ancestors with diverse immune comorbidities to pathogens of more significant concerns. Understanding the spectrum of genetic-epigenetic factors that favour a specific outcome in offspring is also important in increasing the resolution of the details collected during applied immunological research in cancer biology.
Recent in-vitro and in-vivo research has indicated the presence of a link between type I Interferon-based signalling and LINE-1 retro-transposition. Namely, the study suggests that type IFN I and LINE-1 retro-transposons regulate each other, and that an exaggerated type I IFN expression is linked to a higher incidence of autoimmune disease. With regards to SARS-CoV-2 and the spike protein, it was previously indicated that small regions of the viral genome undergo LINE-1 reverse transcription and integration into various parts of the junk and functional DNA of the host cell and, given that long COVID is a result of a delayed and exaggerated extent of IFN-I autocrine and paracrine signalling, it is possible that the viral infection results in a less regulated propagation of LINE-1 retro-transposons. According to evidence collected from genetic studies, it is possible that substantially small, but still concerning, parts of the viral genome have laboratory origins. There are various circulating theories, including that the virus having had natural origins beforehand underwent gain-of-function research to test the way the immune system of bats would react to it. Furthermore, the activity of LINE-1 Reverse Transcription upon roughly 1% of the SARS-CoV-2 genome is new, given that all members of the Betacoronavirus genus are widely different and unrelated to the members of the Betaretrovirus genus. SARS-CoV-2 has outstanding evolutionary abilities of suppressing Type I Interferon-based transmission, which is outstandingly concerning, no matter whether it has fully natural origins or a number of inserts from other pathogens. Furthermore, given the likelihood that the timing and extent of IFN I production influence the spread of LINE-1 retrotransposons, it is statistically probable that, the more capable a virus is of suppressing IFN-I production and distribution to neighbouring cells, the more capable such a virus is of inserting fragments of its genome into the DNA of the host cell, given that the LINE-1 retrotransposon-encoding DNA represents around 17% of the human genome. Likewise, the more capable a virus is of suppressing first-line immune responses, the higher the risks are that such viral infections will ultimately result in the formation of cancers. As a result, the research is approaching the conclusion that facilitated early and regulated IFN-I responses are paramount not only in maintaining anti-viral immunity and preventing the onset of various autoimmune diseases, but also in preventing an exaggerated spread of LINE-1 retro-transposons and maintaining genomic integrity. The research has also started to directly challenge the safety and long-term effectiveness of spike protein-based mRNA and adenoviral vaccines that had been developed during the midst of the COVID-19 pandemic. We are therefore requesting immediate study and clinical trials involving the efficacy of low-dose Interferon I and III-based nasal sprays in human and animal vaccinology, particularly for viruses and bacteria of significant concern. Additionally, we are proposing that such nasal sprays will play a major role in a collective prophylactic and early therapeutic fight against retroviral infection and also in the prolongation of the host cellular and systemic lifespan.
Given the powerful effects of the combination of restricted C5a complement activation and a robust activation of Type I and III Interferon signalling, updating vaccinology accordingly may slowly reduce the intensity and morbidity of numerous diseases from "incurable" to " flu and common cold-like" as decades and centuries pass. In a number of cases, such an approach could be merged with other discovered approaches to ensure long-term effectiveness (i.e. the protollin-based potential nasal vaccine against Alzheimer's disease). Hence, this may be the case given the central role of immunology in facilitating general human and animal wellbeing. COVID-19 may represent a very important opportunity to discover the hidden power of human natural immunity and to facilitate the inclusion of sharpening natural immunity into the efforts of vaccinology-related medical research. To note, for scientific and academic research, the term "sensationalization" represents one of the most important antagonistic terms, and big words have often been disproven in front of the committee. Likewise, it is of an essence for scientists and academicians to perform their due diligence before making any scientific observation, and this study is not exempt from the obligation to perform prior due diligence.
Numerous kinds of significant infectious diseases implicate a latter hyper-activated interferon set of responses, alongside a severe complement-mediated set of immune signals, which often result in the onset of hemophagocytic lymphohistiocytosis, which is in short known as cytokine storm. The truth is that a desired extent of complement system activation also stimulates an early development of interferon signalling, which means that eventual updates in vaccinology could require the inclusion of approaches sharpening complement system-related immunity. A delayed Type I Interferon-based immune response is often accompanied by an exaggerated Complement C3a and C5a-related immune response, which may indicate that the quality of the complement system’s activation process is not fully autonomous of the quality of the activated interferon system. Clinical research involving the search for methods to prevent exaggerated activations in this part of the complement system could also support the development of an early Interferon I and perhaps Interferon III-based antiviral and anti-inflammatory signals, although it is clear that more evidence needs to be found to confirm that the interferon system also displays a degree of dependency upon the complement system, and not solely the other way around (Shibabaw T., 2020). The reason why the discrepancy in timing is significantly elevated is that the production of interferon-alpha, -beta, -delta, -epsilon, -kappa, -omega and lambda takes place as a result of viral sensing, and when the sensing process is temporarily inhibited, then a sudden process of viral sensing will take place in a pronouncedly higher number of host cells, leading to a much higher number of synthesised interferons, which will only contribute to hyper-apoptotic and hyper-inflammatory processes, leading to a large extent of tissue damage and demand for a replacement of the cells that underwent programmed death. In numerous cases, delayed first and second-line immune signalling will have allowed the virus to replicate and spread to tissues in the lower respiratory tract, which in turn will significantly raise the likelihood of the development of severe inflammatory responses, particularly if viral copies have reached bronchioles and alveoli. There will be a much higher extent of natural killer and dendritic cell recruitment, alongside the activation and specification of significantly more lymphocytes. Pro-inflammatory cytokines will become more active than anti-inflammatory chemokines, often leading to a severely disrupted set of antiviral immune responses, and the focus on antiviral action will become much higher than the focus on synthesising qualitative antibodies. Consequently, the onset of hemophagocytic lymphohistiocytosis will be much more likely, particularly in patients with underlying health conditions, which predispose their immune system to overreact.
Likewise, IFNs can be deemed as a double-edged sword in immunology and cancer biology, and herein lies the importance of ensuring that such chemokine-based signaling takes place significantly during the early stages of the disease, and that prevention methods against the pathogenesis of cancer implicate the stimulation of a solid interferon-based response during such a critical stage, before the disease can advance. Moreover, certain approaches could be merged with this immunological method to bring broader positive changes in the fight against cancer, and certain immunosuppressive methods could be temporarily applied for latter stages of cancers and infectious diseases in order to restrict the extent of induced inflammatory responses. Sharpening interferon-based immune responses on an evolutionary scale is likely still enough to improve general human immunity against pathogens with tricky mechanisms of pathogenesis. Nevertheless, a prophylactic approach against a hyperactive complement system will possibly bring a further shift against pathogens with abilities of immune evasion.
Furthermore, given that correct Interferon I and III-based immune signalling has a central importance in maintaining a fortified and wise immune system, it is possible that developing Interferon I and III-based vaccines will reach areas of efficacy as far as the prolongation of the average human lifespan. Moreover, a high energy metabolism, which is associated with high levels of stress and general poor alimentation, leads to a shorter lifespan, as the increased demand for energy consumption implicates an increased speed of cellular and tissular ageing, despite the recent advancements in scientific and medical research. For example, there is a correlation between an activated hexosamine process and an activated interferon pathway during hyperinflammation (Hugo et al., 2020), meaning that a more sensitised interferon system could prevent exaggerated hexosamine-related metabolic processes, which would play a considerable part in lowering the overall metabolic consumption rate, lower cellular stress and prolong cellular and tissular life expectancy long-term. Furthermore, one mathematical model-based study suggests that early interferon activation via the organisation of the cell cycle and of the mTOR pathway results in the post-transcriptional inhibition of Sox2 and the induction of quiescence, ultimately leading to a decrease of the maturation rate of neuronal cells of substantial importance, which was shown to have good effects for young people and decaying effects for the elderly. Interferons were shown to be major regulators of neuronal STEM cell homeostasis, which could explain the lack of efficacy and rather increased process of ageing for the aged brain due to the generally decreased quality of the immune system, the highly solicited metabolic processes and the overall resulted contribution to neuronal ageing (Ibanez et al., 2023). Likewise, early and local interferon production and activation may be associated with a slower neuronal ageing process in young and middle aged people. A general re-establishment of human lifestyle to a nature-friendly point may involve a lower energy metabolism, as the general levels of stress and energy consumption will significantly decrease. There is a wide availability of resources due to the high number and wide areas of nature-friendly areas worldwide; it is the increase of human and societal corruption that allowed for a progressive development of a general crisis of lifestyle. Non-qualitative food and beverages are epigenetically and ultimately genetically predisposing human metabolism to switch to high-energy consumption, thereby accelerating the process of ageing. It is possible that a repeated set of developed early, regulated interferon-based responses over a longer time reduce the rate of cellular ageing, whilst a repeated set of developed late, exaggerated interferon-based responses increase the speed of cellular ageing.
If the interferon-based approach is proven to be successful as a prophylactic, immunising and early therapeutic method against important pathogenic agents and the onset of cancers, then it will constitute a significant step in prolonging the lifespan of humanity, with an estimated increase of the average human lifespan of at least 10 to 20 years over the following several decades. Interferon I and III will not only increase the wisdom of the immune system, so to speak, but it will significantly strengthen important DNA repair mechanisms and ultimately repair many of the past damages done upon human metabolism as well. A process of immune wisening will not only involve better first and second-line responses due to a combined approach to regulate C5a complement activation and stimulate IFN I and III may result in a decreased sensitivity of complement system activation and an increased sensitivity for Type I and III-based signalling and immune sensing of many problematic microbes, and will lead to the production of more qualitative IgM and IgG antibodies during challenging infections. Research has shown that the administration of interferon-alpha to pregnant mothers is safe for the foetuses and that it increases the probability that the future baby will be immunologically healthy. This possibly means the effectiveness of such interferons will cross the umbilical cord and reach the health state of the foetus as well. Recently, researchers have also suggested the utilisation of recombinant NK and T-lymphocytes, alongside dendritic cells, altogether treated with a fairly low dosage of Type I and possibly Type III Interferons beforehand to possibly increase their state of integrity and ability of immune performance, in the efforts to develop natural immune system-based vaccines and therapeutics against future public health crises, given also the context in which the human immunodeficiency virus (HIV) is known to directly infect and kill major subtypes of T-lymphocytes. In such a particular case, the prior treatment of CD4+ and CD8+ T-lymphocytes with Type I and Type III IFNs could bring the most proportionate extent of evolutionary self-defence against pathogenic agents like HIV (Carp T. et al., 2024). The addition of dendritic cells into the equation would be due to the fact that they represent main “factories” of Type I and Type III IFNs. Given the context of heavy innate immune evasion by polymorphic viruses, it would not be recommended for the research community to focus upon developing adaptive and pathogen-based immunisation candidates, given the risk of immune defences becoming too specific for such particular viruses, and only for their current variants. The objective for vaccine development should include the reach of a balance between innate and adaptive immune methodology as the main objective. Not to mention that the yet not fully known origins of SARS-CoV-2 and of the multi-dimensionally pathological extent of the spike glycoprotein, does not allow the scientific community to rely on claims that the spike glycoprotein-based vaccines are safe and effective for the entire world population, despite the colossal efforts of researchers and clinical scientists to rapidly resolve the problem of health system overload caused by the high rate of viral transmission and by the life-altering symptoms of the disease, more prevalently in the people of an advanced age, with one or more significant comorbidities. Polymorphic viruses in general also discourage clinicians from developing pathogen-based vaccines, and sometimes even vaccines based on adaptive immunity, at least without a major aid of natural immune elements, because the developed antibodies would be specific for certain viral variants and future mutations would likely result in the virus evading the previously developed antibodies, alongside its newly developed methods of quickly evading first- and second-line immune signals. In other words, situations of “going back to square one” would often be reached if specificity would be solely relied upon by therapeutics and vaccine developers, with major research and public health funds proving to be wasted in the process. Likewise, updating and revolutionising human vaccinology in the manner described may bring a level of breakthrough the great minds in the past centuries only dreamt about, and believing in a positive outcome and paying close attention to new, conclusive evidence, represents a major step toward a desired extent of scientific and medical progress.
Conclusion
Given results from NCT data, there is a high number of reasons to believe that low-dose type IFN I and III IFN based nasal sprays could match the definition of a therapeutic vaccine with beneficial effects than prophylactic ones.Therefore, whilst potential vaccine candidates can prevent severe cases of COVID-19 disease, it would have the highest impact when administered in varying populations of vulnerable people potentially with or without comorbidities who became infected and developed symptoms that would be noticeable . Examining further the administration of type I IFN from NCTs clarifies that IFN alpha-2b, epsilon, omega and perhaps IFN beta1, (BAI et al., 2022) and type III (IFN lambda1, lambda2 and lambda3) may act as mimitics of cellular infection and stimulate a robust and natural immune response against either viral or neoplastic pathologies, and likewise, such an intervention fits the definition of a traditional vaccine. Because this likely requires further research into dosages, we might refer to the potential immunisation process as a long-term one. Hence, there is a probability that the medical community does not need to insert any viral fragments to stimulate a desired immune response and ultimately gain immune memory. It is essential to mention that an excess of administered interferon-alpha could give rise to the probability of developing adverse reactions, like epistaxis, headaches and dizziness in this case, since an excess amount of type I in the tissue would be caused. The quantity of administered IFN differs based on the disease. For example, a higher amount of IFNs would generally be needed for COVID-19 than the general flu, and for people experiencing significant immunosuppression. Administered interferons could, on the other hand, contribute to pathology induced by autoimmunity, meaning that caution is likely needed nevertheless. Compared to the dosage of experimental injectable interferons, which generally ranges from 3 to 6 million International Units, which is measured in milligrams, the nasal sprays generally contained an interferon concentration ranging from 200 to 500 International Units when used in past clinical trials, which is measured in nanograms. The best part of this discovery is that a matching dosage of Interferon I might bring us a few steps forward with regards to the preparation for a future potential avian influenza pandemic, but in that case, it would be slightly more useful and safer for the compound to be administered only as soon as symptoms develop or when certain tests indicate the presence of the virus in the organism. With regards to the dosage based on age and health condition groups, we are hypothesising a session of vaccination of two to four weeks, with a daily dosage of 350-375 International Units for almost all adults, and 200-225 International Units for children, given their lower BMI, as well as for adults with blood and autoimmune conditions due to slightly increased risks of epistaxis in children and the aggravation of the autoimmune conditions in the affected adults. Given their widespread and trans-species immune modulatory and stimulatory effects, we hypothesise that low-dose interferon-containing substances could be administered to live birds to prevent the spread of the present avian Influenza A viral variants in chicken, which would in turn save animal lives and plenty of alimentary resources for the human society during the present financial and resource availability crises. Overall, the present scientific paper does not mention medical statements, but scientific hypotheses encouraging clinical researchers to further investigate the matter and collect pieces of evidence that would eventually reach conclusive stages. The dosages measured during clinical trials involving Type I and Type III Interferons only constitute part of preliminary research reports, which implies that further clinical research is required to be performed, by medical doctors, pharmaceutical clinicians and research scientists, to determine more exact amounts of interferons that could be used, depending on factors such as age, health status and type of disease to be prevented or attenuated in the early stages of onset. We strongly encourage further testing of such interferon substances and propose that they will play a major part in improving immunological, metabolic and systemic wellbeing in the human body, as well as in animals.
Author Contributions
Conceptualization:.; methodology:.; formal analysis,.; data curation ; writing—original draft preparation,.; writing—review and editing,.; funding acquisition, no funding was requested. All authors have read and agreed to the published version of the manuscript.
Funding
This article was funded by: N/A
Institutional Review Board Statement
This research did not require ethical approval.
Informed Consent Statement
As far as the authors are aware, cited papers and all authors gave informed consent for this publication as did participants within the cited clinical trials.
Data Availability Statement
All related data is presented in this paper.
List of Abbreviations
SARS-CoV-2 - Severe Acute Respiratory Syndrome-Coronavirus 2
COVID-19 - Coronavirus Infectious Disease 2019
ECM - Extracellular Matrix
ACE2 - Angiotensin Convertase Enzyme 2
IAV - Influenza A Virus
RSV - Respiratory Syncytial Virus
ICM - Intracellular Matrix
ORF - Open Reading Frame
NSP - Non-Structural Protein
IFN - Interferon
PAMP - Pathogen-Associated Molecular Pattern
PRR - Pattern Recognition Receptor
TLR - Toll-Like Receptor
MDA-5 - Melanoma-Differentiation Associated protein 5
RIG-I - Retinoic acid-Inducible Gene I
ADAR - Adenosine Deaminase Acting on RNA / RNA-specific Adenosine Deaminase
ADAR1 - Adenosine Deaminase 1
MyD88 - Myeloid-Differentiation primary response 88
IKK - IkB Kinase
IFNAR - Interferon alpha/beta Receptor
IFNLR1 - Interferon lambda Receptor 1
IL10R1 - Interleukin-10 Receptor 1
STAT - Signal Transducer and Activator of Transcription
IRF - Interferon Regulatory Factor
NLRC - Nucleotide-binding oligomerization domain-Like Receptor C
NF-kB - Nuclear Factor-kappa-light-chain-enhancer of B-Lymphocytes
ISG - Interferon-Stimulated Gene
NK Cell - Natural Killer Cell
IFITM3 - Interferon-Induced Transmembrane Protein 3
cGAS-STING - cyclic AMP-GMP Synthase - Stimulator of Interferon Genes
N7-MTase - C-terminal N7 Methyltransferase
ExoN - N-terminal 3’-5’ Exoribonuclease
IL - Interleukin
TNF - Tumour Necrosis Factor
TGF-α - Tumour Growth Factor-alpha
TGF-β - Transforming Growth Factor-beta
CRP - C-Reactive Protein
CCL - Chemokine (C-C motif) Ligand
CXCL - Chemokine (C-X-C motif) Ligand
CCR - Chemokine (C-C motif) Receptor
CXCR - Chemokine (C-X-C motif) Receptor
NK Cell - Natural Killer Cell
IMM - Inflammatory Monocyte and Macrophage
pDC - plasmacytoid Dendritic Cell
HLA - Human Leukocyte Antigen
MHC - Major Histocompatibility Complex
Th Lymphocyte - T-helper Lymphocyte
CTL - Cytotoxic T-Lymphocyte
CD - Cluster of Differentiation
LINE-1 - Long Interspersed Nuclear Element 1
AIDS - Acute Immunodeficiency Syndrome
Ab - Antibody
Ig - Immunoglobulin
C5a - Complement component 5a
GRP78 - 78 kDa-Glucose-Regulated Protein
HSP - Heat-Shock Protein
ARS - Acute Radiation Syndrome
+ssRNA - positive-sense single-stranded Ribonucleic Acid
BMI - Body-Mass Index
IU - International Unit
References
- Ablasser, A., Bauernfeind, F., Hartmann, G. et al. RIG-I-dependent sensing of poly(dA:dT) through the induction of an RNA polymerase III–transcribed RNA intermediate. Nat Immunol 10, 1065–1072 (2009). [CrossRef]
- Ablasser, A., Hertrich, C., Waßermann, R., & Hornung, V. (2013). Nucleic acid driven sterile inflammation. Clinical immunology (Orlando, Fla.), 147(3), 207–215. [CrossRef]
- Aboulnasr, F., Hazari, S., Nayak, S., Chandra, P. K., Panigrahi, R., Ferraris, P., Chava, S., Kurt, R., Song, K., Dash, A., Balart, L. A., Garry, R. F., Wu, T., & Dash, S. (2015). IFN-λ Inhibits MiR-122 Transcription through a Stat3-HNF4α Inflammatory Feedback Loop in an IFN-α Resistant HCV Cell Culture System. PloS one, 10(12), e0141655. [CrossRef]
- Abushahba, W., Balan, M., Castaneda, I., Yuan, Y., Reuhl, K., Raveche, E., de la Torre, A., Lasfar, A., & Kotenko, S. V. (2010). Antitumor activity of type I and type III interferons in BNL hepatoma model. Cancer immunology, immunotherapy : CII, 59(7), 1059–1071. [CrossRef]
- Acevedo Ospina, H., Guay-Vincent, M.-M., & Descoteaux, A. (2022). Macrophage mitochondrial biogenesis and metabolic reprogramming induced by leishmania donovani require lipophosphoglycan and type I interferon signaling. MBio. [CrossRef]
- Acosta, P. L., Byrne, A. B., Hijano, D. R., & Talarico, L. B. (2020). Human Type I Interferon Antiviral Effects in Respiratory and Re-emerging Viral Infections. Journal of immunology research, 2020, 1372494. [CrossRef]
- Adolfo Garcia-Sastre and Christine A. Biron (2006), Type I Interferons and the Virus-Host Relationship: A lesson in Detente, Science, Vol 312, 5775, pp 879-882. [CrossRef]
- Adusumilli, N. C., Zhang, D., Friedman, J. M., & Friedman, A. J. (2020). Harnessing nitric oxide for preventing, limiting and treating the severe pulmonary consequences of COVID-19. Nitric oxide : biology and chemistry, 103, 4–8. [CrossRef]
- Agy, M. B., Acker, R. L., Sherbert, C. H., & Katze, M. G. (1995). Interferon treatment inhibits virus replication in HIV-1- and SIV-infected CD4+ T-cell lines by distinct mechanisms: evidence for decreased stability and aberrant processing of HIV-1 proteins. Virology, 214(2), 379–386. [CrossRef]
- Aiman, A., Basir, S. F., & Islam, A. (2022). Interferons Horizon Therapeutics. In (Ed.), Basic and Clinical Aspects of Interferon Gamma. IntechOpen. [CrossRef]
- Akiyama, H., Ramirez, N. P., Gibson, G., Kline, C., Watkins, S., Ambrose, Z., & Gummuluru, S. (2017). Interferon-Inducible CD169/Siglec1 Attenuates Anti-HIV-1 Effects of Alpha Interferon. Journal of virology, 91(21), e00972-17. [CrossRef]
- Albrecht, J. D., Ninosu, N., Barry, D., Albrecht, T., Schaarschmidt, M. L., Goerdt, S., & Nicolay, J. P. (2022). Non-pegylated and Pegylated Interferon Alpha-2a in Cutaneous T-cell Lymphoma and the Risk of Severe Ocular Side-effects. Acta dermato-venereologica, 102, adv00722. [CrossRef]
- Aldén M, Olofsson Falla F, Yang D, Barghouth M, Luan C, Rasmussen M, De Marinis Y. Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 mRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line. Current Issues in Molecular Biology. 2022; 44(3):1115-1126. [CrossRef]
- Alexander, M. R., Brice, A. M., Jansen van Vuren, P., Rootes, C. L., Tribolet, L., Cowled, C., Bean, A., & Stewart, C. R. (2021). Ribosome-Profiling Reveals Restricted Post Transcriptional Expression of Antiviral Cytokines and Transcription Factors during SARS-CoV-2 Infection. International journal of molecular sciences, 22(7), 3392. [CrossRef]
- Alfi, O., Yakirevitch, A., Wald, O., Wandel, O., Izhar, U., Oiknine-Djian, E., Nevo, Y., Elgavish, S., Dagan, E., Madgar, O., Feinmesser, G., Pikarsky, E., Bronstein, M., Vorontsov, O., Jonas, W., Ives, J., Walter, J., Zakay-Rones, Z., Oberbaum, M., Panet, A., … Wolf, D. G. (2021). Human Nasal and Lung Tissues Infected Ex Vivo with SARS-CoV-2 Provide Insights into Differential Tissue-Specific and Virus-Specific Innate Immune Responses in the Upper and Lower Respiratory Tract. Journal of virology, 95(14), e0013021. [CrossRef]
- Ali, S., Mann-Nüttel, R., Schulze, A., Richter, L., Alferink, J., & Scheu, S. (2019). Sources of Type I Interferons in Infectious Immunity: Plasmacytoid Dendritic Cells Not Always in the Driver's Seat. Frontiers in immunology, 10, 778. [CrossRef]
- Alibek K, Tskhay A (2020) Ahead of a vaccine: A safe method of protection against COVID-19 exists. Research Ideas and Outcomes 6: e61709. [CrossRef]
- Alice Barbarin, André Herbelin, Jean-Marc Gombert. [The CD8+ T cell innate function in the war against cancer].. médecine/sciences, EDP Sciences, 2017, 33 (11), pp.927-929. 10.1051/med-sci/20173311004. inserm-02508042 (PDF) [The CD8+ T cell innate function in the war against cancer].. Available from: https://www.researchgate.net/publication/339946901_The_CD8_T_cell_innate_function_in_the_war_against_cancer [accessed Oct 05 2022].
- Alicea-Torres, K., Sanseviero, E., Gui, J., Chen, J., Veglia, F., Yu, Q., Donthireddy, L., Kossenkov, A., Lin, C., Fu, S., Mulligan, C., Nam, B., Masters, G., Denstman, F., Bennett, J., Hockstein, N., Rynda-Apple, A., Nefedova, Y., Fuchs, S. Y., & Gabrilovich, D. I. (2021). Immune suppressive activity of myeloid-derived suppressor cells in cancer requires inactivation of the type I interferon pathway. Nature communications, 12(1), 1717. [CrossRef]
- Aliyari, S. R., Quanquin, N., Pernet, O., Zhang, S., Wang, L., & Cheng, G. (2022). The Evolutionary Dance between Innate Host Antiviral Pathways and SARS-CoV-2. Pathogens (Basel, Switzerland), 11(5), 538. [CrossRef]
- Alunno, A., Najm, A., Mariette, X., De Marco, G., Emmel, J., Mason, L., McGonagle, D. G., & Machado, P. M. (2021). Immunomodulatory therapies for SARS-CoV-2 infection: a systematic literature review to inform EULAR points to consider. Annals of the rheumatic diseases, 80(6), 803–815. [CrossRef]
- Al-Zaidan, L., Mestiri, S., Raza, A., Merhi, M., Inchakalody, V. P., Fernandes, Q., Taib, N., Uddin, S., & Dermime, S. (2021). The expression of hACE2 receptor protein and its involvement in SARS-CoV-2 entry, pathogenesis, and its application as potential therapeutic target. Tumour biology : the journal of the International Society for Oncodevelopmental Biology and Medicine, 43(1), 177–196. [CrossRef]
- Amanda J. Lee and Ali A. Ashkar (2018), The Dual Nature of Type I and Type II Interferons, Frontline Immunology. [CrossRef]
- Amanda J. Lee and Ali A. Ashkar (2018), The Dual Nature of Type I and Type II Interferons, Frontline Immunology. [CrossRef]
- Amarante-Mendes, G. P., Adjemian, S., Branco, L. M., Zanetti, L. C., Weinlich, R., & Bortoluci, K. R. (2018). Pattern recognition receptors and the host cell death molecular machinery. Frontiers in immunology, 9, 2379. [CrossRef]
- Amos C Lee, Yunjin Jeong, Sumin Lee, Haewook Jang, Allen Zheng, Sunghoon Kwon and John E. Repine (19/05/2021), Nasopharyngeal Type I-Interferon for Immediately Available Prophylaxis Against Emerging Respiratory Viral Infections, Front. Immunol., 12.660298. [CrossRef]
- Andersson, U., Yang, H., & Harris, H. (2018). High-mobility group box 1 protein (HMGB1) operates as an alarmin outside as well as inside cells. Seminars in immunology, 38, 40–48. [CrossRef]
- Anjum, F. R., Anam, S., Abbas, G., Mahmood, M. S., Rahman, S. U., Goraya, M. U., Abdullah, R. M., Luqman, M., Ali, A., Akram, M. K., & Chaudhry, T. H. (2021). Type I IFNs: A Blessing in Disguise or Partner in Crime in MERS-CoV-, SARS-CoV-, and SARS-CoV-2-Induced Pathology and Potential Use of Type I IFNs in Synergism with IFN-γ as a Novel Antiviral Approach Against COVID-19. Viral immunology, 34(5), 321–329. [CrossRef]
- Ank, N., West, H., Bartholdy, C., Eriksson, K., Thomsen, A. R., & Paludan, S. R. (2006). Lambda interferon (IFN-lambda), a type III IFN, is induced by viruses and IFNs and displays potent antiviral activity against select virus infections in vivo. Journal of virology, 80(9), 4501–4509. [CrossRef]
- Arndt, W. D., Cotsmire, S., Trainor, K., Harrington, H., Hauns, K., Kibler, K. V., Huynh, T. P., & Jacobs, B. L. (2015). Evasion of the Innate Immune Type I Interferon System by Monkeypox Virus. Journal of virology, 89(20), 10489–10499. [CrossRef]
- Arvin, A. M., Fink, K., Schmid, M. A., Cathcart, A., Spreafico, R., Havenar-Daughton, C., Lanzavecchia, A., Corti, D., & Virgin, H. W. (2020). A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nature, 584(7821), 353–363. [CrossRef]
- Avota, E., Gassert, E., & Schneider-Schaulies, S. (2010). Measles virus-induced immunosuppression: from effectors to mechanisms. Medical microbiology and immunology, 199(3), 227–237. [CrossRef]
- Avota, E., Koethe, S., & Schneider-Schaulies, S. (2013). Membrane dynamics and interactions in measles virus dendritic cell infections. Cellular microbiology, 15(2), 161–169. [CrossRef]
- Azamor, T., Cunha, D. P., da Silva, A., Bezerra, O., Ribeiro-Alves, M., Calvo, T. L., Kehdy, F., Manta, F., Pinto, T., Ferreira, L. P., Portari, E. A., Guida, L., Gomes, L., Moreira, M., de Carvalho, E. F., Cardoso, C. C., Muller, M., Ano Bom, A., Neves, P., Vasconcelos, Z., … Moraes, M. O. (2021). Congenital Zika Syndrome Is Associated With Interferon Alfa Receptor 1. Frontiers in immunology, 12, 764746. [CrossRef]
- Babiuch, L., Mian, M., Kamińska, E., Szymańska, B., & Georgiades, J. A. (1993). An interim report on the effect of natural human interferon alpha (IFN-alpha) lozenges in patients seropositive for the human immunodeficiency virus type 1 (HIV-1). Archivum immunologiae et therapiae experimentalis, 41(3-4), 213–219, doi: https://pubmed.ncbi.nlm.nih.gov/7907465/.
- Baechler, E. C., Bilgic, H., & Reed, A. M. (2011). Type I interferon pathway in adult and juvenile dermatomyositis. Arthritis research & therapy, 13(6), 249. [CrossRef]
- BAI, H., KAWAHARA, M., TAKAHASHI, M., & IMAKAWA, K. (2022). Recent progress of Interferon-Tau Research and potential direction beyond pregnancy recognition. Journal of Reproduction and Development, 68(5), 299–306. [CrossRef]
- Barber G. N. (2001). Host defense, viruses and apoptosis. Cell death and differentiation, 8(2), 113–126. [CrossRef]
- Barilli, A., Visigalli, R., Ferrari, F., Recchia Luciani, G., Soli, M., Dall'Asta, V., & Rotoli, B. M. (2022). The JAK1/2 Inhibitor Baricitinib Mitigates the Spike-Induced Inflammatory Response of Immune and Endothelial Cells In Vitro. Biomedicines, 10(9), 2324. [CrossRef]
- Barros-Martins, J., Förster, R. & Bošnjak, B. NK cell dysfunction in severe COVID-19: TGF-β-induced downregulation of integrin beta-2 restricts NK cell cytotoxicity. Sig Transduct Target Ther 7, 32 (2022). [CrossRef]
- Baruch, K., Deczkowska, A., David, E., Castellano, J. M., Miller, O., Kertser, A., Berkutzki, T., Barnett-Itzhaki, Z., Bezalel, D., Wyss-Coray, T., Amit, I., & Schwartz, M. (2014). Aging. Aging-induced type I interferon response at the choroid plexus negatively affects brain function. Science (New York, N.Y.), 346(6205), 89–93. [CrossRef]
- Basler C. F. (2012). Nipah and hendra virus interactions with the innate immune system. Current topics in microbiology and immunology, 359, 123–152. [CrossRef]
- Basler C. F. (2015). Innate immune evasion by filoviruses. Virology, 479-480, 122–130. [CrossRef]
- Basler, C. F., & Amarasinghe, G. K. (2009). Evasion of interferon responses by Ebola and Marburg viruses. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research, 29(9), 511–520. [CrossRef]
- Bastard, P., Rosen, L. B., Zhang, Q., Michailidis, E., Hoffmann, H. H., Zhang, Y., Dorgham, K., Philippot, Q., Rosain, J., Béziat, V., Manry, J., Shaw, E., Haljasmägi, L., Peterson, P., Lorenzo, L., Bizien, L., Trouillet-Assant, S., Dobbs, K., de Jesus, A. A., Belot, A., … Casanova, J. L. (2020). Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science (New York, N.Y.), 370(6515), eabd4585. [CrossRef]
- Batiha, G. E., Al-Gareeb, D., Qusti, S., Alshammari, E. M., Rotimi, D., Adeyemi, O. S., & Al-Kuraishy, H. M. (2021). Common NLRP3 inflammasome inhibitors and Covid- 19. [CrossRef]
- Battistini, C., Ballan, R., Herkenhoff, M. E., Saad, S., & Sun, J. (2020). Vitamin D Modulates Intestinal Microbiota in Inflammatory Bowel Diseases. International journal of molecular sciences, 22(1), 362. [CrossRef]
- Bayati, A., Kumar, R., Francis, V., & McPherson, P. S. (2021). SARS-CoV-2 infects cells after viral entry via clathrin-mediated endocytosis. The Journal of biological chemistry, 296, 100306. [CrossRef]
- Beldarraín, A., Cruz, Y., Cruz, O., Navarro, M. and Gil, M., 2001. Purification and conformational properties of a human interferon α2b produced in Escherichia coli. Biotechnology and applied biochemistry, 33(3), pp.173-182. [CrossRef]
- Bellanti, F., Vendemiale, G., Altomare, E., & Serviddio, G. (2012). The impact of interferon lambda 3 gene polymorphism on natural course and treatment of hepatitis C. Clinical & developmental immunology, 2012, 849373. [CrossRef]
- Bennett, A. L., Smith, D. W., Cummins, M. J., Jacoby, P. A., Cummins, J. M., & Beilharz, M. W. (2013). Low-dose oral interferon alpha as prophylaxis against viral respiratory illness: a double-blind, parallel controlled trial during an influenza pandemic year. Influenza and other respiratory viruses, 7(5), 854–862. [CrossRef]
- Bermejo-Jambrina, M., Blatzer, M., Jauregui-Onieva, P., Yordanov, T. E., Hörtnagl, P., Valovka, T., Huber, L. A., Wilflingseder, D., & Posch, W. (2020). CR4 Signaling Contributes to a DC-Driven Enhanced Immune Response Against Complement-Opsonized HIV-1. Frontiers in immunology, 11, 2010. [CrossRef]
- Bharaj, P. , Wang, Y. E., Dawes, B. E., Yun, T. E., Park, A., Yen, B., Basler, C. F., Freiberg, A. N., Lee, B., & Rajsbaum, R. (2016). The Matrix Protein of Nipah Virus Targets the E3-Ubiquitin Ligase TRIM6 to Inhibit the IKKε Kinase-Mediated Type-I IFN Antiviral Response. PLoS pathogens, 12(9), e1005880. [CrossRef]
- Bian, Q., Lu, J., Zhang, L., Chi, Y., Li, Y., & Guo, H. (2017). Highly pathogenic avian influenza A virus H5N1 non-structural protein 1 is associated with apoptotic activation of the intrinsic mitochondrial pathway. Experimental and therapeutic medicine, 14(5), 4041–4046. [CrossRef]
- Bigay, J., Le Grand, R., Martinon, F., & Maisonnasse, P. (2022). Vaccine-associated enhanced disease in humans and animal models: Lessons and challenges for vaccine development. Frontiers in microbiology, 13, 932408. [CrossRef]
- Birkhoff, M., Leitz, M., & Marx, D. (2009). Advantages of Intranasal Vaccination and Considerations on Device Selection. Indian Journal of Pharmaceutical Sciences, 71(6), 729–731.
- Birkhoff, M. Birkhoff, M., Leitz, M., & Marx, D. (2009). Advantages of Intranasal Vaccination and Considerations on Device Selection. Indian Journal of Pharmaceutical Sciences, 71(6), 729–731.
- Biron, C. A. (2016, February 12). Innate immunity: Recognizing and responding to foreign invaders-no training needed. Viral Pathogenesis (Third Edition). Retrieved October 22, 2022, from https://www.sciencedirect.com/science/article/pii/B9780128009642000045.
- Bittner, Z. A., Schrader, M., George, S. E., & Amann, R. (2022). Pyroptosis and Its Role in SARS-CoV-2 Infection. Cells, 11(10), 1717. [CrossRef]
- Blank, T., & Prinz, M. (2017). Type I interferon pathway in CNS homeostasis and neurological disorders. Glia, 65(9), 1397–1406. [CrossRef]
- Block, J. (2021). Vaccinating people who have had covid-19: why doesn’t natural immunity count in the US?. BMJ, n2101. [CrossRef]
- Bo, Z., Miao, Y., Xi, R., Zhong, Q., Bao, C., Chen, H., Sun, L., Qian, Y., Jung, Y. S., & Dai, J. (2020). PRV UL13 inhibits cGAS-STING-mediated IFN-β production by phosphorylating IRF3. Veterinary research, 51(1), 118. [CrossRef]
- Booth, L., Roberts, J. L., Ecroyd, H., Tritsch, S. R., Bavari, S., Reid, S. P., Proniuk, S., Zukiwski, A., Jacob, A., Sepúlveda, C. S., Giovannoni, F., García, C. C., Damonte, E., González-Gallego, J., Tuñón, M. J., & Dent, P. (2016). AR-12 Inhibits Multiple Chaperones Concomitant With Stimulating Autophagosome Formation Collectively Preventing Virus Replication. Journal of cellular physiology, 231(10), 2286–2302. [CrossRef]
- Borden, E.C. Interferons α and β in cancer: therapeutic opportunities from new insights. Nat Rev Drug Discov 18, 219–234 (2019). [CrossRef]
- Botek, M., Krejčí, J., Valenta, M., McKune, A., Sládečková, B., Konečný, P., Klimešová, I., & Pastucha, D. (2022). Molecular Hydrogen Positively Affects Physical and Respiratory Function in Acute Post-COVID-19 Patients: A New Perspective in Rehabilitation. International journal of environmental research and public health, 19(4), 1992. [CrossRef]
- Bournazos, S., Gupta, A., & Ravetch, J. V. (2020). The role of IgG Fc receptors in antibody-dependent enhancement. Nature reviews. Immunology, 20(10), 633–643. [CrossRef]
- Bousse, T., Chambers, R. L., Scroggs, R. A., Portner, A., & Takimoto, T. (2006). Human parainfluenza virus type 1 but not Sendai virus replicates in human respiratory cells despite IFN treatment. Virus research, 121(1), 23–32. [CrossRef]
- Bowie, A. G., & Unterholzner, L. (2008). Viral evasion and subversion of pattern-recognition receptor signalling. Nature reviews. Immunology, 8(12), 911–922. [CrossRef]
- Bracci, L., La Sorsa, V., Belardelli, F., & Proietti, E. (2008). Type I interferons as vaccine adjuvants against infectious diseases and cancer. Expert review of vaccines, 7(3), 373–381. [CrossRef]
- Bressy, C., Droby, G. N., Maldonado, B. D., Steuerwald, N., & Grdzelishvili, V. Z. (2019). Cell Cycle Arrest in G2/M Phase Enhances Replication of Interferon-Sensitive Cytoplasmic RNA Viruses via Inhibition of Antiviral Gene Expression. Journal of virology, 93(4), e01885-18. [CrossRef]
- Brisse, M., & Ly, H. (2019). Comparative Structure and Function Analysis of the RIG-I-Like Receptors: RIG-I and MDA5. Frontiers in immunology, 10, 1586. [CrossRef]
- Brochet B. (2008). Activité à long terme de l'acétate de glatiramère dans le traitement de la sclérose en plaques : état des connaissances [Long-term effects of glatiramer acetate in multiple sclerosis]. Revue neurologique, 164(11), 917–926. [CrossRef]
- Brown, M., & Bhardwaj, N. (2021). Super(antigen) target for SARS-CoV-2. Nature reviews. Immunology, 21(2), 72. [CrossRef]
- Brown, M., & Bhardwaj, N. (2021). Super(antigen) target for SARS-CoV-2. Nature reviews. Immunology, 21(2), 72. [CrossRef]
- Bruttel, V., Washburne, A., & VanDongen, A. (2022). Endonuclease fingerprint indicates a synthetic origin of SARS-COV-2. [CrossRef]
- Brzózka, K., Finke, S., & Conzelmann, K. K. (2005). Identification of the rabies virus alpha/beta interferon antagonist: phosphoprotein P interferes with phosphorylation of interferon regulatory factor 3. Journal of virology, 79(12), 7673–7681. [CrossRef]
- Burns K. H. (2020). Our Conflict with Transposable Elements and Its Implications for Human Disease. Annual review of pathology, 15, 51–70. [CrossRef]
- Busnadiego, I., Fernbach, S., Pohl, M. O., Karakus, U., Huber, M., Trkola, A., Stertz, S., & Hale, B. G. (2020). Antiviral Activity of Type I, II, and III Interferons Counterbalances ACE2 Inducibility and Restricts SARS-CoV-2. mBio, 11(5), e01928-20. [CrossRef]
- Busnadiego, I., Fernbach, S., Pohl, M. O., Karakus, U., Huber, M., Trkola, A., Stertz, S., & Hale, B. G. (2020). Antiviral Activity of Type I, II, and III Interferons Counterbalances ACE2 Inducibility and Restricts SARS-CoV-2. mBio, 11(5), e01928-20. [CrossRef]
- Busnadiego, I., Fernbach, S., Pohl, M. O., Karakus, U., Huber, M., Trkola, A., Stertz, S., & Hale, B. G. (2020). Antiviral Activity of Type I, II, and III Interferons Counterbalances ACE2 Inducibility and Restricts SARS-CoV-2. mBio, 11(5), e01928-20. [CrossRef]
- Buzhdygan, T. P., DeOre, B. J., Baldwin-Leclair, A., Bullock, T. A., McGary, H. M., Khan, J. A., Razmpour, R., Hale, J. F., Galie, P. A., Potula, R., Andrews, A. M., & Ramirez, S. H. (2020). The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood-brain barrier. Neurobiology of disease, 146, 105131. [CrossRef]
- Cabanillas, B., Akdis, C., & Novak, N. (2021). Allergic reactions to the first COVID-19 vaccine: A potential role of polyethylene glycol?. Allergy, 76(6), 1617-1618. [CrossRef]
- Cadegiani F. A. (2022). Catecholamines Are the Key Trigger of COVID-19 mRNA Vaccine-Induced Myocarditis: A Compelling Hypothesis Supported by Epidemiological, Anatomopathological, Molecular, and Physiological Findings. Cureus, 14(8), e27883. [CrossRef]
- Can Li, Yanxia Chen, Yan Zhao, David Christopher Lung, Zhanhong Ye, Wenchen Song, Fei-Fei Liu, Jian-Piao Cai, Wan-Man Wong, Cyril Chik-Yan Yip, Jasper Fuk-Woo Chan, Kelvin Kai-Wang To, Siddharth Sridhar, Ivan Fan-Ngai Hung, Hin Chu, Kin-Hang Kok, Dong-Yan Jin, Anna Jinxia Zhang, Kwok-Yung Yuen, Intravenous Injection of Coronavirus Disease 2019 (COVID-19) mRNA Vaccine Can Induce Acute Myopericarditis in Mouse Model, Clinical Infectious Diseases, 2021;, ciab707. [CrossRef]
- Cao, L., Ge, X., Gao, Y., Herrler, G., Ren, Y., Ren, X., & Li, G. (2015). Porcine epidemic diarrhea virus inhibits dsRNA-induced interferon-β production in porcine intestinal epithelial cells by blockade of the RIG-I-mediated pathway. Virology journal, 12, 127. [CrossRef]
- Carella, C., Mazziotti, G., Morisco, F., Manganella, G., Rotondi, M., Tuccillo, C., Sorvillo, F., Caporaso, N., & Amato, G. (2001). Long-term outcome of interferon-alpha-induced thyroid autoimmunity and prognostic influence of thyroid autoantibody pattern at the end of treatment. The Journal of clinical endocrinology and metabolism, 86(5), 1925–1929. [CrossRef]
- Carlos, A. J., Ha, D. P., Yeh, D. W., Van Krieken, R., Tseng, C. C., Zhang, P., Gill, P., Machida, K., & Lee, A. S. (2021). The chaperone GRP78 is a host auxiliary factor for SARS-CoV-2 and GRP78 depleting antibody blocks viral entry and infection. The Journal of biological chemistry, 296, 100759. [CrossRef]
- Carp, T. N. (2024). How the Next Pandemic Could Be Handled— Studying the Risks of (A)H5N1 Influenza Human Spillover. Preprints. [CrossRef]
- Carp, T. N. (2024). How the Next Pandemic Could Be Handled— Studying the Risks of (A)H5N1 Influenza Human Spillover. Preprints. [CrossRef]
- Carrero J. A. (2013). Confounding roles for type I interferons during bacterial and viral pathogenesis. International immunology, 25(12), 663–669. [CrossRef]
- Carvajal Ibañez, D., Skabkin, M., Hooli, J., Cerrizuela, S., Göpferich, M., Jolly, A.,... & Martin-Villalba, A. (2023). Interferon regulates neural stem cell function at all ages by orchestrating mTOR and cell cycle. EMBO Molecular Medicine, e16434. [CrossRef]
- Case, J. B., Ashbrook, A. W., Dermody, T. S., & Denison, M. R. (2016). Mutagenesis of S-Adenosyl-l-Methionine-Binding Residues in Coronavirus nsp14 N7-Methyltransferase Demonstrates Differing Requirements for Genome Translation and Resistance to Innate Immunity. Journal of virology, 90(16), 7248–7256. [CrossRef]
- Cassius, C., Amode, R., Delord, M., Battistella, M., Poirot, J., How-Kit, A., Lepelletier, C., Jachiet, M., de Masson, A., Frumholtz, L., Cordoliani, F., Boccara, D., Lehmann-Che, J., Wong, J., Dubanchet, S., Alberdi, A. J., Merandet, M., Bagot, M., Bensussan, A., Bouaziz, J. D., … Le Buanec, H. (2020). MDA5+ Dermatomyositis Is Associated with Stronger Skin Type I Interferon Transcriptomic Signature with Upregulation of IFN-κ Transcript. The Journal of investigative dermatology, 140(6), 1276–1279.e7. [CrossRef]
- Casteleyn, C., Broos, A. M., Simoens, P., & Van den Broeck, W. (2010). NALT (nasal cavity-associated lymphoid tissue) in the rabbit. Veterinary immunology and immunopathology, 133(2-4), 212–218. [CrossRef]
- Castro, L.S., Lobo, G.S., Pereira, P., Freire, M.G., Neves, M.C. and Pedro, A.Q., 2021. Interferon-based biopharmaceuticals: Overview on the production, purification, and formulation. Vaccines, 9(4), p.328. [CrossRef]
- Castro-Jiménez, T. K., Gómez-Legorreta, L. C., López-Campa, L. A., Martínez-Torres, V., Alvarado-Silva, M., Posadas-Mondragón, A., Díaz-Lima, N., Angulo-Mendez, H. A., Mejía-Domínguez, N. R., Vaca-Paniagua, F., Ávila-Moreno, F., García-Cordero, J., Cedillo-Barrón, L., Aguilar-Ruíz, S. R., & Bustos-Arriaga, J. (2022). Variability in Susceptibility to Type I Interferon Response and Subgenomic RNA Accumulation Between Clinical Isolates of Dengue and Zika Virus From Oaxaca Mexico Correlate With Replication Efficiency in Human Cells and Disease Severity. Frontiers in cellular and infection microbiology, 12, 890750. [CrossRef]
- Cataldi, M., Shah, N. R., Felt, S. A., & Grdzelishvili, V. Z. (2015). Breaking resistance of pancreatic cancer cells to an attenuated vesicular stomatitis virus through a novel activity of IKK inhibitor TPCA-1. Virology, 485, 340–354. [CrossRef]
- Cerezo, L., & Macià I Garau, M. (2012). Acute radiation syndrome and Fukushima: A watershed moment?. Reports of practical oncology and radiotherapy : journal of Greatpoland Cancer Center in Poznan and Polish Society of Radiation Oncology, 17(1), 1–3. [CrossRef]
- Cerpa-Cruz, S., Paredes-Casillas, P., Landeros Navarro, E. et al. Adverse events following immunization with vaccines containing adjuvants. Immunol Res 56, 299–303 (2013). [CrossRef]
- Cervantes, J., Weinerman, B., Basole, C. et al. TLR8: the forgotten relative revindicated. Cell Mol Immunol 9, 434–438 (2012). [CrossRef]
- Chan, R., Chan, K., Lui, G., Tsun, J., Chan, K., Yip, J., Liu, S., Yu, M., Ng, R., Chong, K., Wang, M. H., Chan, P., Li, A. M., & Lam, H. S. (2022). Mucosal Antibody Response to SARS-CoV-2 in Paediatric and Adult Patients: A Longitudinal Study. Pathogens (Basel, Switzerland), 11(4), 397. [CrossRef]
- Chattree, V., Singh, K., Singh, K., Goel, A., Maity, A., & Lone, A. (2022). A comprehensive review on modulation of SIRT1 signaling pathways in the immune system of COVID-19 patients by phytotherapeutic melatonin and epigallocatechin-3-gallate. Journal of food biochemistry, e14259. Advance online publication. [CrossRef]
- Chauss, D., Freiwald, T., McGregor, R., Yan, B., Wang, L., Nova-Lamperti, E., Kumar, D., Zhang, Z., Teague, H., West, E. E., Vannella, K. M., Ramos-Benitez, M. J., Bibby, J., Kelly, A., Malik, A., Freeman, A. F., Schwartz, D. M., Portilla, D., Chertow, D. S., John, S., … Afzali, B. (2022). Autocrine vitamin D signaling switches off pro-inflammatory programs of TH1 cells. Nature immunology, 23(1), 62–74. [CrossRef]
- Chen, A. et al. (2021) “Chitinase-3-like 1 protein complexes modulate macrophage-mediated immune suppression in glioblastoma,” Journal of Clinical Investigation, 131(16). [CrossRef]
- Chen, K., Liu, J., & Cao, X. (2017). Regulation of type I interferon signaling in immunity and inflammation: A comprehensive review. Journal of autoimmunity, 83, 1–11. [CrossRef]
- Chen, X., Jiang, L., & Liu, X. (2022). Natural killer cells: the next wave in cancer immunotherapy. Frontiers in immunology, 13, 954804. [CrossRef]
- Chen, Y., & Colonna, M. (2021). Microglia in Alzheimer's disease at single-cell level. Are there common patterns in humans and mice? The Journal of experimental medicine, 218(9), e20202717. [CrossRef]
- Chieux, V., Hober, D., Chehadeh, W., & Wattré, P. (1999). Interféron alpha, protéines antivirales et applications médicales [Alpha interferon, antiviral proteins and their value in clinical medicine]. Annales de biologie clinique, 57(6), 659–666.
- Childs, B., & van Deursen, J. (2019). Inhibition of 'jumping genes' promotes healthy ageing. Nature, 566(7742), 46–48. [CrossRef]
- Cho, C., , Mukherjee, R., Peck, A. R., Sun, Y., McBrearty, N., Katlinski, K. V., Gui, J., Govindaraju, P. K., Puré, E., Rui, H., & Fuchs, S. Y. (2020). Cancer-associated fibroblasts downregulate type I interferon receptor to stimulate intratumoral stromagenesis. Oncogene, 39(38), 6129–6137. [CrossRef]
- Cho, H. J., Hwang, J. A., Yang, E. J., Kim, E. C., Kim, J. R., Kim, S. Y., Kim, Y. Z., Park, S. C., & Lee, Y. S. (2022). Nintedanib induces senolytic effect via STAT3 inhibition. Cell death & disease, 13(9), 760. [CrossRef]
- Choi, H., & Shin, E. C. (2021). Roles of Type I and III Interferons in COVID-19. Yonsei medical journal, 62(5), 381–390. [CrossRef]
- Choo, O.-S., Lee, Y. Y., Kim, Y. S., Kim, Y. J., Lee, D. H., Kim, H., Jang, J. H., & Choung, Y.-H. (2022). Effect of statin on age-related hearing loss via drug repurposing. Biochimica Et Biophysica Acta (BBA) - Molecular Cell Research, 1869(11), 119331. [CrossRef]
- Choudhary, A. (2021). Immune regulatory function of interferon-gamma in acute leukemia. INDIAN JOURNAL OF APPLIED RESEARCH, 75–77. [CrossRef]
- Chung, J. H., Hong, S. H., Seo, N., Kim, T. S., An, H. J., Lee, P., Shin, E. C., & Kim, H. M. (2020). Structure-based glycoengineering of interferon lambda 4 enhances its productivity and anti-viral potency. Cytokine, 125, 154833. [CrossRef]
- Civril, F., Deimling, T., de Oliveira Mann, C. et al. Structural mechanism of cytosolic DNA sensing by cGAS. Nature 498, 332–337 (2013). [CrossRef]
- Claudio G., Gallo, Sirio Fiorino, Giovanni Posabella, Donato Antonacci, Antonio Tropeano, Emanuele Pausini, Carlotta Pausini, Tomasso Guarniero, Marco Zancanaro (2020), COVID-19.
- Corona, A., Fanunza, E., Salata, C., Morwitzer, M. J., Distinto, S., Zinzula, L., Sanna, C., Frau, A., Daino, G. L., Quartu, M., Taglialatela-Scafati, O., Rigano, D., Reid, S., Mirazimi, A., & Tramontano, E. (2022). Cynarin blocks Ebola virus replication by counteracting VP35 inhibition of interferon-beta production. Antiviral research, 198, 105251. [CrossRef]
- Corrales, L., McWhirter, S. M., Dubensky, T. W., Jr, & Gajewski, T. F. (2016). The host STING pathway at the interface of cancer and immunity. The Journal of clinical investigation, 126(7), 2404–2411. [CrossRef]
- Costa Silva, R., Bandeira-Melo, C., Paula Neto, H. A., Vale, A. M., & Travassos, L. H. (2022). COVID-19 diverse outcomes: Aggravated reinfection, type I interferons and antibodies. Medical hypotheses, 167, 110943. [CrossRef]
- Crow M. K. (2010). Long interspersed nuclear elements (LINE-1): potential triggers of systemic autoimmune disease. Autoimmunity, 43(1), 7–16. [CrossRef]
- Csépány, T. Csépány, T., & Bereczki, D. (2004). Immunmoduláns kezelés sclerosis multiplexben [Immunomodulatory therapy in multiple sclerosis]. Ideggyogyaszati szemle.
- Dacia Plant (2021), Interferonate tablets, doi: https://www.daciaplant.ro/interferonat-comprimate.html.
- Dagenais A, Villalba-Guerrero C and Olivier M (2023) Trained immunity: A “new” weapon in the fight against infectious diseases. Front. Immunol. 14:1147476. [CrossRef] [PubMed]
- Daniel B. Stetson and Rusian Medzhitov (2006), Type I Interferons in Host Defense, Immunity, Vol 25, pp 373-381. [CrossRef]
- Das, A., Dinh, P. X., Panda, D., & Pattnaik, A. K. (2014). Interferon-inducible protein IFI35 negatively regulates RIG-I antiviral signaling and supports vesicular stomatitis virus replication. Journal of virology, 88(6), 3103–3113. [CrossRef]
- Dasgupta, A. , Tsay, E., Federman, N., Lechner, M. G., & Su, M. A. (2022). Polyendocrine Autoimmunity and Diabetic Ketoacidosis Following Anti-PD-1 and Interferon α. Pediatrics, 149(4), e2021053363. [CrossRef]
- David Vremec, Meredith O'Keeffe, Hubertus Hochrein, Martina Fuchsberger, Irina Caminschi, Mireille Lahoud, Ken Shortman; Production of interferons by dendritic cells, plasmacytoid cells, natural killer cells, and interferon-producing killer dendritic cells. Blood 2007; 109 (3): 1165–1173. doi:. [CrossRef]
- David Vremec, Meredith O'Keeffe, Hubertus Hochrein, Martina Fuchsberger, Irina Caminschi, Mireille Lahoud, Ken Shortman; Production of interferons by dendritic cells, plasmacytoid cells, natural killer cells, and interferon-producing killer dendritic cells. Blood 2007; 109 (3): 1165–1173. doi:. [CrossRef]
- Davies, K. A., Cooper, E., Voon, V., Tibble, J., Cercignani, M., & Harrison, N. A. (2021). Interferon and anti-TNF therapies differentially modulate amygdala reactivity which predicts associated bidirectional changes in depressive symptoms. Molecular psychiatry, 26(9), 5150–5160. [CrossRef]
- Davoudi-Monfared, E., Rahmani, H., Khalili, H., Hajiabdolbaghi, M., Salehi, M., Abbasian, L., Kazemzadeh, H., & Yekaninejad, M. S. (2020). A Randomized Clinical Trial of the Efficacy and Safety of Interferon β-1a in Treatment of Severe COVID-19. Antimicrobial agents and chemotherapy, 64(9), e01061-20. [CrossRef]
- Daza-Cajigal, V., Albuquerque, A. S., Young, D. F., Ciancanelli, M. J., Moulding, D., Angulo, I., Jeanne-Julien, V., Rosain, J., Minskaia, E., Casanova, J. L., Boisson-Dupuis, S., Bustamante, J., Randall, R. E., McHugh, T. D., Thrasher, A. J., & Burns, S. O. (2022). Partial human Janus kinase 1 deficiency predominantly impairs responses to interferon gamma and intracellular control of mycobacteria. Frontiers in immunology, 13, 888427. [CrossRef]
- De Cecco, M., Ito, T., Petrashen, A. P., Elias, A. E., Skvir, N. J., Criscione, S. W., Caligiana, A., Brocculi, G., Adney, E. M., Boeke, J. D., Le, O., Beauséjour, C., Ambati, J., Ambati, K., Simon, M., Seluanov, A., Gorbunova, V., Slagboom, P. E., Helfand, S. L., Neretti, N., … Sedivy, J. M. (2019). L1 drives IFN in senescent cells and promotes age-associated inflammation. Nature, 566(7742), 73–78. [CrossRef]
- De Cecco, M., Ito, T., Petrashen, A.P. et al. Author Correction: L1 drives IFN in senescent cells and promotes age-associated inflammation. Nature 572, E5 (2019). [CrossRef]
- De Clercq, E., & Neyts, J. (2007). Avian influenza A (H5N1) infection: targets and strategies for chemotherapeutic intervention. Trends in pharmacological sciences, 28(6), 280–285. [CrossRef]
- de Groen, R. A., Groothuismink, Z. M., Liu, B. S., & Boonstra, A. (2015). IFN-λ is able to augment TLR-mediated activation and subsequent function of primary human B cells. Journal of leukocyte biology, 98(4), 623–630. [CrossRef]
- De Maeyer, E., & De Maeyer-Guignard, J. (1998). Type I interferons. International reviews of immunology, 17(1-4), 53–73. [CrossRef]
- de Witte, L. , Abt, M., Schneider-Schaulies, S., van Kooyk, Y., & Geijtenbeek, T. B. (2006). Measles virus targets DC-SIGN to enhance dendritic cell infection. Journal of virology, 80(7), 3477–3486. [CrossRef]
- Deczkowska, A., Baruch, K., & Schwartz, M. (2016). Type I/II Interferon Balance in the Regulation of Brain Physiology and Pathology. Trends in immunology, 37(3), 181–192. [CrossRef]
- DeDiego, M. L., Nogales, A., Lambert-Emo, K., Martinez-Sobrido, L., & Topham, D. J. (2016). NS1 Protein Mutation I64T Affects Interferon Responses and Virulence of Circulating H3N2 Human Influenza A Viruses. Journal of virology, 90(21), 9693–9711. [CrossRef]
- Dempoya, J., Matsumiya, T., Imaizumi, T., Hayakari, R., Xing, F., Yoshida, H., Okumura, K., & Satoh, K. (2012). Double-stranded RNA induces biphasic STAT1 phosphorylation by both type I interferon (IFN)-dependent and type I IFN-independent pathways. Journal of virology, 86(23), 12760–12769. [CrossRef]
- Deng, X. , Buckley, A. C., Pillatzki, A., Lager, K. M., Faaberg, K. S., & Baker, S. C. (2020). Inactivating Three Interferon Antagonists Attenuates Pathogenesis of an Enteric Coronavirus. Journal of virology, 94(17), e00565-20. [CrossRef]
- Desfarges, S. , Ciuffi, A. (2012). Viral Integration and Consequences on Host Gene Expression. In: Witzany, G. (eds) Viruses: Essential Agents of Life. Springer, Dordrecht. [CrossRef]
- Di Martino, J.S., Nobre, A.R., Mondal, C. et al. A tumor-derived type III collagen-rich ECM niche regulates tumor cell dormancy. Nat Cancer 3, 90–107 (2022). [CrossRef]
- Di Palma, F., Daino, G. L., Ramaswamy, V. K., Corona, A., Frau, A., Fanunza, E., Vargiu, A. V., Tramontano, E., & Ruggerone, P. (2019). Relevance of Ebola virus VP35 homo-dimerization on the type I interferon cascade inhibition. Antiviral chemistry & chemotherapy, 27, 2040206619889220. [CrossRef]
- Di Trolio, R. Di Trolio, R., Simeone, E., Di Lorenzo, G., Grimaldi, A. M., Romano, A., Ayala, F., Caracò, C., Mozzillo, N., & Ascierto, P. A. (2012). Update on PEG-interferon α-2b as adjuvant therapy in melanoma. Anticancer research, 3909. [Google Scholar]
- Diana A. Pippig, Johannes C. Hellmuth, Sheng Cui, Axel Kirchhofer, Katja Lammens, Alfred Lammens, Andreas Schmidt, Simon Rothenfusser, Karl-Peter Hopfner, The regulatory domain of the RIG-I family ATPase LGP2 senses double-stranded RNA, Nucleic Acids Research, Volume 37, Issue 6, 1 April 2009, Pages 2014–2025. [CrossRef]
- Dick, A., Graf, L., Olal, D., von der Malsburg, A., Gao, S., Kochs, G., & Daumke, O. (2015). Role of nucleotide binding and GTPase domain dimerization in dynamin-like myxovirus resistance protein A for GTPase activation and antiviral activity. The Journal of biological chemistry, 290(20), 12779–12792. [CrossRef]
- Didangelos A. (2020). COVID-19 Hyperinflammation: What about Neutrophils? mSphere, 5(3), e00367-20. [CrossRef]
- Ding, F., Yin, Z., & Wang, H. R. (2011). Ubiquitination in Rho signaling. Current topics in medicinal chemistry, 11(23), 2879–2887. [CrossRef]
- Ding, J., Aldo, P., Roberts, C. M., Stabach, P., Liu, H., You, Y., Qiu, X., Jeong, J., Maxwell, A., Lindenbach, B., Braddock, D., Liao, A., & Mor, G. (2021). Placenta-derived interferon-stimulated gene 20 controls ZIKA virus infection. EMBO reports, 22(10), e52450. [CrossRef]
- Dipasquale, O. , Cooper, E. A., Tibble, J., Voon, V., Baglio, F., Baselli, G., Cercignani, M., & Harrison, N. A. (2016). Interferon-α acutely impairs whole-brain functional connectivity network architecture - A preliminary study. Brain, behavior, and immunity, 58, 31–39. [CrossRef]
- Doerfler W. (2021). Adenoviral Vector DNA- and SARS-CoV-2 mRNA-Based Covid-19 Vaccines: Possible Integration into the Human Genome - Are Adenoviral Genes Expressed in Vector-based Vaccines? Virus research, 302, 198466. [CrossRef]
- Dohlman, A. B., Klug, J., Mesko, M., Gao, I. H., Lipkin, S. M., Shen, X., & Iliev, I. D. (2022). A pan-cancer mycobiome analysis reveals fungal involvement in gastrointestinal and lung tumors. Cell, 185(20), 3807–3822. [CrossRef]
- Doi, A., Iijima, K., Kano, S., & Ishizaka, Y. (2015). Viral protein R of HIV type-1 induces retrotransposition and upregulates glutamate synthesis by the signal transducer and activator of transcription 1 signaling pathway. Microbiology and immunology, 59(7), 398–409. [CrossRef]
- Dong, L. W., Kong, X. N., Yan, H. X., Yu, L. X., Chen, L., Yang, W., Liu, Q., Huang, D. D., Wu, M. C., & Wang, H. Y. (2008). Signal regulatory protein alpha negatively regulates both TLR3 and cytoplasmic pathways in type I interferon induction. Molecular immunology, 45(11), 3025–3035. [CrossRef]
- Durmaz, V., Köchl, K., Krassnigg, A., Parigger, L., Hetmann, M., Singh, A., Nutz, D., Korsunsky, A., Kahler, U., König, C., Chang, L., Krebs, M., Bassetto, R., Pavkov-Keller, T., Resch, V., Gruber, K., Steinkellner, G., & Gruber, C. C. (2022). Structural bioinformatics analysis of SARS-CoV-2 variants reveals higher hACE2 receptor binding affinity for Omicron B.1.1.529 spike RBD compared to wild type reference. Scientific reports, 12(1), 14534. [CrossRef]
- Dutta, D. Dutta, D., Liu, J., & Xiong, H. (2022). NLRP3 inflammasome activation and SARS-CoV-2-mediated hyperinflammation, cytokine storm and neurological syndromes. International journal of physiology, pathophysiology and pharmacology, 14(3), 138–160.
- Edahiro, Y. , Ohishi, K., Gotoh, A., Takenaka, K., Shibayama, H., Shimizu, T., Usuki, K., Shimoda, K., Ito, M., VanWart, S. A., Zagrijtschuk, O., Qin, A., Kawase, H., Miyachi, N., Sato, T., Komatsu, N., & Kirito, K. (2022). Efficacy and safety of ropeginterferon alfa-2b in Japanese patients with polycythemia vera: an open-label, single-arm, phase 2 study. International journal of hematology, 116(2), 215–227. [CrossRef]
- Edahiro, Y., Yasuda, H., Gotoh, A., Morishita, S., Suzuki, T., Takeda, J., Ando, J., Tsutsui, M., Itakura, A., & Komatsu, N. (2021). Interferon therapy for pregnant patients with essential thrombocythemia in Japan. International journal of hematology, 113(1), 106–111. [CrossRef]
- Effat Davoudi-Monfared, Hamid Rahmani, Hossein Khalili, Mahboubeh Hajiabdolbaghi, Mohamadreza Salehi, Ladan Abbasian, Hossein Kazemzadeh, Mir Saeed Yekaninejad (2020), A Randomized Clinical Trial of the Efficacy and Safety of Interferon Beta-1a in Treatment of Severe COVID-19, ASM Journals, Vol 64, 9. [CrossRef]
- Ehrhardt, C., Wolff, T., Pleschka, S., Planz, O., Beermann, W., Bode, J. G., Schmolke, M., & Ludwig, S. (2007). Influenza A virus NS1 protein activates the PI3K/Akt pathway to mediate antiapoptotic signaling responses. Journal of virology, 81(7), 3058–3067. [CrossRef]
- Ejlerskov, P., Hultberg, J. G., Wang, J., Carlsson, R., Ambjørn, M., Kuss, M., Liu, Y., Porcu, G., Kolkova, K., Friis Rundsten, C., Ruscher, K., Pakkenberg, B., Goldmann, T., Loreth, D., Prinz, M., Rubinsztein, D. C., & Issazadeh-Navikas, S. (2015). Lack of Neuronal IFN-β-IFNAR Causes Lewy Body- and Parkinson's Disease-like Dementia. Cell, 163(2), 324–339. [CrossRef]
- Elkon, K. B., & Santer, D. M. (2012). Complement, interferon and lupus. Current opinion in immunology, 24(6), 665–670. [CrossRef]
- Emiliani, Yuliana and Muzi, Gabriel and Sanchez, Andres and Munera, Marlon, Prediction of Molecular Mimicry between Proteins from Trypanosoma Sp. And Human Antigens Associated with Systemic Lupus Erythematosus. Available at SSRN: https://ssrn.com/abstract=4184590 or. [CrossRef]
- Emilie, D., Burgard, M., Lascoux-Combe, C., Laughlin, M., Krzysiek, R., Pignon, C., Rudent, A., Molina, J. M., Livrozet, J. M., Souala, F., Chene, G., Grangeot-Keros, L., Galanaud, P., Sereni, D., Rouzioux, C., & Primoferon A Study Group (2001). Early control of HIV replication in primary HIV-1 infection treated with antiretroviral drugs and pegylated IFN alpha: results from the Primoferon A (ANRS 086) Study. AIDS (London, England), 15(11), 1435–1437. [CrossRef]
- Emily Mantlo, Natalya Bukreyeva, Junki Maruyama, et al. (2020), Antiviral Activities of Type I interferons to SARS-CoV-2 Infection, Journal of Antiviral Resistance, Vol. 178, 104811, doi:. [CrossRef]
- Ezelle, H. J., Balachandran, S., Sicheri, F., Polyak, S. J., & Barber, G. N. (2001). Analyzing the mechanisms of interferon-induced apoptosis using CrmA and hepatitis C virus NS5A. Virology, 281(1), 124–137. [CrossRef]
- Faleiro RJ, Kumar R, Bunn PT, Singh N, Chauhan SB, et al. (2016) Combined Immune Therapy for the Treatment of Visceral Leishmaniasis. PLOS Neglected Tropical Diseases 10(2): e0004415. [CrossRef]
- Fanunza, E., Frau, A., Corona, A., & Tramontano, E. (2019). Insights into Ebola Virus VP35 and VP24 Interferon Inhibitory Functions and their Initial Exploitation as Drug Targets. Infectious disorders drug targets, 19(4), 362–374. [CrossRef]
- Farinholt, T. , Doddapaneni, H., Qin, X., Menon, V., Meng, Q., Metcalf, G., Chao, H., Gingras, M.C., Avadhanula, V., Farinholt, P., Agrawal, C., Muzny, D.M., Piedra, P.A., Gibbs, R.A., & Petrosino, J. (2021) Transmission event of SARS-CoV-2 delta variant reveals multiple vaccine breakthrough infections. BMC Med, 19(1): 255. [CrossRef]
- Farooq, M. , Khan, A. W., Ahmad, B., Kim, M. S., & Choi, S. (2022). Therapeutic Targeting of Innate Immune Receptors Against SARS-CoV-2 Infection. Frontiers in pharmacology, 13, 915565. [CrossRef]
- Fatemi Naeini, F. , Yazdanpanah, M., Mohaghegh, F., Rajabi, P., & Tabatabaei, E. T. (2022). Interferon α-induced lupus-like reaction in a mycosis fungoides patient: A case report. International journal of clinical pharmacology and therapeutics, 60(7), 306–310. [CrossRef]
- Fazeli, M. R., & Hezarjaribi, N. (2022). A Simplified Process for Purification and Refolding of Recombinant Human Interferon-α2b. Iranian biomedical journal, 26(1), 85–90. [CrossRef]
- Fehniger, T. A., Herbein, G., Yu, H., Para, M. I., Bernstein, Z. P., O'Brien, W. A., & Caligiuri, M. A. (1998). Natural killer cells from HIV-1+ patients produce C-C chemokines and inhibit HIV-1 infection. Journal of immunology (Baltimore, Md. : 1950), 161(11), 6433–6438.
- Felgenhauer, U., Schoen, A., Gad, H. H., Hartmann, R., Schaubmar, A. R., Failing, K., Drosten, C., & Weber, F. (2020). Inhibition of SARS-CoV-2 by type I and type III interferons. The Journal of biological chemistry, 295(41), 13958–13964. [CrossRef]
- Felgenhauer, U. , Schoen, A., Gad, H. H., Hartmann, R., Schaubmar, A. R., Failing, K., Drosten, C., & Weber, F. (2020). Inhibition of SARS-CoV-2 by type I and type III interferons. The Journal of biological chemistry, 295(41), 13958–13964. [CrossRef]
- Felgenhauer, U., Schoen, A., Gad, H. H., Hartmann, R., Schaubmar, A. R., Failing, K., Drosten, C., & Weber, F. (2020). Inhibition of SARS-CoV-2 by type I and type III interferons. The Journal of biological chemistry, 295(41), 13958–13964. [CrossRef]
- Feng, E., Balint, E., Poznanski, S. M., Ashkar, A. A., & Loeb, M. (2021). Aging and Interferons: Impacts on Inflammation and Viral Disease Outcomes. Cells, 10(3), 708. [CrossRef]
- Fenton-May, A. E., Dibben, O., Emmerich, T., Ding, H., Pfafferott, K., Aasa-Chapman, M. M., Pellegrino, P., Williams, I., Cohen, M. S., Gao, F., Shaw, G. M., Hahn, B. H., Ochsenbauer, C., Kappes, J. C., & Borrow, P. (2013). Relative resistance of HIV-1 founder viruses to control by interferon-alpha. Retrovirology, 10, 146. [CrossRef]
- Ferran, M. C., & Lucas-Lenard, J. M. (1997). The vesicular stomatitis virus matrix protein inhibits transcription from the human beta interferon promoter. Journal of virology, 71(1), 371–377. [CrossRef]
- Fitzgerald-Bocarsly, P. , & Feng, D. (2007). The role of type I interferon production by dendritic cells in host defense. Biochimie, 89(6-7), 843–855. [CrossRef]
- Flachenecker P. (2004). Disease-modifying drugs for the early treatment of multiple sclerosis. Expert review of neurotherapeutics, 4(3), 455–463. [CrossRef]
- Fleming, D; A Scientific and Forensic investigation: R.M. (2021). Is COVID-19 a Bioweapon? ISBN 978-1-5107-7019-5.
- Flemming A. (2022). Omicron, the great escape artist. Nature reviews. Immunology, 22(2), 75. [CrossRef]
- Francis, M. L., Meltzer, M. S., & Gendelman, H. E. (1992). Interferons in the persistence, pathogenesis, and treatment of HIV infection. AIDS research and human retroviruses, 8(2), 199–207. [CrossRef]
- Frieman, M., Yount, B., Heise, M., Kopecky-Bromberg, S. A., Palese, P., & Baric, R. S. (2007). Severe acute respiratory syndrome coronavirus ORF6 antagonizes STAT1 function by sequestering nuclear import factors on the rough endoplasmic reticulum/Golgi membrane. Journal of virology, 81(18), 9812–9824. [CrossRef]
- Frisch, S. M., & MacFawn, I. P. (2020). Type I interferons and related pathways in cell senescence. Aging cell, 19(10), e13234. [CrossRef]
- Frumholtz, L. , Bouaziz, J. D., Battistella, M., Hadjadj, J., Chocron, R., Bengoufa, D., Le Buanec, H., Barnabei, L., Meynier, S., Schwartz, O., Grzelak, L., Smith, N., Charbit, B., Duffy, D., Yatim, N., Calugareanu, A., Philippe, A., Guerin, C. L., Joly, B., Siguret, V., … Saint-Louis CORE (COvid REsearch) (2021). Type I interferon response and vascular alteration in chilblain-like lesions during the COVID-19 outbreak. The British journal of dermatology, 185(6), 1176–1185. [CrossRef]
- Fukuda, Y., Homma, T., Inoue, H., Goto, Y., Sato, Y., Ikeda, H., Onitsuka, C., Sato, H., Akimoto, K., Ebato, T., Suganuma, H., Kawahara, T., Mikuni, H., Uchida, Y., Suzuki, S., Tanaka, A., & Sagara, H. (2022). Serum IL-28A/IFN-λ2 is linked to disease severity of COVID-19. Scientific reports, 12(1), 5458. [CrossRef]
- Fukuda, Y., Homma, T., Inoue, H., Onitsuka, C., Ikeda, H., Goto, Y., Sato, Y., Kimura, T., Hirai, K., Ohta, S., Yamamoto, M., Kusumoto, S., Suzuki, S., Tanaka, A., & Sagara, H. (2021). Downregulation of type III interferons in patients with severe COVID-19. Journal of medical virology, 93(7), 4559–4563. [CrossRef]
- Fukuda, Y. , Homma, T., Inoue, H., Onitsuka, C., Ikeda, H., Goto, Y., Sato, Y., Kimura, T., Hirai, K., Ohta, S., Yamamoto, M., Kusumoto, S., Suzuki, S., Tanaka, A., & Sagara, H. (2021). Downregulation of type III interferons in patients with severe COVID-19. Journal of medical virology, 93(7), 4559–4563. [CrossRef]
- Galão, R. P., Wilson, H., Schierhorn, K. L., Debeljak, F., Bodmer, B. S., Goldhill, D., Hoenen, T., Wilson, S. J., Swanson, C. M., & Neil, S. (2022). TRIM25 and ZAP target the Ebola virus ribonucleoprotein complex to mediate interferon-induced restriction. PLoS pathogens, 18(5), e1010530. [CrossRef]
- Gal-Ben-Ari, S., Barrera, I., Ehrlich, M., & Rosenblum, K. (2019). PKR: a kinase to remember. Frontiers in molecular neuroscience, 11, 480.
- Gamdzyk, M., Doycheva, D. M., Araujo, C., Ocak, U., Luo, Y., Tang, J., & Zhang, J. H. (2020). cGAS/STING Pathway Activation Contributes to Delayed Neurodegeneration in Neonatal Hypoxia-Ischemia Rat Model: Possible Involvement of LINE-1. Molecular neurobiology, 57(6), 2600–2619. [CrossRef]
- Gao, L., Yu, S., Chen, Q., Duan, Z., Zhou, J., Mao, C., Yu, D., Zhu, W., Nie, J., & Hou, Y. (2010). A randomized controlled trial of low-dose recombinant human interferons alpha-2b nasal spray to prevent acute viral respiratory infections in military recruits. Vaccine, 28(28), 4445–4451. [CrossRef]
- Gardinassi, L. G., Souza, C., Sales-Campos, H., & Fonseca, S. G. (2020). Immune and Metabolic Signatures of COVID-19 Revealed by Transcriptomics Data Reuse. Frontiers in immunology, 11, 1636. [CrossRef]
- Garg, A. D., & Agostinis, P. (2017). Cell death and immunity in cancer: From danger signals to mimicry of pathogen defense responses. Immunological reviews, 280(1), 126–148. [CrossRef]
- Gargan, S., Ahmed, S., Mahony, R., Bannan, C., Napoletano, S., O'Farrelly, C., Borrow, P., Bergin, C., & Stevenson, N. J. (2018). HIV-1 Promotes the Degradation of Components of the Type 1 IFN JAK/STAT Pathway and Blocks Anti-viral ISG Induction. EBioMedicine, 30, 203–216. [CrossRef]
- Gargan, S., Ahmed, S., Mahony, R., Bannan, C., Napoletano, S., O'Farrelly, C., Borrow, P., Bergin, C., & Stevenson, N. J. (2018). HIV-1 Promotes the Degradation of Components of the Type 1 IFN JAK/STAT Pathway and Blocks Anti-viral ISG Induction. EBioMedicine, 30, 203–216. [CrossRef]
- Gargano, J. W., Wallace, M., Hadler, S.C., Langley, G., Su, J.R., Oster, M.E., Broder, K.R., Gee, J., Weintraub, E., Shimabukuro, T., Scobie, H.M., Moulia, D., Markowitz, L.E., Wharton, M., McNally, V.V., Romero, J.R., Talbot, H.K., Lee, G.M., Daley, M.F., & Oliver, S.E. (2021). Use of mRNA COVID-19 Vaccine After Reports of Myocarditis Among Vaccine Recipients: Update from the Advisory Committee on Immunization Practices - United States, MMWR Morb Mortal Wkly Rep, 70(27): 977-982. [CrossRef]
- Gartlan, C., Tipton, T., Salguero, F. J., Sattentau, Q., Gorringe, A., & Carroll, M. W. (2022). Vaccine-Associated Enhanced Disease and Pathogenic Human Coronaviruses. Frontiers in immunology, 13, 882972. [CrossRef]
- Gaudieri, S., Lucas, M., Lucas, A., McKinnon, E., Albloushi, H., Rauch, A., di Iulio, J., Martino, D., Prescott, S. L., & Tulic, M. K. (2012). Genetic variations in IL28B and allergic disease in children. PloS one, 7(1), e30607. [CrossRef]
- George J and Mattapallil JJ (2018), Interferon-α Subtypes As an Adjunct Therapeutic Approach for Human Immunodeficiency Virus Functional Cure. Front. Immunol. 9, 299. [CrossRef] [PubMed]
- Ghasemitarei, M., Privat-Maldonado, A., Yusupov, M., Rahnama, S., Bogaerts, A., & Ejtehadi, M. R. (2022). Effect of Cysteine Oxidation in SARS-CoV-2 Receptor-Binding Domain on Its Interaction with Two Cell Receptors: Insights from Atomistic Simulations. Journal of chemical information and modeling, 62(1), 129–141. [CrossRef]
- Gigante, M. , Mandic, M., Wesa, A. K., Cavalcanti, E., Dambrosio, M., Mancini, V., Battaglia, M., Gesualdo, L., Storkus, W. J., & Ranieri, E. (2008). Interferon-alpha (IFN-alpha)-conditioned DC preferentially stimulate type-1 and limit Treg-type in vitro T-cell responses from RCC patients. Journal of immunotherapy (Hagerstown, Md. : 1997), 31(3), 254–262. [CrossRef]
- Gil, J., & Esteban, M. (2000). The interferon-induced protein kinase (PKR), triggers apoptosis through FADD-mediated activation of caspase 8 in a manner independent of Fas and TNF-alpha receptors. Oncogene, 19(32), 3665–3674. [CrossRef]
- Gilbert, C., Lefeuvre, C., Preisser, L., Pivert, A., Soleti, R., Blanchard, S., Delneste, Y., Ducancelle, A., Couez, D., & Jeannin, P. (2021). Age-Related Expression of IFN-λ1 Versus IFN-I and Beta-Defensins in the Nasopharynx of SARS-CoV-2-Infected Individuals. Frontiers in immunology, 12, 750279. [CrossRef]
- Gilbert, C., Lefeuvre, C., Preisser, L., Pivert, A., Soleti, R., Blanchard, S., Delneste, Y., Ducancelle, A., Couez, D., & Jeannin, P. (2021). Age-Related Expression of IFN-λ1 Versus IFN-I and Beta-Defensins in the Nasopharynx of SARS-CoV-2-Infected Individuals. Frontiers in immunology, 12, 750279. [CrossRef]
- Gill, N., Deacon, P. M., Lichty, B., Mossman, K. L., & Ashkar, A. A. (2006). Induction of innate immunity against herpes simplex virus type 2 infection via local delivery of Toll-like receptor ligands correlates with beta interferon production. Journal of virology, 80(20), 9943–9950. [CrossRef]
- Gina Kolata, “Ignored AIDS Drug Shows Promise in Small Tests,” New York Times, August 15, 1989; //www: https.
- Giotis, E. S., Robey, R. C., Skinner, N. G., Tomlinson, C. D., Goodbourn, S., & Skinner, M. A. (2016). Chicken interferome: avian interferon-stimulated genes identified by microarray and RNA-seq of primary chick embryo fibroblasts treated with a chicken type I interferon (IFN-α). Veterinary research, 47(1), 75. [CrossRef]
- Glennon, N. B., Jabado, O., Lo, M. K., & Shaw, M. L. (2015). Transcriptome Profiling of the Virus-Induced Innate Immune Response in Pteropus vampyrus and Its Attenuation by Nipah Virus Interferon Antagonist Functions. Journal of virology, 89(15), 7550–7566. [CrossRef]
- Glennon-Alty, L., Moots, R. J., Edwards, S. W., & Wright, H. L. (2021). Type I interferon regulates cytokine-delayed neutrophil apoptosis, reactive oxygen species production and chemokine expression. Clinical and experimental immunology, 203(2), 151–159. [CrossRef]
- Goh, J., & Behringer, M. (2018). Exercise alarms the immune system: A HMGB1 perspective. Cytokine, 110, 222–225. [CrossRef]
- Goldman, S. , Bron, D., Tousseyn, T., Vierasu, I., Dewispelaere, L., Heimann, P., Cogan, E., & Goldman, M. (2021). Rapid progression of angioimmunoblastic T Cell lymphoma following BNT162b2 mRNA vaccine booster shot: a case report. Frontiers in Medicine, 8, 798095. [CrossRef]
- Gorlé, N., & Vandenbroucke, R. E. (2019). Interferons: A molecular switch between damage and repair in ageing and Alzheimer's disease. Mechanisms of ageing and development, 183, 111148. [CrossRef]
- Govek, E. E., Hatten, M. E., & Van Aelst, L. (2011). The role of Rho GTPase proteins in CNS neuronal migration. Developmental neurobiology, 71(6), 528–553. [CrossRef]
- Grant, A. H., Estrada, A., 3rd, Ayala-Marin, Y. M., Alvidrez-Camacho, A. Y., Rodriguez, G., Robles-Escajeda, E., Cadena-Medina, D. A., Rodriguez, A. C., & Kirken, R. A. (2021). The Many Faces of JAKs and STATs Within the COVID-19 Storm. Frontiers in immunology, 12, 690477. [CrossRef]
- Grant, A., Ponia, S. S., Tripathi, S., Balasubramaniam, V., Miorin, L., Sourisseau, M., Schwarz, M. C., Sánchez-Seco, M. P., Evans, M. J., Best, S. M., & García-Sastre, A. (2016). Zika Virus Targets Human STAT2 to Inhibit Type I Interferon Signaling. Cell host & microbe, 19(6), 882–890. [CrossRef]
- Grenfell, B. T., Pybus, O.G., Gog, J.R., Wood, J.L., Daly, J.M., Mumford, J.A., & Holmes, E.C. (2004). Unifying the epidemiological and evolutionary dynamics of pathogens. Science, 303(5656): 327-332. [CrossRef]
- Grifoni, A. , Weiskopf, D., Ramirez, S.I., Mateus, J., Dan, J.M., Moderbacher, C.R., Rawlings, S.A., Sutherland, A., Premkumar, L., Jadi, R.S., Marrama, D., de Silva, A.M., Frazier, A., Carlin, A.F., Greenbaum, J.A., Peters, B., Krammer, F., Smith, D.M., Crotty, S., & Sette, A. (2020). Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell, 181(7):1489-1501.e15. [CrossRef]
- Guo, Y. , Yang, C., Liu, Y., Li, T., Li, H., Han, J., Jia, L., Wang, X., Zhang, B., Li, J., & Li, L. (2022). High Expression of HERV-K (HML-2) Might Stimulate Interferon in COVID-19 Patients. Viruses, 14(5), 996. [CrossRef]
- Gupta, S., Nakabo, S., Chu, J., Hasni, S., & Kaplan, M. J. (2020). Association between anti-interferon-alpha autoantibodies and COVID-19 in systemic lupus erythematosus. medRxiv : the preprint server for health sciences, 2020.10.29.20222000. [CrossRef]
- Haasbach, E., Droebner, K., Vogel, A. B., & Planz, O. (2011). Low-dose interferon Type I treatment is effective against H5N1 and swine-origin H1N1 influenza A viruses in vitro and in vivo. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research, 31(6), 515–525. [CrossRef]
- Habjan, M., Andersson, I., Klingström, J., Schümann, M., Martin, A., Zimmermann, P., Wagner, V., Pichlmair, A., Schneider, U., Mühlberger, E., Mirazimi, A., & Weber, F. (2008). Processing of genome 5' termini as a strategy of negative-strand RNA viruses to avoid RIG-I-dependent interferon induction. PloS one, 3(4), e2032. [CrossRef]
- Hackstadt, T., Chiramel, A. I., Hoyt, F. H., Williamson, B. N., Dooley, C. A., Beare, P. A., de Wit, E., Best, S. M., & Fischer, E. R. (2021). Disruption of the Golgi Apparatus and Contribution of the Endoplasmic Reticulum to the SARS-CoV-2 Replication Complex. Viruses, 13(9), 1798. [CrossRef]
- Haga, R. B., & Ridley, A. J. (2016). Rho GTPases: Regulation and roles in cancer cell biology. Small GTPases, 7(4), 207–221. [CrossRef]
- Hakami A. R. (2022). Targeting the RBD of Omicron Variant (B.1.1.529) with Medicinal Phytocompounds to Abrogate the Binding of Spike Glycoprotein with the hACE2 Using Computational Molecular Search and Simulation Approach. Biology, 11(2), 258. [CrossRef]
- Haller O. (2015). A tribute to Jean Lindenmann, co-discoverer of interferon (1924-2015). Cytokine, 76(1), 113–115. [CrossRef]
- Haller, O., & Kochs, G. (2011). Human MxA protein: an interferon-induced dynamin-like GTPase with broad antiviral activity. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research, 31(1), 79–87. [CrossRef]
- Haller, O., Gao, S., von der Malsburg, A., Daumke, O., & Kochs, G. (2010). Dynamin-like MxA GTPase: structural insights into oligomerization and implications for antiviral activity. The Journal of biological chemistry, 285(37), 28419–28424. [CrossRef]
- Halstead S. B. (2021). Vaccine-Associated Enhanced Viral Disease: Implications for Viral Vaccine Development. BioDrugs : clinical immunotherapeutics, biopharmaceuticals and gene therapy, 35(5), 505–515. [CrossRef]
- Han, L. , Zheng, Y., Deng, J., Nan, M. L., Xiao, Y., Zhuang, M. W., Zhang, J., Wang, W., Gao, C., & Wang, P. H. (2022). SARS-CoV-2 ORF10 antagonizes STING-dependent interferon activation and autophagy. Journal of medical virology, 10.1002/jmv.27965. Advance online publication. [CrossRef]
- Han, L. , Zhuang, M. W., Deng, J., Zheng, Y., Zhang, J., Nan, M. L., Zhang, X. J., Gao, C., & Wang, P. H. (2021). SARS-CoV-2 ORF9b antagonizes type I and III interferons by targeting multiple components of the RIG-I/MDA-5-MAVS, TLR3-TRIF, and cGAS-STING signaling pathways. Journal of medical virology, 93(9), 5376–5389. [CrossRef]
- Harandi A. M. (2004). The potential of immunostimulatory CpG DNA for inducing immunity against genital herpes: opportunities and challenges. Journal of clinical virology : the official publication of the Pan American Society for Clinical Virology, 30(3), 207–210. [CrossRef]
- Harman, A. N., Nasr, N., Feetham, A., Galoyan, A., Alshehri, A. A., Rambukwelle, D., Botting, R. A., Hiener, B. M., Diefenbach, E., Diefenbach, R. J., Kim, M., Mansell, A., & Cunningham, A. L. (2015). HIV Blocks Interferon Induction in Human Dendritic Cells and Macrophages by Dysregulation of TBK1. Journal of virology, 89(13), 6575–6584. [CrossRef]
- Hayden, F. G., & Gwaltney, J. M., Jr (1984). Intranasal interferon-alpha 2 treatment of experimental rhinoviral colds. The Journal of infectious diseases, 150(2), 174–180. [CrossRef]
- Hayden, F. G., Kaiser, D. L., & Albrecht, J. K. (1988). Intranasal recombinant alfa-2b interferon treatment of naturally occurring common colds. Antimicrobial agents and chemotherapy, 32(2), 224–230. [CrossRef]
- He, B. , Tran, J. T., & Sanchez, D. J. (2019). Manipulation of Type I Interferon Signaling by HIV and AIDS-Associated Viruses. Journal of immunology research, 2019, 8685312. [CrossRef]
- He, Z. , Tian, M., & Fu, X. (2021). Reduced expression of miR-30c-5p promotes hepatocellular carcinoma progression by targeting RAB32. Molecular therapy. Nucleic acids, 26, 603–612. [CrossRef]
- Hefti, H. P., Frese, M., Landis, H., Di Paolo, C., Aguzzi, A., Haller, O., & Pavlovic, J. (1999). Human MxA protein protects mice lacking a functional alpha/beta interferon system against La crosse virus and other lethal viral infections. Journal of virology, 73(8), 6984–6991. [CrossRef]
- Herder, V. , Dee, K., Wojtus, J. K., Epifano, I., Goldfarb, D., Rozario, C., Gu, Q., Da Silva Filipe, A., Nomikou, K., Nichols, J., Jarrett, R. F., Stevenson, A., McFarlane, S., Stewart, M. E., Szemiel, A. M., Pinto, R. M., Masdefiol Garriga, A., Davis, C., Allan, J., Graham, S. V., … Boutell, C. (2021). Elevated temperature inhibits SARS-CoV-2 replication in respiratory epithelium independently of IFN-mediated innate immune defenses. PLoS biology, 19(12), e3001065. [CrossRef]
- Higgins, P. G., Al-Nakib, W., Willman, J., & Tyrrell, D. A. (1986). Interferon-beta ser as prophylaxis against experimental rhinovirus infection in volunteers. Journal of interferon research, 6(2), 153–159. [CrossRef]
- Hijano, D. R., Vu, L. D., Kauvar, L. M., Tripp, R. A., Polack, F. P., & Cormier, S. A. (2019). Role of Type I Interferon (IFN) in the Respiratory Syncytial Virus (RSV) Immune Response and Disease Severity. Frontiers in immunology, 10, 566. [CrossRef]
- Hirose, S. , Hara, M., Koda, K., Natori, N., Yokota, Y., Ninomiya, S., & Nakajima, H. (2021). Acute autoimmune transverse myelitis following COVID-19 vaccination: A case report. Medicine (Baltimore). 100(51): e28423. [CrossRef]
- Ho, M.-W. Ho, M.-W. (1999). Genetic Engineering, Dream or Nightmare? The Brave New World of Bad Science and Big Business, 2nd ed. Gateway Books, Dublin (IRL). 8264. [Google Scholar]
- Ho, M. -W. (2013). The new genetics and natural versus artificial genetic modification. Entropy, 15(11), 4748–4781. [CrossRef]
- Hoffman, R. M., & Han, Q. (2020). Oral Methioninase for Covid-19 Methionine-restriction Therapy. In vivo (Athens, Greece), 34(3 Suppl), 1593–1596. [CrossRef]
- Hoffman, R. M., & Han, Q. (2020). Oral Methioninase for Covid-19 Methionine-restriction Therapy. In vivo (Athens, Greece), 34(3 Suppl), 1593–1596. [CrossRef]
- Hossain, A., Akter, S., Rashid, A. A., Khair, S., & Alam, A. (2022). Unique mutations in SARS-CoV-2 Omicron subvariants' non-spike proteins: Potential impacts on viral pathogenesis and host immune evasion. Microbial pathogenesis, 170, 105699. [CrossRef]
- Hotter, D., Bosso, M., Jønsson, K. L., Krapp, C., Stürzel, C. M., Das, A., Littwitz-Salomon, E., Berkhout, B., Russ, A., Wittmann, S., Gramberg, T., Zheng, Y., Martins, L. J., Planelles, V., Jakobsen, M. R., Hahn, B. H., Dittmer, U., Sauter, D., & Kirchhoff, F. (2019). IFI16 Targets the Transcription Factor Sp1 to Suppress HIV-1 Transcription and Latency Reactivation. Cell host & microbe, 25(6), 858–872.e13. [CrossRef]
- Hou, W., Wang, S., Wu, H., Xue, L., Wang, B., Wang, S., & Wang, H. (2022). Small GTPase-A Key Role in Host Cell for Coronavirus Infection and a Potential Target for Coronavirus Vaccine Adjuvant Discovery. Viruses, 14(9), 2044. [CrossRef]
- How, J., & Hobbs, G. (2020). Use of Interferon Alfa in the Treatment of Myeloproliferative Neoplasms: Perspectives and Review of the Literature. Cancers, 12(7), 1954. [CrossRef]
- Hu, W. S., & Hughes, S. H. (2012). HIV-1 reverse transcription. Cold Spring Harbor perspectives in medicine, 2(10), a006882. [CrossRef]
- Huang, J., You, H., Su, C., Li, Y., Chen, S., & Zheng, C. (2018). Herpes Simplex Virus 1 Tegument Protein VP22 Abrogates cGAS/STING-Mediated Antiviral Innate Immunity. Journal of virology, 92(15), e00841-18. [CrossRef]
- Huang, M. , Jiang, J. D., & Peng, Z. (2014). Recent advances in the anti-HCV mechanisms of interferon. Acta pharmaceutica Sinica. B, 4(4), 241–247. [CrossRef]
- Huang, Y., Xie, J., Guo, Y., Sun, W., He, Y., Liu, K., Yan, J., Tao, A., & Zhong, N. (2021). SARS-CoV-2: Origin, Intermediate Host and Allergenicity Features and Hypotheses. Healthcare (Basel, Switzerland), 9(9), 1132. [CrossRef]
- Hui, K., Cheung, M. C., Perera, R., Ng, K. C., Bui, C., Ho, J., Ng, M., Kuok, D., Shih, K. C., Tsao, S. W., Poon, L., Peiris, M., Nicholls, J. M., & Chan, M. (2020). Tropism, replication competence, and innate immune responses of the coronavirus SARS-CoV-2 in human respiratory tract and conjunctiva: an analysis in ex-vivo and in-vitro cultures. The Lancet. Respiratory medicine, 8(7), 687–695. [CrossRef]
- Huyton, T., Göttmann, W., Bade-Döding, C., Paine, A., & Blasczyk, R. (2011). The T/NK cell co-stimulatory molecule SECTM1 is an IFN “early response gene” that is negatively regulated by LPS in human monocytic cells. Biochimica Et Biophysica Acta (BBA) - General Subjects, 1810(12), 1294–1301. [CrossRef]
- Iaconis, G., Jackson, B., Childs, K., Boyce, M., Goodbourn, S., Blake, N., Iturriza-Gomara, M., & Seago, J. (2021). Rotavirus NSP1 Inhibits Type I and Type III Interferon Induction. Viruses, 13(4), 589. [CrossRef]
- Ilaria Sciamanna, Chiara de Luca and Corrado Spadafora (2016), The Reverse Transcriptase Encodes by LINE-1 Retrotransposons in the Genesis, Progression and Therapy of Cancer, Front. Chem., doi:. [CrossRef]
- Ilinykh, P. A., Lubaki, N. M., Widen, S. G., Renn, L. A., Theisen, T. C., Rabin, R. L., Wood, T. G., & Bukreyev, A. (2015). Different Temporal Effects of Ebola Virus VP35 and VP24 Proteins on Global Gene Expression in Human Dendritic Cells. Journal of virology, 89(15), 7567–7583. [CrossRef]
- Illés, Á., Pinczés, L. I., & Egyed, M. (2021). A pharmacokinetic evaluation of ropeginterferon alfa-2b in the treatment of polycythemia vera. Expert opinion on drug metabolism & toxicology, 17(1), 3–7. [CrossRef]
- Inserra, F. , Tajer, C., Antonietti, L., Mariani, J., Ferder, L., & Manucha, W. (2021). Vitamin D supplementation: An alternative to enhance the effectiveness of vaccines against SARS-CoV-2? Vaccine, 39(35): 4930-4931. [CrossRef]
- Islam, A. , Khan, M. A., Ahmed, R., Hossain, M. S., Kabir, S., Islam, M. S., & Siddiki, A. (2021). Transcriptome of nasopharyngeal samples from COVID-19 patients and a comparative analysis with other SARS-CoV-2 infection models reveal disparate host responses against SARS-CoV-2. Journal of translational medicine, 19(1), 32. [CrossRef]
- Ito, H., Morishita, R., Tabata, H., & Nagata, K. (2014). Roles of Rho small GTPases in the tangentially migrating neurons. Histology and histopathology, 29(7), 871–879. [CrossRef]
- Ivashkiv, L. B., & Donlin, L. T. (2014). Regulation of type I interferon responses. Nature reviews. Immunology, 14(1), 36–49. [CrossRef]
- Iverson, E., Griswold, K., Song, D., Gagliardi, T. B., Hamidzadeh, K., Kesimer, M., Sinha, S., Perry, M., Duncan, G. A., & Scull, M. A. (2022). Membrane-Tethered Mucin 1 Is Stimulated by Interferon and Virus Infection in Multiple Cell Types and Inhibits Influenza A Virus Infection in Human Airway Epithelium. mBio, 13(4), e0105522. [CrossRef]
- Ives Charlie-Silva, Amanda P. C. Araújo, Abraão T. B. Guimarães, Flávio P Veras, Helyson L. B. Braz, Letícia G. de Pontes, Roberta J. B. Jorge, Marco A. A. Belo, Bianca H, V. Fernandes, Rafael H. Nóbrega, Giovane Galdino, Antônio Condino-Neto, Jorge Galindo-Villegas, Glaucia M. Machado-Santelli, Paulo R. S. Sanches, Rafael M. Rezende, Eduardo M. Cilli, Guilherme Malafaia, An insight into neurotoxic and toxicity of spike fragments SARS-CoV-2 by exposure environment: A threat to aquatic health?, available at: bioRxiv 2021.01.11.425914; doi:. [CrossRef]
- Jafarzadeh, A., Chauhan, P., Saha, B., Jafarzadeh, S., & Nemati, M. (2020). Contribution of monocytes and macrophages to the local tissue inflammation and cytokine storm in COVID-19: Lessons from SARS and MERS, and potential therapeutic interventions. Life sciences, 257, 118102. [CrossRef]
- Jafarzadeh, A. , Nemati, M., Saha, B., Bansode, Y. D., & Jafarzadeh, S. (2021). Protective Potentials of Type III Interferons in COVID-19 Patients: Lessons from Differential Properties of Type I- and III Interferons. Viral immunology, 34(5), 307–320. [CrossRef]
- Jafarzadeh, A. , Nemati, M., Saha, B., Bansode, Y. D., & Jafarzadeh, S. (2021). Protective Potentials of Type III Interferons in COVID-19 Patients: Lessons from Differential Properties of Type I- and III Interferons. Viral immunology, 34(5), 307–320. [CrossRef]
- Jiang, S. (2020). Don’t rush to deploy COVID-19 vaccines and drugs without sufficient safety guarantees. Nature, 579, 321. [CrossRef]
- Jiao, P., Tian, G., Li, Y., Deng, G., Jiang, Y., Liu, C., Liu, W., Bu, Z., Kawaoka, Y., & Chen, H. (2008). A single-amino-acid substitution in the NS1 protein changes the pathogenicity of H5N1 avian influenza viruses in mice. Journal of virology, 82(3), 1146–1154. [CrossRef]
- The Journal of Infectious Diseases, 128; //www: (2), 261–264. http.
- Jureidini J, McHenry L B. The illusion of evidence based medicine BMJ 2022; 376 :o702. [CrossRef]
- Kalashnyk, O., Lykhmus, O., Izmailov, M., Koval, L., Komisarenko, S., & Skok, M. (2021). SARS-Cov-2 spike protein fragment 674-685 protects mitochondria from releasing cytochrome c in response to apoptogenic influence. Biochemical and biophysical research communications, 561, 14–18. [CrossRef]
- Kalyuzhin, O. V., Ponezheva, L. O., Turapova, A. N., Nurtazina, A. Y., Bykov, A. S., & Karaulov, A. V. (2022). Interferons alpha and Gamma, pidotimod, and tilorone in the treatment of acute respiratory infections in patients with allergic rhinitis: A prospective, cohort clinical and immunological study. Bulletin of Siberian Medicine, 21(2), 48–59. [CrossRef]
- Karlowitz, R. , Stanifer, M. L., Roedig, J., Andrieux, G., Bojkova, D., Bechtel, M., Smith, S., Kowald, L., Schubert, R., Boerries, M., Cinatl, J., Jr, Boulant, S., & van Wijk, S. (2022). USP22 controls type III interferon signaling and SARS-CoV-2 infection through activation of STING. Cell death & disease, 13(8), 684. [CrossRef]
- Katlinski, K. V., Gui, J., Katlinskaya, Y. V., Ortiz, A., Chakraborty, R., Bhattacharya, S., Carbone, C. J., Beiting, D. P., Girondo, M. A., Peck, A. R., Puré, E., Chatterji, P., Rustgi, A. K., Diehl, J. A., Koumenis, C., Rui, H., & Fuchs, S. Y. (2017). Inactivation of Interferon Receptor Promotes the Establishment of Immune Privileged Tumor Microenvironment. Cancer cell, 31(2), 194–207. [CrossRef]
- Kato, A. , Ohnishi, Y., Kohase, M., Saito, S., Tashiro, M., & Nagai, Y. (2001). Y2, the smallest of the Sendai virus C proteins, is fully capable of both counteracting the antiviral action of interferons and inhibiting viral RNA synthesis. Journal of virology, 75(8), 3802–3810. [CrossRef]
- 2021; -9. [CrossRef]
- Khaledi, M., Sameni, F., Yahyazade, S., Radandish, M., Owlia, P., Bagheri, N., Afkhami, H., Mahjoor, M., Esmaelpour, Z., Kohansal, M., & Aghaei, F. (2022). COVID-19 and the potential of Janus family kinase (JAK) pathway inhibition: A novel treatment strategy. Frontiers in medicine, 9, 961027. [CrossRef]
- Khalil, B. A., Elemam, N. M., & Maghazachi, A. A. (2021). Chemokines and chemokine receptors during COVID-19 infection. Computational and structural biotechnology journal, 19, 976–988. [CrossRef]
- Khanmohammadi, S., Rezaei, N., Khazaei, M. et al. A Case of Autosomal Recessive Interferon Alpha/Beta Receptor Alpha Chain (IFNAR1) Deficiency with Severe COVID-19. J Clin Immunol 42, 19–24 (2022). [CrossRef]
- Khatamzas, E. , Hipp, M. M., Gaughan, D., Pichulik, T., Leslie, A., Fernandes, R. A., Muraro, D., Booth, S., Zausmer, K., Sun, M. Y., Kessler, B., Rowland-Jones, S., Cerundolo, V., & Simmons, A. (2017). Snapin promotes HIV-1 transmission from dendritic cells by dampening TLR8 signaling. The EMBO journal, 36(20), 2998–3011. [CrossRef]
- Kho, V. M., Mekers, V. E., Span, P. N., Bussink, J., & Adema, G. J. (2021). Radiotherapy and cGAS/STING signaling: Impact on MDSCs in the tumor microenvironment. Cellular immunology, 362, 104298. [CrossRef]
- Kieseier, B. C., & Hartung, H. P. (2003). Current disease-modifying therapies in multiple sclerosis. Seminars in neurology, 23(2), 133–146. [CrossRef]
- Kim, E. S., Jeon, M. T., Kim, K. S., Lee, S., Kim, S., & Kim, D. G. (2021). Spike Proteins of SARS-CoV-2 Induce Pathological Changes in Molecular Delivery and Metabolic Function in the Brain Endothelial Cells. Viruses, 13(10), 2021. [CrossRef]
- Kim, H., Jang, J., Song, M. J., Kim, G., Park, C. H., Lee, D. H., Lee, S. H., & Chung, J. H. (2022). Attenuation of intrinsic ageing of the skin via elimination of senescent dermal fibroblasts with senolytic drugs. Journal of the European Academy of Dermatology and Venereology : JEADV, 36(7), 1125–1135. [CrossRef]
- Kim, H., Jang, J., Song, M. J., Park, C. H., Lee, D. H., Lee, S. H., & Chung, J. H. (2022). Inhibition of matrix metalloproteinase expression by selective clearing of senescent dermal fibroblasts attenuates ultraviolet-induced photoaging. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie, 150, 113034. [CrossRef]
- Kim, J. C., Park, T. J., & Kang, H. Y. (2022). Skin-Aging Pigmentation: Who Is the Real Enemy? Cells, 11(16), 2541. [CrossRef]
- Kim, J. H., Kim, D. H., Baek, S. H., Lee, H. J., Kim, M. R., Kwon, H. J., & Lee, C. H. (2006). Rengyolone inhibits inducible nitric oxide synthase expression and nitric oxide production by down-regulation of NF-kappaB and p38 MAP kinase activity in LPS-stimulated RAW 264.7 cells. Biochemical pharmacology, 71(8), 1198–1205. [CrossRef]
- Kim, J. Y., Kwon, J. S., Bae, S., Cha, H. H., Lim, J. S., Kim, M. C., Chung, J. W., Park, S. Y., Lee, M. J., Kim, B. N., Jung, J., Kim, M. J., Shin, E. C., & Kim, S. H. (2021). SARS-CoV-2-Specific Antibody and T Cell Response Kinetics According to Symptom Severity. The American journal of tropical medicine and hygiene, 105(2), 395–400. [CrossRef]
- Kim, M. A., Lee, Y.W., Kim, S.R., Kim, J.H., Min, T.K., Park, H.S., Shin, M., Ye, Y.M., Lee, S., Lee, J., Choi, J.H., Jang, G.C., & Chang, Y.S. (2021). COVID-19 vaccine-associated anaphylaxis and allergic reactions: consensus statements of the KAAACI Urticaria/Angioedema/Anaphylaxis Working Group. Allergy Asthma Immunol Res, 13(4):526-544. [CrossRef]
- Kim, Y. M., & Shin, E. C. (2021). Type I and III interferon responses in SARS-CoV-2 infection. Experimental & molecular medicine, 53(5), 750–760. [CrossRef]
- Kim, YM., Shin, EC. Type I and III interferon responses in SARS-CoV-2 infection. Exp Mol Med 53, 750–760 (2021). [CrossRef]
- Kirchdoerfer, R. N., Wang, N., Pallesen, J., Wrapp, D., Turner, H. L., Cottrell, C. A., Corbett, K. S., Graham, B. S., McLellan, J. S., & Ward, A. B. (2018). Stabilized coronavirus spikes are resistant to conformational changes induced by receptor recognition or proteolysis. Scientific reports, 8(1), 15701. [CrossRef]
- Klinkhammer, J., Schnepf, D., Ye, L., Schwaderlapp, M., Gad, H. H., Hartmann, R., Garcin, D., Mahlakõiv, T., & Staeheli, P. (2018). IFN-λ prevents influenza virus spread from the upper airways to the lungs and limits virus transmission. eLife, 7, e33354. [CrossRef]
- Molecular biotherapy, 2; //pubmed: (2), 91–95, doi: https.
- Koga, R. , Ohno, S., Ikegame, S., & Yanagi, Y. (2010). Measles virus-induced immunosuppression in SLAM knock-in mice. Journal of virology, 84(10), 5360–5367. [CrossRef]
- Kong, Z., Yin, H., Wang, F., Liu, Z., Luan, X., Sun, L., Liu, W., & Shang, Y. (2022). Pseudorabies virus tegument protein UL13 recruits RNF5 to inhibit STING-mediated antiviral immunity. PLoS pathogens, 18(5), e1010544. [CrossRef]
- Korth, M. J., Taylor, M. D., & Katze, M. G. (1998). Interferon inhibits the replication of HIV-1, SIV, and SHIV chimeric viruses by distinct mechanisms. Virology, 247(2), 265–273. [CrossRef]
- Kosciuczuk, E. M., Mehrotra, S., Saleiro, D., Kroczynska, B., Majchrzak-Kita, B., Lisowski, P., Driehaus, C., Rogalska, A., Turner, A., Lienhoop, T., Gius, D., Fish, E. N., Vassilopoulos, A., & Platanias, L. C. (2019). Sirtuin 2-mediated deacetylation of cyclin-dependent kinase 9 promotes STAT1 signaling in type I interferon responses. The Journal of biological chemistry, 294(3), 827–837. [CrossRef]
- Kostoff, R. N., Calina, D., Kanduc, D., Briggs, M.B., Vlachoyiannopoulos, P., Svistunov, A.A., & Tsatsakis, A. (2021). Why are we vaccinating children against COVID-19? Toxicol Rep 8, 1665–1684. [CrossRef]
- Kotenko, S. V., Rivera, A., Parker, D., & Durbin, J. E. (2019). Type III IFNs: Beyond antiviral protection. Seminars in immunology, 43, 101303. [CrossRef]
- Kounis, N. G., Koniari, I., Mplani, V., Plotas, P., & Velissaris, D. (2022). Hypersensitivity myocarditis and the pathogenetic conundrum of COVID 19 Vaccine Related Myocarditis. Cardiology, 10.1159/000524224. Advance online publication. [CrossRef]
- Kouwaki, T. , Nishimura, T., Wang, G., & Oshiumi, H. (2021). RIG-I-Like Receptor-Mediated Recognition of Viral Genomic RNA of Severe Acute Respiratory Syndrome Coronavirus-2 and Viral Escape From the Host Innate Immune Responses. Frontiers in immunology, 12, 700926. [CrossRef]
- Kowarz, E. , Krutzke, L., Reis, J., Bracharz, S., Kochanek, S., & Marschalek, R. (2021). Vaccine-induced COVID-19 mimicry syndrome: splice reactions within the SARS-CoV-2 spike open reading frame result in spike protein variants that may cause thromboembolic events in patients immunized with vector-based vaccines. Nature-portfolio [preprint]. [CrossRef]
- Krutzke, L. , Roesler, R., Wiese, S., & Kochanek, S. (2021). Process-related impurities in the ChAdOx1 nCov-19 vaccine. Nature-portfolio [preprint]. [CrossRef]
- Kumar R, Singh N, Gautam S, Singh OP, Gidwani K, et al. (2014) Leishmania Specific CD4 T Cells Release IFNγ That Limits Parasite Replication in Patients with Visceral Leishmaniasis. PLOS Neglected Tropical Diseases 8(10): e3198. [CrossRef]
- Kumar, R., Bunn, P. T., Singh, S. S., Ng, S. S., Montes de Oca, M., De Labastida Rivera, F., Chauhan, S. B., Singh, N., Faleiro, R. J., Edwards, C. L., Frame, T. C. M., Sheel, M., Austin, R. J., Lane, S. W., Bald, T., Smyth, M. J., Hill, G. R., Best, S. E., Haque, A., … Engwerda, C. R. (2020). Type I interferons suppress anti-parasitic immunity and can be targeted to improve treatment of visceral leishmaniasis. Cell Reports, 30(8). [CrossRef]
- Kunz, N., & Kemper, C. (2021). Complement Has Brains-Do Intracellular Complement and Immunometabolism Cooperate in Tissue Homeostasis and Behavior? Frontiers in immunology, 12, 629986. [CrossRef]
- Kuriyama, Y., Shimizu, A., Kanai, S. et al. Coordination of retrotransposons and type I interferon with distinct interferon pathways in dermatomyositis, systemic lupus erythematosus and autoimmune blistering disease. Sci Rep 11, 23146 (2021). [CrossRef]
- Kuriyama, Y., Shimizu, A., Kanai, S., Oikawa, D., Tokunaga, F., Tsukagoshi, H., & Ishikawa, O. (2021). The synchronized gene expression of retrotransposons and type I interferon in dermatomyositis. Journal of the American Academy of Dermatology, 84(4), 1103–1105. [CrossRef]
- Kuroda, M., Halfmann, P. J., Hill-Batorski, L., Ozawa, M., Lopes, T., Neumann, G., Schoggins, J. W., Rice, C. M., & Kawaoka, Y. (2020). Identification of interferon-stimulated genes that attenuate Ebola virus infection. Nature communications, 11(1), 2953. [CrossRef]
- Kuypers F. A. (2022). Hyperinflammation, apoptosis, and organ damage. Experimental biology and medicine (Maywood, N.J.), 247(13), 1112–1123. [CrossRef]
- Kuzmin, I. V., Schwarz, T. M., Ilinykh, P. A., Jordan, I., Ksiazek, T. G., Sachidanandam, R., Basler, C. F., & Bukreyev, A. (2017). Innate Immune Responses of Bat and Human Cells to Filoviruses: Commonalities and Distinctions. Journal of virology, 91(8), e02471-16. [CrossRef]
- Kwon, J. S., Kim, J. Y., Kim, M. C., Park, S. Y., Kim, B. N., Bae, S., Cha, H. H., Jung, J., Kim, M. J., Lee, M. J., Choi, S. H., Chung, J. W., Shin, E. C., & Kim, S. H. (2020). Factors of Severity in Patients with COVID-19: Cytokine/Chemokine Concentrations, Viral Load, and Antibody Responses. The American journal of tropical medicine and hygiene, 103(6), 2412–2418. [CrossRef]
- La Bonnardière, C., Lefèvre, F., & Charley, B. (1994). Interferon response in pigs: molecular and biological aspects. Veterinary immunology and immunopathology, 43(1-3), 29–36. [CrossRef]
- Lagunas-Rangel, F. A., & Chávez-Valencia, V. (2020). High IL-6/IFN-γ ratio could be associated with severe disease in COVID-19 patients. Journal of medical virology, 92(10), 1789–1790. [CrossRef]
- LaSalle, T. J., Gonye, A. L. K., Freeman, S. S., Kaplonek, P., Gushterova, I., Kays, K. R., Manakongtreecheep, K., Tantivit, J., Rojas-Lopez, M., Russo, B. C., Sharma, N., Thomas, M. F., Lavin-Parsons, K. M., Lilly, B. M., Mckaig, B. N., Charland, N. C., Khanna, H. K., Lodenstein, C. L., Margolin, J. D., Blaum, E. M., … Sade-Feldman, M. (2021). Longitudinal characterization of circulating neutrophils uncovers distinct phenotypes associated with disease severity in hospitalized COVID-19 patients. bioRxiv : the preprint server for biology, 2021.10.04.463121. [CrossRef]
- Lasfar, A., Abushahba, W., Balan, M., & Cohen-Solal, K. A. (2011). Interferon lambda: a new sword in cancer immunotherapy. Clinical & developmental immunology, 2011, 349575. [CrossRef]
- Lasfar, A., de laTorre, A., Abushahba, W., Cohen-Solal, K. A., Castaneda, I., Yuan, Y., Reuhl, K., Zloza, A., Raveche, E., Laskin, D. L., & Kotenko, S. V. (2016). Concerted action of IFN-α and IFN-λ induces local NK cell immunity and halts cancer growth. Oncotarget, 7(31), 49259–49267. [CrossRef]
- Lasfar, A., Gogas, H., Zloza, A., Kaufman, H. L., & Kirkwood, J. M. (2016). IFN-λ cancer immunotherapy: new kid on the block. Immunotherapy, 8(8), 877–888. [CrossRef]
- Lasfar, A., Zloza, A., & Cohen-Solal, K. A. (2016). IFN-lambda therapy: current status and future perspectives. Drug discovery today, 21(1), 167–171. [CrossRef]
- Laskovs, M., Partridge, L., & Slack, C. (2022). Molecular inhibition of RAS signalling to target ageing and age-related health. Disease models & mechanisms, 15(10), dmm049627. [CrossRef]
- Laviada-Molina, H. A., Leal-Berumen, I., Rodriguez-Ayala, E., & Bastarrachea, R. A. (2020). Working Hypothesis for Glucose Metabolism and SARS-CoV-2 Replication: Interplay Between the Hexosamine Pathway and Interferon RF5 Triggering Hyperinflammation. Role of BCG Vaccine?. Frontiers in endocrinology, 11, 514. [CrossRef]
- Lazear, H. M., Schoggins, J. W., & Diamond, M. S. (2019). Shared and Distinct Functions of Type I and Type III Interferons. Immunity, 50(4), 907–923. [CrossRef]
- Le Coupanec, A., Desforges, M., Kaufer, B., Dubeau, P., Côté, M., & Talbot, P. J. (2021). Potential differences in cleavage of the S protein and type-1 interferon together control human coronavirus infection, propagation, and neuropathology within the central nervous system. Journal of virology, 95(10), e00140-21. Advance online publication. [CrossRef]
- Le, H., Spearman, P., Waggoner, S. N., & Singh, K. (2022). Ebola virus protein VP40 stimulates IL-12- and IL-18-dependent activation of human natural killer cells. JCI insight, 7(16), e158902. [CrossRef]
- Lee AJ, Mian F, Poznanski SM, Stackaruk M, Chan T, Chew MV and Ashkar AA (2019) Type I Interferon Receptor on NK Cells Negatively Regulates Interferon-γ Production. Front. Immunol 10, 1261. [CrossRef]
- Lee, J.S., Shin, EC. The type I interferon response in COVID-19: implications for treatment. Nat Rev Immunol 20, 585–586 (2020). [CrossRef]
- Lee-Kirsch M. A. (2017). The Type I Interferonopathies. Annual review of medicine, 68, 297–315. [CrossRef]
- Lefèvre, F., Guillomot, M., D'Andréa, S., Battegay, S., & La Bonnardière, C. (1998). Interferon-delta: the first member of a novel type I interferon family. Biochimie, 80(8-9), 779–788. [CrossRef]
- Lekkerkerker, A. N., van Kooyk, Y., & Geijtenbeek, T. B. (2006). Viral piracy: HIV-1 targets dendritic cells for transmission. Current HIV research, 4(2), 169–176. [CrossRef]
- Letarov, A. V., Babenko, V. V., & Kulikov, E. E. (2021). Free SARS-CoV-2 Spike Protein S1 Particles May Play a Role in the Pathogenesis of COVID-19 Infection. Biochemistry. Biokhimiia, 86(3), 257–261. [CrossRef]
- Leung, L. W., Park, M. S., Martinez, O., Valmas, C., López, C. B., & Basler, C. F. (2011). Ebolavirus VP35 suppresses IFN production from conventional but not plasmacytoid dendritic cells. Immunology and cell biology, 89(7), 792–802. [CrossRef]
- Levine, M. E., Lu, A. T., Quach, A., Chen, B. H., Assimes, T. L., Bandinelli, S., Hou, L., Baccarelli, A. A., Stewart, J. D., Li, Y., Whitsel, E. A., Wilson, J. G., Reiner, A. P., Aviv, A., Lohman, K., Liu, Y., Ferrucci, L., & Horvath, S. (2018). An epigenetic biomarker of aging for lifespan and healthspan. Aging, 10(4), 573–591. [CrossRef]
- Li, B., Raghwani, J., Hill, S. C., Francois, S., Lefrancq, N., Liang, Y.,... & Tian, H. (2023). Association of poultry vaccination with the interspecies transmission and molecular evolution of H5 subtype avian influenza virus. bioRxiv, 2023-12. [CrossRef]
- Li, Q., Humphries, F., Girardin, R. C., Wallace, A., Ejemel, M., Amcheslavsky, A., McMahon, C. T., Schiller, Z. A., Ma, Z., Cruz, J., Dupuis, A. P., Payne, A. F., Maryam, A., Yilmaz, N. K., McDonough, K. A., Pierce, B. G., Schiffer, C. A., Kruse, A. C., Klempner, M. S., Cavacini, L. A., … Wang, Y. (2022). Mucosal nanobody IgA as inhalable and affordable prophylactic and therapeutic treatment against SARS-CoV-2 and emerging variants. Frontiers in immunology, 13, 995412. [CrossRef]
- Li, S.-fang, Gong, M.-jiao, Zhao, F.-rong, Shao, J.-jun, Xie, Y.-li, Zhang, Y.-guang, & Chang, H.-yun. (2018). Type I interferons: Distinct biological activities and current applications for viral infection. Cellular Physiology and Biochemistry, 51(5), 2377–2396. [CrossRef]
- Li, W., Wang, G., Zhang, H., Xin, G., Zhang, D., Zeng, J., Chen, X., Xu, Y., Cui, Y., & Li, K. (2010). Effects of NS1 variants of H5N1 influenza virus on interferon induction, TNFalpha response and p53 activity. Cellular & molecular immunology, 7(3), 235–242. [CrossRef]
- Li, X. , Li, Y., Fang, S., Su, J., Jiang, J., Liang, B., Huang, J., Zhou, B., Zang, N., Ho, W., Li, J., Li, Y., Chen, H., Ye, L., & Liang, H. (2017). Downregulation of autophagy-related gene ATG5 and GABARAP expression by IFN-λ1 contributes to its anti-HCV activity in human hepatoma cells. Antiviral research, 140, 83–94. [CrossRef]
- Li, X., Xie, J., Li, D., Li, H., Niu, Y., Wu, B., Yang, Y., Yan, Z., Zhang, X., Chen, L., & Feng, R. (2022). HSP27 Attenuates cGAS-Mediated IFN-β Signaling through Ubiquitination of cGAS and Promotes PRV Infection. Viruses, 14(9), 1851. [CrossRef]
- Li, Y., Li, C., Xue, P., Zhong, B., Mao, A. P., Ran, Y., Chen, H., Wang, Y. Y., Yang, F., & Shu, H. B. (2009). ISG56 is a negative-feedback regulator of virus-triggered signaling and cellular antiviral response. Proceedings of the National Academy of Sciences of the United States of America, 106(19), 7945–7950. [CrossRef]
- Li, Z., Jiang, Y., Jiao, P., Wang, A., Zhao, F., Tian, G., Wang, X., Yu, K., Bu, Z., & Chen, H. (2006). The NS1 gene contributes to the virulence of H5N1 avian influenza viruses. Journal of virology, 80(22), 11115–11123. [CrossRef]
- Lim, U., & Song, M. A. (2018). DNA Methylation as a Biomarker of Aging in Epidemiologic Studies. Methods in molecular biology (Clifton, N.J.), 1856, 219–231. [CrossRef]
- Lin, J., Cao, Y., Shah, A. U., Zuo, J., Zhang, S., Yu, Q., Chong, M., & Yang, Q. (2022). Inhibition of the antigen-presenting ability of dendritic cells by non-structural protein 2 of influenza A virus. Veterinary microbiology, 267, 109392. [CrossRef]
- Liu, J., Qian, C., & Cao, X. (2016). Post-Translational Modification Control of Innate Immunity. Immunity, 45(1), 15–30. [CrossRef]
- Liu, N., Pang, X., Zhang, H., & Ji, P. (2022). The cGAS-STING Pathway in Bacterial Infection and Bacterial Immunity. Frontiers in immunology, 12, 814709. [CrossRef]
- Liu, X. , Yin, L., Xue, M., Chen, J., Li, L., Fu, F., Feng, L., & Liu, P. (2022). Coronavirus Porcine Deltacoronavirus Upregulates MHC Class I Expression through RIG-I/IRF1-Mediated NLRC5 Induction. Journal of virology, 96(7), e0015822. [CrossRef]
- Locke, M., Lythe, G., López-García, M., Muñoz-Fontela, C., Carroll, M., & Molina-París, C. (2021). Quantification of Type I Interferon Inhibition by Viral Proteins: Ebola Virus as a Case Study. Viruses, 13(12), 2441. [CrossRef]
- Lodi, L. , Mastrolia, M. V., Bello, F., Rossi, G. M., Angelotti, M. L., Crow, Y. J., Romagnani, P., & Vaglio, A. (2022). Type I interferon-related kidney disorders. Kidney international, 101(6), 1142–1159. [CrossRef]
- Lopez, L., Sang, P. C., Tian, Y., & Sang, Y. (2020). Dysregulated Interferon Response Underlying Severe COVID-19. Viruses, 12(12), 1433. [CrossRef]
- Low, Z. Y., Zabidi, N. Z., Yip, A., Puniyamurti, A., Chow, V., & Lal, S. K. (2022). SARS-CoV-2 Non-Structural Proteins and Their Roles in Host Immune Evasion. Viruses, 14(9), 1991. [CrossRef]
- Lu, C. , Klement, J. D., Ibrahim, M. L., Xiao, W., Redd, P. S., Nayak-Kapoor, A., Zhou, G., & Liu, K. (2019). Type I interferon suppresses tumor growth through activating the STAT3-granzyme B pathway in tumor-infiltrating cytotoxic T lymphocytes. Journal for immunotherapy of cancer, 7(1), 157. [CrossRef]
- Lu, Y. , Cai, H., Lu, M., Ma, Y., Li, A., Gao, Y., Zhou, J., Gu, H., Li, J., & Gu, J. (2020). Porcine Epidemic Diarrhea Virus Deficient in RNA Cap Guanine-N-7 Methylation Is Attenuated and Induces Higher Type I and III Interferon Responses. Journal of virology, 94(16), e00447-20. [CrossRef]
- Lubaki, N. M., Younan, P., Santos, R. I., Meyer, M., Iampietro, M., Koup, R. A., & Bukreyev, A. (2016). The Ebola Interferon Inhibiting Domains Attenuate and Dysregulate Cell-Mediated Immune Responses. PLoS pathogens, 12(12), e1006031. [CrossRef]
- Luecke, S. , & Paludan, S. R. (2017). Molecular requirements for sensing of intracellular microbial nucleic acids by the innate immune system. Cytokine, 98, 4–14. [CrossRef]
- Luisetto, M. Luisetto, M., Almukthar, N., & Tarro, G., Intracellular Reverse Transcription of COVID-19 mRNA Vaccine, LAP LAMBERT Academic Publishing. 2022; 1, 3-67. 9: ISBN, 3157. [Google Scholar]
- Luo, K., Li, N., Ye, W., Gao, H., Luo, X., & Cheng, B. (2022). Activation of Stimulation of Interferon Genes (STING) Signal and Cancer Immunotherapy. Molecules (Basel, Switzerland), 27(14), 4638. [CrossRef]
- Lurie, N. , Saville, M., Hatchett, R., & Halton, J. (2020). Developing COVID-19 vaccines at pandemic speed. N Engl J Med, 382, 1969–1973. [CrossRef]
- Lv, L., Cao, M., Bai, J., Jin, L., Wang, X., Gao, Y., Liu, X., & Jiang, P. (2020). PRV-encoded UL13 protein kinase acts as an antagonist of innate immunity by targeting IRF3-signaling pathways. Veterinary microbiology, 250, 108860. [CrossRef]
- Lv, L., Cao, M., Bai, J., Jin, L., Wang, X., Gao, Y., Liu, X., & Jiang, P. (2020). PRV-encoded UL13 protein kinase acts as an antagonist of innate immunity by targeting IRF3-signaling pathways. Veterinary microbiology, 250, 108860. [CrossRef]
- Lyons-Weiler, J. (2020). Pathogenic priming likely contributes to serious and critical illness and mortality in COVID-19 via autoimmunity. J Transl Autoim, 3, 100051. [CrossRef]
- Lyudmila Shalamova, Ulrike Felgenhauer, Andreas R. Schaubmar, Kathrin Buettner, Marek Widera, Sandra Ciesek, Friedemann Weber (2022), Omicron variant of SARS-CoV-2 exhibits an increased resilience to the antiviral type I interferon response, doi:. [CrossRef]
- Ma, D. Y., & Suthar, M. S. (2015). Mechanisms of innate immune evasion in re-emerging RNA viruses. Current opinion in virology, 12, 26–37. [CrossRef]
- Ma, Y. , Su, X. Z., & Lu, F. (2020). The Roles of Type I Interferon in Co-infections With Parasites and Viruses, Bacteria, or Other Parasites. Frontiers in immunology, 11, 1805. [CrossRef]
- Ma, Y. , Wang, C., Xue, M., Fu, F., Zhang, X., Li, L., Yin, L., Xu, W., Feng, L., & Liu, P. (2018). The Coronavirus Transmissible Gastroenteritis Virus Evades the Type I Interferon Response through IRE1α-Mediated Manipulation of the MicroRNA miR-30a-5p/SOCS1/3 Axis. Journal of virology, 92(22), e00728-18. [CrossRef]
- Ma, Z. , Qin, Y., Jia, Y., Xie, Y., Qi, X., Guo, Y., He, J., Zhang, Y., Li, F., Yu, J., Zhu, H., Yang, F., Zhang, Y., Mao, R., & Zhang, J. (2022). Thyroid dysfunction incidence and risk factors in Chinese chronic hepatitis B patients treated with pegylated interferon alpha: A long-term follow-up study. Journal of viral hepatitis, 29(6), 412–419. [CrossRef]
- Magalhaes, J., Tresse, E., Ejlerskov, P., Hu, E., Liu, Y., Marin, A., Montalant, A., Satriano, L., Rundsten, C. F., Carlsen, E., Rydbirk, R., Sharifi-Zarchi, A., Andersen, J. B., Aznar, S., Brudek, T., Khodosevich, K., Prinz, M., Perrier, J. M., Sharma, M., Gasser, T., … Issazadeh-Navikas, S. (2021). PIAS2-mediated blockade of IFN-β signaling: a basis for sporadic Parkinson disease dementia. Molecular psychiatry, 26(10), 6083–6099. [CrossRef]
- Mahalapbutr, P. Mahalapbutr, P., Kongtaworn, N., & Rungrotmongkol, T. (2020). Structural insight into the recognition of S-adenosyl-L-homocysteine and sinefungin in SARS-CoV-2 Nsp16/Nsp10 RNA cap 2′-O-Methyltransferase. Computational and structural biotechnology journal, 2765. [Google Scholar]
- Makrinioti, H., Bush, A., Gern, J., Johnston, S. L., Papadopoulos, N., Feleszko, W., Camargo, C. A., Jr, Hasegawa, K., & Jartti, T. (2021). The Role of Interferons in Driving Susceptibility to Asthma Following Bronchiolitis: Controversies and Research Gaps. Frontiers in immunology, 12, 761660. Coomes, E. A., & Haghbayan, H. (2020). Frontiers in immunology, 12, 761660. [CrossRef]
- Malik, G., & Zhou, Y. (2020). Innate Immune Sensing of Influenza A Virus. Viruses, 12(7), 755. [CrossRef]
- Malmgaard L. (2004). Induction and regulation of IFNs during viral infections. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research, 24(8), 439–454. [CrossRef]
- Malur, M., Gale, M., Jr, & Krug, R. M. (2012). LGP2 downregulates interferon production during infection with seasonal human influenza A viruses that activate interferon regulatory factor 3. Journal of virology, 86(19), 10733–10738. [CrossRef]
- Manan, A., Pirzada, R. H., Haseeb, M., & Choi, S. (2022). Toll-like Receptor Mediation in SARS-CoV-2: A Therapeutic Approach. International journal of molecular sciences, 23(18), 10716. [CrossRef]
- Mangia, A., Mottola, L., & Santoro, R. (2013). Interleukin 28B polymorphisms as predictor of response in hepatitis C virus genotype 2 and 3 infected patients. World journal of gastroenterology, 19(47), 8924–8928. [CrossRef]
- Margolis, S. R., Wilson, S. C., & Vance, R. E. (2017). Evolutionary Origins of cGAS-STING Signaling. Trends in immunology, 38(10), 733–743. [CrossRef]
- Marquis, K. A., Becker, R. L., Weiss, A. N., Morris, M. C., & Ferran, M. C. (2020). The VSV matrix protein inhibits NF-κB and the interferon response independently in mouse L929 cells. Virology, 548, 117–123. [CrossRef]
- Marsh, G. A., & Wang, L. F. (2012). Hendra and Nipah viruses: why are they so deadly?. Current opinion in virology, 2(3), 242–247. [CrossRef]
- Martel, J., Ko, Y. F., Young, J. D., & Ojcius, D. M. (2020). Could nasal nitric oxide help to mitigate the severity of COVID-19? Microbes and infection, 22(4-5), 168–171. [CrossRef]
- Martins, D. , Dipasquale, O., Davies, K., Cooper, E., Tibble, J., Veronese, M., Frigo, M., Williams, S., Turkheimer, F., Cercignani, M., & Harrison, N. A. (2022). Transcriptomic and cellular decoding of functional brain connectivity changes reveal regional brain vulnerability to pro- and anti-inflammatory therapies. Brain, behavior, and immunity, 102, 312–323. [CrossRef]
- Mary Hongying Cheng, She Zhang, Rebecca A. Porritt, Magali Noval Rivas, Lisa Paschold, Edith Willscher, Mascha Binder, Mosche Arditi and Ivet Bahar (2020), Superantigenic character of an insert unique to SARS-CoV-2 spike supported by skewed TCR repertoire in patients with hyperinflammation, Proceedings of the National Academy of Sciences of the United States of America, 117(41), pp 25254-25262, doi:. [CrossRef]
- Mateo, M., Reid, S. P., Leung, L. W., Basler, C. F., & Volchkov, V. E. (2010). Ebolavirus VP24 binding to karyopherins is required for inhibition of interferon signaling. Journal of virology, 84(2), 1169–1175. [CrossRef]
- Mathern, D. R., & Heeger, P. S. (2015). Molecules Great and Small: The Complement System. Clinical journal of the American Society of Nephrology : CJASN, 10(9), 1636–1650. [CrossRef]
- Mathieu, E. , Ritchie, H., Ortiz-Ospina, E., Roser, M., Hasell, J., Appel, C., Giattino, C., & Ortiz-Ospina, E. (2021). A global database of COVID-19 vaccinations. Nat Hum Behav 5, 947–953, Data on COVID-19 (coronavirus) vaccinations by Our World in Data, accessible: https://github.com/owid/COVID-19-data/tree/master/public/data/vaccinations. [CrossRef]
- Matic, S. Matic, S., Popovic, S., Djurdjevic, P., Todorovic, D., Djordjevic, N., Mijailovic, Z.,... & Baskic, D. (2020). SARS-CoV-2 infection induces mixed M1/M2 phenotype in circulating monocytes and alterations in both dendritic cell and monocyte subsets. PLoS One, 0241. [Google Scholar]
- Matos, P., Pereira, J., & Jordan, P. (2022). Targeting Cancer by Using Nanoparticles to Modulate RHO GTPase Signaling. Advances in experimental medicine and biology, 1357, 115–127. [CrossRef]
- McCullough, P. A., Kelly, R.J., Ruocco, G., Lerma, E., Tumlin, J., Wheelan, K.R., Katz, N., Lepor, N.E., Vijay, K., Carter, H., Singh, B., McCullough, S.P., Bhambi, B.K., Palazzuoli, A., De Ferrari, G.M., Milligan, G.P., Safder, T., Tecson, K.M., Wang, D.D., McKinnon, J.E., O'Neill, W.W., Zervos, M., & Risch, H.A. (2021). Pathophysiological basis and rationale for early outpatient treatment of SARS-CoV-2 (COVID-19) Infection. Am J Med, 134(1):16-22. [CrossRef]
- McDonough, A., Lee, R. V., & Weinstein, J. R. (2017). Microglial Interferon Signaling and White Matter. Neurochemical research, 42(9), 2625–2638. [CrossRef]
- McKellar, J., Rebendenne, A., Wencker, M., Moncorgé, O., & Goujon, C. (2021). Mammalian and Avian Host Cell Influenza A Restriction Factors. Viruses, 13(3), 522. [CrossRef]
- McNab, F. , Mayer-Barber, K., Sher, A., Wack, A., & O'Garra, A. (2015). Type I interferons in infectious disease. Nature reviews. Immunology, 15(2), 87–103. [CrossRef]
- Meng Zhongji, Wang Tongyu, Chen Li, Chen Xinhe, Li Longti, Qin Xueqin, Li Hai *, Luo Jie *, The Effect of Recombinant Human Interferon Alpha Nasal Drops to Prevent COVID-19 Pneumonia for Medical Staff in an Epidemic Area, Current Topics in Medicinal Chemistry 2021; 21(10). d.
- Menni, C. , Klaser, K., May, A., Polidori, L., Nguzen, L.H., Drew, D.A., Merino, J., Hu, C., Selvachandran, S., Antonelli, M., Murray, B., Canas, L.S., Molteni, E., Graham, M.S., Modat, M., Joshi, A.D., Mangino, M., Hammers, A., Goodman, A.L., Chan, A.T., Wolf, J., Steves, C.J., Valdes, A.M., Ourselin, S., & Spector, T.D. (2021). Vaccine side-effects and SARS-CoV-2 infection after vaccination in users of the COVID Symptom Study app in the UK: a prospective observational study. Lancet Infect Dis, 21(7): 939-949. [CrossRef]
- Mesev, E. V., LeDesma, R. A., & Ploss, A. (2019). Decoding type I and III interferon signalling during viral infection. Nature microbiology, 4(6), 914–924. [CrossRef]
- Mesquita, S. D., Ferreira, A. C., Gao, F., Coppola, G., Geschwind, D. H., Sousa, J. C., Correia-Neves, M., Sousa, N., Palha, J. A., & Marques, F. (2015). The choroid plexus transcriptome reveals changes in type I and II interferon responses in a mouse model of Alzheimer's disease. Brain, behavior, and immunity, 49, 280–292. [CrossRef]
- Mestecky J. (1987). The common mucosal immune system and current strategies for induction of immune responses in external secretions. Journal of clinical immunology, 7(4), 265–276. [CrossRef]
- Mestecky, J., & McGhee, J. R. (1992). Prospects for human mucosal vaccines. Advances in experimental medicine and biology, 327, 13–23. [CrossRef]
- Metz-Zumaran, C. , Kee, C., Doldan, P., Guo, C., Stanifer, M. L., & Boulant, S. (2022). Increased Sensitivity of SARS-CoV-2 to Type III Interferon in Human Intestinal Epithelial Cells. Journal of virology, 96(7), e0170521. [CrossRef]
- Meyer, K., Patra, T., Vijayamahantesh, & Ray, R. (2021). SARS-CoV-2 Spike Protein Induces Paracrine Senescence and Leukocyte Adhesion in Endothelial Cells. Journal of virology, 95(17), e0079421. [CrossRef]
- Meylan, P. R., Guatelli, J. C., Munis, J. R., Richman, D. D., & Kornbluth, R. S. (1993). Mechanisms for the inhibition of HIV replication by interferons-alpha, -beta, and -gamma in primary human macrophages. Virology, 193(1), 138–148. [CrossRef]
- Michael W. Russell, Zina Moldoveanu, Pearay L. Ogra and Jiri Mesteacky, Mucosal Immunity in COVID- 19. [CrossRef]
- Minich, D. M., Henning, M., Darley, C., Fahoum, M., Schuler, C. B., & Frame, J. (2022). Is Melatonin the "Next Vitamin D"?: A Review of Emerging Science, Clinical Uses, Safety, and Dietary Supplements. Nutrients, 14(19), 3934. [CrossRef]
- Mizutani, T., Fukushi, S., Saijo, M., Kurane, I., & Morikawa, S. (2004). Importance of Akt signaling pathway for apoptosis in SARS-CoV-infected Vero E6 cells. Virology, 327(2), 169–174. [CrossRef]
- Mo, S., Tang, W., Xie, J., Chen, S., Ren, L., Zang, N., Xie, X., Deng, Y., Gao, L., & Liu, E. (2021). Respiratory syncytial virus activates rab5a to suppress IRF1-dependent lambda interferon production, subverting the antiviral defense of airway epithelial cells. Journal of Virology, 95(8). [CrossRef]
- Moerdyk-Schauwecker, M., Shah, N. R., Murphy, A. M., Hastie, E., Mukherjee, P., & Grdzelishvili, V. Z. (2013). Resistance of pancreatic cancer cells to oncolytic vesicular stomatitis virus: role of type I interferon signaling. Virology, 436(1), 221–234. [CrossRef]
- Moozhipurath, R.K., Kraft, L. & Skiera, B. Evidence of protective role of Ultraviolet-B (UVB) radiation in reducing COVID-19 deaths. Sci Rep 10, 17705 (2020). [CrossRef]
- Moozhipurath, R.K., Kraft, L. Association of lockdowns with the protective role of ultraviolet-B (UVB) radiation in reducing COVID-19 deaths. Sci Rep 11, 22851 (2021). [CrossRef]
- Mordstein, M., Kochs, G., Dumoutier, L., Renauld, J. C., Paludan, S. R., Klucher, K., & Staeheli, P. (2008). Interferon-lambda contributes to innate immunity of mice against influenza A virus but not against hepatotropic viruses. PLoS pathogens, 4(9), e1000151. [CrossRef]
- Mordstein, M., Michiels, T., & Staeheli, P. (2010). What have we learned from the IL28 receptor knockout mouse? Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research, 30(8), 579–584. [CrossRef]
- Mordstein, M., Neugebauer, E., Ditt, V., Jessen, B., Rieger, T., Falcone, V., Sorgeloos, F., Ehl, S., Mayer, D., Kochs, G., Schwemmle, M., Günther, S., Drosten, C., Michiels, T., & Staeheli, P. (2010). Lambda interferon renders epithelial cells of the respiratory and gastrointestinal tracts resistant to viral infections. Journal of virology, 84(11), 5670–5677. [CrossRef]
- Moreno, L., Quereda, C., Moreno, A., Perez-Elías, M. J., Antela, A., Casado, J. L., Dronda, F., Mateos, M. L., Bárcena, R., & Moreno, S. (2004). Pegylated interferon alpha2b plus ribavirin for the treatment of chronic hepatitis C in HIV-infected patients. AIDS (London, England), 18(1), 67–73. [CrossRef]
- Morita, N., Tanaka, Y., Odkhuu, E., Naiki, Y., Komatsu, T., & Koide, N. (2020). Sendai virus V protein decreases nitric oxide production by inhibiting RIG-I signaling in infected RAW264.7 macrophages. Microbes and infection, 22(8), 322–330. [CrossRef]
- Mortezaee, K., & Majidpoor, J. (2022). Cellular immune states in SARS-CoV-2-induced disease. Frontiers in Immunology, 13, doi:. [CrossRef]
- m: Cevik, Krutika Kuppalli, Jason Kindrachuk and Malik Peiris (2020), Virology, pathology and pathogenesis of SARS-CoV-2, BMJ, 371, 2020; -2. [CrossRef]
- Müller, L., Aigner, P., & Stoiber, D. (2017). Type I Interferons and Natural Killer Cell Regulation in Cancer. Frontiers in immunology, 8, 304. [CrossRef]
- Munir S, Hillyer P, Le Nouën C, Buchholz UJ, Rabin RL, et al. (2011) Respiratory Syncytial Virus Interferon Antagonist NS1 Protein Suppresses and Skews the Human T Lymphocyte Response. PLOS Pathogens 7(4): e1001336. [CrossRef]
- Murata, M., Nagai, M., Bando, S., Dobashi, H., & Takahara, J. (1993). Emergence of acute interstitial pneumonia following high dose interferon delta treatment in a case of chronic myelogenous leukemia. Internal medicine (Tokyo, Japan), 32(9), 716–718. [CrossRef]
- Murray, C.J.L. The Global Burden of Disease Study at 30 years. Nat Med 28, 2019–2026 (2022). [CrossRef]
- Mustelin, T., & Ukadike, K. C. (2020). How Retroviruses and Retrotransposons in Our Genome May Contribute to Autoimmunity in Rheumatological Conditions. Frontiers in immunology, 11, 593891. [CrossRef]
- Mustelin, T., & Ukadike, K. C. (2020). How Retroviruses and Retrotransposons in Our Genome May Contribute to Autoimmunity in Rheumatological Conditions. Frontiers in immunology, 11, 593891. [CrossRef]
- Myasnikov, A. L., Berns, S. A., Talyzin, P. A., & Ershov, F. I. (2021). Voprosy virusologii, 66(1), 47–54. [CrossRef]
- Naggie, S., Osinusi, A., Katsounas, A., Lempicki, R., Herrmann, E., Thompson, A. J., Clark, P. J., Patel, K., Muir, A. J., McHutchison, J. G., Schlaak, J. F., Trippler, M., Shivakumar, B., Masur, H., Polis, M. A., & Kottilil, S. (2012). Dysregulation of innate immunity in hepatitis C virus genotype 1 IL28B-unfavorable genotype patients: impaired viral kinetics and therapeutic response. Hepatology (Baltimore, Md.), 56(2), 444–454. [CrossRef]
- Naito, Y. , Takagi, T., Yamamoto, T., & Watanabe, S. (2020). Association between selective IgA deficiency and COVID-19. J Clin Biochem Nutr, 67(2): 122-125. [CrossRef]
- Narunsky-Haziza, L., Sepich-Poore, G. D., Livyatan, I., Asraf, O., Martino, C., Nejman, D., Gavert, N., Stajich, J. E., Amit, G., González, A., Wandro, S., Perry, G., Ariel, R., Meltser, A., Shaffer, J. P., Zhu, Q., Balint-Lahat, N., Barshack, I., Dadiani, M., … Straussman, R. (2022). Pan-cancer analyses reveal cancer-type-specific fungal ecologies and bacteriome interactions. Cell, 185(20), 3789–3806. [CrossRef]
- NatBiotech, (2016). Research not fit to print. Nat Biotechnol, 34(2): 115. [CrossRef]
- Navaratnarajah, C. K., Pease, D. R., Halfmann, P. J., Taye, B., Barkhymer, A., Howell, K. G., Charlesworth, J. E., Christensen, T. A., Kawaoka, Y., Cattaneo, R., Schneider, J. W., & Wanek Family Program for HLHS-Stem Cell Pipeline (2021). Highly Efficient SARS-CoV-2 Infection of Human Cardiomyocytes: Spike Protein-Mediated Cell Fusion and Its Inhibition. Journal of virology, 95(24), e0136821. [CrossRef]
- Nchioua, R. , Kmiec, D., Müller, J. A., Conzelmann, C., Groß, R., Swanson, C. M., Neil, S., Stenger, S., Sauter, D., Münch, J., Sparrer, K., & Kirchhoff, F. (2020). SARS-CoV-2 Is Restricted by Zinc Finger Antiviral Protein despite Preadaptation to the Low-CpG Environment in Humans. mBio, 11(5), e01930-20. [CrossRef]
- Nelson, B. R., Roby, J. A., Dobyns, W. B., Rajagopal, L., Gale, M., Jr, & Adams Waldorf, K. M. (2020). Immune Evasion Strategies Used by Zika Virus to Infect the Fetal Eye and Brain. Viral immunology, 33(1), 22–37. [CrossRef]
- Ngunjiri, J. M., Buchek, G. M., Mohni, K. N., Sekellick, M. J., & Marcus, P. I. (2013). Influenza virus subpopulations: exchange of lethal H5N1 virus NS for H1N1 virus NS triggers de novo generation of defective-interfering particles and enhances interferon-inducing particle efficiency. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research, 33(3), 99–107. [CrossRef]
- Nguyen, A. A., Habiballah, S. B., Platt, C. D., Geha, R. S., Chou, J. S., & McDonald, D. R. (2020). Immunoglobulins in the treatment of COVID-19 infection: Proceed with caution! Clinical immunology (Orlando, Fla.), 216, 108459. [CrossRef]
- Nie, Y., Yang, D., & Oppenheim, J. J. (2016). Alarmins and Antitumor Immunity. Clinical therapeutics, 38(5), 1042–1053. [CrossRef]
- Nogales, A., Martinez-Sobrido, L., Topham, D. J., & DeDiego, M. L. (2017). NS1 Protein Amino Acid Changes D189N and V194I Affect Interferon Responses, Thermosensitivity, and Virulence of Circulating H3N2 Human Influenza A Viruses. Journal of virology, 91(5), e01930-16. [CrossRef]
- Nogales, A., Villamayor, L., Utrilla-Trigo, S., Ortego, J., Martinez-Sobrido, L., & DeDiego, M. L. (2021). Natural Selection of H5N1 Avian Influenza A Viruses with Increased PA-X and NS1 Shutoff Activity. Viruses, 13(9), 1760. [CrossRef]
- Odendall, C., & Kagan, J. C. (2015). The unique regulation and functions of type III interferons in antiviral immunity. Current opinion in virology, 12, 47–52. [CrossRef]
- Odkhuu, E., Komatsu, T., Naiki, Y., Koide, N., & Yokochi, T. (2014). Sendai virus C protein inhibits lipopolysaccharide-induced nitric oxide production through impairing interferon-β signaling. International immunopharmacology, 23(1), 267–272. [CrossRef]
- Odnokoz, O. , Yu, P., Peck, A. R., Sun, Y., Kovatich, A. J., Hooke, J. A., Hu, H., Mitchell, E. P., Rui, H., & Fuchs, S. Y. (2020). Malignant cell-specific pro-tumorigenic role of type I interferon receptor in breast cancers. Cancer biology & therapy, 21(7), 629–636. [CrossRef]
- Ogando, N. S., Zevenhoven-Dobbe, J. C., van der Meer, Y., Bredenbeek, P. J., Posthuma, C. C., & Snijder, E. J. (2020). The Enzymatic Activity of the nsp14 Exoribonuclease Is Critical for Replication of MERS-CoV and SARS-CoV-2. Journal of virology, 94(23), e01246-20. [CrossRef]
- Oh, S. J., & Shin, O. S. (2021). SARS-CoV-2 Nucleocapsid Protein Targets RIG-I-Like Receptor Pathways to Inhibit the Induction of Interferon Response. Cells, 10(3), 530. [CrossRef]
- Oliva, A. , Kinter, A. L., Vaccarezza, M., Rubbert, A., Catanzaro, A., Moir, S., Monaco, J., Ehler, L., Mizell, S., Jackson, R., Li, Y., Romano, J. W., & Fauci, A. S. (1998). Natural killer cells from human immunodeficiency virus (HIV)-infected individuals are an important source of CC-chemokines and suppress HIV-1 entry and replication in vitro. The Journal of clinical investigation, 102(1), 223–231. [CrossRef]
- Olson, G. S., Murray, T. A., Jahn, A. N., Mai, D., Diercks, A. H., Gold, E. S., & Aderem, A. (2021). Type I interferon decreases macrophage energy metabolism during mycobacterial infection. Cell reports, 35(9), 109195. [CrossRef]
- Oshiumi, H., Matsumoto, M., Funami, K., Akazawa, T., & Seya, T. (2003). TICAM-1, an adaptor molecule that participates in Toll-like receptor 3-mediated interferon-beta induction. Nature immunology, 4(2), 161–167. [CrossRef]
- Pabst R. (2015). Mucosal vaccination by the intranasal route. Nose-associated lymphoid tissue (NALT)-Structure, function and species differences. Vaccine, 33(36), 4406–4413. [CrossRef]
- Padayachee, Y., Flicker, S., Linton, S., Cafferkey, J., Kon, O. M., Johnston, S. L., Ellis, A. K., Desrosiers, M., Turner, P., Valenta, R., & Scadding, G. K. (2021). Review: The Nose as a Route for Therapy. Part 2 Immunotherapy. Frontiers in allergy, 2, 668781. [CrossRef]
- Paidi, R. K., Jana, M., Mishra, R. K., Dutta, D., & Pahan, K. (2021). Selective Inhibition of the Interaction between SARS-CoV-2 Spike S1 and ACE2 by SPIDAR Peptide Induces Anti-Inflammatory Therapeutic Responses. Journal of immunology (Baltimore, Md. : 1950), 207(10), 2521–2533. [CrossRef]
- Paidi, R. K., Jana, M., Mishra, R. K., Dutta, D., Raha, S., & Pahan, K. (2021). ACE-2-interacting Domain of SARS-CoV-2 (AIDS) Peptide Suppresses Inflammation to Reduce Fever and Protect Lungs and Heart in Mice: Implications for COVID-19 Therapy. Journal of neuroimmune pharmacology : the official journal of the Society on NeuroImmune Pharmacology, 16(1), 59–70. [CrossRef]
- Paidi, R. K., Jana, M., Raha, S., McKay, M., Sheinin, M., Mishra, R. K., & Pahan, K. (2021). Eugenol, a Component of Holy Basil (Tulsi) and Common Spice Clove, Inhibits the Interaction Between SARS-CoV-2 Spike S1 and ACE2 to Induce Therapeutic Responses. Journal of neuroimmune pharmacology : the official journal of the Society on NeuroImmune Pharmacology, 16(4), 743–755. [CrossRef]
- Pan S, Manabe N, Yamaguchi Y (2021). 3D Structures of IgA, IgM, and Components. International Journal of Molecular Sciences; 22(23):12776. [CrossRef]
- Pantazi, I., Al-Qahtani, A. A., Alhamlan, F. S., Alothaid, H., Matou-Nasri, S., Sourvinos, G., Vergadi, E., & Tsatsanis, C. (2021). SARS-CoV-2/ACE2 Interaction Suppresses IRAK-M Expression and Promotes Pro-Inflammatory Cytokine Production in Macrophages. Frontiers in immunology, 12, 683800. [CrossRef]
- Paparisto, E., Hunt, N. R., Labach, D. S., Coleman, M. D., Di Gravio, E. J., Dodge, M. J., Friesen, N. J., Côté, M., Müller, A., Hoenen, T., & Barr, S. D. (2021). Interferon-Induced HERC5 Inhibits Ebola Virus Particle Production and Is Antagonized by Ebola Glycoprotein. Cells, 10(9), 2399. [CrossRef]
- Pardi N. , Weissman D. (2017) Nucleoside Modified mRNA Vaccines for Infectious Diseases. In: Kramps T., Elbers K. (eds) RNA Vaccines. Methods in Molecular Biology, vol 1499. Humana Press, New York, NY. [CrossRef]
- Park, A. , & Iwasaki, A. (2020). Type I and Type III Interferons - Induction, Signaling, Evasion, and Application to Combat COVID-19. Cell host & microbe, 27(6), 870–878. [CrossRef]
- Park, G. J., Osinski, A., Hernandez, G., Eitson, J. L., Majumdar, A., Tonelli, M., Henzler-Wildman, K., Pawłowski, K., Chen, Z., Li, Y., Schoggins, J. W., & Tagliabracci, V. S. (2022). The mechanism of RNA capping by SARS-CoV-2. Nature, 609(7928), 793–800. [CrossRef]
- Pavel Kovarik, Castiglia Virginia, Ivin Masa, Ebner Florian (2016), Type I Interferons in Bacterial Infections: A Balancing Act, Frontiers in Immunology, Vol. 7, pp 652, doi: https://www.frontiersin.org/article/10. Frontiers in Immunology; //www: , Vol. 7, pp 652, doi: https, 3389. [Google Scholar]
- Peng, M. Y., Liu, W. C., Zheng, J. Q., Lu, C. L., Hou, Y. C., Zheng, C. M., Song, J. Y., Lu, K. C., & Chao, Y. C. (2021). Immunological Aspects of SARS-CoV-2 Infection and the Putative Beneficial Role of Vitamin-D. International journal of molecular sciences, 22(10), 5251. [CrossRef]
- Pereda, R., González, D., Rivero, H. B., Rivero, J. C., Pérez, A., López, L., Mezquia, N., Venegas, R., Betancourt, J. R., & Domínguez, R. E. (2020). Therapeutic Effectiveness of Interferon-α2b Against COVID-19: The Cuban Experience. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research, 40(9), 438–442. [CrossRef]
- Perwitasari, O., Cho, H., Diamond, M. S., & Gale, M., Jr (2011). Inhibitor of κB kinase epsilon (IKK(epsilon)), STAT1, and IFIT2 proteins define novel innate immune effector pathway against West Nile virus infection. The Journal of biological chemistry, 286(52), 44412–44423. [CrossRef]
- Pestka, S., Krause, C. D., & Walter, M. R. (2004). Interferons, interferon-like cytokines, and their receptors. Immunological reviews, 202, 8–32. [CrossRef]
- Phillpotts, R. J., Scott, G. M., Higgins, P. G., Wallace, J., Tyrrell, D. A., & Gauci, C. L. (1983). An effective dosage regimen for prophylaxis against rhinovirus infection by intranasal administration of HuIFN-alpha 2. Antiviral research, 3(2), 121–136. [CrossRef]
- Pierangeli, A., Gentile, M., Oliveto, G., Frasca, F., Sorrentino, L., Matera, L., Nenna, R., Viscido, A., Fracella, M., Petrarca, L., D'Ettorre, G., Ceccarelli, G., Midulla, F., Antonelli, G., & Scagnolari, C. (2022). Comparison by Age of the Local Interferon Response to SARS-CoV-2 Suggests a Role for IFN-ε and -ω. Frontiers in immunology, 13, 873232. [CrossRef]
- Pierce, C. A., Preston-Hurlburt, P., Dai, Y., Aschner, C. B., Cheshenko, N., Galen, B., Garforth, S. J., Herrera, N. G., Jangra, R. K., Morano, N. C., Orner, E., Sy, S., Chandran, K., Dziura, J., Almo, S. C., Ring, A., Keller, M. J., Herold, K. C., & Herold, B. C. (2020). Immune responses to SARS-CoV-2 infection in hospitalized pediatric and adult patients. Science translational medicine, 12(564), eabd5487. [CrossRef]
- Pieretti, J. C., Rubilar, O., Weller, R. B., Tortella, G. R., & Seabra, A. B. (2021). Nitric oxide (NO) and nanoparticles - Potential small tools for the war against COVID-19 and other human coronavirus infections. Virus research, 291, 198202. [CrossRef]
- Planas, D., Saunders, N., Maes, P., Guivel-Benhassine, F., Planchais, C., Buchrieser, J., Bolland, W. H., Porrot, F., Staropoli, I., Lemoine, F., Péré, H., Veyer, D., Puech, J., Rodary, J., Baele, G., Dellicour, S., Raymenants, J., Gorissen, S., Geenen, C., Vanmechelen, B., … Schwartz, O. (2022). Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature, 602(7898), 671–675. [CrossRef]
- Poli, G. , Biswas, P., & Fauci, A. S. (1994). Interferons in the pathogenesis and treatment of human immunodeficiency virus infection. Antiviral research, 24(2-3), 221–233. [CrossRef]
- Pons, J. C., Lebon, P., Frydman, R., & Delfraissy, J. F. (1995). Pharmacokinetics of interferon-alpha in pregnant women and fetoplacental passage. Fetal diagnosis and therapy, 10(1), 7–10. [CrossRef]
- Posch, W., Bermejo-Jambrina, M., Lass-Flörl, C., & Wilflingseder, D. (2020). Role of Complement Receptors (CRs) on DCs in Anti-HIV-1 Immunity. Frontiers in immunology, 11, 572114. [CrossRef]
- Posch, W., Bermejo-Jambrina, M., Steger, M., Witting, C., Diem, G., Hörtnagl, P., Hackl, H., Lass-Flörl, C., Huber, L. A., Geijtenbeek, T., & Wilflingseder, D. (2021). Complement Potentiates Immune Sensing of HIV-1 and Early Type I Interferon Responses. mBio, 12(5), e0240821. [CrossRef]
- Prabhakara, C., Godbole, R., Sil, P., Jahnavi, S., Gulzar, S. E., van Zanten, T. S., Sheth, D., Subhash, N., Chandra, A., Shivaraj, A., Panikulam, P., U, I., Nuthakki, V. K., Puthiyapurayil, T. P., Ahmed, R., Najar, A. H., Lingamallu, S. M., Das, S., Mahajan, B., Vemula, P., … Mayor, S. (2021). Strategies to target SARS-CoV-2 entry and infection using dual mechanisms of inhibition by acidification inhibitors. PLoS pathogens, 17(7), e1009706. [CrossRef]
- Prieto-Dominguez, N., Parnell, C., & Teng, Y. (2019). Drugging the Small GTPase Pathways in Cancer Treatment: Promises and Challenges. Cells, 8(3), 255. [CrossRef]
- Prokop, J. W., Hartog, N. L., Chesla, D., Faber, W., Love, C. P., Karam, R., Abualkheir, N., Feldmann, B., Teng, L., McBride, T., Leimanis, M. L., English, B. K., Holsworth, A., Frisch, A., Bauss, J., Kalpage, N., Derbedrossian, A., Pinti, R. M., Hale, N., Mills, J., … Rajasekaran, S. (2021). High-Density Blood Transcriptomics Reveals Precision Immune Signatures of SARS-CoV-2 Infection in Hospitalized Individuals. Frontiers in immunology, 12, 694243. [CrossRef]
- Puhl, A. C., Gomes, G. F., Damasceno, S., Fritch, E. J., Levi, J. A., Johnson, N. J., Scholle, F., Premkumar, L., Hurst, B. L., Lee-Montiel, F., Veras, F. P., Batah, S. S., Fabro, A. T., Moorman, N. J., Yount, B. L., Dickmander, R. J., Baric, R. S., Pearce, K. H., Cunha, F. Q., Alves-Filho, J. C., … Ekins, S. (2022). Vandetanib Blocks the Cytokine Storm in SARS-CoV-2-Infected Mice. ACS omega, 7(36), 31935–31944. [CrossRef]
- Qi, Y. Y., Zhou, X. J., Cheng, F. J., Hou, P., Ren, Y. L., Wang, S. X., Zhao, M. H., Yang, L., Martinez, J., & Zhang, H. (2018). Increased autophagy is cytoprotective against podocyte injury induced by antibody and interferon-α in lupus nephritis. Annals of the rheumatic diseases, 77(12), 1799–1809. [CrossRef]
- Qiong Zhou, Virginia Chen, et al. (2020), “Interferon-a2b Treatment for COVID-19,” Frontiers in Immunology, Vol 11, Article 1601, doi:. [CrossRef]
- Quinones, Q. J., de Ridder, G. G., & Pizzo, S. V. (2008). GRP78: a chaperone with diverse roles beyond the endoplasmic reticulum. Histology and histopathology, 23(11), 1409–1416. [CrossRef]
- Radim Nencka, Jan Silhan, Martin Klima, Tomas Otava, Hugo Kocek, Petra Krafcikova, Evzen Boura (2022), Coronaviral RNA-methyltransferases: function, structure and inhibition, Nucleic Acids Research, 50(2), 635–650, https://doi.org/10. Nucleic Acids Research; //doi: , 50(2), 635–650, https, 1093. [Google Scholar]
- Radzikowska, U., Ding, M., Tan, G., Zhakparov, D., Peng, Y., Wawrzyniak, P., Wang, M., Li, S., Morita, H., Altunbulakli, C., Reiger, M., Neumann, A. U., Lunjani, N., Traidl-Hoffmann, C., Nadeau, K. C., O'Mahony, L., Akdis, C., & Sokolowska, M. (2020). Distribution of ACE2, CD147, CD26, and other SARS-CoV-2 associated molecules in tissues and immune cells in health and in asthma, COPD, obesity, hypertension, and COVID-19 risk factors. Allergy, 75(11), 2829–2845. [CrossRef]
- Raghavan, S., Kenchappa, D. B., & Leo, M. D. (2021, May 19). SARS-COV-2 spike protein induces degradation of junctional proteins that maintain endothelial barrier integrity. Frontiers. Retrieved December 5, 2022, from https://www.frontiersin.org/articles/10. SARS-COV-2 spike protein induces degradation of junctional proteins that maintain endothelial barrier integrity; //www: Frontiers. Retrieved December 5, 2022, from https, 3389. [Google Scholar]
- Rah, B., Rather, R. A., Bhat, G. R., Baba, A. B., Mushtaq, I., Farooq, M., Yousuf, T., Dar, S. B., Parveen, S., Hassan, R., Mohammad, F., Qassim, I., Bhat, A., Ali, S., Zargar, M. H., & Afroze, D. (2022). JAK/STAT Signaling: Molecular Targets, Therapeutic Opportunities, and Limitations of Targeted Inhibitions in Solid Malignancies. Frontiers in pharmacology, 13, 821344. [CrossRef]
- Ramasamy, S., & Subbian, S. (2021). Critical Determinants of Cytokine Storm and Type I Interferon Response in COVID-19 Pathogenesis. Clinical microbiology reviews, 34(3), e00299-20. [CrossRef]
- Rashid, F., Xie, Z., Suleman, M., Shah, A., Khan, S., & Luo, S. (2022). Roles and functions of SARS-CoV-2 proteins in host immune evasion. Frontiers in immunology, 13, 940756. [CrossRef]
- Rashid, F., Xie, Z., Suleman, M., Shah, A., Khan, S., & Luo, S. (2022). Roles and functions of SARS-CoV-2 proteins in host immune evasion. Frontiers in immunology, 13, 940756. [CrossRef]
- Ratajczak, M. Z., & Kucia, M. (2020). SARS-CoV-2 infection and overactivation of Nlrp3 inflammasome as a trigger of cytokine "storm" and risk factor for damage of hematopoietic stem cells. Leukemia, 34(7), 1726–1729. [CrossRef]
- Rauchhaus, J., Robinson, J., Monti, L., & Di Antonio, M. (2022). G-quadruplexes Mark Sites of Methylation Instability Associated with Ageing and Cancer. Genes, 13(9), 1665. [CrossRef]
- Rautela, J., & Huntington, N. D. (2017). IL-15 signaling in NK cell cancer immunotherapy. Current opinion in immunology, 44, 1–6. [CrossRef]
- Rayner, J. O., Roberts, R. A., Kim, J., Poklepovic, A., Roberts, J. L., Booth, L., & Dent, P. (2020). AR12 (OSU-03012) suppresses GRP78 expression and inhibits SARS-CoV-2 replication. Biochemical pharmacology, 182, 114227. [CrossRef]
- Reizis B. (2019). Plasmacytoid Dendritic Cells: Development, Regulation, and Function. Immunity, 50(1), 37–50. [CrossRef]
- Ren, Y., Cui, G., & Gao, Y. (2021). Research progress on inflammatory mechanism of primary Sjögren syndrome. Zhejiang da xue xue bao. Yi xue ban = Journal of Zhejiang University. Medical sciences, 50(6), 783–794. [CrossRef]
- Rezaie, A., Melmed, G.Y., Leite, G. et al. Endotracheal Application of Ultraviolet A Light in Critically Ill Patients with Severe Acute Respiratory Syndrome Coronavirus 2: A First-in-Human Study. Adv Ther 38, 4556–4568 (2021). [CrossRef]
- Richardson, G; recent developments and future strategies: , & Tate, B. (2000). Hormonal and pharmacological manipulation of the circadian clock.
- Ricke D. O. (2021). Two Different Antibody-Dependent Enhancement (ADE) Risks for SARS-CoV-2 Antibodies. Frontiers in immunology, 12, 640093. [CrossRef]
- Rios, C. I., Cassatt, D. R., Hollingsworth, B. A., Satyamitra, M. M., Tadesse, Y. S., Taliaferro, L. P., Winters, T. A., & DiCarlo, A. L. (2021). Commonalities Between COVID-19 and Radiation Injury. Radiation research, 195(1), 1–24. [CrossRef]
- Robbins, M. A., Maksumova, L., Pocock, E., & Chantler, J. K. (2003). Nuclear factor-kappaB translocation mediates double-stranded ribonucleic acid-induced NIT-1 beta-cell apoptosis and up-regulates caspase-12 and tumor necrosis factor receptor-associated ligand (TRAIL). Endocrinology, 144(10), 4616–4625. [CrossRef]
- Rodriguez, J. J., Parisien, J. P., & Horvath, C. M. (2002). Nipah virus V protein evades alpha and gamma interferons by preventing STAT1 and STAT2 activation and nuclear accumulation. Journal of virology, 76(22), 11476–11483. [CrossRef]
- Roers, A. , Hiller, B., & Hornung, V. (2016). Recognition of Endogenous Nucleic Acids by the Innate Immune System. Immunity, 44(4), 739–754. [CrossRef]
- Romano, M., Ruggiero, A., Squeglia, F., Maga, G., & Berisio, R. (2020). A Structural View of SARS-CoV-2 RNA Replication Machinery: RNA Synthesis, Proofreading and Final Capping. Cells, 9(5), 1267. [CrossRef]
- Roskoski R., Jr (2016). Janus kinase (JAK) inhibitors in the treatment of inflammatory and neoplastic diseases. Pharmacological research, 111, 784–803. [CrossRef]
- Roumenina, L. T., Daugan, M. V., Noé, R., Petitprez, F., Vano, Y. A., Sanchez-Salas, R., Becht, E., Meilleroux, J., Clec'h, B. L., Giraldo, N. A., Merle, N. S., Sun, C. M., Verkarre, V., Validire, P., Selves, J., Lacroix, L., Delfour, O., Vandenberghe, I., Thuilliez, C., Keddani, S., … Fridman, W. H. (2019). Tumor Cells Hijack Macrophage-Produced Complement C1q to Promote Tumor Growth. Cancer immunology research, 7(7), 1091–1105. [CrossRef]
- Rubio-Casillas, A., Redwan, E. M., & Uversky, V. N. (2022). SARS-CoV-2: A Master of Immune Evasion. Biomedicines, 10(6), 1339. [CrossRef]
- Saadeldin, M. K., Abdel-Aziz, A. K., & Abdellatif, A. (2021). Dendritic cell vaccine immunotherapy; the beginning of the end of cancer and COVID-19. A hypothesis. Medical hypotheses, 146, 110365. [CrossRef]
- Safadi DE, Lebeau G, Lagrave A, Mélade J, Grondin L, Rosanaly S, Begue F, Hoareau M, Veeren B, Roche M, Hoarau J-J, Meilhac O, Mavingui P, Desprès P, Viranaïcken W, Krejbich-Trotot P. Extracellular Vesicles Are Conveyors of the NS1 Toxin during Dengue Virus and Zika Virus Infection. Viruses. 2023; 15(2):364. [CrossRef]
- Saito, H. Saito, H., Takenaka, H., Yoshida, S., Tsubokawa, T., Ogata, A., Imanishi, F., & Imanishi, J. (1985). Prevention from naturally acquired viral respiratory infection by interferon nasal spray. Rhinology.
- Saito, S., Ogino, T., Miyajima, N., Kato, A., & Kohase, M. (2002). Dephosphorylation failure of tyrosine-phosphorylated STAT1 in IFN-stimulated Sendai virus C protein-expressing cells. Virology, 293(2), 205–209. [CrossRef]
- Sakai, K., Ueda, A., Hasegawa, M., & Ueda, Y. (2018). Efficacy and safety of interferon alpha for essential thrombocythemia during pregnancy: two cases and a literature review. International journal of hematology, 108(2), 203–207. [CrossRef]
- Sakai, S., & Shichita, T. (2022). Role of alarmins in poststroke inflammation and neuronal repair. Seminars in immunopathology, 10.1007/s00281-022-00961-5. Advance online publication. [CrossRef]
- Salerno, F., Guislain, A., Cansever, D., & Wolkers, M. C. (2016). TLR-Mediated Innate Production of IFN-γ by CD8+ T Cells Is Independent of Glycolysis. Journal of immunology (Baltimore, Md. : 1950), 196(9), 3695–3705. [CrossRef]
- Saliu, T. P. Saliu, T. P., Umar, H. I., Ogunsile, O. J., Okpara, M. O., Yanaka, N., & Elekofehinti, O. O. (2021). Molecular docking and pharmacokinetic studies of phytocompounds from nigerian medicinal plants as promising inhibitory agents against SARS-CoV-2 methyltransferase (nsp16). Journal of Genetic Engineering and Biotechnology.
- Sampaio, N. G., Chauveau, L., Hertzog, J., Bridgeman, A., Fowler, G., Moonen, J. P., Dupont, M., Russell, R. A., Noerenberg, M., & Rehwinkel, J. (2021). The RNA sensor MDA5 detects SARS-CoV-2 infection. Scientific reports, 11(1), 13638. [CrossRef]
- Sánchez-Aparicio, M. T., Garcin, D., Rice, C. M., Kolakofsky, D., García-Sastre, A., & Baum, A. (2017). Loss of Sendai virus C protein leads to accumulation of RIG-I immunostimulatory defective interfering RNA. The Journal of general virology, 98(6), 1282–1293. [CrossRef]
- Sangaletti, P., Doe, J., Gatti, A., Arvay, C., Giuliani, L., & Lettner, H. (2022). SARS-CoV-2 and the Vaccination Hype. International Journal of Vaccine Theory, Practice, and Research, 2(1), 173–207. https://doi.org/10. International Journal of Vaccine Theory, Practice, and Research, 2; //doi: (1), 173–207. https, 5609. [Google Scholar] [CrossRef]
- Santer, D.M., Li, D., Ghosheh, Y. et al. Interferon-λ treatment accelerates SARS-CoV-2 clearance despite age-related delays in the induction of T cell immunity. Nat Commun 13, 6992 (2022). [CrossRef]
- Santerre, M., Arjona, S. P., Allen, C. N., Shcherbik, N., & Sawaya, B. E. (2021). Why do SARS-CoV-2 NSPs rush to the ER? Journal of neurology, 268(6), 2013–2022. [CrossRef]
- Santini, S. M., Lapenta, C., Santodonato, L., D'Agostino, G., Belardelli, F., & Ferrantini, M. (2009). IFN-alpha in the generation of dendritic cells for cancer immunotherapy. Handbook of experimental pharmacology, (188), 295–317. [CrossRef]
- Saramago, M., Bárria, C., Costa, V. G., Souza, C. S., Viegas, S. C., Domingues, S.,... & Matos, R. G. (2021). New targets for drug design: importance of nsp14/nsp10 complex formation for the 3’-5’exoribonucleolytic activity on SARS-CoV-2. The FEBS journal, 288(17), 5130-5147. [CrossRef]
- Satarker, S., Tom, A. A., Shaji, R. A., Alosious, A., Luvis, M., & Nampoothiri, M. (2021). JAK-STAT Pathway Inhibition and their Implications in COVID-19 Therapy. Postgraduate medicine, 133(5), 489–507. [CrossRef]
- Savulescu, J. , Giubilini, A., & Danchin, M. (2021). Global Ethical Considerations Regarding Mandatory Vaccination in Children. The Journal of pediatrics, 231, 10–16. [CrossRef]
- Schaefer, A., Unterberger, C., Frankenberger, M., Lohrum, M., Staples, K. J., Werner, T., Stunnenberg, H., & Ziegler-Heitbrock, L. (2009). Mechanism of interferon-gamma mediated down-regulation of interleukin-10 gene expression. Molecular Immunology, 46(7), 1351–1359. [CrossRef]
- Schardey, J., Globig, A. M., Janssen, C., Hofmann, M., Manegold, P., Thimme, R., & Hasselblatt, P. (2019). Vitamin D Inhibits Pro-Inflammatory T Cell Function in Patients With Inflammatory Bowel Disease. Journal of Crohn's & colitis, 13(12), 1546–1557. [CrossRef]
- Schmidt, A. , Rothenfusser, S., & Hopfner, K. P. (2012). Sensing of viral nucleic acids by RIG-I: from translocation to translation. European journal of cell biology, 91(1), 78–85. [CrossRef]
- Schmidt, N., Lareau, C. A., Keshishian, H., Ganskih, S., Schneider, C., Hennig, T., Melanson, R., Werner, S., Wei, Y., Zimmer, M., Ade, J., Kirschner, L., Zielinski, S., Dölken, L., Lander, E. S., Caliskan, N., Fischer, U., Vogel, J., Carr, S. A., Bodem, J., … Munschauer, M. (2021). The SARS-CoV-2 RNA-protein interactome in infected human cells. Nature microbiology, 6(3), 339–353. [CrossRef]
- Schoenmaker, L., Witzigmann, D., Kulkarni, J. A., Verbeke, R., Kersten, G., Jiskoot, W., & Crommelin, D. (2021). mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. International journal of pharmaceutics, 601, 120586. [CrossRef]
- Schomacker, H., Hebner, R. M., Boonyaratanakornkit, J., Surman, S., Amaro-Carambot, E., Collins, P. L., & Schmidt, A. C. (2012). The C proteins of human parainfluenza virus type 1 block IFN signaling by binding and retaining Stat1 in perinuclear aggregates at the late endosome. PloS one, 7(2), e28382. [CrossRef]
- Schreiber G. (2020). The Role of Type I Interferons in the Pathogenesis and Treatment of COVID-19. Frontiers in immunology, 11, 595739. [CrossRef]
- Scott, L. J., & Perry, C. M. (2002). Interferon-alpha-2b plus ribavirin: a review of its use in the management of chronic hepatitis C. Drugs, 62(3), 507–556. [CrossRef]
- Sebők C, Walmsley S, Tráj P, Mackei M, Vörösházi J, et al. (2022) Immunomodulatory effects of chicken cathelicidin-2 on a primary hepatic cell co-culture model. PLOS ONE 17(10): e0275847. [CrossRef]
- Segreto, R., & Deigin, Y. (2021). The genetic structure of SARS-CoV-2 does not rule out a laboratory origin: SARS-COV-2 chimeric structure and furin cleavage site might be the result of genetic manipulation. BioEssays : news and reviews in molecular, cellular and developmental biology, 43(3), e2000240. [CrossRef]
- Seneff, S; //ijvtpr.com/index: , Nigh, G. (2021). Worse than the disease? Reviewing some possible unintended consequences of the mRNA vaccines against COVID-19. International Journal of Vaccine Theory, Practice, and Research, 2(1): 38-79. Retrieved from https.
- Seneff, S., Nigh, G., Kyriakopoulos, A. M., & McCullough, P. A. (2022). Innate immune suppression by SARS-CoV-2 mRNA vaccinations: The role of G-quadruplexes, exosomes, and MicroRNAs. Food and chemical toxicology : an international journal published for the British Industrial Biological Research Association, 164, 113008. [CrossRef]
- Setaro, A. C., & Gaglia, M. M. (2021). All hands on deck: SARS-CoV-2 proteins that block early anti-viral interferon responses. Current research in virological science, 2, 100015. [CrossRef]
- Sette, A., & Crotty, S. (2021). Adaptive immunity to SARS-CoV-2 and COVID-19. Cell, 184(4), 861–880. [CrossRef]
- Seya, T., Matsumoto, M., Ebihara, T., & Oshiumi, H. (2009). Functional evolution of the TICAM-1 pathway for extrinsic RNA sensing. Immunological reviews, 227(1), 44–53. [CrossRef]
- Shabunin,S. ,Gritsyuk,V.,Vostroilova,G.,Shabanov,D.,Khokhlova,N. & Korchagina,A.(2022).Study of Mutagenic and Antitoxic Properties of Gentabiferon-B. Macedonian Veterinary Review,45(1) 79-87. [CrossRef]
- Shafiq, A., Zubair, F., Ambreen, A., Suleman, M., Yousafi, Q., Rasul Niazi, Z., Anwar, Z., Khan, A., Mohammad, A., & Wei, D. Q. (2022). Investigation of the binding and dynamic features of A.30 variant revealed higher binding of RBD for hACE2 and escapes the neutralizing antibody: A molecular simulation approach. Computers in biology and medicine, 146, 105574. [CrossRef]
- Shan, C., Miao, S., Liu, C. et al. Induction of macrophage pyroptosis-related factors by pathogenic E. coli high pathogenicity island (HPI) in Yunnan Saba pigs. BMC Vet Res 17, 114 (2021). [CrossRef]
- Shao, S. C., Wang, C.H., Chang, K.C., Hung, M.J., Chen, H.Y., & Liao, S.C. (2021). Guillain-Barré Syndrome Associated with COVID-19 Vaccination. Emerg Infect Dis, 27(12): 3175-3178. [CrossRef]
- Shaver, K. A., Croom-Perez, T. J., & Copik, A. J. (2021). Natural Killer Cells: The Linchpin for Successful Cancer Immunotherapy. Frontiers in immunology, 12, 679117. [CrossRef]
- Shaw, M. L., García-Sastre, A., Palese, P., & Basler, C. F. (2004). Nipah virus V and W proteins have a common STAT1-binding domain yet inhibit STAT1 activation from the cytoplasmic and nuclear compartments, respectively. Journal of virology, 78(11), 5633–5641. [CrossRef]
- Sheng, L., Chen, X., Wang, Q., Lyu, S., & Li, P. (2020). Interferon-α2b enhances survival and modulates transcriptional profiles and the immune response in melanoma patients treated with dendritic cell vaccines. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie, 125, 109966. [CrossRef]
- Shepherd, J., Brodin, H., Cave, C., Waugh, N., Price, A., & Gabbay, J. (2004). Pegylated interferon alpha-2a and -2b in combination with ribavirin in the treatment of chronic hepatitis C: a systematic review and economic evaluation. Health technology assessment (Winchester, England), 8(39), iii–125. [CrossRef]
- Shepherd, J., Jones, J., Hartwell, D., Davidson, P., Price, A., & Waugh, N. (2007). Interferon alpha (pegylated and non-pegylated) and ribavirin for the treatment of mild chronic hepatitis C: a systematic review and economic evaluation. Health technology assessment (Winchester, England), 11(11), 1–iii. [CrossRef]
- Sherer, N. M., & Mothes, W. (2008). Cytonemes and tunneling nanotubules in cell-cell communication and viral pathogenesis. Trends in cell biology, 18(9), 414–420. [CrossRef]
- Shi, J., Zhou, J., Zhang, X., Hu, W., Zhao, J. F., Wang, S., Wang, F. S., & Zhang, J. Y. (2021). Single-Cell Transcriptomic Profiling of MAIT Cells in Patients With COVID-19. Frontiers in immunology, 12, 700152. [CrossRef]
- Shi, Z. , & Puyo, C.A. (2020). Acetylcysteine to Combat COVID- 19. [CrossRef]
- Shibabaw T, Molla MD, Teferi B, Ayelign B. Role of IFN and Complements System: Innate Immunity in SARS-CoV-2. J Inflamm Res. 2020; 13, 507–518. [CrossRef]
- Shibabaw, T., Molla, M. D., Teferi, B., & Ayelign, B. (2020). Role of IFN and Complements System: Innate Immunity in SARS-CoV-2. Journal of inflammation research, 13, 507–518. [CrossRef]
- Shimoyama, S. , Kanisawa, Y., Ono, K., Souri, M., & Ichinose, A. (2021). First and fatal case of autoimmune acquired factor XIII/13 deficiency after COVID-19/SARS-CoV-2 vaccination. Am J Hematol, 1–3. [CrossRef]
- Shin, J., Toyoda, S., Fukuhara, A., & Shimomura, I. (2022). GRP78, a Novel Host Factor for SARS-CoV-2: The Emerging Roles in COVID-19 Related to Metabolic Risk Factors. Biomedicines, 10(8), 1995. [CrossRef]
- Shin, J., Toyoda, S., Nishitani, S., Fukuhara, A., Kita, S., Otsuki, M., & Shimomura, I. (2021). Possible Involvement of Adipose Tissue in Patients With Older Age, Obesity, and Diabetes With SARS-CoV-2 Infection (COVID-19) via GRP78 (BIP/HSPA5): Significance of Hyperinsulinemia Management in COVID-19. Diabetes, 70(12), 2745–2755. [CrossRef]
- Shirazi, Y. , & Pitha, P. M. (1993). Interferon alpha-mediated inhibition of human immunodeficiency virus type 1 provirus synthesis in T-cells. Virology, 193(1), 303–312. [CrossRef]
- Sidahmed, A. M., León, A. J., Bosinger, S. E., Banner, D., Danesh, A., Cameron, M. J., & Kelvin, D. J. (2012). CXCL10 contributes to p38-mediated apoptosis in primary T lymphocytes in vitro. Cytokine, 59(2), 433–441. [CrossRef]
- Singh, P. K., Kulsum, U., Rufai, S. B., Mudliar, S. R., & Singh, S. (2020). Mutations in SARS-CoV-2 Leading to Antigenic Variations in Spike Protein: A Challenge in Vaccine Development. Journal of laboratory physicians, 12(2), 154–160. [CrossRef]
- Skene, D. J. Skene, D. J., Deacon, S., & Arendt, J. (1996). Use of melatonin in circadian rhythm disorders and following phase shifts. Acta neurobiologiae experimentalis.
- Smith G. L. (2018). Vaccinia Virus Protein C6: A Multifunctional Interferon Antagonist. Advances in experimental medicine and biology, 1052, 1–7. [CrossRef]
- Smith, B. L., Chen, G., Wilke, C. O., & Krug, R. M. (2018). Avian Influenza Virus PB1 Gene in H3N2 Viruses Evolved in Humans To Reduce Interferon Inhibition by Skewing Codon Usage toward Interferon-Altered tRNA Pools. mBio, 9(4), e01222-18. [CrossRef]
- Smolensky, M. H., Sackett-Lundeen, L. L., & Portaluppi, F. (2015). Nocturnal light pollution and underexposure to daytime sunlight: Complementary mechanisms of circadian disruption and related diseases. Chronobiology international, 32(8), 1029–1048. [CrossRef]
- Sodano, F., Gazzano, E., Fruttero, R., & Lazzarato, L. (2022). NO in Viral Infections: Role and Development of Antiviral Therapies. Molecules (Basel, Switzerland), 27(7), 2337. [CrossRef]
- Sodeifian, F, Nikfarjam, M, Kian, N, Mohamed, K, Rezaei, N. The role of type I interferon in the treatment of COVID-19. J Med Virol. 2021; 94, 63. [CrossRef]
- Sodeifian, F., Nikfarjam, M., Kian, N., Mohamed, K., & Rezaei, N. (2022). The role of type I interferon in the treatment of COVID-19. Journal of medical virology, 94(1), 63–81. [CrossRef]
- Solórzano, A., Webby, R. J., Lager, K. M., Janke, B. H., García-Sastre, A., & Richt, J. A. (2005). Mutations in the NS1 protein of swine influenza virus impair anti-interferon activity and confer attenuation in pigs. Journal of virology, 79(12), 7535–7543. [CrossRef]
- Solov'ev V. D. (1969). The results of controlled observations on the prophylaxis of influenza with interferon. Bulletin of the World Health Organization.
- Soy, M., Keser, G., Atagündüz, P., Tabak, F., Atagündüz, I., & Kayhan, S. (2020). Cytokine storm in COVID-19: pathogenesis and overview of anti-inflammatory agents used in treatment. Clinical rheumatology, 39(7), 2085–2094. [CrossRef]
- Sperber, S. J., Levine, P. A., Innes, D. J., Mills, S. E., & Hayden, F. G. (1988). Tolerance and efficacy of intranasal administration of recombinant beta serine interferon in healthy adults. The Journal of infectious diseases, 158(1), 166–175. [CrossRef]
- Stanifer, M. L., Guo, C., Doldan, P., & Boulant, S. (2020). Importance of Type I and III Interferons at Respiratory and Intestinal Barrier Surfaces. Frontiers in immunology, 11, 608645. [CrossRef]
- Stawicki SP. Could tracheo-bronchial ultraviolet C irradiation be a valuable adjunct in the management of severe COVID-19 pulmonary infections?. Int J Acad Med [serial online] 2020 [cited 2022 Feb 28]; 6, 156–8.
- Steinman, R. M., Granelli-Piperno, A., Pope, M., Trumpfheller, C., Ignatius, R., Arrode, G., Racz, P., & Tenner-Racz, K. (2003). The interaction of immunodeficiency viruses with dendritic cells. Current topics in microbiology and immunology, 276, 1–30. [CrossRef]
- Sträter, J., & Möller, P. (2004). TRAIL and viral infection. Vitamins and hormones, 67, 257–274. [CrossRef]
- Su, C., & Zheng, C. (2017). Herpes Simplex Virus 1 Abrogates the cGAS/STING-Mediated Cytosolic DNA-Sensing Pathway via Its Virion Host Shutoff Protein, UL41. Journal of virology, 91(6), e02414-16. [CrossRef]
- Sugai, A. , Sato, H., Takayama, I., Yoneda, M., & Kai, C. (2017). Nipah and Hendra Virus Nucleoproteins Inhibit Nuclear Accumulation of Signal Transducer and Activator of Transcription 1 (STAT1) and STAT2 by Interfering with Their Complex Formation. Journal of virology, 91(21), e01136-17. [CrossRef]
- Sumi, T. , Nagahisa, Y., Matsuura, K., Sekikawa, M., Yamada, Y., Nakata, H., & Chiba, H. (2021). Lung squamous cell carcinoma with hemoptysis after vaccination with tozinameran (BNT162b2, Pfizer-BioNTech). Thorac Cancer, 12, 3072–3075.
- Sun, J., Tang, X., Bai, R., Liang, C., Zeng, L., Lin, H., Yuan, R., Zhou, P., Huang, X., Xiong, Q., Peng, J., Cui, F., Ke, B., Su, J., Liu, Z., Lu, J., Tian, J., Sun, R., & Ke, C. (2020). The kinetics of viral load and antibodies to SARS-CoV-2. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases, 26(12), 1690.e1–1690.e4. [CrossRef]
- Sun, X., Belser, J. A., & Maines, T. R. (2020). Adaptation of H9N2 Influenza Viruses to Mammalian Hosts: A Review of Molecular Markers. Viruses, 12(5), 541. [CrossRef]
- Sung, R. Y., Yin, J., Oppenheimer, S. J., Tam, J. S., & Lau, J. (1993). Treatment of respiratory syncytial virus infection with recombinant interferon alfa-2a. Archives of disease in childhood, 69(4), 440–442. [CrossRef]
- Swiecki, M., & Colonna, M. (2011). Type I interferons: diversity of sources, production pathways and effects on immune responses. Current opinion in virology, 1(6), 463–475. [CrossRef]
- Takaki, H., Oshiumi, H., Sasai, M., Kawanishi, T., Matsumoto, M., & Seya, T. (2009). Oligomerized TICAM-1 (TRIF) in the cytoplasm recruits nuclear BS69 to enhance NF-kappaB activation and type I IFN induction. European journal of immunology, 39(12), 3469–3476. [CrossRef]
- Tan, X. , Sun, L., Chen, J., & Chen, Z. J. (2018). Detection of Microbial Infections Through Innate Immune Sensing of Nucleic Acids. Annual review of microbiology, 72, 447–478. [CrossRef]
- Tang, Y., Zhang, P., Liu, Q., Cao, L., & Xu, J. (2022). Pyroptotic Patterns in Blood Leukocytes Predict Disease Severity and Outcome in COVID-19 Patients. Frontiers in immunology, 13, 888661. [CrossRef]
- Tannock, G. A., Gillett, S. M., Gillett, R. S., Barry, R. D., Hensley, M. J., Herd, R., Reid, A. L., & Saunders, N. A. (1988). A study of intranasally administered interferon A (rIFN-alpha 2A) for the seasonal prophylaxis of natural viral infections of the upper respiratory tract in healthy volunteers. Epidemiology and infection, 101(3), 611–621. [CrossRef]
- Tannock, G. A., Gillett, S. M., Gillett, R. S., Barry, R. D., Hensley, M. J., Herd, R., Reid, A. L., & Saunders, N. A. (1988). A study of intranasally administered interferon A (rIFN-alpha 2A) for the seasonal prophylaxis of natural viral infections of the upper respiratory tract in healthy volunteers. Epidemiology and infection, 101(3), 611–621. [CrossRef]
- Tarasova O, Ivanov S, Filimonov DA, Poroikov V. Data and Text Mining Help Identify Key Proteins Involved in the Molecular Mechanisms Shared by SARS-CoV-2 and HIV-1. Molecules. 2020; 25(12):2944. [CrossRef]
- Taylor, M. D., Korth, M. J., & Katze, M. G. (1998). Interferon treatment inhibits the replication of simian immunodeficiency virus at an early stage: evidence for a block between attachment and reverse transcription. Virology, 241(1), 156–162. [CrossRef]
- Time Magazine (1980), The Big IF in Cancer, https://time. 6855.
- Time Magazine (2023), A One-Shot COVID-19 Treatment Shows Promise, https://time. 6254.
- Timothy P. Riley, Hui-Ting Chou, Ruozhen Hu, Krzysztof P. Bzymek, Ana R. Correia, Alexander C. Partin, Danqing Li, Danyang Gong, Zhulun Wang, Xinchao Yu, Paolo Manzanillo and Fernando Garces (2021), Enhancing the Prefusion Conformational Stability of SARS-CoV-2 Spike Protein Through Structure-Guided Design, Frontiers in Immunology, doi:. [CrossRef]
- Tintoré M. (2009). New options for early treatment of multiple sclerosis. Journal of the neurological sciences, 277 Suppl 1, S9–S11. [CrossRef]
- Tonutti, A. , Motta, F., Ceribelli, A., Isailovic, N., Selmi, C., & De Santis, M. (2022). Anti-MDA5 Antibody Linking COVID-19, Type I Interferon, and Autoimmunity: A Case Report and Systematic Literature Review. Frontiers in immunology, 13, 937667. [CrossRef]
- Tresse, E., Riera-Ponsati, L., Jaberi, E., Sew, W., Ruscher, K., & Issazadeh-Navikas, S. (2021). IFN-β rescues neurodegeneration by regulating mitochondrial fission via STAT5, PGAM5, and Drp1. The EMBO journal, 40(11), e106868. [CrossRef]
- Tsuji, R. F., Geba, G. P., Wang, Y., Kawamoto, K., Matis, L. A., & Askenase, P. W. (1997). Required early complement activation in contact sensitivity with generation of local C5-dependent chemotactic activity, and late T cell interferon gamma: a possible initiating role of B cells. The Journal of experimental medicine, 186(7), 1015–1026. [CrossRef]
- Tufan, A. B., Lazarow, K., Kolesnichenko, M., Sporbert, A., von Kries, J. P., & Scheidereit, C. (2022). tsg101 associates with parp1 and is essential for parylation and dna damage-induced nf-κb activation. The EMBO Journal. [CrossRef]
- Tunbak, H., Enriquez-Gasca, R., Tie, C., Gould, P. A., Mlcochova, P., Gupta, R. K., Fernandes, L., Holt, J., van der Veen, A. G., Giampazolias, E., Burns, K. H., Maillard, P. V., & Rowe, H. M. (2020). The HUSH complex is a gatekeeper of type I interferon through epigenetic regulation of LINE-1s. Nature communications, 11(1), 5387. [CrossRef]
- Uriu, K. , Kimura, I., Shirakawa, K., Takaori-Kondo, A., Nakada, T.A., Kaneda, A., The Genotype to Phenotype Japan (G2P-Japan) Consortium, Nakagawa, S., & Sato, K. (2021). Ineffective neutralization of the SARS-CoV-2 Mu variant by convalescent and vaccine sera. bioRxiv, 2021.09.06.459005. [CrossRef]
- van den Berg, D. F., & Te Velde, A. A. (2020). Severe COVID-19: NLRP3 Inflammasome Dysregulated. Frontiers in immunology, 11, 1580. [CrossRef]
- Vanderheiden, A., Ralfs, P., Chirkova, T., Upadhyay, A. A., Zimmerman, M. G., Bedoya, S., Aoued, H., Tharp, G. M., Pellegrini, K. L., Manfredi, C., Sorscher, E., Mainou, B., Lobby, J. L., Kohlmeier, J. E., Lowen, A. C., Shi, P. Y., Menachery, V. D., Anderson, L. J., Grakoui, A., Bosinger, S. E., … Suthar, M. S. (2020). Type I and Type III Interferons Restrict SARS-CoV-2 Infection of Human Airway Epithelial Cultures. Journal of virology, 94(19), e00985-20. [CrossRef]
- Vanderheiden, A. , Ralfs, P., Chirkova, T., Upadhyay, A. A., Zimmerman, M. G., Bedoya, S., Aoued, H., Tharp, G. M., Pellegrini, K. L., Manfredi, C., Sorscher, E., Mainou, B., Lobby, J. L., Kohlmeier, J. E., Lowen, A. C., Shi, P. Y., Menachery, V. D., Anderson, L. J., Grakoui, A., Bosinger, S. E., … Suthar, M. S. (2020). Type I and Type III Interferons Restrict SARS-CoV-2 Infection of Human Airway Epithelial Cultures. Journal of virology, 94(19), e00985-20. [CrossRef]
- Vanderheiden, A., Ralfs, P., Chirkova, T., Upadhyay, A. A., Zimmerman, M. G., Bedoya, S., Aoued, H., Tharp, G. M., Pellegrini, K. L., Manfredi, C., Sorscher, E., Mainou, B., Lobby, J. L., Kohlmeier, J. E., Lowen, A. C., Shi, P. Y., Menachery, V. D., Anderson, L. J., Grakoui, A., Bosinger, S. E., … Suthar, M. S. (2020). Type I and Type III Interferons Restrict SARS-CoV-2 Infection of Human Airway Epithelial Cultures. Journal of virology, 94(19), e00985-20. [CrossRef]
- Vandoorn, E., Stadejek, W., Parys, A., Chepkwony, S., Chiers, K., & Van Reeth, K. (2022). Pathobiology of an NS1-Truncated H3N2 Swine Influenza Virus Strain in Pigs. Journal of virology, 96(11), e0051922. [CrossRef]
- Varble, A. J., Ried, C. D., Hammond, W. J., Marquis, K. A., Woodruff, M. C., & Ferran, M. C. (2016). The vesicular stomatitis virus matrix protein inhibits NF-κB activation in mouse L929 cells. Virology, 499, 99–104. [CrossRef]
- Vidy, A., Chelbi-Alix, M., & Blondel, D. (2005). Rabies virus P protein interacts with STAT1 and inhibits interferon signal transduction pathways. Journal of virology, 79(22), 14411–14420. [CrossRef]
- Vignau, J. , Karila, L., Costisella, O., & Canva, V. (2005). Hépatite C, Interféron alpha et dépression: principales hypothèses physiopathologiques [Hepatitis C, interferon a and depression: main physiopathologic hypothesis]. L'Encephale, 31(3), 349–357. [CrossRef]
- Villarreal, L; Kolb V (ed): P. (2015) Virolution Can Help Us Understand the Origin of Life. In; ISBN 13-978-1-4665-8462-4.
- Villarreal, L. P., Witzany, G. (2021) Social networking of quasi-species consortia drive virolution via persistence. AIMS Microbiology, 7(2): 138-162. [CrossRef]
- Vishnubalaji, R. , Shaath, H., & Alajez, N. M. (2020). Protein Coding and Long Noncoding RNA (lncRNA) Transcriptional Landscape in SARS-CoV-2 Infected Bronchial Epithelial Cells Highlight a Role for Interferon and Inflammatory Response. Genes, 11(7), 760. [CrossRef]
- Vital, E. M., Merrill, J. T., Morand, E. F., Furie, R. A., Bruce, I. N., Tanaka, Y., Manzi, S., Kalunian, K. C., Kalyani, R. N., Streicher, K., Abreu, G., & Tummala, R. (2022). Anifrolumab efficacy and safety by type I interferon gene signature and clinical subgroups in patients with SLE: post hoc analysis of pooled data from two phase III trials. Annals of the rheumatic diseases, 81(7), 951–961. [CrossRef]
- W. A. Robinson, T. I. Mughal, M. R. Thomas, Melinda Johnson, R. J. Spiegel (1986), Treatment of Metastatic Malignant Melanoma with Recombinant Interferon Alpha 2, Immunobiology, Vol 172, 3-5, pp 275-282,. [CrossRef]
- Wan, D., Jiang, W., & Hao, J. (2020). Research Advances in How the cGAS-STING Pathway Controls the Cellular Inflammatory Response. Frontiers in immunology, 11, 615. [CrossRef]
- Wang, C. , Shan, L., Qu, S., Xue, M., Wang, K., Fu, F., Wang, L., Wang, Z., Feng, L., Xu, W., & Liu, P. (2020). The Coronavirus PEDV Evades Type III Interferon Response Through the miR-30c-5p/SOCS1 Axis. Frontiers in microbiology, 11, 1180. [CrossRef]
- Wang, C., Wang, T., Duan, L., Chen, H., Hu, R., Wang, X., Jia, Y., Chu, Z., Liu, H., Wang, X., Zhang, S., Xiao, S., Wang, J., Dang, R., & Yang, Z. (2022). Evasion of Host Antiviral Innate Immunity by Paramyxovirus Accessory Proteins. Frontiers in microbiology, 12, 790191. [CrossRef]
- Yao xue xue bao = Acta pharmaceutica Sinica, 49; //pubmed: (11), 1547–1553, available at: https.
- Yao xue xue bao = Acta pharmaceutica Sinica, 49; //pubmed: (11), 1547–1553, doi: https.
- Wang, M., Liu, Y., Qin, C., Lang, Y., Xu, A., Yu, C., Zhao, Z., Zhang, R., Yang, J., & Tang, J. (2022). Pseudorabies Virus EP0 Antagonizes the Type I Interferon Response via Inhibiting IRF9 Transcription. Journal of virology, 96(13), e0217121. [CrossRef]
- Wang, N., Zhan, Y., Zhu, L., Hou, Z., Liu, F., Song, P., Qiu, F., Wang, X., Zou, X., Wan, D., Qian, X., Wang, S., Guo, Y., Yu, H., Cui, M., Tong, G., Xu, Y., Zheng, Z., Lu, Y., & Hong, P. (2020). Retrospective Multicenter Cohort Study Shows Early Interferon Therapy Is Associated with Favorable Clinical Responses in COVID-19 Patients. Cell host & microbe, 28(3), 455–464.e2. [CrossRef]
- Wang, Y., Luo, J., Alu, A., Han, X., Wei, Y., & Wei, X. (2020). cGAS-STING pathway in cancer biotherapy. Molecular cancer, 19(1), 136. [CrossRef]
- Wang, Y., Qian, G., Zhu, L., Zhao, Z., Liu, Y., Han, W., Zhang, X., Zhang, Y., Xiong, T., Zeng, H., Yu, X., Yu, X., Zhang, X., Xu, J., Zou, Q., & Yan, D. (2022). HIV-1 Vif suppresses antiviral immunity by targeting STING. Cellular & molecular immunology, 19(1), 108–121. [CrossRef]
- Wang, Y., Sun, Y., Wu, A., Xu, S., Pan, R., Zeng, C., Jin, X., Ge, X., Shi, Z., Ahola, T., Chen, Y., & Guo, D. (2015). Coronavirus nsp10/nsp16 Methyltransferase Can Be Targeted by nsp10-Derived Peptide In Vitro and In Vivo To Reduce Replication and Pathogenesis. Journal of virology, 89(16), 8416–8427. [CrossRef]
- Warfield, K. L., Perkins, J. G., Swenson, D. L., Deal, E. M., Bosio, C. M., Aman, M. J., Yokoyama, W. M., Young, H. A., & Bavari, S. (2004). Role of natural killer cells in innate protection against lethal ebola virus infection. The Journal of experimental medicine, 200(2), 169–179. [CrossRef]
- Webb, L. G., & Fernandez-Sesma, A. (2022). RNA viruses and the cGAS-STING pathway: reframing our understanding of innate immune sensing. Current opinion in virology, 53, 101206. [CrossRef]
- Wei, X., Xie, F., Zhou, X., Wu, Y., Yan, H., Liu, T., Huang, J., Wang, F., Zhou, F., & Zhang, L. (2022). Role of pyroptosis in inflammation and cancer. Cellular & molecular immunology, 19(9), 971–992. [CrossRef]
- West, E. E., Kunz, N., & Kemper, C. (2020). Complement and human T cell metabolism: Location, location, location. Immunological reviews, 295(1), 68–81. [CrossRef]
- Woo, J., Shin, S., Cho, E., Ryu, D., Garandeau, D., Chajra, H., Fréchet, M., Park, D., & Jung, E. (2021). Senotherapeutic-like effect of Silybum marianum flower extract revealed on human skin cells. PloS one, 16(12), e0260545. [CrossRef]
- Woo, S. R., Corrales, L., & Gajewski, T. F. (2015). Innate immune recognition of cancer. Annual review of immunology, 33, 445–474. [CrossRef]
- Woolsey, C., Menicucci, A. R., Cross, R. W., Luthra, P., Agans, K. N., Borisevich, V., Geisbert, J. B., Mire, C. E., Fenton, K. A., Jankeel, A., Anand, S., Ebihara, H., Geisbert, T. W., Messaoudi, I., & Basler, C. F. (2019). A VP35 Mutant Ebola Virus Lacks Virulence but Can Elicit Protective Immunity to Wild-Type Virus Challenge. Cell reports, 28(12), 3032–3046.e6. [CrossRef]
- Xia X. (2020). Extreme Genomic CpG Deficiency in SARS-CoV-2 and Evasion of Host Antiviral Defense. Molecular biology and evolution, 37(9), 2699–2705. [CrossRef]
- Xia, H., & Shi, P. Y. (2020). Antagonism of Type I Interferon by Severe Acute Respiratory Syndrome Coronavirus 2. Journal of interferon & cytokine research : the official journal of the International Society for Interferon and Cytokine Research, 40(12), 543–548. [CrossRef]
- Xia, H., Cao, Z., Xie, X., Zhang, X., Chen, J. Y., Wang, H., Menachery, V. D., Rajsbaum, R., & Shi, P. Y. (2020). Evasion of Type I Interferon by SARS-CoV-2. Cell reports, 33(1), 108234. [CrossRef]
- Xia, P., Wang, S., Gao, P., Gao, G., & Fan, Z. (2016). DNA sensor cGAS-mediated immune recognition. Protein & cell, 7(11), 777–791. [CrossRef]
- Xiang, Y., Wang, M., Chen, H., & Chen, L. (2021). Potential therapeutic approaches for the early entry of SARS-CoV-2 by interrupting the interaction between the spike protein on SARS-CoV-2 and angiotensin-converting enzyme 2 (ACE2). Biochemical pharmacology, 192, 114724. [CrossRef]
- Xing, F., Matsumiya, T., Shiba, Y., Hayakari, R., Yoshida, H., & Imaizumi, T. (2016). Non-Canonical Role of IKKα in the Regulation of STAT1 Phosphorylation in Antiviral Signaling. PloS one, 11(12), e0168696. [CrossRef]
- Xu, J. , Mercado-López, X., Grier, J. T., Kim, W. K., Chun, L. F., Irvine, E. B., Del Toro Duany, Y., Kell, A., Hur, S., Gale, M., Jr, Raj, A., & López, C. B. (2015). Identification of a Natural Viral RNA Motif That Optimizes Sensing of Viral RNA by RIG-I. mBio, 6(5), e01265-15. [CrossRef]
- Xu, L. , Li, M., Yang, Y., Zhang, C., Xie, Z., Tang, J., Shi, Z., Chen, S., Li, G., Gu, Y., Wang, X., Zhang, F., Wang, Y., & Shen, X. (2022). Salmonella Induces the cGAS-STING-Dependent Type I Interferon Response in Murine Macrophages by Triggering mtDNA Release. mBio, 13(3), e0363221. [CrossRef]
- Xu, Z., Chen, Y., & Chen, Y. (2019). Spatiotemporal Regulation of Rho GTPases in Neuronal Migration. Cells, 8(6), 568. [CrossRef]
- Xu, Z., Choi, J. H., Dai, D. L., Luo, J., Ladak, R. J., Li, Q., Wang, Y., Zhang, C., Wiebe, S., Liu, A. C. H., Ran, X., Yang, J., Naeli, P., Garzia, A., Zhou, L., Mahmood, N., Deng, Q., Elaish, M., Lin, R., Mahal, L. K., … Sonenberg, N. (2022). SARS-CoV-2 impairs interferon production via NSP2-induced repression of mRNA translation. Proceedings of the National Academy of Sciences of the United States of America, 119(32), e2204539119. [CrossRef]
- Xue, B., Li, H., Liu, S., Feng, Q., Xu, Y., Deng, R., Chen, S., Wang, J., Li, X., Wan, M., Tang, S., & Zhu, H. (2022). The redox cycling of STAT2 maintains innate immune homeostasis. Cell Reports, 40(7), 111215. [CrossRef]
- Yamada, T. , Sato, S., Sotoyama, Y., Orba, Y., Sawa, H., Yamauchi, H., Sasaki, M., & Takaoka, A. (2021). RIG-I triggers a signaling-abortive anti-SARS-CoV-2 defense in human lung cells. Nature immunology, 22(7), 820–828. [CrossRef]
- Yang, Q. , Lin, F., Wang, Y., Zeng, M., & Luo, M. (2021). Long Noncoding RNAs as Emerging Regulators of COVID-19. Frontiers in immunology, 12, 700184. [CrossRef]
- Yang, Y., Jing, Y., Yang, J., & Yang, Q. (2018). Effects of intranasal administration with Bacillus subtilis on immune cells in the nasal mucosa and tonsils of piglets. Experimental and therapeutic medicine, 15(6), 5189–5198. [CrossRef]
- Yao, H. , Dittmann, M., Peisley, A., Hoffmann, H. H., Gilmore, R. H., Schmidt, T., Schmidt-Burgk, J., Hornung, V., Rice, C. M., & Hur, S. (2015). ATP-dependent effector-like functions of RIG-I-like receptors. Molecular cell, 58(3), 541–548. [CrossRef]
- Yazdani Brojeni, P., Matok, I., Garcia Bournissen, F., & Koren, G. (2012). A systematic review of the fetal safety of interferon alpha. Reproductive toxicology (Elmsford, N.Y.), 33(3), 265–268. [CrossRef]
- Yin, L. , Liu, X., Hu, D., Luo, Y., Zhang, G., & Liu, P. (2022). Swine Enteric Coronaviruses (PEDV, TGEV, and PDCoV) Induce Divergent Interferon-Stimulated Gene Responses and Antigen Presentation in Porcine Intestinal Enteroids. Frontiers in immunology, 12, 826882. [CrossRef]
- Yin, X. , Riva, L., Pu, Y., Martin-Sancho, L., Kanamune, J., Yamamoto, Y., Sakai, K., Gotoh, S., Miorin, L., De Jesus, P. D., Yang, C. C., Herbert, K. M., Yoh, S., Hultquist, J. F., García-Sastre, A., & Chanda, S. K. (2021). MDA5 Governs the Innate Immune Response to SARS-CoV-2 in Lung Epithelial Cells. Cell reports, 34(2), 108628. [CrossRef]
- Yin, Y., Romero, N., & Favoreel, H. W. (2021). Pseudorabies Virus Inhibits Type I and Type III Interferon-Induced Signaling via Proteasomal Degradation of Janus Kinases. Journal of virology, 95(20), e0079321. [CrossRef]
- Yockey, L. J., Jurado, K. A., Arora, N., Millet, A., Rakib, T., Milano, K. M., Hastings, A. K., Fikrig, E., Kong, Y., Horvath, T. L., Weatherbee, S., Kliman, H. J., Coyne, C. B., & Iwasaki, A. (2018). Type I interferons instigate fetal demise after Zika virus infection. Science immunology, 3(19), eaao1680. [CrossRef]
- Yoo, J. S., Sasaki, M., Cho, S. X., Kasuga, Y., Zhu, B., Ouda, R., Orba, Y., de Figueiredo, P., Sawa, H., & Kobayashi, K. S. (2021). SARS-CoV-2 inhibits induction of the MHC class I pathway by targeting the STAT1-IRF1-NLRC5 axis. Nature communications, 12(1), 6602. [CrossRef]
- Yoo, JS., Sasaki, M., Cho, S.X. et al. SARS-CoV-2 inhibits induction of the MHC class I pathway by targeting the STAT1-IRF1-NLRC5 axis. Nat Commun 12, 6602 (2021). [CrossRef]
- Yoshida, A. , Kawabata, R., Honda, T., Sakai, K., Ami, Y., Sakaguchi, T., & Irie, T. (2018). A Single Amino Acid Substitution within the Paramyxovirus Sendai Virus Nucleoprotein Is a Critical Determinant for Production of Interferon-Beta-Inducing Copyback-Type Defective Interfering Genomes. Journal of virology, 92(5), e02094-17. [CrossRef]
- Yoshida, Y., Katsurada, T., Nakabou, Y., & Kawabata, H. (2017). Efficacy of interferon-alpha in essential thrombocythemia during pregnancy. Annals of hematology, 96(5), 877–878. [CrossRef]
- Zhonghua shi yan he lin chuang bing du xue za zhi = Zhonghua shiyan he linchuang bingduxue zazhi = Chinese journal of experimental and clinical virology, 19; //pubmed: (3), 216–219, doi: https.
- Yu, Q., Carbone, C. J., Katlinskaya, Y. V., Zheng, H., Zheng, K., Luo, M., Wang, P. J., Greenberg, R. A., & Fuchs, S. Y. (2015). Type I interferon controls propagation of long interspersed element-1. The Journal of biological chemistry, 290(16), 10191–10199. [CrossRef]
- Yu, R. , Zhu, B., & Chen, D. (2022). Type I interferon-mediated tumor immunity and its role in immunotherapy. Cellular and molecular life sciences : CMLS, 79(3), 191. [CrossRef]
- Yu, S., Ge, H., Li, S., & Qiu, H. J. (2022). Modulation of Macrophage Polarization by Viruses: Turning Off/On Host Antiviral Responses. Frontiers in microbiology, 13, 839585. [CrossRef]
- Yu, W., Wu, X., Zhao, Y., Chen, C., Yang, Z., Zhang, X., Ren, J., Wang, Y., Wu, C., Li, C., Chen, R., Wang, X., Zheng, W., Liao, H., & Yuan, X. (2021). Computational Simulation of HIV Protease Inhibitors to the Main Protease (Mpro) of SARS-CoV-2: Implications for COVID-19 Drugs Design. Molecules (Basel, Switzerland), 26(23), 7385. [CrossRef]
- Yuanlin Ma, Xin-Zhuan Su and Fangli Lu (2020), The Roles of Type I Interferon in Co-infections With Parasites and Viruses, Bacteria, or Other Parasites, Front. Immunol., doi:. [CrossRef]
- Yuen, C. K., Lam, J. Y., Wong, W. M., Mak, L. F., Wang, X., Chu, H., Cai, J. P., Jin, D. Y., To, K. K., Chan, J. F., Yuen, K. Y., & Kok, K. H. (2020). SARS-CoV-2 nsp13, nsp14, nsp15 and orf6 function as potent interferon antagonists. Emerging microbes & infections, 9(1), 1418–1428. [CrossRef]
- Zanoni, I., Granucci, F., & Broggi, A. (2017). Interferon (IFN)-λ Takes the Helm: Immunomodulatory Roles of Type III IFNs. Frontiers in immunology, 8, 1661. [CrossRef]
- Zhang L, Richards A, Khalil A, Wogram E, Ma H, Young RA, Jaenisch R. SARS-CoV-2 RNA reverse-transcribed and integrated into the human genome. bioRxiv [Preprint]. 2020 Dec 13, 2020. [CrossRef]
- Zhang, C., Yang, Y., Zhou, X., Liu, X., Song, H., He, Y., & Huang, P. (2010). Highly pathogenic avian influenza A virus H5N1 NS1 protein induces caspase-dependent apoptosis in human alveolar basal epithelial cells. Virology journal, 7, 51. [CrossRef]
- Zhang, C., Yang, Y., Zhou, X., Yang, Z., Liu, X., Cao, Z., Song, H., He, Y., & Huang, P. (2011). The NS1 protein of influenza A virus interacts with heat shock protein Hsp90 in human alveolar basal epithelial cells: implication for virus-induced apoptosis. Virology journal, 8, 181. [CrossRef]
- Zhang, J. , Zhao, C., & Zhao, W. (2021). Virus Caused Imbalance of Type I IFN Responses and Inflammation in COVID-19. Frontiers in immunology, 12, 633769. [CrossRef]
- Zhang, J., Zhao, C., & Zhao, W. (2021). Virus Caused Imbalance of Type I IFN Responses and Inflammation in COVID-19. Frontiers in immunology, 12, 633769. [CrossRef]
- Zhang, M. , Zheng, S., & Liang, J. Q. (2022). Transcriptional and reverse transcriptional regulation of host genes by human endogenous retroviruses in cancers. Frontiers in microbiology, 13, 946296. [CrossRef]
- Zhang, Q. , & Yoo, D. (2016). Immune evasion of porcine enteric coronaviruses and viral modulation of antiviral innate signaling. Virus research, 226, 128–141. [CrossRef]
- Zhang, Q. , Ke, H., Blikslager, A., Fujita, T., & Yoo, D. (2018). Type III Interferon Restriction by Porcine Epidemic Diarrhea Virus and the Role of Viral Protein nsp1 in IRF1 Signaling. Journal of virology, 92(4), e01677-17. [CrossRef]
- Zhang, Q., Wang, C., Ma, F., Yao, L., Gao, H., Zhu, L. and Zheng, L., 2020. Development and biological activity of long-acting recombinant human interferon-α2b. BMC biotechnology, 20, pp.1-9. [CrossRef]
- Zhang, R., Xu, A., Qin, C., Zhang, Q., Chen, S., Lang, Y., Wang, M., Li, C., Feng, W., Zhang, R., Jiang, Z., & Tang, J. (2017). Pseudorabies Virus dUTPase UL50 Induces Lysosomal Degradation of Type I Interferon Receptor 1 and Antagonizes the Alpha Interferon Response. Journal of virology, 91(21), e01148-17. [CrossRef]
- Zhang, R., Xu, A., Qin, C., Zhang, Q., Chen, S., Lang, Y., Wang, M., Li, C., Feng, W., Zhang, R., Jiang, Z., & Tang, J. (2017). Pseudorabies Virus dUTPase UL50 Induces Lysosomal Degradation of Type I Interferon Receptor 1 and Antagonizes the Alpha Interferon Response. Journal of virology, 91(21), e01148-17. [CrossRef]
- Zhang, S. Y., Boisson-Dupuis, S., Chapgier, A., Yang, K., Bustamante, J., Puel, A., Picard, C., Abel, L., Jouanguy, E., & Casanova, J. L. (2008). Inborn errors of interferon (IFN)-mediated immunity in humans: insights into the respective roles of IFN-alpha/beta, IFN-gamma, and IFN-lambda in host defense. Immunological reviews, 226, 29–40. [CrossRef]
- Zhang, T., Liu, Y., Chen, Y., Wang, J., Feng, H., Wei, Q., Zhao, S., Yang, S., Ma, H., Liu, D., & Zhang, G. (2021). Antiviral activity of porcine interferon delta 8 against pseudorabies virus in vitro. International journal of biological macromolecules, 177, 10–18. [CrossRef]
- Zhang, X., Bai, X. C., & Chen, Z. J. (2020). Structures and Mechanisms in the cGAS-STING Innate Immunity Pathway. Immunity, 53(1), 43–53. [CrossRef]
- Zhang, X., Lu, S., Li, H., Wang, Y., Lu, Z., Liu, Z., Lai, Q., Ji, Y., Huang, X., Li, Y., Sun, J., Wu, Y., Xu, X., & Hou, J. (2020). Viral and Antibody Kinetics of COVID-19 Patients with Different Disease Severities in Acute and Convalescent Phases: A 6-Month Follow-Up Study. Virologica Sinica, 35(6), 820–829. [CrossRef]
- Zhang, Y. , Chen, S., Jin, Y., Ji, W., Zhang, W., & Duan, G. (2021). An Update on Innate Immune Responses during SARS-CoV-2 Infection. Viruses, 13(10), 2060. [CrossRef]
- Zhang, Y., Chen, S., Jin, Y., Ji, W., Zhang, W., & Duan, G. (2021). An Update on Innate Immune Responses during SARS-CoV-2 Infection. Viruses, 13(10), 2060. [CrossRef]
- Zhang, Y., Chen, Y., Li, Y., Huang, F., Luo, B., Yuan, Y., Xia, B., Ma, X., Yang, T., Yu, F., Liu, J., Liu, B., Song, Z., Chen, J., Yan, S., Wu, L., Pan, T., Zhang, X., Li, R., Huang, W., … Zhang, H. (2021). The ORF8 protein of SARS-CoV-2 mediates immune evasion through down-regulating MHC-Ι. Proceedings of the National Academy of Sciences of the United States of America, 118(23), e2024202118. [CrossRef]
- Zhang, Y., Gargan, S., Roche, F. M., Frieman, M., & Stevenson, N. J. (2022). Inhibition of the IFN-α JAK/STAT Pathway by MERS-CoV and SARS-CoV-1 Proteins in Human Epithelial Cells. Viruses, 14(4), 667. [CrossRef]
- Zhao, J. Zhao, J., Zhu, F. C., Shu, Y. L., Zhou, R., Liu, L. Q., Zhang, L. L., Shi, Z. Y., Tang, Z., Lin, L. Z., Yu, Z. A., Zhang, L. P., Zhang, B., & Hou, Y. D. (2005). Zhonghua shi yan he lin chuang bing du xue za zhi = Zhonghua shiyan he linchuang bingduxue zazhi = Chinese journal of experimental and clinical virology.
- Zhao, J. Zhao, J., Zhu, F. C., Shu, Y. L., Zhou, R., Liu, L. Q., Zhang, L. L., Shi, Z. Y., Tang, Z., Lin, L. Z., Yu, Z. A., Zhang, L. P., Zhang, B., & Hou, Y. D. (2005). Zhonghua shi yan he lin chuang bing du xue za zhi = Zhonghua shiyan he linchuang bingduxue zazhi = Chinese journal of experimental and clinical virology.
- Zhao, X., Zhao, Y., Du, J., Gao, P., & Zhao, K. (2021). The Interplay Among HIV, LINE-1, and the Interferon Signaling System. Frontiers in immunology, 12, 732775. [CrossRef]
- Zheng, J., Mo, J., Zhu, T., Zhuo, W., Yi, Y., Hu, S., Yin, J., Zhang, W., Zhou, H., & Liu, Z. (2020). Comprehensive elaboration of the cGAS-STING signaling axis in cancer development and immunotherapy. Molecular cancer, 19(1), 133. [CrossRef]
- Zheng, Y., An, H., Yao, M., Hou, J., Yu, Y., Feng, G., & Cao, X. (2010). Scaffolding adaptor protein Gab1 is required for TLR3/4- and RIG-I-mediated production of proinflammatory cytokines and type I IFN in macrophages. Journal of immunology (Baltimore, Md. : 1950), 184(11), 6447–6456. [CrossRef]
- Zheng, Y. , Deng, J., Han, L., Zhuang, M. W., Xu, Y., Zhang, J., Nan, M. L., Xiao, Y., Zhan, P., Liu, X., Gao, C., & Wang, P. H. (2022). SARS-CoV-2 NSP5 and N protein counteract the RIG-I signaling pathway by suppressing the formation of stress granules. Signal transduction and targeted therapy, 7(1), 22. [CrossRef]
- Zheng, Y. , Zhuang, M. W., Han, L., Zhang, J., Nan, M. L., Zhan, P., Kang, D., Liu, X., Gao, C., & Wang, P. H. (2020). Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) membrane (M) protein inhibits type I and III interferon production by targeting RIG-I/MDA-5 signaling. Signal transduction and targeted therapy, 5(1), 299. [CrossRef]
- Zhou, H., Zhu, J., Tu, J., Zou, W., Hu, Y., Yu, Z., Yin, W., Li, Y., Zhang, A., Wu, Y., Yu, Z., Chen, H., & Jin, M. (2010). Effect on virulence and pathogenicity of H5N1 influenza A virus through truncations of NS1 eIF4GI binding domain. The Journal of infectious diseases, 202(9), 1338–1346. [CrossRef]
- Zhou, Q., Chen, V., Shannon, C. P., Wei, X. S., Xiang, X., Wang, X., Wang, Z. H., Tebbutt, S. J., Kollmann, T. R., & Fish, E. N. (2020). Interferon-α2b Treatment for COVID-19. Frontiers in immunology, 11, 1061. [CrossRef]
- Zhou, Q., Chen, V., Shannon, C. P., Wei, X. S., Xiang, X., Wang, X., Wang, Z. H., Tebbutt, S. J., Kollmann, T. R., & Fish, E. N. (2020). Corrigendum: Interferon-α2b Treatment for COVID-19. Frontiers in immunology, 11, 615275. [CrossRef]
- Zhou, Q., MacArthur, M. R., He, X., Wei, X., Zarin, P., Hanna, B. S., Wang, Z. H., Xiang, X., & Fish, E. N. (2020). Interferon-α2b Treatment for COVID-19 Is Associated with Improvements in Lung Abnormalities. Viruses, 13(1), 44. [CrossRef]
- Zhou, R., Zheng, S. X., Tang, W., He, P. L., Li, X. Y., Yang, Y. F., Li, Y. C., Geng, J. G., & Zuo, J. P. (2006). Inhibition of inducible nitric-oxide synthase expression by (5R)-5-hydroxytriptolide in interferon-gamma- and bacterial lipopolysaccharide-stimulated macrophages. The Journal of pharmacology and experimental therapeutics, 316(1), 121–128. [CrossRef]
- Zhou, Y., Wang, M., Li, Y., Wang, P., Zhao, P., Yang, Z., Wang, S., Zhang, L., Li, Z., Jia, K., Zhong, C., Li, N., Yu, Y., & Hou, J. (2021). SARS-CoV-2 Spike protein enhances ACE2 expression via facilitating Interferon effects in bronchial epithelium. Immunology letters, 237, 33–41. [CrossRef]
- Zitvogel, L. , Galluzzi, L., Kepp, O., Smyth, M. J., & Kroemer, G. (2015). Type I interferons in anticancer immunity. Nature reviews. Immunology, 15(7), 405–414. [CrossRef]
- Znaidia, M., Demeret, C., van der Werf, S., & Komarova, A. V. (2022). Characterization of SARS-CoV-2 Evasion: Interferon Pathway and Therapeutic Options. Viruses, 14(6), 1247. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



