
Submitted:
16 January 2023
Posted:
19 January 2023
You are already at the latest version
Abstract
G-protein coupled receptors (GPCRs), one of the largest superfamilies of cell-surface receptors, are heptahelical integral membrane proteins that play critical roles in virtually every organ system. G-protein-coupled receptors operate in membranes rich in cholesterol, with an imbalance in cholesterol level within the vicinity of GPCR transmembrane domains affecting the structure and/or function of many GPCRs, a phenomenon that has been linked to several diseases. These effects of cholesterol could result in indirect changes by altering the mechanical properties of the lipid environment or direct changes by binding to specific sites on the protein. There are a number of studies and reviews on how cholesterol modulates class A GPCRs, however, this area of study is yet to be explored for class C GPCRs, which are characterized by a large extracellular region and often form constitutive dimers. This review highlights specific sites of interaction, functions, and structural dynamics involved in the cholesterol recognition of the class C GPCRs. We summarize recent data from some typical family members to explain the effects of membrane cholesterol on the structural features and functions of Class C GPCRs and speculate on their corresponding therapeutic potential.
Keywords:
G-protein-coupled-receptors
; GPCRs
; Membrane protein
; Protein-lipid interactions
; Cholesterol
; Class C GPCRs
1. Introduction
Many recent studies are geared towards deciphering the structures of G-protein coupled receptors (GPCRs) through several methods, most commonly crystallography and cryogenic electron microscopy (Cryo-EM). This is simply because a large number of medications are designed to target GPCRs due to their central role in many biological functions. Some members of GPCRs exist and function as monomers, especially within the class A GPCRs. However, most GPCRs in a lipid bilayer are not stand-alone molecules; rather, they function as dimers and oligomers with themselves or other GPCRs, and with lipids and sterols from the bilayer[1], most notably cholesterol. Most recently, several determined GPCR structures often existing as dimers, appear to indicate the presence of cholesterol. A notable example is the 2-adrenergic receptor (2AR), a class A GPCR which has been crystallized with cholesterol molecules, and a component of the dimer interface consisting of the addition of post-translational palmitate groups from each protomer[2]. This suggests that in GPCR dimerization, there are other responsibilities for lipids and sterols besides protein-protein interactions[3]. GPCRs also thrive within cholesterol-rich membranes, and an insufficient or excessive amount of cholesterol within the membrane could induce conformational changes in many GPCRs which would result in various diseases[4]. These effects of cholesterol could result in indirect changes by altering the mechanical properties of the lipid environment or direct changes by binding to specific sites on the protein[5,6,7]. There are a number of studies and reviews on how cholesterol modulates class A GPCRs, but this area of study is yet to be fully explored for class C GPCRs. Recent advances in experimental and computational power have enabled researchers to investigate the role of lipids in various membranes and solvable proteins, at the atomic level using molecular dynamics simulation[8,9,10,11,12,13].
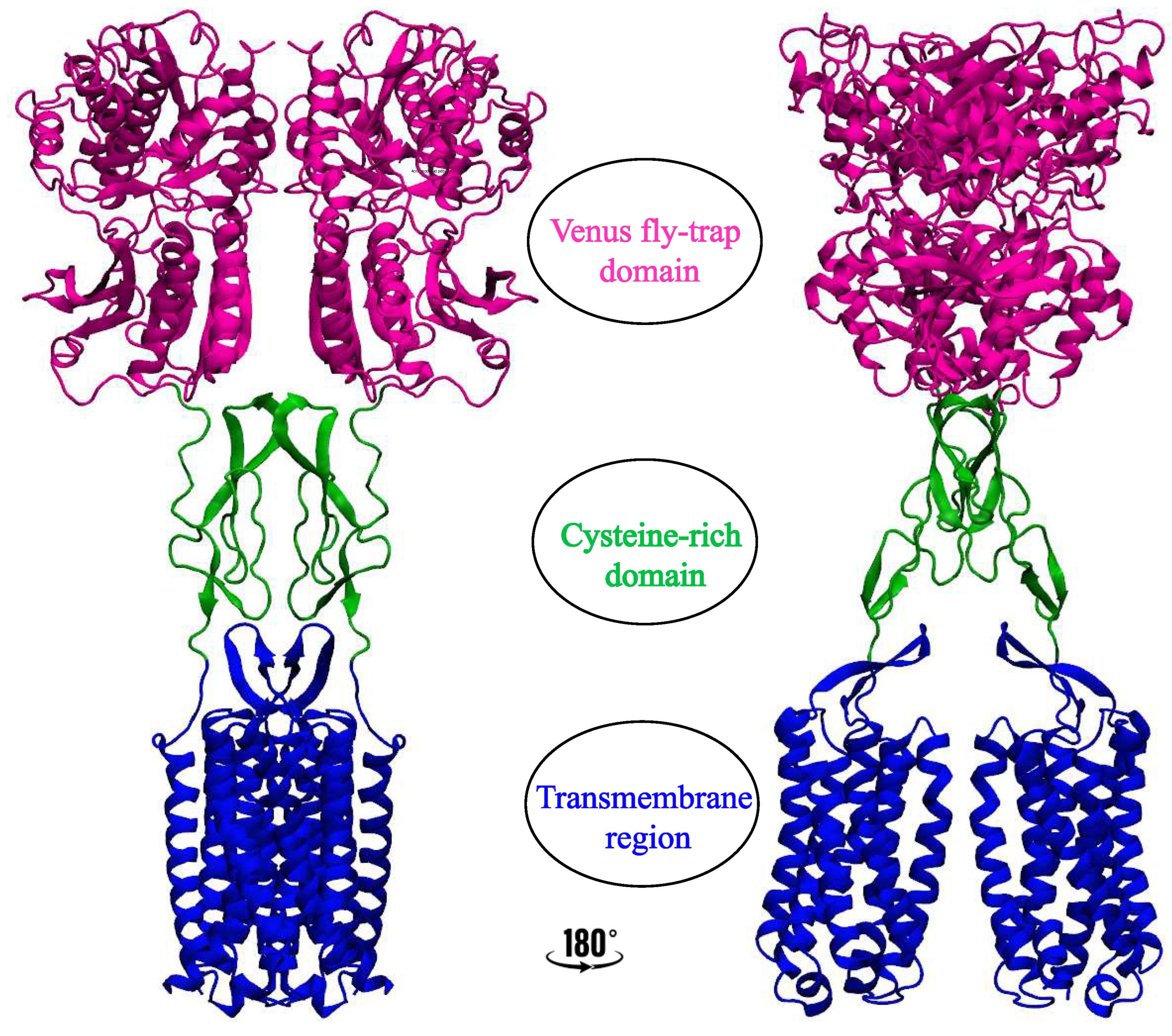
Class C GPCRs comprise about 23 receptors with a unique characterization compared to other GPCR classes and are responsible for neurotransmitters, glutamate, GABA, sweet and umami taste, and calcium receptors. They are obligate homo-, (e.g. mGluRs)[14,15,16] or hetero-, (e.g. ) dimers[17,18,19], and are made up of 3 unique structural elements: a seven-transmembrane domain which is responsible for allosteric ligand recognition but is uniquely dimeric in the case of class C GPCRS[20]; an unusually large extracellular venus flytrap domain (VFT) which has a double-lobed structure with a crevice between them to serve as the orthosteric binding site; and a cysteine-rich domain (CRD) that links the VFT region to the 7TM region (Figure 1)[21]. However, some class C GPCRs, like receptor[22,23], lack the cysteine-rich domain. Due to these distinct structural features and mandatory dimerization, the class C GPCRs have been the most complex of the GPCRs in terms of understanding their activation mechanism[24,25]. Structures of over 20 class C GPCRs have been solved to date, comprising eight metabotropic glutamate receptors (mGluR1–8), gamma-aminobutyric acid receptors (1 and GABA2), calcium-sensing receptor (CaS)[24], retinoic acid-inducible orphan G-protein-coupled receptors (RAIG), taste receptors (TAS1R1–TAS1R3)[26] and orphan receptors (GPR158, GPR179, GPR156 ). Similarly to other GPCR structures, some of the determined class C GPCR structures include cholesterol or cholesteryl hemisuccinate (CHS) during crystallization (Table 1). In contrast, some others include bound cholesterol acting as ligands to the already determined structures (Table 1). The argument for cholesterol addition varies from the stabilization of the protein to aiding dimerization. Experimental analysis and, most recently molecular dynamics simulations[8,9,10,11,12,27] have been used to decipher the possible role of cholesterol in these protein structures. In this review, we will discuss the relevance and position of cholesterol molecules in class C GPCR structures and functions.
1.1. Cholesterol-Membrane Interactions
The plasma membrane of eukaryotic cells consists of various lipids displaying high biochemical variability in both their apolar moiety and their polar head[28]. Sterols are a class of lipids that are a key component of the plasma membrane and, are characterized by their steroid hydrocarbon ring structure. One specific sterol, cholesterol, makes up a vital part of the plasma membrane of eukaryotic cells. Cholesterol is crucial for membrane dynamics and organization and it is also necessary for viability and cell proliferation. The structural features of cholesterol qualify it to interact with proteins and other membrane lipids in several different ways through a variety of different interaction domains[28]. Cholesterol consists of a tetracyclic fused ring skeleton with a single hydroxyl group, a double bond, and a flexible iso-octyl hydrocarbon sidechain[29] which allows it to take on a wide array of conformations[28]. The hydroxyl group is said to contribute significantly to the amphiphilic behavior of cholesterol, causing it to orient in membranes[30]. It is also essential in the hydrogen bond formation between cholesterol and water, as well as other lipid membranes in the cell. The hydroxyl group can form two distinct types of hydrogen bonds (acceptor and donor) with a polar group belonging to either a membrane lipid or a protein. Cholesterol is able to affect the physical behavior and dynamics of the cell membrane by interacting with membranes rich in sphingolipids such as lipid “rafts”[31,32,33], or by being present in the liquid disordered (Ld) phase of membranes which contain a large number of glycerophospholipids such as phosphatidylcholine[34]. As a result of this, cholesterol is able to alter the properties and dynamics of proteins in the membrane[29,35,36]. When bound to cholesterol, some integral membrane proteins could become activated or inactivated [36,37,38]. Cholesterol has been known to affect the properties of the membrane directly or indirectly by modulating the physical properties of the membrane[29,39,40,41,42]. In recent times, there has been considerable interest in cholesterol interaction sites in membrane proteins. Due to various structural characteristics of membrane proteins, not all proteins will react with cholesterol, and those that do may do so in different manners or locations[43]. For example, some proteins associate with cholesterol-rich domains while others have direct interactions with cholesterol through their transmembrane domains. The more common cholesterol binding sites in membrane proteins include the cholesterol recognition/interaction amino acid consensus (CRAC)/CARC domain[44], and the sterol-sensing domain (SSD)[45,46]. All of these listed are structural features in proteins that could result in preferential involvement with cholesterol. Several studies have shown that protein-cholesterol interactions are more common in proteins with sequences comprising of the CRAC motif[47], a short peptide segment at the tail of a transmembrane helix comprising of 5-13 amino acid residues. The CRAC motif consists of a well-defined linear sequence of amino acids[28,43,47,48,49,50,51] identified by the following pattern: a leucine or valine residue, 1-5 non-specific amino acid residues, tyrosine, another 1-5 residues of any amino acid, and finally a lysine or arginine residue [-L/V-(X)1–5-Y-(X)1–5-R/K-, with (X)1–5 representing between one and five residues of any amino acid] [43,47,48,49]. HIV-1 transmembrane protein gp41[47,52] and mammalian seminal plasma protein PDC-109[47,53] are examples of proteins that have been identified with the CRAC motif recognition site. The major difference between the CARC and CRAC motif is that one exhibits a preference for the outer membrane leaflet (CARC) while the mirror sequence (CRAC) is located in the inner membrane leaflet[54,55]. A double CARC-CRAC motif has been identified within the transmembrane domains of some membrane proteins[54,56]; however, the limiting factor of the CRAC/CARC sequence is that they are based on a linear (1D) sequence motif, as opposed to cholesterol-binding sites which consist of a three-dimensional (3D) structure[57]. Additionally, the sterol sensing domain is another significant cholesterol recognition motif with a larger protein segment and comprises five transmembrane helices. The sterol-sensing domains usually consist of a tetrapeptide amino acid sequence: tyrosine; isoleucine; tyrosine; and phenylalanine (YIYF), which has been found to be present in other lipid-raft associated proteins without the SSD motif[43]. Studies have shown that the presence of the YIYF amino acid sequence alone can interact with cholesterol-rich domains[43]. Finally, START proteins have also been identified as a cholesterol binding motif, with the transport of cholesterol molecules being their primary function[58,59]. Proteins with the START domain are able to transfer lipids between membranes and also interact with cholesterol[43]. While all of these are cholesterol-binding motifs in membrane proteins, the CARC-CRAC motif is the major cholesterol interaction site that has been observed in GPCRs.
1.2. GPCR-Cholesterol Interactions
G-protein-coupled receptors are a superfamily of integral membrane proteins in the human genome, constituting one of the largest classes of clinical drug targets[60,61,62,63]. Often distinguished by a characteristic seven transmembrane helices plus an eighth helix that lies underneath the surface of the layer, GPCRs depend on a relationship with the lipid membranes in their physical environment to perform their function[64,65] . As per the phylogenetic investigation, most GPCRs belong to one of four classes, i.e. A, B, C, and Frizzled. The class-C GPCR family contains metabotropic glutamate receptors (mGluR), -aminobutyric corrosive B receptors, a few taste-detecting receptors (e.g., TAS1R1), and a Ca2+-detecting receptor (CaS)[66]. One trademark highlight of the class-C GPCRs is their dimerization, either into homo- or hetero-dimers, which is requisite for their proper functioning[15]. Cholesterol assumes an essential role in the function of a significant number of GPCR structures[67]. It does this by binding to many GPCR structures at both canonical and non-canonical binding sites, consequently altering their ligand binding activity allosterically, which could result in the activation or inactivation of the protein. As such, cholesterol can influence GPCR dependability, oligomerization, and ligand proclivity[68]. Two mechanisms have been proposed by which cholesterol might influence the structure and function of GPCRs: directly, through specific interactions with the GPCRs; indirectly, by altering the physical properties of the membrane; or perhaps some combination of the two[47]. Functional implications of cholesterol interactions have been determined for a number of GPCRs, including rhodopsin, oxytocin, galanin, and serotonin 1A receptors. Recently, several GPCR structures have been determined through X-ray diffraction and even more through cryo-EM. A large percentage of these structures have been stabilized by site-specific cholesterol binding, although it is uncertain if these cholesterol associations are due to recurring cholesterol-binding motifs or if the experimental technique used determines the method of cholesterol binding. A comprehensive study by Taghon et al.[69], showed that cholesterol binding in both X-ray and cryo-EM structures is much the same. They also indicate that about 92% of cholesterol molecules on GPCR surfaces are located in visible regions that do not require cholesterol-binding motifs[69]. The importance of cholesterol in GPCRs structural dynamics has been identified in some GPCR structures, especially within the class A family. In some cases, CHS has been used to substitute cholesterol in GPCRs: although, the validity of this replacement has been contested[48,70,71,72]. The CRAC motif has been established as a characteristic feature of the serotonin(1A) receptor, the 2-adrenergic receptor, rhodopsin, cannabinoid (CB1) receptor, etc[47,48,73], indicating that the interaction of cholesterol with GPCRs could be specific in nature. However, another group of researchers, suggested that the presence of CRAC/CARC motifs does not automatically prove that cholesterol interacts within those binding motifs[74]. On the flip side, their impact on class C GPCRs is yet to be fully explored[17]. The significance of cholesterol to GPCR structures and their functional dynamics is an ongoing question that is yet to be fully elucidated[48].

Table 1.
Solved class C GPCR structures with bound cholesterol.
| Name | PDB ID* | # of Choleseterols in TM |
|---|---|---|
| mGluR1 | 4OR2 [17] | 6 CLR |
| mGluR5 | 7FD8 [16] | 2 CHS |
| () | 6WIV [75] | 10 CLR |
| 7CUM [76] | 16 CLR | |
| 7CA3 [76] | 3 CLR | |
| Orphan receptor (GPR158) | 7SHF [77] | 22 CLR |
| 7SHE [77] | 22 CLR** |
* Data obtained from protein data bank (PDB) database (https://www.rcsb.org). Citations to the PDB structures are included. ** CLR cholesterol; CHS cholesterol hemisuccinate).
Figure 1.
Representation of Class C GPCRs showing the different regions as: VFT (magenta), CRD (green) and 7TM region (blue)
Figure 1.
Representation of Class C GPCRs showing the different regions as: VFT (magenta), CRD (green) and 7TM region (blue)
2. Significance and Interaction Sites of Cholesterol in Class C GPCRs
2.1. Metabotropic Glutamate Receptors (mGluRs)
Metabotropic glutamate receptors are transmembrane proteins that belong to the family of GTP-binding proteins named GPCRs[78]. The GPCRs are membrane-bound proteins expressed in the central nervous system (CNS), and their physiological functions are dependent on their lipid environment[79]. There are three groups with eight subtypes of mGluRs that are classified based on G-protein coupling and ligand selectivity[15,80]. Group I consists of mGluR1 and mGluR5 which are linked to the activation of phospholipase C (PLC) to increase diacylglycerol (DAG) and inositol triphosphate (IP3), Group II includes mGluR2 and 3[81], Group III comprises of mGluR4, 6, 7 and 8. These last two groups are linked to the inhibition of adenylyl cyclases (AC)[82,83]. In mammalian cells, cholesterol is highly concentrated in the plasma membrane but low in the intracellular membrane[84]. Cholesterol affects receptors function by affecting the membrane’s fluidity or interacting with the receptor’s binding site[85] and, moves freely between the inner and outer leaflets[86]. In lipid rafts, plasma membrane are rich in cholesterol and sphingolipids within their lipid domains, and the cholesterol forms specific interactions with GPCRs including mGluR1 and mGluR2. However, the mechanisms underlying their recruitment to these membrane domains still need to be discovered. There has been a lasting argument as to why cholesterol is included in the membrane of mGluRs. While a general consensus agrees that it is strictly due to crystal packing, studies have shown that there could be additional reasons. Research has shown that cholesterol aids the dimerization of mGluR2 and mGluR5 through interactions with the TM4/TM5 domains and also through the TM1/TM2 domains of mGluR1[87,88,89]. In mGluRs, investigations from several scientists have suggested specific interaction sites for mGluR1 and mGluR2[15,90]. A study has revealed the presence of a CRAC motif in the transmembrane helix 5 domain of mGluR1, which is conserved for all mGluRs. The CRAC motif located in TM5 plays an important role in supporting mGluR1 recruitment to the lipid raft as a result of agonist binding[88]. It has been reported that mutations in this motif affect both signaling and the association of mGluR1 with cholesterol-rich membrane domains[88]. Another group has experimentally determined that within the transmembrane domain of mGluR1, cholesterol is localized within the helix I homodimer interface. Intriguingly, this was observed through analyses on the crystal structure of the transmembrane domain of mGluR1, bound by six cholesterol molecules mediating the dimer interface, which in this case is mainly composed of the TM1 helices from both protomers (Figure 2). These cholesterol molecules have been suggested as stabilizing the dimerization of mGluR1 (PDB:4OR2)[17]. In addition, by increasing cholesterol levels, mGluR1 signaling efficiency is enhanced upon stimulation by an agonist, while by lowering cholesterol levels, Extracellular signal-regulated kinase-mitogen-activated protein kinase (ERK-MAPK) activation via mGluR1 is inhibited[88,91]. In this way, lipid rafts and membrane cholesterol act as positive allosteric modulators of the group I mGluR signaling pathway. Therefore, it is possible to modulate abnormal group I mGluR behavior in neuropsychiatric conditions (fragile X syndrome and autism) through the use of drugs such as statins and cyclodextrins, which affect membrane cholesterol levels[88]. Furthermore, the role of cholesterol has also been considered for class II members of mGluRs. A number of neuropsychiatric conditions, including depression, Alzheimer’s disease, and Parkinson’s disease, as well as different types of cancer, have been treated with these same classes of drugs[92,93,94]. The binding of glutamate to mGluR2 dimers results in the transmission of a signal across the transmembrane domain of the receptor that prevents the activity of adenylate cyclase via the protein[80]. In a study that utilized molecular dynamics simulations[95,96,97], biochemical approaches, and photocrosslinking experiments, the interaction of cholesterol with mGluR2 was demonstrated across 2 to 5 sites in the transmembrane domain of mGluR2[80]. It was observed that mGluR2 is modulated by their surrounding lipid environment, particularly cholesterol, through an unknown mechanism. The CRAC/CARC motif and a cholesterol consensus motif (CCM) were suggested as cholesterol-binding motifs in GPCRs[28,98]. A central aromatic amino acid that interacts with sterols is a common characteristic of some of the motifs described[80]. A recent study conducted by Bruno et al.,[99] found that the conformational differences observed in the helical structure of the mGluR2-TM8 domain can be used as an indicator to detect the presence of cholesterol in metabotropic glutamate receptors and GPCRs. They observed that the inclusion of higher levels of cholesterol in the membrane stabilizes the transmembrane helix 8 (TM8) of mGluR2, while a lack of cholesterol results in destabilization of the TM8 domain[99]. However, the role of cholesterol in the third group of mGluRs remains unknown.
2.2. GABAB Receptors
In mammals, GABA (-Aminobutyric acid) is one of the major inhibitory neurotransmitters. In order for GABA to exert their effects, it must bind to at least two different receptor classes: and . Approximately 20 to 50% of the brain’s synapses contain receptors[100]. They are pentameric receptors belonging to a superfamily of ligand-gated ion channels[101]. GABA acts as an agonist by binding to the -benzodiazepine receptor complex, causing conformational changes and increasing the permeability of the central pore to chloride ions. Consequently, the chloride flux affects the neurons by hyperpolarizing them, reducing their excitability, and inhibiting their activity in general[102]. receptors have faster responses (milliseconds) compared to receptors (hundreds of milliseconds)[103,104,105]. Unlike , receptors are members of class C GPCRs with the typical classification of an N-terminal VFT region; a 7TM domain, and a C-terminal intracellular domain[106,107]. receptors function as inhibitor receptors by opening potassium channels, reducing the activity of adenylate cyclase and calcium channels[108]. There are few solved structures of receptors containing cholesterol, deposited on the protein data bank (Figure 3), and subsequently, there is little knowledge of the effect of membrane cholesterol on the receptors. Experimental investigations have shown that cholesterol enrichment and depletion both decrease GABA potency, resulting in an increase of up to fourfold in EC50[109]. The structures of receptors with cholesterol were determined based on ligand type; thus there was no cholesterol present in the structures which were not bound to ligands. However, for systems bound to an antagonist, 10 and 16 molecules of cholesterol[75,76] were bound between the protomers of the transmembrane dimers[67], while 3 cholesterol molecules were attached to the receptor bound to a positive allosteric modulator[76] (Figure 3).
2.3. Taste Receptor
TAS1R1 and TAS1R2 were among the first determined subfamilies of taste-related GPCRs. Prior to identifying their physiological ligands, they were originally classified as orphan receptors[110]. Subsequently, some scientists identified a member, TAS1R3, through a fusion of molecular biological and genetic approaches[111]. These three members: TAS1R1-3, code for sweet and umami tastes and are classified as class C GPCRs. The sweet taste signals are activated by TAS1R2 and TAS1R3 heterodimers, while the umami taste signals are transduced by heterodimers of TAS1R1 and TAS1R3[112]. Therefore, the class C taste receptors consist of either TAS1R1 or TAS1R2, linked by a common subunit TAS1R3. Similar to other class C GPCRs, they exist as obligate dimers and are characterized by a large extracellular N-terminus, which houses the orthosteric ligand-binding site, while the allosteric binding sites are present in the cysteine-rich domain and/or transmembrane region[111]. Due to these multiple binding sites, a single taste receptor is able to function for various stimuli[113]. The sweet taste receptor is able to interact with various compounds at a lower sensitivity, unlike most GPCRs, which are highly selective to specific high-affinity ligands. Cholesterol has been shown to regulate GPCR signaling in sweet taste receptors[114,115]. A study showing the presence of a CRAC motif in T2R4 (a subset of GPCRs responsible for bitter taste receptors[116]), explains that taste receptors are crucial to cholesterol sensitivity[43] and become more sensitive to cholesterol through a cellular mechanism[47]. Furthermore, they observe electrostatic interactions between the 3-hydroxyl group of cholesterol and the positively charged residue in the cholesterol binding motif[117,118]. Site-directed mutagenesis and functional assays have been optimized in the study of putative cholesterol-binding motifs (CRAC and CARC) to determine the mechanism of cholesterol binding to taste receptors. A comparison of the dynamics of wild-type T2R14 receptors and mutant T2R14 receptors revealed that the amino acid residues K110, F236, and L239 are required for the receptor to function appropriately when cholesterol is present. Based on this study, it could be suggested that cholesterol influences taste receptors by directly interacting with the receptor[119].
2.4. Calcium Sensing Receptor-Related Receptor
As a G-protein-coupled receptor, the calcium-sensing receptor (CaSR) is essential for controlling calcium homeostasis[120] in humans. CaSR is a Ca2+-sensing protein found on the surface of cells[121], that exists as an obligate homodimer and belongs to class C GPCRs[122]. Each protomer has a Ca2+-binding extracellular domain and a seven-transmembrane-helix domain (7TM) that activates heterotrimeric G-proteins[24]. The classical calcium-sensing receptor is known to be involved in the pathophysiology of parathyroid and renal-related diseases by sensing calcium ions in extracellular fluid[113,120]. Increasing receptor sensitivity to Ca2+ as a result of homodimer interactions between CaS-TM domains is consistent with the role of a positive allosteric modulator in stabilizing the active conformation. CaS subunits interact with cholesterol molecules near the extra- and intracellular membranes. Cholesterol-mediated interactions occur in the extracellular domain and involve the TM7 elements surrounding the direct TM6 dimer interface. These peripheral dimer interactions involving TM6 residues are not necessary for forming a TM6-centered homodimer interface. Due to the separation of the TM6 helices on the intracellular side, cholesterol molecules (CLR4 and CLR6) mediate nearly all homodimer interactions. However, this is different in the middle of TM6, where two Ile816 side chains pack against each other and make indirect dimer contacts through cholesterol. Each substitution of alanine for a different residue and leucine for Phe809 resulted in a significant decrease in Ca2+ potency and efficiency. The inactivating F809L substitution is another notable mutation which causes disease[123]. The TM6-TM6 dimer interface is stabilized by cholesterol, which has been theorized to play a crucial role in receptor activation. Cholesterol depletion has been shown to negatively impact receptor function by decreasing basal activity and Ca2+ sensitivity. Parathyroid cells have been found to contain the CaS receptor in their lipid- and cholesterol-rich membranes[124]. A study showed that vascular smooth muscle cells (VSMCs)[125] are expressed in CaSR and can be altered by cholesterol[126]. They further indicated that plaque stability can be affected due to CaSR[127] mediating MMP-2 (Matrix metalloproteinase-2) production in the presence of cholesterol via the phosphatidylinositol 3-kinase (PI3K)/Akt signal pathway[128,129]. In addition, activation of CaSR in VSMCs increases cell proliferation and survival via the phospholipase C (PLC)-IP3 and MAPK-ERK1/2 pathways[130].
2.5. Orphan Receptor
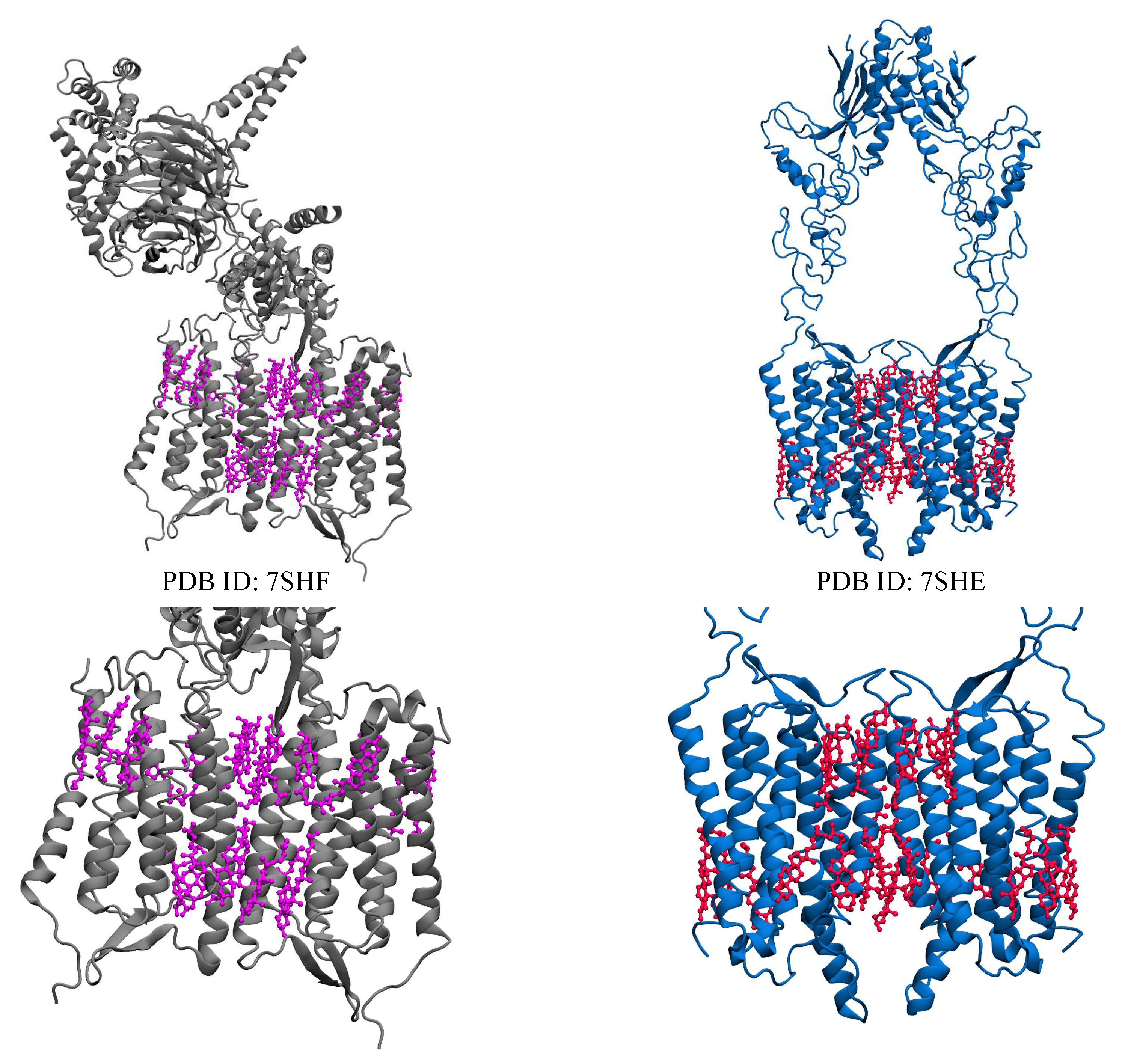
Orphan receptors are membrane-bound receptors that mediate communication between cells and molecules outside the cell; however, they are yet to be thoroughly characterized due to distinct structural features[131,132]. Despite extensive reorganization efforts, there are hundreds of receptors within the GPCR family that have yet to be fully identified[133,134,135,136], many of which are olfactory[137] or taste receptors[138]. The orphan nuclear receptor family is a primary classification of orphan receptors, which are mainly located in the cytosol[139]. To function as transcription factors, these receptors must first bind to their respective ligands, at which point they undergo a spatial conformational shift and move to specific regions inside the nucleus. These factors regulate gene expression in response to various physiological, developmental, and environmental signals[140]. Specific nuclear receptors can also mediate nongenomic effects that are too fast to require changes in gene transcription. Typically, nuclear receptors will have four distinct functional subunits: ligand binding domains, DNA binding domains, modulator domains, and hinge regions[141]. Some examples of orphan nuclear receptors include: the Retinoic acid receptor-related orphan receptors (RORs), Farnesoid X (FX), Liver X (LX)[142], Pregnane X (PX), Dax1, and the short heterodimeric partner (Shp) receptors[143]. A subfamily of nuclear receptors consists of the retinoic acid-related orphan receptors alpha, beta, and gamma (ROR- encoded by RORA-C or NR1F1-3)[144]. An N-terminal domain, a highly conserved DNA-binding domain (DBD) with two C2-C2 zinc finger motifs, a ligand-binding domain (LBD), and a hinge domain between the DBD and LBD[14] are all characteristic of RORs[145,146]. ROR response elements (ROREs) are sequences that contain the RGGTCA consensus and are typically preceded by an A/T-rich sequence, which are recognized by the DBD of RORs [147]. By binding as monomers to ROREs in the regulatory regions of target genes and then recruiting co-activators or co-repressors[148], RORs regulate transcription[149]. The role of RORs as ligand-dependent transcription factors has been noted[150]. Different sterols and synthetic ligands can bind to RORs and act as agonists or inverse agonists to alter ROR / transcriptional activity[151]. GPR156, GPR158, GPR179, GPRC5A, GPRC5B, GPRC5C, GPRC5D, and GPRC6 all make up the orphan receptor of class C GPCRs. Cholesterol and their metabolites, are bioactive lipids that control many proteins and signaling pathways. ROR and members of the ligand-dependent nuclear receptor superfamily, have been found to have surprisingly broad binding specificity for a variety of sterols. Several cholesterol metabolites and intermediates work with ROR and ROR as agonists or inverse agonists. Changes in cholesterol homeostasis that affect the amount or type of sterol metabolites in cells can change the physiological processes that RORs control, such as different immune responses and metabolic pathways[148,152]. Recently, two structures of GPR158 which are bound to cholesterol molecules have been determined[77]. In both initial structures, cholesterol interacts between the protomers and groves of the transmembrane helix (Figure 4).
3. Conclusions
Through this review and the accompanying table and figures, we have described the interaction sites of cholesterol in specific receptors of class C GPCR structures. Through the collective study of class C GPCR structures, we notice that cholesterol is mostly bound between the transmembrane dimers of the receptors and also within the surrounding groves of the transmembrane helices, which could explain why it seems to aid dimerization. Furthermore, this review highlights the significance of cholesterol within specific class C GPCRs. A consideration of several studies revealed that cholesterol is important for oligomerization, organization, function and dynamics of class C GPCRs. In general, we see that cholesterol could affect ligand binding, G-protein coupling, and intracellular signaling of GPCRs. With the possible emergence of more cholesterol-bound GPCRs structures and analyses, we picture an exciting and enlightening future in the knowledge of cholesterol-GPCR interactions. We expect that this information will help provide insight into the molecular mechanisms of cholesterol molecules bound to particular receptors of class C GPCRs.
Author Contributions
Conceptualization, U.H.I.; and M.M.; methodology, U.H.I.; and M.M.; software, U.H.I.; and M.M.; validation, U.H.I.; S.B.; E.K.; and M.M.; formal analysis, U.H.I.; S.B.; E.K.; investigation, U.H.I.; S.B.; E.K.; and M.M.; resources, U.H.I.; S.B.; E.K.; and M.M.; data curation, U.H.I.; S.B.; E.K.; and M.M.; writing—original draft preparation, U.H.I.; S.B.; E.K.; and M.M.; writing—review and editing, U.H.I.; and M.M.; visualization, U.H.I.; S.B.; E.K.; and M.M.; supervision, M.M.; project administration, U.H.I.; and M.M.; funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institute of General Medical Sciences of the National Institutes of Health under award number R35GM147423. This work was also supported by the Arkansas Biosciences Institute.
Data Availability Statement
No new data is reported in this review article
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| GPCR | G-protein-coupled receptors |
| CHS | Cholesteryl hemisuccinate |
| CLR | Cholesterol |
| mGluR | Metabotropic glutamate receptors |
| CRAC | Cholesterol Recognition/Interaction Amino Acid Consensus (CRAC) |
| MD(S) | Molecular dynamics (simulation) |
| GABA | Gamma-Aminobutyric acid |
| TAS1R1 | Taste 1 receptor member 1 |
| TAS1R2 | Taste 1 receptor member 2 |
| TAS1R3 | Taste 1 receptor member 3 |
| RORs | Retinoid-related orphan receptors |
| CASR | calcium-sensing receptor |
| TM(D) | Transmembrane (domain) |
| NTD | N-terminal domain |
| VSMCs | Vascular smooth muscle cells |
| PLC | phospholipase C |
| MMP-2 | Matrix metalloproteinase-2 |
| ERK | Extracellular signal-regulated kinase |
| MAPK | Mitogen-activated protein kinase |
| AMPK | AMP-activated protein kinase |
References
- Maurice, P.; Kamal, M.; Jockers, R. Asymmetry of GPCR oligomers supports their functional relevance. Trends in pharmacological sciences 2011, 32, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Goddard, A.D.; Watts, A. Regulation of G protein-coupled receptors by palmitoylation and cholesterol, 2012. [CrossRef]
- Cherezov, V.; Rosenbaum, D.M.; Hanson, M.A.; Rasmussen, S.G.; Thian, F.S.; Kobilka, T.S.; Choi, H.J.; Kuhn, P.; Weis, W.I.; Kobilka, B.K.; others. High-resolution crystal structure of an engineered human β2-adrenergic G protein–coupled receptor. science 2007, 318, 1258–1265. [Google Scholar] [CrossRef]
- Soffientini, U.; Graham, A. Intracellular cholesterol transport proteins: roles in health and disease. Clinical Science 2016, 130, 1843–1859. [Google Scholar] [CrossRef]
- Lee, A.G. Interfacial Binding Sites for Cholesterol on G Protein-Coupled Receptors. Biophysical Journal 2019, 116, 1586–1597. [Google Scholar] [CrossRef] [PubMed]
- Zakany, F.; Kovacs, T.; Panyi, G.; Varga, Z. Direct and indirect cholesterol effects on membrane proteins with special focus on potassium channels. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2020, 1865, 158706. [Google Scholar] [CrossRef]
- Zocher, M.; Zhang, C.; Rasmussen, S.G.; Kobilka, B.K.; Müller, D.J. Cholesterol increases kinetic, energetic,and mechanical stability of the human β2-adrenergic receptor. Proceedings of the National Academy of Sciences of the United States of America 2012, 109. [Google Scholar] [CrossRef] [PubMed]
- Polasa, A.; Moradi, M. Deciphering the Inter-domain Decoupling in the Gram-negative Bacterial Membrane Insertase. bioRxiv 2022. [Google Scholar] [CrossRef]
- Govind Kumar, V.; Polasa, A.; Agrawal, S.; Kumar, T.K.S.; Moradi, M. Binding affinity estimation from restrained umbrella sampling simulations. Nature Computational Science 2022. [Google Scholar] [CrossRef]
- Polasa, A.; Mosleh, I.; Losey, J.; Abbaspourrad, A.; Beitle, R.; Moradi, M. Developing a rational approach to designing recombinant proteins for peptide-directed nanoparticle synthesis. Nanoscale Adv. 2022, 4, 3161–3171. [Google Scholar] [CrossRef]
- Govind Kumar, V.; Ogden, D.S.; Isu, U.H.; Polasa, A.; Losey, J.; Moradi, M. Prefusion spike protein conformational changes are slower in SARS-CoV-2 than in SARS-CoV-1. Journal of Biological Chemistry 2022, 298, 101814. [Google Scholar] [CrossRef]
- Polasa, A.; Hettige, J.; Kalyan, I.; Moradi, M. An investigation of the YidC-mediated membrane insertion of Pf3 coat protein using molecular dynamics simulations. Frontiers in Molecular Biosciences 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Immadisetty, K.; Moradi, M. Mechanistic Picture for Chemomechanical Coupling in a Bacterial Proton-Coupled Oligopeptide Transporter from Streptococcus Thermophilus. The Journal of Physical Chemistry B 2021, 125, 9738–9750. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Kim, T.K.; Hobrath, J.V.; Oak, A.S.W.; Tang, E.K.Y.; Tieu, E.W.; Li, W.; Tuckey, R.C.; Jetten, A.M. Endogenously produced nonclassical vitamin D hydroxy-metabolites act as “biased” agonists on VDR and inverse agonists on RORα and RORγ. The Journal of Steroid Biochemistry and Molecular Biology 2017, 173, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: physiology, pharmacology, and disease. Annual review of pharmacology and toxicology 2010, 50, 295. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, C.; Cannone, G.; Briot, J.; Rottier, K.; Berizzi, A.E.; Huang, C.Y.; Quast, R.B.; Hoh, F.; Banères, J.L.; Malhaire, F.; Berto, L.; Dumazer, A.; Font-Ingles, J.; Gómez-Santacana, X.; Catena, J.; Kniazeff, J.; Goudet, C.; Llebaria, A.; Pin, J.P.; Vinothkumar, K.R.; Lebon, G. Agonists and allosteric modulators promote signaling from different metabotropic glutamate receptor 5 conformations. Cell Reports 2021, 36, 109648. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, C.; Gregory, K.J.; Han, G.W.; Cho, H.P.; Xia, Y.; Niswender, C.M.; Katritch, V.; Meiler, J.; Cherezov, V.; Conn, P.J.; Stevens, R.C. Structure of a class C GPCR metabotropic glutamate receptor 1 bound to an allosteric modulator. Science 2014. [Google Scholar] [CrossRef] [PubMed]
- Mazzitelli, M.; Palazzo, E.; Maione, S.; Neugebauer, V. Group II Metabotropic Glutamate Receptors: Role in Pain Mechanisms and Pain Modulation. Frontiers in Molecular Neuroscience 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Shaye, H.; Ishchenko, A.; Lam, J.H.; Han, G.W.; Xue, L.; Rondard, P.; Pin, J.P.; Katritch, V.; Gati, C.; Cherezov, V. Structural basis of the activation of a metabotropic GABA receptor. Nature 2020, 584, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Vafabakhsh, R.; Levitz, J.; Isacoff, E.Y. Conformational dynamics of a class C G-protein-coupled receptor. Nature 2015, 524, 497–501. [Google Scholar] [CrossRef]
- Chun, L.; Zhang, W.h.; Liu, J.f. Structure and ligand recognition of class C GPCRs. Acta Pharmacologica Sinica 2012, 33, 312–323. [Google Scholar] [CrossRef]
- Kunishima, N.; Shimada, Y.; Tsuji, Y.; Sato, T.; Yamamoto, M.; Kumasaka, T.; Nakanishi, S.; Jingami, H.; Morikawa, K. Structural basis of glutamate recognition by a dimeric metabotropic glutamate receptor. Nature 2000. [Google Scholar] [CrossRef]
- Muto, T.; Tsuchiya, D.; Morikawa, K.; Jingami, H. Structures of the extracellular regions of the group II/III metabotropic glutamate receptors. Proceedings of the National Academy of Sciences of the United States of America 2007. [Google Scholar] [CrossRef]
- Møller, T.C.; Moreno-Delgado, D.; Pin, J.P.; Kniazeff, J. Class CG protein-coupled receptors: reviving old couples with new partners. Biophysics reports 2017, 3, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Salon, J.A.; Lodowski, D.T.; Palczewski, K. The Significance of G Protein-Coupled Receptor Crystallography for Drug Discovery. Pharmacological Reviews 2011, 63, 901–937. [Google Scholar] [CrossRef]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.G.; Schiöth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Molecular Pharmacology. [CrossRef]
- Harkey, T.; Govind Kumar, V.; Hettige, J.; Tabari, S.H.; Immadisetty, K.; Moradi, M. The Role of a Crystallographically Unresolved Cytoplasmic Loop in Stabilizing the Bacterial Membrane Insertase YidC2. Scientific Reports 2019. [Google Scholar] [CrossRef]
- Fantini, J.; Barrantes, F. How cholesterol interacts with membrane proteins: An exploration of cholesterol-binding sites including CRAC, CARC, and tilted domains. Frontiers in physiology 2013, 4, 31. [Google Scholar] [CrossRef]
- Ohvo-Rekilä, H.; Ramstedt, B.; Leppimäki, P.; Peter Slotte, J. Cholesterol interactions with phospholipids in membranes, 2002. [CrossRef]
- McMullen, T.P.W.; Lewis, R.N.A.H.; McElhaney, R.N. Cholesterol–phospholipid interactions, the liquid-ordered phase and lipid rafts in model and biological membranes. Current Opinion in Colloid & Interface Science 2004, 8, 459–468. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef]
- Anderson, R.G.W.; Jacobson, K. A Role for Lipid Shells in Targeting Proteins to Caveolae, Rafts, and Other Lipid Domains. Science 2002, 296, 1821–1825. [Google Scholar] [CrossRef]
- Fantini, J.; Yahi, N. Molecular insights into amyloid regulation by membrane cholesterol and sphingolipids: common mechanisms in neurodegenerative diseases. Expert Reviews in Molecular Medicine 2010, 12, e27. [Google Scholar] [CrossRef]
- Fantini, J.; Garmy, N.; Mahfoud, R.; Yahi, N. Lipid rafts: structure, function and role in HIV, Alzheimer’s and prion diseases. Expert Reviews in Molecular Medicine 2002, 4, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Yeagle, P.L. Cholesterol and the cell membrane. Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes 1985, 822, 267–287. [Google Scholar] [CrossRef]
- Yeagle, P.L. Modulation of membrane function by cholesterol. Biochimie 1991, 73, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Peränen, J.; Schreiner, R.; Wieland, F.; Kurzchalia, T.V.; Simons, K. VIP21/caveolin is a cholesterol-binding protein. Proceedings of the National Academy of Sciences 1995, 92, 10339–10343. [Google Scholar] [CrossRef]
- Porter, J.A.; Young, K.E.; Beachy, P.A. Cholesterol Modification of Hedgehog Signaling Proteins in Animal Development. Science 1996, 274, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Edwards, P.A.; Ericsson, J. Signaling molecules derived from the cholesterol biosynthetic pathway: mechanisms of action and possible roles in human disease. Current Opinion in Lipidology 1998, 9. [Google Scholar] [CrossRef] [PubMed]
- Paila, Y.D.; Chattopadhyay, A. The function of G-protein coupled receptors and membrane cholesterol: Specific or general interaction? Glycoconjugate Journal, 2009. [CrossRef]
- Paila, Y.D.; Tiwari, S.; Chattopadhyay, A. Are specific nonannular cholesterol binding sites present in G-protein coupled receptors?, 2009. [CrossRef]
- Paila, Y.D.; Chattopadhyay, A. Membrane cholesterol in the function and organization of g-protein coupled receptors. Sub-Cellular Biochemistry, 2010. [Google Scholar] [CrossRef]
- Epand, R.M. Cholesterol and the interaction of proteins with membrane domains. Progress in Lipid Research 2006, 45, 279–294. [Google Scholar] [CrossRef]
- Sutter, M.L.; Console, L.; Fahner, A.F.; Samodelov, S.L.; Gai, Z.; Ciarimboli, G.; Indiveri, C.; Kullak-Ublick, G.A.; Visentin, M. The role of cholesterol recognition (CARC/CRAC) mirror codes in the allosterism of the human organic cation transporter 2 (OCT2, SLC22A2). Biochemical Pharmacology 2021, 194, 114840. [Google Scholar] [CrossRef]
- Kuwabara, P.E.; Labouesse, M. The sterol-sensing domain: multiple families, a unique role? Trends in Genetics 2002, 18, 193–201. [Google Scholar] [CrossRef]
- Martín, V.; Carrillo, G.; Torroja, C.; Guerrero, I. The sterol-sensing domain of Patched protein seems to control Smoothened activity through Patched vesicular trafficking. Current biology : CB 2001, 11, 601–607. [Google Scholar] [CrossRef]
- Jafurulla, M.; Tiwari, S.; Chattopadhyay, A. Identification of cholesterol recognition amino acid consensus (CRAC) motif in G-protein coupled receptors. Biochemical and biophysical research communications 2011, 404, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Chattopadhyay, A. Cholesterol in GPCR Structures: Prevalence and Relevance. The Journal of Membrane Biology 2022, 255, 99–106. [Google Scholar] [CrossRef]
- Li, H.; Yao, Z.x.; Degenhardt, B.; Teper, G.; Papadopoulos, V. Cholesterol binding at the cholesterol recognition/ interaction amino acid consensus (CRAC) of the peripheral-type benzodiazepine receptor and inhibition of steroidogenesis by an HIV TAT-CRAC peptide. Proceedings of the National Academy of Sciences 2001, 98, 1267–1272. [Google Scholar] [CrossRef]
- Sarkar, P.; Mozumder, S.; Bej, A.; Mukherjee, S.; Sengupta, J.; Chattopadhyay, A. Structure, dynamics and lipid interactions of serotonin receptors: excitements and challenges. Biophysical Reviews 2021, 13, 101–122. [Google Scholar] [CrossRef]
- Prasad, R.; Paila, Y.D.; Jafurulla, M.; Chattopadhyay, A. Membrane cholesterol depletion from live cells enhances the function of human serotonin1A receptors. Biochemical and Biophysical Research Communications 2009, 389, 333–337. [Google Scholar] [CrossRef]
- Vincent, N.; Genin, C.; Malvoisin, E. Identification of a conserved domain of the HIV-1 transmembrane protein gp41 which interacts with cholesteryl groups. Biochimica et Biophysica Acta (BBA) - Biomembranes 2002, 1567, 157–164. [Google Scholar] [CrossRef]
- Scolari, S.; Müller, K.; Bittman, R.; Herrmann, A.; Müller, P. Interaction of Mammalian Seminal Plasma Protein PDC-109 with Cholesterol: Implications for a Putative CRAC Domain. Biochemistry 2010, 49, 9027–9031. [Google Scholar] [CrossRef]
- Fantini, J.; Di Scala, C.; Evans, L.S.; Williamson, P.T.F.; Barrantes, F.J. A mirror code for protein-cholesterol interactions in the two leaflets of biological membranes. Scientific Reports 2016, 6, 21907. [Google Scholar] [CrossRef]
- Di Scala, C.; Baier, C.; Evans, L.; Williamson, P.; Fantini, J.; Barrantes, F. Relevance of CARC and CRAC Cholesterol-Recognition Motifs in the Nicotinic Acetylcholine Receptor and Other Membrane-Bound Receptors. Current Topics in Membranes 2017. [Google Scholar] [CrossRef]
- Sengupta, D.; Chattopadhyay, A. Molecular dynamics simulations of GPCR-cholesterol interaction: An emerging paradigm, 2015. [CrossRef]
- Azzaz, F.; Chahinian, H.; Yahi, N.; Di Scala, C.; Baier, C.J.; Barrantes, F.J.; Fantini, J. Chapter 7 - Cholesterol-recognizing amino acid consensus motifs in transmembrane proteins: Comparative analysis of in silico studies and structural data. In Cholesterol; Bukiya, A.N.; Dopico, A.M., Eds.; Academic Press, 2022; pp. 127–145. [CrossRef]
- Bukiya, A.N.; Dopico, A.M. Common structural features of cholesterol binding sites in crystallized soluble proteins. Journal of Lipid Research 2017, 58, 1044–1054. [Google Scholar] [CrossRef]
- Rone, M.B.; Fan, J.; Papadopoulos, V. Cholesterol transport in steroid biosynthesis: Role of protein–protein interactions and implications in disease states. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2009, 1791, 646–658. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, X.; Hua, T.; Liu, Z.J.; Liu, H.; Zhao, S. MD Simulations Revealing Special Activation Mechanism of Cannabinoid Receptor 1. Frontiers in Molecular Biosciences 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Nieto Gutierrez, A.; McDonald, P.H. GPCRs: Emerging anti-cancer drug targets. Cellular Signalling 2018, 41, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nature Reviews Drug Discovery 2017. [Google Scholar] [CrossRef] [PubMed]
- Sriram, K.; Insel, P.A. GPCRs as targets for approved drugs: How many targets and how many drugs? Molecular Pharmacology 2018. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Prasanna, X.; Mohole, M.; Chattopadhyay, A. exploring GPCR-lipid interactions by Molecular dynamic simulation. J. Phys. Chem. B 2018. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, X.; Mohole, M.; Chattopadhyay, A.; Sengupta, D. Role of cholesterol-mediated effects in GPCR heterodimers. Chemistry and Physics of Lipids 2020, 227, 104852. [Google Scholar] [CrossRef] [PubMed]
- Niswender, C.M.; Jones, C.K.; Lin, X.; Bubser, M.; Thompson Gray, A.; Blobaum, A.L.; Engers, D.W.; Rodriguez, A.L.; Loch, M.T.; Daniels, J.S.; others. Development and antiparkinsonian activity of VU0418506, a selective positive allosteric modulator of metabotropic glutamate receptor 4 homomers without activity at mGlu2/4 heteromers. ACS chemical neuroscience 2016, 7, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Moreau, C.J.; Audic, G.; Lemel, L.; García-Fernández, M.D.; Nieścierowicz, K. Interactions of cholesterol molecules with GPCRs in different states: A comparative analysis of GPCRs’ structures. Biochimica et Biophysica Acta (BBA) - Biomembranes 2023, 1865, 184100. [Google Scholar] [CrossRef]
- Jakubík, J.; El-Fakahany, E.E. Allosteric Modulation of GPCRs of Class A by Cholesterol, 2021. [CrossRef]
- Taghon, G.; Rowe, J.; Kapolka, N.; Isom, D. Predictable cholesterol binding sites in GPCRs lack consensus motifs. Structure 2021. [Google Scholar] [CrossRef]
- Augustyn, B.; Stepien, P.; Poojari, C.; Mobarak, E.; Polit, A.; Wisniewska-Becker, A.; Róg, T. Cholesteryl Hemisuccinate Is Not a Good Replacement for Cholesterol in Lipid Nanodiscs. The Journal of Physical Chemistry B 2019, 123, 9839–9845. [Google Scholar] [CrossRef] [PubMed]
- Kulig, W.; Tynkkynen, J.; Javanainen, M.; Manna, M.; Rog, T.; Vattulainen, I.; Jungwirth, P. How well does cholesteryl hemisuccinate mimic cholesterol in saturated phospholipid bilayers? Journal of Molecular Modeling 2014, 20, 2121. [Google Scholar] [CrossRef] [PubMed]
- Kulig, W.; Jurkiewicz, P.; Olżyńska, A.; Tynkkynen, J.; Javanainen, M.; Manna, M.; Rog, T.; Hof, M.; Vattulainen, I.; Jungwirth, P. Experimental determination and computational interpretation of biophysical properties of lipid bilayers enriched by cholesteryl hemisuccinate. Biochimica et Biophysica Acta (BBA) - Biomembranes 2015, 1848, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Dainese, E.; Sandiford, S.; Fezza, F.; Lanuti, M.; Chiurchiù, V.; Totaro, A.; Catanzaro, G.; Barcaroli, D.; De Laurenzi, V.; Centonze, D.; Mukhopadhyay, S.; Selent, J.; Howlett, A.; Maccarrone, M. Effects of palmitoylation of Cys 415 in helix 8 of the CB 1 cannabinoid receptor on membrane localization and signalling. British Journal of Pharmacology 2012, 165, 2635–2651. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Chattopadhyay, A. Cholesterol interaction motifs in G protein-coupled receptors: Slippery hot spots? Wiley Interdisciplinary Reviews: Systems Biology and Medicine 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Fu, Z.; Frangaj, A.; Liu, J.; Mosyak, L.; Shen, T.; Slavkovich, V.N.; Ray, K.M.; Taura, J.; Cao, B.; Geng, Y.; Zuo, H.; Kou, Y.; Grassucci, R.; Chen, S.; Liu, Z.; Lin, X.; Williams, J.P.; Rice, W.J.; Eng, E.T.; Huang, R.K.; Soni, R.K.; Kloss, B.; Yu, Z.; Javitch, J.A.; Hendrickson, W.A.; Slesinger, P.A.; Quick, M.; Graziano, J.; Yu, H.; Fiehn, O.; Clarke, O.B.; Frank, J.; Fan, Q.R. Structure of human GABAB receptor in an inactive state. Nature 2020, 584, 304–309. [Google Scholar] [CrossRef]
- Kim, Y.; Jeong, E.; Jeong, J.H.; Kim, Y.; Cho, Y. Structural Basis for Activation of the Heterodimeric GABAB Receptor. Journal of Molecular Biology 2020, 432, 5966–5984. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.N.; Singh, S.; Laboute, T.; Strutzenberg, T.S.; Qiu, X.; Wu, D.; Novick, S.J.; Robinson, C.V.; Griffin, P.R.; Hunt, J.F.; Izard, T.; Singh, A.K.; Martemyanov, K.A. Cryo-EM structure of human GPR158 receptor coupled to the RGS7-Gβ5 signaling complex. Science 2022, 375, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Pin, J.P.; De Colle, C.; Bessis, A.S.; Acher, F. New perspectives for the development of selective metabotropic glutamate receptor ligands. European journal of pharmacology 1999, 375, 277–294. [Google Scholar] [CrossRef]
- Sejdiu, B.I.; Tieleman, D.P. Lipid-protein interactions are a unique property and defining feature of G protein-coupled receptors. Biophysical journal 2020, 118, 1887–1900. [Google Scholar] [CrossRef]
- Kurth, M.; Lolicato, F.; Sandoval, A.; Amaya-Espinosa, H.; Teslenko, A.; Sinning, I.; Beck, R.; Brügger, B.; Aponte-Santamaria, C. Cholesterol Localization Around the Metabotropic Glutamate Receptor 2. The Journal of Physical Chemistry B 2020, 124. [Google Scholar] [CrossRef]
- Liauw, B.W.H.; Foroutan, A.; Schamber, M.R.; Lu, W.; Samareh Afsari, H.; Vafabakhsh, R. Conformational fingerprinting of allosteric modulators in metabotropic glutamate receptor 2. eLife 2022, 11, e78982. [Google Scholar] [CrossRef]
- Bordi, F.; Ugolini, A. Group I metabotropic glutamate receptors: implications for brain diseases. Progress in neurobiology 1999, 59, 55–79. [Google Scholar] [CrossRef]
- Liauw, B.W.H.; Afsari, H.S.; Vafabakhsh, R. Conformational rearrangement during activation of a metabotropic glutamate receptor. Nature Chemical Biology 2021, 17, 291–297. [Google Scholar] [CrossRef]
- Yeagle, P. The biophysics and cell biology of cholesterol: an hypothesis for the essential role of cholesterol in mammalian cells. Cholesterol in Membrane Models, 1993; 1–12. [Google Scholar]
- Sooksawate, T.; Simmonds, M. Influence of membrane cholesterol on modulation of the GABAA receptor by neuroactive steroids and other potentiators. British journal of pharmacology 2001, 134, 1303–1311. [Google Scholar] [CrossRef]
- Schroeder, F.; Jefferson, J.R.; Kier, A.B.; Knittel, J.; Scallen, T.J.; Wood, W.G.; Hapala, I. Membrane cholesterol dynamics: cholesterol domains and kinetic pools. Proceedings of the Society for Experimental Biology and Medicine 1991, 196, 235–252. [Google Scholar] [CrossRef]
- Marlow, B.; Kuenze, G.; Li, B.; Sanders, C.R.; Meiler, J. Structural determinants of cholesterol recognition in helical integral membrane proteins. Biophysical Journal 2021, 120, 1592–1604. [Google Scholar] [CrossRef]
- Kumari, R.; Castillo, C.; Francesconi, A. Agonist-dependent signaling by group I metabotropic glutamate receptors is regulated by association with lipid domains. Journal of Biological Chemistry 2013, 288, 32004–32019. [Google Scholar] [CrossRef]
- Lei, T.; Hu, Z.; Ding, R.; Chen, J.; Li, S.; Zhang, F.; Pu, X.; Zhao, N. Exploring the Activation Mechanism of a Metabotropic Glutamate Receptor Homodimer via Molecular Dynamics Simulation. ACS Chemical Neuroscience 2020, 11, 133–145. [Google Scholar] [CrossRef]
- Thibado, J.K.; Tano, J.Y.; Lee, J.; Salas-Estrada, L.; Provasi, D.; Strauss, A.; Marcelo Lamim Ribeiro, J.; Xiang, G.; Broichhagen, J.; Filizola, M.; Lohse, M.J.; Levitz, J. Differences in interactions between transmembrane domains tune the activation of metabotropic glutamate receptors. eLife 2021, 10, e67027. [Google Scholar] [CrossRef]
- Francesconi, A.; Kumari, R.; Zukin, R.S. Regulation of group I metabotropic glutamate receptor trafficking and signaling by the caveolar/lipid raft pathway. Journal of Neuroscience 2009, 29, 3590–3602. [Google Scholar] [CrossRef]
- Caraci, F.; Nicoletti, F.; Copani, A. Metabotropic glutamate receptors: the potential for therapeutic applications in Alzheimer’s disease. Current Opinion in Pharmacology 2018, 38, 1–7. [Google Scholar] [CrossRef]
- Nicoletti, F.; Bruno, V.; Ngomba, R.T.; Gradini, R.; Battaglia, G. Metabotropic glutamate receptors as drug targets: what’s new? Current Opinion in Pharmacology 2015, 20, 89–94. [Google Scholar] [CrossRef]
- Lumeng, J.Y.; Wall, B.A.; Wangari-Talbot, J.; Chen, S. Metabotropic glutamate receptors in cancer. Neuropharmacology 2017, 115, 193–202. [Google Scholar]
- Immadisetty, K.; Hettige, J.; Moradi, M. What Can and Cannot Be Learned from Molecular Dynamics Simulations of Bacterial Proton-Coupled Oligopeptide Transporter GkPOT? The Journal of Physical Chemistry B 2017, 121, 3644–3656. [Google Scholar] [CrossRef]
- Moradi, M.; Babin, V.; Roland, C.; Sagui, C. The Adaptively Biased Molecular Dynamics method revisited: New capabilities and an application. Journal of Physics: Conference Series 2015, 640, 12020. [Google Scholar] [CrossRef]
- Govind Kumar, V.; Agrawal, S.; Kumar, T.K.S.; Moradi, M. Mechanistic Picture for Monomeric Human Fibroblast Growth Factor 1 Stabilization by Heparin Binding. The Journal of Physical Chemistry B 2021, 125, 12690–12697. [Google Scholar] [CrossRef]
- Baier, C.J.; Fantini, J.; Barrantes, F.J. Disclosure of cholesterol recognition motifs in transmembrane domains of the human nicotinic acetylcholine receptor. Scientific reports 2011, 1, 1–7. [Google Scholar] [CrossRef]
- Bruno, A.; Costantino, G.; de Fabritiis, G.; Pastor, M.; Selent, J. Membrane-sensitive conformational states of helix 8 in the metabotropic Glu2 receptor, a class C GPCR. PLoS ONE 2012. [Google Scholar] [CrossRef]
- Nutt, D.J.; Malizia, A.L. New insights into the role of the GABAA–benzodiazepine receptor in psychiatric disorder. The British Journal of Psychiatry 2001, 179, 390–396. [Google Scholar] [CrossRef]
- Shaye, H.; Stauch, B.; Gati, C.; Cherezov, V. Molecular mechanisms of metabotropic GABAB receptor function. Science Advances 2021, 7, eabg3362. [Google Scholar] [CrossRef]
- Maconochie, D.J.; Zempel, J.M.; Steinbach, J.H. How quickly can GABAA receptors open? Neuron 1994, 12, 61–71. [Google Scholar] [CrossRef]
- Sieghart, W. Structure, pharmacology, and function of GABAA receptor subtypes. Advances in pharmacology 2006, 54, 231–263. [Google Scholar]
- Soltesz, I.; Mody, I. Patch-clamp recordings reveal powerful GABAergic inhibition in dentate hilar neurons. Journal of Neuroscience 1994, 14, 2365–2376. [Google Scholar] [CrossRef]
- Otis, T.S.; De Koninck, Y.; Mody, I. Characterization of synaptically elicited GABAB responses using patch-clamp recordings in rat hippocampal slices. The Journal of physiology 1993, 463, 391–407. [Google Scholar] [CrossRef]
- Burmakina, S.; Geng, Y.; Chen, Y.; Fan, Q.R. Heterodimeric coiled-coil interactions of human GABAB receptor. Proceedings of the National Academy of Sciences 2014, 111, 6958–6963. [Google Scholar] [CrossRef]
- Geng, Y.; Bush, M.; Mosyak, L.; Wang, F.; Fan, Q.R. Structural mechanism of ligand activation in human GABAB receptor. Nature 2013, 504, 254–259. [Google Scholar] [CrossRef]
- Bassetti, D. Keeping the Balance: GABAB Receptors in the Developing Brain and Beyond. Brain Sciences 2022, 12, 419. [Google Scholar] [CrossRef]
- Sooksawate, T.; Simmonds, M. Effects of membrane cholesterol on the sensitivity of the GABAA receptor to GABA in acutely dissociated rat hippocampal neurones. Neuropharmacology 2001, 40, 178–184. [Google Scholar] [CrossRef]
- Hoon, M.A.; Adler, E.; Lindemeier, J.; Battey, J.F.; Ryba, N.J.; Zuker, C.S. Putative mammalian taste receptors: a class of taste-specific GPCRs with distinct topographic selectivity. Cell 1999, 96, 541–551. [Google Scholar] [CrossRef]
- Vigues, S.; Dotson, C.D.; Munger, S.D. The receptor basis of sweet taste in mammals. Results and problems in cell differentiation 2009, 47, 187–202. [Google Scholar] [CrossRef]
- Prasad Pydi, S.; Singh, N.; Upadhyaya, J.; Pal Bhullar, R.; Chelikani, P. The third intracellular loop plays a critical role in bitter taste receptor activation. Biochimica et Biophysica Acta (BBA) - Biomembranes 2014, 1838, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Dalziel, J.E. G Protein-Coupled Receptors in Taste Physiology and Pharmacology. Frontiers in Pharmacology 2020, 11, 587664. [Google Scholar] [CrossRef] [PubMed]
- Pydi, S.P.; Jafurulla, M.; Wai, L.; Bhullar, R.P.; Chelikani, P.; Chattopadhyay, A. Cholesterol modulates bitter taste receptor function. Biochimica et Biophysica Acta (BBA)-Biomembranes 2016, 1858, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Ilegems, E.; Iwatsuki, K.; Kokrashvili, Z.; Benard, O.; Ninomiya, Y.; Margolskee, R.F. REEP2 Enhances Sweet Receptor Function by Recruitment to Lipid Rafts. The Journal of Neuroscience 2010, 30, 13774LP–13783. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, J.; Mueller, K.L.; Hoon, M.A.; Adler, E.; Feng, L.; Guo, W.; Zuker, C.S.; Ryba, N.J. T2Rs function as bitter taste receptors. Cell 2000, 100, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Jamin, N.; Neumann, J.M.; Ostuni, M.A.; Vu, T.K.N.; Yao, Z.X.; Murail, S.; Robert, J.C.; Giatzakis, C.; Papadopoulos, V.; Lacapere, J.J. Characterization of the cholesterol recognition amino acid consensus sequence of the peripheral-type benzodiazepine receptor. Molecular endocrinology 2005, 19, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.F.; Thomas, A.; Brasseur, R.; Vishwanathan, S.A.; Hunter, E.; Epand, R.M. Juxtamembrane protein segments that contribute to recruitment of cholesterol into domains. Biochemistry 2006, 45, 6105–6114. [Google Scholar] [CrossRef] [PubMed]
- Shaik, F.A.; Medapati, M.R.; Chelikani, P. Cholesterol modulates the signaling of chemosensory bitter taste receptor T2R14 in human airway cells. American Journal of Physiology-Lung Cellular and Molecular Physiology 2019, 316, L45–L57. [Google Scholar] [CrossRef]
- Brown, E.M.; MacLeod, R.J. Extracellular calcium sensing and extracellular calcium signaling. Physiological reviews 2001, 81, 239–297. [Google Scholar] [CrossRef]
- Hannan, F.M.; Kallay, E.; Chang, W.; Brandi, M.L.; Thakker, R.V. The calcium-sensing receptor in physiology and in calcitropic and noncalcitropic diseases. Nature Reviews Endocrinology 2019, 15, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Schamber, M.R.; Vafabakhsh, R. Mechanism of sensitivity modulation in the calcium-sensing receptor via electrostatic tuning. Nature Communications 2022, 13, 2194. [Google Scholar] [CrossRef]
- Hendy, G.N.; Guarnieri, V.; Canaff, L. Calcium-sensing receptor and associated diseases. Progress in molecular biology and translational science 2009, 89, 31–95. [Google Scholar] [PubMed]
- Kifor, O.; Diaz, R.; Butters, R.; Kifor, I.; Brown, E.M. The calcium-sensing receptor is localized in caveolin-rich plasma membrane domains of bovine parathyroid cells. Journal of Biological Chemistry 1998, 273, 21708–21713. [Google Scholar] [CrossRef] [PubMed]
- Augé, N.; Maupas-Schwalm, F.; Elbaz, M.; Thiers, J.C.; Waysbort, A.; Itohara, S.; Krell, H.W.; Salvayre, R.; Nègre-Salvayre, A. Role for matrix metalloproteinase-2 in oxidized low-density lipoprotein–induced activation of the sphingomyelin/ceramide pathway and smooth muscle cell proliferation. Circulation 2004, 110, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, M.; Yamamoto, M.; Azuma, H.; Nagashima, G.; Niimi, Y.; Tamaki, M.; Hirakawa, K.; Yamamoto, K. Immunolocalization of matrix metalloproteinases in rabbit carotid arteries after balloon denudation. Histochemistry and cell biology 1998, 109, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Sundararaman, S.S.; van der Vorst, E.P. Calcium-Sensing Receptor (CaSR), Its Impact on Inflammation and the Consequences on Cardiovascular Health. International journal of molecular sciences 2021, 22, 2478. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, L.; Zielke, H.R.; Cheng, L.; Xiao, R.; Crow, M.T.; Stetler-Stevenson, W.G.; Froehlich, J.; Lakatta, E.G. Increased expression of 72-kd type IV collagenase (MMP-2) in human aortic atherosclerotic lesions. The American journal of pathology 1996, 148, 121. [Google Scholar] [PubMed]
- Molostvov, G.; Fletcher, S.; Bland, R.; Zehnder, D. Extracellular calcium-sensing receptor mediated signalling is involved in human vascular smooth muscle cell proliferation and apoptosis. Cellular Physiology and Biochemistry 2008, 22, 413–422. [Google Scholar] [CrossRef]
- Li, H.X.; Kong, F.J.; Bai, S.Z.; He, W.; Xing, W.J.; Xi, Y.H.; Li, G.W.; Guo, J.; Li, H.Z.; Wu, L.Y.; others. Involvement of calcium-sensing receptor in oxLDL-induced MMP-2 production in vascular smooth muscle cells via PI3K/Akt pathway. Molecular and cellular biochemistry 2012, 362, 115–122. [Google Scholar] [CrossRef]
- Yoshida, M.; Miyazato, M.; Kangawa, K. Orphan GPCRs and methods for identifying their ligands. Methods in enzymology 2012, 514, 33–44. [Google Scholar] [PubMed]
- Mullican, S.E.; DiSpirito, J.R.; Lazar, M.A. The orphan nuclear receptors at their 25-year reunion. Journal of molecular endocrinology 2013, 51, T115–T140. [Google Scholar] [CrossRef] [PubMed]
- Schmid, E.M.; Ford, M.G.J.; Burtey, A.; Praefcke, G.J.K.; Peak-Chew, S.Y.; Mills, I.G.; Benmerah, A.; McMahon, H.T. Role of the AP2 β-appendage hub in recruiting partners for clathrin-coated vesicle assembly. PLoS biology 2006, 4, e262. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.l.; Wang, Y.; Li, D.l.; Luo, J.; Liu, M.y. Orphan G protein-coupled receptors (GPCRs): biological functions and potential drug targets. Acta Pharmacologica Sinica 2012, 33, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhou, Q.; Labroska, V.; Qin, S.; Darbalaei, S.; Wu, Y.; Yuliantie, E.; Xie, L.; Tao, H.; Cheng, J.; others. G protein-coupled receptors: Structure-and function-based drug discovery. Signal transduction and targeted therapy 2021, 6, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: new agents, targets and indications. Nature reviews Drug discovery 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, R.; Bhutani, I.; Somavarapu, A.K.; Mahajan, S.; Parkesh, R.; Gupta, P. ONRLDB—manually curated database of experimentally validated ligands for orphan nuclear receptors: insights into new drug discovery. Database 2015, 2015. [Google Scholar] [CrossRef]
- Giguère, V. Orphan nuclear receptors: from gene to function. Endocrine reviews 1999, 20, 689–725. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Mani, S. Orphan Nuclear Receptors as Targets for Drug Development. Pharmaceutical Research 2010, 27, 1439–1468. [Google Scholar] [CrossRef]
- de Vera, I.M.S. Advances in orphan nuclear receptor pharmacology: a new era in drug discovery, 2018.
- Zhou, J.; Wang, Y.; Wu, D.; Wang, S.; Chen, Z.; Xiang, S.; Chan, F.L. Orphan nuclear receptors as regulators of intratumoral androgen biosynthesis in castration-resistant prostate cancer. Oncogene 2021, 40, 2625–2634. [Google Scholar] [CrossRef]
- Edwards, P.A.; Kennedy, M.A.; Mak, P.A. LXRs;: Oxysterol-activated nuclear receptors that regulate genes controlling lipid homeostasis. Vascular pharmacology 2002, 38, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Mi, L.Z.; Devarakonda, S.; Harp, J.M.; Han, Q.; Pellicciari, R.; Willson, T.M.; Khorasanizadeh, S.; Rastinejad, F. Structural basis for bile acid binding and activation of the nuclear receptor FXR. Molecular cell 2003, 11, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Jetten, A.M. Retinoid-related orphan receptors (RORs): critical roles in development, immunity, circadian rhythm, and cellular metabolism. Nuclear receptor signaling 2009, 7, nrs–07003. [Google Scholar] [CrossRef] [PubMed]
- Solt, L.A.; Burris, T.P. Action of RORs and their ligands in (patho) physiology. Trends in Endocrinology & Metabolism 2012, 23, 619–627. [Google Scholar]
- Jetten, A.M.; Kang, H.S.; Takeda, Y. Retinoic acid-related orphan receptors α and γ: key regulators of lipid/glucose metabolism, inflammation, and insulin sensitivity. Frontiers in endocrinology 2013, 4, 1. [Google Scholar] [CrossRef]
- Giguere, V.; McBroom, L.; Flock, G. Determinants of target gene specificity for ROR alpha 1: monomeric DNA binding by an orphan nuclear receptor. Molecular and Cellular Biology 1995, 15, 2517–2526. [Google Scholar] [CrossRef] [PubMed]
- Jetten, A.M.; Takeda, Y.; Slominski, A.; Kang, H.S. Retinoic acid-related orphan receptor γ (RORγ): Connecting sterol metabolism to regulation of the immune system and autoimmune disease. Current Opinion in Toxicology 2018, 8, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, A.; Yan, Z.H.; Hirose, T.; Giguère, V.; Jetten, A.M. Cloning of a cDNA encoding the murine orphan receptor RZR/RORγ and characterization of its response element. Gene 1996, 181, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.N.; Kang, H.S.; Jetten, A.M. Retinoic acid-related orphan receptors (RORs): regulatory functions in immunity, development, circadian rhythm, and metabolism. Nuclear receptor research 2015, 2. [Google Scholar] [CrossRef]
- Kallen, J.; Schlaeppi, J.M.; Bitsch, F.; Delhon, I.; Fournier, B. Crystal structure of the human RORα ligand binding domain in complex with cholesterol sulfate at 2.2 Å. Journal of Biological Chemistry 2004, 279, 14033–14038. [Google Scholar] [CrossRef]
- Shahoei, S.H.; Nelson, E.R. Nuclear receptors, cholesterol homeostasis and the immune system. The Journal of Steroid Biochemistry and Molecular Biology 2019, 191, 105364. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Crystal structures of mGluR1 (grey) and mGluR5 (blue) determined with 6 molecules of cholesterol (red) and 2 molecules of CHS (magenta) respectively
Figure 2.
Crystal structures of mGluR1 (grey) and mGluR5 (blue) determined with 6 molecules of cholesterol (red) and 2 molecules of CHS (magenta) respectively
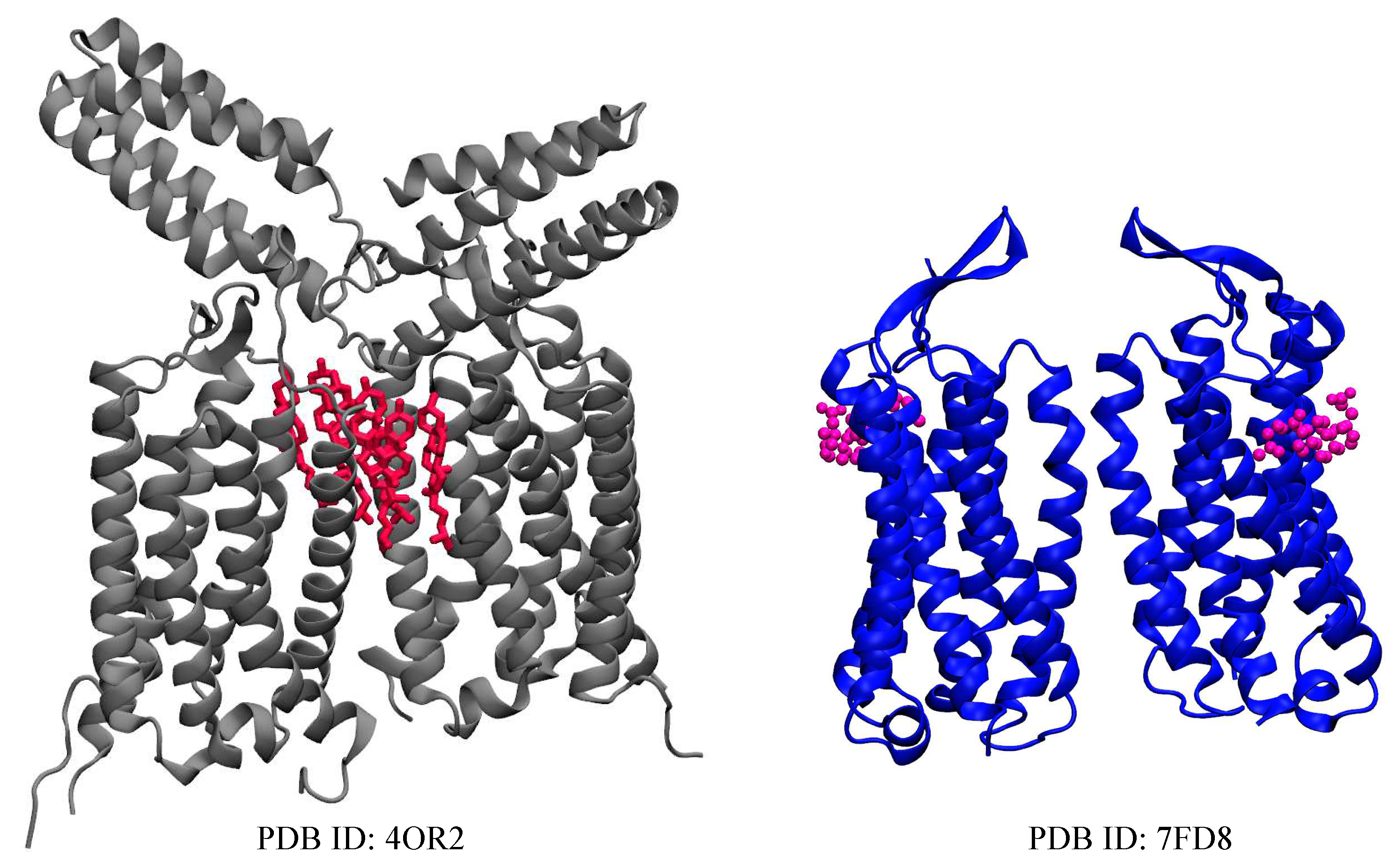
Figure 3.
Visual representations of solved receptors (red, orange, grey) bound with 17, 2, and 16 cholesterol molecules respectively within the transmembrane region. The figures here show the transmembrane region only bound to cholesterol (cholesterol molecules are shown as cyan and green sticks)
Figure 3.
Visual representations of solved receptors (red, orange, grey) bound with 17, 2, and 16 cholesterol molecules respectively within the transmembrane region. The figures here show the transmembrane region only bound to cholesterol (cholesterol molecules are shown as cyan and green sticks)
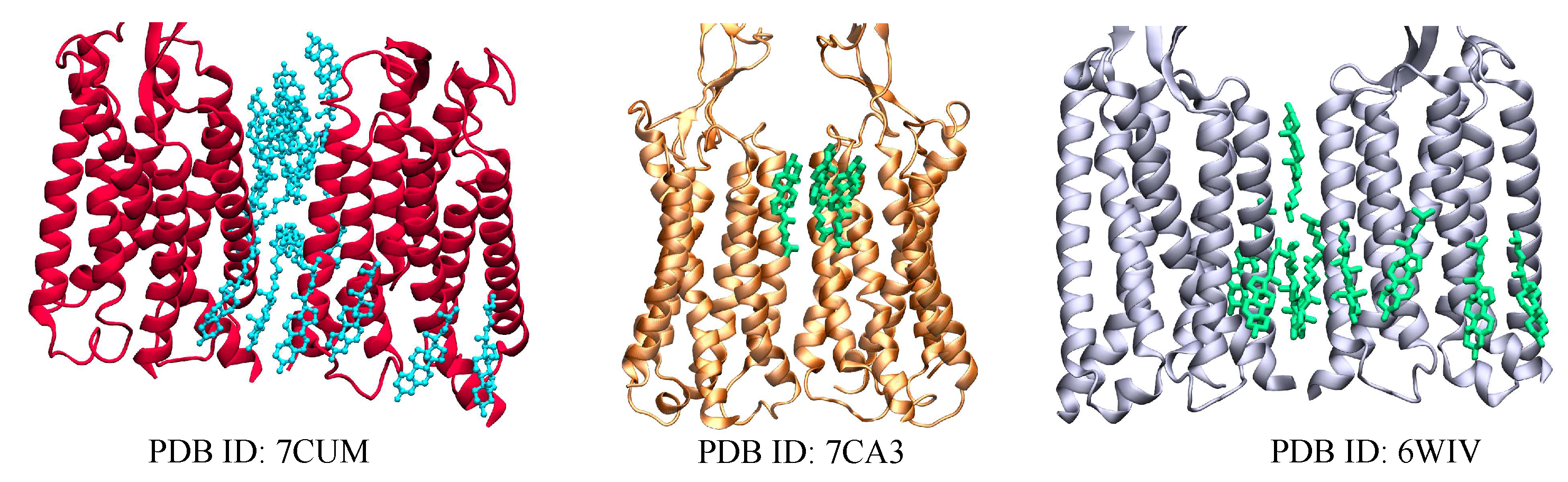
Figure 4.
Visual representations of GPR158 class C orphan receptors (grey and blue) bound with 22 cholesterol molecules each within the transmembrane region. The upper figures show the whole protein while the lower figures only show the transmembrane region (cholesterol molecules are shown as violet and red sticks).
Figure 4.
Visual representations of GPR158 class C orphan receptors (grey and blue) bound with 22 cholesterol molecules each within the transmembrane region. The upper figures show the whole protein while the lower figures only show the transmembrane region (cholesterol molecules are shown as violet and red sticks).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



