
Submitted:
22 February 2023
Posted:
23 February 2023
You are already at the latest version
Abstract
An iconic feature of insects is the apparent lack of legs on the abdomen, which is believed to be due to the repression of the leg-patterning gene Distalless (Dll) by abdominal Hox genes. However, in contrast to these molecular observations, it is not widely appreciated that the embryos of most insect groups do in fact form paired protrusions on most abdominal segments that appear to be homologous to the thoracic legs. However, these degenerate before hatching to form the abdominal body wall. To resolve this discordance between molecular and morphological observations, the expression patterns of pannier and araucan, genes known to distinguish proximal leg segments in all arthropods, are examined in embryos of the flour beetle Tribolium castaneum. In Tribolium embryos, all pregenital abdominal segments develop leg-like paired protrusions, and the stripes of pannier and araucan expression that delineate the proximal leg segments of the thorax are also expressed in the same configuration around these abdominal protrusions. This suggests that insect abdominal legs are homologous to only the proximal portion of the thoracic legs, which in insect adults forms the body wall (lateral tergum and pleura). These cryptic, truncated abdominal legs – likely inherited from their crustacean ancestors – appear to be an important wellspring for new functions in insects, such as caterpillar prolegs, gills, and structures for camouflage16 and aposematic warning.
Keywords:
arthropod
; leg
; evo-devo
; exite
1. Introduction
Insects are the most successful group of animals on the planet, due in part to the plethora of outgrowths that decorate their bodies with functions including flight, camouflage, and respiration. An iconic feature of the insect body plan is the presence of six walking legs, which gives the group its name, Hexapoda. It is commonly assumed in molecular and evo devo circles that insects lack legs on the pre-genital abdomen, except for the pleuropod on the first abdominal segment in embryos of certain insect groups18–22. In insect groups where larvae or adults have abdominal appendages, such as caterpillars or male sepsid flies, it has been proposed that these abdominal legs arose by re-evolution of the leg program19,21,23. By contrast, crustaceans (i.e. non-insect pancrustaceans), from which insects evolved24, generally have a pair of legs on all abdominal segments. The loss of these ancestral abdominal legs in the insect lineage is thought to have evolved when posterior Hox genes such as Ultrabithorax (Ubx) and abdominal-A (abdA) gained the ability to suppress the leg-patterning gene Distalless (Dll) in the insect abdomen18,25.
However, morphologists since 18441 have noted that, in the embryos of most insect groups, a pair of nubs forms on most abdominal segments which appear to be homologous to the thoracic legs1–9. These abdominal nubs flatten into the body wall to form the abdominal body wall (lateral tergum, pleura, and coxosternites) before the embryo hatches. But how could abdominal legs form in insects when Dll is suppressed by Hox genes?
By comparing a century of previous morphological work with the expression and function of several leg- and wing-patterning genes between insects, crustaceans, and arachnids – representing three of the four main groups of arthropods – Bruce and Patel 202010, 202111, and 202212 concluded that arthropods ancestrally have a total of 8 leg segments, but many arthropods have incorporated proximal leg segments into the body wall (Figure 1). Insects, for example, have incorporated proximal leg segments 7 and 8, which now form the body wall (pleura and lateral tergum, respectively)9, resulting in the familiar six (free) leg segments of insects: pretarsus/claw (1), tarsus (2), tibia (3), femur (4), trochanter (5), and coxa (6). In the embryos of all arthropods examined to date, representing three of the four major living arthropod groups — Drosophila melanogaster (fruit fly; insect)26–29, Tribolium castaneum (flour beetle; insect)10–12, Parhyale hawaiensis (beach scud; “crustacean”)10–12, Daphnia magna (water flea; “crustacean”)12, and Acanthoscurria geniculata (tarantula; chelicerate)11 — the Iroquois complex gene araucan (ara) is expressed in two stripes that bracket the incorporated 8th leg segment, and the GATA factor pannier (pnr) is expressed in the dorsal-most tissue and marks the bona fide body wall that is not leg-derived (Figure 1 and Figure 2). Thus, in contrast to other leg patterning genes30, the expression patterns of pnr and ara are highly conserved across arthropods. As such, they can be used to identify proximal leg segments even if the leg segments now function as body wall.
2. Results
In Tribolium embryos, we found that all pregenital abdominal segments develop leg-like paired protrusions. In situ Hybridization Chain Reaction (HCR)31,32 reveals that the stripes of pannier and araucan expression that delineate the proximal leg segments of the thorax are expressed in the same configuration around these abdominal protrusions (Figure 2 and Figure 3) as follows. In both the thorax and abdomen, pnr is expressed in the dorsal-most tissue, and this dorsal stripe of pnr is adjacent to two stripes of ara expression. The region bracketed by ara is highly similar between the thorax and abdomen: two armbands of ara surround one spiracle10,12,33 along with one eave-like protrusion (paranotal lobe), which is marked with vestigial and will later form a tergal plate or a wing. This configuration of gene expression and morphological structures is a hallmark of leg segment 8, which in adult insects forms the body wall (lateral tergum)10–12,33,34.
Previous experiments have shown that the insect abdomen does not express Dll25,35–39, which marks leg segments 1 – 5 (Figure 1)10–12,18,38, but does express buttonhead (btd) in leg-like, paired circular domains40, which in the thoracic legs marks segments 1 – 610–12,41,42. Based on this molecular deduction, the small abdominal protrusions ventral to leg segment 8 may represent leg segment 7 alone, or leg segments 6 and 7. Thus, rather than being completely limbless, the insect abdomen has a pair of legs on all pregenital abdominal body segments, but these abdominal legs are truncated, consisting of just the proximal two or three leg segments 6, 7, and 8 (coxa, pleura, and lateral tergum, respectively). Based on their similar positioning, embryonic development, and gene expression1–8, these abdominal leg nubs appear to be serially homologous with the proximal portions of thoracic legs.
3. Discussion
The results presented here answer the question of how legs can form on the insect abdomen despite the repression of Dll by posterior Hox genes in the insect lineage: only the distal leg, represented by leg segments 1 – 5 (claw to trochanter), is repressed by the Hox genes; the three proximal leg segments that do not depend on Dll function41,42, i.e., leg segments 6 – 8, are still generated. This is consistent with previous findings that a) loss of Dll does not delete the entire insect leg35–39; and more importantly, b) Dll is not sufficient to initiate leg development41,42,44. Together, these observations indicate that leg initiation must be achieved by other, more upstream genes. Candidates that could potentially initiate the entire arthropod leg (i.e., leg segments 1 – 8, which also includes the wing) are genes such as btd44 and Sp6-9 (Sp1 in Drosophila)41,42, and the juxtaposition of dorsal dpp with ventral wg22,45,46. Notably all of these genes have similar expression in the thorax and abdomen: btd is expressed in leg-like, paired, circular domains in both the thorax and the abdomen of insects40, and the intersecting stripes of dpp and wg that initiate leg development in the thorax are similarly expressed in the abdomen. This lends further support to the existence of cryptic insect abdominal legs.
Why truncate these ancestral abdominal legs instead of simply deleting the whole structure? One reason is that several essential structures develop from these proximal leg segments, such as the respiratory system (the spiracle and tracheae)2,47,48 as well as various exocrine glands like defensive scent glands49 and oenocytes2, which perform lipid processing, pheromone secretion, and developmental signaling50. In addition to these essential structures, many other useful structures are also carried on this leg-derived abdominal body wall, including tergal plates, gin traps51, knob-like pupal support structures52, dorsal “umbilical cord”-like structures in embryos of viviparous earwigs53, rod-like sensory organs in certain hemipterans54, and larval gills15,55 (Figure 4). Furthermore, in some insect lineages, the embryonic abdominal legs do not degenerate and instead form prolegs in caterpillars14,56, sawflies13, and Dipteran watersnipes19, as well as the adult sepsid fly male sternal brushes used in courtship23. Many of these insect abdominal structures have been called novel structures, which are commonly defined as structures that are not derived from, or homologous to, any structure in the ancestor nor any other structure in the individual57. However, rather than lacking homology, all of these structures likely derived from abdominal leg exites (leg lobes like gills and tergal plates) and legs inherited from their crustacean ancestors that have persisted in a cryptic state in insect embryos10,12,58. A similar molecular approach could be used to assay for cryptic abdominal legs in the paraphyletic “entomostracan” crustaceans which, like insects, also appear to lack abdominal legs59,60.
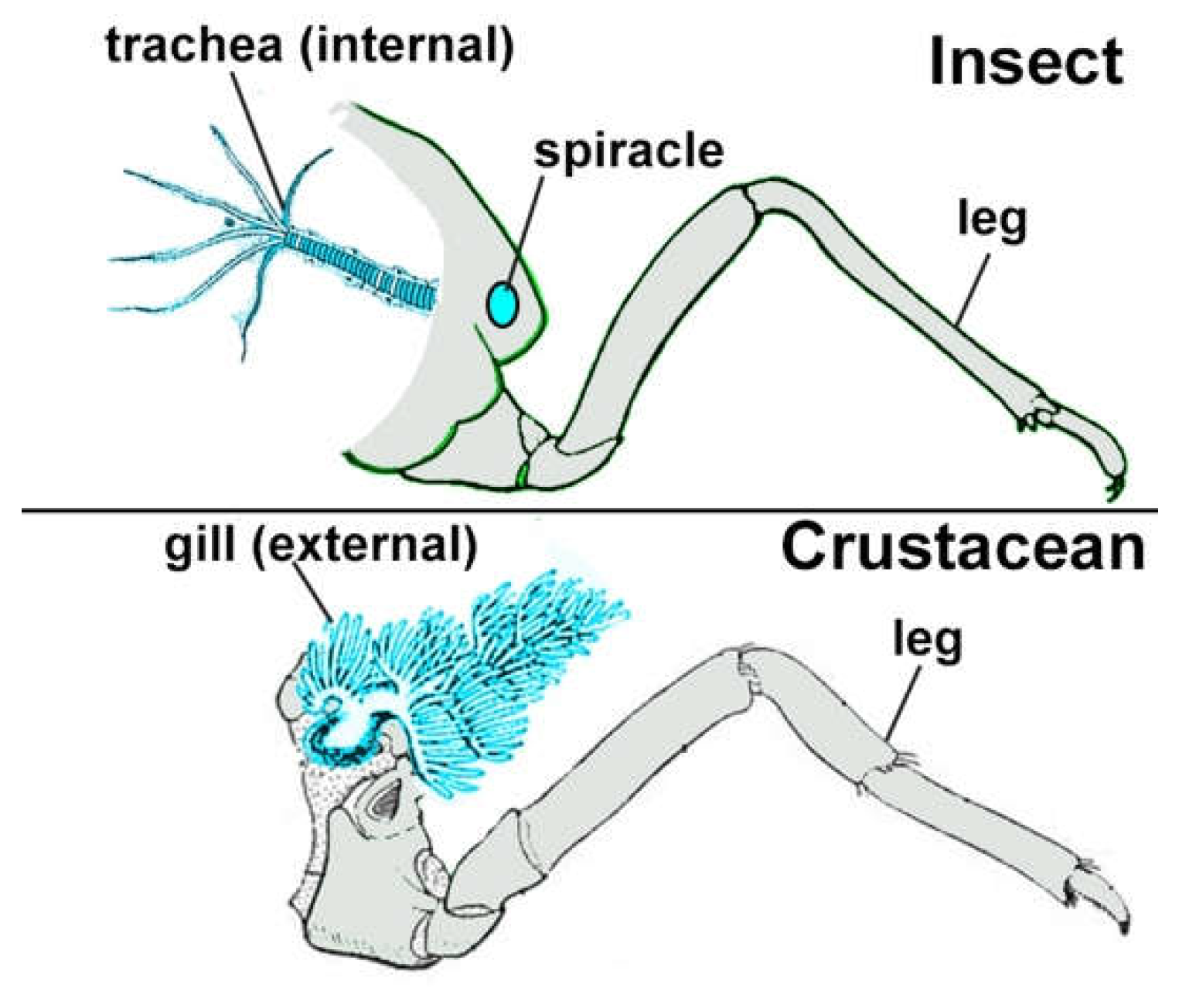
If insect abdominal legs were inherited from their crustacean ancestors, then the functional structures on these legs may also have been inherited from crustaceans10,61. Insect tracheae may be internalized crustacean gills (Figure 5)44,62; insect wings, tergal plates, helmets, horns, and other ectodermal outgrowths likely evolved from crustacean plate-type outgrowths10,12,63; and insect secretory glands (salivary, endocrine, exocrine, etc.) may have evolved from similar glands in crustaceans49,62,64. Surprisingly, respiratory organs and secretory glands can be homeotically transformed into each other49,62,64 and plate-type outgrowths arise from the same tissue as respiratory organs65, therefore all three types of structures may have arisen from a common embryonic exite-like structure on the lateral side of the proximal 8th leg segment10–12 that was inherited from the ancestor of all arthropods. Future studies may determine whether and how the different functional types of exites can be interconverted in nature.
Notably, multiple exites may emerge from one leg segment in crustaceans, like the anterior and posterior gills (arthrobranchs) of decapods34,66,67, and these multiple exites may even have different functions, such as the protective plate, respiratory gill, and brood-care lobe (oostegite) on leg segment 7 (coxa) of amphipod crustaceans like Parhyale34,65,67. Therefore, it is unsurprising if insects also have multiple exites with divergent functions emerging from the same leg segment, like the wing and spiracle that emerge from leg segment 8 that now forms body wall (lateral tergum)2,10,33. It will be interesting to determine whether each leg segment is limited to a set number of exites at restricted locations, or if any number of exites can arise in any location of the leg segment. In the latter case, it may be difficult to track the homology of individual exites within a leg segment over large phylogenetic distances.
This perspective of ancient homology plus divergence, rather than concepts like “partial homology”, explains why structures that have clearly different functions, such as wings and gills, often share some genes but not others: they are anciently homologous as exites, but not as wings, horns, tracheae, etc1,15,52,68–72. Similarly, it is likely that familiar genes such as vestigial, trachealess, ventral veins lacking, blistered, and apterous confer specific functions and shapes to exites rather than positional identity44,63,73–75. While useful for determining whether a structure is derived from an exite, these and other exite-specifying genes are probably less informative for determining positional homology between different arthropods44,63,73,74, in contrast to the well-conserved proximal-distal positional markers pnr and ara, along with joint markers like odd-skipped10–12.
The above perspective also provides an alternative interpretation of other insect abdominal structures, for example the posterior lobe on the genitalia of male Drosophila flies. The posterior lobe has been proposed as a novel structure that resulted when spiracle genes became co-opted into an unrelated structure, the genitalia76. However, given that genitals appear to be serially homologous to legs77–80, and respiratory structures like spiracles/tracheae are likely derived from the leg, then perhaps the genital “leg” program retains the ability to activate the spiracle/tracheae program. Given that respiratory structures need not be internal (crustacean gills are external lobes and the Drosophila larval posterior spiracle is external), it is plausible that the posterior lobe is an external spiracular structure. Rather than arising through the co-option of genes by an unrelated tissue, the posterior lobe may be the result of de-repression or re-activation of a serial homolog. This hypothesis would be supported if the posterior lobe emerges from the proximal-lateral side of the genital “leg” and if Iroquois genes like ara are expressed dorsal and ventral to the lobe.
In summary, the retention of proximal leg segments in the insect abdomen for essential functions like respiration and secretion appears to have allowed the non-essential plate-like outgrowths to become elaborated into new, useful structures like gin traps and camouflage. Thus, cryptic, truncated abdominal legs appear to serve as an important wellspring of new structures and functions in insects.
Author Contributions
HSB conceived, designed, and performed experiments and wrote manuscript. NHP edited manuscript.
Acknowledgements
HSB thanks Brendon E. Boudinot, Thierry Deuve, and Yukimasa Kobayashi for helpful discussion and comments.
Declaration of interests
The authors declare no competing interests.
References
- Matsuda, R. Morphology and evolution of the insect abdomen: with special reference to developmental patterns and their bearings upon systematics. (Pergamon Press, 1976).
- Kobayashi, Y.; Niikura, K.; Oosawa, Y.; Takami, Y. Embryonic development of Carabus insulicola (Insecta, Coleoptera, Carabidae) with special reference to external morphology and tangible evidence for the subcoxal theory. J. Morphol. 2013, 274, 1323–1352. [Google Scholar] [CrossRef] [PubMed]
- Brusca, R.C.; Brusca, G.J. Invertebrates. (Sinauer Associates Incorporated, 2003).
- Anderson, D.T. Embryology And Phylogeny In Annelids And Arthropods. (Pergamon Press Ltd., 1973).
- Roonwal, M.L. Studies on the embryology of the African migratory locust, Locusta migratoria migratorioides Reiche and Frm.(Orthoptera, Acrididae). II. Organogeny. Philos. Trans. R. Soc. London. Ser. B: Biol. Sci. 1937, 227. [Google Scholar]
- Ikeda, Y.; Machida, R. Embryogenesis of the dipluran Lepidocampa weberi Oudemans (Hexapoda, Diplura, Campodeidae): external morphology. J. Morphol. 1998, 237, 101–115. [Google Scholar] [CrossRef]
- Machida, R. External features of embryonic development of a jumping bristletail, Pedetontus unimaculatus Machida (Insecta, Thysanura, Machilidae). J. Morphol. 1981, 168, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Uchifune, T.; Machida, R. Embryonic development of Galloisiana yuasai Asahina, with special reference to external morphology (insecta: Grylloblattodea). J. Morphol. 2005, 266, 182–207. [Google Scholar] [CrossRef] [PubMed]
- Deuve, T. What is the epipleurite? A contribution to the subcoxal theory as applied to the insect abdomen. Ann. Soc. Entomol. Fr. 2018, 54, 1–26. [Google Scholar] [CrossRef]
- Bruce, H.S.; Patel, N.H. Knockout of crustacean leg patterning genes suggests that insect wings and body walls evolved from ancient leg segments. Nat. Ecol. Evol. 2020, 4, 1703–1712. [Google Scholar] [CrossRef]
- Bruce, H.S. How to align arthropod leg segments. bioRxiv. 2021. [CrossRef]
- Bruce, H.S.; Patel, N.H. The Daphnia carapace and other novel structures evolved via the cryptic persistence of serial homologs. Curr. Biol. 2022, 32. [Google Scholar] [CrossRef]
- Suzuki, Y.; Palopoli, M. Evolution of insect abdominal appendages: are prolegs homologous or convergent traits? Dev. Genes Evol. 2001, 211, 486–492. [Google Scholar] [CrossRef]
- Panganiban, G.; Nagy, L.; Carroll, S.B. The role of the Distal-less gene in the development and evolution of insect limbs. Curr. Biol. 1994, 4, 671–675. [Google Scholar] [CrossRef]
- Almudi, I.; et al. Genomic adaptations to aquatic and aerial life in mayflies and the origin of insect wings. Nat. Commun. 2020, 11, 2631. [Google Scholar] [CrossRef] [PubMed]
- Bank, S.; et al. A tree of leaves: Phylogeny and historical biogeography of the leaf insects (Phasmatodea: Phylliidae). Commun. Biol. 2021, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Motyka, M.; et al. Conspicuousness, phylogenetic structure, and origins of Müllerian mimicry in 4000 lycid beetles from all zoogeographic regions. Sci. Rep. 2021, 11, 5961. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.L.; DeCamillis, M.; Bennett, R.L. Distinct roles of the homeotic genes Ubx and abd-A in beetle embryonic abdominal appendage development. Proc. Natl. Acad. Sci. USA 2000, 97, 4504–4509. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.M.; Grbić; M. Cell Lineages in Larval Development and Evolution of Holometabolous Insects. in The Origin and Evolution of Larval Forms 275–300 (Elsevier, 1999). [CrossRef]
- Galant, R.; Carroll, S.B. Evolution of a transcriptional repression domain in an insect Hox protein. Nature 2002, 415, 910–913. [Google Scholar] [CrossRef]
- Jockusch, E.L.; Smith, F.W. Hexapoda: Comparative Aspects of Later Embryogenesis and Metamorphosis. in (null) 111–208 (Springer Vienna, 2015). [CrossRef]
- Estella, C. A dynamic network of morphogens and transcription factors patterns the fly leg. Curr. Top. Dev. Biol. 2012, 98, 173–198. [Google Scholar]
- Bowsher, J.H.; Nijhout, H.F. Partial co-option of the appendage patterning pathway in the development of abdominal appendages in the sepsid fly Themira biloba. Dev. Genes Evol. 2010, 219, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Fernandez, J.; et al. Pancrustacean Evolution Illuminated by Taxon-Rich Genomic-Scale Data Sets with an Expanded Remipede Sampling. Genome Biol. Evol. 2019, 11, 2055–2070. [Google Scholar] [CrossRef] [PubMed]
- Ronshaugen, M.; McGinnis, N.; McGinnis, W. Hox protein mutation and macroevolution of the insect body plan. Nature 2002, 415, 914–917. [Google Scholar] [CrossRef]
- Ikmi, A.; Netter, S.; Coen, D. Prepatterning the Drosophila notum: the three genes of the iroquois complex play intrinsically distinct roles. Dev. Biol. 2008, 317, 634–648. [Google Scholar] [CrossRef]
- Calleja, M.; et al. Generation of medial and lateral dorsal body domains by the pannier gene of Drosophila. Development 2000, 127, 3971–3980. [Google Scholar] [CrossRef] [PubMed]
- Diez del Corral, R.; Aroca, P.; Gomez-Skarmeta, J.L.; Cavodeassi, F.; Modolell, J. The Iroquois homeodomain proteins are required to specify body wall identity in Drosophila. Genes Dev. 1999, 13, 1754–1761. [Google Scholar] [CrossRef] [PubMed]
- Cavodeassi, F.; Diez del Corral, R.; Campuzano, S.; Domínguez, M. Compartments and organising boundaries in the Drosophila eye: the role of the homeodomain Iroquois proteins. Development 1999, 126, 4933–4942. [Google Scholar] [CrossRef] [PubMed]
- Abzhanov, A.; Kaufman, T.C. Homologs of Drosophila appendage genes in the patterning of arthropod limbs. Dev. Biol. 2000, 227, 673–689. [Google Scholar] [CrossRef]
- Choi, H.M.T.; et al. Third-generation in situ hybridization chain reaction: multiplexed, quantitative, sensitive, versatile, robust. Development 2018, 145, dev165753–122. [Google Scholar] [CrossRef] [PubMed]
- Bruce, H.S. et al. Hybridization Chain Reaction (HCR) In Situ Protocol v1. protocols.io Hybridization Chain Reaction (HCR) In Situ Protocol v1. 2021. [CrossRef]
- Kobayashi, Y. et al. Paranotal Lobes are Appendicular in Origin: Elucidation by Micro-CT Analysis of the Thoracic Muscular System in the Larvae of Carabus insulicola (Insecta, Coleoptera). Proceedings of the Arthropodan Embryological Society of Japan (2022).
- Boxshall, G.A. The evolution of arthropod limbs. Biol. Rev. 2004, 79, 253–300. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Jürgens, G. Proximal-distal pattern formation in Drosophila: cell autonomous requirement for Distal-less gene activity in limb development. EMBO J. 1989, 8, 2045–2055. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Simcox, A.A.; Cohen, S.M. Allocation of the thoracic imaginal primordia in the Drosophila embryo. Development 1993, 117, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.; Tomlinson, A. The roles of the homeobox genes aristaless and Distal-less in patterning the legs and wings of Drosophila. Development 1998, 125, 4483–4493. [Google Scholar] [CrossRef]
- Beermann, A.; et al. The Short antennae gene of Tribolium is required for limb development and encodes the orthologue of the Drosophila Distal-less protein. Development 2001, 128, 287–297. [Google Scholar] [CrossRef]
- Angelini, D.R.; Kaufman, T.C. Functional analyses in the hemipteran Oncopeltus fasciatus reveal conserved and derived aspects of appendage patterning in insects. Dev. Biol. 2004, 271, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Schaeper, N.D.; Prpic, N.-M.; Wimmer, E.A. A clustered set of three Sp-family genes is ancestral in the Metazoa: evidence from sequence analysis, protein domain structure, developmental expression patterns and chromosomal location. BMC Evol. Biol. 2010, 10, 88. [Google Scholar] [CrossRef] [PubMed]
- Estella, C.; Rieckhof, G.; Calleja, M.; Morata, G. The role of buttonhead and Sp1 in the development of the ventral imaginal discs of Drosophila. Development 2003, 130, 5929–5941. [Google Scholar] [CrossRef] [PubMed]
- Estella, C.; Mann, R.S. Non-Redundant Selector and Growth-Promoting Functions of Two Sister Genes, buttonhead and Sp1, in Drosophila Leg Development. PLoS Genet. 2010, 6, e1001001. [Google Scholar] [CrossRef] [PubMed]
- Klann, M.; Schacht, M.I.; Benton, M.A.; Stollewerk, A. Functional analysis of sense organ specification in the Tribolium castaneum larva reveals divergent mechanisms in insects. BMC Biol. 2021, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Franch-Marro, X.; Martín, N.; Averof, M.; Casanova, J. Association of tracheal placodes with leg primordia in Drosophila and implications for the origin of insect tracheal systems. Development 2006, 133, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.; Tomlinson, A. Initiation of the proximodistal axis in insect legs. Development 1995. [Google Scholar] [CrossRef] [PubMed]
- Letizia, A.; Barrio, R.; Campuzano, S. Antagonistic and cooperative actions of the EGFR and Dpp pathways on the iroquois genes regulate Drosophila mesothorax specification and patterning. Development 2007, 134, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Chavez, C.; Andrew, D.J. Trachealess (Trh) regulates all tracheal genes during Drosophila embryogenesis. Dev. Biol. 2011, 360, 160–172. [Google Scholar] [CrossRef]
- Snodgrass, R.E. Morphology and mechanism of the insect thorax. vol. 80 (City of Washington, Smithsonian Institution, 1927).
- Hanna, L.; Popadić, A. A hemipteran insect reveals new genetic mechanisms and evolutionary insights into tracheal system development. PNAS 2020, 121, 201908975. [Google Scholar] [CrossRef]
- Burns, K.A.; Gutzwiller, L.M.; Tomoyasu, Y.; Gebelein, B. Oenocyte development in the red flour beetle Tribolium castaneum. Dev. Genes Evol. 2012, 222, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Linz, D.M.; Tomoyasu, Y. Dual evolutionary origin of insect wings supported by an investigation of the abdominal wing serial homologs in Tribolium. 2018; 115, E658–E667. [Google Scholar]
- Hu, Y.; Moczek, A.P. Wing serial homologues and the diversification of insect outgrowths: insights from the pupae of scarab beetles. Proc. R. Soc. B. 2021, 288, 20202828. [Google Scholar] [CrossRef] [PubMed]
- Tworzydlo, W.; Jaglarz, M.K.; Pardyak, L.; Bilinska, B.; Bilinski, S.M. Evolutionary origin and functioning of pregenital abdominal outgrowths in a viviparous insect, Arixenia esau. Sci. Rep. 2019, 9, 16090. [Google Scholar] [CrossRef] [PubMed]
- Hoch, H. et al. Non-sexual abdominal appendages in adult insects challenge a 300 million year old bauplan. Curr. Biol. 24, R16-7 (2014).
- Komatsu, S.; Kobayashi, Y. Embryonic development of a whirligig beetle, Dineutus mellyi, with special reference to external morphology (insecta: Coleoptera, Gyrinidae). J. Morphol. 2012, 273, 541–560. [Google Scholar] [CrossRef]
- Warren, R.W.; Nagy, L.; Selegue, J.; Gates, J.; Carroll, S. Evolution of homeotic gene regulation and function in flies and butterflies. Nature 1994, 372, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.B.; Wagner, G.P. Novelty in evolution: restructuring the concept. Annu. Rev. Ecol. Syst. 1991, 22, 229–56. [Google Scholar] [CrossRef]
- Sadier, A.; Sears, K.E.; Womack, M. Unraveling the heritage of lost traits. J Exp Zool (Mol Dev Evol) jez.b.23030 (2021). [CrossRef]
- Fritsch, M.; Richter, S. How body patterning might have worked in the evolution of arthropods—A case study of the mystacocarid Derocheilocaris remanei (Crustacea, Oligostraca). J. Exp. Zool. Pt. B 2022, 338, 342–359. [Google Scholar] [CrossRef] [PubMed]
- McCarthy-Taylor, J.B.; et al. Expression of Abdominal-B in the brine shrimp, Artemia franciscana, expands our evolutionary understanding of the crustacean abdomen. Dev. Biol. 2022, 489, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Tomoyasu, Y. What crustaceans can tell us about the evolution of insect wings and other morphologically novel structures. Curr. Opin. Genet. Dev. 2021, 69, 48–55. [Google Scholar] [CrossRef]
- Grillo, M.; Casanova, J.; Averof, M. Development: A Deep Breath for Endocrine Organ Evolution. Curr. Biol. 2014, 24, R38–R40. [Google Scholar] [CrossRef]
- Fisher, C.R.; Kratovil, J.D.; Angelini, D.R.; Jockusch, E.L. Out from under the wing: reconceptualizing the insect wing gene regulatory network as a versatile, general module for body-wall lobes in arthropods. Proc. R. Soc. B 2021, 288, 20211808. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Higueras, C.; Sotillos, S.; Castelli-Gair Hombría, J. Common origin of insect trachea and endocrine organs from a segmentally repeated precursor. Curr. Biol. 2014, 24, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Ungerer, P.; Wolff, C. External morphology of limb development in the amphipod Orchestia cavimana (Crustacea, Malacostraca, Peracarida). Zoomorphology 2005, 124, 89–99. [Google Scholar] [CrossRef]
- Hong, S.Y. Development of epipods and gills in some pagurids and brachyurans. J. Nat. Hist. 1988, 22, 1005–1040. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Jaume, D. Exopodites, epipodites and gills in Crustaceans. Arthropod Syst. Phylogeny 2009, 67, 229–254. [Google Scholar]
- Hu, Y.; Linz, D.M.; Moczek, A.P. Beetle horns evolved from wing serial homologs. Science 2019, 366, 1004–1007. [Google Scholar] [CrossRef] [PubMed]
- Tomoyasu, Y.; Ohde, T.; Clark-Hachtel, C. What serial homologs can tell us about the origin of insect wings. F1000Res 2017, 6, 268–11. [Google Scholar] [CrossRef] [PubMed]
- Clark-Hachtel, C.M.; Tomoyasu, Y. Exploring the origin of insect wings from an evo-devo perspective. Curr. Opin. Insect. Sci. 2016, 13, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Clark-Hachtel, C.M.; Linz, D.M.; Tomoyasu, Y. Insights into insect wing origin provided by functional analysis of vestigial in the red flour beetle, Tribolium castaneum. 2013; 110, 16951–16956. [Google Scholar]
- DiFrisco, J.; Wagner, G.P.; Love, A.C. Reframing research on evolutionary novelty and co-option: Character identity mechanisms versus deep homology. Seminars in Cell & Developmental Biology S1084952122001033 (2022). [CrossRef]
- Averof, M.; Cohen, S.M. Evolutionary origin of insect wings from ancestral gills. Nature 1997, 385, 627–630. [Google Scholar] [CrossRef]
- Damen WG, M.; Saridaki, T.; Averof, M. Diverse adaptations of an ancestral gill: a common evolutionary origin for wings, breathing organs, and spinnerets. Curr. Biol. 2002, 12, 1711–1716. [Google Scholar] [CrossRef]
- Fisher, C.R.; Wegrzyn, J.L.; Jockusch, E.L. Co-option of wing-patterning genes underlies the evolution of the treehopper helmet. Nat. Ecol. Evol. 2020, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Glassford, W.J.; et al. Co-option of an Ancestral Hox-Regulated Network Underlies a Recently Evolved Morphological Novelty. Dev. Cell 2015, 34, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Estrada, B.; Sánchez-Herrero, E. Abdominal-B antagonizes leg development. Development 2001, 9. [Google Scholar]
- Gorfinkiel, N.; Sánchez, L.; Guerrero, I. Drosophila terminalia as an appendage-like structure. Mech. Dev. 1999, 86, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Casares, F.; Sánchez, L.; Guerrero, I.; Sánchez-Herrero, E. The genital disc of Drosophila melanogaster. Dev. Genes Evol. 1997, 207, 216–228. [Google Scholar] [CrossRef]
- Boudinot, B.E. A general theory of genital homologies for the Hexapoda (Pancrustacea) derived from skeletomuscular correspondences, with emphasis on the Endopterygota. Arthropod Struct. Dev. 2018, 47, 563–613. [Google Scholar] [CrossRef]
Figure 1.
Leg segment alignment of arthropod legs based on expression and function of leg genes. From Bruce 2020 and 2021.
Figure 1.
Leg segment alignment of arthropod legs based on expression and function of leg genes. From Bruce 2020 and 2021.
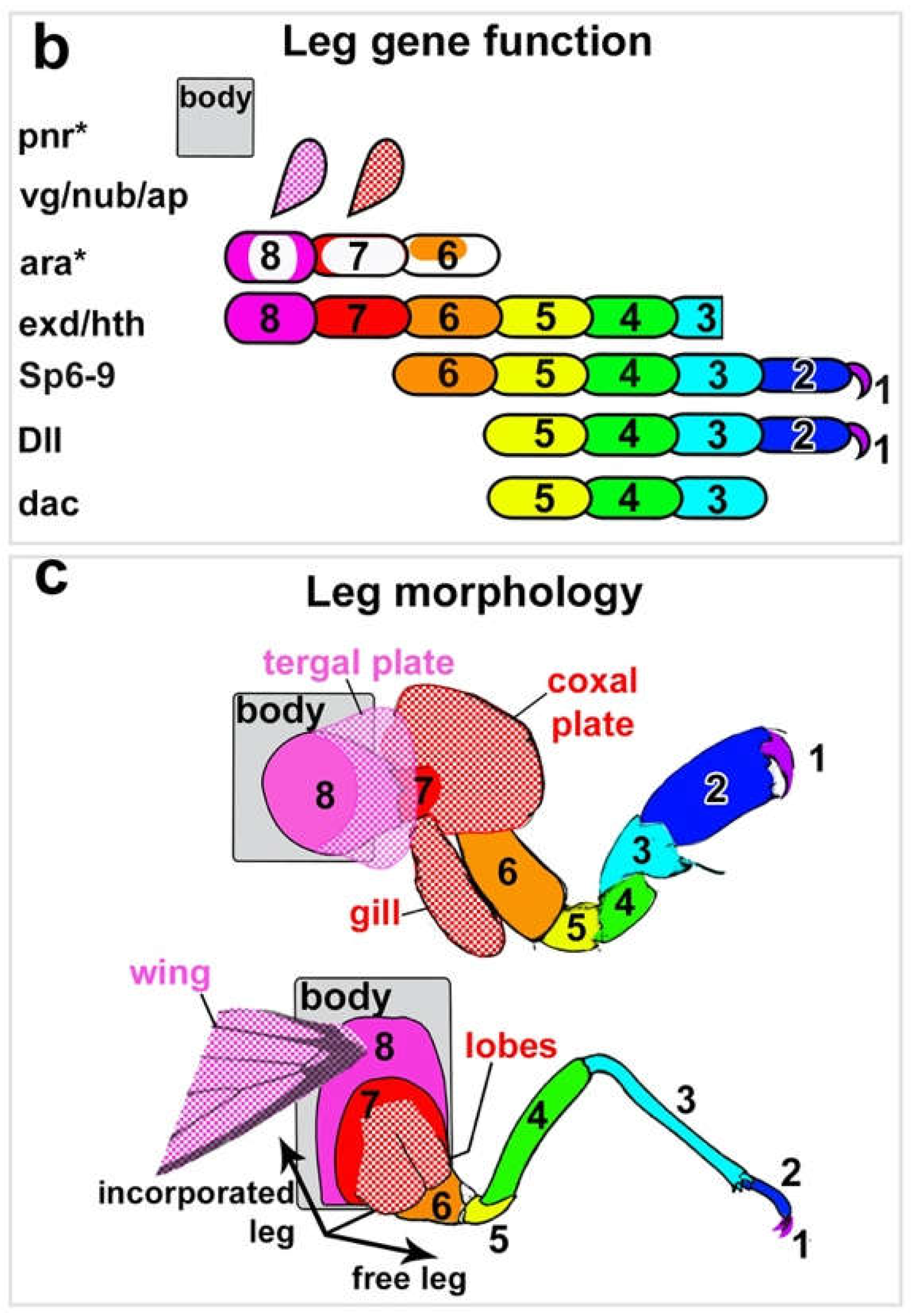
Figure 2.
In all arthropods examined, araucan (ara) brackets the incorporated 8th leg segment, and pannier (pnr) is expressed in the dorsal-most tissue and marks the true body wall. A, B. Parhyale, crustacean. C, D. Tribolium, insect. E, F. Acanthoscurria, chelicerate. From Bruce 2021.
Figure 2.
In all arthropods examined, araucan (ara) brackets the incorporated 8th leg segment, and pannier (pnr) is expressed in the dorsal-most tissue and marks the true body wall. A, B. Parhyale, crustacean. C, D. Tribolium, insect. E, F. Acanthoscurria, chelicerate. From Bruce 2021.
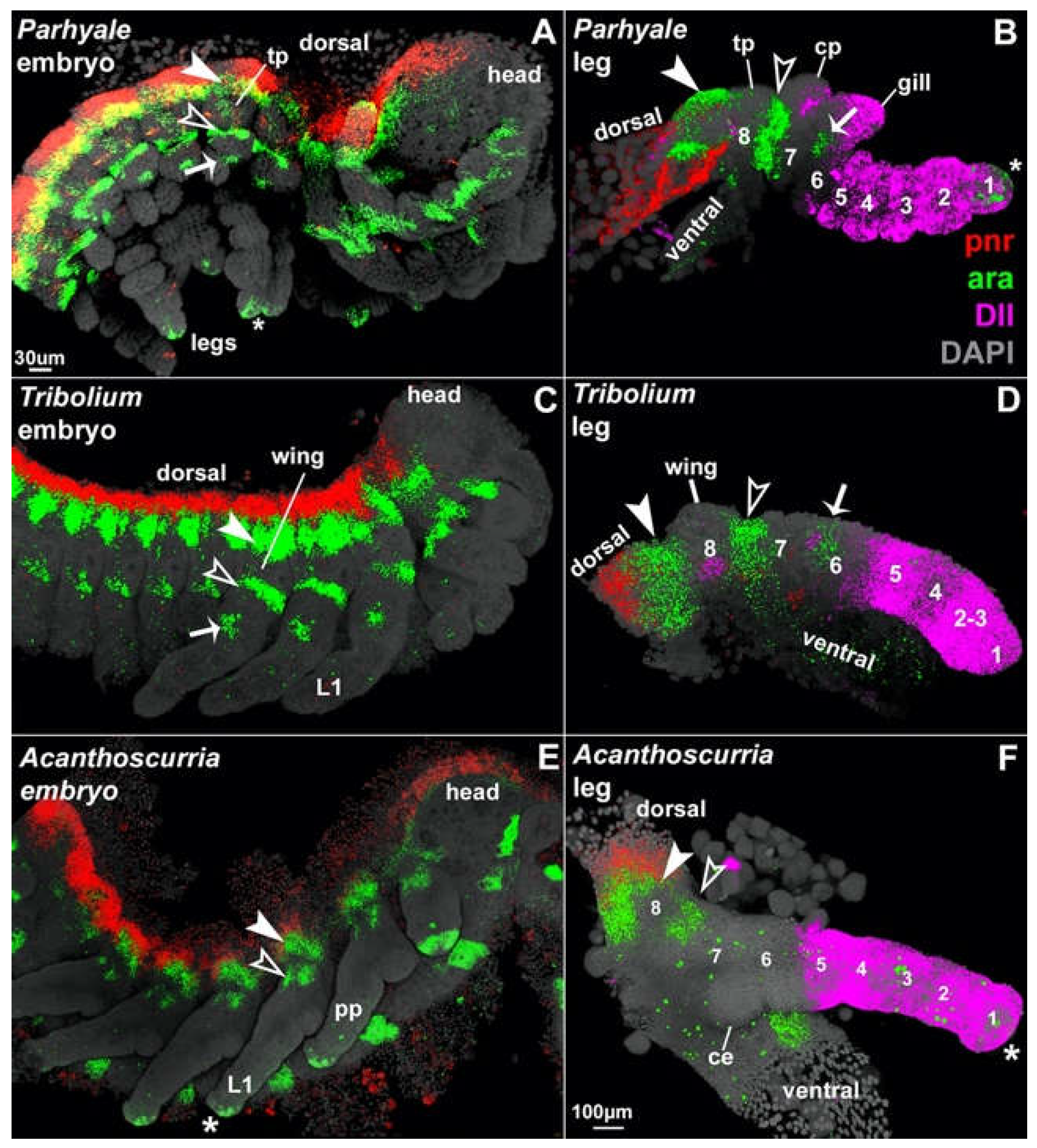
Figure 3.
Segment identity of abdominal leg nubs in Tribolium embryos. Top: approximately stage NS14.1 (Klann 2021)43. Bottom: Stage NS15.4 (Klann 2021). Arrow points to 4th abdominal leg nub that later degenerates into the body wall. araucan (ara, green) is expressed in two stripes down the length of the embryo, one dorsal stripe and one lateral stripe, as well as a circular patch on leg segment 6 (coxa) of each thoracic leg. The two stripes bracket the proximal-most 8th leg segment that carries both the wing and the spiracle. vestigial (vg, pink) marks the future wing serial homologs: the wing, elytra, and tergal plates, as well as certain cells in the ventral nerve cord. Note that, in addition to differences in their shape and axial position, cells in the ventral nerve cord are larger and less compact than cells of the leg nub, thus the two are readily distinguished. Gray, DAPI, marks all cell nuclei.
Figure 3.
Segment identity of abdominal leg nubs in Tribolium embryos. Top: approximately stage NS14.1 (Klann 2021)43. Bottom: Stage NS15.4 (Klann 2021). Arrow points to 4th abdominal leg nub that later degenerates into the body wall. araucan (ara, green) is expressed in two stripes down the length of the embryo, one dorsal stripe and one lateral stripe, as well as a circular patch on leg segment 6 (coxa) of each thoracic leg. The two stripes bracket the proximal-most 8th leg segment that carries both the wing and the spiracle. vestigial (vg, pink) marks the future wing serial homologs: the wing, elytra, and tergal plates, as well as certain cells in the ventral nerve cord. Note that, in addition to differences in their shape and axial position, cells in the ventral nerve cord are larger and less compact than cells of the leg nub, thus the two are readily distinguished. Gray, DAPI, marks all cell nuclei.
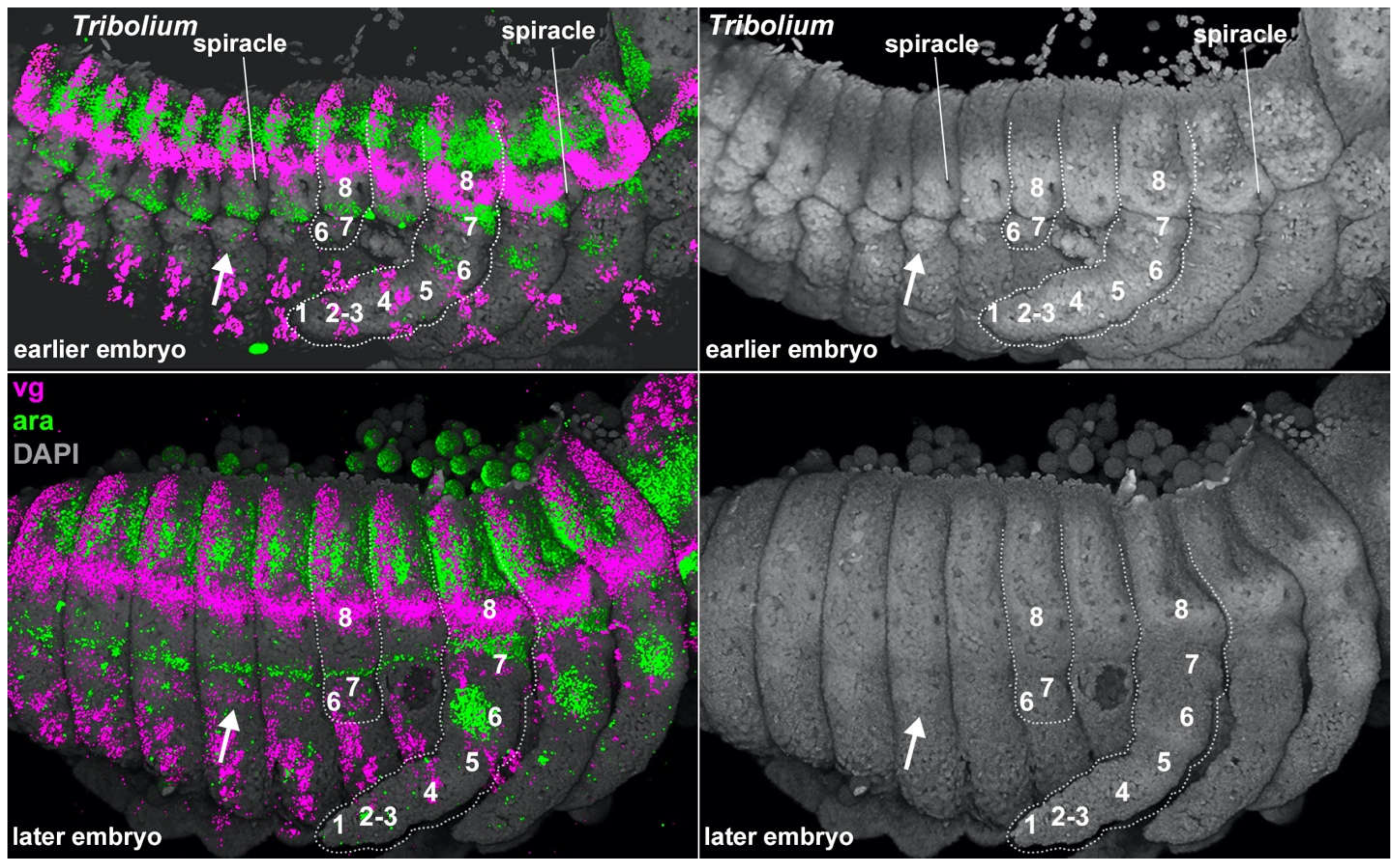
Figure 4.
Model of potential exite locations in arthropods. Top: Generalized arthropod ancestor indicating leg segments (colored rectangles) and regions where leg exites may potentially form (colored circles). Bottom: Generalized insect indicating leg segments and regions where leg exites may potentially form as well as examples of specific structures that are here proposed to be derived from cryptic, truncated abdominal legs. In insect: 1=claw, 2=tarsus, 3=tibia, 4=femur, 5=trochanter, 6=coxa, 7=subcoxa, 8=precoxa. Note that 7 and 8 now form lateral body wall in insects. “Proleg” here refers to Lepidopteran prolegs.
Figure 4.
Model of potential exite locations in arthropods. Top: Generalized arthropod ancestor indicating leg segments (colored rectangles) and regions where leg exites may potentially form (colored circles). Bottom: Generalized insect indicating leg segments and regions where leg exites may potentially form as well as examples of specific structures that are here proposed to be derived from cryptic, truncated abdominal legs. In insect: 1=claw, 2=tarsus, 3=tibia, 4=femur, 5=trochanter, 6=coxa, 7=subcoxa, 8=precoxa. Note that 7 and 8 now form lateral body wall in insects. “Proleg” here refers to Lepidopteran prolegs.
Figure 5.
Similarity of internal insect tracheae and external crustacean gills. Modified from Snodgrass 1935 and Boxshall 2009.
Figure 5.
Similarity of internal insect tracheae and external crustacean gills. Modified from Snodgrass 1935 and Boxshall 2009.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



