
Submitted:
30 December 2022
Posted:
05 January 2023
You are already at the latest version
Abstract
The selectivity in selective autophagy pathways is achieved via the selective autophagy receptors (SARs) – proteins that bind a ligand on the substrate to be degraded and the Atg8-family protein on the growing autophagic membrane, phagophore, effectively bridging them. In mammals, the most common ligand of SARs is ubiquitin, a small protein modifier that tags substrates for their preferential degradation by autophagy. Consequently, the most common SARs are the ubiquitin-binding SARs, such as SQSTM1/p62 (sequestosome 1). Surprisingly, there is only one SAR of this type in yeast – Cue5, which acts as a receptor for aggrephagy and proteaphagy – pathways that remove the ubiquitinated protein aggregates and proteasomes, respectively. However, recent studies described the ubiquitin-dependent autophagic pathways that do not require Cue5, e.g. stationary phase lipophagy for intracellular lipid droplets and nitrogen starvation-induced mitophagy for mitochondria. What is the role of ubiquitin in these pathways? Here, we propose that the ubiquitinated lipid droplets and mitochondria are recognized by the alternative ubiquitin-binding SARs. Our analysis identifies the proteins that could potentially fulfill this role in yeast. We think that matching of the ubiquitin-dependent (but Cue5-independent) autophagic pathways with the ubiquitin-and-Atg8-binding proteins enlisted here might uncover the novel ubiquitin-binding SARs in yeast.
Keywords:
Atg8
; autophagic receptor
; Cue5
; SAR
; selective autophagy
; selective autophagy receptor
; ubiquitin
; ubiquitin-binding domain
; ubiquitin-binding protein
; ubiquitin-binding receptor
1. Introduction
Tagging of substrates for degradation by the selective autophagy pathways can proceed either without or with ubiquitin. The ubiquitin-independent cargo recognition is more prevalent in yeast and can be exemplified by the first autophagic receptor that was identified, Atg19 (autophagy related 19) [1,2]. This protein binds Ape1 (aminopeptidase 1) in cytoplasm and ensures its selective delivery to the vacuole via the cytoplasm-to-vacuole targeting pathway or via (otherwise) non-selective autophagy. In contrast, the ubiquitin-dependent recognition of substrates is more widespread in higher eukaryotes and can be represented by the first mammalian autophagic receptor, SQSTM1/p62 (sequestosome 1) [3,4]. This receptor binds a protein modifier, ubiquitin, promotes aggregation of the ubiquitinated proteins and their targeting to the lysosome via aggrephagy, the selective autophagy of protein aggregates. Since the ubiquitin-binding SQSTM1-like receptors can recognize a variety of ubiquitinated proteins at a surface of different structures, their role in selective autophagy is much broader than that of the Atg19-like receptors and involves simultaneous functioning in several autophagic pathways, including (since recently) lipophagy, the selective autophagy of lipid droplets (LDs) [5,6]. However, the two types of receptors share a common step – bridging of the non-ubiquitinated or ubiquitinated substrate with the Atg8-family protein that lines up a membrane of phagophore, the autophagic transport vesicle (autophagosome) precursor. For example, Atg19 bridges Ape1 with Atg8 in yeast [7,8] and SQSTM1 brings together the ubiquitinated protein aggregates and MAP1LC3B (microtubule associated protein 1 light chain 3 beta) in mammalian cells [3,4].
2. Cue5, the SQSTM1-like autophagic receptor in yeast
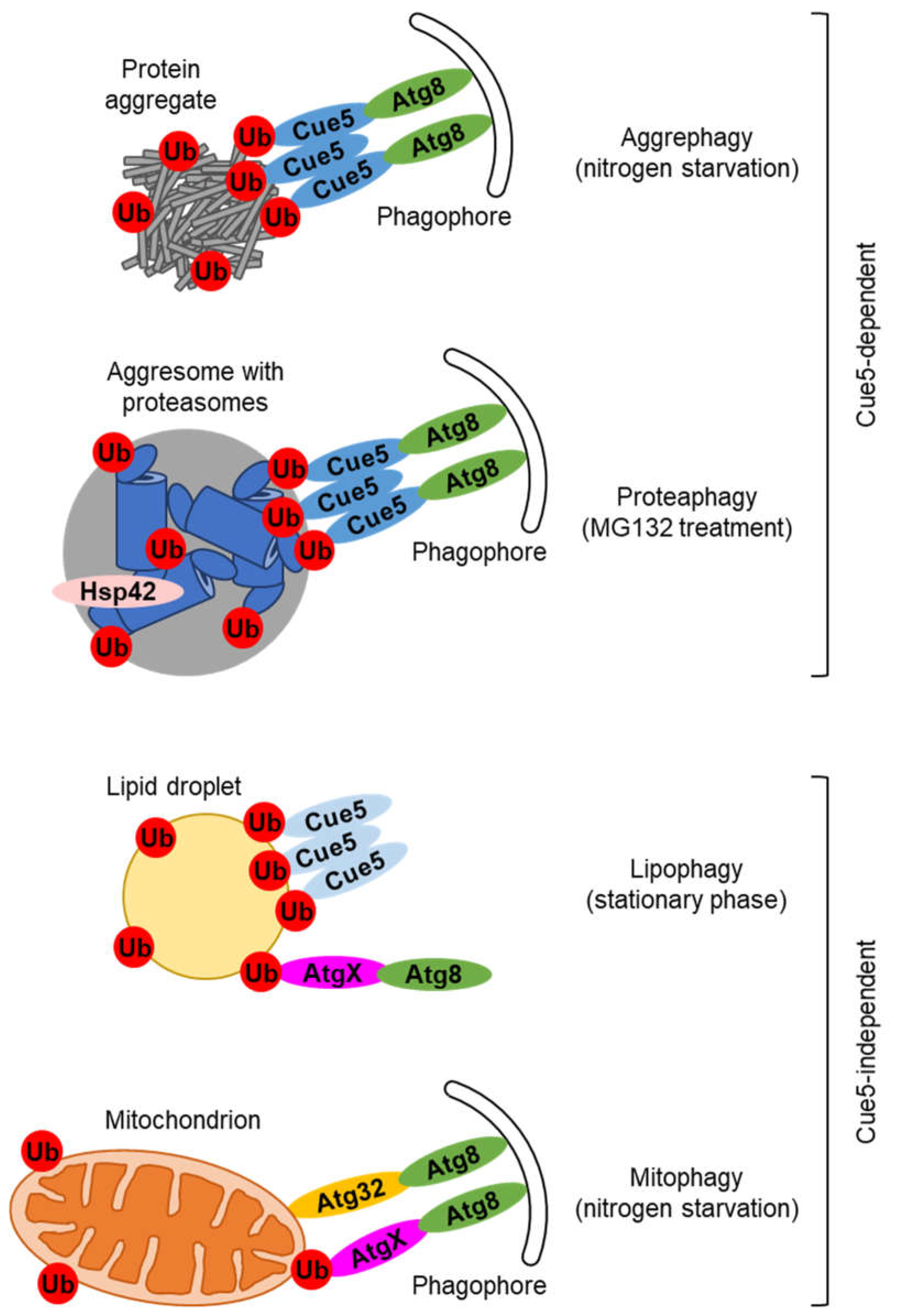
At present, Cue5 is the only known ubiquitin-binding autophagic receptor in yeast. It drives two autophagic pathways, aggrephagy and proteaphagy (selective autophagy of proteasomes) (Figure 1) [9,10]. In aggrephagy, it satisfies all the requirements of the selective autophagy receptor (SAR). Specifically, the Saccharomyces cerevisiae Cue5: 1) binds ubiquitinated proteins via its ubiquitin-binding “coupling of ubiquitin conjugation to ER degradation” (CUE) domain, 2) bridges them with the membrane-bound Atg8 through the Atg8-interacting motif (AIM), 3) is co-degraded with ubiquitinated proteins in the vacuole, and 4) is required for their vacuolar degradation under nitrogen starvation conditions [9]. Moreover, because of the low affinity of CUE domain to ubiquitin and its ability to oligomerize, Cue5 can target only the ubiquitinated protein aggregates that, unlike ubiquitinated soluble proteins, are rich in ubiquitin for Cue5 oligomers to bind with high avidity [11]. By its oligomerization, Cue5 resembles the mammalian SQSTM1-like receptors even though they do not have this affinity problem with ubiquitin and oligomerize rather to cluster the ubiquitinated proteins into protein aggregates [5]. In proteaphagy, Cue5 satisfies the requirements of a SAR for the ubiquitinated 26S proteasomes (note that proteasomes are ubiquitinated after their inactivation with MG132; they are not ubiquitinated and do not require Cue5 for their autophagic degradation under nitrogen starvation conditions) [10]. Precisely, the S. cerevisiae Cue5: 1) binds ubiquitinated proteasomes, 2) bridges them with Atg8 using CUE and AIM domains, which are both essential for proteaphagy, and 3) is required for the vacuolar degradation of ubiquitinated proteasomes. Importantly, the inactivated 26S proteasomes must first be aggregated by Hsp42 chaperone into the cytoplasmic aggresomes before they can be degraded by the Cue5-mediated process [10]. This makes the Cue5-dependent aggrephagy and proteaphagy very similar and suggests that MG132 might also induce aggrephagy that coalesces with proteaphagy.
3. Ubiquitin-dependent but Cue5-independent autophagic pathways in yeast
Recently, Cue5 was also probed for the involvement in lipophagy [12,13]. Similar to the Cue5-dependent aggrephagy and proteaphagy (but unlike the Cue5-independent pathways, including the nitrogen starvation-induced proteaphagy) [9,10], lipophagy does not require the autophagic scaffold protein, Atg11 [14-17]. The similarity does not end there, as Komagataella phaffii Cue5 binds LDs via its CUE domain and is co-degraded with them in the vacuole (note that this happens in the stationary phase of growth, but not under nitrogen starvation conditions) [13]. However, both K. phaffii and S. cerevisiae Cue5 proteins are dispensable for the stationary phase lipophagy [12,13]. This means that in the stationary phase, Cue5 is just another substrate (and not a SAR) for lipophagy (Figure 1). Since Prl1 (positive regulator of lipophagy 1) is essential for Cue5 recruitment to LDs (most likely via LD ubiquitination) and for stationary phase lipophagy, we hypothesized that another ubiquitin-binding protein (AtgX) could be a SAR for the stationary phase lipophagy and, as such, would act as a lipophagy effector of the Prl1 regulator [13,17]. Here, we make a broader case for the existence and possible identities of such alternative ubiquitin-binding SARs in yeast.
Another argument for the presence of such receptors in yeast came from the studies of mitophagy in S. cerevisiae [18,19]. During mitophagy induced by the treatment of cells with rapamycin, the Ubp3-Bre5 deubiquitination complex is recruited from cytosol to the mitochondria where it inhibits mitophagy suggesting that ubiquitination of mitochondria plays an important positive role in their degradation [18]. Interestingly, the subsequent study found that ubiquitination of the two mitochondrial outer membrane proteins, Mdm12 and Mdm34, which are components of the endoplasmic reticulum-mitochondria encounter structure (ERMES), is required for efficient mitophagy under a variety of conditions, including rapamycin treatment, stationary phase of growth and nitrogen starvation [19]. However, despite a widespread involvement of Mdm12 and Mdm34 ubiquitination in mitophagy, the ubiquitin-binding Cue5 is not a part of the mechanism, at least under nitrogen starvation conditions [9], indicating that another ubiquitin-binding SAR must fulfil the role of additional receptor (the main one is a well-characterized Atg19-like receptor, Atg32 [20,21]) (Figure 1).
4. Candidates for alternative ubiquitin-binding SARs in yeast
Since at least two autophagic pathways (lipophagy and mitophagy) are ubiquitin-dependent but Cue5-independent, we looked for the proteins that, similar to Cue5, could bind both ubiquitin and Atg8. For this, we compared the physical interactomes of ubiquitin (179 proteins in Saccharomyces Genome Database (SGD)) and Atg8 (143 proteins in SGD and 92 proteins in [22]), and found 16 overlaps, including Cue5 (Table 1). The other 15 proteins are Arc40, Ccr4, Cdc48, Dhh1, Dsk2, Ede1, Ent1, Ent2, Nab2, Nup84, Rsp5, Shp1, Sla1, Ubx5 and Ydj1. Similar to Cue5, many of them are capable of self-interaction. Unlike Cue5, three of them (Ccr4, Dhh1 and Ede1) can bind Atg11. Interestingly, Ccr4 and Dhh1 affect autophagy in two directions: decrease it under growth conditions and increase it during nitrogen starvation by down- and up-regulating the expression of different ATG genes, respectively [23-25]. However, both Ccr4 and Dhh1, as well as another candidate, Nab2, are very promiscuous proteins with thousands of binding partners (Table 1). Therefore, they are unlikely to serve the highly specific roles of the SARs.

Table 1.
Proteins that physically interact with both ubiquitin and Atg8 in yeast*.
| Protein | Brief description | Ubiquitin-binding domain | Physical interactions | Autophagy regulator | ||
|---|---|---|---|---|---|---|
| Atg11 | Self | Total | ||||
| Arc40 | Subunit of the Arp2/3 complex | - | - | - | 51 | - |
| Ccr4 | Component of the Ccr4-Not complex | - | Yes | Yes | 3,035 | Yes [23] |
| Cdc48 | AAA ATPase with protein-unfoldase activity | - | - | Yes | 200 | Yes [29-31] |
| Cue5 | Ubiquitin-binding protein | CUE | - | Yes | 29 | Yes [9,10] |
| Dhh1 | DEAD-box helicase/mRNA decapping activator | - | Yes | Yes | 3,608 | Yes [24,25] |
| Dsk2 | Ubiquitin-like polyubiquitin-binding protein | UBA | - | Yes | 43 | Yes [26] |
| Ede1 | Endocytic adaptor | UBA | Yes [28] | Yes | 62 | Yes [28] |
| Ent1 | Epsin-like protein | UIM (x2) | - | - | 20 | - |
| Ent2 | Epsin-like protein | UIM (x2) | - | - | 48 | - |
| Nab2 | Nuclear polyadenylated RNA-binding protein | - | - | Yes | 2,641 | - |
| Nup84 | Subunit of the Nup84 subcomplex of NPC | - | - | Yes | 74 | - |
| Rsp5 | NEDD4 family E3 ubiquitin ligase | HECT | - | Yes | 354 | Yes [9,19,32] |
| Shp1 | UBX domain-containing protein, binds Cdc48 | UBA | - | - | 65 | Yes [30] |
| Sla1 | Cytoskeletal protein binding protein | - | - | Yes | 189 | - |
| Ubx5 | UBX domain-containing protein, binds Cdc48 | UBA, UIM [22] | - | - | 26 | Yes [22] |
| Ydj1 | Type I Hsp40 co-chaperone | - | - | Yes | 133 | Yes [27] |
*This analysis was performed based on the data in SGD (accessed on 11/22/2022) and reference [22], unless stated otherwise.
Importantly, 7 proteins, in addition to Cue5, have at least one known ubiquitin-binding domain: (1) Dsk2, Ede1, Shp1 and Ubx5 possess the ubiquitin-associated (UBA) domain, (2) Ent1 and Ent2 each carry two ubiquitin-interacting motifs (UIMs), and (3) Rsp5 has the ubiquitin-transfering “homologous to the E6-AP carboxyl terminus” (HECT) domain (Table 1). Interestingly, all UBA and HECT domain-containing proteins have been already implicated in autophagy. For example, Dsk2 (as well as another candidate, Ydj1) stimulates the formation of the mutated huntingtin inclusion body, which is then efficiently degraded by autophagy in yeast [26,27]. Similar to this artificial scenario, Dsk2 and Ydj1 could also promote the assembly and autophagic clearance of endogenous substrates. In addition, the third Atg11-binding protein, Ede1, through its oligomerization and condensation achieves the formation of the endocytic protein deposits in plasma membrane, and then, serves as a SAR for their autophagic degradation [28]. The Ede1’s UBA domain seems to be not involved in this process, but we cannot exclude the possibility that it might bring Ede1 to some other substrates for a similar phase separation into condensates that will be degraded by selective autophagy. Next, since another candidate, Cdc48, and its UBA domain-containing cofactor, Shp1, promote both selective and non-selective autophagy [29-31], their functioning as SARs is highly unlikely (all yeast SARs are dispensable for non-selective autophagy). Therefore, the Cdc48-Shp1 pair must play some other, more generic role in autophagic pathways that is not related to the selection of substrates. Finally, Ubx5 is another UBA domain-containing cofactor of Cdc48 that targets dysfunctional Cdc48 for autophagic degradation as a SAR, but is not involved in the autophagic degradation of functional Cdc48, piecemeal microautophagy of the nucleus and non-selective autophagy under nitrogen starvation conditions [22,30]. Interestingly, the UBA domain of Ubx5 is not required for the selective autophagy of dysfunctional Cdc48, which is bridged with Atg8 via the ubiquitin regulatory X (UBX) and UIM domains, respectively [22]. However, it might be required for the selective autophagy of ubiquitinated substrates yet to be identified.
Finally, the HECT domain-containing E3 ubiquitin ligase, Rsp5, regulates several autophagic pathways, including the Cue5-dependent aggrephagy, proteaphagy, and Cue5-independent mitophagy, but not a non-selective autophagy [9,19,32]. In aggrephagy, it is required for ubiquitination of proteins, their recognition by Cue5 and their vacuolar degradation. Interestingly, Rsp5 also ubiquitinates the Cue5 receptor itself, and is co-degraded with it and its aggrephagy substrates [9]. However, since autophagic degradation of Rsp5 heavily relies on Cue5 [9], it is unlikely to act as an alternative aggrephagy SAR. In proteaphagy, Rsp5 ubiquitinates the inactivated 26S proteasomes in cytoplasmic aggresomes created by Hsp42 and promotes their vacuolar clearance [32]. Similarly, Rsp5 also ubiquitinates the ERMES components, Mdm12 and Mdm34, and supports efficient vacuolar degradation of mitochondria via mitophagy [19]. In the latter two cases, the role of Rsp5 was studied only in the frame of ubiquitination, but since Rsp5 binds hundreds of proteins in addition to ubiquitin and Atg8 (Table 1), it is unlikely to function as a SAR in these or other pathways and bring enough selectivity to the process.
5. Conclusions
In summary, we think that a topic of the ubiquitin-binding autophagic receptors is understudied in yeast. While one such receptor (Cue5) has been identified and implicated in two autophagic pathways (aggrephagy and proteaphagy), our analysis suggests the existence of ubiquitin-dependent, but Cue5-independent, pathways (lipophagy and mitophagy) that could engage other ubiquitin- and Atg8-binding proteins as ubiquitin-binding SARs. Some of these proteins (Ede1 and Ubx5) act as the Atg19-like SARs in corresponding autophagic pathways. Therefore, they are poised to function also as the ubiquitin-binding SARs for other substrates. It will be interesting to see further developments in this exciting area of research.
Funding
This work was supported by the NIH grant, GM119571, to Taras Y. Nazarko.
Disclosure Statement
No potential conflict of interest was reported by the authors.
References
- Leber, R.; Silles, E.; Sandoval, I.V.; Mazón, M.J. Yol082p, a Novel CVT Protein Involved in the Selective Targeting of Aminopeptidase I to the Yeast Vacuole. J. Biol. Chem. 2001, 276, 29210–29217. [Google Scholar] [CrossRef]
- Scott, S.V.; Guan, J.; Hutchins, M.U.; Kim, J.; Klionsky, D.J. Cvt19 Is a Receptor for the Cytoplasm-to-Vacuole Targeting Pathway. Mol. Cell 2001, 7, 1131–1141. [Google Scholar] [CrossRef]
- Bjorkoy, G.; Lamark, T.; Brech, A.; et al. p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. J. Cell Biol. 2005, 171, 603–14. [Google Scholar] [CrossRef]
- Pankiv, S.; Clausen, T.H.; Lamark, T.; et al. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 2007, 282, 24131–45. [Google Scholar] [CrossRef] [PubMed]
- Kirkin, V.; Rogov, V.V. A Diversity of Selective Autophagy Receptors Determines the Specificity of the Autophagy Pathway. Mol. Cell 2019, 76, 268–285. [Google Scholar] [CrossRef]
- Shroff, A.; Nazarko, T.Y. SQSTM1, lipid droplets and current state of their lipophagy affairs. Autophagy 2022, 19, 720–723. [Google Scholar] [CrossRef]
- Kim, J.; Huang, W.P.; Stromhaug, P.E.; et al. Convergence of multiple autophagy and cytoplasm to vacuole targeting components to a perivacuolar membrane compartment prior to de novo vesicle formation. J. Biol. Chem. 2002, 277, 763–73. [Google Scholar] [CrossRef]
- Shintani, T.; Huang, W.-P.; Stromhaug, P.E.; Klionsky, D.J. Mechanism of Cargo Selection in the Cytoplasm to Vacuole Targeting Pathway. Dev. Cell 2002, 3, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Psakhye, I.; Jentsch, S. Autophagic Clearance of PolyQ Proteins Mediated by Ubiquitin-Atg8 Adaptors of the Conserved CUET Protein Family. Cell 2014, 158, 549–563. [Google Scholar] [CrossRef]
- Marshall, R.S.; McLoughlin, F.; Vierstra, R.D. Autophagic Turnover of Inactive 26S Proteasomes in Yeast Is Directed by the Ubiquitin Receptor Cue5 and the Hsp42 Chaperone. Cell Rep. 2016, 16, 1717–1732. [Google Scholar] [CrossRef]
- Lu, K.; den Brave, F.; Jentsch, K.L.F.D.B.S. Receptor oligomerization guides pathway choice between proteasomal and autophagic degradation. Nat. Cell Biol. 2017, 19, 732–739. [Google Scholar] [CrossRef]
- Robichaud, S.; Fairman, G.; Vijithakumar, V.; Mak, E.; Cook, D.P.; Pelletier, A.R.; Huard, S.; Vanderhyden, B.C.; Figeys, D.; Lavallée-Adam, M.; et al. Identification of novel lipid droplet factors that regulate lipophagy and cholesterol efflux in macrophage foam cells. Autophagy 2021, 17, 3671–3689. [Google Scholar] [CrossRef]
- Kumar, R.; Shroff, A.; Nazarko, T.Y. Komagataella phaffii Cue5 Piggybacks on Lipid Droplets for Its Vacuolar Degradation during Stationary Phase Lipophagy. Cells 2022, 11, 215. [Google Scholar] [CrossRef]
- van Zutphen, T.; Todde, V.; de Boer, R.; Kreim, M.; Hofbauer, H.F.; Wolinski, H.; Veenhuis, M.; van der Klei, I.J.; Kohlwein, S.D. Lipid droplet autophagy in the yeast Saccharomyces cerevisiae. Mol. Biol. Cell 2014, 25, 290–301. [Google Scholar] [CrossRef]
- Wang, C.-W.; Miao, Y.-H.; Chang, Y.-S. A sterol-enriched vacuolar microdomain mediates stationary phase lipophagy in budding yeast. J. Cell Biol. 2014, 206, 357–366. [Google Scholar] [CrossRef]
- Seo, A.Y.; Lau, P.W.; Feliciano, D.; et al. AMPK and vacuole-associated Atg14p orchestrate mu-lipophagy for energy production and long-term survival under glucose starvation. Elife 2017, 6. [Google Scholar] [CrossRef]
- Kumar, R.; Rahman, M.A.; Nazarko, T.Y. Nitrogen Starvation and Stationary Phase Lipophagy Have Distinct Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 9094. [Google Scholar] [CrossRef]
- Müller, M.; Kötter, P.; Behrendt, C.; Walter, E.; Scheckhuber, C.Q.; Entian, K.-D.; Reichert, A.S. Synthetic Quantitative Array Technology Identifies the Ubp3-Bre5 Deubiquitinase Complex as a Negative Regulator of Mitophagy. Cell Rep. 2015, 10, 1215–1225. [Google Scholar] [CrossRef]
- Belgareh-Touzé, N.; Cavellini, L.; Cohen, M.M. Ubiquitination of ERMES components by the E3 ligase Rsp5 is involved in mitophagy. Autophagy 2017, 13, 114–132. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Kondo-Okamoto, N.; Ohsumi, Y. Mitochondria-Anchored Receptor Atg32 Mediates Degradation of Mitochondria via Selective Autophagy. Dev. Cell 2009, 17, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Kanki, T.; Wang, K.; Cao, Y.; Baba, M.; Klionsky, D.J. Atg32 Is a Mitochondrial Protein that Confers Selectivity during Mitophagy. Dev. Cell 2009, 17, 98–109. [Google Scholar] [CrossRef]
- Marshall, R.S.; Hua, Z.; Mali, S.; et al. ATG8-Binding UIM Proteins Define a New Class of Autophagy Adaptors and Receptors. Cell 2019, 177, 766–781. [Google Scholar] [CrossRef]
- Yin, Z.; Zhang, Z.; Lei, Y.; Klionsky, D.J. Bidirectional roles of the Ccr4-Not complex in regulating autophagy before and after nitrogen starvation. Autophagy 2022, 19, 415–425. [Google Scholar] [CrossRef]
- Hu, G.; McQuiston, T.; Bernard, A.; Park, Y.-D.; Qiu, J.; Vural, A.; Zhang, N.; Waterman, S.R.; Blewett, N.H.; Myers, T.G.; et al. A conserved mechanism of TOR-dependent RCK-mediated mRNA degradation regulates autophagy. Nature 2015, 17, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yao, Z.; Jin, M.; Namkoong, S.; Yin, Z.; Lee, J.H.; Klionsky, D.J. Dhh1 promotes autophagy-related protein translation during nitrogen starvation. PLOS Biol. 2019, 17, e3000219. [Google Scholar] [CrossRef]
- Chuang, K.-H.; Liang, F.; Higgins, R.; Wang, Y. Ubiquilin/Dsk2 promotes inclusion body formation and vacuole (lysosome)-mediated disposal of mutated huntingtin. Mol. Biol. Cell 2016, 27, 2025–2036. [Google Scholar] [CrossRef]
- Higgins, R.; Kabbaj, M.-H.; Hatcher, A.; Wang, Y. The absence of specific yeast heat-shock proteins leads to abnormal aggregation and compromised autophagic clearance of mutant Huntingtin proteins. PLOS ONE 2018, 13, e0191490. [Google Scholar] [CrossRef]
- Wilfling, F.; Lee, C.-W.; Erdmann, P.S.; Zheng, Y.; Sherpa, D.; Jentsch, S.; Pfander, B.; Schulman, B.A.; Baumeister, W. A Selective Autophagy Pathway for Phase-Separated Endocytic Protein Deposits. Mol. Cell 2020, 80, 764–778. [Google Scholar] [CrossRef]
- Ossareh-Nazari, B.; Bonizec, M.; Cohen, M.; Dokudovskaya, S.; Delalande, F.; Schaeffer, C.; Van Dorsselaer, A.; Dargemont, C. Cdc48 and Ufd3, new partners of the ubiquitin protease Ubp3, are required for ribophagy. Embo Rep. 2010, 11, 548–554. [Google Scholar] [CrossRef]
- Krick, R.; Bremer, S.; Welter, E.; et al. Cdc48/p97 and Shp1/p47 regulate autophagosome biogenesis in concert with ubiquitin-like Atg8. J. Cell Biol. 2010, 190, 965–73. [Google Scholar] [CrossRef] [PubMed]
- Buchan, J.R.; Kolaitis, R.M.; Taylor, J.P.; et al. Eukaryotic stress granules are cleared by autophagy and Cdc48/VCP function. Cell 2013, 153, 1461–74. [Google Scholar] [CrossRef]
- Marshall, R.S.; Vierstra, R.D. A trio of ubiquitin ligases sequentially drives ubiquitylation and autophagic degradation of dysfunctional yeast proteasomes. Cell Rep. 2022, 38, 110535. [Google Scholar] [CrossRef]
Figure 1.
The ubiquitin-dependent selective autophagy pathways in yeast. Cue5 is the only known ubiquitin-binding SAR that functions in aggrephagy and proteaphagy. However, the Cue5-independent pathways of lipophagy and mitophagy suggest the existence of other ubiquitin-binding SARs (depicted as AtgX). For potential identities of AtgX, see Table 1.
Figure 1.
The ubiquitin-dependent selective autophagy pathways in yeast. Cue5 is the only known ubiquitin-binding SAR that functions in aggrephagy and proteaphagy. However, the Cue5-independent pathways of lipophagy and mitophagy suggest the existence of other ubiquitin-binding SARs (depicted as AtgX). For potential identities of AtgX, see Table 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



