Submitted:
11 January 2023
Posted:
12 January 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Pharmacovigilance databases are showing evidence of injury in the context of the COVID-19 modified mRNA shots. According to recent publications, adverse event reports linked to the mRNA COVID-19 products largely point to the spike protein as an aetiological agent of adverse events, but we propose that the platform itself may be culpable. To assess the safety of current and future mRNA vaccines, further analysis on the risks due to the platform itself, and not specifically the expressed antigen. If harm can be exclusively and conclusively attributed to the spike protein, then it is possible that future mRNA vaccines expressing other antigens will be safe. If harms are attributable to the platform itself, then regardless of the toxicity, or lack thereof, of the chosen payload therein, the platform may be inherently unsafe, pending modification. In this work, we examine previous studies of RNA-based delivery by a lipid nanoparticle and break down the possible etiological elements of harm.
Keywords:
COVID-19 vaccination
; mRNA vaccines
; Clinical Trials
; Safety Assessment
; Novel Technologies
; Spike protein
1. Introduction
Pharmaceutical and medical device approvals are based in structured approval processes and historically, the track record for approval has been far from ideal. There are many examples of over-turnings of approvals of pharmaceuticals post facto, due to emergence of oversights of particular safety factors that had occurred during the approval process [1]. The former examples represent failures in regulatory bodies to sufficiently assess safety during this process and this can be quite costly both in terms of potential economic and individual harms [2]. To put this ever-present issue into perspective, of 309 novel cardiovascular, orthopaedic and neurologic devices approved in the EU between 2005 and 2010, 73 (24%) were subject to either a safety alert or product recall [3], consistent with other reported rates [4]. Importantly, as complexities and novelties of products are increasing, approval success rates have been decreasing [5] and in the face of new drug approvals currently marred by low Phase III trial success rates (~10%), this is an issue in dire need of acknowledgement and remedy [6]. The precautionary principle dictates that caution must be exercised in the context of potential safety issues with novel drugs and technologies, and thus due to the low success rates of novel and unprecedented drugs [6,7,8], and the potential risks to the population, it is important to adopt the precautionary principle [9] when approving any pharmacological products, especially those given to large populations. COVID-19 mRNA products are novel with regard to their delivery system and their content as the first mRNA vaccines approved for use in humans, as well as the first approved coronavirus vaccine in humans. The speed at which they were designed, developed, approved and administered is also unprecedented in pharmaceutical history [10] and defies traditional timelines for testing of biological products for use in humans.
Given the approval of the mRNA platform by health regulators across the globe, industry is poised to develop new vaccines using mRNA, as it is a versatile platform which only requires the genetic sequence of the target antigen. The administration of billions of doses has resulted in great industry enthusiasm for the platform, and other mRNA products are being developed using the same core technology [11,12].
To assess the novelty of COVID-19 mRNA products, we look back to the history of mRNA vaccines, which begins with experiments on in-vitro transcribed RNA, i.e., delivering RNA to a cell for expression of a protein of interest. Synthetic RNA technology has a wide variety of applications, from the delivery of small interfering RNAs (siRNAs) to reduce gene expression, or messenger RNAs (mRNAs) to encode for a protein of therapeutic value, or to encode for an antigen to stimulate an immune response, as in the strategy of mRNA vaccination.
Early attempts to express proteins from injected mRNA faced several challenges. First, bare RNA produces an inflammatory response, limiting the expression potential of the RNA, as it is broken down. Secondly, it is difficult for the bare RNA to enter through a cell membrane. These issues were addressed through the processes of pseudouridinylation and encapsulation in a lipid nanoparticle (LNP) respectively. The former discovery decreased the lability of RNA, enabling it to remain in the body for longer periods of time. The latter discovery not only shielded the RNA from the host’s immune response as well as RNAses, it also enabled efficient uptake by cells, where it could be efficiently translated by host ribosomes. Pseudouridine was later replaced by m1-methyl-psuedouridine, owing to its greater translation fidelity, higher expression, and better evasion of the host immune response.
LNP development was improved through two innovations, PEGylation, and the use of cationic lipids. LNP surface modifications by poly-ethylene glycol (PEG) enabled lipid nanoparticles to survive for longer lengths of time still, so that their package contents could be delivered to cells to provoke an immune response when the antigen is expressed. Another important development for LNPs is the use of cationic lipids; enabling efficient self-assembly and encapsulation of the mRNA. Cationic lipids can additionally be modified to deliver drugs to certain cell types, an important consideration when delivering mRNA.
There is a prior history of drug delivery by lipid nanoparticles (LNP), and one approved drug for the delivery of a small interfering RNA (Onpattro [13]). Questions remain pertaining to the safety of mRNA vaccines, as several assumptions on which they were rapidly approved, have been either challenged or overturned by experimental [14] and clinical evidence [15]. Quoted theoretical safety advantages were the ease of production without contamination (mRNA vaccines do not require live virus production), and lower (in theory non-existent) risks of infection or host genome integration [16]. Beforehand, concerns existed over the induction of Type I interferon responses by mRNA vaccines [17,18], which are associated with inflammation and autoimmunity [19,20].
For example, the dual assumptions that LNPs remain at the injection site, and that the mRNA degrades quickly have been shown to be false; biodistribution and bioaccumulation data indicate that LNPs can enter the bloodstream [14,21,22,23,24,25], and peer-reviewed studies have shown durability of both mRNA and spike protein for in vivo [23] and up to 4 months post injection for spike protein [21]. Given the novelty of mRNA vaccines, and the increasing evidence of harm from clinical reports [15], epidemiology [26] as well as laboratory science [27], there are open safety concerns to be addressed by future research.
Multiorgan vasculitis, with notable involvement of small vessels, is now emerging as a common theme underlying adverse events following COVID-19 vaccination. Affliction of capillaries with thrombus formation in brain and heart are repeatedly being observed [28,29,30]. This pattern of disease in small and smallest vessels is novel and deemed by the authors to be characteristic for the action of mRNA-based vaccines.
The outcome with future mRNA vaccines against other pathogens will be much the same as we have witnessed with the COVID-19 vaccines. It is true that the spike protein itself can promote blood clotting and inflammation without any help from the immune system [31]. Nevertheless, the available evidence indicates that the grave, widespread and sustained injury to tissues and to blood vessels is mostly caused by the immune attack on spike protein-producing cells. This attack occurs simply because the spike protein is a non-self antigen; and since every other mRNA vaccine will encode its own non-self antigen, derived from whichever particular microbe it targets, we must expect that it will cause harm by the same mechanism and to a similar extent. These scenarios will worsen with each successive booster.
This review summarizes known mechanisms of harm specific to mRNA vaccines, where we examine historical data on mRNA vaccines to determine if safety signals were apparent during production or testing. Prior to the trials on COVID-19 vaccines involving tens of thousands of people, public data existed on only 285 people administered mRNA vaccines, with the earliest trials finishing in 2018 and exhibiting high rates (>10%) of severe adverse events (Supplementary Table S1). The novelty of mRNA/LNP products must be stressed in guiding their safety assessment, as current approvals still leave many questions unanswered, and serious risks cannot be definitively ruled out based on current evidence.
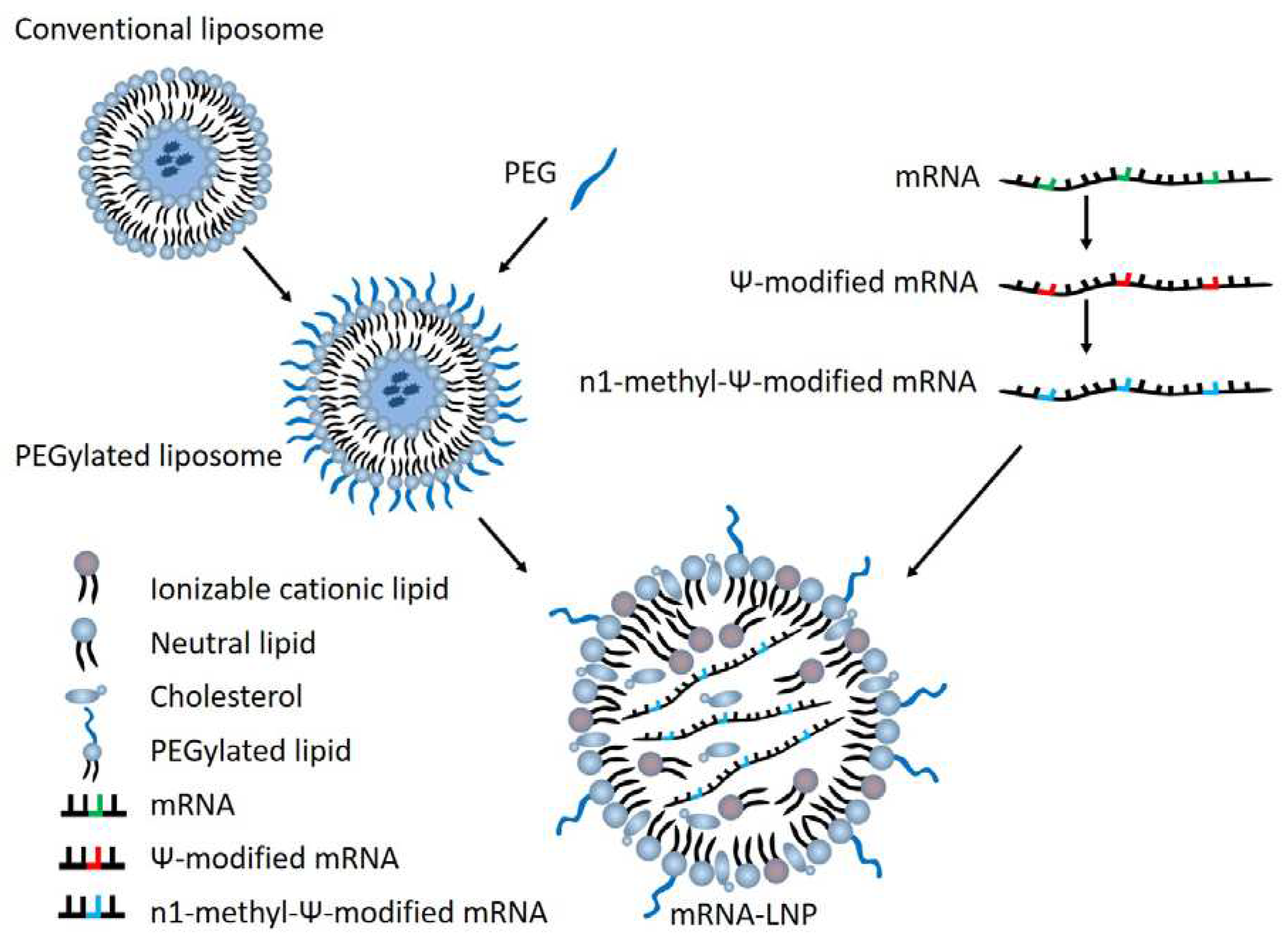
Figure 1.
Overview of mRNA-LNP vaccine components.
In this review, we summarize what is known about the individual components of mRNA vaccines individually, by reviewing literature on past therapeutics. Additionally, we review the known safety impacts of mRNA vaccines prior to COVID-19, as well as other coronavirus vaccines which, while using a non-mRNA platform, inform us of safety risks when vaccinating against coronaviruses.
2. mRNA Vaccine Elements and Potential for Harm
2.1. Harms due to Lipid Nanoparticle (LNP)
Lipid nanoparticles have been used in the delivery of drugs for decades, beginning with the 1990 EU approval of the drug AmBisome (LNP-encapsulated Amphotericin B) for fungal infections [13]. In the US environment, the first LNP administered drugs were Doxil (LNP-encapsulated Doxorubicin) for Kaposi’s sarcoma and Abelcet (LNP-encapsulated Amphotericin B) for asperillosis [13].
The simplest form of LNP is a liposome, which is produced endogenously. This consists merely of a lipid bilayer which separates the contents from the outside environment. While simple liposomes are detected and destroyed by the body’s immune system, the addition of polyethylene glycol (PEG), which enables the liposome to evade the host’s immune response and last longer in the body to deliver the encapsulated product.
While PEG is thought to be inert in the body, the injection of PEG does elicit anti-IgM antibodies, and subsequent injections are cleared faster due to this immune response [32]. Additionally, a small proportion of the population has an allergy to PEG, and injection can trigger anaphylaxis, as has happened for several people receiving COVID-19 vaccines [33,34,35,36].While PEG is thought to be inert in the body, the injection of PEG does elicit anti-IgM antibodies, and subsequent injections are cleared faster due to this immune response [e2].
The safety of DSPC, a component of the LNP used in both the Pfizer and Moderna COVID-19 vaccines, has been studied for its safety [37]. Animal studies in mice ruled that it was likely not toxic to humans, as no clinical manifestations were present. LNPs have been claimed safe for delivery of therapeutic agents, according to a review [38]. However, pro-inflammatory concerns remain over LNPs, even in isolation [39,40].
2.2. Harms due to Exogenous RNA
Extracellular RNA exists as a pro-coagulant [41], and increases the permeability of the endothelial cells in brain microvasculature [42]. .Foreign RNA of course triggers an inflammatory response, as a toll like receptor (TLR) [43] and retinoic acid-inducible gene I (RIG-I) [44] are activated. In fact, the initial reason for modification of the RNA by pseudouridinylation was to bypass activation of TLR [45]. As pseudouridinylated RNA was translated at lower fidelity than RNA [46], the nucleosides were modified to N1-methyl-pseudouridine, which brought translation fidelity to near that of RNA [47].
The properties of both ΨRNA and N1-mΨRNA have been studied in some depth, though questions still remain. ΨRNA exists in nature and comprises between 0.2% and 0.6% of uridine content in human cell lines, and has biologically significant differences from RNA [48]. While N1-mΨRNA exists in nature, and is found within the archaea [49], studies on its properties go back only as recently as 2015 [50]. Additionally, important biological differences exist between unmodified and modified RNA. For example, through some application of the central dogma of molecular biology, it is assumed that RNA vaccines cannot be incorporated into the genome. This statement is not supported by the research [51], and is in fact contradicted by experiments showing reverse transcription of the Pfizer BioNTech COVID-19 mRNA vaccine into a human liver cell line [27].
2.3. Harms due to In Vitro Transcribed (IVT) RNA
One level of complexity over delivery of a non-expressed RNA are therapeutics which deliver an mRNA which is expressed by the host translational system. For these applications, typically you are replacing a damaged protein of interest by supplying it exogenously. Using an LNP-mRNA platform here is better than supplying the protein itself, as a protein expressed from IVT RNA is more likely to have the correct post transcriptional modifications (and subsequent conformation) for its cell type than an exogenously supplied protein [52]. For these applications, it is typically necessary for the drug to be administered repetitively over long time periods [52,53]. With repetitive dosing, safety is very important, as even a low per-dose AE rate can compound over the many doses of the treatment.
Most studies of this therapeutic modality so far focus on drug efficacy, and limited safety data exists. In a 2021 review of non-immunologic application of mRNA, all studies using LNP-mRNA as protein replacement therapy demonstrated liver toxicity or lacked safety data [52]. Several studies also saw the development of anti-drug antibodies (ADAs) [54,55,56], which can deactivate the drug and prevent treatment [57,58,59,60]. Immune-mediated toxicity is also a cause for concern [61,62]. Most studies of this therapeutic modality so far focus on drug efficacy, and limited safety data exists. In a 2021 review of non-immunologic application of mRNA, all studies using LNP-mRNA as protein replacement therapy demonstrated liver toxicity or lacked safety data [52]. Several studies also saw the development of anti-drug antibodies (ADAs) [54,55,56], which can deactivate the drug and prevent treatment [57,58,59,60]. Immune-mediated toxicity is also a cause for concern [61,62].Recent work has demonstrated a class switch towards an IgG4 antibody response, observed after three doses of Pfizer BNT162b2 (COVID-19 vaccine) and not adenoviral vector COVID-19 vaccines [63], which raises concerns over possible immune tolerance, which is linked to an IgG4-dominated response [64,65].
With ADAs, there is also the concern of the development of cross-reactivity to endogenous proteins, which can occur if the endogenous protein possesses similar structural motifs to the protein expressed from the administered mRNA [66]. Thromboembolic events have been observed in ADA reactions [58]. Typically, ADA reactions are decreased in cases where the encoded protein is a ‘self’ protein, as opposed to an exogenous protein [67].
2.4. Harms of RNA Vaccination
In addition to the other harms present in IVT RNAs, RNA vaccines also have the additional safety challenges of expressing an exogenous protein for the express purpose of generating an immune response and immune memory [68]. Of the RNA therapeutic systems introduced so far in this review, the mRNA vaccines are the most complex drug-like therapeutic biologic.
Limited safety data exists on RNA vaccines against infection [69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89] (Supplementary Table S1). Prior to the trials for COVID-19 vaccines, there was data on 285 patients, with the earliest trials on a non-HIV vaccine only completed in 2018. The serious adverse event (SAE) rate of these exploratory trials was 14±2% (Grade 3 or above1). As a comparison, a post-marketing surveillance study of influenza vaccines in the UK found an SAE rate of 0.16% [90]:, almost 100 times less than the SAE rate for mRNA vaccines.
In addition, study results that might capture long-term effects are not available, or limited. such as those mediated by the immune response with later variants of the virus. While the type of vaccination (i.e., attenuated live virus, inactivated virus, mRNA) should not have a significant impact on the IgG antibodies produced, an important consideration must be mentioned; mRNA vaccines, in encoding for a limited number of proteins (a single antigen in most cases), better enable immune escape than a broader antibody response including other proteins. Recent evidence shows a subclass switch from IgG1 to IgG4 in the context the Comirnaty mRNA product, which may have consequences with regard to cancer [91], pregnancy [92] and IgG4-related diseases [63,93]. COVID-19 mRNA vaccines are commonly used in Europe and North America; these encode specifically and exclusively for the spike (S) protein [94,95]. Since the introduction of vaccines, mutations have occurred lessening the neutralizing capacity of these vaccines [96,97].
2.5. Harms of Coronavirus Vaccination
In addition to the considerations on the novelty of mRNA vaccines, the C19 mRNA vaccines are also unprecedented on another quality, namely, they are the first coronavirus vaccines approved in humans. Following the 2002/2003 outbreak of SARS-CoV [98] and the 2012 outbreak of MERS-CoV [99], In addition to the considerations on the novelty of mRNA vaccines, the C19 mRNA vaccines are also unprecedented on another quality, namely, they are the first coronavirus vaccines approved in humans. Following the 2002/2003 outbreak of SARS-CoV [98] and the 2012 outbreak of MERS-CoV [99], vaccines against coronaviruses infecting humans gained more attention [100]. Coronavirus vaccines were tested in both animal models as well as human subjects [100].
Animal trials on four SARS vaccine candidates in ferrets demonstrated an initial protective period against infection, followed by hypersensitivity to rechallenge with SARS-CoV. The ferrets developed histopathological changes in the lungs induced from virus challenge after all four vaccine candidates, suggesting immune mediated damage [101]. One 2004 study on ferrets given a vaccine against SARS-CoV-2 showed enhanced hepatitis [102].
Mice given an inactivated virus later developed a pro-inflammatory pulmonary response upon challenge [103]. Furthermore, anti-spike IgG antibodies, as are produced by all of the mRNA vaccines, in addition to most other Covid-19 vaccines, are demonstrated to cause severe acute lung injury on re-exposure to the virus, suggesting a negative impact of a narrow immune response [104]. MERS-CoV vaccines have been tested in mice and rhesus macaques [105], revealing protection without visible histopathology.
Immune mediated danger from vaccines has been widely acknowledged to be an extant issue in the development of coronavirus vaccines [106,107,108,109,110,111,112], and is supported by current evidence [113]. During the rapid development of Covid-19 vaccines, it was an issue of concern that sufficient long-term monitoring for antibody-dependent enhancement (ADE) effects be put into place [114,115]. Unfortunately, as of this writing, there is not data available on the long term impacts of Covid-19 vaccines, including effects resulting from rechallenge with the virus. Immune mediated danger from vaccines has been widely acknowledged to be an extant issue in the development of coronavirus vaccines [106,107,108,109,110,111,112], and is supported by current evidence [113].
Other, more distantly related coronaviruses have veterinary vaccines available. These have been summarized in a recent review [116]. Evidence of immune dependent enhancement was present for cell culture experiments on vaccination against feline coronaviruses [117,118,119]. Additionally, ADE is a concern for avian infectious bronchitis virus (IBV), a coronavirus [120,121]. In IBV, suboptimal vaccination alters the evolutionary dynamics of the viruses and can contribute to the production of escape mutants [122,123,124]. Finding broadly neutralizing IBV vaccines remains a significant challenge for the poultry industry [125,126,127,128,129].
Early canine coronavirus vaccines were withdrawn due to neurological symptoms [130,131], though current vaccines do not carry the same safety issues [132,133]. Bovine coronavirus vaccinations often fail to provide immunity against subsequent reinfections [134,135,136]. Immunizations against transmissible gastroenteritis virus (TGEV) in swine have historically had issues in inducing immune protection [137,138], but are widely used now. Too frequent exposure to vaccine antigens can lower the immune response against TGEV [139].Another swine coronavirus porcine epidemic diarrhea virus (PEDV) is widely used. Extant safety concerns are minor, and mostly deal with lack of efficacy; these are summarized in a review [140].Bovine coronavirus vaccinations often fail to provide immunity against subsequent reinfections [134,135,136]. Immunizations against transmissible gastroenteritis virus (TGEV) in swine have historically had issues in inducing immune protection [137,138], but are widely used now. Too frequent exposure to vaccine antigens can lower the immune response against TGEV [139].Another swine coronavirus porcine epidemic diarrhea virus (PEDV) is widely used. Extant safety concerns are minor, and mostly deal with lack of efficacy; these are summarized in a review [140].
Human trials of coronavirus vaccines have also taken place prior to the approval of Covid-19 vaccines. In addition to the endemic coronaviruses which infect humans, several epidemic strains of coronaviruses have occurred in the past two decades, namely the coronaviruses associated with severe acute respiratory syndrome (SARS-CoV) in 2003 [98] and middle eastern respiratory syndrome (MERS-CoV) in 2012 [141]. These outbreaks impelled the production of coronavirus vaccine candidates, summarized in a recent review [100] (Table 1). In total, before the development of the Covid-19 vaccines, data existed on a total of 179 human participants given a SARS or MERS vaccine candidate, of which, 7 (4±2%) experienced a serious adverse event (Table 1). A human trial of 63 adults for a MERS vaccine candidate showed no severe adverse events, but infections in 36% of participants [142,143].
2.6. Harms of RNA Vaccination with SARS-CoV-2 Spike (S) Antigen
There is reason to believe that vaccines encoding the spike (S) protein of SARS-CoV-2 have additional mechanisms of harm, owing to the biological impacts of S protein specifically. There is a wide literature [150,151,152,153], and it is beyond the scope of this review to cover this in significant depth. However, the addition of spike protein adds another factor in assessing the complexity of RNA vaccines. The complexity, as well as uncertainties about possible harms, are non-trivial and cannot be dismissed based on current data. This section will briefly cover some of the hypothesized mechanisms of harm from spike protein encoding mRNA vaccines and the evidence for each from a clinical/epidemiological outlook as well as any mechanistic data from laboratory work.
Several observations have been made which contradict fundamental claims of RNA vaccine safety. For example, it was assumed that the RNA was relatively labile and transient in the cell. However, studies observe spike protein and vaccine mRNA up to 4 months post-injection [21,23]. Spike protein has been shown in laboratory settings to cause inflammation [154,155], vascular damage [156], and to act as a seed for amyloid formation [157].
The use of the spike protein for an immune response is also concerning as it is not only the pathogenic part of covid 19 but also has been shown to enter the nucleus [158]. Having both one of the most pathogenic and lethal proteins ever discovered found within the nucleus of human cells with its genetic code is a concerning discovery. It was also shown to suppress anticancer mechanisms in the body. Singh and Singh demonstrated Spike protein models that anticipated an interaction with tumor suppressor genes P53 and BRCA1 [159].
2.6.1. Harms of Cross Reactivity and Autoimmunity
Cross reactivity has been considered one of the original problems of any vaccination. For the covid spike protein it was found that 60 pentapeptides are shared by SARS-CoV-2 spike glycoprotein (gp) and human proteins that when altered, mutated, deficient or, however, improperly functioning, may cause vascular diseases, and thromboembolic complications [160,161].
Other concerns exist over possible immune damage. One preprint study demonstrated a change in both the adaptive and innate immune responses following a full series of Pfizer BNT162b2 [162]. Combined with the demonstration of Ig class switch to IgG4 following a BNT162b2 booster dose [63], immune mediated harm is a very real possibility.
3. Discussion
There is limited information to make a safety assessment of mRNA vaccines. In the category of mRNA vaccines, there are patient data for 385 patients. For mRNA vaccines against an infection, there was patient data for 285 patients. The rate of serious adverse events was 64 out of 385 for the broad category of RNA vaccines (including cancer vaccines), or 17%; restricting the definition to vaccines against infection, the rate of SAEs is 41/285 or 14%. While high levels can be expected for trials of a novel technology where dosage levels must be determined (many of these trials are phase I) [163], these findings showcase the relative immaturity of mRNA vaccination as a strategy. Given the low efficacy and short duration of protection of SARS-CoV-2 mRNA products [164,165], and the low risk of many populations from COVID-19 complications [166], it may be advisable for the suspension of mRNA vaccines in certain risk cohorts.
The key to the reactivity of mRNA vaccines is the fact that they express a foreign antigen, for which the antigen presenting cells are marked for destruction. While the lipid nanoparticle exhibits an acute inflammatory response by itself [39,40,167], the trials using LNPs so far have not found a large safety signal when using LNPs to deliver small molecules, non-expressing RNAs, or RNAs for endogenous proteins [39,40,167].
In addition to there being harms attributable to the general immune response from an LNP-RNA delivery system, there are also some harms specific to the spike protein. Several of these mechanisms are supported by laboratory experiments and clinical findings but need more investigation. Medicine is replete with cases for which safety was assumed without adequate evidence at the time, which later regretfully led to loss of health and life. mRNA vaccines are demonstrating great unintended harms, and these harms demand further investigation into mechanism, which is important for identifying treatment modalities.
Novel biomedical technologies can bring relief to a wide variety of conditions and diseases. However, their use must take into consideration their possible harms. Here, we argue that the mRNA technology is novel enough that safety concerns in current and future products cannot be definitively ruled out, and further research must be performed to ensure their safety for current and future users. Considering the lack of data on the platform itself, we recommend a robust, independent and wide-ranging safety audit of mRNA-LNP formulations and call on regulators to hold manufacturers to high safety standards, especially for products used prophylactically in the general population.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Safety profile of previous LNP-mRNA products.
Author Contributions
Conceptualization, M.T.J.H.; writing—original draft preparation, M.T.J.H.; research, M.T.J.H, J.R. and T.L.; writing—review and editing, M.T.J.H, J.R., A.J. and T.L.; supervision, M.T.J.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We thank Cristoph Plothe for his comments.
Conflicts of Interest
M.T.J.H, A.J and T.L. are members of World Council for Health, a non-profit organization for holistic health promotion.
| 1 | Event classification available at: https://rsc.niaid.nih.gov/sites/default/files/corrected-grading-table-v-2-1-with-all-changes-highlighted.pdf. |
References
- Nagaich, U.; Sadhna, D. Drug Recall: An Incubus for Pharmaceutical Companies and Most Serious Drug Recall of History. Int J Pharm Investig 2015, 5, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Gagne, J.J.; Choudhry, N.K. The Epidemiology of Drug Recalls in the United States. Arch Intern Med 2012, 172, 1109–1110. [Google Scholar] [CrossRef] [PubMed]
- Hwang, T.J.; Sokolov, E.; Franklin, J.M.; Kesselheim, A.S. Comparison of Rates of Safety Issues and Reporting of Trial Outcomes for Medical Devices Approved in the European Union and United States: Cohort Study. BMJ 2016, 353, i3323. [Google Scholar] [CrossRef] [PubMed]
- Vajapey, S.P.; Li, M. Medical Device Recalls in Orthopedics: Recent Trends and Areas for Improvement. J Arthroplasty 2020, 35, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Young, R.; Bekele, T.; Gunn, A.; Chapman, N.; Chowdhary, V.; Corrigan, K.; Dahora, L.; Martinez, S.; Permar, S.; Persson, J.; et al. Developing New Health Technologies for Neglected Diseases: A Pipeline Portfolio Review and Cost Model 2020.
- Dowden, H.; Munro, J. Trends in Clinical Success Rates and Therapeutic Focus. Nature Reviews Drug Discovery 2019, 18, 495–496. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.; Siah, K.W.; Lo, A.W. Estimation of Clinical Trial Success Rates and Related Parameters. Biostatistics 2019, 20, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, E.; Nelson, G.M.; Haynes, M.; Sargeant, F. Success Rates for Product Development Strategies in New Drug Development. J Clin Pharm Ther 2016, 41, 198–202. [Google Scholar] [CrossRef]
- Hayes, A.W. The Precautionary Principle. Arh Hig Rada Toksikol 2005, 56, 161–166. [Google Scholar] [PubMed]
- Ball, P. The Lightning-Fast Quest for COVID Vaccines — and What It Means for Other Diseases. Nature 2020, 589, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Le, T.; Sun, C.; Chang, J.; Zhang, G.; Yin, X. MRNA Vaccine Development for Emerging Animal and Zoonotic Diseases. Viruses 2022, 14, 401. [Google Scholar] [CrossRef]
- Ladak, R.J.; He, A.J.; Huang, Y.-H.; Ding, Y. The Current Landscape of MRNA Vaccines Against Viruses and Cancer-A Mini Review. Front Immunol 2022, 13, 885371. [Google Scholar] [CrossRef] [PubMed]
- Akinc, A.; Maier, M.A.; Manoharan, M.; Fitzgerald, K.; Jayaraman, M.; Barros, S.; Ansell, S.; Du, X.; Hope, M.J.; Madden, T.D.; et al. The Onpattro Story and the Clinical Translation of Nanomedicines Containing Nucleic Acid-Based Drugs. Nat. Nanotechnol. 2019, 14, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Ogata, A.F.; Cheng, C.-A.; Desjardins, M.; Senussi, Y.; Sherman, A.C.; Powell, M.; Novack, L.; Von, S.; Li, X.; Baden, L.R.; et al. Circulating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Vaccine Antigen Detected in the Plasma of MRNA-1273 Vaccine Recipients. Clinical Infectious Diseases 2022, 74, 715–718. [Google Scholar] [CrossRef]
- Deb, A.; Abdelmalek, J.; Iwuji, K.; Nugent, K. Acute Myocardial Injury Following COVID-19 Vaccination: A Case Report and Review of Current Evidence from Vaccine Adverse Events Reporting System Database. J Prim Care Community Health 2021, 12, 21501327211029230. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines — a New Era in Vaccinology. Nat Rev Drug Discov 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Pepini, T.; Pulichino, A.-M.; Carsillo, T.; Carlson, A.L.; Sari-Sarraf, F.; Ramsauer, K.; Debasitis, J.C.; Maruggi, G.; Otten, G.R.; Geall, A.J.; et al. Induction of an IFN-Mediated Antiviral Response by a Self-Amplifying RNA Vaccine: Implications for Vaccine Design. The Journal of Immunology 2017, 198, 4012–4024. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.K.; Jasny, E.; Yoon, H.; Horscroft, N.; Schanen, B.; Geter, T.; Fotin-Mleczek, M.; Petsch, B.; Wittman, V. Adjuvant Effects of a Sequence-Engineered MRNA Vaccine: Translational Profiling Demonstrates Similar Human and Murine Innate Response. Journal of Translational Medicine 2017, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Theofilopoulos, A.N.; Baccala, R.; Beutler, B.; Kono, D.H. TYPE I INTERFERONS (α/β) IN IMMUNITY AND AUTOIMMUNITY. Annual Review of Immunology 2005, 23, 307–335. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Conrad, C.; Tun-Kyi, A.; Homey, B.; Gombert, M.; Boyman, O.; Burg, G.; Liu, Y.-J.; Gilliet, M. Plasmacytoid Predendritic Cells Initiate Psoriasis through Interferon-α Production. Journal of Experimental Medicine 2005, 202, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.; Perincheri, S.; Fleming, T.; Poulson, C.; Tiffany, B.; Bremner, R.M.; Mohanakumar, T. Cutting Edge: Circulating Exosomes with COVID Spike Protein Are Induced by BNT162b2 (Pfizer–BioNTech) Vaccination Prior to Development of Antibodies: A Novel Mechanism for Immune Activation by MRNA Vaccines. The Journal of Immunology 2021, 207, 2405–2410. [Google Scholar] [CrossRef] [PubMed]
- Magen, E.; Mukherjee, S.; Bhattacharya, M.; Detroja, R.; Merzon, E.; Blum, I.; Livoff, A.; Shlapobersky, M.; Baum, G.; Talisman, R.; et al. Clinical and Molecular Characterization of a Rare Case of BNT162b2 MRNA COVID-19 Vaccine-Associated Myositis. Vaccines (Basel) 2022, 10, 1135. [Google Scholar] [CrossRef]
- Röltgen, K.; Nielsen, S.C.A.; Silva, O.; Younes, S.F.; Zaslavsky, M.; Costales, C.; Yang, F.; Wirz, O.F.; Solis, D.; Hoh, R.A.; et al. Immune Imprinting, Breadth of Variant Recognition, and Germinal Center Response in Human SARS-CoV-2 Infection and Vaccination. Cell 2022, 185, 1025–1040.e14. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kase, M.; Sano, H.; Kamijima, R.; Sano, S. Persistent Varicella Zoster Virus Infection Following MRNA COVID-19 Vaccination Was Associated with the Presence of Encoded Spike Protein in the Lesion. Journal of Cutaneous Immunology and Allergy n/a. [CrossRef]
- SARS-CoV-2 MRNA Vaccine (BNT162, PF-07302048); Japanese Government; pp. 1–30.
- Welsh, K.J.; Baumblatt, J.; Chege, W.; Goud, R.; Nair, N. Thrombocytopenia Including Immune Thrombocytopenia after Receipt of MRNA COVID-19 Vaccines Reported to the Vaccine Adverse Event Reporting System (VAERS). Vaccine 2021, 39, 3329–3332. [Google Scholar] [CrossRef] [PubMed]
- Aldén, M.; Olofsson Falla, F.; Yang, D.; Barghouth, M.; Luan, C.; Rasmussen, M.; De Marinis, Y. Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 MRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line. Curr Issues Mol Biol 2022, 44, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Mörz, M. A Case Report: Multifocal Necrotizing Encephalitis and Myocarditis after BNT162b2 MRNA Vaccination against COVID-19. Vaccines 2022, 10, 1651. [Google Scholar] [CrossRef] [PubMed]
- Killingley, B.; Mann, A.J.; Kalinova, M.; Boyers, A.; Goonawardane, N.; Zhou, J.; Lindsell, K.; Hare, S.S.; Brown, J.; Frise, R.; et al. Safety, Tolerability and Viral Kinetics during SARS-CoV-2 Human Challenge in Young Adults. Nat Med 2022, 28, 1031–1041. [Google Scholar] [CrossRef]
- Choi, S.; Lee, S.; Seo, J.-W.; Kim, M.-J.; Jeon, Y.H.; Park, J.H.; Lee, J.K.; Yeo, N.S. Myocarditis-Induced Sudden Death after BNT162b2 MRNA COVID-19 Vaccination in Korea: Case Report Focusing on Histopathological Findings. J Korean Med Sci 2021, 36, e286. [Google Scholar] [CrossRef] [PubMed]
- Letarov, A.V.; Babenko, V.V.; Kulikov, E.E. Free SARS-CoV-2 Spike Protein S1 Particles May Play a Role in the Pathogenesis of COVID-19 Infection. Biochemistry (Mosc) 2021, 86, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ishida, T.; Kiwada, H. Anti-PEG IgM Elicited by Injection of Liposomes Is Involved in the Enhanced Blood Clearance of a Subsequent Dose of PEGylated Liposomes. Journal of Controlled Release 2007, 119, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, B.M. Rare PEG Allergy Triggered Postvaccination Anaphylaxis. JAMA 2021, 325, 1931. [Google Scholar] [CrossRef]
- Cox, F.; Khalib, K.; Conlon, N. PEG That Reaction: A Case Series of Allergy to Polyethylene Glycol. The Journal of Clinical Pharmacology 2021, 61, 832–835. [Google Scholar] [CrossRef]
- Sellaturay, P.; Nasser, S.; Ewan, P. Polyethylene Glycol-Induced Systemic Allergic Reactions (Anaphylaxis). J Allergy Clin Immunol Pract 2021, 9, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Castells, M.C.; Phillips, E.J. Maintaining Safety with SARS-CoV-2 Vaccines. N Engl J Med 2020, NEJMra2035343. [CrossRef]
- Ohgoda, O.; Robinson, I. Toxicological Evaluation of DSPC (1,2-Distearoyl-Sn-Glycero- 3-Phosphocholine). Fundamental Toxicological Sciences 2020, 7. [Google Scholar] [CrossRef]
- Doktorovová, S.; Kovačević, A.B.; Garcia, M.L.; Souto, E.B. Preclinical Safety of Solid Lipid Nanoparticles and Nanostructured Lipid Carriers: Current Evidence from in Vitro and in Vivo Evaluation. Eur J Pharm Biopharm 2016, 108, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S.M.; Simberg, D. Pro-Inflammatory Concerns with Lipid Nanoparticles. Molecular Therapy 2022, 30, 2109–2110. [Google Scholar] [CrossRef] [PubMed]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyártó, B.Z. The MRNA-LNP Platform’s Lipid Nanoparticle Component Used in Preclinical Vaccine Studies Is Highly Inflammatory. iScience 2021, 24. [Google Scholar] [CrossRef] [PubMed]
- Kannemeier, C.; Shibamiya, A.; Nakazawa, F.; Trusheim, H.; Ruppert, C.; Markart, P.; Song, Y.; Tzima, E.; Kennerknecht, E.; Niepmann, M.; et al. Extracellular RNA Constitutes a Natural Procoagulant Cofactor in Blood Coagulation. Proceedings of the National Academy of Sciences 2007, 104, 6388–6393. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Gerriets, T.; Wessels, C.; Walberer, M.; Kostin, S.; Stolz, E.; Zheleva, K.; Hocke, A.; Hippenstiel, S.; Preissner, K.T. Extracellular RNA Mediates Endothelial-Cell Permeability via Vascular Endothelial Growth Factor. Blood 2007, 110, 2457–2465. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Kawai, T. Toll-Like Receptor Signaling Pathways. Front Immunol 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed]
- Wienert, B.; Shin, J.; Zelin, E.; Pestal, K.; Corn, J.E. In Vitro–Transcribed Guide RNAs Trigger an Innate Immune Response via the RIG-I Pathway. PLOS Biology 2018, 16, e2005840. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Eyler, D.E.; Franco, M.K.; Batool, Z.; Wu, M.Z.; Dubuke, M.L.; Dobosz-Bartoszek, M.; Jones, J.D.; Polikanov, Y.S.; Roy, B.; Koutmou, K.S. Pseudouridinylation of MRNA Coding Sequences Alters Translation. Proc Natl Acad Sci U S A 2019, 116, 23068–23074. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Q.; Burgute, B.D.; Tzeng, S.-C.; Jing, C.; Jungers, C.; Zhang, J.; Yan, L.L.; Vierstra, R.D.; Djuranovic, S.; Evans, B.S.; et al. N1-Methylpseudouridine Found within COVID-19 MRNA Vaccines Produces Faithful Protein Products. Cell Rep 2022, 40, 111300. [Google Scholar] [CrossRef]
- Borchardt, E.K.; Martinez, N.M.; Gilbert, W.V. Regulation and Function of RNA Pseudouridylation in Human Cells. Annu Rev Genet 2020, 54, 309–336. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.; Ihara, M.; Kuchino, Y.; Nishimura, S.; Gupta, R.; Woese, C.R.; McCloskey, J.A. Structure of a Modified Nucleoside in Archaebacterial TRNA Which Replaces Ribosylthymine. 1-Methylpseudouridine. J Biol Chem 1982, 257, 3589–3592. [Google Scholar] [CrossRef] [PubMed]
- Andries, O.; Mc Cafferty, S.; De Smedt, S.C.; Weiss, R.; Sanders, N.N.; Kitada, T. N(1)-Methylpseudouridine-Incorporated MRNA Outperforms Pseudouridine-Incorporated MRNA by Providing Enhanced Protein Expression and Reduced Immunogenicity in Mammalian Cell Lines and Mice. J Control Release 2015, 217, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Domazet-Lošo, T. MRNA Vaccines: Why Is the Biology of Retroposition Ignored? Genes (Basel) 2022, 13, 719. [Google Scholar] [CrossRef] [PubMed]
- Vlatkovic, I. Non-Immunotherapy Application of LNP-MRNA: Maximizing Efficacy and Safety. Biomedicines 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, P.S.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the Messenger: Advances in Technologies for Therapeutic MRNA Delivery. Molecular Therapy 2019, 27, 710–728. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yin, L.; Theisen, M.; Zhuo, J.; Siddiqui, S.; Levy, B.; Presnyak, V.; Frassetto, A.; Milton, J.; Salerno, T.; et al. Systemic MRNA Therapy for the Treatment of Fabry Disease: Preclinical Studies in Wild-Type Mice, Fabry Mouse Model, and Wild-Type Non-Human Primates. Am J Hum Genet 2019, 104, 625–637. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Schneller, J.L.; Frassetto, A.; Liang, S.; Zhu, X.; Park, J.-S.; Theisen, M.; Hong, S.-J.; Zhou, J.; Rajendran, R.; et al. Systemic Messenger RNA Therapy as a Treatment for Methylmalonic Acidemia. Cell Rep 2018, 24, 2520. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Berraondo, P.; Jericó, D.; Guey, L.T.; Sampedro, A.; Frassetto, A.; Benenato, K.E.; Burke, K.; Santamaría, E.; Alegre, M.; et al. Systemic Messenger RNA as an Etiological Treatment for Acute Intermittent Porphyria. Nat Med 2018, 24, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Bartelds, G.M.; Krieckaert, C.L.M.; Nurmohamed, M.T.; van Schouwenburg, P.A.; Lems, W.F.; Twisk, J.W.R.; Dijkmans, B.A.C.; Aarden, L.; Wolbink, G.J. Development of Antidrug Antibodies against Adalimumab and Association with Disease Activity and Treatment Failure during Long-Term Follow-Up. JAMA 2011, 305, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Moots, R.J.; Xavier, R.M.; Mok, C.C.; Rahman, M.U.; Tsai, W.-C.; Al-Maini, M.H.; Pavelka, K.; Mahgoub, E.; Kotak, S.; Korth-Bradley, J.; et al. The Impact of Anti-Drug Antibodies on Drug Concentrations and Clinical Outcomes in Rheumatoid Arthritis Patients Treated with Adalimumab, Etanercept, or Infliximab: Results from a Multinational, Real-World Clinical Practice, Non-Interventional Study. PLOS ONE 2017, 12, e0175207. [Google Scholar] [CrossRef] [PubMed]
- Pratt, K.P. Anti-Drug Antibodies: Emerging Approaches to Predict, Reduce or Reverse Biotherapeutic Immunogenicity. Antibodies 2018, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Krishna, M.; Nadler, S.G. Immunogenicity to Biotherapeutics – The Role of Anti-Drug Immune Complexes. Front Immunol 2016, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.B. Mechanisms of Adverse Drug Reactions to Biologics. Handb Exp Pharmacol 2010, 453–474. [Google Scholar] [CrossRef] [PubMed]
- Pichler, W.J. Adverse Side-Effects to Biological Agents. Allergy 2006, 61, 912–920. [Google Scholar] [CrossRef]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schäfer, S.; Zhong, J.; et al. Class Switch towards Non-Inflammatory, Spike-Specific IgG4 Antibodies after Repeated SARS-CoV-2 MRNA Vaccination. Sci Immunol 2022, eade2798. [Google Scholar] [CrossRef] [PubMed]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG Subclasses and Allotypes: From Structure to Effector Functions. Front Immunol 2014, 5, 520. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, R.; Roth-Walter, F.; Ohradanova-Repic, A.; Flicker, S.; Hufnagl, K.; Fischer, M.B.; Stockinger, H.; Jensen-Jarolim, E. IgG4 Drives M2a Macrophages to a Regulatory M2b-like Phenotype: Potential Implication in Immune Tolerance. Allergy 2019, 74, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Sathish, J.G.; Sethu, S.; Bielsky, M.-C.; de Haan, L.; French, N.S.; Govindappa, K.; Green, J.; Griffiths, C.E.M.; Holgate, S.; Jones, D.; et al. Challenges and Approaches for the Development of Safer Immunomodulatory Biologics. Nat Rev Drug Discov 2013, 12, 306–324. [Google Scholar] [CrossRef] [PubMed]
- Banugaria, S.G.; Prater, S.N.; Ng, Y.-K.; Kobori, J.A.; Finkel, R.S.; Ladda, R.L.; Chen, Y.-T.; Rosenberg, A.S.; Kishnani, P.S. The Impact of Antibodies on Clinical Outcomes in Diseases Treated with Therapeutic Protein: Lessons Learned from Infantile Pompe Disease. Genet Med 2011, 13, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. MRNA Vaccines for Infectious Diseases: Principles, Delivery and Clinical Translation. Nat Rev Drug Discov 2021, 20, 817–838. [Google Scholar] [CrossRef] [PubMed]
- Suhr, O.B.; Coelho, T.; Buades, J.; Pouget, J.; Conceicao, I.; Berk, J.; Schmidt, H.; Waddington-Cruz, M.; Campistol, J.M.; Bettencourt, B.R.; et al. Efficacy and Safety of Patisiran for Familial Amyloidotic Polyneuropathy: A Phase II Multi-Dose Study. Orphanet J Rare Dis 2015, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Balwani, M.; Sardh, E.; Ventura, P.; Peiró, P.A.; Rees, D.C.; Stölzel, U.; Bissell, D.M.; Bonkovsky, H.L.; Windyga, J.; Anderson, K.E.; et al. Phase 3 Trial of RNAi Therapeutic Givosiran for Acute Intermittent Porphyria. N Engl J Med 2020, 382, 2289–2301. [Google Scholar] [CrossRef] [PubMed]
- Alberer, M.; Gnad-Vogt, U.; Hong, H.S.; Mehr, K.T.; Backert, L.; Finak, G.; Gottardo, R.; Bica, M.A.; Garofano, A.; Koch, S.D.; et al. Safety and Immunogenicity of a MRNA Rabies Vaccine in Healthy Adults: An Open-Label, Non-Randomised, Prospective, First-in-Human Phase 1 Clinical Trial. Lancet 2017, 390, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, C.; Leroux-Roels, I.; Huang, K.B.; Bica, M.A.; Loeliger, E.; Schoenborn-Kellenberger, O.; Walz, L.; Leroux-Roels, G.; von Sonnenburg, F.; Oostvogels, L. Proof-of-Concept of a Low-Dose Unmodified MRNA-Based Rabies Vaccine Formulated with Lipid Nanoparticles in Human Volunteers: A Phase 1 Trial. Vaccine 2021, 39, 1310–1318. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.A.; Fuhr, R.; Smolenov, I.; Mick Ribeiro, A.; Panther, L.; Watson, M.; Senn, J.J.; Smith, M.; Almarsson, Ӧrn; Pujar, H.S.; et al. MRNA Vaccines against H10N8 and H7N9 Influenza Viruses of Pandemic Potential Are Immunogenic and Well Tolerated in Healthy Adults in Phase 1 Randomized Clinical Trials. Vaccine 2019, 37, 3326–3334. [CrossRef] [PubMed]
- Bahl, K.; Senn, J.J.; Yuzhakov, O.; Bulychev, A.; Brito, L.A.; Hassett, K.J.; Laska, M.E.; Smith, M.; Almarsson, Ö.; Thompson, J.; et al. Preclinical and Clinical Demonstration of Immunogenicity by MRNA Vaccines against H10N8 and H7N9 Influenza Viruses. Molecular Therapy 2017, 25, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.M.; Routy, J.-P.; Welles, S.; DeBenedette, M.; Tcherepanova, I.; Angel, J.B.; Asmuth, D.M.; Stein, D.K.; Baril, J.-G.; McKellar, M.; et al. Dendritic Cell Immunotherapy for HIV-1 Infection Using Autologous HIV-1 RNA: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J Acquir Immune Defic Syndr 2016, 72, 31–38. [Google Scholar] [CrossRef] [PubMed]
- de Jong, W.; Aerts, J.; Allard, S.; Brander, C.; Buyze, J.; Florence, E.; van Gorp, E.; Vanham, G.; Leal, L.; Mothe, B.; et al. IHIVARNA Phase IIa, a Randomized, Placebo-Controlled, Double-Blinded Trial to Evaluate the Safety and Immunogenicity of IHIVARNA-01 in Chronically HIV-Infected Patients under Stable Combined Antiretroviral Therapy. Trials 2019, 20, 361. [Google Scholar] [CrossRef] [PubMed]
- Leal, L.; Guardo, A.C.; Morón-López, S.; Salgado, M.; Mothe, B.; Heirman, C.; Pannus, P.; Vanham, G.; van den Ham, H.J.; Gruters, R.; et al. Phase I Clinical Trial of an Intranodally Administered MRNA-Based Therapeutic Vaccine against HIV-1 Infection. AIDS 2018, 32, 2533–2545. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, R.T.; Kwon, D.S.; Macklin, E.A.; Shopis, J.R.; McLean, A.P.; McBrine, N.; Flynn, T.; Peter, L.; Sbrolla, A.; Kaufmann, D.E.; et al. Immunization of HIV-1-Infected Persons With Autologous Dendritic Cells Transfected With MRNA Encoding HIV-1 Gag and Nef: Results of a Randomized, Placebo-Controlled Clinical Trial. JAIDS Journal of Acquired Immune Deficiency Syndromes 2016, 71, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Oehm, P.; Derhovanessian, E.; Jabulowsky, R.A.; Vormehr, M.; Gold, M.; Maurus, D.; Schwarck-Kokarakis, D.; Kuhn, A.N.; Omokoko, T.; et al. An RNA Vaccine Drives Immunity in Checkpoint-Inhibitor-Treated Melanoma. Nature 2020, 585, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Papachristofilou, A.; Hipp, M.M.; Klinkhardt, U.; Früh, M.; Sebastian, M.; Weiss, C.; Pless, M.; Cathomas, R.; Hilbe, W.; Pall, G.; et al. Phase Ib Evaluation of a Self-Adjuvanted Protamine Formulated MRNA-Based Active Cancer Immunotherapy, BI1361849 (CV9202), Combined with Local Radiation Treatment in Patients with Stage IV Non-Small Cell Lung Cancer. J Immunother Cancer 2019, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Eigentler, T.; Bauernfeind, F.G.; Becker, J.C.; Brossart, P.; Fluck, M.; Heinzerling, L.; Krauss, J.; Mohr, P.; Ochsenreither, S.; Schreiber, J.S.; et al. A Phase I Dose-Escalation and Expansion Study of Intratumoral CV8102 as Single-Agent or in Combination with Anti-PD-1 Antibodies in Patients with Advanced Solid Tumors. JCO 2020, 38, 3096–3096. [Google Scholar] [CrossRef]
- Doener, F.; Hong, H.S.; Meyer, I.; Tadjalli-Mehr, K.; Daehling, A.; Heidenreich, R.; Koch, S.D.; Fotin-Mleczek, M.; Gnad-Vogt, U. RNA-Based Adjuvant CV8102 Enhances the Immunogenicity of a Licensed Rabies Vaccine in a First-in-Human Trial. Vaccine 2019, 37, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Translate Bio Announces Results from Second Interim Data Analysis from Ongoing Phase 1/2 Clinical Trial of MRT5005 in Patients with Cystic Fibrosis (CF). Available online: https://www.biospace.com/article/translate-bio-announces-results-from-second-interim-data-analysis-from-ongoing-phase-1-2-clinical-trial-of-mrt5005-in-patients-with-cystic-fibrosis-cf-/ (accessed on 13 October 2022).
- Burris, H.A.; Patel, M.R.; Cho, D.C.; Clarke, J.M.; Gutierrez, M.; Zaks, T.Z.; Frederick, J.; Hopson, K.; Mody, K.; Binanti-Berube, A.; et al. A Phase I Multicenter Study to Assess the Safety, Tolerability, and Immunogenicity of MRNA-4157 Alone in Patients with Resected Solid Tumors and in Combination with Pembrolizumab in Patients with Unresectable Solid Tumors. JCO 2019, 37, 2523–2523. [Google Scholar] [CrossRef]
- Costello, C.L.; Gregory, T.K.; Ali, S.A.; Berdeja, J.G.; Patel, K.K.; Shah, N.D.; Ostertag, E.; Martin, C.; Ghoddusi, M.; Shedlock, D.J.; et al. Phase 2 Study of the Response and Safety of P-Bcma-101 CAR-T Cells in Patients with Relapsed/Refractory (r/r) Multiple Myeloma (MM) (PRIME). Blood 2019, 134, 3184–3184. [Google Scholar] [CrossRef]
- Gregory, T.; Cohen, A.D.; Costello, C.L.; Ali, S.A.; Berdeja, J.G.; Ostertag, E.M.; Martin, C.; Shedlock, D.J.; Resler, M.L.; Spear, M.A.; et al. Efficacy and Safety of P-Bcma-101 CAR-T Cells in Patients with Relapsed/Refractory (r/r) Multiple Myeloma (MM). Blood 2018, 132, 1012. [Google Scholar] [CrossRef]
- Anttila, V.; Saraste, A.; Knuuti, J.; Jaakkola, P.; Hedman, M.; Svedlund, S.; Lagerström-Fermér, M.; Kjaer, M.; Jeppsson, A.; Gan, L.-M. Synthetic MRNA Encoding VEGF-A in Patients Undergoing Coronary Artery Bypass Grafting: Design of a Phase 2a Clinical Trial. Mol Ther Methods Clin Dev 2020, 18, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Late-Breaking Science Abstracts and Featured Science Abstracts From the American Heart Association’s Scientific Sessions 2021 and Late-Breaking Abstracts in Resuscitation Science From the Resuscitation Science Symposium 2021. Circulation 2021, 144, e564–e593. [CrossRef] [PubMed]
- Gan, L.-M.; Lagerström-Fermér, M.; Carlsson, L.G.; Arfvidsson, C.; Egnell, A.-C.; Rudvik, A.; Kjaer, M.; Collén, A.; Thompson, J.D.; Joyal, J.; et al. Intradermal Delivery of Modified MRNA Encoding VEGF-A in Patients with Type 2 Diabetes. Nat Commun 2019, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- de Lusignan, S.; Damaso, S.; Ferreira, F.; Byford, R.; McGee, C.; Pathirannehelage, S.; Shende, V.; Yonova, I.; Schmidt, A.; Schuind, A.; et al. Brand-Specific Enhanced Safety Surveillance of GSK’s Fluarix Tetra Seasonal Influenza Vaccine in England: 2017/2018 Season. Hum Vaccin Immunother 2020, 16, 1762–1771. [Google Scholar] [CrossRef] [PubMed]
- Crescioli, S.; Correa, I.; Karagiannis, P.; Davies, A.M.; Sutton, B.J.; Nestle, F.O.; Karagiannis, S.N. IgG4 Characteristics and Functions in Cancer Immunity. Curr Allergy Asthma Rep 2016, 16, 7. [Google Scholar] [CrossRef] [PubMed]
- Schlaudecker, E.P.; McNeal, M.M.; Dodd, C.N.; Ranz, J.B.; Steinhoff, M.C. Pregnancy Modifies the Antibody Response to Trivalent Influenza Immunization. J Infect Dis 2012, 206, 1670–1673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lu, H.; Peng, L.; Zhou, J.; Wang, M.; Li, J.; Liu, Z.; Zhang, W.; Zhao, Y.; Zeng, X.; et al. The Role of PD-1/PD-Ls in the Pathogenesis of IgG4-Related Disease. Rheumatology 2022, 61, 815–825. [Google Scholar] [CrossRef]
- Pfizer-BioNTech COVID-19 Vaccine, COMIRNATY® (Tozinameran). Available online: https://www.who.int/publications/m/item/comirnaty-covid-19-mrna-vaccine (accessed on 30 December 2022).
- Moderna MRNA-1273, COVID-19 Vaccine. Available online: https://www.who.int/publications/m/item/moderna-covid-19-vaccine-(mrna-1273) (accessed on 30 December 2022).
- Chakraborty, C.; Bhattacharya, M.; Sharma, A.R. Present Variants of Concern and Variants of Interest of Severe Acute Respiratory Syndrome Coronavirus 2: Their Significant Mutations in S-Glycoprotein, Infectivity, Re-Infectivity, Immune Escape and Vaccines Activity. Reviews in Medical Virology 2022, 32, e2270. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Bhattacharya, M.; Lee, S.-S. A Detailed Overview of Immune Escape, Antibody Escape, Partial Vaccine Escape of SARS-CoV-2 and Their Emerging Variants With Escape Mutations. Front Immunol 2022, 13, 801522. [Google Scholar] [CrossRef] [PubMed]
- Zhong, N.; Zheng, B.; Li, Y.; Poon, L.; Xie, Z.; Chan, K.; Li, P.; Tan, S.; Chang, Q.; Xie, J.; et al. Epidemiology and Cause of Severe Acute Respiratory Syndrome (SARS) in Guangdong, People’s Republic of China, in February, 2003. The Lancet 2003, 362, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Gastañaduy, P.A. Update: Severe Respiratory Illness Associated with Middle East Respiratory Syndrome Coronavirus (MERS-CoV) — Worldwide, 2012–2013. MMWR Morb Mortal Wkly Rep 2013, 62, 480–483. [Google Scholar]
- Li, Y.-D.; Chi, W.-Y.; Su, J.-H.; Ferrall, L.; Hung, C.-F.; Wu, T.-C. Coronavirus Vaccine Development: From SARS and MERS to COVID-19. Journal of Biomedical Science 2020, 27, 104. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.-T.; Sbrana, E.; Iwata-Yoshikawa, N.; Newman, P.C.; Garron, T.; Atmar, R.L.; Peters, C.J.; Couch, R.B. Immunization with SARS Coronavirus Vaccines Leads to Pulmonary Immunopathology on Challenge with the SARS Virus. PLoS One 2012, 7, e35421. [Google Scholar] [CrossRef] [PubMed]
- Weingartl, H.; Czub, M.; Czub, S.; Neufeld, J.; Marszal, P.; Gren, J.; Smith, G.; Jones, S.; Proulx, R.; Deschambault, Y.; et al. Immunization with Modified Vaccinia Virus Ankara-Based Recombinant Vaccine against Severe Acute Respiratory Syndrome Is Associated with Enhanced Hepatitis in Ferrets. J Virol 2004, 78, 12672–12676. [Google Scholar] [CrossRef] [PubMed]
- Bolles, M.; Deming, D.; Long, K.; Agnihothram, S.; Whitmore, A.; Ferris, M.; Funkhouser, W.; Gralinski, L.; Totura, A.; Heise, M.; et al. A Double-Inactivated Severe Acute Respiratory Syndrome Coronavirus Vaccine Provides Incomplete Protection in Mice and Induces Increased Eosinophilic Proinflammatory Pulmonary Response upon Challenge. J Virol 2011, 85, 12201–12215. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wei, Q.; Lin, Q.; Fang, J.; Wang, H.; Kwok, H.; Tang, H.; Nishiura, K.; Peng, J.; Tan, Z.; et al. Anti-Spike IgG Causes Severe Acute Lung Injury by Skewing Macrophage Responses during Acute SARS-CoV Infection. JCI Insight 2019, 4, 123158. [Google Scholar] [CrossRef] [PubMed]
- Doremalen, N. van; Haddock, E.; Feldmann, F.; Meade-White, K.; Bushmaker, T.; Fischer, R.J.; Okumura, A.; Hanley, P.W.; Saturday, G.; Edwards, N.J.; et al. A Single Dose of ChAdOx1 MERS Provides Broad Protective Immunity against a Variety of MERS-CoV Strains 2020, 2020. 04.13.03 6293.
- Lee, W.S.; Wheatley, A.K.; Kent, S.J.; DeKosky, B.J. Antibody-Dependent Enhancement and SARS-CoV-2 Vaccines and Therapies. Nat Microbiol 2020, 5, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Cheng, Y.; Ling, R.; Dai, Y.; Huang, B.; Huang, W.; Zhang, S.; Jiang, Y. Antibody-Dependent Enhancement of Coronavirus. Int J Infect Dis 2020, 100, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Ma, Z.; Li, Y.; Pang, Z.; Xiao, S. Antibody Dependent Enhancement: Unavoidable Problems in Vaccine Development. Adv Immunol 2021, 151, 99–133. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Zuno, G.A.; Matuz-Flores, M.G.; González-Estevez, G.; Nicoletti, F.; Turrubiates-Hernández, F.J.; Mangano, K.; Muñoz-Valle, J.F. A Review: Antibody-Dependent Enhancement in COVID-19: The Not so Friendly Side of Antibodies. Int J Immunopathol Pharmacol 2021, 35, 20587384211050200. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Smatti, M.K.; Ouhtit, A.; Cyprian, F.S.; Almaslamani, M.A.; Thani, A.A.; Yassine, H.M. Antibody-Dependent Enhancement (ADE) and the Role of Complement System in Disease Pathogenesis. Mol Immunol 2022, 152, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Ricke, D.O. Two Different Antibody-Dependent Enhancement (ADE) Risks for SARS-CoV-2 Antibodies. Front Immunol 2021, 12, 640093. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Shang, J.; Sun, S.; Tai, W.; Chen, J.; Geng, Q.; He, L.; Chen, Y.; Wu, J.; Shi, Z.; et al. Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry. J Virol 2020, 94, e02015–19. [Google Scholar] [CrossRef]
- Shimizu, J.; Sasaki, T.; Koketsu, R.; Morita, R.; Yoshimura, Y.; Murakami, A.; Saito, Y.; Kusunoki, T.; Samune, Y.; Nakayama, E.E.; et al. Reevaluation of Antibody-Dependent Enhancement of Infection in Anti-SARS-CoV-2 Therapeutic Antibodies and MRNA-Vaccine Antisera Using FcR- and ACE2-Positive Cells. Sci Rep 2022, 12, 15612. [Google Scholar] [CrossRef] [PubMed]
- Lurie, N.; Saville, M.; Hatchett, R.; Halton, J. Developing Covid-19 Vaccines at Pandemic Speed. N Engl J Med 2020, 382, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- London, A.J.; Kimmelman, J. Against Pandemic Research Exceptionalism. Science 2020, 368, 476–477. [Google Scholar] [CrossRef] [PubMed]
- Tizard, I.R. Vaccination against Coronaviruses in Domestic Animals. Vaccine 2020, 38, 5123–5130. [Google Scholar] [CrossRef] [PubMed]
- Olsen, C.W.; Corapi, W.V.; Ngichabe, C.K.; Baines, J.D.; Scott, F.W. Monoclonal Antibodies to the Spike Protein of Feline Infectious Peritonitis Virus Mediate Antibody-Dependent Enhancement of Infection of Feline Macrophages. J Virol 1992, 66, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Hohdatsu, T.; Yamada, M.; Tominaga, R.; Makino, K.; Kida, K.; Koyama, H. Antibody-Dependent Enhancement of Feline Infectious Peritonitis Virus Infection in Feline Alveolar Macrophages and Human Monocyte Cell Line U937 by Serum of Cats Experimentally or Naturally Infected with Feline Coronavirus. J Vet Med Sci 1998, 60, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Nakaguchi, M.; Doki, T.; Hohdatsu, T. Antibody-Dependent Enhancement of Serotype II Feline Enteric Coronavirus Infection in Primary Feline Monocytes. Arch Virol 2017, 162, 3339–3345. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, Z.; Simpson, B.; Donald Reynolds, D.V.M. Antibody Dependent Enhancement of Infectious Bronchitis Virus in Poultry. UCARE Research Products 2022. [Google Scholar]
- H, T.; D, P.; Ra, G.; Vl, van S. ; Fw, van G.; J, Z.; Ks, J. Infectious Bronchitis Virus Subpopulations in Vaccinated Chickens after Challenge. Avian diseases 2012, 56. [Google Scholar] [CrossRef] [PubMed]
- Brandão, P.E.; Berg, M.; Silva, S.O.S.; Taniwaki, S.A. Emergence of Avian Coronavirus Escape Mutants Under Suboptimal Antibody Titers. J Mol Evol 2022, 90, 176–181. [Google Scholar] [CrossRef] [PubMed]
- F, B.; Ss, A.; Mh, B.; H, M.; Ar, O. Progress and Challenges toward the Development of Vaccines against Avian Infectious Bronchitis. Journal of immunology research 2015, 2015. [Google Scholar] [CrossRef]
- Eldemery, F.; Li, Y.; Yu, Q.; van Santen, V.L.; Toro, H. Infectious Bronchitis Virus S2 of 4/91 Expressed from Recombinant Virus Does Not Protect Against Ark-Type Challenge. Avian Dis 2017, 61, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, R.; Chan, J.; Prabakaran, M. Vaccines against Major Poultry Viral Diseases: Strategies to Improve the Breadth and Protective Efficacy. Viruses 2022, 14, 1195. [Google Scholar] [CrossRef] [PubMed]
- Shao, G.; Chen, T.; Feng, K.; Zhao, Q.; Zhang, X.; Li, H.; Lin, W.; Xie, Q. Efficacy of Commercial Polyvalent Avian Infectious Bronchitis Vaccines against Chinese QX-like and TW-like Strain via Different Vaccination Strategies. Poult Sci 2020, 99, 4786–4794. [Google Scholar] [CrossRef] [PubMed]
- Sjaak de Wit, J.J.; Cook, J.K.A.; van der Heijden, H.M.J.F. Infectious Bronchitis Virus Variants: A Review of the History, Current Situation and Control Measures. Avian Pathol 2011, 40, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Severe Acute Respiratory Syndrome Vaccine Development: Experiences of Vaccination against Avian Infectious Bronchitis Coronavirus. Avian Pathol 2003, 32, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Legnardi, M.; Tucciarone, C.M.; Franzo, G.; Cecchinato, M. Infectious Bronchitis Virus Evolution, Diagnosis and Control. Vet Sci 2020, 7, E79. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.B.; Holladay, J.A.; Cave, J.S. A Neurologic Syndrome Associated with Use of a Canine Coronavirus-Parvovirus Vaccine in Dogs. The Compendium on continuing education for the practicing veterinarian (USA) 1986. [Google Scholar]
- Martin, M. Canine Coronavirus Enteritis and a Recent Outbreak Following Modified Live Virus Vaccination. Compendium on continuing education for the practicing veterinarian 1985, 7, 1012–1017. [Google Scholar]
- Pratelli, A.; Tinelli, A.; Decaro, N.; Martella, V.; Camero, M.; Tempesta, M.; Martini, M.; Carmichael, L.E.; Buonavoglia, C. Safety and Efficacy of a Modified-Live Canine Coronavirus Vaccine in Dogs. Vet Microbiol 2004, 99, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, A. High-cell-passage Canine Coronavirus Vaccine Providing Sterilising Immunity. J Small Anim Pract 2007, 48, 574–578. [Google Scholar] [CrossRef]
- Cho, K.-O.; Hasoksuz, M.; Nielsen, P.R.; Chang, K.-O.; Lathrop, S.; Saif, L.J. Cross-Protection Studies between Respiratory and Calf Diarrhea and Winter Dysentery Coronavirus Strains in Calves and RT-PCR and Nested PCR for Their Detection. Arch Virol 2001, 146, 2401–2419. [Google Scholar] [CrossRef] [PubMed]
- Heckert, R.A.; Saif, L.J.; Hoblet, K.H.; Agnes, A.G. A Longitudinal Study of Bovine Coronavirus Enteric and Respiratory Infections in Dairy Calves in Two Herds in Ohio. Vet Microbiol 1990, 22, 187–201. [Google Scholar] [CrossRef]
- Fulton, R.W.; d’Offay, J.M.; Landis, C.; Miles, D.G.; Smith, R.A.; Saliki, J.T.; Ridpath, J.F.; Confer, A.W.; Neill, J.D.; Eberle, R.; et al. Detection and Characterization of Viruses as Field and Vaccine Strains in Feedlot Cattle with Bovine Respiratory Disease. Vaccine 2016, 34, 3478–3492. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Bruszewski, J.; Smalling, R.; Browne, J.K. Studies of TGEV Spike Protein Gp195 Expressed in E. Coli and by a TGE-Vaccinia Virus Recombinant. Adv Exp Med Biol 1985, 185, 63–82. [Google Scholar] [CrossRef]
- Gómez, N.; Wigdorovitz, A.; Castañón, S.; Gil, F.; Ordá, R.; Borca, M.V.; Escribano, J.M. Oral Immunogenicity of the Plant Derived Spike Protein from Swine-Transmissible Gastroenteritis Coronavirus. Arch Virol 2000, 145, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- Lamphear, B.J.; Streatfield, S.J.; Jilka, J.M.; Brooks, C.A.; Barker, D.K.; Turner, D.D.; Delaney, D.E.; Garcia, M.; Wiggins, B.; Woodard, S.L.; et al. Delivery of Subunit Vaccines in Maize Seed. J Control Release 2002, 85, 169–180. [Google Scholar] [CrossRef]
- Gerdts, V.; Zakhartchouk, A. Vaccines for Porcine Epidemic Diarrhea Virus and Other Swine Coronaviruses. Vet Microbiol 2017, 206, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Al-Abdallat, M.M.; Payne, D.C.; Alqasrawi, S.; Rha, B.; Tohme, R.A.; Abedi, G.R.; Al Nsour, M.; Iblan, I.; Jarour, N.; Farag, N.H.; et al. Hospital-Associated Outbreak of Middle East Respiratory Syndrome Coronavirus: A Serologic, Epidemiologic, and Clinical Description. Clinical Infectious Diseases 2014, 59, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Yoon, I.-K.; Kim, J.H. First Clinical Trial of a MERS Coronavirus DNA Vaccine. The Lancet Infectious Diseases 2019, 19, 924–925. [Google Scholar] [CrossRef] [PubMed]
- Modjarrad, K.; Roberts, C.C.; Mills, K.T.; Castellano, A.R.; Paolino, K.; Muthumani, K.; Reuschel, E.L.; Robb, M.L.; Racine, T.; Oh, M.; et al. Safety and Immunogenicity of an Anti-Middle East Respiratory Syndrome Coronavirus DNA Vaccine: A Phase 1, Open-Label, Single-Arm, Dose-Escalation Trial. The Lancet Infectious Diseases 2019, 19, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-T.; Zhang, J.-S.; Su, N.; Xu, J.-G.; Wang, N.; Chen, J.-T.; Chen, X.; Liu, Y.-X.; Gao, H.; Jia, Y.-P.; et al. Safety and Immunogenicity from a Phase I Trial of Inactivated Severe Acute Respiratory Syndrome Coronavirus Vaccine. Antivir Ther 2007, 12, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.E.; Louder, M.K.; Holman, L.A.; Gordon, I.J.; Enama, M.E.; Larkin, B.D.; Andrews, C.A.; Vogel, L.; Koup, R.A.; Roederer, M.; et al. A SARS DNA Vaccine Induces Neutralizing Antibody and Cellular Immune Responses in Healthy Adults in a Phase I Clinical Trial. Vaccine 2008, 26, 6338–6343. [Google Scholar] [CrossRef] [PubMed]
- Koch, T.; Dahlke, C.; Fathi, A.; Kupke, A.; Krähling, V.; Okba, N.M.A.; Halwe, S.; Rohde, C.; Eickmann, M.; Volz, A.; et al. Safety and Immunogenicity of a Modified Vaccinia Virus Ankara Vector Vaccine Candidate for Middle East Respiratory Syndrome: An Open-Label, Phase 1 Trial. The Lancet Infectious Diseases 2020, 20, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Fathi, A.; Dahlke, C.; Krähling, V.; Kupke, A.; Okba, N.M.A.; Raadsen, M.P.; Heidepriem, J.; Müller, M.A.; Paris, G.; Lassen, S.; et al. Increased Neutralization and IgG Epitope Identification after MVA-MERS-S Booster Vaccination against Middle East Respiratory Syndrome. Nat Commun 2022, 13, 4182. [Google Scholar] [CrossRef] [PubMed]
- Folegatti, P.M.; Bittaye, M.; Flaxman, A.; Lopez, F.R.; Bellamy, D.; Kupke, A.; Mair, C.; Makinson, R.; Sheridan, J.; Rohde, C.; et al. Safety and Immunogenicity of a Candidate Middle East Respiratory Syndrome Coronavirus Viral-Vectored Vaccine: A Dose-Escalation, Open-Label, Non-Randomised, Uncontrolled, Phase 1 Trial. The Lancet Infectious Diseases 2020, 20, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Bosaeed, M.; Balkhy, H.H.; Almaziad, S.; Aljami, H.A.; Alhatmi, H.; Alanazi, H.; Alahmadi, M.; Jawhary, A.; Alenazi, M.W.; Almasoud, A.; et al. Safety and Immunogenicity of ChAdOx1 MERS Vaccine Candidate in Healthy Middle Eastern Adults (MERS002): An Open-Label, Non-Randomised, Dose-Escalation, Phase 1b Trial. Lancet Microbe 2022, 3, e11–e20. [Google Scholar] [CrossRef] [PubMed]
- Buzhdygan, T.P.; DeOre, B.J.; Baldwin-Leclair, A.; Bullock, T.A.; McGary, H.M.; Khan, J.A.; Razmpour, R.; Hale, J.F.; Galie, P.A.; Potula, R.; et al. The SARS-CoV-2 Spike Protein Alters Barrier Function in 2D Static and 3D Microfluidic in-Vitro Models of the Human Blood-Brain Barrier. Neurobiol Dis 2020, 146, 105131. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, C.B.; Zhang, L.; Bhushan, A.; Swanson, B.; Zhang, L.; Mamede, J.I.; Voigt, R.M.; Shaikh, M.; Engen, P.A.; Keshavarzian, A. The SARS-CoV-2 S1 Spike Protein Promotes MAPK and NF-KB Activation in Human Lung Cells and Inflammatory Cytokine Production in Human Lung and Intestinal Epithelial Cells. Microorganisms 2022, 10, 1996. [Google Scholar] [CrossRef] [PubMed]
- Bhargavan, B.; Kanmogne, G.D. SARS-CoV-2 Spike Proteins and Cell-Cell Communication Inhibits TFPI and Induces Thrombogenic Factors in Human Lung Microvascular Endothelial Cells and Neutrophils: Implications for COVID-19 Coagulopathy Pathogenesis. Int J Mol Sci 2022, 23, 10436. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-Y.; Park, J.H.; Jo, C.; Kim, K.-C.; Koh, Y.H. SARS-CoV-2 Spike S1 Subunit Protein-Mediated Increase of Beta-Secretase 1 (BACE1) Impairs Human Brain Vessel Cells. Biochem Biophys Res Commun 2022, 626, 66–71. [Google Scholar] [CrossRef]
- Hsu, A.C.-Y.; Wang, G.; Reid, A.T.; Veerati, P.C.; Pathinayake, P.S.; Daly, K.; Mayall, J.R.; Hansbro, P.M.; Horvat, J.C.; Wang, F.; et al. SARS-CoV-2 Spike Protein Promotes Hyper-Inflammatory Response That Can Be Ameliorated by Spike-Antagonistic Peptide and FDA-Approved ER Stress and MAP Kinase Inhibitors in Vitro 2020, 2020. 09.30.31 7818.
- Khan, S.; Shafiei, M.S.; Longoria, C.; Schoggins, J.W.; Savani, R.C.; Zaki, H. SARS-CoV-2 Spike Protein Induces Inflammation via TLR2-Dependent Activation of the NF-ΚB Pathway. Elife 2021, 10, e68563. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Zhang, J.; Schiavon, C.R.; He, M.; Chen, L.; Shen, H.; Zhang, Y.; Yin, Q.; Cho, Y.; Andrade, L.; et al. SARS-CoV-2 Spike Protein Impairs Endothelial Function via Downregulation of ACE 2. Circulation Research 2021, 128, 1323–1326. [Google Scholar] [CrossRef]
- Nyström, S.; Hammarström, P. Amyloidogenesis of SARS-CoV-2 Spike Protein. J Am Chem Soc 2022, 144, 8945–8950. [Google Scholar] [CrossRef]
- Sattar, S.; Kabat, J.; Jerome, K.; Feldmann, F.; Bailey, K.; Mehedi, M. Nuclear Translocation of Spike MRNA and Protein Is a Novel Pathogenic Feature of SARS-CoV-2 2022, 2022. 09.27.50 9633.
- Singh, N.; Bharara Singh, A. S2 Subunit of SARS-NCoV-2 Interacts with Tumor Suppressor Protein P53 and BRCA: An in Silico Study. Transl Oncol 2020, 13, 100814. [Google Scholar] [CrossRef]
- Kanduc, D. From SARS-CoV-2 to Myocarditis and Sudden Death via Molecular Mimicry and Immunologic Memory. Current Practice in Medical Science Vol. 4 2022, 129–139. [Google Scholar] [CrossRef]
- Nunez-Castilla, J.; Stebliankin, V.; Baral, P.; Balbin, C.A.; Sobhan, M.; Cickovski, T.; Mondal, A.M.; Narasimhan, G.; Chapagain, P.; Mathee, K.; et al. Potential Autoimmunity Resulting from Molecular Mimicry between SARS-CoV-2 Spike and Human Proteins. Viruses 2022, 14, 1415. [Google Scholar] [CrossRef] [PubMed]
- Föhse, K.; Geckin, B.; Overheul, G.; van de Maat, J.; Kilic, G.; Bulut, O.; Dijkstra, H.; Lemmers, H.; Sarlea, A.; Reijnders, M.; et al. The BNT162b2 MRNA Vaccine against SARS-CoV-2 Reprograms Both Adaptive and Innate Immune Response 2021.
- Muglia, J.J.; DiGiovanna, J.J. Phase 1 Clinical Trials. J Cutan Med Surg 1998, 2, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Arndt, L.; Schwarz, T.; Loyal, L.; Henze, L.; Kruse, B.; Dingeldey, M.; Gürcan, K.; Uyar-Aydin, Z.; Müller, M.A.; Drosten, C.; et al. Cutting Edge: Serum but Not Mucosal Antibody Responses Are Associated with Pre-Existing SARS-CoV-2 Spike Cross-Reactive CD4+ T Cells Following BNT162b2 Vaccination in the Elderly. J Immunol 2022, 208, 1001–1005. [Google Scholar] [CrossRef]
- Chau, N.V.V.; Ngoc, N.M.; Nguyet, L.A.; Quang, V.M.; Ny, N.T.H.; Khoa, D.B.; Phong, N.T.; Toan, L.M.; Hong, N.T.T.; Tuyen, N.T.K.; et al. Transmission of SARS-CoV-2 Delta Variant Among Vaccinated Healthcare Workers, Vietnam 2021.
- Pezzullo, A.M.; Axfors, C.; Contopoulos-Ioannidis, D.G.; Apostolatos, A.; Ioannidis, J.P.A. Age-Stratified Infection Fatality Rate of COVID-19 in the Non-Elderly Population. Environmental Research 2023, 216, 114655. [Google Scholar] [CrossRef] [PubMed]
- Kedmi, R.; Ben-Arie, N.; Peer, D. The Systemic Toxicity of Positively Charged Lipid Nanoparticles and the Role of Toll-like Receptor 4 in Immune Activation. Biomaterials 2010, 31, 6867–6875. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Summary of human trials of non-covid-19 coronavirus vaccines. Adapted from [100].
Table 1.
Summary of human trials of non-covid-19 coronavirus vaccines. Adapted from [100].
| Platform | Vaccine | Group | Status | Severe adverse events | NCT ID | Study |
|---|---|---|---|---|---|---|
| SARS Vaccine Clinical Trials | ||||||
| Inactivated virus | Inactivated SARS-CoV vaccine (ISCV) | Sinovac | Phase I, completed | [0/24, 0%] | No NCT ID | [144] |
| DNA vaccine | VRC-SRSDNA015-00-VP | NIAID | Phase I, completed | [0/9, 0%] | NCT00099463 | [145] |
| MERS Vaccine Clinical Trials | ||||||
| DNA vaccine | GLS-5300 (INO-4700) | GeneOne Life Science/Inovio Pharmaceuticals/International Vaccine Institute | Phase I, completed | [0/75, 0%] *Infections in 36% of participants |
NCT02670187 | [143] |
| DNA vaccine | GLS-5300 (INO-4700) | GeneOne Life Science/Inovio Pharmaceuticals/International Vaccine Institute | Phase I/IIa, completed | No results available | NCT03721718 | |
| Viral vector vaccine | MVA-MERS-S | CTC North GmbH & Co. KG | Phase I, completed | [0/23,0%] | NCT03615911 | [146] |
| Viral vector vaccine | MVA-MERS-S_DF1 | CTC North GmbH & Co. KG | Phase Ib, not yet recruiting | No data | NCT04119440 | [147] |
| Viral vector vaccine | ChAdOx1 MERS | University of Oxford | Phase I, recruiting | [1/24, 4%] | NCT03399578 | [148] |
| Viral vector vaccine | ChAdOx1 MERS | King Abdullah International Medical Research Center/University of Oxford | Phase I, recruiting | [6/24, 25%] | NCT04170829 | [149] |
| Viral vector vaccine | BVRS-GamVac-Combi | Gamaleya Research Institute of Epidemiology and Microbiology/Acellena Contract Drug Research and Development | Phase I/II, recruiting | No data | NCT04128059 | |
| Viral vector vaccine | BVRS-GamVac | Gamaleya Research Institute of Epidemiology and Microbiology | Phase I/II, recruiting | No data | NCT04130594 | |
Studies of coronavirus vaccines have a limited number of human participants and still represent a novel technique. The recent implementation of large-scale vaccination programs for Covid-19 has greatly increased the level of data available to assess the safety of human coronavirus vaccines.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



