
Submitted:
25 January 2023
Posted:
26 January 2023
You are already at the latest version
Abstract
The World Health Organization has estimated an annual occurrence of approximately 392 million Dengue virus (DENV) infections in more than 100 countries where the virus is endemic, and this represents a serious threat to humanity. DENV is a serologic group with four distinct serotypes (DENV-1, DENV-2, DENV-3, and DENV-4) belonging to the genus Flavivirus, in the family Flaviviridae. Dengue is the most widespread mosquito-borne disease in the world. The ~10.7 kb DENV genome encodes three structural proteins (capsid [C], pre-membrane [prM], and envelope [E]) and seven non-structural (NS) proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5). The NS1 protein is a membrane-associated dimer and a secreted, lipid-associated hexamer. Dimeric NS1 is found on membranes both in cellular compartments and cell surfaces. Secreted NS1 (sNS1) is often present in patient serum at very high levels, which correlates with severe dengue symptoms. This study was conducted to discover how NS1 protein, microRNAs-15/16 (miRNAs-15/16), and apoptosis are related during DENV-4 infection in human liver cell lines. Huh 7.5 and HepG2 cells were infected with DENV-4, and miRNAs-15/16, viral load, NS1 protein, and caspases-3/7 were quantified after different times of infection. This study demonstrated that miRNAs-15/16 are overexpressed during infection of HepG2 and Huh 7.5 cells by DENV-4 and have a relationship with NS1 protein expression, viral load, and activity of caspases-3/7, thus making these miRNAs potential injury markers during DENV infection in human hepatocytes.
Keywords:
Apoptosis
; Dengue Virus
; microRNAs
; Viral Nonstructural Proteins
1. Introduction
The World Health Organization (WHO) states that about 392 million dengue virus (DENV) infections happen every year in more than 100 countries where the virus is common, being a severe threat to public health. DENV causes the most common mosquito-borne disease in the world. It consists of a group of four different serotypes (DENV-1, DENV-2, DENV-3, and DENV-4) that belong to the genus Flavivirus, in the family Flaviviridae. Dengue fever (DF) and severe dengue (SD) (formerly called dengue hemorrhagic fever/dengue shock syndrome [DHF/DSS]) are major causes of illness in tropical and subtropical areas [1,2,3].
DENV is spread by the Aedes aegypti mosquito, which must feed on a person with the virus during the first 5 days of high viremia when the person is just starting to feel sick. Asymptomatic individuals can still infect mosquitoes, and infected mosquitoes incubate the virus. The virus remains in the mosquitoes for 8–12 extra days (extrinsic incubation period) before it can be passed on to a susceptible human. The mosquito is infected for its whole life and, more importantly, can spread DENV to humans more than once. There is a lack of evidence that the virus can be passed on in unusual ways, like through organ transplants, blood transfusions, or from an infected pregnant woman to her unborn child [4].
DENV has a genome of about ~10.7 kb that encodes a single polyprotein which is post-translationally cleaved into three structure proteins (capsid [C], pre-membrane [prM], and envelope [E]) and seven non-structural (NS) proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5). They are 50-nm, icosahedral, lipid-wrapped particles with the same structure and pathogenic properties but different genetic and serological properties [5].
The NS1 protein is a membrane-associated dimer and a secreted, lipid-associated hexamer (sNS1). Dimeric NS1 is found on membranes in cellular compartments and on the cell surfaces of virus-infected cells. It has been shown that serum from patients contains sNS1, and the levels of this protein are often very high, which is linked to the start of SD. NS1 also affects the innate immune response and has been seen in multiple interactions with the complement system through factor H [6,7,8,9].
Identifying regulatory ~22 nucleotides (nt) non-coding RNAs has been one of the significant molecular biology tasks. MicroRNAs (miRNAs) are produced from transcripts that form a fold-back hairpin structure, which is cropped by the RNase III enzyme Drosha to a ~70 nt long pre-miRNA [7,10,11].
This structure is exported to the cytoplasm and further processed by a second RNase III enzyme, Dicer, into the mature ~22 nt miRNA and its complementary star sequence. Mature miRNAs control gene expression by directing effector complexes of Argonauts (Ago) proteins to their cognate targets, which stops translation or speeds up mRNA degradation [12]. Regulation depends on base pairing between the miRNA and target sites on the mRNA. The seed sequence, nucleotides 2–7 at the 5' end of the miRNA, is significant for this interaction [10,13].
Here, we show a correlation between the expression profile of the NS1 protein and the expression of miRNAs-15/16 during infection by DENV-4 in the human liver cell lines HepG2 and Huh 7.5.
2. Materials and Methods
2.1. Cell Culture
HepG2 and Huh 7.5 hepatic human cell lines were cultured at 37 °C in a nutrient Dulbecco's modified Eagle's medium (DMEM) (Sigma-Aldrich) containing L-glutamine, HEPES buffer and 10% fetal bovine serum (FBS, Gibco).
2.2. Viral Samples
At first, the DENV-4 (BeH778494) isolate was grown in C6/36 cells with 5% FBS in Leibovitz 15 (L-15) medium at 28 °C. On the seventh day after infection, the culture supernatant was harvested and stored at -80 °C before infections of Huh 7.5 and HepG2 cell types.
2.3. Infection of Hepatic Human Cells
The HepG2 and Huh 7.5 cells were infected with DENV-4 using the adsorption method for one hour at 37 °C (MOI=1). Subsequently, the cell cultures were washed with phosphate-buffered saline (PBS), put in a new medium, and grown again for the experiment. The extracted RNAs were kept at -80 °C until needed. The supernatants of the virus-infected cells were collected daily up to 120 hours post-infection (hpi).
2.4. RNA Extraction
The cell supernatant was subjected to viral RNA extraction for viral load quantification using the commercial Maxwell 16 Viral Total Nucleic Acid Purification kit (Promega, Madison, WI, USA). The samples were then quantified using the commercial Qubit RNA High Sensitivity (HS) kit (Thermofisher, USA) on the Qubit 3.0 platform (Thermofisher, USA). The extracted RNAs were kept at -80 °C until needed.
2.5. Quantification of Viral Load by RT-qPCR
The cell supernatant was used in each of the analyzed periods. Detection of DENV genome was performed by RT-qPCR using the method described by Johnson et al. [14] with the commercial kit GoTaq Probe 1-Step RT-qPCR System (Promega, USA) on the ViiA 7 platform (Life Technologies, California, USA). To quantify the viral load, a standard curve based on DENV genome previously cloned in the pGEM Easy plasmid (Promega, USA) was used.
2.6. Quantification of miRNA Levels by RT-qPCR
The commercial kit TaqMan MicroRNA Cells-to-CT (Ambion, California, USA) was used to measure the levels of miRNAs-15/16, according to the manufacturer’s instructions. Endogenous controls were made from the targets of RNU48 (SNORD48) and RNU58a (the gene for ribosomal protein L17). The quantitative PCR was performed on the ViiA 7 platform (Life Technologies, California, USA).
Relative quantification was performed using the method described by Livak et al. [15]. To visualize the miRNA quantification data, Expression Suite v1.0 (Applied Biosystems, California, USA) and the R Project's qPCR.Ct package were used. Each of the time points analyzed was performed in biological triplicates to ensure reliability of the data.
2.7. NS1 Quantification
The amount of NS1 protein was measured with the commercial kit Platelia Dengue NS1 Ag (Bio-Rad, Hercules, California, USA) and a protein curve built with a synthetic protein made from an expression plasmid. This quantification was performed using supernatants from cells infected with DENV-4.
2.8. Apoptosis Detection
To quantify the activity of caspases-3/7, a plate assay was performed with DENV-infected cells and the commercial Caspase-Glo 3/7 Assay kit (Promega, /USA) on the Glomax-Multi+ platform (Promega, USA), according to the manufacturer's instructions. The values of uninfected control (MOCK) were compared with the infection times. This experiment was carried out in technical and biological triplicates.
2.9. NS1 Fluorescent Imaging
Up to 120 hours after DENV-4 infection, Huh 7.5 cells were moved to a microscope slide and fixed with 3.7% formaldehyde. Subsequently, 3% bovine serum albumin (Sigma-Aldrich) was used to block nonspecific antibody binding sites for 1 hour. The cells were then put in a 1:20 solution of FITC-conjugated DENV NS1 polyclonal antibody (Biorbyt) at 4 °C for 16 hours. After putting the coverslip on, samples were visualized on an LSM 510 META laser-scanning confocal fluorescence microscope (Carl Zeiss, Colony, Germany) with excitation at 488 nm and emission collected from 500 to 550 nm. Images were processed using ImageJ v1.48 (National Institutes of Health, Bethesda, USA).
2.10. Statistical Analysis
Expression Suite v1.0 (Applied Biosystems, California, USA) and the R Project's qPCR.Ct package were used to statistically analyze the data. Analysis of variance (ANOVA) and Pearson correlation were performed on the Jamovi Project 2.3 platform (jamovi.org). P values less than 0.05 were considered statistically significant.
3. Results
3.1. Viral Load Profile During Infection
At first, we looked at the amount of virus particles in DENV-4-infected cell lines up to 120 hpi, to find out when the number of viruses was increasing the most (Figure 1). As observed, 72 hpi was the time point when both cell lines had the highest viral load.
3.2. Expression Levels of miRNAs-15/16 During Infection
At different time points after DENV-4 infection, qRT-PCR was used to assess the levels of miRNAs-15/16 in the culture supernatants of Huh 7.5 and HepG2 cells.
Expression of miRNA-15 showed a slight change between 24 and 48 hpi. However, the most significant increase was at 72 hpi; after this substantial increase, the miRNA-15 levels remained stable (Figure 2A,B).
Expression of miRNA-16 showed a significant increase at 48 hpi, which continued until reaching the highest level at 72 hpi, in both studied cells. However, there was a significant reduction at 96 hpi, reaching the lowest miRNA-16 level at 120 hpi (Figure 2C,D). Even though both cell lines showed negative expression values for this miRNA at 24 hpi, there was no statistically significant difference between these values and those at the time point before infection, which we called 0 hpi.
3.3. Activation of Caspases-3/7 During Infection
Next, we investigated how the apoptosis pathway was turned on during DENV-4 infection by measuring the activity of caspases-3/7 (Figure 3). The results showed that caspase-3/7 activity was highest at 72 hpi in both cell lines. This activity then went down at 96 hpi and reached its lowest level at 120 hpi.
3.4. NS1 Expression During Infection
Afterwards, we evaluated the expression of NS1 protein during DENV-4 infection in HepG2 and Huh 7.5 cells (Figure 4). As observed, the NS1 levels were similar for both cell lines in this experiment, continuously increasing throughout the course of the viral infection.
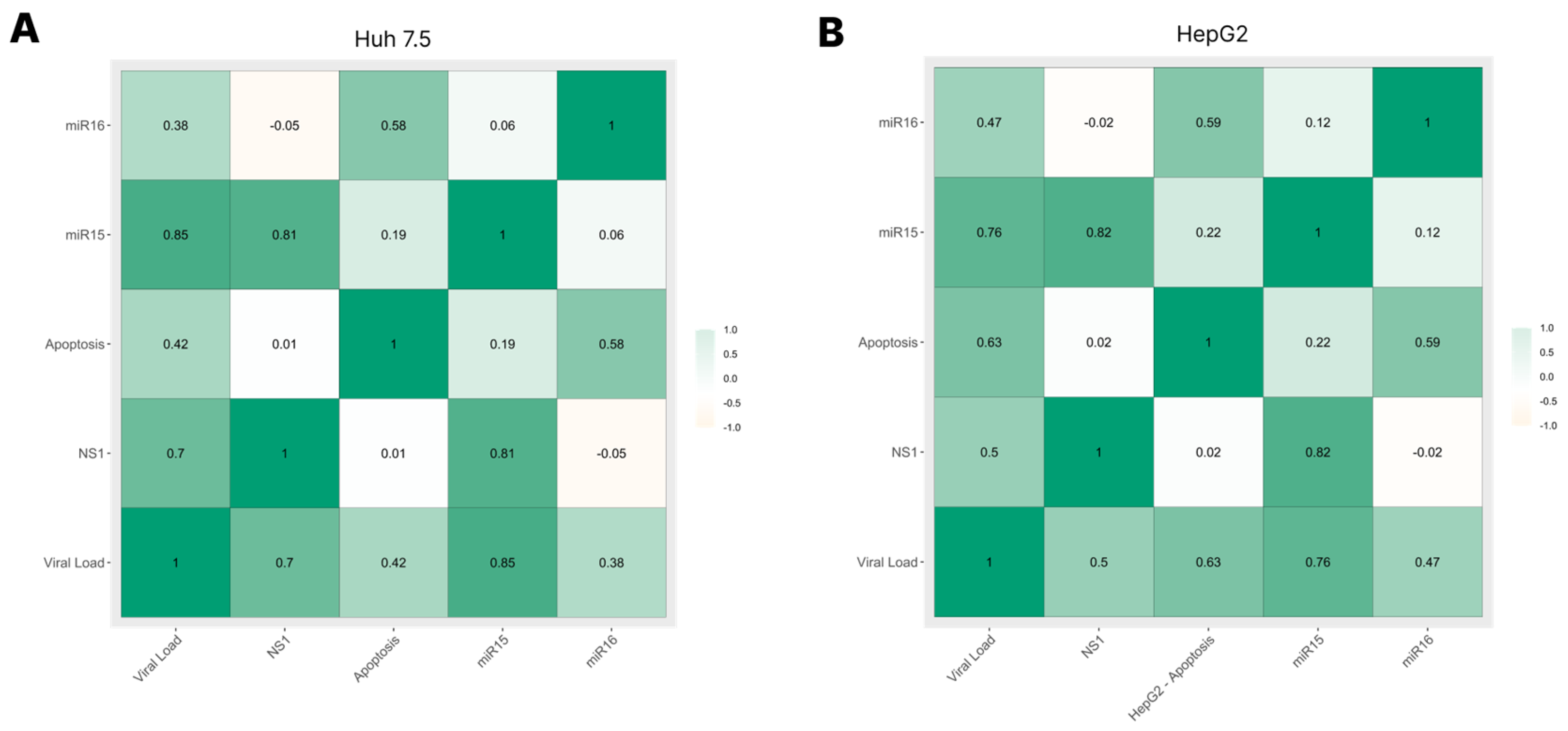
3.5. Correlation among Viral Load, NS1 Levels, miRNA-15/16 Expression, and Caspase-3/7 Activity
Then, an analysis of correlation among the previous data was performed (Figure 5). We observed that Huh 7.5 and HepG2 cell lines behaved similarly, with a high positive correlation between miRNA-15 expression and both NS1 levels and viral load and a moderate positive correlation between miRNA-16 expression and caspase-3/7 activity.
4. Discussion
DENV is a flavivirus with a vast worldwide dispersion. Even with the advances in knowledge about this virus, several questions remain obscure: along with the role of miRNA in DENV infection, one of them is whether viral proteins can act as signaling factors.
Studies such as those by Casseb et al. [16] and Castillo and Urcuqui-Inchim [17] showed that DENV infection alters the way in which the main proteins related to miRNA biogenesis are controlled. Differential expression of these proteins during infection by other flaviviruses were previously observed, as in the work by Holanda et al. [18], who observed a significant change in AGO2 during YFV infection.
Schafer et al. [19] described that members of the miRNA-15/16 family, more specifically miR-424, are able to suppress the expression of the E3 ubiquitin ligase SIAH1, which is normally induced during DENV-2 infection through activation of the unfolded protein response (UPR), thus suggesting that miRNAs belonging to the miRNA-15/16 family have an important role during DENV replication.
It should be noted that miRNAs are important in regulatory networks that shape effective T-cell responses by fine-tuning thousands of genes. Studies in mice demonstrate that miRNAs of the miRNA-15/16 family restrict the cycle, survival, and differentiation of memory T cells [20]. Despite not being the focus of our work, this demonstrates how this family of miRNAs plays an important role in cell cycle and survival.
Interestingly, viral proteins may be related to miRNA [21]. One of these proteins is NS1, which is released from infected cells and can be uptaken and stored by hepatocytes. As there is evidence that DENV affects liver function, especially in SD, and NS1 interacts with hepatocytes, these cells are suitable for studying and testing how NS1 are involved in dengue immunopathology. In addition, there is evidence that NS1 can alter cell signaling pathways to help produce more viruses and bypass host defenses [9,22,23].
This study analyzed how miRNA-15/16 expression and NS1 protein levels change over time during infection of Huh 7.5 and HepG2 cells with DENV-4. When the virus replicated in these human hepatocyte cell lines, we observed viral load profiles similar to those described by Alhoot et al. [24].
It has been shown that miRNA-15 is a key regulator of apoptosis, and low expression of this miRNA has been linked to cancer [25,26]. Papillomavirus infections have also been shown to alter the levels of miRNA-15 in cells, having important effects on the p53 signaling pathway [27,28,29]. Even though studies have shown that DENV infection promotes apoptosis [30,31], the molecular basis of this induction is not well understood.
As miRNA-15, miRNA-16 is also involved in apoptotic pathways and cell growth during oncogenic processes [20,30,32,33]. The miRNA-16 levels have also been investigated in viral infections. A recent study by El-Abd et al. [34] showed that its regulatory function is diminished when Hepatitis C virus causes a long-term infection, which can lead to hepatocellular carcinoma. In this study, however, we showed that the expression of both miRNAs goes up during acute DENV-4 infection, along with an increase in the expression of NS1 protein and the activity of the apoptosis markers caspases-3/7.
According to [35], miRNAs-15/16 promote downregulation of Bcl-2 – a family of proteins that prevent cell death – by a direct (miRNA-mRNA complementarity) or indirect interaction, which would lead to a change in the cell cycle and then apoptosis. In our study, DENV-4 NS1 levels increased along with caspase-3/7 activity up to 72 hpi (Figure 3), suggesting that this viral protein plays a role in apoptosis during DENV-4 infection in liver cells. This finding may be clinically important and may help explain why apoptosis is observed in the liver cells of people who died from dengue and yellow fever [26,36].
Furthermore, NS1 can interact with Beclin-1 during DENV infection, and this interaction attenuates Beclin-1 cleavage and facilitates autophagy to prevent cell apoptosis [37]. However, as shown by Lu et al. [38], elevated caspases trigger apoptosis by degrading Beclin-1 in the late stage of infection.
Antibodies against NS1 can produce a protective immune response and induce complement-fixing activity. When released into the extracellular medium, NS1 acts as a pathogen-associated molecular pattern (PAMP), which directly activates macrophages and peripheral blood mononuclear cells (PBMCs) via Toll-like receptor 4 (TLR-4), causing them to produce and release pro-inflammatory cytokines and chemokines [23,39].
It was previously shown that, during DENV-1 infection in HepG2 cells, the NS1 protein can be found in lipid raft domains on the host cell surface, suggesting its involvement in signal transduction events [40]. In addition, the authors of this study found that the NF-kB p65 protein moves to the nucleus when NS1 is expressed in HepG2 cells. More DENV infections have recently been linked to NF-kB activation, which leads to endothelial cell death by apoptosis and bleeding symptoms [28,41].
5. Conclusions
This study showed that miRNAs-15/16 are overexpressed during DENV-4 infection in human hepatocyte cell lines. NS1 protein production, viral load, and apoptosis are linked to this finding, suggesting a critical interplay between them. Thus, we consider that miRNAs-15/16 have a significant potential for use as disease markers during DENV infection in human hepatocytes.
Author Contributions
Samir Casseb: Methodology, data analysis, and manuscript writing. Karla Melo: Methodology and data analysis. Carlos Carvalho: Methodology and manuscript revision. Carolina Santos: data analysis. Edna Franco: manuscript writing; Pedro Vasconcelos: manuscript revision.
Funding
This study was supported by grants from CNPq/CAPES/FAPESPA 573739/2008-0 and CNPq 301641/2010-2.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors are grateful to Evandro Chagas Institute for providing laboratory facilities, assay reagents and viral strains.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Horstick, O.; Tozan, Y.; Wilder-Smith, A. Reviewing dengue: still a neglected tropical disease? PLoS Negl. Trop. Dis. 2015, 9, e0003632. [Google Scholar] [CrossRef]
- Li, G.; Khandekar, A.; Yin, T.; Hicks, S.C.; Guo, Q.; Takahashi, K.; Lipovsky, C.E.; Brumback, B.D.; Rao, P.K.; Weinheimer, C.J.; Rentschler, S.L. Differential Wnt-mediated programming and arrhythmogenesis in right versus left ventricles. J. Mol. Cell. Cardiol. 2018, 123, 92–107. [Google Scholar] [CrossRef]
- Siqueira, J.B.; Martelli, C.M.T.; Coelho, G.E.; Simplicio, A.C. da R.; Hatch, D.L. Dengue and dengue hemorrhagic fever, Brazil, 1981-2002. Emerging Infect. Dis. 2005, 11, 48–53. [Google Scholar] [CrossRef]
- Azeredo, E.L.; Dos Santos, F.B.; Barbosa, L.S.; Souza, T.M.A.; Badolato-Corrêa, J.; Sánchez-Arcila, J.C.; Nunes, P.C.G.; de-Oliveira-Pinto, L.M.; de Filippis, A.M.; Dal Fabbro, M.; Hoscher Romanholi, I.; Venancio da Cunha, R. Clinical and Laboratory Profile of Zika and Dengue Infected Patients: Lessons Learned From the Co-circulation of Dengue, Zika and Chikungunya in Brazil. PLoS Curr. Influenza 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Díaz, D.A.; Gutiérrez-Díaz, A.A.; Orozco-García, E.; Puerta-González, A.; Bermúdez-Santana, C.I.; Gallego-Gómez, J.C. Dengue virus potentially promotes migratory responses on endothelial cells by enhancing pro-migratory soluble factors and miRNAs. Virus Res. 2019, 259, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Hafirassou, M.L.; Meertens, L.; Umaña-Diaz, C.; Labeau, A.; Dejarnac, O.; Bonnet-Madin, L.; Kümmerer, B.M.; Delaugerre, C.; Roingeard, P.; Vidalain, P.-O.; Amara, A. A global interactome map of the dengue virus NS1 identifies virus restriction and dependency host factors. Cell Rep. 2017, 21, 3900–3913. [Google Scholar] [CrossRef] [PubMed]
- Yen, P.-S.; Chen, C.-H.; Sreenu, V.; Kohl, A.; Failloux, A.-B. Assessing the potential interactions between cellular miRNA and arboviral genomic RNA in the yellow fever mosquito, Aedes aegypti. Viruses 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S. Recent advances in understanding dengue. [version 1; peer review: 2 approved]. F1000Res. 2019, 8. [Google Scholar] [CrossRef]
- Halstead, S.B.; Russell, P.K.; Brandt, W.E. NS1, Dengue’s Dagger. J. Infect. Dis. 2020, 221, 857–860. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microrna biogenesis, mechanisms of actions, and circulation. Front Endocrinol (Lausanne) 2018, 9, 402. [Google Scholar] [CrossRef]
- Huang, Y.; Shen, X.J.; Zou, Q.; Wang, S.P.; Tang, S.M.; Zhang, G.Z. Biological functions of microRNAs: a review. J. Physiol. Biochem. 2011, 67, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Dou, Y.; Chen, L.; Wang, J.; Jiang, N.; Guo, C.; Yao, Q.; Wang, C.; Liu, L.; Yu, B.; Zheng, B.; Chekanova, J.A.; Ma, J.; Ren, G. Degradation of unmethylated miRNA/miRNA*s by a DEDDy-type 3’ to 5’ exoribonuclease Atrimmer 2 in Arabidopsis. Proc Natl Acad Sci USA 2018, 115, E6659–E6667. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Yu, X.; Hu, S.; Yu, J. A brief review on the mechanisms of miRNA regulation. Genomics Proteomics Bioinformatics 2009, 7, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.W.; Russell, B.J.; Lanciotti, R.S. Serotype-specific detection of dengue viruses in a fourplex real-time reverse transcriptase PCR assay. J. Clin. Microbiol. 2005, 43, 4977–4983. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Casseb, S.M.M.; Simith, D.B.; Melo, K.F.L.; Mendonça, M.H.; Santos, A.C.M.; Carvalho, V.L.; Cruz, A.C.R.; Vasconcelos, P.F.C. Drosha, DGCR8, and Dicer mRNAs are down-regulated in human cells infected with dengue virus 4, and play a role in viral pathogenesis. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Castillo, J.A.; Urcuqui-Inchima, S. Letter to the Editor “Drosha, DGCR8, and Dicer mRNAs are downregulated in human cells infected with dengue virus 4” - Genet. Mol. Res. 15 (2): gmr.15027891 - Drosha, Dicer, and TRBP mRNA are downregulated in Vero cells with the 3’UTR of Dengue virus. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Holanda, G.M.; Casseb, S.M.M.; Mello, K.F.L.; Vasconcelos, P.F.C.; Cruz, A.C.R. Yellow Fever Virus Modulates the Expression of Key Proteins Related to the microRNA Pathway in the Human Hepatocarcinoma Cell Line HepG2. Viral Immunol. 2017, 30, 336–341. [Google Scholar] [CrossRef]
- Murphy Schafer, A.R.; Smith, J.L.; Pryke, K.M.; DeFilippis, V.R.; Hirsch, A.J. The E3 ubiquitin ligase SIAH1 targets myd88 for proteasomal degradation during dengue virus infection. Front. Microbiol. 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, J.D.; Kageyama, R.; Shehata, H.M.; Fassett, M.S.; Mar, D.J.; Wigton, E.J.; Johansson, K.; Litterman, A.J.; Odorizzi, P.; Simeonov, D.; Laidlaw, B.J.; Panduro, M.; Patel, S.; Jeker, L.T.; Feeney, M.E.; McManus, M.T.; Marson, A.; Matloubian, M.; Sanjabi, S.; Ansel, K.M. miR-15/16 Restrain Memory T Cell Differentiation, Cell Cycle, and Survival. Cell Rep. 2019, 28, 2169–2181.e4. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Kumar, A.; Ingle, H.; Kumar, H. The Interplay Between Viral-Derived miRNAs and Host Immunity During Infection. Front. Immunol. 2019, 10, 3079. [Google Scholar] [CrossRef]
- Acosta-Ampudia, Y.; Monsalve, D.M.; Castillo-Medina, L.F.; Rodríguez, Y.; Pacheco, Y.; Halstead, S.; Willison, H.J.; Anaya, J.-M.; Ramírez-Santana, C. Autoimmune neurological conditions associated with zika virus infection. Front. Mol. Neurosci. 2018, 11, 116. [Google Scholar] [CrossRef]
- Modhiran, N.; Watterson, D.; Muller, D.A.; Panetta, A.K.; Sester, D.P.; Liu, L.; Hume, D.A.; Stacey, K.J.; Young, P.R. Dengue virus NS1 protein activates cells via Toll-like receptor 4 and disrupts endothelial cell monolayer integrity. Sci. Transl. Med. 2015, 7, 304ra142. [Google Scholar] [CrossRef]
- Alhoot, M.A.; Wang, S.M.; Sekaran, S.D. RNA interference mediated inhibition of dengue virus multiplication and entry in HepG2 cells. PLoS ONE 2012, 7, e34060. [Google Scholar] [CrossRef] [PubMed]
- Aqeilan, R.I.; Calin, G.A.; Croce, C.M. miR-15a and miR-16-1 in cancer: discovery, function and future perspectives. Cell Death Differ. 2010, 17, 215–220. [Google Scholar] [CrossRef]
- de Oliveira, L.F.; de Andrade, A.A.S.; Pagliari, C.; de Carvalho, L.V.; Silveira, T.S.; Cardoso, J.F.; Silva, A.L.T.E.; de Vasconcelos, J.M.; Moreira-Nunes, C.A.; Burbano, R.M.R.; Nunes, M.R.T.; Dos Santos, E.J.M.; Júnior, J.L. da S.G.V. Differential expression analysis and profiling of hepatic miRNA and isomiRNA in dengue hemorrhagic fever. Sci. Rep. 2021, 11, 5554. [Google Scholar] [CrossRef]
- Chen, C.-L.; Lin, C.-F.; Wan, S.-W.; Wei, L.-S.; Chen, M.-C.; Yeh, T.-M.; Liu, H.-S.; Anderson, R.; Lin, Y.-S. Anti-dengue virus nonstructural protein 1 antibodies cause NO-mediated endothelial cell apoptosis via ceramide-regulated glycogen synthase kinase-3β and NF-κB activation. J. Immunol. 2013, 191, 1744–1752. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-C.; Lin, S.-C.; Chen, W.-Y.; Yen, Y.-T.; Lai, C.-W.; Tao, M.-H.; Lin, Y.-L.; Miaw, S.-C.; Wu-Hsieh, B.A. Dengue viral protease interaction with NF-κB inhibitor α/β results in endothelial cell apoptosis and hemorrhage development. J. Immunol. 2014, 193, 1258–1267. [Google Scholar] [CrossRef]
- Ouyang, X.; Jiang, X.; Gu, D.; Zhang, Y.; Kong, S.K.; Jiang, C.; Xie, W. Dysregulated Serum MiRNA Profile and Promising Biomarkers in Dengue-infected Patients. Int. J. Med. Sci. 2016, 13, 195–205. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Tsukiyama-Kohara, K. Mammalian animal models for dengue virus infection: a recent overview. Arch. Virol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Nanaware, N.; Banerjee, A.; Mullick Bagchi, S.; Bagchi, P.; Mukherjee, A. Dengue virus infection: A tale of viral exploitations and host responses. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Wong, R.R.; Abd-Aziz, N.; Affendi, S.; Poh, C.L. Role of microRNAs in antiviral responses to dengue infection. J. Biomed. Sci. 2020, 27, 4. [Google Scholar] [CrossRef] [PubMed]
- Linsley, P.S.; Schelter, J.; Burchard, J.; Kibukawa, M.; Martin, M.M.; Bartz, S.R.; Johnson, J.M.; Cummins, J.M.; Raymond, C.K.; Dai, H.; Chau, N.; Cleary, M.; Jackson, A.L.; Carleton, M.; Lim, L. Transcripts targeted by the microRNA-16 family cooperatively regulate cell cycle progression. Mol. Cell. Biol. 2007, 27, 2240–2252. [Google Scholar] [CrossRef]
- El-Abd, N.E.; Fawzy, N.A.; El-Sheikh, S.M.; Soliman, M.E. Circulating miRNA-122, miRNA-199a, and miRNA-16 as Biomarkers for Early Detection of Hepatocellular Carcinoma in Egyptian Patients with Chronic Hepatitis C Virus Infection. Mol. Diagn. Ther. 2015, 19, 213–220. [Google Scholar] [CrossRef]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; Rassenti, L.; Alder, H.; Volinia, S.; Liu, C.-G.; Kipps, T.J.; Negrini, M.; Croce, C.M. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc Natl Acad Sci USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef]
- Pagliari, C.; Quaresma, J.A.S.; Fernandes, E.R.; Stegun, F.W.; Brasil, R.A.; de Andrade, H.F.; Barros, V.; Vasconcelos, P.F.C.; Duarte, M.I.S. Immunopathogenesis of dengue hemorrhagic fever: contribution to the study of human liver lesions. J. Med. Virol. 2014, 86, 1193–1197. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.; Gyeltshen, S.; Chaijaroenkul, W.; Na-Bangchang, K. Significance of autophagy in dengue virus infection: A brief review. Am. J. Trop. Med. Hyg. 2019, 100, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.-Y.; Cheng, M.-H.; Yu, C.-Y.; Lin, Y.-S.; Yeh, T.-M.; Chen, C.-L.; Chen, C.-C.; Wan, S.-W.; Chang, C.-P. Dengue Nonstructural Protein 1 Maintains Autophagy through Retarding Caspase-Mediated Cleavage of Beclin-1. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Arboleda, J.F.; Fernandez, G.J.; Urcuqui-Inchima, S. Vitamin D-mediated attenuation of miR-155 in human macrophages infected with dengue virus: Implications for the cytokine response. Infect. Genet. Evol. 2019, 69, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.M.; Sousa, L.P.; Gomes-Ruiz, A.C.; Leite, F.G.G.; Teixeira, M.M.; da Fonseca, F.G.; Pimenta, P.F.P.; Ferreira, P.C.P.; Kroon, E.G.; Bonjardim, C.A. The dengue virus nonstructural protein 1 (NS1) increases NF-κB transcriptional activity in HepG2 cells. Arch. Virol. 2011, 156, 1275–1279. [Google Scholar] [CrossRef]
- Puerta-Guardo, H.; Glasner, D.R.; Espinosa, D.A.; Biering, S.B.; Patana, M.; Ratnasiri, K.; Wang, C.; Beatty, P.R.; Harris, E. Flavivirus NS1 Triggers Tissue-Specific Vascular Endothelial Dysfunction Reflecting Disease Tropism. Cell Rep. 2019, 26, 1598–1613.e8. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Quantification of viral load in the culture supernatant by RT-qPCR as a function of time after DENV-4 infection in Huh 7.5 (A) and HepG2 (B) cells. Statistical analysis was performed using ANOVA with Tukey's post-test.
Figure 1.
Quantification of viral load in the culture supernatant by RT-qPCR as a function of time after DENV-4 infection in Huh 7.5 (A) and HepG2 (B) cells. Statistical analysis was performed using ANOVA with Tukey's post-test.
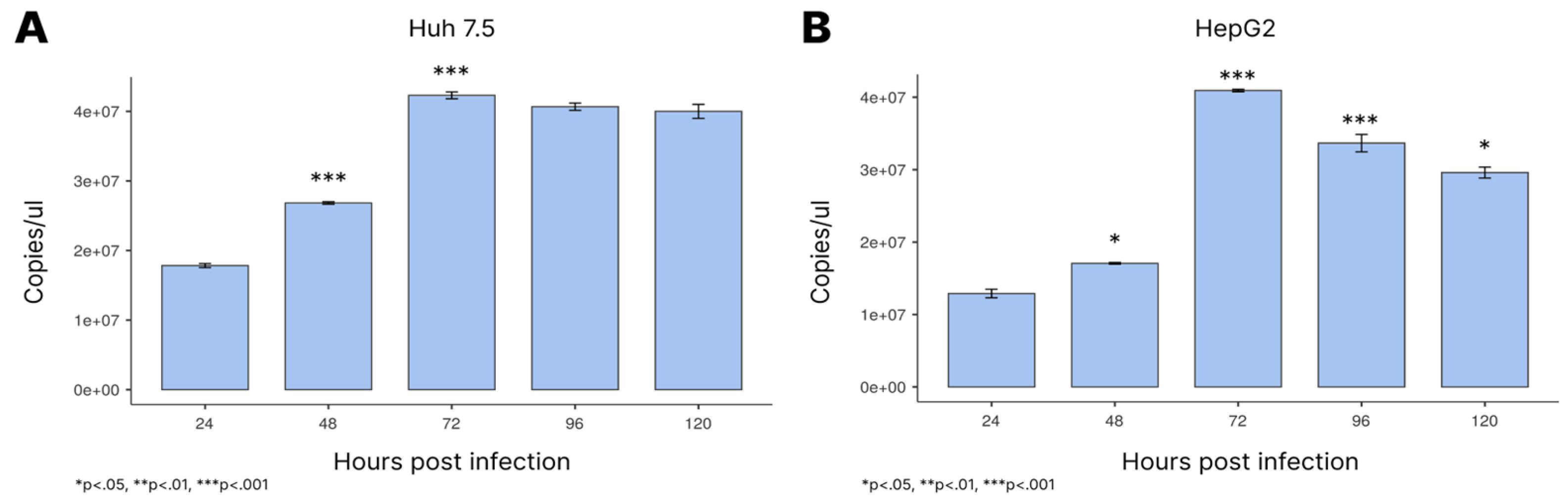
Figure 2.
Quantification of miRNA-15 (A and B) and miRNA-16 (C and D) levels as a function of time after infection of Huh 7.5 (A and C) and HepG2 (B and D) cells with DENV-4 by RT-qPCR. Statistical analysis was performed using ANOVA with Tukey's post-test.
Figure 2.
Quantification of miRNA-15 (A and B) and miRNA-16 (C and D) levels as a function of time after infection of Huh 7.5 (A and C) and HepG2 (B and D) cells with DENV-4 by RT-qPCR. Statistical analysis was performed using ANOVA with Tukey's post-test.
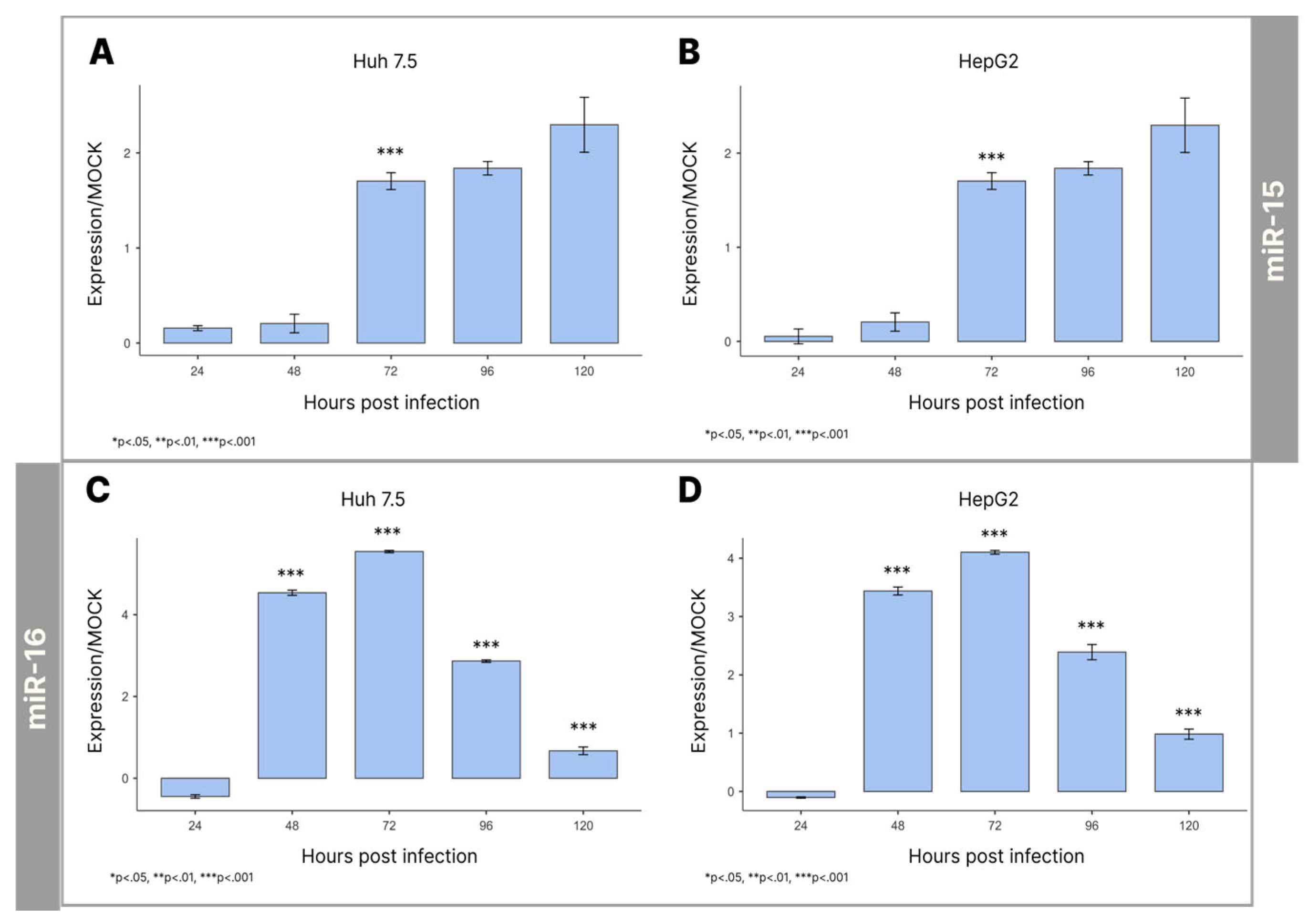
Figure 3.
Quantification of caspase-3/7 activity as a function of time after infection of Huh 7.5 (A) and HepG2 (B) cells with DENV-4 by luminescent assay. Statistical analysis was performed using ANOVA with Tukey's post-test.
Figure 3.
Quantification of caspase-3/7 activity as a function of time after infection of Huh 7.5 (A) and HepG2 (B) cells with DENV-4 by luminescent assay. Statistical analysis was performed using ANOVA with Tukey's post-test.
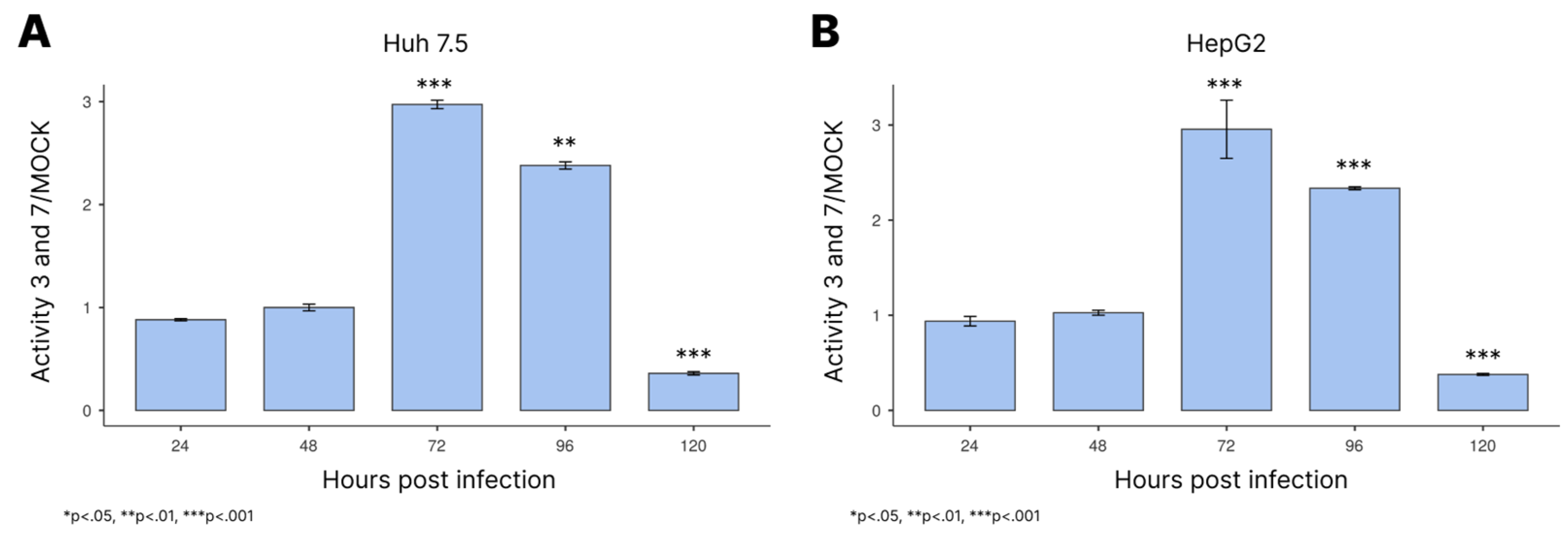
Figure 4.
Quantification of NS1 levels in Huh 7.5 (A and C) and HepG2 (B) cells following DENV-4 infection by enzyme immunoassay (A and B) and laser-scanning confocal fluorescence microscopy (C). Statistical analysis was performed using ANOVA with Tukey's post-test.
Figure 4.
Quantification of NS1 levels in Huh 7.5 (A and C) and HepG2 (B) cells following DENV-4 infection by enzyme immunoassay (A and B) and laser-scanning confocal fluorescence microscopy (C). Statistical analysis was performed using ANOVA with Tukey's post-test.
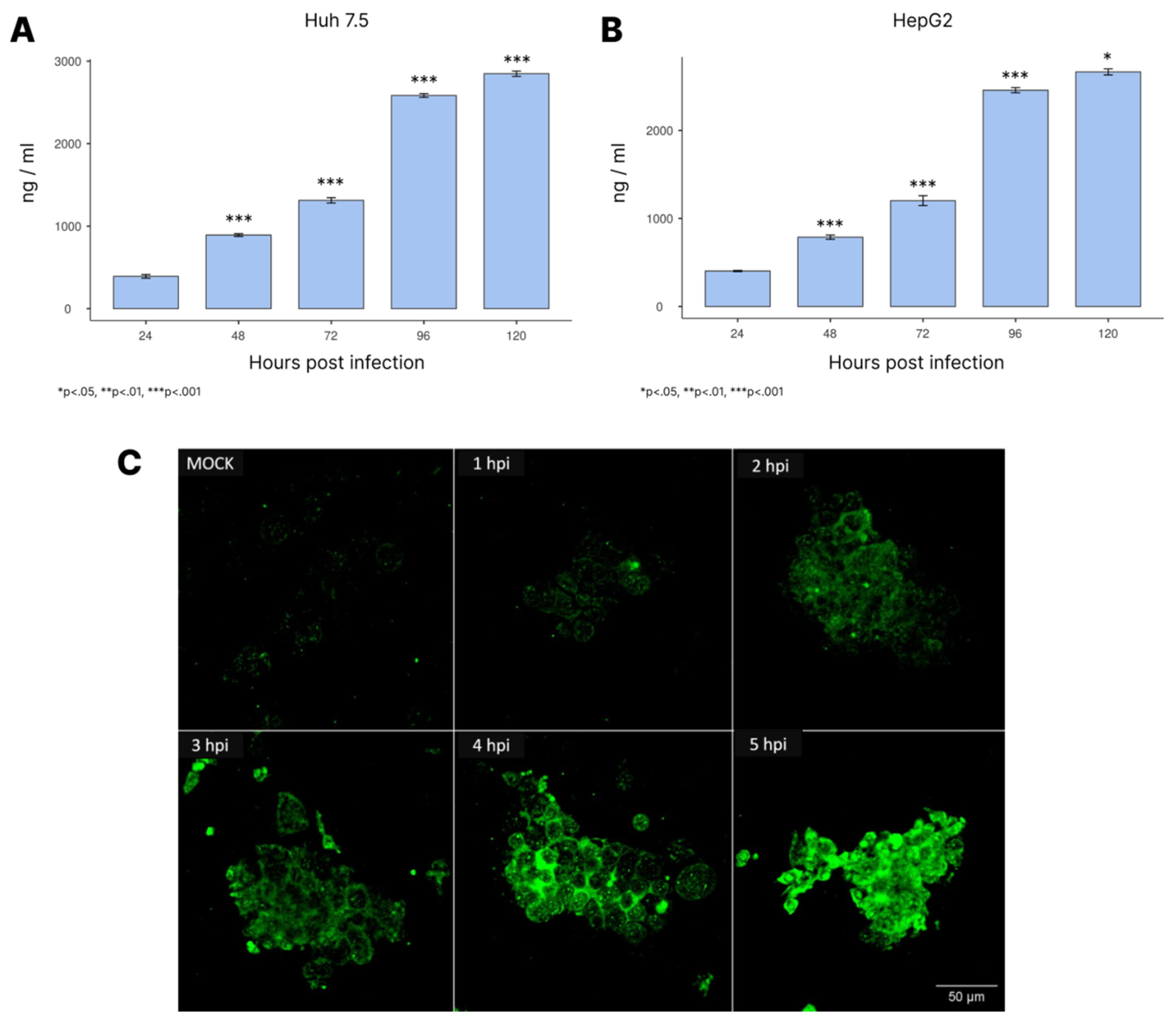
Figure 5.
Pearson correlation analysis between viral load, NS1 levels, miRNA-15/16 expression, and caspase-3/7 activity during DENV-4 infection in Huh 7.5 (A) and HepG2 (B) cell lines.
Figure 5.
Pearson correlation analysis between viral load, NS1 levels, miRNA-15/16 expression, and caspase-3/7 activity during DENV-4 infection in Huh 7.5 (A) and HepG2 (B) cell lines.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



