Submitted:
29 December 2022
Posted:
03 January 2023
You are already at the latest version
Abstract
Background: Visceral leishmaniasis affects approximately 12 million people worldwide. Current existing drugs cause several side effects and resistance, the identification of new therapeutic targets for the development of new drugs is necessary. The vesicular trafficking in the parasite flagellar pocket is essential for parasite survival and proliferation, but little is known about the proteins involved in the endocytic/secretory pathways. In this work, we are describing the characterization of a transporter cargo protein SFT-2/Got-1 like in Leishmania infantum. Material and Methods: Using bioinformatics tools, the putative Got-1 protein A4HSK3 was characterized structurally and phylogenetically. Molecular biology was used to clone and express the protein and confocal fluorescence to determine its subcellular localization. Results: The Got-1 like protein presents four alpha helix TM conserved domains (aa38-aa147) characteristics of the Got-1/SFT-2 family proteins and an important Pen-2 conserved motif LGDYXXF located in the extracellular loop. Confocal microscopy suggests that Got-1/SFT-2 like is localized in the flagellar pocket, and is partially co-localized with the L. infantum presenilin-like protein. Phylogenetically, the Leishmania sp Got-1 like is contained in a subgroup of proteins clearly detached from the other Trypanosomatidae, such as T. cruzi and T. brucei. Conclusions: Our data show the presence of an important transported proteins, and its colocalization with the presenilin aspartyl protease in the flagellar pocket of the L. infantum open new perspectives to identify novel targets, which can be an alternative therapeutic strategy against leishmaniasis.
Keywords:
Leishmania infantum
; STF2/Got-1 family
; cloning
; cellular localization
; membrane topology
1. Introduction
Currently, it is estimated 50 000 to 90 000 new cases of Visceral Leishmaniasis (VL) occur worldwide annually, it is transmitted by many species of sandfly and different Leishmania species [1]. VL is highly lethal if not treated, symptoms include fever, weight loss, hepatosplenomegaly and anemia [1]. It is endemic in some regions of South America, Indian subcontinent and East Africa [1].
Membrane compartmentalization is a fundamental mechanism for the eukaryotic homeostasis, in which cells can separate and control the biological processes. The growth, multiplication, development, and adaptation of leishmaniasis to biotic and abiotic stress rely on a highly dynamic of endomembrane system. However, very little is known, in trypanosomatids, regarding the proteins that constitute this system and the regulation of molecular membrane traffic [2]. In trypanossomatids the fagellar pocket is the main subcellular domain of secretory and endocytic pathways. Information about proteins that constitute and regulate this process are still limited. Although, genomic analysis revealed that Leishmania encode many putative membrane transporters during parasite life cycle, few were characterized [2].
Although there are other soluble cargo secretory pathways [3] the Golgi apparatus is the main organelle in which newly synthesized and glycated transmembrane proteins are translocated to plasmatic membrane. Recent studies highlight the secretory pathways as relevant virulence factor, lipophosphoglycan and GP63 are released by the parasite and redistribute within the host cell, GP63 cleaves components of the host cell membrane fusion machinery [4]. In trypanassomatids, fagellar pocket membrane transporter interacts with transmembrane peptidase complexes [5]. Proteases are intensively studied as therapeutic targets for treatment of Leishmaniasis [6]. In MEROPS database, there are just two Leishmania homologous proteins belonging to Clans AD and AA are described in the MEROPS [7], Presenilin-1 (PS) and Peptide Peptidase Signal (PPS) that cleave type I and II membrane proteins, respectively. The aspartic proteases found in Leishmania spp belong to the clans AA and AD. The AA clan contains most of the enzymes with classical activity, related to pH and microenvironment content, while the enzymes from the AD clan, are proteases that hydrolyze peptide bonds within biological membranes.
Presenilin (A22 family) is part of the clan AD and is the catalytic component of the gamma-secretase complex, necessary for the proteolysis of type I transmembrane proteins (PTMs), such as the protein amyloid precursor. Functional studies have shown that the two forms of human presenilins (PS1 and PS2) form the catalytic subunit of a complex called gamma-secretase (GSC). The GSC is located integrally in the reticular membranes and has as substrate several transmembrane proteins (PTM) type 1, including Amyloid Precursor Protein (APP) [8]. Mutations in the PSs lead to aberrant APP processing. Recently, the importance of of PSs in Alzheimer's Disease (AD) and of the GSC activator protein which is degraded via the proteasome, and a possible deregulation of this system, affects the activity of the complex, contributing to the emergence of AD in addition to other neurodegenerative diseases [8]. The complex is formed by a presenilin (PS1 or PS2 isoform) an intramembrane aspartyl protease as the catalytic component, along with nicastrin (Nct), anterior pharynx defective-1 (Aph1) and presenilin enhancer-2 (Pen-2).
Different cellular functions have been attributed to PS, including the promotion or reduced susceptibility of neurons to apoptosis [9], regulation of the calcium signaling mechanism [10], regulation of stability of catenins, traffic of membrane proteins, cleavage of APP-secretase in different cell types and action on several others substrates such as nectin-1 [11], E-cadherin-catenin [12], CD44 [13,14], Erb-4, epidermal growth factor receptor [15], involvement in catalytic activities of the Notch cascade [16] and regulation of calcium homeostasis [17]. Pen-2 is important for complex stability and trafficking, it is conserved retaining 56% similarity between Homo sapiens and Arabidopsis thaliana [18], but no similar protein is found in Leishmnia sp. Thus, we hypothesized that other protein may function as Pen-2.
Intracellular trafficking is important for parasite-host interplay, immune system evasion and nutrient uptake. The SNAREs (soluble N-ethylmaleimide-sensitive factor adaptor proteins receptors) family of proteins are important for docking and fusion of vesicles with organelles and contribute to organelle specificity [19]. Bioinformatic and genomic analysis revealed several L. major SNARE domain-containing proteins [20].
Got1 is a protein that facilitate fusion events of Sed5, a member of the syntaxin family of SNARE proteins. It is an evolutionary conserved non-essential protein [22], and a membrane topology with four putative transmembrane (TM) domains similar to that of Sft2. Both Got1 and Sft2 is required for vesicle fusion with the Golgi [23].
Although much is known about the involvement of presenilins with signal transduction in humans, very little is known in unicellular organisms and parasites. Recently, the PS-like protein [24] and some components of the GSC [25] was characterized in Trypanosoma cruzi and other parasites [26] by our group. During our characterization studies of this group of membrane protein in Leishmania sp, we did not identify any protein with significant similarity to Pen-2. However, we identified in a putative Got1 transporter protein a domain characteristic of Pen-2 proteins (Figure S3). Thus, we considered essential the characterization of this protein, since the interaction of TM and vesicular transport proteins can provide knowledge about the functions of these macromolecules in the life cycle of the parasite or on its infectivity process, pathophysiological role, parasite-host relationship and provide targets for therapeutic strategies.
2. Results
2.1. Homology search
The L. infantum presenilin-like gene has an identity of 31% and similarity of 2e-41 to orthologous human presenilin-2 (P49810), despite the low similarity index there was no doubt about the selection of the gene, due to the identification of conserved domains among presenilins. No similar proteins in L. infantum was found for Aph-1 and Nicastrin using Basic Local Alignment Search Tool (BLAST). However, a search employing the human Pen-2 sequence (Q9NZ42) did not result in selection of a sequence with high homology and/or similarity scores. But a putative protein, classified as Sft2/Got-1 vesicular transport protein (A4HSK3) presented some identity (35%; Figure S1). Therefore, we selected this gene and cloned to demonstrate its conservation and possible function in gamma-secretase complex (Table 1).
2.2. Genomic DNA extraction and Got-1-like protein cloning
Genomic DNA was extracted from L.infantum culture (Strain IOC-579). The sequence of the gene from Got-1-like was obtained from GenBank and primers were designed. The selected region of Got-1-like genes were successfully cloned into the pGEM-T easy vector (Promega, Madison, Wisconsin, USA), with nine positive colonies for Got-1-like with its sequence confirmed by SANGER sequencing. From all the colonies selected, seven clones of each gene did not show mutations. The Got-1-like gene isolated from L. infantum consisted of an ORF of 492 bp encoding a polypeptide of 163 amino acids and with a calculated molecular weight of 17 kDa. One of the seven clones sequenced carrying the Got-1-like gene were subcloned into pNUS-GFP (Green Fluorescent Protein) expression vector, these plasmid was chosen for the following reasons (1) they have the ability to be expressed in the parasite itself, due to the existence of a replication region recognized in Leishmania and (2) knowledge of bacteria limitations to express eukaryotic proteins [23]. Ligation results are shown in Figure 1 and show that they were successful, as the expected molecular weight for the PCR amplification was around 500 bp.
Due to the limitations of transfection and maintenance of a gene within the parasite, a large-scale plasmid DNA extraction was performed to purify pNUS-GFP-Got-1-like. After confirming the ligation between the Got-1-like gene to the expression vectors, they were submitted to the protocol for extracting and purifying DNA in higher concentration. The ligation of the Got-1-like gene yielded 245 μg and the same way, the presence of the gene was confirmed by PCR. Approximately 10 µg of the plasmid fused to the Got-1 gene of L. infantum was used in the transfection for promastigotes of L. infantum. Four conditions were used to evaluate the experiment. (1) one wild type (WT), (2) one MOCK, (3) one using the empty plasmid and (4) one using the gene-fused to the plasmid. The period of recovery of the parasites after the stress undertaken by the technique of electroporation (in medium without selection antibiotic) was 24 hours. after this period, the selection antibiotic G418 was added at a concentration of 50 µg/mL and, after 48h of incubation, no viable individuals were detected both in the WT and MOCK samples and only samples with empty plasmid and with plasmid pNUS-GFP fused to Got-1-like showed cell growth, giving these individuals resistance to the antibiotic. The results of PCR from this analysis are shown in Figure 2 and confirm that the region that confers antibiotic resistance, of 127 bp, is not present in individuals wild.
2.2. Cellular localization of PS-like and Got-1-like
The cellular localization of Got-1/Sft2 like protein of L. infantum was determined by fluorescence microscopy using the Got-1/Sft2 GFP (Figure 3), and its colocalization with the presenilin was performed using antibodies anti-Leishmnia-PS-like. GFP fluorescence was observed distributed along the body of promastigotes suggesting distinct subdomains.. Promastigotes displayed a stronger staining signal for Got-1/Sft2 GFP protein in the anterior region near the kinetoplast and flagellum (Figure 3; Figure S2). Additionally, PS imunnolabeled staining was also found in the anterior region of promastigotes near the kinetoplast. Colocalization analysis revealed a certain degree of colocalization (30%, yellow), suggesting that proteins could interact in the same subcellular location (Figure S4).
2.3. Topology and conserved motifs of SFT2/Got-1-like
All Got-1 family members have conserved domains composed of four alpha helix transmembrane regions (aa38 –aa147). Using the Uniprot predicted structural model the four conserved membrane spanning regions were higlighted (TM1 = 40-59; TM2 = 65-89; TM3 = 101-122; TM4 = 128 -149). The model presented in Figure 4 was generated using Alphafold and analysis with the Orientation Membrane Protein access showed that some residues (aa 48– aa 54 and 58) could attach to the membrane, large intracelular membrane loops were identified, but few residues were found extracellularly. Sequence alignment revealed that L. infantum Got1/Sft2 like protein have a Pen-2 important conserved motif LGDYXXF located in the extracellular loop (Figure 4, left side and Figure S3).
2.4. Phylogenetic analysis
A phylogenetic tree using sequences of L. infantum Got1/Sft2 like protein was performed first on the Uniprot server (http://www.uniprot. Org; Accessed on 12 October 2020). Information of the sequence A4HSK3 in Uniprot server shows “Vesicle transport protein” and protein status is predicted with domains showing to be SFT-2 family and Got-1. Multiple sequence analysis with A4HSK3 homolog sequence was performed retrieving data from InParanoid. The tree with the highest log likelihood is shown in Figure 5. L. infantum Got1/Sft2 seems to be a very divergent protein, from other protozoan (Entamoeba histolytica and Trichomonas vaginalis), probably assuming other roles or biochemical pathways. L. infantum Got1/Sft2 like protein was found in the same clade with other trypanosomatids, but phylogenetic distance from others organism and its human orthologs.
3. Discussion
The flagellar pocket is a site of intense vesicular trafficking, we identify in Leishmania presenilin like protein one component of the gamma-secretase complex. In higher eukaryotes the complex is composed of four proteins; Presenilin 1 or Presenilin 2, NIC, APH1 and PEN-2, whose presence is necessary for the physiological activity of the complex. As in human, PS can exist as two separate homologs, one of them suffering alternative splicing, it appears that the CGS of human cells may be present in six possible combinations of PS1 or PS2 [27,28,29]. In addition, one or more proteins regulators [eg. transmembrane trafficking protein, 21-kD TMP21[30] and GSAP activating protein-C-secretase [31], whose presence is not essential, can interact with the subset of the complex. GSC regulates intramembrane proteolysis of several types of proteins membranes [32,33]. As this subject is little known in lower eukaryotes, such as Leishmania sp, we proposed in this work to continue the characterization studies of the GSC initiated in our laboratory. In silico analysis carried out in this work using genomic banks of Leishmania sp, indicated the presence of PS, however no gene was further identified coding for Pen-2. It is known that the flagellar pocket of trypanosomatids is a region with intense proteolytic and secretory activity [34,35]. This is the first study to describe the presence of the PS-like enzyme, the complex gamma-secretase and a SNARE like protein Sft2/Got-1 like in this compartment. In this work, this protein was initially identified as Pen-2 through homology. However, during the development of our study and using several other tools, we concluded that the sequence corresponded to the Sft2/Got-1 protein. Pen-2 is important for GSC stabilization but is not essential for PS activity [18]. Mutagenesis studies suggest that the conserved motif DYLSF at the C terminus of PEN-2 (residues 90–94) is important Pen-2 binding to presenilin and other proteins in GSC complex [36]. Sequence alignment identified this conserved Pen-2 motif in Sft2/Got-1 protein, the DYXXF is in extracellular loop, equally topology studies demonstrated that C-terminal of Pen-2 containing this motif mediate interactions with the large ectodomain of Nct and the extracellular loops of both PS and APH-1 [37].
Several studies show that the GSC of human cells is located preferentially in the endoplasmic reticulum and Golgi complex, as well as in other membrane compartments, such as in mitochondria, cell membrane, nuclear envelope and lysosomes [38,39,40,41]. This location is consistent with the activities assigned to the complex, both processing transmembrane proteins as part of the secretory pathway [42]. In the promastigote forms of Leishmania sp, as well as in other trypanosomatids, the secretory pathway involves the reticulum endoplasmic membrane, the Golgi complex and the flagellar pocket region, these being last two very close structures [42]. Equally close is the flagellar pocket and basal body in the kinetoplast, a characteristic region of trypanosomatid mitochondria rich in kDNA. The cellular localization assays of PS-like and Got-1 proteins from L. infantum using anti-PS serum and GFP-labeled Got-1 by confocal microscopy clearly showed that the two proteins are superimposed, however, they do not overlap punctually in the region of the flagellar pocket. Therefore, these results showed for the first time that the two proteins could be located in the same cellular compartment. This location is similar to that found in T. cruzi [24] and suggests the involvement of the complex in processes of the secretory pathway of this organism. Another interesting point was the demonstration that the Got-1-like protein is also present in this structure, in a PS-like complexed way. In fact, presenilin interaction with vesicular protein synaptotagmin 1 (Syt1) can modulate intracellular trafficking and facilitate exocytosis [43].
Vesicle transport and trafficking is essential for Leishmania differentiation, growth, and parasite-host interplay. Vesicle trafficking in leishmania is related to exossomes/ extracellular vesicles release with virulence factors that contribute to parasite infection and immune evasion [44]. Proteomic data revealed that many transport proteins are upregulated during parasite life cycle and in miltefosine resistant parasites [45]. Vesicle fusion to membrane is an essential mechanism for exocytosis and endocytosis through flagellar pocket, an important family of proteins involved in this process are SNAREs. Proteomic analysis revealed that L. major present more than 20 SNARE proteins, these proteins were located in the Golgi, late endosome/lysosome and near the flagellar pocket [20]. Got1/Stf2 family represent proteins that facilitate fusion events of Sed5, a member of the syntaxin family of SNARE proteins [23]. In Trypanosoma brucei Got1/Stf2 like protein (Tb927. 11.10980) were upregulated in bloodstream form, and evidence suggest this protein is essential and contribute to gain of fitness [46]. A deeper look at composition of L. infantum and L. amazonensis membrane showed the expression of Got1/Stf2 like protein in L. amazonensis [47]. Leishmania proteins involved in intracellular trafficking is not yet characterized. However, recent reports emerging indicate in trypanosomatids that some Rabs and SNAREs are conserved [48].
As mentioned before, in Leishmania sp the mechanisms of secretion involve a network of endomembrane vesicles, the endoplasmic reticulum, the Golgi apparatus and the region of the flagellar pocket, these last two organelles being found one next to the other. Next to these is the kinetoplast, a portion of the mitochondria of kDNA-rich trypanosomatids. The process of macroautophagy is crucial for the differentiation of procyclic promastigotes and metacyclics, in exponential or stationary growth phase respectively [49], as well as in the transition between promastigote and amastigote forms [49]. Autophagy is important in the degradation of proteins and organelles during cell differentiation and also as a defense mechanism in times of stress, but it can also be induced by pharmacological treatment [50]. There are hypotheses of the presence of a peculiar mechanism of cell death in Leishmania [51] and that the dying cell exhibits increased autophagy. Human PS-1 has been described as an important element in the maturation of the autophagosome and fusion with the lysosome [52]. Like the PS-like of L. infantum shares important properties with the human ortholog, perhaps it can also play a possible role in the macroautophagy process. Thus, there is a possibility of Presenilin from Leishmania sp and other trypanosomatids being involved in cell differentiation processes. The study and characterization of flagellar pocket proteins and trafficking proteins can improve the knowledge of physiological, biochemical and parasite-host interplay, providing possible novel targets for therapeutic strategies.
4. Materials and Methods
4.1. Parasites
Promastigote forms of L. infantum were maintained in vitro at 26° C in BHI medium (Bovine Brain and Heart Infusion; Oxoid, São Paulo, Brazil), pH 7.5, supplemented with 10% inactivated fetal bovine serum (FBS) and antibiotics penicillin/streptomycin at 20 mg/mL (Sigma-Aldrich, Saint Louis MO, USA), according to protocol standardized in our laboratory. For transfection assays, the parasites were maintained in synthetic M199 medium (Gibco, Waltham, Massachusetts, USA) supplemented with 1 mM HEPES, 10 mM adenine, 5000u/ml penicillin, 5 mg/ml streptomycin, 0.25% hemin, 2 mM L-glutamine, 6 mM NaHCO3 supplemented with 10% FBS. For the maintenance of mutant parasites, antibiotic G418 (Sigma, St Louis, MO, USA) was used at varying concentrations and 10 mL of culture medium.
4.2. DNA extraction and oligonucleotide synthesis
Approximately 7x107 parasites in the promastigote form, obtained in the log phase of growth, were washed three times with PBS (8 mM Na2PO4, 1.5 mM KH2PO4, 140 mM NaCl and 2.7 mM KCl) by centrifugation at 3000 X g for 15 min at 4°C. DNA extraction was performed using the commercial kit Wizard® Genomic DNA Purification Kit (Promega, Madison, Wisconsin, USA), as instructed by the manufacturer. Acquired SFT2/Got-1-specific oligonucleotide primers of L. infantum from Integrated DNA Technologies (IDT; Coralville, Iowa, USA) from the sequences available in the SwissProt protein database. The SciTools tool (IDT, Coralville, Iowa, USA). The sequences designed for the Got1-like gene were named Got 1(F) (5`-AGATCTATGGATAAGCTTACCTCCAT CA TC-3), containing the site of restriction enzyme BglII (underlined) and Got1 (R) (5`-GGT ACCGCGGTTCATGAGACG-3) containing KpnI restriction enzyme site (underli ned). The insertion of restriction sites was programmed and added in the sequence of protein oligonucleotides as a facilitator for the strategy in the later step of subcloning in the expression vector.
4.3. Amplification and cloning
The PCR fragments corresponding to the Got-1-like gene were amplified from the genomic DNA of L. infantum using the GoTaq® Hot Start Polymerase (Promega, Madison, Wisconsin, USA) and the Got1(F)/Got1(R) primers, as recommended by the manufacturer. The PCR products were purified using the Wizard SV Gel and PCR Clean-Up kit (Promega, Madison, Wisconsin, USA) following the manufacturer's instructions and then ligated into the pGEM-T cloning vector easy vector (Thermo-Fischer, Waltham, MA, USA) using the TA strategy. Therm shock protocol transformed the ligation into chemically competent bacteria (E. coli DH5α-TOP10). After this step, 100 μL of the transformation were plated on Luria-Bertani (LB)-Tryptone-Agar 1% (85mM NaCl, yeast – 0.5%, 15% Agar containing 100 μg ampicillin/ ml). Plasmid DNA from bacteria containing the clones was purified using the PureYield Plasmid Miniprep System Kit (Thermo-Fischer, Waltham, MA, USA), and SANGER sequenced. The plasmid DNAs were then digested with the BglII and KpnI enzymes (New England Biolabs, Massachusetts, USA) following the manufacturer's guidelines, and digestions were resolved in 0.8% agarose gel for the PS-like and 1.5% for the Got1-like gene. The bands referring to the inserts were purified using the Wizard SV Gel Kit and PCR Clean-Up (Promega, Madison, Wisconsin, USA). pNUS-GFP (Green Fluorescent Protein) (Figure 3) expression vector was used and digested with Bglll and KpnI enzymes, as they were designed to contain, among others, the same sites as the restriction enzymes operated. They were later purified using the as-me kit. The inserts were ligated to the plasmids through the strategy of cohesive ends, using the enzyme T4 DNA Ligase (Promega, Madison, Wisconsin, USA). Heat shock transformed the binding into chemically competent E. coli DH5α-TOP10. The plasmid DNA of the bacteria containing the clones was purified using the kit Plasmid Maxi Prep (QIAGEN, Germantown, MD, USA). The generation of mutant protozoa was performed by transfection using the high-voltage electroporation method [53]. Briefly, three tubes containing 5x106 wild-type promastigote cells (L. infantum– IOC-579) were washed by centrifugation three times in an M199 medium and resuspended in 100 μL of electroporation solution from the Were analyzed under three conditions: MOCK [without plasmid and Human T Cell NucleofectorTM kit (Lonza Bioscience, Morrisville, USA)] undergoing electroporation), WT (without plasmid and without undergoing electroporation), and TEST (with plasmid and undergoing electroporation); 0.2 cm electrodes were used in Amaxa Nucleofector II electroporator (Lonza Bioscience, Morrisville, USA) and the U-033 program preset in the equipment. The parasites were then transferred to 10 mL of medium supplemented with 10% FBS and incubated at 26°C for 24h without a selection antibiotic. The parasites were collected by centrifugation and resuspended in 100 µL of freshly prepared M199 culture medium with 10% FBS and inoculated into 25 cm2 bottles containing 5 ml of M199 medium and selection antibiotics G418 at 100 mg/mL.
4.4. Polyclonal antibodies anti-presenilin like protein
A peptide sequence located in a loop between the domain TM2 and TM3 (VLLYHFHLQFVLYG) of L. amazonensis PS protein was chosen to obtain polyclonal antibodies (Figure S4). This epitope was identified by microarray of peptides using patient’s sera and an Auto-Spot Robot ASP-222 (Intavis Bioanalytical Instruments AG, Köln, Germany) [41]. A multi antigen peptide (MAP4) was produced using the F-moc strategy in a synthesizer machine (MultiPep-1, CEM Corp, Charlotte, NC, USA) and used to immunize a rabbit using previously defined protocol [24]. The specificity of the rabbit sera anti-PS was checked by dot blot (Figure S5).
4.5. Fluorescence microscopy
Parasites 5x104 were washed three times with PBS and fixed with 2% paraformaldehyde solution in PBS. Then, they were deposited on microscopy slides, allowed to dry (16 h at 25° C), and stored at -20°C. For the immunological reaction, the slides were thawed at room temperature for 5 min and washed with PBS containing 0.05% Tween 20 (PBS-T) under gentle horizontal agitation for 5 min. Next, the slides were incubated in a humid chamber (90 min at 37°C) with rabbit serum anti-PS (1:150 prepared in PBS-T containing 1% skim milk). After three washes with PBS-T for 10 min, the slides were incubated for 30 min at 37oC in the same humid chamber with goat antibody anti-rabbit IgG-Alexa fluor 546 labeled (1:500; prepared in T-PBS containing 1% skim milk). Finally, a new cycle of five washes in PBS-T of 10 min was performed, and after the last washing, slides were mounted with 90% glycerol in PBS and analyzed in a confocal fluorescence microscope LSM-710 (AxioCam; Carls-Zeiss Microscopy GmbH, Jena, Germany). Captured images with ZEN Black software (Carls-Zeiss).
4.6. Phylogenetic analysis
The L. infantum SFT2/Got-1 sequence (A4HSK3) was prospected in Blast and accessing InParanoid (https://inparanoid.sbc.su.se/cgi-bin/index.cgi), followed by multiple sequence alignment using ClustalW. Then, inferred the evolutionary history using the Maximum Likelihood method based on the JTT matrix-based model. The initial tree(s) for the heuristic search were obtained automatically by applying Neighbor Join algorithms to a matrix of pairwise distances estimated using a JTT model and then selecting the topology with a superior log-likelihood value. Finally, evolutionary analyses were conducted in MEGA7 [54].
4.7. Bioinformatic analysis
Retrieved the search for the sequence of the Got-1 (XP_001463044.1) from GenBank. Performed the databank search for L. infantum sequence (A4HSK3) in UniProt (http://www.uniprot.org/, Accessed on 21 August 2020). The prediction of the secondary structure of the protein was performed by the PSIPRED servers (http://bioinf.cs.ucl.ac.uk//, Accessed on 10 February 2021), and TMD visualization was achieved using the Protter platform (https://wlab.ethz.ch/protter/). The tertiary structure prediction and the transmembrane domain by Alphafold (https://alphafold.ebi.ac.uk/; Accessed on 15 November 2021). Obtained the protein transmembrane orientation by inserting protein structure in the direction of the membrane protein database (OMP, (https://opm.phar. umich. edu/, Accessed on 12 November 2021) using the PPM server [55].
5. Conclusions
The Got-1 protein identified in L. infantum presents a conserved motif in human Pen-2. However, due to its drastic structural differences, it is difficult to think that it can perform the same functions attributed to human Pen-2 of helping to function of presenilins. Regardless, our data show the presence of an important protein transported close to or co-located to the gamma-secretase complex and which, in some yet unknown way, can act synergistically or concomitant in the processing and transport of proteins or perform functions not yet known. On the other hand, the location of Got-1 in the flagellar pocket, where there is an intense metabolic transit and extremely important for the parasite's nutrition, suggests that this protein represents an important alternative to be evaluated as a new therapeutic target.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization and Investigation: S.G.D.-S.; Methodology: L.A.T-P; N.-P., and G.C.L.; Funding: S.G.D-S. and C.M.M.. Computational analysis: L.A.T-P and G.C.L.; Writing— original draft: L.A.T-P.; Review and editing: S.G.D.-S and G.C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Brazilian Council for Scientific Research (CNPq #30515- 2020-5) and Carlos Chagas Filho Foundation of Research Support of the State of Rio de Janeiro (FAPERJ #200.960-2022). Funding was also provided by FAPERJ (#210.003/2018) through the National Institutes of Science and Technology Program (INCT) to C.M.M. (INCT-IDPN).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
P.N-P and G.C.L are CAPES/FIOCRUZ PostDoc fellow.
Conflicts of Interest
The authors declare no conflict of interest
References
- WHO. Leishmaniasis. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (Accessed on 20 June 2022).
- Akpunarlieva, S.; Burchmore, R. The role of membrane transporters in Leishmania virulence. Emerg Top Life Sci 2017, 1, 601-611. [CrossRef]
- Field, M.C.; Natesan, S.K.; Gabernet-Castello, C.; Koumandou, V.L. Intracellular trafficking in the trypanosomatids. Traffic 2007, 8, 629-639. [CrossRef]
- Matte, C.; Descoteaux, A. Exploitation of the host cell membrane fusion machinery by leishmania is part of the infection process. PLoS Pathog. 2016, 12, e1005962. [CrossRef]
- Landfear, S.M.; Ignatushchenko, M. The flagellum and flagellar pocket of trypanosomatids. Mol Biochem Parasitol 2001, 115, 1-17. [CrossRef]
- Machado, P.A.; Carneiro, M.P.D.; Sousa-Batista, A.J.; Lopes, F.J.P.; Lima, A.P.C.A.; Chaves, S.P.; Sodero, A.C.R.; de Matos Guedes, H.L. Leishmanicidal therapy targeted to parasite proteases. Life Sci 2019, 219, 163-181. [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Finn, R. Twenty years of the MEROPS database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res 2016, 44, D343-D350. [CrossRef]
- Yang, G.; Zhou, R.; Guo, X.; Yan, C.; Lei, J.; Shi, Y. Structural basis of γ-secretase inhibition and modulation by small molecule drugs. Cell 2021, 184, 521-533.e14. [CrossRef]
- Welander, H.; Tjernberg, L.O.; Karlström, H.; Ankarcrona, M. γ-Secretase complexes containing caspase-cleaved presenilin-1 increase intracellular Aβ(42) /Aβ(40) ratio. J Cell Mol Med 2011, 15, 2150-63. [CrossRef]
- Leissring, M.A.; Yamasaki, T.R.; Wasco, W.; Buxbaum, J.D.; Parker, I.; LaFerla, F.M. Calsenilin reverses presenilin-mediated enhancement of calcium signaling. Proc Natl Acad Sci USA 2000, 97, 8590-8593. [CrossRef]
- Kim, D.Y.; Ingano, L.A.; Kovacs, D.M. Nectin-1 alpha, an immunoglobulin-like receptor involved in the formation of synapses, is a substrate for presenilin/gamma-secretase-like cleavage. J Biol Chem 2002, 277, 49976-49981. [CrossRef]
- Marambaud, P.; Shioi, J.; Serban, G.; Georgakopoulos, A., Sarner, S.; Nagy, V.; Baki, L.; Wen, P.; Efthimiopoulos, S.; Shao, Z.; Wisniewski, T.; Robakis, N.K. A presenilin-1/gamma-secretase cleavage releases the E-cadherin intracellular domain and regulates disassembly of adherens junctions. EMBO J 2002, 21, 1948-1956. [CrossRef]
- Lammich, S.; Okochi, M.; Takeda, M.; Kaether, C.; Capell, A.; Zimmer, A.K.; Edbauer, D.; Walter, J.; Steiner, H.; Haass, C. Presenilin-dependent intramembrane proteolysis of CD44 leads to the liberation of its intracellular domain and the secretion of an a beta-like peptide. J Biol Chem 2002, 277, 44754-44759. [CrossRef]
- Cai, T.; Tomita, T. Structure-activity relationship of presenilin in γ-secretase-mediated intramembrane cleavage. Semin Cell Dev Biol. 2020, 105, 102-109. [CrossRef]
- Nasamu, A.S.; Polino, A.J.; Istvan, E.S.; Goldberg, D.E. Malaria parasite plasmepsins: More than just plain old degradative pepsins. J Biol Chem 2020, 295, 8425-8441. [CrossRef]
- Johnson, D.S.; Li, Y.M.; Pettersson, M.; St George-Hyslop, P.H. Structural and chemical biology of presenilin complexes. Cold Spring Harb Perspect Med. 2017, 7, a024067. [CrossRef]
- Sarasija, S.; Norman, K.R. A gamma-secretase independent role for presenilin in calcium homeostasis impacts mitochondrial function and morphology in Caenorhabditis elegans. Genetics 2015. 201, 1453-1466. [CrossRef]
- Holmes, O.; Paturi, S.; Selkoe, D.J.; Wolfe, M.S. Pen-2 is essential for γ-secretase complex stability and trafficking but partially dispensable for endoproteolysis. Biochemistry 2014, 53(27):4393-4406. [CrossRef]
- Yoon, T.Y.; Munson, M. SNARE complex assembly and disassembly. Curr Biol. 2018, 28, R397-R401. [CrossRef]
- Besteiro, S.; Coombs, G.H.; Mottram, J.C. The SNARE protein family of Leishmania major. BMC Genomics 2006, 7, 250. [CrossRef]
- Conchon, S.; Cao, X.; Barlowe, C.; Pelham, H.R. Got1p and Sft2p: membrane proteins involved in traffic to the Golgi complex. EMBO J 1999, 18, 3934-3946. [CrossRef]
- Banfield, D.K.; Lewis, M.J.; Pelham, H.R.; A SNARE-like protein required for traffic through the Golgi complex. Nature 1995, 375, 806-809. [CrossRef]
- Suga, K.; Saito, A.; Tomiyama, T.; Mori, H.; Akagawa, K. Syntaxin 5 interacts specifically with presenilin holoproteins and affects processing of betaAPP in neuronal cells. J Neurochem 2005, 94, 425-439. [CrossRef]
- Lechuga, G.C.; Bottino, C.C.G.; Pinho, R.T.; Souza, A.L.A.; Provance-Jr, D.W.; De-Simone, S.G. Trypanosoma cruzi presenilin-like transmembrane aspartyl protease: Characterization and cellular localization. Biomolecules 2020, 10, 1564-1584. [CrossRef]
- Lechuga, G.C.; Napoleão-Pêgo, P.; Gomes, L.R.; Durans, A.M.; Provance-Jr, D.W.; De-Simone, S.G. Nicastrin-like, a novel transmembrane protein from Trypanosoma cruzi associated to the flagellar pocket. Microorganisms 2021, 9, 1750. [CrossRef]
- De-Simone, S.G.; Napoleão-Pêgo, P.; Gonçalves, P.S.; Lechuga, G.C.; Mandonado-Jr, A.; Graeff-Teixeira, C.; Provance-Jr, D.W. Angiostrongylus cantonensis an atypical presenilin: epitope mapping, characterization, and development of an ELISA peptide assay for specific diagnostic of angiostrongyliasis. Membranes 2022, 12, 108. [CrossRef]
- Wolfe, M.S.; Miao, Y. Structure and mechanism of the γ-secretase intramembrane protease complex. Curr Opin Struct Biol. 2022, 74,102373. [CrossRef]
- Zhao, G.; Cui, M.Z.; Mao, G.; Dong, Y.; Tan, J.; Sun, L.; Xu, X. gamma-Cleavage is dependent on zeta-cleavage during the proteolytic processing of amyloid precursor protein within its transmembrane domain. J Biol Chem 2005, 280, 37689-37697. [CrossRef]
- Wolfe, M.S. Structure and function of the γ-secretase complex. Biochemistry 2019, 58, 2953-2966. [CrossRef]
- Chen, F.; Hasegawa, H.; Schmitt-Ulms, G.; Kawarai, T.; Bohm, C.; Katayama, T.; Gu, Y.; Sanjo, N.; Glista, M.; Rogaeva, E.; et al.. TMP21 is a presenilin complex component that modulates gamma-secretase but not epsilon-secretase activity. Nature 2006, 440, 1208-1212. [CrossRef]
- Wolfe, M.S. Probing mechanisms and therapeutic potential of γ-secretase in Alzheimer's disease. Molecules 2021, 26, 388. [CrossRef]
- Svedružić, Ž.M.; Vrbnjak, K.; Martinović, M.; Miletić, V. Structural Analysis of the simultaneous activation and inhibition of γ-secretase activity in the development of drugs for Alzheimer's disease. Pharmaceutics 2021, 13, 514. [CrossRef]
- Marambaud, P.; Robakis, N.K. Genetic and molecular aspects of Alzheimer's disease shed light on new mechanisms of transcriptional regulation. Genes Brain Behav, 2005, 4, 134-146. [CrossRef]
- Besteiro, S.; Williams, R.A.; Coombs, G.H.; Mottram, J.C. Protein turnover and differentiation in Leishmania. Int J Parasitol 2007, 37, 1063-1075. [CrossRef]
- Arango Duque, G.; Jardim, A.; Gagnon, É.; Fukuda, M.; Descoteaux, A. The host cell secretory pathway mediates the export of Leishmania virulence factors out of the parasitophorous vacuole. PLoS Pathog 2019, 15, e1007982. [CrossRef]
- Hasegawa, H.; Sanjo, N.; Chen, F.; Gu, Y.J.; S,hier, C.; Petit, A.; Kawarai, T.; Katayama, T.; Schmidt, S.D.; Mathews, P.M. et al.. Both the sequence and length of the C terminus of PEN-2 are critical for intermolecular interactions and function of presenilin complexes. J Biol Chem 2004, 279, 46455-46463. [CrossRef]
- Crystal, A.S.; Morais, V.A.; Pierson, T.C.; Pijak, D.S.; Carlin, D.; Lee, V.M.; Doms, R.W. Membrane topology of gamma-secretase component PEN-2. J Biol Chem 2003, 278, 20117-20123. [CrossRef]
- Wheeler, R.J.; Sunter, J.D.; Gull, K. Flagellar pocket restructuring through the Leishmania life cycle involves a discrete flagellum attachment zone. J Cell Sci 2016, 129, 854-867. [CrossRef]
- Brickman, M.J.; Balber, A.E.Trypanosoma brucei rhodesiense: membrane glycoproteins localized primarily in endosomes and lysosomes of bloodstream forms. Exp Parasitol 1993, 76, 329-344. [CrossRef]
- Müller, M.T.; Schempp, R.; Lutz, A.; Felder, T.; Felder, E.; Miklavc, P. Interaction of microtubules and actin during the post-fusion phase of exocytosis. Sci Rep 2019, 9, 11973. [CrossRef]
- McConville, M.J.; Mullin, K.A.; Ilgoutz, S.C.; Teasdale, R.D. Secretory pathway of trypanosomatid parasites. Microbiol Mol Biol Rev 2002, 66, 122-54. [CrossRef]
- Clayton, C.; Hausler,T.; Blattner, J. Protein trafficking in kinetoplastid protozoa. Microbiol Rev 1995, 59, 325-344. [CrossRef]
- Zoltowska, K.M.; Maesako, M.; Lushnikova, I; Takeda, S.; Keller, L.J.; Skibo, G.; Hyman, B.T.; Berezovska, O. Dynamic presenilin 1 and synaptotagmin 1 interaction modulates exocytosis and amyloid β production. Mol Neurodegener 2017, 12, 15. [CrossRef]
- Arango Duque, G.; Jardim, A.; Gagnon, É.; Fukuda, M.; Descoteaux, A. The host cell secretory pathway mediates the export of Leishmania virulence factors out of the parasitophorous vacuole. PLoS Pathog 2019, 15, e1007982. [CrossRef]
- Saboia-Vahia, L.; Cuervo, P.; Wiśniewski, J.R; Dias-Lopes, G.; Pinho, N.; Padrón, G.; de Pilla Varotti, F.; Murta, S.M.F. In-depth quantitative proteomics characterization of in vitro selected miltefosine resistance in Leishmania infantum. Proteomes 2022, 10,10. [CrossRef]
- Alsford, S.; Turner, D.J.; Obado, S.O.; Sanchez-Flores, A.; Glover, L.; Berriman, M.; Hertz-Fowler, C.; Horn, D. High-throughput phenotyping using parallel sequencing of RNA interference targets in the African trypanosome. Genome Res 2011, 21, 915-924. [CrossRef]
- Oliveira, I.H.R.; Figueiredo, H.C.P.; Rezende, C.P.; Verano-Braga, T.; Melo-Braga, M.N.; Reis Cunha, J.L.; de Andrade, H.M. Assessing the composition of the plasma membrane of Leishmania (Leishmania) infantum and L. (L.) amazonensis using label-free proteomics. Exp Parasitol 2020, 218, 107964. [CrossRef]
- Venkatesh, D.; Boehm, C.; Barlow, L.D.; Nankissoor, N.N.; O'Reilly, A.; Kelly, S.; Dacks, J.B.; Field, M.C. Evolution of the endomembrane systems of trypanosomatids-conservation and specialisation. J Cell Sci 2017, 130, 1421-1434. [CrossRef]
- Gossage, S.M.; Roger, M.E.; Bates, P.A. Two separate growth phases during the development of Leishmania in sand flies: implications for understanding the life cycle. Int J Parasitol 2003, 33, 1027-1034. [CrossRef]
- Mesquita-Rodrigues, C.; Menna-Barreto, R.F.; Sabóia-Vahia, L.; Da-Silva, S.A.; de Souza, E.M.; Waghabi, M.C.; Cuervo, P.; De Jesus, J.B. Cellular growth and mitochondrial ultrastructure of Leishmania (Viannia) braziliensis promastigotes are affected by the iron chelator 2,2-dipyridyl. PLoS Negl Trop Dis 2013, 7, e2481. [CrossRef]
- Taylor-Brown, E.; Hurd, H. The first suicides: a legacy inherited by parasitic protozoans from prokaryote ancestors. Parasit Vectors. 2013, 6, 108. [CrossRef]
- Sharma, B.; Pal, D.; Sharma, U.; Kumar, A. Mitophagy: An emergence of new player in Alzheimer's disease. Front Mol Neurosci. 2022,15, 921908. [CrossRef]
- Planelles, L.; Marañón, C.; Requena, J.M.; López, M.C. Phage recovery by electroporation of naked DNA into host cells avoids the use of packaging extracts. Anal Biochem 1999, 267, 234-235. [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for Bigger Datasets. Mol Biol Evol 2016, 33, 1870–1874. [CrossRef]
- Lomize, M.A.; Pogozheva, I.D.; Joo, H.; Mosberg, H.I.; Lomize, A.L. OPM database and PPM web server: Resources for positioning of proteins in membranes. Nucleic Acids Res 2011, 40, D370–D376. [CrossRef]
Figure 1.
Analysis by agarose gel electrophoresis (0.8%) of the genes of the colonies after ligation with pGEM-T easy vector. Got-1-like gene have a molecular weight of 492bp.
Figure 1.
Analysis by agarose gel electrophoresis (0.8%) of the genes of the colonies after ligation with pGEM-T easy vector. Got-1-like gene have a molecular weight of 492bp.

Figure 2.
Analysis of the subcloning of the Got1-like gene + pNUS-GFP in agarose gel. 1-1Kb DNA ladder; 2 - Purification of the gene-fused pNUS-GPF plasmid Got1-like; 3 - PCR of the purification of the pNUS-GPF plasmid fused to the Got1-like gene with primers targeting the Got1-like gene; 4 - Negative control of the PCR reaction (Water DNAse free); 5 - Positive control of the PCR reaction (genomic DNA).
Figure 2.
Analysis of the subcloning of the Got1-like gene + pNUS-GFP in agarose gel. 1-1Kb DNA ladder; 2 - Purification of the gene-fused pNUS-GPF plasmid Got1-like; 3 - PCR of the purification of the pNUS-GPF plasmid fused to the Got1-like gene with primers targeting the Got1-like gene; 4 - Negative control of the PCR reaction (Water DNAse free); 5 - Positive control of the PCR reaction (genomic DNA).
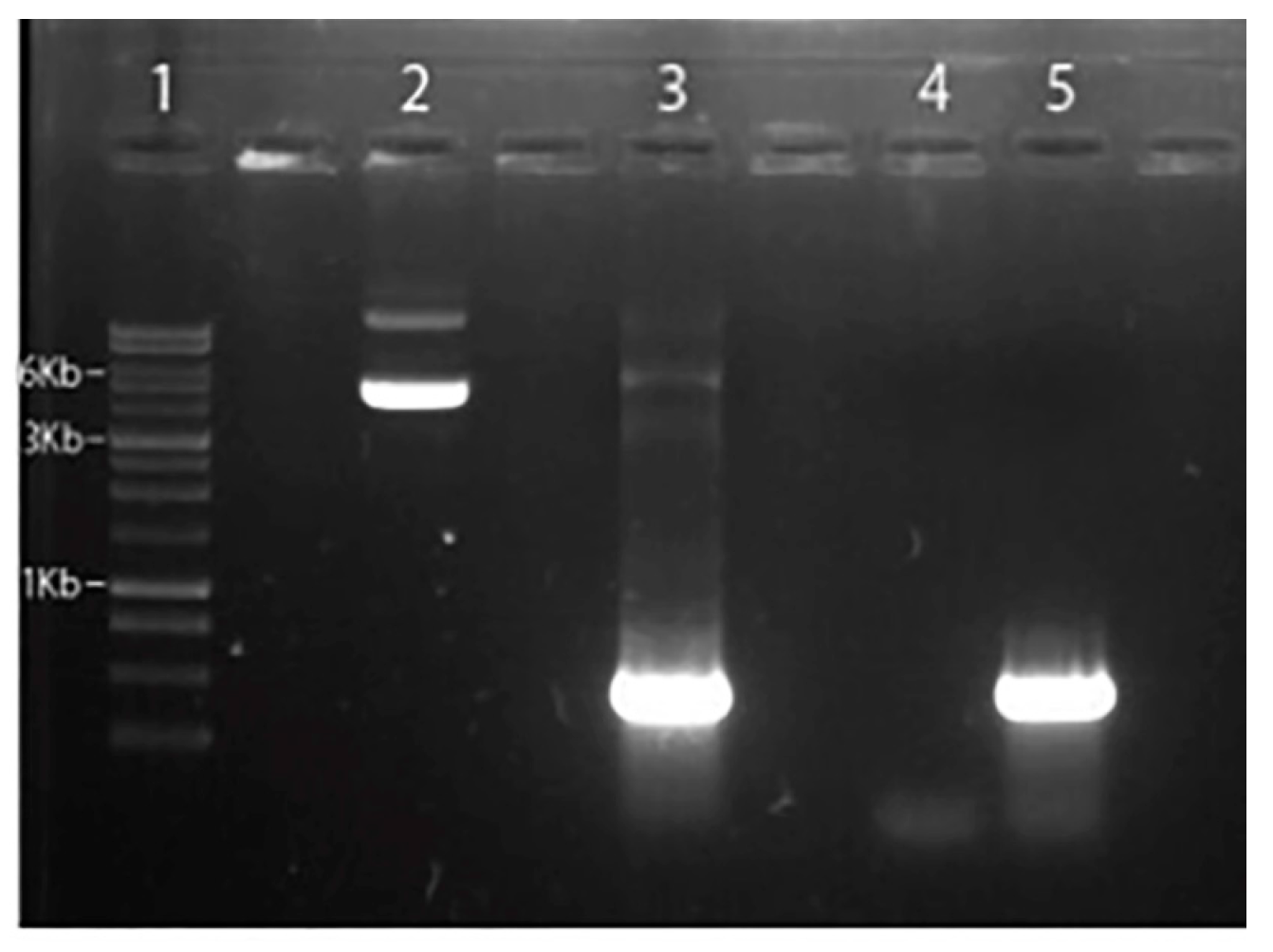
Figure 3.
Immunofluorescence subcellular localization of Got-1/Sft2 like protein and presenilin-like of L. infantumi. The expression of Got1/Sft2-recombinant linked to GFP in promastigotes is shown in green and the immunolabeled with anti-PS polyclonal rabbit serum labeled in red (Alexa 546). The images were superimposed. Dic, differential interference contrast microscopy.
Figure 3.
Immunofluorescence subcellular localization of Got-1/Sft2 like protein and presenilin-like of L. infantumi. The expression of Got1/Sft2-recombinant linked to GFP in promastigotes is shown in green and the immunolabeled with anti-PS polyclonal rabbit serum labeled in red (Alexa 546). The images were superimposed. Dic, differential interference contrast microscopy.
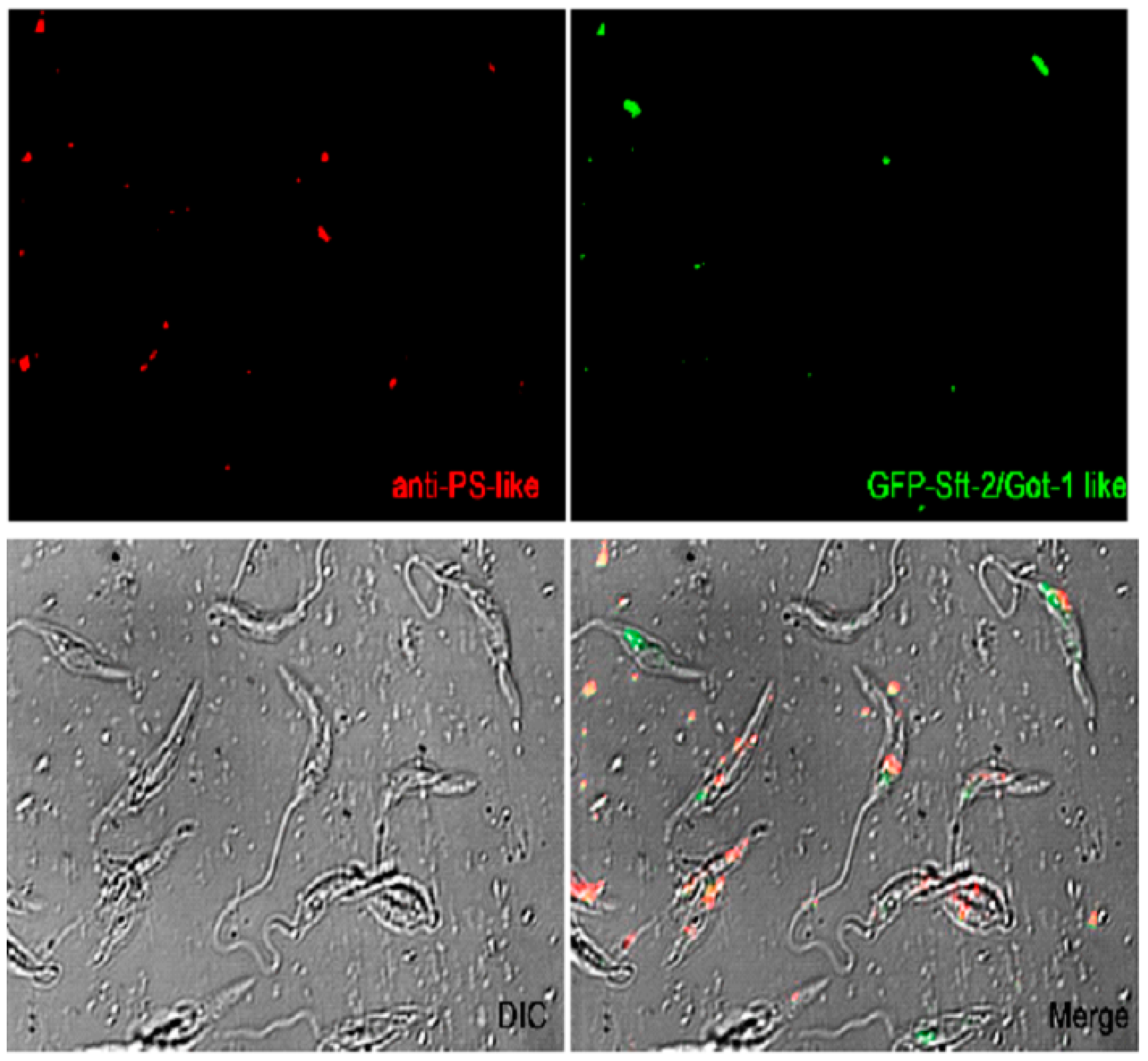
Figure 4.
Model of L. infantum Got1/Sft2 like (A4HSK3) protein with transmembrane domain analysis. (A) Membrane-spanning model of Got1/Sft2-like protein with the four TM conserved domains (aa38 –aa147). (B) Protein orientation on the lipid bilayer was performed and showed the four alpha helix transmembrane conserved domains and site of membrane interaction with embedded residues, determined using the Orientation Membrane Protein (https://opm.phar.umich. edu/; Accessed on 1 October 2021).
Figure 4.
Model of L. infantum Got1/Sft2 like (A4HSK3) protein with transmembrane domain analysis. (A) Membrane-spanning model of Got1/Sft2-like protein with the four TM conserved domains (aa38 –aa147). (B) Protein orientation on the lipid bilayer was performed and showed the four alpha helix transmembrane conserved domains and site of membrane interaction with embedded residues, determined using the Orientation Membrane Protein (https://opm.phar.umich. edu/; Accessed on 1 October 2021).
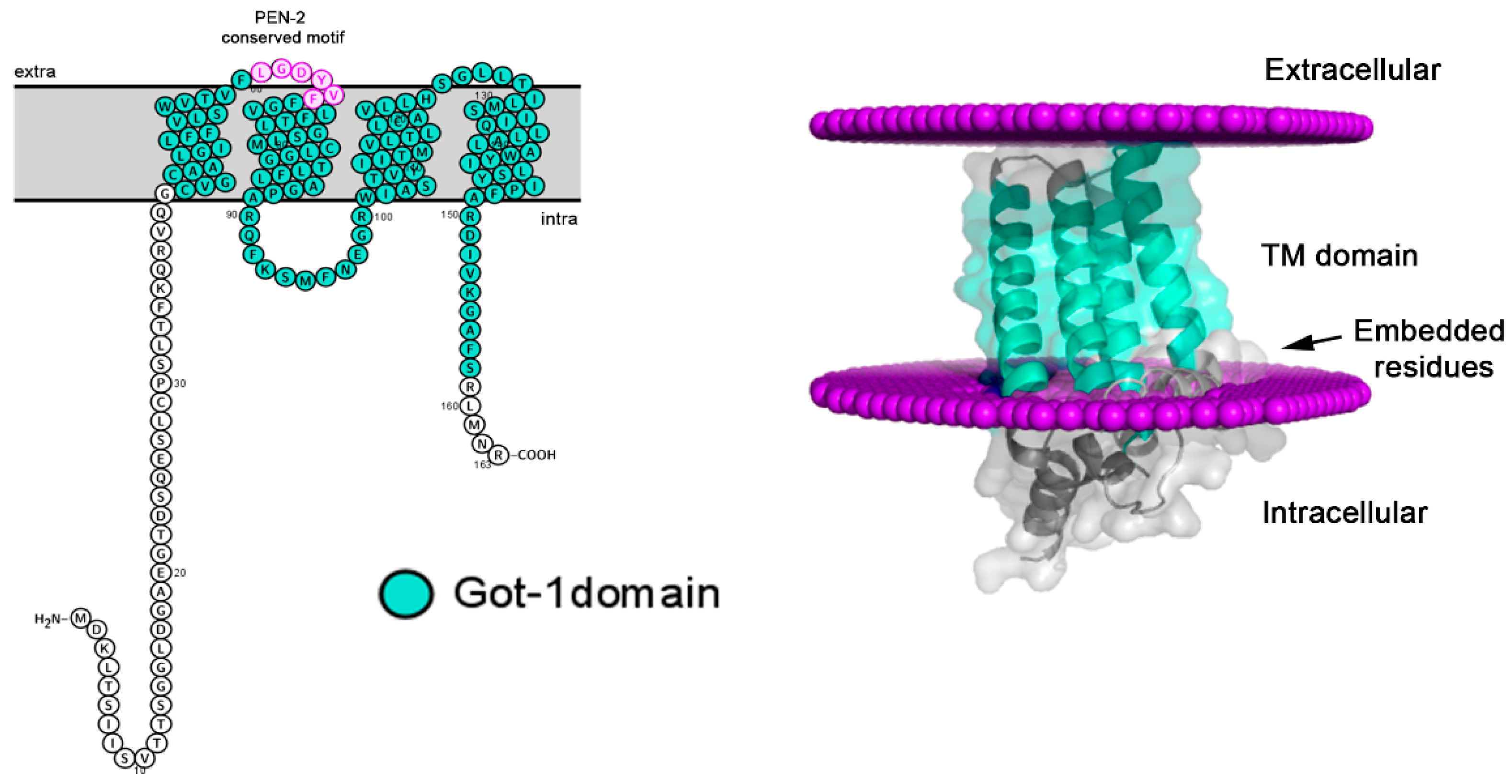
Figure 5.
Phylogenetic relationship of L. infantum Sft2/Got-1 like protein (A4HSK3). The evolutionary history was inferred by using the Maximum Likelihood method based on the JTT matrix-based model [27]. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using a JTT model, and then selecting the topology with superior log likelihood value. Evolutionary analyses were conducted in MEGA7.
Figure 5.
Phylogenetic relationship of L. infantum Sft2/Got-1 like protein (A4HSK3). The evolutionary history was inferred by using the Maximum Likelihood method based on the JTT matrix-based model [27]. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using a JTT model, and then selecting the topology with superior log likelihood value. Evolutionary analyses were conducted in MEGA7.
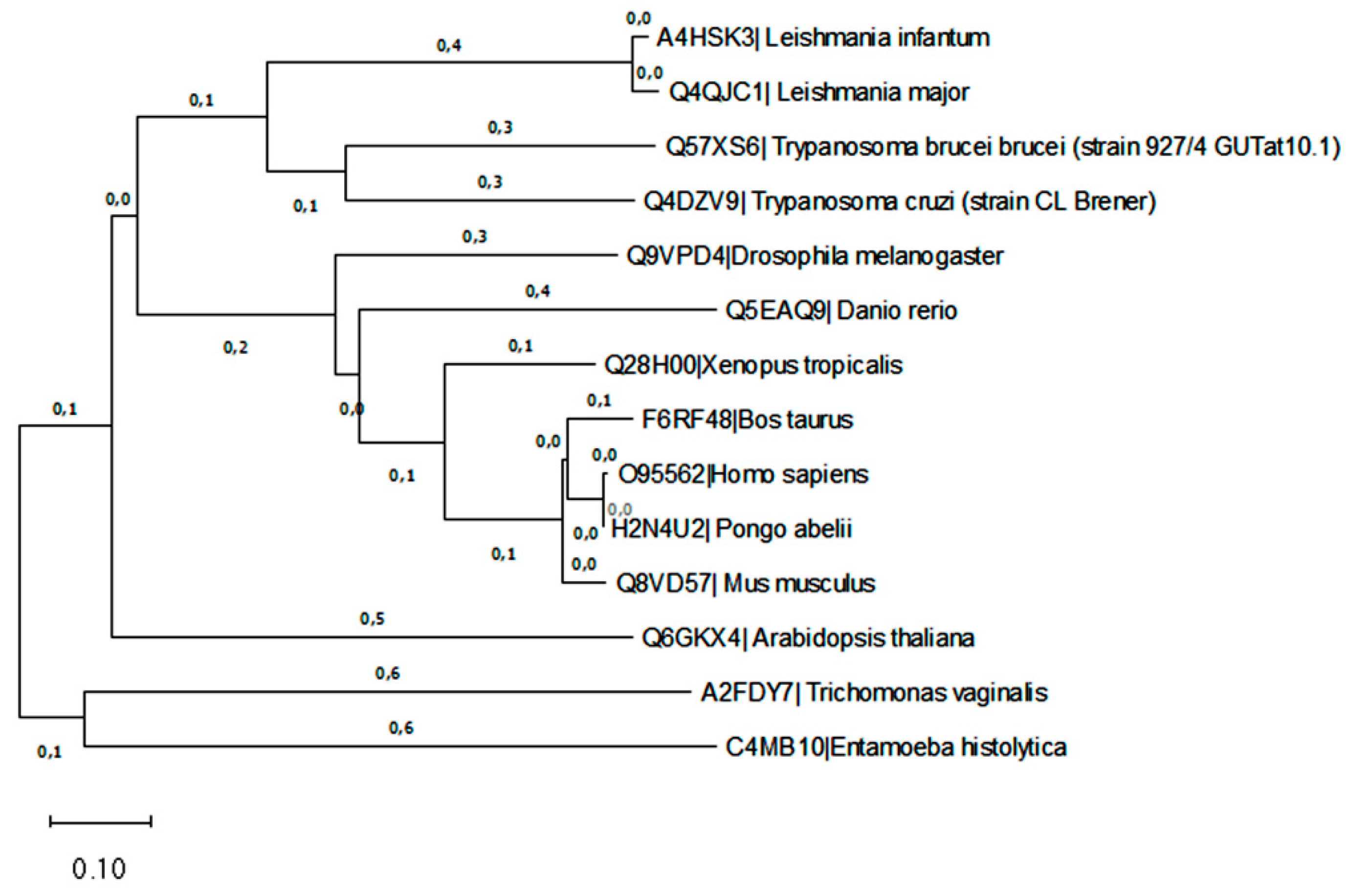

Table 1.
Homologous proteins of L. infantum annotated in the Uniprot database (http://www. uniprot.org/) as Presenilin, NIC, APH1 and PEN2.
Table 1.
Homologous proteins of L. infantum annotated in the Uniprot database (http://www. uniprot.org/) as Presenilin, NIC, APH1 and PEN2.
| Code (UNIPROT) | Human protein | L. infantum protein | Identity (%) | L. infantum code |
| P49768 | Presenilin-1 | - | - | - |
| P49810 | Presenilin-2 | Putative presenilin-like aspartic peptidase | 31 | A4HWP2 |
| Q92542 | Nicastrin | - | - | - |
| Q96BI3/Q8WW43 | Aph1-a/b | APH-1 family | No significant | A4HV58 |
| - | - | - | ||
| Q9NZ42 | Pen-2 | Putative Sft2/Got-1 like family | 35 | A4HSK3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



