
Submitted:
10 January 2023
Posted:
13 January 2023
You are already at the latest version
Abstract
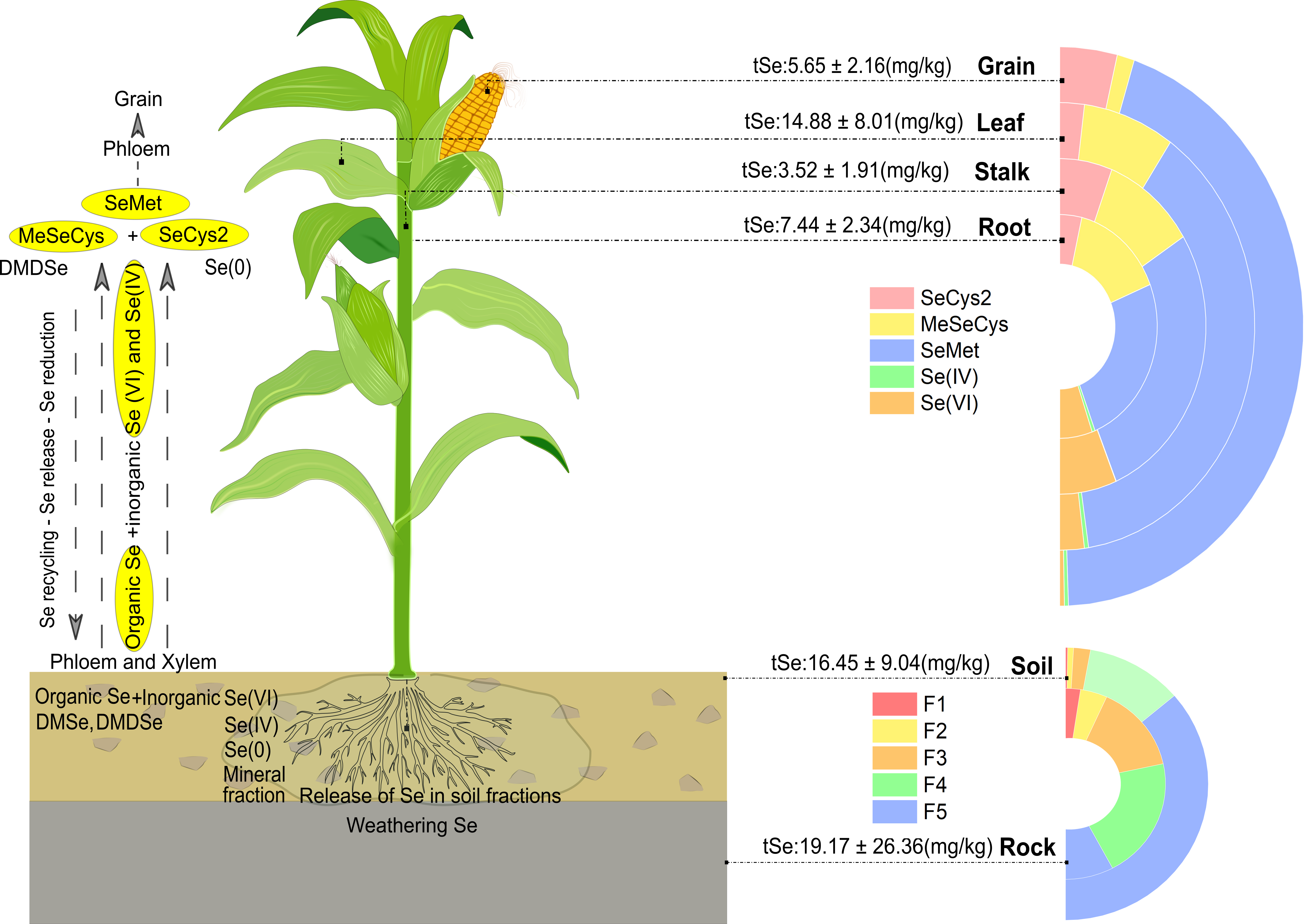
Maize (Zea mays) is one of the most important staple food and primary source of livestock feed in the world. As the consumption of maize grown on the selenium-enriched soils of Naore Valley is one of the apparent causes of selenosis in the area, this work collected and analyzed total Se, Se fractions, and Se species distributions in maize plant samples, including grains, leaves, stalks, roots, rhizosphere soils, and the most representative parent rock materials from Naore Valley, Ziyang County, China. The Se distribution in soils markedly correlated with the weathered Se-enriched bedrocks, but most of the Se in the analyzed soils is enclosed as recalcitrant residual Se and organic-sulfide bound Se. In contrast, Se in rocks had a comparatively higher bioavailability and is bounded mainly to organic matter and sulfides minerals, with very few of the Se enclosed in the residual fraction. Maize plants might take a large amount of Se from the organic-sulfide bound Se fraction in the Se-rich soils, the weathered products from bedrocks or plant litters. Total Se concentrations in the collected samples were observed in descending order soil>leaf>root>grain>stalk. The predominant Se species detected in maize plants was SeMet. Se inorganic forms, mainly Se(VI), decreased from root to grain and were possibly assimilated into organic forms. Se (IV) was barely present. The natural increases in Se concentration affected mainly leaf and root dry-weight biomass as they are the organs that coped with the highest Se accumulation. This paper offers an insight into the uptake, accumulation, and distribution of Se forms in natural Se-rich maize crops and an opportunity for shifting Se-rich soils from menaces to valuable resources for growing Se-rich agricultural products.
Keywords:
Selenium species
; Maize plants
; Se fractions
; Rock-soil-plant interface
; Selenium accumulation
1. Introduction
Selenium (Se) is considered an essential micronutrient for humans and animals (Rayman, 2000) and plays a significant role in maintaining a healthy immune system, reducing cancer and cardiovascular diseases, and is of great concern for its narrow range between dietary essentiality and toxicity for human beings (Navarro-Alarcón and López-Martínez, 2000). However, in many regions of central China, Finland, and Sweden, the dietary Se intake from food consumption is lower than the 40 μg day-1 recommended by the World Health Organization (WHO) (Combs, 2001). Generally, Se is widely distributed in rocks, soils, sediments, waters, and plants in the environment system, and Se in soils represents the major source for Se in plants.
As a trace element, the selenium content in the earth's crust is very heterogeneous and low, with an abundance of 0.13 mg kg-1 (Rudnick and Gao, 2003). Soil Se concentrations worldwide are also considerably low, mainly varying from 0.01 to 2 mg kg-1, with a mean value of 0.4 mg kg-1 (Fordyce, 2007). Similarly, soil Se concentrations in China mostly vary from 0.02 to 3.8 mg kg-1, with a mean value of 0.24 mg kg-1 (Tan et al., 2002). Soil Se mass fractions in some Se-rich areas of China can reach 87.3 mg kg-1 in Enshi Prefecture, Hubei Province (Qin et al., 2013) and 36.1 mg kg-1 in Ziyang County, Shaanxi Province (Cui et al., 2017), making these two regions the most seleniferous areas in China. However, soil available Se depends not only on the total Se but also on its speciation and fractions in soils. Se in soil mainly occurs in four states (-II, 0, IV, VI), of which inorganic Se(VI) is more accessible to plants for its high solubility and mobility, whereas Se(IV) has a higher affinity than Se(VI) for adsorption into charged surfaces of clay minerals, thus reducing its availability for plants (Hartikainen, 2005). Under more strongly reducing conditions, selenite tends to reduce into elemental Se(0) and selenides Se(-II), which are poorly mobile, presenting as stable and insoluble forms (Supriatin et al., 2015). Different forms of organic Se in humic and fulvic acid fractions of soil organic matter might also serve as potential plant-available Se in soil (Supriatin et al., 2016). Apart from different chemical forms in the soil, Se also exists in different geochemical fractionations based on their difference in binding capacity with various soil components, separating by sequential extraction procedures (SEPs) for extracting soil or rock Se fractions (Martens and Suarez, 1997; Kulp and Pratt, 2004). Particularly, Soluble Se and exchangeable Se are commonly considered plant-available Se, but organic-bound Se is also potentially available for plant uptake (Li et al., 2016).
Maize (Zea mays), also known as corn, is the most widely grown crop in the study area of Ziyang County. Wang et al. (2012) reported that the total Se concentrations in different maize tissues in Ziyang County decreased in the order of leaf > root > seed > stalk, of which the highest concentration could reach 8.7 mg kg-1, 4 mg kg-1, 3.8 mg kg-1, and 2.3 mg kg-1, respectively. Wang et al. (2019) also reported that Se concentrations of maize grain samples in Shuang'an Town, Ziyang County, were 0.9 ± 5.4 mg kg-1, 62% of the analyzed samples exceeding the Se toxicity standard (> 1 mg kg-1), and 30% of which were higher than 3 mg kg-1. Nevertheless, in other countries or regions, Se content is too low in maize grains to meet the dietary Se requirement. For instance, Zhang et al. (2019) reported that the Se concentrations in the maize plant tissues in the Wumeng Mountain area, Guizhou, China were 179-232 μg kg-1 in roots, 23-87 μg kg-1 in stalks, 86-166 μg kg-1 in leaves, and 10-90 μg kg-1 in grains. Thus, Se biofortification has been used to increase Se concentration in crops (Alfthan et al., 2015). It is necessary to understand the transfer patterns of Se from rock to soil, soil to plant, and from root to grain, because the accumulation of Se in edible crop parts is directly related to Se in soil (Sun et al., 2010). However, there have been few reports about Se distribution, accumulation, and translocation in the whole maize plant, e.g., root, stalk, leaf, and grain, under field conditions. Thus, the relationships between the Se concentrations in different tissues of maize plants and their total Se and Se fraction in rhizosphere soils and rocks are still unclear, which is important to understand the Se uptake, translocation, and accumulation by the maize plant.
In addition to total Se concentrations in plant food, the interest in Se species in plants is because of the different health benefits of antioxidant and anticarcinogenic properties of certain species (Zhou et al., 2018). Merely using total Se concentration to assess the Se-enriched crop qualities is insufficient, as the intake of Se species is important in determining its toxicity, nutritional value, and metabolic pathways in crop tissues (Gergely et al., 2006). Se species in different crop types have been investigated, and SeMet was confirmed to be the major Se species in wheat, rice, and other grain crops (Hart et al., 2011). However, Se species percentages in different tissues of maize grown in soils of seleniferous areas have rarely been studied.
In this study, we investigated the total Se (tSe) concentrations and Se species in different maize organs and tSe and Se fraction concentrations in their corresponding rhizosphere soils and parental rocks from the seleniferous area of Naore Valley in Ziyang County, China. The purpose of this research was to elucidate the uptake, accumulation, translocation, and distribution of excessive amounts of tSe and Se species in maize plants from a natural Se-rich soil environment and the geological input of Se in the crop system.
2. Materials and Methods
2.1. Location and geological settings of the study area
Naore Village, a village of few inhabitants in the Naore valley, is located 3 km northeast of Shuang'an town, Ziyang County, Ankang City, Shaanxi Province (Figure 1). Ziyang county is geographically situated between the Qinling Mountains and the Bashan Mountains. It has a typical north subtropical humid condition, monsoon climate, with an average annual temperature of 15.1 ℃ and average annual precipitation of 1066mm.
The Lujiaping Formation of lower Cambrian strata with particular Se geological enrichment is partly exposed in the Ziyang area. It consists of black shales containing pyrite, intercalated stone coal, and pyritic tuff, as well as a set of light metamorphic and sedimentary shallow fine clastic rocks and argillaceous rocks derived from the faults system. Vanadium and phosphorous nodules are found in the siliceous limestones. The total thickness of the whole stratigraphic record is 700-900 m, but the Se-bearing strata are 250-400 m, averaging 20 mg kg-1 (Luo, 2006). The Naore valley is aligned structurally with a monoclinal structure with a northwest trend. The soil in the study area (yellow-brown soil) has a shallow thickness that varies from 10 to 30 cm and was developed from the weathering of the rocks mentioned above and has Se average concentration of 26 mg kg-1 (Luo et al., 2004).
The specific lithology strata distribution found in the 11 exposed outcrops studied was as follows: sites 12,13,18 weathered carbonaceous shales with occasional fine calcite lamination and high quartz contents, similarly in site 21 carbonaceous slate with high amounts of pyrite and iron oxides. In sites 14 and 22, small layers of yellow argillaceous shale and limestone, from site 16, carbonaceous stone coal lenses, in sites 15, and 17, weathered and fresh gray carbonaceous shales interspersed with quartz masses and veins, while in site19 non-pyritic gray tuffaceous slate high in carbonate contents.
2.2. Collection and preparation of rock, soil, and plant samples
Sampling was performed in August 2020, which was the harvest season for the maize previously sown in late April. Mature maize plants, rhizosphere soil around roots, and corresponding parent rock material were collected from 11 field sampling points in a parallel transect alongside the upper part of the Naore valley. Because the Se concentration in the lower part of the valley is uneven (Wang et al., 2012), selenium fertilizer has been applied to compensate for these Se differences, so these soils are not anymore pristine.
According to the technical specification for soil environmental monitoring (HJ/T 166-2004) and the land quality geochemical assessment (DZ/T 0295-2016), soil samples were collected at 0–20 cm soil depth, mainly representing the plow layer from 3 subpoints around the plant, combined as one representative sample, and kept in cloth bags for transportation. At each of the 11 sampling plots, plant samples were collected as a whole by digging up the roots, and parental rock samples were taken from nearby exposed outcrops for the geochemical characterization of the sampling sites and the main lithology. All the soil samples were yellow-brown tillage rhizosphere soils with small amounts of partially weathered rock debris and plant roots, identified as Luvisols, referring to the World Reference Base (WRB) for Soil Resources (FAO, 2014).
After removing dirt, maize samples were washed with deionized water, separated root, stalk, leaves, and grains, weighed and oven-dried at 50 ºC during 2 days. The dried plant parts were weighed again, ground to a fine powder using a plant grinder, and stored at -20 ºC until analysis. After roots and detrital materials were removed, the rhizosphere soil and rock samples were ground to pass through a 0.074 mm nylon sieve and homogenized. The pH values were measured using an FE28-pH meter at a ratio of 1:2 (soil: water ratio). Soil and rock contents of TOC were analyzed by the K2Cr2O7-H2SO4 oxidation method with 1 g of samples (Nelson and Sommers, 1996).
2.3. Total selenium (tSe) determination in soil, plant, and rock samples
A portion of 0.5 g of dried and powdered maize samples were mixed with 2 mL concentrated HNO3 and 1 mL H2O2 in polytetrafluoroethylene PTFE vessels, allowed to react for 4 h, and then digested for 6 h at 160 °C according to Tang et al. (2021). The remained acid solutions were reduced to Se(IV) as described below. For soil and rock digestion, a 0.1 g sample was treated with 3 mL of a concentrated mixed acid (HNO3: HF: HClO4, 2: 0.5: 0.5, v/v) in polytetrafluoroethylene (PTFE) line bomb vessels and digested overnight at 180 °C. The remained acid solutions were evaporated to almost dryness, and 2 mL of 6 mol/L HCl solution was added and heated at (<90 °C) for 1 h to reduce Se(VI) to Se(IV) before analysis of tSe. The obtained solutions were diluted with 5% HCl and stored at 4°C until being analyzed by HG-AFS (AFS-9230, Beijing Jitian Instrument Co, China). Reagent blanks and standard reference material GBW10045 were used for quality control.
2.4. Chemical fractions of Se in soil and rock samples
Five operationally defined Se fractions were determined by a method modified from Kulp and Pratt, (2004). Soil samples (1.0 g) were placed into 50 mL centrifuge tubes, and the analytical procedure was as follows: For the first extraction step (F1= Water soluble Se), 20 mL of ultrapure water was added and shaken on a horizontal shaker at room temperature for 1.5 h. For the second step (F2= Exchangeable Se), 10 mL of 0.1 mol L-1 KH2PO4-K2HPO4 buffer solution was added and shaken for 0.5 h. The third step (F3= Alkali-soluble Se) integrates the extraction of hydrolyzable "labile" organic matter associated with humic substances and manganese, iron, and aluminum hydroxides and was extracted through the addition of 0.1 mol L-1 NaOH and placed in a water bath at 90°C for 2 h. In the fourth step, (F4= Acid-soluble Se) it is targeted the extraction of Se bound to acid-soluble labile organic matter that was not solubilized in the previous fraction, and Se bound to sulfides, the residue left from F3 was oven dried (50°C), and 0.5 g KClO3 and 10 mL of concentrated HCl were added and mixed for 1 h with occasional shaking. Afterward, all the samples were centrifuged at 4000 rpm for 20 min, and the supernatant was filtered with 0.45 μm membrane filters. Then, 10 ml of ultrapure water was added to the tube, and centrifuging was repeated. The two supernatants were combined and stored at 4°C until analysis. The tSe concentration in the extraction solution was measured by hydride generation-atomic fluorescence spectrometry (AFS-9230, Beijing Jitian Instrument Co, China). The extraction quality was assessed with the certified reference materials GBW07901 and GBW07400 for soil and rock, respectively. Standard and blank solutions were prepared following the same procedure together with the samples. The sequential extraction resulted in recoveries between 80 to 120%.
2.5. Enzyme-soluble selenium species extraction from plant organs
Enzymatic hydrolysis according to the ERM-BC210A standard material matching method (ERM_certificate) was used to extract five enzyme-soluble chemical selenium forms; selenium cysteine SeCys2, methylselenocysteine MeSeCys, selenomethionine SeMet, Selenium tetravalent Se(IV) and Selenium hexavalent Se(VI), from maize plants collected in the field, as follows: 0.5 g sample was weighed in triplicates into 20 mL centrifuge tubes, and 60 mg protease XIV, 30 mg lipase and 10 mL 30 Mm TrIS-HCl (pH = 7.5) were added for ultrasonic degassing. After that, they were cultured in darkness at 37° C for 20 hours, mixed well, and oscillated in a water bath oscillator at 150 r/min. The hydrolyzed sample was centrifuged at 10000 r/min at 4°C for 30 min. The supernatant was filtered and stored at -20 °C, and the remaining residue was added to 5 mL of hydrolysate containing 100 mg collapse enzyme, and the above steps were repeated. The two supernatants were combined and diluted and immediately injected in an HCLP-ICP-MS.
The concentrations of extracted Se species were determined using an ICP-MS (NexION 2000 triple quadrupole) under dynamic reaction cell mode, which was coupled to an HPLC system (Agilent 1000) equipped with an ion-exchange anion exchange chromatography column (Hamilton PRP-X 100). The optimized mobile phase was mixed with ultra-pure water (87%), methanol (3%), and 50 mmol L-1 pH = 5.5 citric acid (10%). The higher resolution mode of the ICP-QQQ allows 80Se to be chosen as the reported Se isotope due to its highest abundance. CH4 was used as the reaction gas to avoid 40Ar40Ar+ ion interference.
The precision and accuracy obtained for tSe and Se species were assessed using ERM-BC210a for Se and SeMet analysis with enzymolysis recoveries of 90.9% ± 9.1. Spike additions (0.5,1 and 2 mg kg-1) also showed good agreement determination of the selenium forms with recoveries that ranged from 86% to 108%. Detection limits of the method (LOD) ranged from 0.87 μg L-1 to 1.70 μg L-1, among which the SeCys2 was the lowest (0.87 μg L-1), while the SeMet was the highest (1.70 μg L-1) (Table S1, Supplementary Data).
2.6. Statistical analysis
Relationships between tSe in maize organs, rocks, soils, Se fractions, and Se species were identified using Pearson and Spearman correlation coefficients (r), and significance probabilities (p), as well as linear regressions and graphical analysis, were performed by Origin 2017. All values presented are expressed as mean ± standard deviation in triplicates. Reported numbers are rounded for results beyond 0.1 (tSe) and 1 (Se species). Differences were considered not significant at values of p > 0.05.
3. Results and Discussion
3.1. Selenium fractions in rock-soils interface and translocation potential
The Se distribution in the different rock parent material samples and the soils derived from these strata are presented in Figure 2. The coal lenses intercalated within the black-gray shales were identified as the most important selenium carriers in Naore Valley and have the highest potential to mobilize Se through soil-forming processes to maize crops. The Se concentrations in the rock strata were unevenly distributed (from 1.3 ± 0.5 to 85.5 ± 3.4 mg kg-1). Gray-black shales, yellow argillaceous shales, and limestones all had high tSe, while the lowest tSe content was found in the gray non-pyritic tuffaceous slate (Table 1).
These results were consistent with our previous study in Tian et al. (2020), which demonstrated that Se in the Lower Cambrian Lujiaping Formation in Naore Village was mainly associated with pyrite-bearing black shales, while in contrast, the non-pyritic tuff has low Se concentrations. The rhizosphere soil showed a range of Se concentrations (from 7.8 ± 0.1 to 29.9 ± 2.1 mg kg-1), 20 to 70-fold higher than the average of 0.4 mg kg-1 (Fordyce, 2007), thus classified as ultra-high Se-rich soil in China.
The pH ranged from 6.5 to 7.9 and 6.5 to 9.3, while the TOC concentrations ranged from 2.2 to 5.7 % and 0.2 to 14.8% for soils and rocks, respectively. The TOC concentrations were significantly positively correlated with the tSe in soils (r = 0.61 p < 0.05) but not in rocks (Table 1). This indicates the role of biogeochemical cycling in the investigated soil-plant system where mobile-available Se forms were transferred from weathered rocks to soils, taken up by plants, and possibly returned to the soil in the form of organically bound Se with dead plant material. The biological accumulation of Se and its release into the topsoil from fallen litter upon decomposition have been shown as an important Se input in soils of typical selenosis area (Li et al., 2008). However, high TOC contents do not necessarily suggest a higher accumulation of mobile-available Se forms, as organic matter increases the organic bound and residual fractions and conversely declines the soluble and exchangeable Se fractions (Sun et al., 2021). The Se encapsulated in less mobile forms can be remobilized back by root exudates, as maize roots induce localized dissociation and release of mobile Se (Wang et al., 2019) which can lead to the oxidation of reduced Se species along with the mineralization of Se from organic matter that increases Se mobility (Schilling et al., 2015).
In the rhizosphere of plants, the microbial activity is higher than that in the bulk soil or weathered rocks (Wang et al., 2022), so chemical forms of Se in the rhizosphere are governed by the release of primary and secondary metabolites from root-microbiome exudates that alter the chemical speciation, mobility and plant availability of Se in the vicinity of roots, making the Se more accessible for plant uptake. This might explain why the chemical Se fractions distribution in parental rock materials substantially differed from the pattern found in soil samples.
The collected rock samples had more mobile Se content (F1+F2) than soils, averaging 10.8% vs. 1.8 %, respectively. The highest proportion of Se fractions in rocks, about 76.7%, were found in F3+F4, which accounts for the Se bound to hydrolyzable organic matter, recalcitrant organic matter, and pyrite-sulfide minerals. In contrast, most of tSe in soil was found in F5, averaging 71.1%, trapped as immobile residual Se in soil minerals, and only about 21.6% of soil Se was found in F4 (results are listed in Table S2, Supplementary Data). Correlations analysis between soil and rock Se fractions showed the dominant fractions (F4+F5) in soils, as well as tSe, had a significant positive correlation with the dominant fractions (F3+F4) in rocks, (r = 0.828, p < 0.001 and r = 0.906, p < 0.001, respectively) (Table S3, Supplementary Data), which indicates the geological source of Se. Similarly, the organic bound Se in Se-rich soils and weathered stone coal has been proven as the potential source of bioavailable Se in selenosis areas of Enshi in Hubei province (Qin et al., 2011). However, it is reasonable to think that Se in agricultural soils is influenced not only by bedrock weathering but also by tillage (Qin et al., 2017) and other agricultural management measures (Yu et al., 2014).
Unfortunately, we did not consider the chemical fractionation of Se in non-rhizosphere soil or other inputs; thus, our data does not allow further interpretation. Nonetheless, the link between human activities as the main cause of high Se in cropland soils has already been assessed by Zhu et al. (2008), who suggested that in tandem with natural factors (weathering and leaching of Se-enriched rock), the areas of higher soil Se concentrations in Yutangba are located where the soils have been heavily disturbed by human activities. The mechanical action of tillage most probably influences the transfer of partially weathered rock debris from bottom to topsoil and impacts the amount of soil organic matter (Table 1) and dead plant roots that have been raised up, accelerating the oxidation of sulfide/selenide fractions in cultivated soils and thereby influencing tSe and labile Se forms in soils.
3.2. Selenium accumulation in maize depending on tSe and Se fractions in soil
Since the tSe in soils can be a misleading predictor of the bioavailable Se for plants, to measure the relationship between tSe and Se fractions in soils against tSe and Se species in maize organs and obtain information on the fractions influencing Se accumulation in maize, Pearson's and Spearman correlation analyses were performed. There were significant positive correlations between water-soluble Se(F1) + exchangeable Se(F2), and tSe in leaves (r = 0.691 p < 0.05), but there were not found significant correlations with tSe in soil. Alternatively, Spearman correlations with F2 showed that tSe in leaf, grain, and stalk were significantly correlated with correlation coefficients of 0.781, 0.709, and 0.681, respectively (Table S4, Supplementary Data). While these results are in agreement with other authors (Wang et al., 2019; De Feudis et al., 2018), the low concentration of these fractions that are generally regarded as easily bioavailable for plants, indicates that despite in Naore village, most of the soil Se is not readily bioavailable, maize plants could be taken up Se from less bioavailable pools.
The tSe concentration in the different compartments considered was in the descending order rock>soil>leaf>root>grain>stalk (Figure 3), with the tSe concentrations in leaves being 2-fold higher than roots, indicating a high efficiency of Se accumulation in the leaves and roots, and high Se translocation from roots into shoots. These results are consistent with the findings of Wang et al. (2012), who previously investigated tSe in soil and plants from the Naore Valley. High tSe concentrations in maize leaves have also been found in Enshi (Zhu et al., 2000), suggesting a high capacity for accumulating Se in the above-ground organs of maize.
As the uptake and accumulation of Se in maize plants are mainly influenced by the uptake of Se from the soil via sulfate transporters and the translocation of organic and inorganic species, the Se depending on cellular level concentration can influence a variety of metabolic processes that can affect plant vigor, growth and biomass production. Thus, the tSe concentrations in soils and plants were compared against the dry-weight accumulation in each organ to investigate whether decreases in dry-matter accumulation were related to tSe (Table S5, Supplementary Data). Negative Pearson's correlations were found between tSe in soils when compared with leaf and root biomass (r= -0.672, p < 0.01) and (r= -0.370, p < 0.05), respectively, as well as with the different soil Se fractions. Root biomass was also negatively correlated with its tSe (r= -0.410, p < 0.05), and for the case of grain biomass, it was negatively correlated with its tSe (r= -0.440, p < 0.05), and the tSe in stalks (r= -0.346 p < 0.05), but positively correlated with stalk biomass (r= 0.372 p < 0.05) and root biomass (r= 0.501 p < 0.05). It is likely that high Se concentrations negatively affected the dry-matter accumulation in maize plants, concretely the organs that coped with the highest Se accumulation, roots, and leaves in this study. One possible reason is that toxic amounts of Se can inhibit the root system elongation in maize plants and dramatically decrease biomass accumulation in above-ground organs (Hawrylak-Nowak, 2008), as Se specifically decreases the ability for photosynthesis and photosynthate translocation in leaves (Jiang et al., 2017), negatively affecting maize plant development (Placzek et al., 2021).
3.3. Selenium organ-specific species distribution in maize
A large extent of the tSe in roots, stalks, and leaves was enzyme-insoluble. Figure 4 reveals a clear different pattern of Se species distribution in each maize organ. The results for Se species in the enzymatic hydrolysis extract of maize organs are summarized in Table 2 and listed in detail in Table S6 (Supplementary Data). The average concentration of organic Se species (SeMet, MeSeCys, and SeCys2) were comparatively 7, 25, and 100-fold higher than the inorganic species Se(IV)+Se(VI), in roots-stalks, leaves, and grains, respectively. Unknown peaks were observed on each column, however, they were ignored as the largest unknown peak represented less than 2% of Se in the sample.
The enzyme-soluble Se in maize was mainly as SeMet, with an average increase from 41.8% in roots up to 86.7% in grains. SeMet is the Se analog of methionine, and its levels highly variate between maize genotypes (Lai and Messing, 2002). Still, it has in common that maize that thrives in seleniferous soils stores most of Se in tissue as SeMet (Beilstein et al., 1991). Inorganic species instead were more predominant in stalks and roots, for around 20% but decreased on leaves to an average of 7.3% and to 2.3% in grains. These inorganic Se species were found mainly Se(VI) and very few as Se (IV).
Other identified organic species were MeSeCys which decreased from 32.7% in roots to 2.6% in grains, and SeCys2, with the lowest average proportion found in leaves, 4.0%, while the highest, 12.0%, found in stalks. This changing distribution in the accumulation of enzyme-soluble organic Se species seems to link SeCys2 and its methyl forms as key precursors for forming SeMet (Duncan et al., 2017), while, MeSeCys was an important component of Se in the root, probably had been developed mainly from biotic processes happening in the below-ground system of the maize crop (Pilon-Smits and Quinn 2010), as this form of Se can safely be accumulated in high concentrations since it is not incorporated into proteins (Neuhierl et al., 1999), and it is also the precursor for DMDSe (dimethyldiselenium), a key specie for the plant metabolic mechanism to excrete Se excessive toxic accumulations (Ellis et al., 2003).
Enzyme-soluble Se species in the stalk, leaf, and grain were also positively correlated with tSe, except for inorganic Se in grains (Figure 5), suggesting that most of the inorganic Se forms might be transposed to organic bound Se or further reduced. As maize is a high-stalk plant, the Se uptake exhibits a similar trend to that observed in rice, where it has been demonstrated that organic Se species, mainly SeMet, MeSeCys, and inorganic Se(VI) are rapidly loaded into the phloem and xylem vessels, yet organic species are delivered to the above-ground organs far more efficiently than Se(VI) (Carey et al., 2012). While, Se(IV) form is scarce in the maize plant as it is readily assimilated into organic forms in the root (Yu et al., 2011) and translocated or rapidly reduced to Se(0) enzymatically by sulfite reductase or non-enzymatically by reduced glutathione (White, 2016 ), whereas Se(VI), if not reduced, is delivered immediately.
Moreover, all these linear relationships indicated that the concentrations of Se species in maize could be predicted by tSe concentrations in any above-ground part of maize plants (i.e., stalk, leaf, grain), except for the root that as a result of poor enzyme-soluble Se concentrations, there was a generalized absence of correlation with tSe, suggesting that uptake and conversion of Se species in roots were different and the extraction of enzyme-soluble Se species was possibly affected by other factors.
The gradual decrease of Se extraction recoveries seen from grain to root might be a consequence of the reaction of Se species with phenolic compounds in the matrix components, an increase in insoluble-fiber content, degradations that occurred during the enzymatic extraction, or the sum of all these factors. Reactions of Se with phenols during the extraction process have been demonstrated as the main factor affecting Se speciation (Cuderman et al., 2010). Phenolics are resistant to decomposition and primarily present in the enzyme-insoluble fraction (Salinas-Moreno et al., 2017), and in maize is notorious the increase of lignin from leaf to root, a phenol that in maize is more pronounced than in other cereals (Abiven et al., 2011). While the lowest insoluble fiber content is found in maize grains, which explains why easily extracted Se bonds were generally higher in grains (Smrkolj et al., 2005). Future studies on maize or other plants should focus on Se enzyme-soluble species extraction in roots to isolate the effects of matrix components that might lead to low extractions recoveries.
In the present study, although there were large differences in the tSe concentrations of the different maize organs and tSe concentrations in soils between sampling areas, the distribution and translocation pattern of Se species in maize plants growing in natural Se-enriched soils were comparatively uniform. In general, the proportion of organic species displayed increases of SeMet but diminutions of MeSeCys from roots to grains, and variable SeCys proportions, while inorganic Se species showed a diminution in leaves and grains. Unlike other organic forms, SeMet is barely lost after cooking (Lu et al., 2018). This implies that from a raw portion of only 100 grams of the analyzed maize grains, roughly 560 µg of Se could be ingested, of which up to 70% could be bioavailable to the human gastrointestinal tract in the main form of SeMet (Muleya et al., 2021). Thus, considering that Se daily intake in Naore Village is still excessive (1801 μg Se), with 46.65% coming from cereal consumption (Cui et al. 2017). The consumption of locally grown maize can significantly contribute to Se ingestion beyond the tolerable upper intake (300 µg day-1) (SCF, 2000), presenting still a latent risk for human health.
However, the current situation differs from the past prevalence years of human selenosis outbreak. Contrary to the drought period that triggered it, villagers nowadays do not use maize stalks as fuel, nor their ashes as soil fertilizer, and have access to a more diversified diet from non-seleniferous regions. So the exposure to highly toxic amounts of Se by ingestion, inhalation, contact of Se with foodstuff by its smoke, and Se recycling in soils probably decreased, and maize is no anymore a selenosis aggravating factor. Local consumer exposure to toxic Se concentrations from maize grains-based products can be mitigated by limiting direct consumption and reducing the Se levels through blending low and high Se grains or inter-cropping. Nevertheless, new cases of chronic human selenium poisoning caused by ingesting locally produced seleniferous food are rare, yet Se deficiencies and low Se-intakes are more likely to happen. Thus crops grown in Se-rich areas can alternately be economically rentable and promoted as Se-enriched agricultural resources as the cost of Se-rich products and the environmental impact of Se fertilizer usage can be reduced, becoming a valuable and unexpected opportunity,
4. Conclusions
The particular geological and pedological enrichment of Se implies that maize crops cultivated in the area end up accumulating large extents of Se. Five Se species, including SeMet, SeCys2, MeSeCys, Se(IV), and Se(VI), were detected in the maize plant samples with SeMet as the predominant specie, while Se inorganic forms decreased in importance from root to grain. Further studies are needed to verify if these distributions are similar for different maize plant characteristics or genotypes. Particular attention should be paid to the impact of soil Se on maize plant yield, as there was a negative correlation between maize plant biomasses and excessive soil Se concentrations. And care has to be taken to avoid low enzymatic extraction recoveries when related to increases in insoluble-fibers contents of the maize plant, as the present study also found overall decreasing extraction efficiencies from grain to root.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
This study was financially supported by China Geological Survey (No. DD20190475), the National Natural Science Foundation of China (42003035), the Youth Science and Technology New Star Project of Shaanxi Province (2021KJXX-98, AK2020-FX01-9), and Natural Science Foundation of Shaanxi Province (2021JQ-956 and 2018JQ3064), and also supported by the Fundamental Research Funds for the Central Universities, China University of Geosciences (Wuhan) (No. G1323520058 and CUG170104). The authors also appreciate the constructive comments and suggestions of the anonymous reviewers and journal editors for editorial handling.
References
- Abiven, S.; Heim, A.; Schmidt, M.W.I. Lignin content and chemical characteristics in maize and wheat vary between plant organs and growth stages: Consequences for assessing lignin dynamics in soil. Plant Soil 2011, 343, 369–378. [Google Scholar] [CrossRef]
- Alfthan, G.; Eurola, M.; Ekholm, P.; Venäläinen, E.R.; Root, T.; Korkalainen, K.; Hartikainen, H.; Salminen, P.; Hietaniemi, V.; Aspila, P.; Aro, A. Effects of nationwide addition of Selenium to fertilizers on foods, and animal and human health in Finland: From deficiency to optimal selenium status of the population. J. Trace Elem. Med. Biol. 2015, 31, 142–147. [Google Scholar] [CrossRef]
- Beilstein, M.; Whanger, P.; Yang, G. Chemical forms of Selenium in corn and rice grown in a high selenium area of China. Biomed Env. Sci. 1991, 4, 392. [Google Scholar]
- Carey, A.M.; Scheckel, K.G.; Lombi, E.; Newville, M.; Choi, Y.; Norton, G.J.; Price, A.H.; Meharg, A.A. Grain accumulation of selenium species in rice (Oryza sativa L.). Environ. Sci. Technol. 2012, 46, 5557–5564. [Google Scholar] [CrossRef] [PubMed]
- Combs, G.F. Selenium in global food systems. Br. J. Nutr. 2001, 85, 517–547. [Google Scholar] [CrossRef] [PubMed]
- Cuderman, P.; Stibilj, V. Stability of Se species in plant extracts rich in phenolic substances. Anal. Bioanal. Chem. 2010, 396, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Huang, J.; Peng, Q.; Yu, D.; Wang, S.; Liang, D. Risk assessment for human health in a seleniferous area, Shuang'an, China. Environ. Sci. Pollut. Res. 2017, 24, 17701–17710. [Google Scholar] [CrossRef] [PubMed]
- De Feudis, M.; D’Amato, R.; Businelli, D.; Guiducci, M. Fate of selenium in soil: A case study in a maize (Zea mays L.) field under two irrigation regimes and fertilized with sodium selenite. Sci. Total Environ. 2019, 659, 131–139. [Google Scholar] [CrossRef]
- Duncan, E.G.; Maher, W.A.; Jagtap, R.; Krikowa, F.; Roper, M.M.; O'Sullivan, C.A. Selenium speciation in wheat grain varies in the presence of nitrogen and sulphur fertilisers. Environ. Geochem. Health 2017, 39, 955–966. [Google Scholar] [CrossRef]
- Ellis, D.R.; Salt, D.E. Plants, Selenium and human health. Curr. Opin. Plant Biol. 2003, 6, 273–279. [Google Scholar] [CrossRef]
- ERM_certificate, ERM_certificate: http://www.speciation.net/Database/Materials/LGC-Ltd/ERMBC210a--Wheat-Flour--Selenium-and-Selenomethionine-;i1203 (available on 90.09.2022). Google Scholar.
- Food and Agriculture Organization of the United Nations (FAO)., 2014. World reference base for soil resources 2014. International soil classification system for naming soils and creating legends for soil maps. FAO, Rome.
- Fordyce, F. Selenium Geochemestry. Int. Symp. Med. Geol. 2007, 18–19. [Google Scholar] [CrossRef]
- Gergely, V.; Kubachka, K.M.; Mounicou, S.; Fodor, P.; Caruso, J.A. Selenium speciation in Agaricus bisporus and Lentinula edodes mushroom proteins using multi-dimensional chromatography coupled to inductively coupled plasma mass spectrometry. J. Chromatogr. A 2006, 1101, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Gupta, S. An overview of selenium uptake, metabolism, and toxicity in plants. Front. Plant Sci. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hartikainen, H. Biogeochemistry of Selenium and its impact on food chain quality and human health. J. Trace Elem. Med. Biol. 2005, 18, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Hart, D.J.; Fairweather-Tait, S.J.; Broadley, M.R.; Dickinson, S.J.; Foot, I.; Knott, P.; McGrath, S.P.; Mowat, H.; Norman, K.; Scott, P.R.; Stroud, J.L.; Tucker, M.; White, P.J.; Zhao, F.J.; Hurst, R. Selenium concentration and speciation in biofortified flour and bread: Retention of Selenium during grain biofortification, processing and production of Se-enriched food. FOOD CHEM 2011, 126, 1771–1778. [Google Scholar] [CrossRef] [PubMed]
- Hawrylak-Nowak, B. Effect of Selenium on Salt Tolerance in Maize Plants. J. Environ. Sci. 2008, 49, 91–124. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na + accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kulp, T.R.; Pratt, L.M. Speciation and weathering of Selenium in Upper Cretaceous chalk and shale from South Dakota and Wyoming, USA. Geochim. Cosmochim. Acta 2004, 68, 3687–3701. [Google Scholar] [CrossRef]
- Lai, J.; Messing, J. Increasing maize seed methionine by mRNA stability. Plant J. 2002, 30, 395–402. [Google Scholar] [CrossRef]
- Li, J.; Peng, Q.; Liang, D.; Liang, S.; Chen, J.; Sun, H.; Li, S.; Lei, P. Effects of aging on the fraction distribution and bioavailability of Selenium in three different soils. Chemosphere 2016, 144, 2351–2359. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Luo, K.; Li, H. Environmental behaviors of selenium in soil of typical selenosis area, China. J. Environ. Sci. 2008, 20, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; He, Z.; Lin, Z.; Zhu, Y.; Yuan, L.; Liu, Y.; Yin, X. Effects of Chinese cooking methods on the content and speciation of selenium in selenium bio-fortified cereals and soybeans. Nutrients 2018, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Luo, K. The Lujiaping Formtion of Northern Daba Mountain. J. Stratigraphy, 2006, 30(2): 149–156 (in Chinese with English Abstract).
- Luo, K.; Xu, L.; Tan, J.; Wang, D.; Xiang, L. Selenium source in the selenosis area of the Daba region, South Qinling Mountain, China. Environ. Geol. 2004, 45, 426–432. [Google Scholar] [CrossRef]
- Martens, Dean A.; Suarez, Donald L. Selenium Speciation of Soil/ Sediment Determined with Sequential Extractions and Hydride Generation Atomic Absorption Spectrophotometry. Environ. Sci. Technol. 1997, 31, 133-139. Retrieved from https://digitalcommons.unl.edu/usdaarsfacpub/502.
- Ministry of Ecology and Environment of the People's Republic of China (MEEPRC)., 2004. Technical specification for soil environmental monitoring (HJ/T 166-2004). Beijing, 44 pp. (in Chinese).
- Ministry of Natural Resources of the People's Republic of China (MNRPRC)., 2016. Specification of land quality geochemical assessment (DZ/T 0295-2016). Beijing, 52 pp. (in Chinese).
- Muleya, M.; Young, S.D.; Reina, S.V.; Ligowe, I.S.; Broadley, M.R.; Joy, E.J.M.; Chopera, P.; Bailey, E.H. 2021. Selenium speciation and bioaccessibility in Se-fertilised crops of dietary importance in Malawi. J. Food Compos. Anal. 103841. [CrossRef]
- Nelson, D.W.; Sommers, L.E. 1996. Total carbon, organic carbon, and organic matter. Methods Soil Anal. Part 3 Chem. Methods 961–1010. [CrossRef]
- Neuhierl, B.; Thanbichler, M.; Lottspeich, F.; Böck, A. A family of S-methylmethionine-dependent thiol/selenol methyltransferases. Role in selenium tolerance and evolutionary relation. J. Biol. Chem. 1999, 274, 5407–5414. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Alarcón, M.; López-Martínez, M.C. Essentiality of Selenium in the human body: Relationship with different diseases. Sci. Total Environ. 2000, 249, 347–371. [Google Scholar] [CrossRef] [PubMed]
- Pilon-smits, E.A.H.; Quinn, C.F. 2010. Selenium Metabolism in Plants In Progress in botany (pp. 93–107). Cham, Switzerland.Springer. [CrossRef]
- Płaczek, A.; Patorczyk-Pytlik, B. The dynamics of selenium uptake by maize (Zea mays L.). Agronomy 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Qin, H. B, Zhu, J. ming, Liang, L.; Wang, M. shi, Su, H. The bioavailability of selenium and risk assessment for human selenium poisoning in high-Se areas, China. Environ. Int. 2013, 52, 66–74. [CrossRef]
- Qin, H.B.; Zhu, J.M.; Lin, Z.Q.; Xu, W.P.; Tan, D.C.; Zheng, L.R.; Takahashi, Y. Selenium speciation in seleniferous agricultural soils under different cropping systems using sequential extraction and X-ray absorption spectroscopy. Environ. Pollut. 2017, 225, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.B.; Zhu, J.M.; Su, H. Selenium fractions in organic matter from Se-rich soils and weathered stone coal in selenosis areas of China. Chemosphere 2011, 86, 626–633. [Google Scholar] [CrossRef]
- Rayman, M.P. The influence of Selenium on human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef]
- Rudnick, R.L. and Gao, S. 2003 Composition of the Continental Crust. In: Rudnick, R.L.; Ed., The Crust, Elsevier-Pergamon, 1-64. [CrossRef]
- Salinas-Moreno, Y.; García-Salinas, C.; Ramírez-Díaz, J.L.; Alemán-de la Torre, I. 2017. Phenolic Compounds in Maize Grains and Its Nixtamalized Products. Phenolic Compd. - Nat. Sources, Importance Appl. [CrossRef]
- SCF (Scientific Committee on Food)., 2000. Opinion on the Scientific Committee on Food on the tolerable upper intake level of selenium. SCF/CS/NUT/UPPLEV/25 Final. 18 pp.
- Schilling, K.; Johnson, T.M.; Dhillon, K.S.; Mason, P.R.D. Fate of Selenium in Soils at a Seleniferous Site Recorded by High Precision Se Isotope Measurements. Environ. Sci. Technol. 2015, 49, 9690–9698. [Google Scholar] [CrossRef]
- Smrkolj, P.; Stibilj, V.; Kreft, I.; Kapolna, E. Selenium species determination in selenium-enriched pumpkin (Cucurbita pepo L.) seeds by HPLC-UV-HG-AFS. Anal. Sci. 2005, 21, 1501–1504. [Google Scholar] [CrossRef]
- Sun, C.; Yang, Y.; Zeeshan, M.; Qin, S.; Ma, J.; Liu, L.; Yang, J.; Zhou, X.; Huang, J. Arbuscular mycorrhizal fungi reverse selenium stress in Zea mays seedlings by improving plant and soil characteristics. Ecotoxicol. Environ. Saf. 2021, 228, 113000. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.X.; Liu, X.; Williams, P.N.; Zhu, Y.G. Distribution and translocation of Selenium from soil to grain and its speciation in Paddy Rice (Oryza sativa L.). Environ. Sci. Technol. 2010, 44, 6706–6711. [Google Scholar] [CrossRef] [PubMed]
- Supriatin, S.; Weng, L.; Comans, R.N.J. Selenium speciation and extractability in Dutch agricultural soils. Sci. Total Environ. 2015, 532, 368–382. [Google Scholar] [CrossRef] [PubMed]
- Supriatin, S.; Weng, L.; Comans, R.N.J. Selenium-rich dissolved organic matter determines selenium uptake in wheat grown on Low-selenium arable land soils. Plant Soil 2016, 408, 73–94. [Google Scholar] [CrossRef]
- Tan, J.; Zhu, W.; Wang, W.; Li, R.; Hou, S.; Wang, D.; Yang, L. Selenium in soil and endemic diseases in China. Sci. Total Environ. 2002, 284, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Lu, G.; Fan, B.; Xiang, W.; Bao, Z. Bioaccumulation and risk assessment of heavy metals in soil-crop systems in Liujiang karst area, Southwestern China. Environ. Sci. Pollut. Res. 2021, 28, 9657–9669. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Xie, S.; Carranza, E.J.M.; Bao, Z.; Zhang, H.; Wu, S.; Wei, C.; Ma, Z. Distributions of Selenium and related elements in high pyrite and Se-enriched rocks from Ziyang, Central China. J. Geochemical Explor. 2020, 212, 106506. [Google Scholar] [CrossRef]
- Wang, M.; Cui, Z.; Xue, M.; Peng, Q.; Zhou, F.; Wang, D.; Dinh, Q.T.; Liu, Y.; Liang, D. Assessing the uptake of selenium from naturally enriched soils by maize (Zea mays L.) using diffusive gradients in thin-films technique (DGT) and traditional extractions. Sci. Total Environ. 2019, 689, 1–9. [Google Scholar] [CrossRef]
- Wang, S.; Liang, D.; Wang, D.; Wei, W.; Fu, D.; Lin, Z. Selenium fractionation and speciation in agriculture soils and accumulation in corn (Zea mays L.) under field conditions in Shaanxi Province, China. Sci. Total Environ. 2012, 427–428, 159–164. [CrossRef]
- Wang, Y.; Shi, X.; Huang, X.; Huang, C.; Wang, H.; Yin, H.; Shao, Y.; Li, P. Linking microbial community composition to farming pattern in selenium-enriched region: Potential role of microorganisms on Se geochemistry. J. Environ. Sci. (China) 2022, 112, 269–279. [Google Scholar] [CrossRef]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Yang, Z.; Lv, Y.; Hou, Q.; Xia, X.; Feng, H.; Zhang, M.; Jin, L.; Kan, Z. The origin and geochemical cycle of soil selenium in a Se-rich area of China. J. Geochemical Explor. 2014, 139, 97–108. [Google Scholar] [CrossRef]
- Yu, Y.; Luo, L.; Yang, K.; Zhang, S. Influence of mycorrhizal inoculation on the accumulation and speciation of Selenium in maize growing in selenite and selenate spiked soils. Pedobiologia (Jena). 2011, 54, 267–272. [Google Scholar] [CrossRef]
- Zhang, Z.; Yuan, L.; Qi, S.; Yin, X. The threshold effect between the soil bioavailable molar Se:Cd ratio and the accumulation of Cd in corn (Zea mays L.) from natural Se-Cd rich soils. Sci. Total Environ. 2019, 688, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yang, W.; Wang, M.; Miao, Y.; Cui, Z.; Li, Z.; Liang, D. Effects of selenium application on Se content and speciation in Lentinula edodes. Food Chem. 2018, 265, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zheng, B.; Wang, Z.; Xiao, H.; Mao, D.; Su, H. Distribution of Selenium in corn and its relationship with soil selenium in yutangba mini-landscape. Chinese J. Geochemistry 2000, 20, 161–166. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, N.; Li, S.; Li, L.; Su, H.; Liu, C. Distribution and transport of Selenium in Yutangba, China: Impact of human activities. Sci. Total Environ. 2008, 392, 252–261. [Google Scholar] [CrossRef]
Figure 1.
Location of the study area and sampling sites. Map of China based on GS(2019)182.
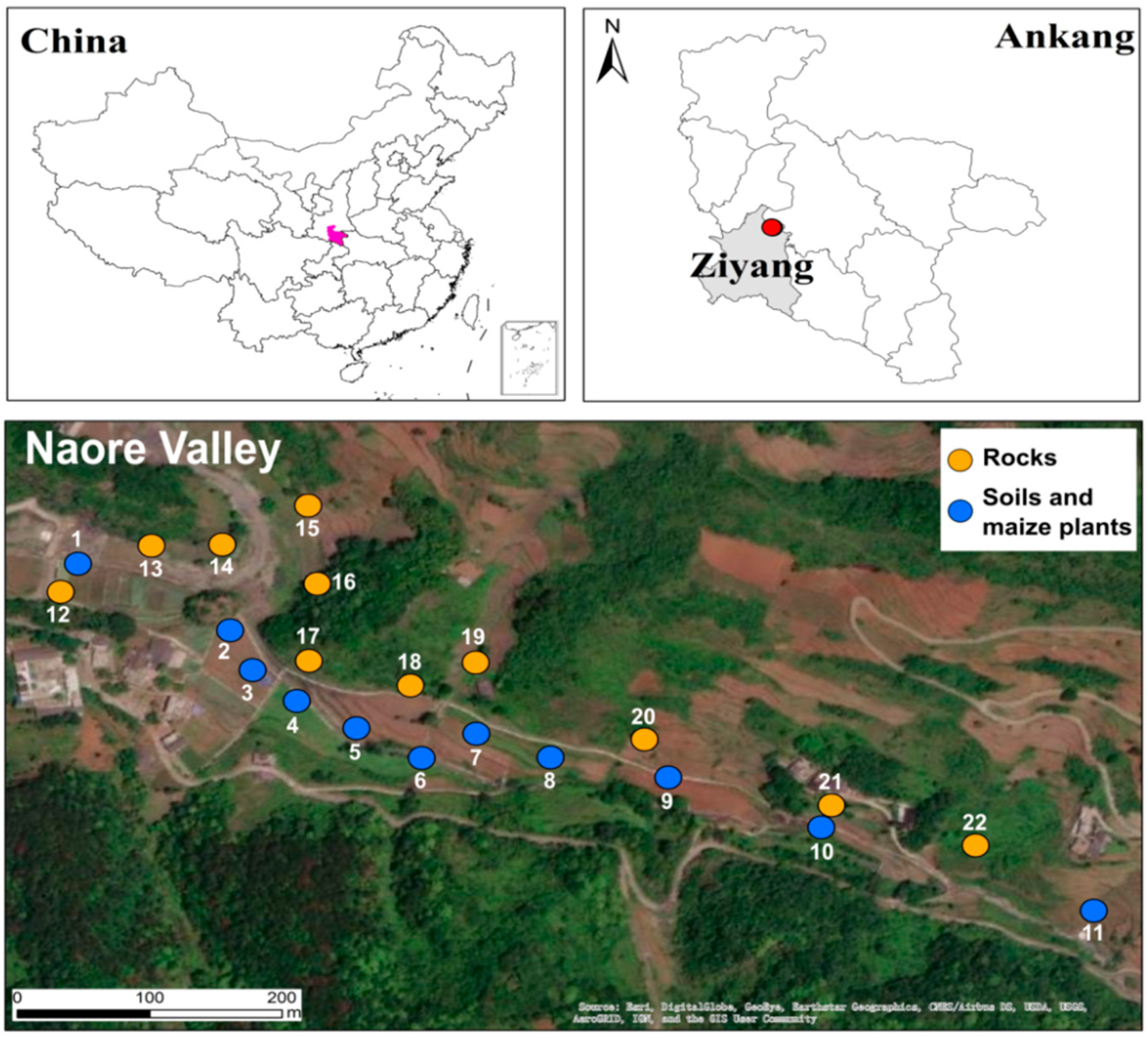
Figure 2.
Total selenium concentrations and proportions of Se in the five operationally defined Se fractions in soil and rock samples across the study area. The bars show means ± sd (n = 3). F1: water-soluble Se, F2: Exchangeable Se, F3: Alkali-soluble Se, F4: Acid soluble Se, and F5: Residual Se.
Figure 2.
Total selenium concentrations and proportions of Se in the five operationally defined Se fractions in soil and rock samples across the study area. The bars show means ± sd (n = 3). F1: water-soluble Se, F2: Exchangeable Se, F3: Alkali-soluble Se, F4: Acid soluble Se, and F5: Residual Se.
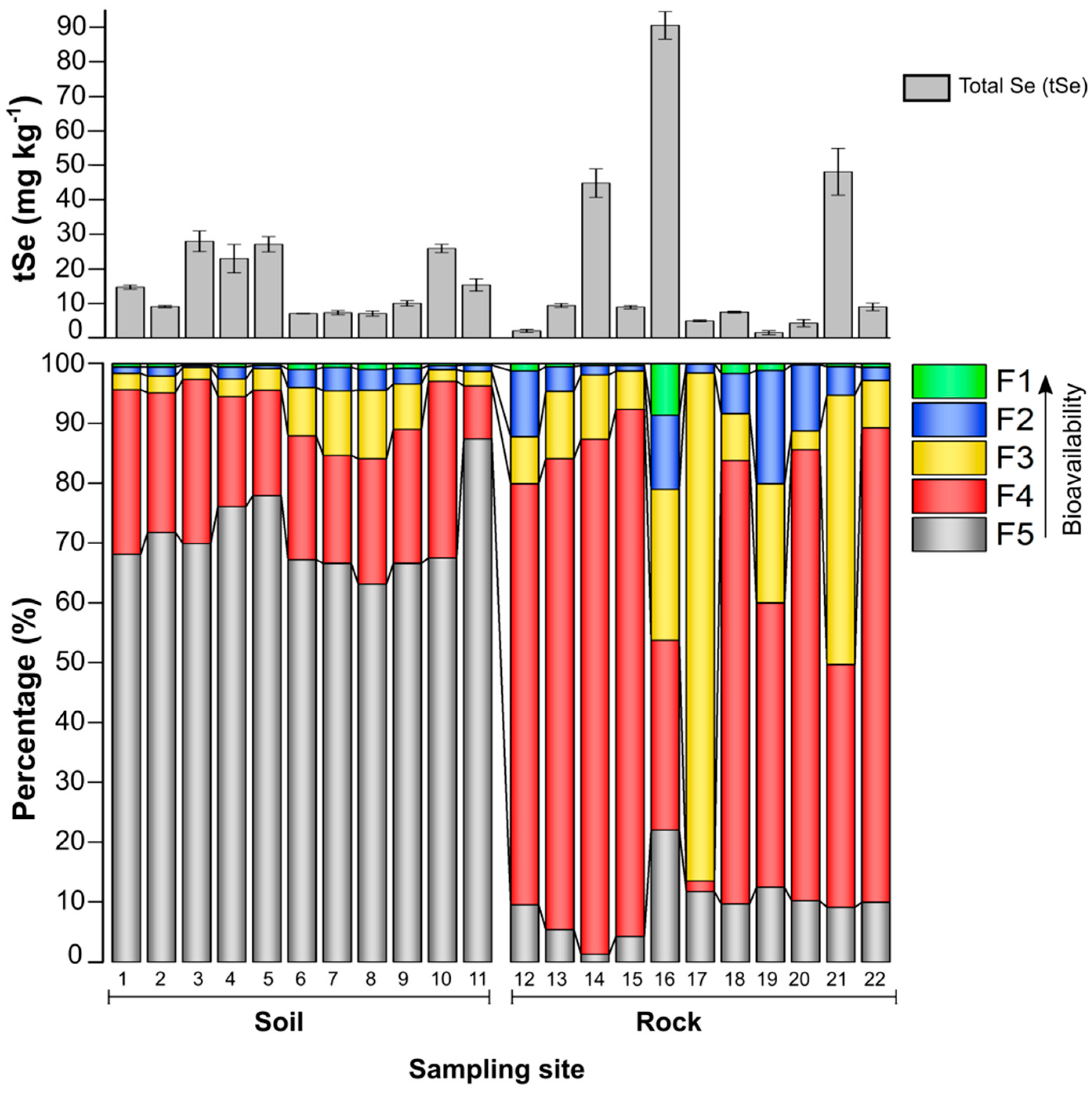
Figure 3.
Total Se concentrations in grain, leaf, stalk, root, soil, and rocks for (n=11) samples in triplicate. The square is the mean value. The limits represent the minimum and maximum values, and the horizontal bottom, middle, and top lines correspond to the 10th percentile, median, and 90th percentile values, respectively.
Figure 3.
Total Se concentrations in grain, leaf, stalk, root, soil, and rocks for (n=11) samples in triplicate. The square is the mean value. The limits represent the minimum and maximum values, and the horizontal bottom, middle, and top lines correspond to the 10th percentile, median, and 90th percentile values, respectively.
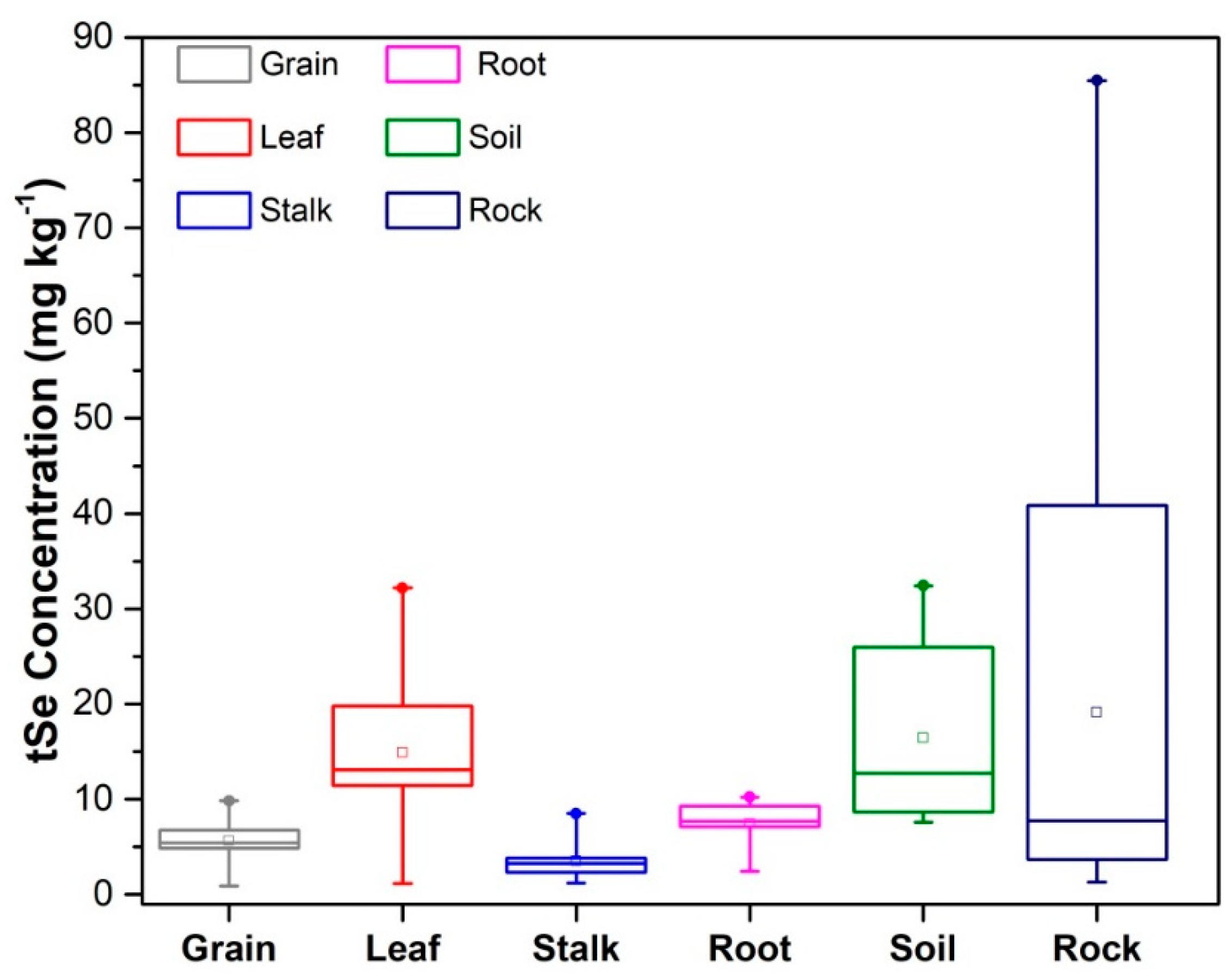
Figure 4.
Distribution of Se enzyme-soluble species (extracted), their sum, and tSe in each maize organ.
Figure 4.
Distribution of Se enzyme-soluble species (extracted), their sum, and tSe in each maize organ.
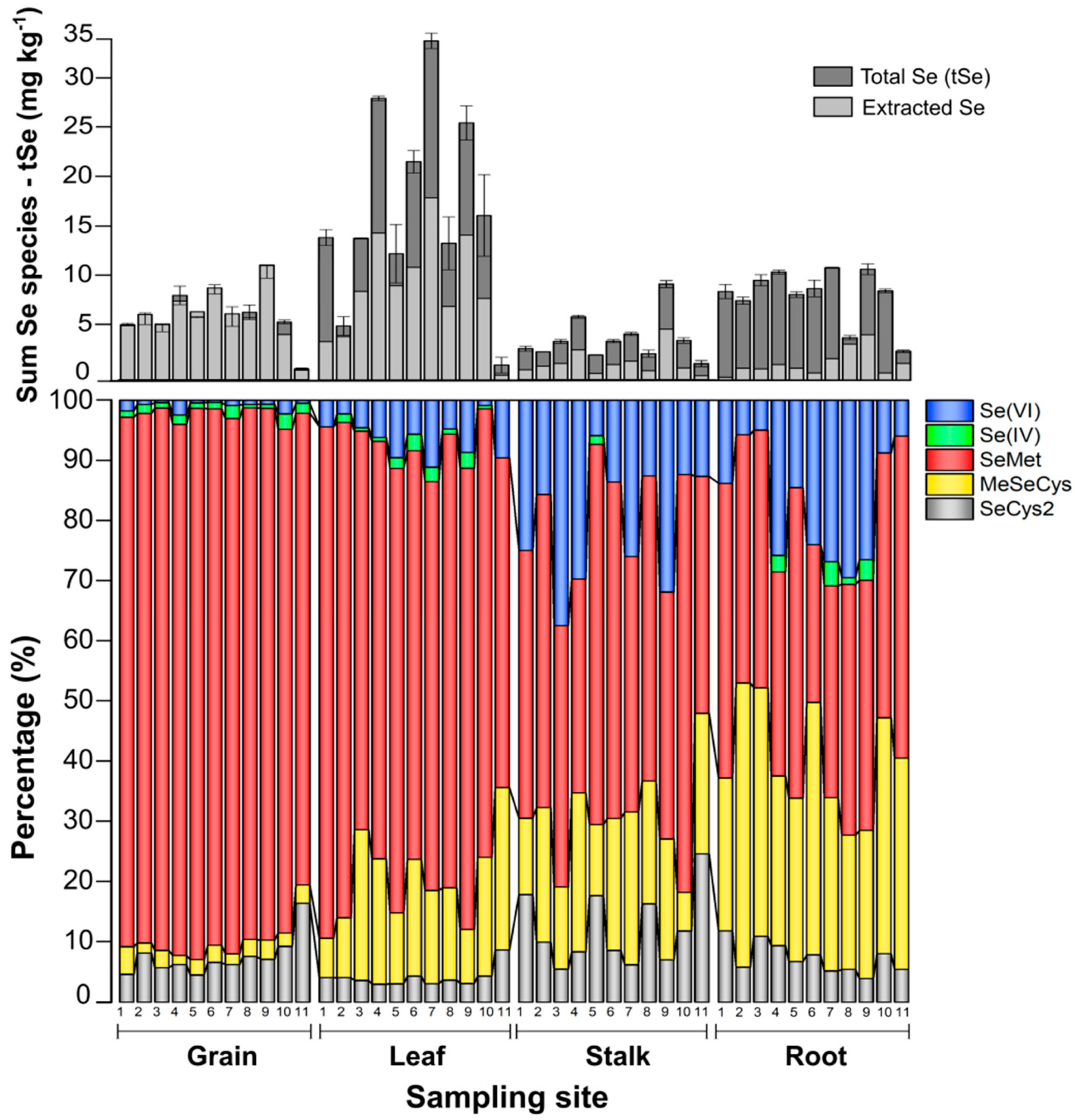
Figure 5.
Regressions between organic and inorganic Se species when compared to tSe contents (*p < 0.05, **p < 0.01, correlation coefficient).
Figure 5.
Regressions between organic and inorganic Se species when compared to tSe contents (*p < 0.05, **p < 0.01, correlation coefficient).
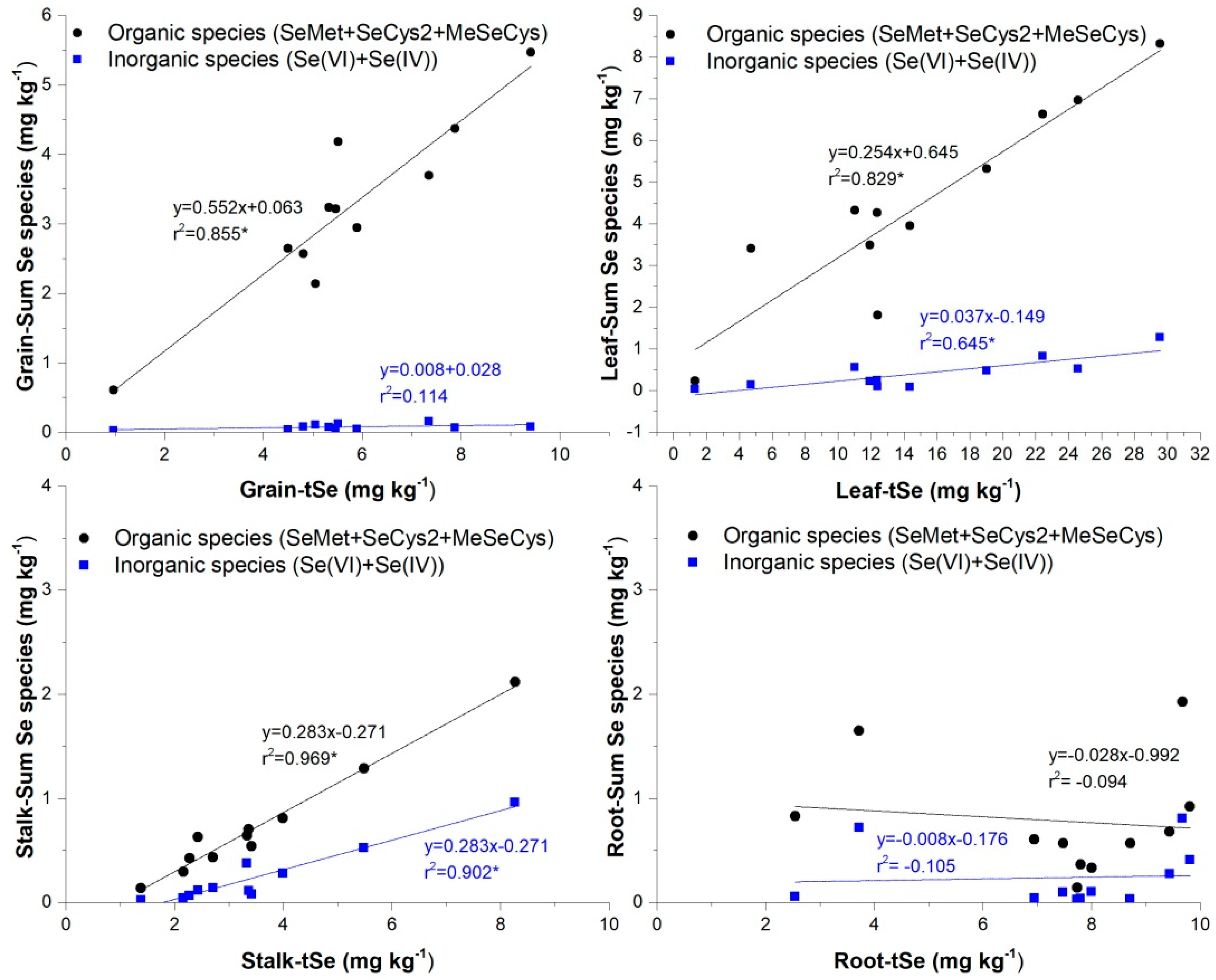

Table 1.
The concentration of tSe in soils, rocks, and pH and TOC, as well as tSe in maize organs and biomasses (dry weight per plant).
Table 1.
The concentration of tSe in soils, rocks, and pH and TOC, as well as tSe in maize organs and biomasses (dry weight per plant).
| Samples | Parameter | Plot 1 | Plot 2 | Plot 3 | Plot 4 | Plot 5 | Plot 6 | Plot 7 | Plot 8 | Plot 9 | Plot 10 | Plot 11 |
| Soil | No. | S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | S11 |
| tSe (mg/kg) | 13.3 ± 0.6 | 8.4 ± 0.3 | 29.4 ± 3.0 | 22.0 ± 4.1 | 29.9 ± 2.2 | 7.8 ± 0.1 | 8.0 ± 0.6 | 8.8 ± 0.7 | 11.1 ± 0.8 | 27.8 ± 1.2 | 14.2 ± 1.8 | |
| pH | 7.5 | 7.7 | 7.9 | 8.7 | 7.7 | 7.6 | 7.7 | 7.3 | 6.5 | 7.5 | 7.6 | |
| TOC | 4.5 | 3.3 | 4.9 | 5.7 | 7.0 | 2.2 | 5.0 | 3.2 | 5.2 | 4.2 | 5.1 | |
| Rock | No. | R12 | R13 | R14 | R15 | R16 | R17 | R18 | R19 | R20 | R21 | R22 |
| tSe (mg/kg) | 2.0 ± 0.4 | 10.0 ± 0.5 | 41.5 ±4.1 | 7.7 ±0.5 | 85.5 ± 4.0 | 3.8 ± 1.0 | 6.1 ± 0.2 | 1.3±0.3 | 3.7 ± 0.6 | 40.9 ± 6.8 | 7.7 ± 1.1 | |
| pH | 9.3 | 9.0 | 8.5 | 8.7 | 8.0 | 8.3 | 9.1 | 9.3 | 9.3 | 6.5 | 8.8 | |
| TOC | 2.1 | 2.0 | 1.2 | 8.4 | 13.1 | 3.4 | 3.1 | 8.1 | 3.8 | 15.0 | 0.2 | |
| Maize | No. | M1 | M2 | M3 | M4 | M5 | M6 | M7 | M8 | M9 | M10 | M11 |
| tSe-grain (mg/kg) | 4.8 ± 0.1 | 5.3 ± 0.5 | 4.5 ± 0.3 | 7.3 ± 0.8 | 5.5 ± 0.05 | 7.9 ± 0.4 | 5.5 ± 0.8 | 5.9 ± 0.63 | 9.4 ± 0.5 | 5.0 ± 0.2 | 1.0 ± 0.08 | |
| tSe-leaf (mg/kg) | 12.4 ± 0.8 | 4.7 ± 0.05 | 12.4 ± 0.2 | 24.6 ± 2.5 | 11.0 ± 1.0 | 19.0 ± 0.7 | 29.6 ± 2.3 | 11.9 ± 1.5 | 22.4 ± 3.5 | 14.3 ± 0.7 | 1.3 ± 0.2 | |
| tSe-stalk (mg/kg) | 2.7 ± 0.2 | 2.4 ± 0.2 | 3.3 ± 0.1 | 5.5 ± 0.03 | 2.2 ± 0.13 | 3.4 ± 0.1 | 4.0 ± 0.3 | 2.3 ± 0.06 | 8.3 ± 0.2 | 3.4 ± 0.04 | 1.4 ± 0.2 | |
| tSe-root (mg/kg) | 7.7 ± 0.6 | 6.9 ± 0.3 | 8.7 ± 0.5 | 9.4 ± 0.2 | 7.5 ± 0.2 | 8.0 ± 0.7 | 9.8 ± 0.05 | 3.7 ± 0.2 | 9.7 ± 0.5 | 7.8 ± 0.2 | 2.5 ±0.1 | |
| Biomass-grain (g) | 414.8 | 385.1 | 282.1 | 298.2 | 101.6 | 322.2 | 369.9 | 356.3 | 515.8 | 188.7 | 415.0 | |
| Biomass-leaf (g) | 94.9 | 106.3 | 88.9 | 63.2 | 100.1 | 146.3 | 128.7 | 125.8 | 145.1 | 84.5 | 143.6 | |
| Biomass-stalk (g) | 94.5 | 93.5 | 136.6 | 121.5 | 98.3 | 106.5 | 124.0 | 116.2 | 114.4 | 145.8 | 133.5 | |
| Biomass-root (g) | 63.0 | 109.62 | 34.2 | 33.8 | 39.3 | 99.5 | 35.8 | 49.0 | 56.2 | 85.5 | 96.2 |
Table 2.
Se species in enzymatic hydrolysis extracts and tSe of maize samples (in Se mg kg-1 and % Se of species for tSe content).
Table 2.
Se species in enzymatic hydrolysis extracts and tSe of maize samples (in Se mg kg-1 and % Se of species for tSe content).
| Maize organ |
tSe | SUM-Se extracted | Organic species | Inorganic species | |||||||
| SeMet | SeCys2 | MeSeCys | Se(IV)+Se(VI) | Extraction efficiency | |||||||
| (mg/kg) | (mg /Kg) | (mg/kg) | (%) | (mg/kg) | (%) | (mg/kg) | (%) | (mg/kg) | (%) | (%) | |
| Grain | 5.6 ± 2.2 | 5.6 ± 2.3 | 5.0 ± 2.1 | 86.7 ± 3.5 | 0.43 ± 0.17 | 7.4 ± 3.3 | 0.13 ± 0.1 | 2.62 ± 0.86 | 0.06 ± 0.03 | 2.3 ± 1.3 | 98.7 ± 9.8 |
| Leaf | 14.9 ± 8.4 | 8.0 ± 4.6 | 6.2 ± 3.5 | 80.0 ± 8.3 | 0.33 ± 0.15 | 4.0 ± 1.6 | 1.2 ± 0.82 | 16.1 ± 6.5 | 0.30 ± 0.28 | 7.3 ± 3.8 | 54.1 ± 15.5 |
| Stalk | 3.5 ± 1.9 | 1.5 ± 1.2 | 0.81 ± 0.57 | 48.1 ± 10.4 | 0.17 ± 0.1 | 12.0 ± 6.0 | 0.30 ± 0.31 | 18.3 ± 6.3 | 0.19 ± 0.21 | 20.1 ± 10.0 | 37.5 ± 10.6 |
| Root | 7.4 ± 2.3 | 1.5 ± 1.1 | 0.76 ± 0.58 | 41.8 ± 7.9 | 0.11 ± 0.07 | 7.3 ± 2.5 | 0.46 ± 0.29 | 32.7 ± 8.4 | 0.18 ± 0.21 | 17.9 ± 10.7 | 25.4 ± 25.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



