
Submitted:
13 January 2023
Posted:
17 January 2023
You are already at the latest version
Abstract
The emergence of drug-resistant tuberculosis forced the development of new drugs and screnning of more effective or less toxic analogs. Mycolic acids biosynthesis is targeted by several antituberculosis drugs toped by the isoniazid being one of the most important in tuberculosis therapy. Recently, perchlozone, acting on another step in FAS-II cycle, received official approval and was included in the Russian national clinical guidelines. Using the serial dilution method on agar 7H10 plates for perchlozone and Sensititre MYCOTB microdilution plate we analyzed the phenotypic properties of primary M. tuberculosis clinical isolates and analyzed the molecular determinants of resistance to isoniazid, ethionamide and perchlozone. We found a wide variation in the MIC of perchlozone from 2 to 64 mg/L, correlating with the overall resistance profile: the MIC was higher for MDR and pre-XDR isolates. The cross-resistance between the ethionamide and the perchlozone was driven by mutations in ethA gene encoding monooxygenase responsible for activation of both drugs. Presumably susceptible to perchlozone wild-type strains had MICs ranged 2–4 mg/L, and the breakpoint estimated to be at 4 or 8 mg/L. In conclusion, susceptibility to perchlozone retained for a part of MDR strains, as does suceptibility to ethionamide, providing the possibility of therapy for such cases based on phenotypic or molecular analysis.
Keywords:
tuberculosis
; drug resistance
; molecular determinants
; perchlozone
; ethionamide
; isoniazid
; thioacetozone
1. Introduction
Tuberculosis currently remains a major public health problem worldwide. The transmission of drug-resistant strains of M. tuberculosis are becoming common in many regions of the world. The numbers of multidrug-resistant (MDR) and extensively drug-resistant (XDR) tuberculosis primary cases are steadily increasing. The chemotherapy regimens for such patients need to be modified in a timely manner to include effective drugs.
Thiosemicarbazones are a group of chemical compounds with mycobactericidal activity, discovered upon the systematic investigation of sulfonamides and thiazoles as anti-tuberculosis agents [1]. The best known representative of this group was thioacetazone, also known as Tibione or Tb 1, the first synthetic anti-tuberculosis drug, which began be used in the late 1940s. It was widely tested and used in postwar Europe, despite adverse reactions such as severe gastrointestinal, hepatic, and renal disorders. Subsequently, the high toxicity of tibone forced the search for less toxic analogs. In 1952, three isomeric forms of nicotinaldehyde thiosemicarbazone in the alpha, beta and gamma positions were produced [2]. The betta and gamma forms were less toxic than thioacetazone. The thiosemicarbazone isonicotinaldehyde (gamma isomer) obtained at that time is the precursor of the modern perchlozone. However, simultaneously, the high anti-tuberculosis activity of isonicotine hydrazine (isoniazid), which has lower toxicity, was also shown [3]. This led to an intensive study on isoniazid as one of the most effective anti-tuberculosis drugs of the time.
The emergence of drug-resistant tuberculosis has pushed forward the development of new drugs, also as the studies of repurposed and previously described less effective drugs [4]. Thioureidominomethylpyridinium perchlorate or perchlozone received official approval in November 2012 and was included in the Russian national clinical guidelines as a treatment for multidrug-resistant and extensively drug-resistant tuberculosis patients. However, only a few studies on perchlozone MICs for clinical M. tuberculosis strains and their molecular mechanisms of action have been performed [5].
The mechanism of action of the perchlozone was revealed by Pooja Gopal and Thomas Dick, who found that HadABC is the target of perchlozone activated in the cell by EthA (Figure 1) [6]. The activation path by EthA monooxygenase is also shared by thioacetazone and ethionamide, and the possibility of cross-resistance between perchlozone, thioacetazone and ethionamide by inactivating mutations in the ethA locus has also been proposed [6].
In this report, we describe the phenotypic and genetic characteristics of clinical isolates of M. tuberculosis from the Moscow region with variable susceptibility to perchlozone, ethionamide, and isoniazid. Combining phenotypic and molecular data, we estimated the proportion of drug-resistant isolates suitable for perchlozone treatment.
2. Results
Clinical isolates from 22 patients were analyzed in this study. Samples were obtained in 2017–2018 from patients who attended the Moscow Research and Clinical Center for Tuberculosis Control. Eight of the 22 patients had susceptible form of tuberculosis, twelve patients had MDR-TB, and two had pre-XDR-TB, detrmined as additional resistance to fluoroquinolones (Table S1).
Isolates were genotyped by the 24-loci MIRU-VNTR method, and all samples belonged to the L2 Beijing lineage. Most of the isolates were from the modern clonal complexes CC1 and CC2 (Table S1) [7]. Seven isolates Central Asian/Russian Beijing cluster 94-32 were predominately susceptible, whereas six isolates of Beijing B0/W-148 cluster (MIRU type 100-32) were MDR of pre-XDR.
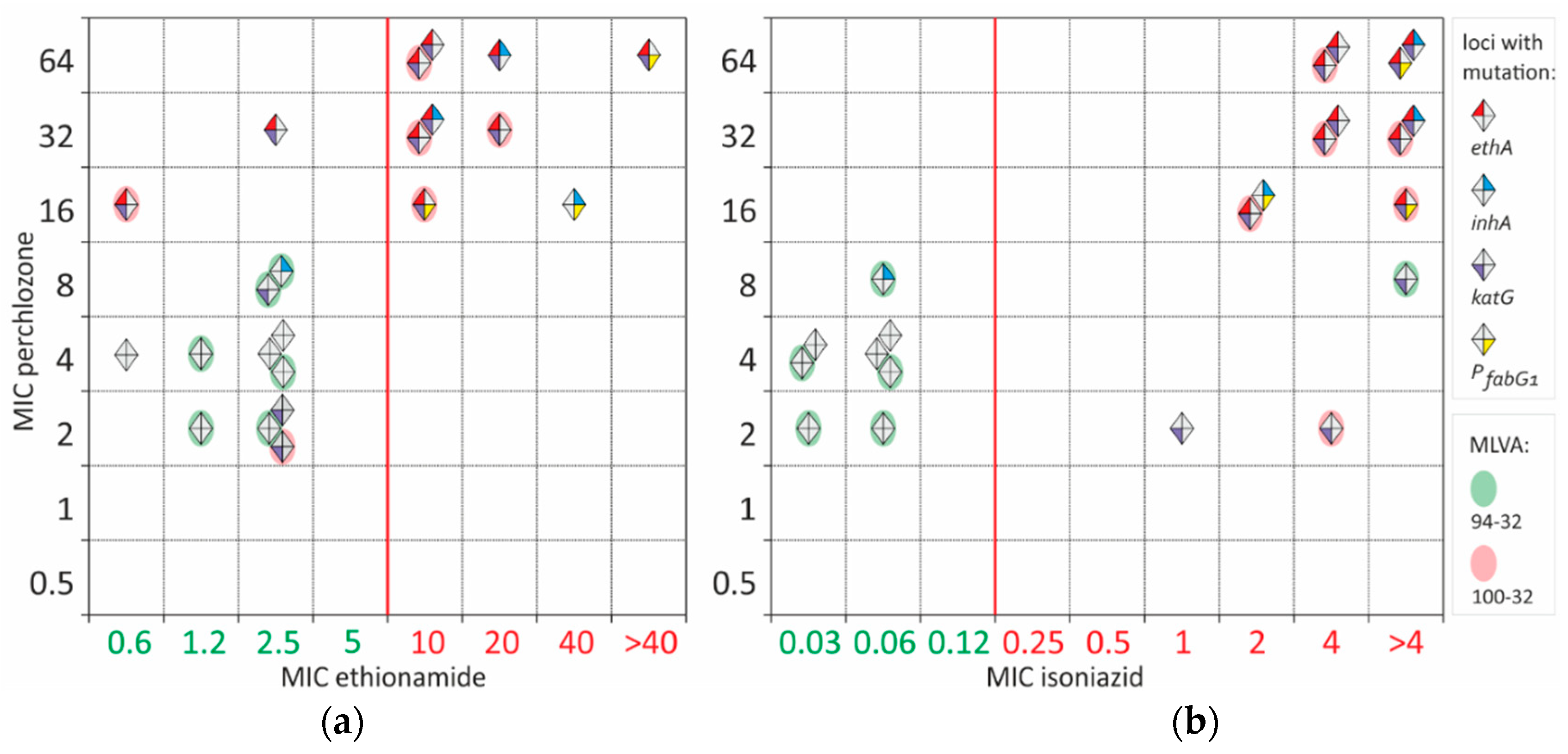
Resistance to ethionamide and high MICs of perchlozone were identified only in MDR isolates. The observed MIC of perchlozone spanned from 2 to 64 mg/L, with a statistically insignificant difference in isolate numbers at each concentration.
Common activation path of perchlozone and ethionamide by EthA lead to at least partial cross-resistance between these drugs (Figure 2). All ethionamide-resistant isolates had also high perchlozone MIC. Most of resistant isolates had an altered ethA sequence. However, two samples with mutations in ethA and a high MIC of perchlozone (16 and 32 mg/L) retained low ethionamide MIC of 0.6 and 2.5 mg/L, which were below the breakpoint. One of them bore EthA L440P substitution with an unknown effect on protein function, while the other had a frameshift mutation at 880 position. The latter was resistant to ethionamide as determined by critical concentration at 5 mg/L.
Pairwise correlation of isoniazid and perchlozone MIC revealed partial cross-correlation (Figure 2b). Isoniazid resistance could be explained by KatG S315T substitution in 13 of 14 isolates. Only one resistant isolate had PfabG1 mutation and InhA I194V substituon. Two isoniazid-resistant and presumably perchlozone-susceptible isolates had perchlozone MIC of 2 mg/L. No other mutations besides KatG S315T were found.
Two isolates had an ‘intermediate’ perchlozone MIC of 8 mg/L. One was susceptible to isoniazid and had F97L substitution in InhA. It was not previously described in resistant isolates and could be neutral. The second was isoniazid-resistant and had S315T substitution in KatG.
Some discrepancies between ethA mutations and phenotype existed for three isolates with perchlozone MIC of 16mg/L. One strain had no mutations in the ethA gene, and high MICs of ethionamide and isoniazid could be explained by frequently identified promoter PfabG1 mutation c(-15)t and InhA substituton I194V. Additional sequencing of the hadABC locus did not reveal any mutations, thus, high perchlozone MIC is caused by alterations in other genomic loci.
Nine presumable susceptible to perchlozone isolates with MICs of 2-4 mg/L had no mutations in ethA. They are also characterized by low ethionamide MICs ranging from 0.6 to 2.5 mg/L (Figure 2). Control M. tuberculosis H37Rv strain had perchlozone MIC of 4 mg/L.
3. Discussion
Treatment of MDR-TB is challenging due to the limited number of effective drugs available and severe adverse drug reactions. Additionaly, cross-resistance between antituberculosis drugs may limit their efficacy. Besides a common target of action for structurally similar drugs, such as in the rifamycins or fluoroquinolones groups, cross-resistance mechanisms may be due to common pathways of drug efflux or prodrug activation. For example, mutations in the transcriptional repressor gene rv0678 lead to cross-resistance to bedaquiline and clofazimine due to derepression of mmpL5-mmpS5 efflux pump operon [8]. Even more frightening is a recent report that the same type of mutations may also lead to low-level resistance to promising new drugs in the benzothiazinone group PBTZ169 and BTZ043 [9].
Perchlozone is a relatively new drug approved and used in the Russian Federation since 2012. Being a member of the thiosemicarbazone family, it has a mechanism of action similar to that of thioacetozone, according to the experiments on in vitro selection of resistant mutants [6]. Perchlozone is a prodrug and is activated by EthA monooxygenase also as ethionamide, thioacetazone, and isoxyl [10], so cross-resistance between currently used in tuberculosis treatment perchlozone and ethionamide has been suggested [6].
We found a wide variation in the MIC of perchlozone from 2 to 64 mg/L, correlating with the overall resistance profile of unexposed clinical strain: the MIC was higher for MDR and pre-XDR isolates. For presumably susceptible wild-type strains, the MIC of perchlozone was 2–4 mg/L. Gopal and Dick reported the perchlozone MIC50 value measured by optical density in 7H9 liquid medium equal to 0.04 mg/L for M. tuberculosis and 3.6 mg/L for M. bovis BCG. These values are underestimated in comparison to the MIC defined in the standard way [6]. The data for agar medium were based on 99% growth inhibition and for M. bovis the MIC of perchlozone was 18.2 mg/L. In an in vitro efficacy study of thioacetasone analogs, the MIC of for a structurally matched compound (ID=7) for M. tuberculosis strain H37Rv was 0.8 mg/L, which is comparable to our data [11]. It is worth noting that perchlozone was 20 times less effective than thioacetazone in the Gopal study [6] and 8 times less effective in the Alahari study [11]. The average plasma concentrations from pharmacokinetic studies at doses ranging from 800 to 1600 mg/day for patients of different body weights was 23.4 mg/L [12]. Thus, the preliminary breakpoint concentration of perchlozone on Middlebrook 7H10 agar medium could be estimated as 4-8 mg/L.
In our work, we showed the cross-resistance between the ethionamide and the perchlozone in the primary non-exposed to perchlozone M. tuberculosis clinical isolates. Mutations associated with resistance were localized in the gene encoding EthA monooxygenase, and all strains with a high MIC of ethionamide also had a high MIC of the perchlozone. A previous study on a set of clinical isolates lacked phenotypic data on perchlozone susceptibility, which limits the interpretation of the results [13]. However, in one case, in concordance with the proposal by Gopal and Dick and our findings, an additional frameshift mutation in the ethA gene emerged during treatment with the perchlozone. It could be proposed that isolates with elevated MIC of ethionamide and frameshift mutations in the ethA gene should be considered resistant and such cases should not be treated with the perchlozone.
The observed incomplete cross-resistance between isoniazid and perchlozone was due to the properties of the strains chosen for the study, since all strains with high MIC of isoniazid and perchlozone carry mutations in ethA and were resistant to ethionamide. Generally, susceptibility to perchlozone retained for a part of MDR strains, as does suceptibility to ethionamide [14,15,16], providing the possibility of therapy for such cases based on phenotypic or molecular analysis.
Other mutations could be the cause of elevated perchlozone MIC. We analyzed the sequence of HadABC locus in one isolate with MIC of 8 mg/L and wild-type ethA, but it was also unmutated. Another possible mechanism could be caused by the structural similarity of perchlozone and thioacetazone. Alahari, 2007 showed the effect of thioacetazone on the cyclopropane mycolic acid synthases family of mycobacterial enzymes responsible for the synthesis of different subtypes of mycolic acids. In the proposed model, thioacetozone inhibits MmaA2 and activated by MmaA4 [11,17], which are involved in the production of the mycolic keto acids. An independent confirmation was obtained by in vitro selection of resistant M. tuberculosis strains, and obtained strains bore mutations in hadA, hadC, and mmaA4 [18].
A limitation of this study is the relatively small sample size of 22 isolates. Nevertheless, this is the first study of clinical M. tuberculosis clinical isolates, that combined phenotypic and molecular data and confirmed the basic assumptions about the mechanisms of perchlozone resistance.
4. Materials and Methods
4.1. Mycobacterium tuberculosis Strains
The M. tuberculosis isolates were obtained from clinical specimens collected from patients with TB at the Moscow Research and Clinical Center for Tuberculosis Control. All clinical strains were isolated in 2017–2018 from sputum of newly diagnosed patients before the start of anti-tuberculosis therapy or its long-term lasting no more than one month. The laboratory reference strain M. tuberculosis Н37Rv was used as a control strain. For molecular analysis samples were isolated sequentially, one isolate from one patient, before treatment initiation. Drug susceptibility testing for rifampicin, isoniazid, streptomycin, ethambutol, pyrazinamide, ofloxacin, moxifloxacin, kanamycin, capreomycin, and amikacin, PAS, ethionamide was performed using Bactec MGIT 960 as previously described [19,20]. Sensititre MYCOTB MIC determination was performed as described in [21]. The MIC of perchlozone was determined using serial dilutions method on Middlerook 7H10 agar plates supple-mented with the drug at the following concentrations: 0.5, 1, 2, 4, 8, 16, 32, 64 mg/L.
4.2. DNA Isolation and Sequencing
DNA isolation and sequencing of the ethA, inhA and PfabG1 fragments were performed as previously described [48]. The following PCR primers were used: ethAR-F1: 5′-cgacgttgaaatcacgctgg-3′, ethAR-R1: 5′-gtgaccgacaccattgaacg-3′; ethAR-F2: 5′-ttcaaccccgttgcggtaat-3′; ethAR-R2: 5′-ctctttctgtgcagcggcta-3′; ethAR-F3: 5′-atgatcggcccgacgaaatc-3′; ethAR-R3: 5′-ccctggcagcttactacgtg-3′; PfabG1-F: 5′-cctcgctgcccagaaaggga-3′; PfabG1-R: 5′-atcccccggtttcctccggt-3′; inhA-F2: 5′-gagctatatctccggtgcgg-3′; inhA-R2: 5′-gcgaccgtcatccagttgta-3′; inhA-F3: 5′-ccacatctcggcgtattcgt-3′; inhA-R3: 5′-cggtgataccccaccgaaat-3′.
4.3. MIRU-VNTR typing
Twenty-four loci MIRU-VNTR typing was made according to [22]. Profiles in the article are given in the following order: MIRU04(ETRD-1), MIRU26, MIRU40, MIRU10, MIRU16, MIRU31(ETRE), Mtub04, ETRC, ETR-A, Mtub30, Mtub39, Qub4156, Qub11b, Mtub21, Qub26, MIRU02, MIRU23, MIRU39, MIRU20, MIRU24, MIRU27 (Qub5), Mtub29, ETRB, Mtub34. The MLVA types were compared to MIRU-VNTRplus online (http://www.miru-vntrplus.org/) [23].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Summary of characterized clinical M. tuberculosis isolates.
Author Contributions
Conceptualization, D.Z. and S.S.; methodology, D.G., Y.M. and A.U.; validation, A.U.; formal analysis, A.U., Y.M. and L.K.; investigation, A.U., Y.M. and L.K.; data curation, M.M. and S.S.; writing—original draft preparation, A.U., review and editing, D.Z.; funding acquisition, D.Z.; supervision, S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Foundation for Basic Research, grant number 20-015-00463.
Institutional Review Board Statement
The study was approved by the Ethics Committee of the Moscow Government Health Department. The Ethics Committee waived the need for patient consent because the study did not include personal identifiers or clinical data and the samples were analyzed anonymously.
Data Availability Statement
Data is contained within the article or Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Domagk, G. Investigations on the Antituberculous Activity of the Thiosemicarbazones in Vitro and in Vivo. Am Rev Tuberc 1950, 61, 8–19.
- Fox, H.H. SYNTHETIC TUBERCULOSTATS. III. ISONICOTINALDEHYDE THIOSEMICARBAZONE AND SOME RELATED COMPOUNDS. J. Org. Chem. 1952, 17, 555–562. [CrossRef]
- Fox, H.H. The Chemical Attack on Tuberculosis. Trans N Y Acad Sci 1953, 15, 234–242. [CrossRef]
- Barry, C.E.; Slayden, R.A.; Sampson, A.E.; Lee, R.E. Use of Genomics and Combinatorial Chemistry in the Development of New Antimycobacterial Drugs. Biochem Pharmacol 2000, 59, 221–231. [CrossRef]
- Malík, I.; Čižmárik, J.; Pecháčová, M. Focus on Perchlozone, an Anti-Tuberculosis Drug from the Russian Federation. Ceska Slov Farm 2020, 69, 203–210.
- Gopal, P.; Dick, T. The New Tuberculosis Drug Perchlozone® Shows Cross-Resistance with Thiacetazone. Int J Antimicrob Agents 2015, 45, 430–433. [CrossRef]
- Merker, M.; Blin, C.; Mona, S.; Duforet-Frebourg, N.; Lecher, S.; Willery, E.; Blum, M.G.B.; Rüsch-Gerdes, S.; Mokrousov, I.; Aleksic, E.; et al. Evolutionary History and Global Spread of the Mycobacterium Tuberculosis Beijing Lineage. Nat Genet 2015, 47, 242–249. [CrossRef]
- Hartkoorn, R.C.; Uplekar, S.; Cole, S.T. Cross-Resistance between Clofazimine and Bedaquiline through Upregulation of MmpL5 in Mycobacterium Tuberculosis. Antimicrob Agents Chemother 2014, 58, 2979–2981. [CrossRef]
- Poulton, N.C.; Azadian, Z.A.; DeJesus, M.A.; Rock, J.M. Mutations in Rv0678 Confer Low-Level Resistance to Benzothiazinone DprE1 Inhibitors in Mycobacterium Tuberculosis. Antimicrob Agents Chemother 2022, 66, e0090422. [CrossRef]
- DeBarber, A.E.; Mdluli, K.; Bosman, M.; Bekker, L.G.; Barry, C.E. Ethionamide Activation and Sensitivity in Multidrug-Resistant Mycobacterium Tuberculosis. Proc Natl Acad Sci U S A 2000, 97, 9677–9682. [CrossRef]
- Alahari, A.; Trivelli, X.; Guérardel, Y.; Dover, L.G.; Besra, G.S.; Sacchettini, J.C.; Reynolds, R.C.; Coxon, G.D.; Kremer, L. Thiacetazone, an Antitubercular Drug That Inhibits Cyclopropanation of Cell Wall Mycolic Acids in Mycobacteria. PLoS One 2007, 2, e1343. [CrossRef]
- Yablonskiy, P.K.; Vinogradova, T.I.; Levashev, Yu.N.; Pavlova, M.V.; Zilber, E.K.; Starshinova, A.A.; Sapozhnikova, N.V.; Chernokhaeva, I.V.; Archakova, L.I.; Zabolotnykh, N.V.; et al. Preclinical and Clinical Trials of the New Tuberculosis Drug Perchlozon. Ter. arkh. 2016, 88, 111. [CrossRef]
- Mokrousov, I.; Vyazovaya, A.; Akhmedova, G.; Solovieva, N.; Turkin, E.; Zhuravlev, V. Genetic Variation Putatively Associated with Mycobacterium Tuberculosis Resistance to Perchlozone, a New Thiosemicarbazone: Clues from Whole Genome Sequencing and Implications for Treatment of Multidrug-Resistant Tuberculosis. Antibiotics (Basel) 2020, 9, E669. [CrossRef]
- Morlock, G.P.; Metchock, B.; Sikes, D.; Crawford, J.T.; Cooksey, R.C. EthA, InhA, and KatG Loci of Ethionamide-Resistant Clinical Mycobacterium Tuberculosis Isolates. Antimicrob Agents Chemother 2003, 47, 3799–3805. [CrossRef]
- Vilchèze, C.; Jacobs, W.R. Resistance to Isoniazid and Ethionamide in Mycobacterium Tuberculosis: Genes, Mutations, and Causalities. Microbiol Spectr 2014, 2, MGM2-0014–2013. [CrossRef]
- Ushtanit, A.; Kulagina, E.; Mikhailova, Y.; Makarova, M.; Safonova, S.; Zimenkov, D. Molecular Determinants of Ethionamide Resistance in Clinical Isolates of Mycobacterium Tuberculosis. Antibiotics (Basel) 2022, 11, 133. [CrossRef]
- Alahari, A.; Alibaud, L.; Trivelli, X.; Gupta, R.; Lamichhane, G.; Reynolds, R.C.; Bishai, W.R.; Guerardel, Y.; Kremer, L. Mycolic Acid Methyltransferase, MmaA4, Is Necessary for Thiacetazone Susceptibility in Mycobacterium Tuberculosis. Mol Microbiol 2009, 71, 1263–1277. [CrossRef]
- Coxon, G.D.; Craig, D.; Corrales, R.M.; Vialla, E.; Gannoun-Zaki, L.; Kremer, L. Synthesis, Antitubercular Activity and Mechanism of Resistance of Highly Effective Thiacetazone Analogues. PLoS One 2013, 8, e53162. [CrossRef]
- Sharma, M.; Thibert, L.; Chedore, P.; Shandro, C.; Jamieson, F.; Tyrrell, G.; Christianson, S.; Soualhine, H.; Wolfe, J. Canadian Multicenter Laboratory Study for Standardized Second-Line Antimicrobial Susceptibility Testing of Mycobacterium Tuberculosis. J Clin Microbiol 2011, 49, 4112–4116. [CrossRef]
- Barrera, L.; Cooreman, E.; de Dieu Iragena, J.; Drobniewski, F.; Duda, P.; Havelkova, M.; Hoffner, S.; Kam, K.M.; Kim, S.J.; Labelle, S.; et al. Policy Guidance on Drug-Susceptibility Testing (DST) of Second-Line Antituberculosis Drugs; WHO Guidelines Approved by the Guidelines Review Committee; World Health Organization: Geneva, 2008;
- Nosova, E.Y.; Zimenkov, D.V.; Khakhalina, A.A.; Isakova, A.I.; Krylova, L.Y.; Makarova, M.V.; Galkina, K.Y.; Krasnova, M.A.; Safonova, S.G.; Litvinov, V.I.; et al. A Comparison of the Sensititre MycoTB Plate, the Bactec MGIT 960, and a Microarray-Based Molecular Assay for the Detection of Drug Resistance in Clinical Mycobacterium Tuberculosis Isolates in Moscow, Russia. PLoS One 2016, 11, e0167093. [CrossRef]
- Supply, P.; Allix, C.; Lesjean, S.; Cardoso-Oelemann, M.; Rüsch-Gerdes, S.; Willery, E.; Savine, E.; de Haas, P.; van Deutekom, H.; Roring, S.; et al. Proposal for Standardization of Optimized Mycobacterial Interspersed Repetitive Unit-Variable-Number Tandem Repeat Typing of Mycobacterium Tuberculosis. J Clin Microbiol 2006, 44, 4498–4510. [CrossRef]
- Allix-Béguec, C.; Harmsen, D.; Weniger, T.; Supply, P.; Niemann, S. Evaluation and Strategy for Use of MIRU-VNTRplus, a Multifunctional Database for Online Analysis of Genotyping Data and Phylogenetic Identification of Mycobacterium Tuberculosis Complex Isolates. J Clin Microbiol 2008, 46, 2692–2699. [CrossRef]
Figure 1.
Simplified model of isoniazid, ethionamide, perchlozone an thioacetazone action on M. tuberculosis. The drugs are activated either by EthA monooxygenase or KatG katalase-peroxidase activities. Ethionamide is also could be activated by MymA, Rv0565c, and Rv077c, thioaceatazone could also be activated by MmaA4. Toxic drug adducts with NADH inhibit the enzymes of FAS-II cycle of mycolic acid biosynthesis. The transcription of the inhA gene is driven by the promoter located upstream the fabG1 gene. Pyridine ans thiosemicarbazone groups are marked with red and blue colors, respectvely.
Figure 1.
Simplified model of isoniazid, ethionamide, perchlozone an thioacetazone action on M. tuberculosis. The drugs are activated either by EthA monooxygenase or KatG katalase-peroxidase activities. Ethionamide is also could be activated by MymA, Rv0565c, and Rv077c, thioaceatazone could also be activated by MmaA4. Toxic drug adducts with NADH inhibit the enzymes of FAS-II cycle of mycolic acid biosynthesis. The transcription of the inhA gene is driven by the promoter located upstream the fabG1 gene. Pyridine ans thiosemicarbazone groups are marked with red and blue colors, respectvely.
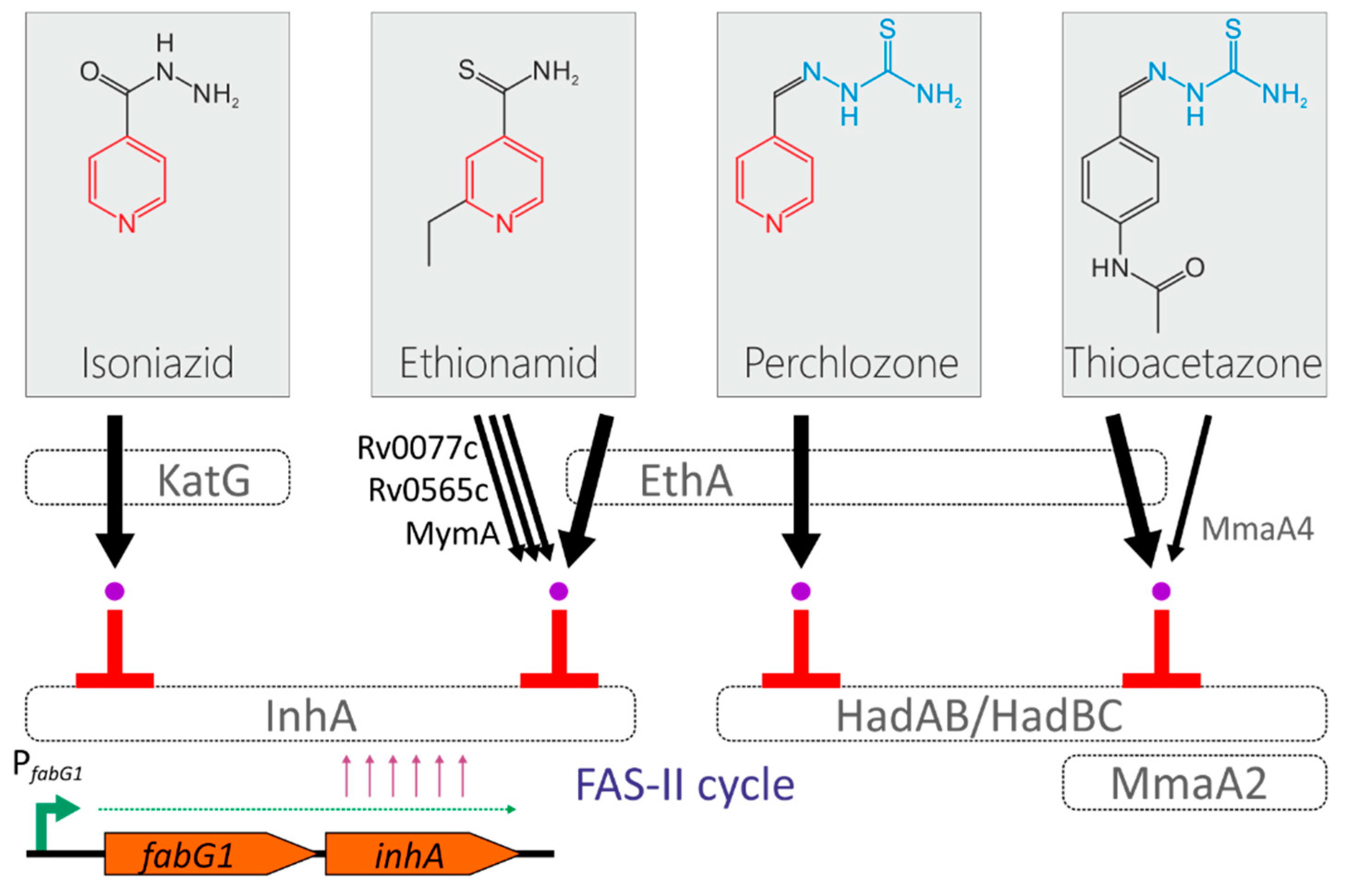
Figure 2.
Pairwise MIC correlation of perchlozone with ethionamide (a) and isoniazid (b). Each isolate is shown with diamond. Isolates with wild-type sequences are marked with gray. Isolates with mutations in ethA are represented by a gray diamond with orange label. Isolates with mutations in inhA are given in blue label. Isolates with mutations in katG are given in purple label and with mutations in fabG1 promoter are given in green label. Isolates with two main multi-locus variable number tandem repeat analysis (MLVA) types 94-32 and 100-32 are marked with green and red ovals.
Figure 2.
Pairwise MIC correlation of perchlozone with ethionamide (a) and isoniazid (b). Each isolate is shown with diamond. Isolates with wild-type sequences are marked with gray. Isolates with mutations in ethA are represented by a gray diamond with orange label. Isolates with mutations in inhA are given in blue label. Isolates with mutations in katG are given in purple label and with mutations in fabG1 promoter are given in green label. Isolates with two main multi-locus variable number tandem repeat analysis (MLVA) types 94-32 and 100-32 are marked with green and red ovals.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



