
Submitted:
12 January 2023
Posted:
23 January 2023
You are already at the latest version
Abstract
Common root rot caused by Bipolaris sorokiniana infestation of wheat is one of the main reasons of yield reduction in wheat crops worldwide. In current study, strain JK-25 was isolated from soil of wheat rhizosphere and identified as Bacillus halotolerans based on morphological, physiological, biochemical characteristics and molecular identification. The strain showed significant antagonism to B.sorokiniana and broad-spectrum resistance to Fusarium oxysporum, Fusarium graminearum and Rhizoctonia zeae. Inhibition of Bipolaris sorokiniana mycelial dry weight and spore germination rate by JK-25 fermentation supernatant reached 60% and 88% respectively. The crude extract of JK-25 was found by MALDI-TOF-MS to contain the surfactin that exerted an inhibitory effect on B.sorokiniana. The disruption of mycelial cell membranes was observed under microscopy (LSCM) after treatment of B.sorokiniana mycelium with the crude extract. The antioxidant enzyme activity of B.sorokiniana was significantly reduced and the oxidation product MDA content increased after treatment with the crude extract. The incidence of root rot was significantly reduced in pot experiments with the addition of JK-25 culture ferment, which had a significant biological control effect of 72.06%. Its ability to produce siderophores may help to promote wheat growth, and the production of proteases and pectinases may also be part of the strain's role in suppressing pathogens. These results demonstrate the excellent antagonistic effect of JK-25 against B.sorokiniana and suggest that this strain has great potential as a resource for biological control of wheat root rot strains.
Keywords:
Bipolaris sorokiniana
; Bacillus halotolerans
; common root rot
; biocontrol agent
; wheat
1. Introduction
Wheat is one of the world's top three crops, with the second-highest yield after maize [1]. The pathogens associated with common root rot diseases have become the cause of wheat, especially durum wheat, production serious constraints to yield and quality losses in [2,3]. Common root rot symptoms are characterized by necrotic lesions on the shoot root and usually followed by the shoot crown, which are dark brown to black in color [4]. Root rot infestation is turning into a serious threat to wheat cultivation worldwide, and common root decay has been documented in many wheat-growing locations such as Australia, China, etc [5,6].
Bipolaris sorokiniana as a typical wheat root rot causal agent has been reported to be more threatening to durum wheat[4]. B.sorokiniana can infest not only the roots of wheat but also the leave and stems as well as the wheat grains[7] . In general, there are two aspects of B.sorokiniana infestation, one is to reduce seedling establishment by infesting wheat seeds with mold and rot, and the other is to damage various plant tissues, including wheat rootstocks, for the duration of the growth of wheat seedlings[8].
Currently, the most effective way to manipulate root rot of wheat brought on by B.sorokiniana infestation is to use chemical control fungicides in combination with agronomic measures[9]. It has been reported that the incidence of root rot of wheat can be significantly reduced by crop rotation of wheat with Brassica carinata [10]. Organic agriculture considerably decreases the population of pathogenic fungi linked to root deterioration in wheat soils within the Canadian locale [11]. However, these agricultural practices are costly and require a high degree of mechanized agricultural development, which is not suitable for the actual farming environment and arable land requirements in China. Chemical antimicrobial agents are significantly effective in controlling root rot, but they are not in line with the concept of sustainable agriculture because they cause serious pathogenic bacteria resistance and crop chemical residues [12].
In current years, organic manipulate techniques have received increasing consideration, and the development of effective biocontrol agents is important to complement agricultural management practices and reduce the use of chemical antimicrobials [4,13]. A number of antagonistic strains inhibiting B.sorokiniana have been reported. Several antagonistic strains (e.g., Trichoderma harzianum, Bacillus subtilis, Ochrobactrum pseudogrignonense, Pseudomonas mediterranea, etc.) inhibiting B.sorokiniana have been reported and these strains obtained by screening from soil are considered secure for the growing environment and mammals [14-18]. Among them, Bacillus are considered the greatest potential to be developed as a biocide because of its broad-spectrum antibacterial properties and significant growth-promoting effect. Bacillus, as a bacterium, has an advantage over other antagonistic microorganisms in that it has a unique ability to form endospores in response to various adverse environments as well as can proliferate rapidly [19]. Some Bacillus promote crop growth by producing siderophore and indoleacetic acid as well as playing an remarkable role in activation of crop immune mechanism by triggering the induced systemic resistance (ISR) [20,21]. It additionally has been reported that Bacillus can inhibit the growth of plant pathogenic fungi by producing some extracellular active molecules such as lytic enzymes, antibiotics and lipopeptides [22]. In conclusion, Bacillus sp. as a potential development resource for biocontrol agents has been widely researched and concerned. For example, the fermentation supernatant of strain XZ34-1 was reported to be effective in controlling root rot of wheat under greenhouse conditions as well as having a significant growth-promoting effect [23]. Furthermore, Bacillus velezensis LHSB1 Inhibition of root-rot pathogens in peanut growth through the production of three antimicrobial lipopeptide substances [24].
In this study, we screened for a strain of Bacillus sp. JK-25 with a strong antagonistic effect against B. sorokiniana.The inhibition effect of this strain against B. sorokiniana and the biological control effect in wheat potted plants under greenhouse conditions were investigated. It was revealed that the mechanism of inhibition of B. sorokiniana by the antagonistic strain JK-25 and its growth-promoting characteristics were investigated.
2. Results
2.1. Screening of Antagonistic Bacteria
A total of 74 bacterial strains were screened in the collected soil of wheat field, and five of them showed significant inhibitory effects against common wheat pathogens. Table 1 shows that out of the five bacterial strains, JK-25 exhibited the best antagonistic effect against B. sorokiniana. with an inhibition of 82.63%. JK-25 also showed the best inhibition against common wheat bacterial pathogens other than B. sorokiniana. including Fusarium oxysporum, Fusarium graminearum, and Rhizoctonia zeae with inhibition rates of 80.47%, 75.33%, and 59.73%, respectively. Because the superior inhibition effect of JK-25 has shown this bacterium to have great potential for the biological control of wheat root rot, the strain was chosen for further identification and analysis.
2.2. Identification of Antagonistic Strain JK-25
Twenty-four hours after incubation on solid LB medium, strain JK-25 formed round, creamy white colonies without obvious pigmentation, with an overall smooth surface, rough margin and opaque. The strain JK-25 was found to be gram-positive under microscope, and the rod cells contained endospores. It was found to be capable of utilizing citrate and fermenting glucose, reducing nitrate, producing contactase and catalase, hydrolyzing starch and gelatin, and withstand in a high salt environment with concentrations of 10% NaCl (Table 2). The 16SrDNA sequence of strain JK-25 was uploaded to Genbank (accession number OP804109) and a Neighbor-joining phylogenyetic tree was constructed (Figure 1). The strain JK-25 showed the highest homology (99.72%) with B. halotolerans (accession number MH036321), and the strains were clustered on the same evolutionary branch as B. halotolerans. The morphological analysis, physiological and biochemical tests and molecular identification of the strain JK-25 were combined to determine that the strain is a B. halotolerans.
2.3. Inhibitory Effect of the Antagonistic Strain JK-25 CF on B. sorokiniana
JK-25 showed a very significant antagonistic effect on B. sorokiniana by plate confrontation (Figure 2). The CF of strain JK-25 was found to have a significant inhibitory effect on mycelial growth and spore germination of B. sorokiniana (Figure 3). The results in the figure showed a decrease of 63.97% in mycelial dry weight and 87.52% in spore germination rate of B. sorokiniana after the addition of JK-25 CF. This indicates that the supernatant contains some antimicrobial substances produced by JK-25 fermentation, which can have a significant antagonistic effect on both mycelium and spores of B. sorokiniana.
2.4. Detection and identification of antifungal metabolites
MALDI-TOF MS was used for the efficient determination and analysis of the lipopeptides produced by the antagonist strain JK-25. Based on the analysis of the results, surfactin was successfully detected in the crude extract of the fermentation broth. As shown in Figure 4, Ions of m/z values 1016.545, 1030.542, 1044.530, 1058.577 were detected, corresponding to C12-17 surfactin [M+H]+ as previous reported (Dhanarajan et al. 2016), and ions of m/z values 1074.547 1080.558, 1102.564 correspond to C14-17 surfactin [M+Na]+. The heterogeneity among peaks was differ in molecular weight by 14 Da, suggesting the presence of varied lengths of fatty acid chains within surfactin.
2.5. Effect of crude extract on Cell Membrane of B. sorokiniana
Under laser scanning confocal microscope (LSCM), it was observed that the surface of B. sorokiniana mycelium treated with JK-25 crude extract became rougher and many mycelium showed abnormal twisted state and more branching nodes, while the morphology of untreated B. sorokiniana mycelium was more uniform and did not show the above morphology (Figure 5). The treated B. sorokiniana mycelium could be observed to have obvious red fluorescence after PI staining, while the untreated mycelium did not show fluorescence, indicating that the crude extract of JK-25 significantly disrupted the cell membrane integrity of the mycelium.
2.6. Detection of the effect of crude extract on the antioxidant activity of B. sorokiniana
Activities of superoxide dismutase (SOD), catalase (CAT), peroxidase (POD) and malondialdehyde (MDA) concentrations in the hyphae of B. sorokiniana were detected after different treatment with different levels of JK-25 crude extract. Figure 6A-C showed the three aforementioned antioxidant enzyme activities, respectively, and as can be seen, the enzyme activities in the T1 (1% crude extract) and T2 (2% crude extract) treated groups were significantly lower than those in the control (untreated group). By comparing antioxidant enzyme activities following treatment with different crude extract levels, the reduction in the activities of all three antioxidant enzymes was evident as the crude extract concentration increased. MDA is one of the compounds produced by the oxidative degradation of unsaturated fatty acids on mycelial membrane system by the action of oxygen radicals. Figure 6D reflects a significant increase in MDA content with increasing crude extract in the treated group compared to the control, indicating that the B. sorokiniana mycelial membrane system was disrupted.
2.7. Biocontrol of wheat rot under greenhouse conditions
Wheat was evaluated for root rot incidence and the effectiveness of birth control after twenty-eight days of growth in pots. As shown in Figure 7A1, B1, in the control group the wheat seedlings showed significant wilting and black necrotic lesions (black spots) on the roots after being infected with B. sorokiniana. A significant increase in wheat growth status and root health was observed in the treatment groups supplemented with 50% carbendazim solution and supernatant of strain JK-25 fermentation solution (Figure 7A2,A3,B2,B3). The addition of JK-25 supernatant significantly suppressed the incidence of root rot compared to the sterile water treatment control, with a biological control effect of 72.06%, although this result was slightly lower than that of the chemical fungicide carbendazim (Table 3). The result suggested that treatment with JK-25 culture broth was useful for the control of wheat common root rot.
2.8. Production of extracellular enzymes and Growth Promotion
To explore the ability of this strain to produce extracellular enzymes, JK-25 was inoculated onto different types of plates. The results showed that strain JK-25 was able to produce transparent zone after staining on M9 medium spiked with pectin and cellulose, indicating that the strain was able to secrete pectinase and cellulase. JK-25 was also observed to produce protease and siderophores. (Table 4) However, the strain unable to hydrolyse chitin on the plate, suggesting it could not produce chitinase. Also, the results for IAA indicate that JK-25 does not have the ability to produce IAA.
3. Discussion
Common root rot diseases of wheat can lead to significant reductions in wheat yields and cause huge economic losses. It is currently mostly controlled with chemical fungicides, but is not in line with the concept of green agriculture and sustainable development. More and more biological control methods have been applied to the control of wheat root rot control in recent times. Sharma et al. reported that actinomycetes isolated from wheat seeds were effective in suppressing soil-borne diseases caused by pathogens including B. sorokiniana[25]. Yi et al. reported that Bacillus amyloliquefaciens XZ34-1 could inhibit B. sorokiniana mycelia dry weight and spore germination through culture filtrate[23]. In currently study, five bacterial strains with antagonistic effect against B. sorokiniana were screened from wheat soil, among which JK-25 showed the highest inhibitory activity against the pathogenic bacterium B. sorokiniana. JK-25 also showed an inhibitory effect on B. sorokiniana mycelial dry weight and spore germination in this study.
Different species of antagonistic bacteria inhibit pathogenic bacteria and promote plant growth in different ways, and analysis of the specific mechanisms of action of strain JK-25 could help to improve its biological control in wheat crops. Bacillus is a potential antagonist strain due to its ability to suppress plant pathogens by producing a variety of secondary metabolites and antibiotics [26-29]. Many studies have reported the production of lipopeptides by Bacillus as an effective biocontrol agent against plant pathogens, for instance, Bacillus atrophaeus FA12 and B. cabrialesii FA26 was showed to be able to co-produce the lipopeptides iturins and fengycin, which were responsible for the effective antifungal effects against Xanthomonas oryzae pv. oryzae [30], while mycosubtilin was proved to be responsible for the biocontrol activity of Bacillus subtilis strain Z15 against plant pathogen Verticillium dahliae 991 [31]. By analysing the genome of the strain, Villa-Rodriguez et al. found that Bacillus cabrialesii TE3T was able to encode the production of surfactin, fengycin, and rhizocticin A to effectively suppress B. sorokiniana [32]. Some reports have also analysed the ability of an antagonistic strain to produce multiple antimicrobial lipopeptides that act synergistically, enhancing the antimicrobial effect of the strain and broadening the spectrum of inhibition [28,33,34]. Our results show that analysis of the crude extract of JK-25 by MALDI-TOF-MS revealed the presence of surfactin, which are key substances in the antagonism of B. sorokiniana by JK-25. It has also been reported that surfactin can trigger the Induced Systemic Resistance (ISR) in plants to enhance self-protection mechanisms and thus strengthen their own antagonistic effects against phytopathogens[35]. An important mechanism of action of the lipopeptides produced by Bacillus sp. to inhibit the growth of plant pathogens is to disrupt their cell membrane system [36]. In this study significant red fluorescence was observed in the cell membranes of B. sorokiniana mycelia treated with crude extracts after PI staining, and cell membrane integrity and function were disrupted, which is consistent with the findings reported previously.
Antagonistic bacteria are able to secrete secondary metabolites that disrupt the cellular integrity of pathogens and destabilise their antioxidant systems [37]. The crude extract of the fermentation broth of Bacillus velezensis HY19 significantly reduced the enzymatic activities of CAT, SOD and POD of the pathogenic fungus, thereby inhibiting its growth [38]. Reactive oxygen species (ROS) can cause oxidative damage to biological cells through peroxidation, and antioxidant enzymes can scavenge ROS to maintain cellular stability [3]. Reduced antioxidant enzyme activity after treatment of pathogen mycelium with crude extracts of fermentation broth, leading to accumulation of ROS and thus destruction of pathogen cells [39]. Therefore, as oxidative damage intensifies, the cell membrane system is disrupted and the level of malondialdehyde (MDA), one of the main products of the oxidation of lipid substances in the membrane system, increases [40]. In our results, after treatment of B. sorokiniana mycelium with different levels of crude extracts of strain JK-25, the mycelial antioxidant enzyme activity was significantly reduced compared to the control and the MDA content was significantly increased, which is consistent with previous studies. The increase in mycelial MDA content after treatment also further explains the disruption of mycelial cell membranes observed under the microscopy as a result of oxidative damage.
In the present study,the potential of JK-25 as a highly effective resource of biocontrol was demonstrated by the 72.06% control effect of common root rot in wheat pot experiments conducted under greenhouse conditions. Siderophores have a high affinity for Fe3+ and are capable of binding Fe3+ to form complexes, which make Fe easier for uptaking by microorganisms, as well as being taken up by plants to increase iron content within the tissues and promote plant growth [41]. A study by Mageshwaran et al. revealed that one of the factors that Bacillus subtilis TRO4 promotes the growth of chickpea plants is the production of siderophores [42]. In line with previous reports, the ability of JK-25 to produce siderophores was verified in this study by the CAS blue agar plate method as one of the reasons for the ability to promote wheat growth. The ability of antagonistic bacteria to act on fungal cell structures, generally through the secretion of extracellular hydrolases, is one of their important mechanisms of action. LI et al. reported that Bacillus velezensis HY19 is capable of producing proteases, pectinases and cellulases to act on fungal to achieve disease control [38]. The present investigation showed that strain JK-25 could produce pectinase and protease,it suggested that the hydrolase might reduce the ability of the pathogen to invasion the wheat tissues of the host. Overall, the antagonistic mechanism of action of strain JK-25 against common root rot of wheat is mainly the production of inhibitory secondary metabolites that directly affect the growth and disrupt the cellular stability of B.sorokiniana, in addition to enhancing the plant's protection against the pathogen after infection by triggering ISR of wheat.
4. Materials and Methods
4.1. Materials
Soil samples for screening the isolated antagonistic bacteria were collected from arable land where wheat is grown year-round in Zhoukou, Henan Province. The wheat pathogenic bacteria including B. sorokiniana, Fusarium graminearum, Fusarium oxysporum, Aspergillus niger and Aspergillus flavus involved in this study were all kept in the laboratory of the School of Biological Engineering, Henan University of Technology. The medium was LB medium ( 0.5% yeast extract, 1% peptone, 1% NaCl, solid medium need 2% agar, pH 7) and potato dextrose agar (PDA) medium ( 2% glucose, 20% potato, 2% agar, pH 7).
4.2. Isolation and Screening of Antagonistic Bacteria
The sample soil was obtained from the target area using a five-point sampling method at a depth of approximately 10 cm from the surface and then quickly transferred to the laboratory in sterile bags for temporary storage in a -20°C refrigerator. Soil samples were prepared into different concentrations of sample suspensions using the serial dilution method, followed by three dilutions of 10-5, 10-6 and 10-7 in LB medium for coating and incubation, and the individual distinct colonies were selected for numbering and preservation [43].
Plate confrontation method using for Screening the strain with good antagonistic effect against wheat pathogenic fungi [44]. Briefly, the wheat root rot-causing fungus was pre-cultured on PDA medium at 30°C for 5d, and a 0.5-cm-diameter plug taken from Pathogens was placed at center of 9-cm-diameter PDA plate, then candidate strains were inoculated to the plate with 2.5 cm away from the plug. Plates without any other strains were used as controls, and all plates were culture at 30°C until the colonies in the control plate had grown all over the plate. The inhibition rate(IR) was used to express the antagonistic activity of the screened strains which was calculated using the following equation [24]:
where A is the diameter of the control fungus, a is the diameter of the treatment fungus, the 0.5 is the diameter of the inoculated plug of pathogens.
The strain with the best antagonistic effect was numberd as JK-25 and saved in 30% glycerol at -80°C for further analysis.
4.3. Identification of Strain JK-25
Morphological analyses as well as physiological and biochemical tests were performed and combined with molecular methods to identify antagonistic bacterial species. Morphological analysis is mainly to observe the colony characteristics including color, size, growth characteristics, microscopic morphology and other indicators. Then physiological and biochemical tests according to Bergey’s Manual of Determinative Bacteriology [45].
For molecular distinguishing proof, genomic DNA was obtained from the strain utilizing commercial bacterial genomic DNA extraction kit (Tiangen, China), and the 16SrRNA gene region was amplified using bacterial universal primers 27F(5'-AGA GTT TGA TCA TGG CTC AG-3') and 1492R (5'-ACG GTT ACC TTG TTA CGA CTT-3'). Polymerase chain reaction (PCR) amplification volumes was 25 µl(12.5 µl of PCR MIX (2×), 5 µl of bacterial DNA, 1µl of each primer,and5.5 µl sterile deionized water ) with the following conditions: 94 ℃ for 5 min,30 cycles at 94 ℃ for 30 s,55℃ for 30 s and 72 ℃ for 100 s, followed by a final extension at 72 ℃ for 10 min. Then, the PCR product were sent to Biotech Co., Ltd. (Shanghai, China) for sequencing. The DNA sequences compared for homology through the BLAST tool of the NCBI (USA; http://www.Ncbi.nlm.nih.gov) bacterial database, the phylogenetic tree was inferred using neighbor joining method in MEGA7.0 software using the Maximum Likelihood method with bootstrap values were calculated based on 1,000 replicates [46].
4.4. Inhibitory Effect of the Antagonistic Strain JK-25 culture filtrate (CF) on B.sorokiniana
The antagonistic Strain JK-25 was cultured for 24 h at 37 ℃ and 180r/min in a 250 mL flask containing 100 mL of LB medium [47].The obtained culture broth was centrifuged at 10,000 g and 4 ℃ for 10 minutes to remove the bacterial cells, and then the supernatant which was filtered through a sterile 0.22μm membrane and then the sterile culture filtrate was collected and preserved at 4 ℃.
The spore solution of B. sorokiniana (1 × 106cfu/mL) was accessed in a 250 ml conical flask containing 97 ml of potato dextrose liquid medium at 2% inoculum, 1 ml of the above culture filtrate was added, and then incubated at 30 °C and 180 r/min for 5 days, the treatment with the addition of sterile water was served as the control group. The fermentation broth in the conical flasks of different treatments was placed in 50 ml centrifuge tubes at 4 °C and 8000 g for 10 min to collect the B. sorokiniana mycelial precipitate. The mycelium was washed three times with sterile water, then dried at 55°C to a constant weight and weighed. Inoculation of B. sorokiniana spore suspension 6 h followed by microscopic observation of spore germination status and calculation of germination rate.
4.5. Detection and identification of antagonistic metabolites
To investigate the antimicrobial metabolites produced by the antagonistic bacterium JK-25, a sterile culture filtrate was first obtained according to the treatment method described in 2.4. The pH of the culture filtrate was adjusted to 2 with 6 N HCl and placed at 4°C overnight to allow the crude lipopeptides were precipitated. After that, the precipitate was collected by centrifugation at 4°C and 11000g for fifteen minutes and then extracted three times with methanol. The methanol extract was filtered and evaporated to one-tenth of the original volume under vacuum. The above crude extracts were further analyzed for antibacterial metabolites using MALDI-TOF-MS. The crude extract of the fermentation broth was determined using MALDI-TOF-MS referring to the previously reported method [48]. Mass spectrometry was performed on a instrument of Bruker Daltonik Reflex MALDI-TOF with a 337 nm nitrogen laser for desorption and ionisation using α-cyano-4-hydroxycinnamic acid as a matrix [30,46].
4.6. Effect of crude extract on Cell Membrane of B. sorokiniana
The effect of the antagonist JK-25 crude extract on B. sorokiniana cell membrane integrity was assessed by Propidium iodide (PI) staining. The B. sorokiniana was inoculated in a 250 ml conical flask containing 100 ml of PDB medium and incubated at 28°C for 72 h. The culture medium was centrifuged at 8000 r/min for 10 min to collect the mycelial sediment and the appropriate amount of mycelium was incubated at 28℃ for 12h following resuspended with 900 μl PBS and 100 μl bacterial crude extract .The treatment with equal volume of LB medium was used as the control group. The treated mycelial precipitates were collected by centrifugation at 8000 g for ten minutes and resuspended with 1ml PBS, then stained with PI dye at 37℃ in the dark for 20 minutes. After staining, all treated groups were washed twice with PBS and then observed under laser scanning confocal microscopy.
4.7. Detection of the effect of crude extract on the antioxidant activity of B. sorokiniana
The effects on the antioxidant activity and the inhibition mechanism of antibacterial substances were investigated by measuring the activities of superoxide dismutase (SOD), peroxidase (POD), catalase (CAT) and malondialdehyde (MDA) in B. sorokiniana mycelium treated with different levels of lipopeptide crude extracts. Briefly, 0.2 g of B. sorokiniana mature mycelium was taken and added to 1% and 2% of lipopeptide crude extract liquid as described before, respectively, and PBS was supplemented to 1ml and incubated for 2h at 28°C. The treatment group with only 1 ml of PBS added was used as a control, and the experiment was repeated three times for each treatment group. The mycelial sediment was collected by centrifugation at 4°C and 10,000g for 10 minutes and ground well in liquid nitrogen. The SOD and POD enzyme activities as well as the MDA content were determined by referring to the previous report by Yi et al [23]. The CAT enzyme activity was measured by referring to Zhou et al [49].
4.8. Pot experiments
Pot experiment using wheat seeds of Zhengmai 103 variety (purchased from Henan Qiule Seed Technology Co., Ltd., China). The efficacy of the antagonistic bacterium JK-25 for the control of common root rots was evaluated in wheat pot experiments under greenhouse conditions. Before sowing, the wheat was soaked in 0.5% sodium hypochlorite for half an hour to disinfect the surface and washed twice with sterile water, then sown in plastic pots (15 cm in diameter) containing 3.5 L of soil. Potted plants were grown for 14 days after sowing in greenhouse conditions (25°C -28°C and 30%-70% for temperature and humidity, respectively). After 14 days, a root rot spore solution (1 × 106 cfu/ml) was irrigated near the soil of wheat roots in pots, followed by 20 ml of bacterial culture filtrate. An equal volume of 50% chemical fungicide diluted 1000 times was added as a positive control, and a treatment with an equal volume of sterile water was added as a negative control. Six parallel trials were conducted for each treatment group, with each potted replicate guaranteed to have 10 or more wheat plants.
Ten days after treatment growth all wheat roots were uprooted and washed under a tap with running water for assessment of disease severity. There were five score levels: level 0 for healthy and disease free, level 1 for ≤25% disease, level 2 for >25% to 50% disease, level 3 for >50% to 75% disease , and level 4 for >75% disease. The disease incidence rate (DIR), disease index (DI) and control efficacy (CE) were calculated as follows [50]:
where n indicates the number of wheat seedlings with infected roots, N is the total number of surveyed wheat seedlings, Ni denotes the number of infected wheat seedlings of a certain level of disease, i denotes a certain level of disease, 4 denotes the highest level of disease, DIck denotes the disease index of the control group, and DIt denotes the disease index of the treatment group.
4.9. Production of extracellular enzymes and Growth Promotion
Plate test method for assessing the growth-promoting properties of antagonistic strain JK25, includes the production of indoleacetic acid (IAA), extracellular enzymes (protease, pectinase, cellulase, and chitinase), and siderophores. Where the IAA was determined by referring to the method of Alfiky et al and extracellular enzymes were detected by the methods described by Khedher et al [46,51]. Colonies of strain JK-25 were inoculated on CAS blue agar to determine siderophore production [52].
4.10. Statistical analysis
Each experiment was performed at least three times and the results were expressed as mean ± standard deviation. All data were analyzed using SPSS 22.0 with Duncan's new multiple range test, P-values < 0.05 were considered statistically significant. All statistical analyses were carried out with the help of Graph pad prism software (version 9.1.2).
5. Conclusions
The B. halotolerans JK-25 showed a very pronounced antagonistic effect against B. sorokiniana and broad-spectrum resistance to wheat pathogens. This strain has shown significant biocontrol effects on wheat pots under greenhouse conditions and has revealed a preliminary mechanism of action. Further analysis revealed that the strain had phytopromotional properties including the ability to produce substances such as siderophores. The presence of lipopeptides including surfactin in the crude extract of JK-25 fermentation broth was detected by MALDI-TOF-MS analysis. The antioxidant capacity of B.sorokiniana treated with JK-25 crude extract was significantly reduced and oxidative damage increased. In summary, antagonistic strain JK-25 has broad application prospect in biological control of common root rot in wheat and needs further research and development.
Author Contributions
Conceptualization, Y.H. and K.K.; methodology, K.K. and W.Z.; validation, K.K., Z.N. and W.Z.; formal analysis, S.W. and Y.L.; investigation, K.K and Z.N.; resources, Y.H.; data curation, K.K.; writing—original draft preparation, K.K. and W.Z.; writing—review and editing, S.W.,Y.L. and Y.H.; visualization, Z.N.; supervision, K.K..; funding acquisition, Y.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Major Science and Technology Projects in Henan Province(221100110700).
Data Availability Statement
The relevant data for this study are presented in the article for the reader's own reference.
Acknowledgments
We acknowledge Henan Agricultural University for providing some of the pathogenic fungi for this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alomari, D.Z.; Eggert, K.; von Wiren, N.; Alqudah, A.M.; Polley, A.; Plieske, J.; Ganal, M.W.; Pillen, K.; Roder, M.S. Identifying Candidate Genes for Enhancing Grain Zn Concentration in Wheat. Front Plant Sci 2018, 9, 1313. [Google Scholar] [CrossRef] [PubMed]
- Dhanarajan, G.; Rangarajan, V.; Sridhar, P.R.; Sen, R. Development and Scale-up of an Efficient and Green Process for HPLC Purification of Antimicrobial Homologues of Commercially Important Microbial Lipopeptides. ACS Sustainable Chemistry & Engineering 2016, 4, 6638–6646. [Google Scholar] [CrossRef]
- Su, J.; Zhao, J.; Zhao, S.; Li, M.; Pang, S.; Kang, Z.; Zhen, W.; Chen, S.; Chen, F.; Wang, X. Genetics of Resistance to Common Root Rot (Spot Blotch), Fusarium Crown Rot, and Sharp Eyespot in Wheat. Front Genet 2021, 12, 699342. [Google Scholar] [CrossRef]
- Al-Sadi, A.M. Bipolaris sorokiniana-Induced Black Point, Common Root Rot, and Spot Blotch Diseases of Wheat: A Review. Front Cell Infect Microbiol 2021, 11, 584899. [Google Scholar] [CrossRef] [PubMed]
- Poole, G.J.; Harries, M.; Huberli, D.; Miyan, S.; MacLeod, W.J.; Lawes, R.; McKay, A. Predicting Cereal Root Disease in Western Australia Using Soil DNA and Environmental Parameters. Phytopathology 2015, 105, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Yang, G.; Wang, J.; Song, Y.; Liu, L.; Zhao, K.; Li, Y.; Han, Z. Spatial Distribution of Root and Crown Rot Fungi Associated With Winter Wheat in the North China Plain and Its Relationship With Climate Variables. Front Microbiol 2018, 9, 1054. [Google Scholar] [CrossRef]
- Allali, K.; Goudjal, Y.; Zamoum, M.; Bouznada, K.; Sabaou, N.; Zitouni, A. Nocardiopsis dassonvillei strain MB22 from the Algerian Sahara promotes wheat seedlings growth and potentially controls the common root rot pathogen Bipolaris sorokiniana. J. Plant Pathol. 2019, 101, 1115–1125. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Dürr, C.; Schwanck, A.A.; Robin, M.-H.; Sarthou, J.-P.; Cellier, V.; Messéan, A.; Aubertot, J.-N. Integrated management of damping-off diseases. A review. Agronomy for Sustainable Development 2017, 37. [Google Scholar] [CrossRef]
- O'Sullivan, C.A.; Roper, M.M.; Myers, C.A.; Thatcher, L.F. Developing Actinobacterial Endophytes as Biocontrol Products for Fusarium pseudograminearum in Wheat. Front Bioeng Biotechnol 2021, 9, 691770. [Google Scholar] [CrossRef]
- Campanella, V.; Mandalà, C.; Angileri, V.; Miceli, C. Management of common root rot and Fusarium foot rot of wheat using Brassica carinata break crop green manure. Crop Protect. 2020, 130. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Ulrich, D.; Brandt, S.A.; Zentner, R.P.; Wang, H.; Thomas, A.G.; Olfert, O. Crop Management Effects on Root and Crown Rot of Wheat in West-Central Saskatchewan, Canada. Agron. J. 2011, 103, 756–765. [Google Scholar] [CrossRef]
- Le, C.N.; Mendes, R.; Kruijt, M.; Raaijmakers, J.M. Genetic and Phenotypic Diversity of Sclerotium rolfsii in Groundnut Fields in Central Vietnam. Plant Dis. 2012, 96, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Sahu, P.K.; Singh, S.; Gupta, A.; Singh, U.B.; Brahmaprakash, G.P.; Saxena, A.K. Antagonistic potential of bacterial endophytes and induction of systemic resistance against collar rot pathogen Sclerotium rolfsii in tomato. Biol. Control 2019, 137. [Google Scholar] [CrossRef]
- Sarkar, J.; Chakraborty, U.; Chakraborty, B.N. Induced defense response in wheat plants against Bipolaris sorokiniana following application of Bacillus safensis and Ochrobactrum pseudogrignonense. Indian Phytopathology 2018, 71, 49–58. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Kumar, M.; Sahu, P.K.; Singh, H.V.; Kumar, S.; Roy, M.; Imran, M.; Rai, J.P.; et al. Trichoderma harzianum- and Methyl Jasmonate-Induced Resistance to Bipolaris sorokiniana Through Enhanced Phenylpropanoid Activities in Bread Wheat (Triticum aestivum L.). Front Microbiol 2019, 10, 1697. [Google Scholar] [CrossRef]
- Ullah, H.; Yasmin, H.; Mumtaz, S.; Jabeen, Z.; Naz, R.; Nosheen, A.; Hassan, M.N. Multitrait Pseudomonas spp. Isolated from Monocropped Wheat (Triticum aestivum) Suppress Fusarium Root and Crown Rot. Phytopathology 2020, 110, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Villa-Rodríguez, E.; Parra-Cota, F.; Castro-Longoria, E.; López-Cervantes, J.; de los Santos-Villalobos, S. Bacillus subtilis TE3: A promising biological control agent against Bipolaris sorokiniana, the causal agent of spot blotch in wheat (Triticum turgidum L. subsp. durum). Biol. Control 2019, 132, 135–143. [Google Scholar] [CrossRef]
- Zhao, L.; Xu, Y.; Lai, X.H.; Shan, C.; Deng, Z.; Ji, Y. Screening and characterization of endophytic Bacillus and Paenibacillus strains from medicinal plant Lonicera japonica for use as potential plant growth promoters. Braz. J. Microbiol. 2015, 46, 977–989. [Google Scholar] [CrossRef]
- Miljakovic, D.; Marinkovic, J.; Balesevic-Tubic, S. The Significance of Bacillus spp. in Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8. [Google Scholar] [CrossRef]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Villarreal-Delgado, M.F.; Villa-Rodríguez, E.D.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Parra-Cota, F.I.; De los Santos-Villalobos, S. El género Bacillus como agente de control biológico y sus implicaciones en la bioseguridad agrícola. Revista Mexicana de Fitopatología, Mexican Journal of Phytopathology 2018, 36. [Google Scholar] [CrossRef]
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: a review. Biotechnology & Biotechnological Equipment 2017, 31, 446–459. [Google Scholar] [CrossRef]
- Yi, Y.; Shan, Y.; Liu, S.; Yang, Y.; Liu, Y.; Yin, Y.; Hou, Z.; Luan, P.; Li, R. Antagonistic Strain Bacillus amyloliquefaciens XZ34-1 for Controlling Bipolaris sorokiniana and Promoting Growth in Wheat. Pathogens 2021, 10. [Google Scholar] [CrossRef]
- Chen, L.; Wu, Y.D.; Chong, X.Y.; Xin, Q.H.; Wang, D.X.; Bian, K. Seed-borne endophytic Bacillus velezensis LHSB1 mediate the biocontrol of peanut stem rot caused by Sclerotium rolfsii. J. Appl. Microbiol. 2020, 128, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Aswini, K.; Sai Prasad, J.; Kumar, N.; Pathak, D.; Gond, S.; Venkadasamy, G.; Suman, A. Characterization of actinobacteria from wheat seeds for plant growth promoting traits and protection against fungal pathogens. J. Basic Microbiol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Backman, P.A.; Sikora, R.A. Endophytes: An emerging tool for biological control. Biol. Control 2008, 46, 1–3. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Shahid, M.A.; Mustafa, A.; Sayyed, R.Z.; Cura, J.A. Insights into the Interactions among Roots, Rhizosphere, and Rhizobacteria for Improving Plant Growth and Tolerance to Abiotic Stresses: A Review. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liang, J.; Zhang, C.; Wang, L.; Gao, W.; Jiang, J. Bacillus megaterium WL-3 Lipopeptides Collaborate Against Phytophthora infestans to Control Potato Late Blight and Promote Potato Plant Growth. Front Microbiol 2020, 11, 1602. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Liang, J.; Wu, L.; Gao, W.; Jiang, J. Iturin A Extracted From Bacillus subtilis WL-2 Affects Phytophthora infestans via Cell Structure Disruption, Oxidative Stress, and Energy Supply Dysfunction. Front Microbiol 2020, 11, 536083. [Google Scholar] [CrossRef]
- Rajer, F.U.; Samma, M.K.; Ali, Q.; Rajar, W.A.; Wu, H.; Raza, W.; Xie, Y.; Tahir, H.A.S.; Gao, X. Bacillus spp.-Mediated Growth Promotion of Rice Seedlings and Suppression of Bacterial Blight Disease under Greenhouse Conditions. Pathogens 2022, 11. [Google Scholar] [CrossRef]
- Lin, R.; Zhang, Q.; Yin, L.; Zhang, Y.; Yang, Q.; Liu, K.; Wang, Y.; Han, S.; Zhao, H.; Zhao, H. Isolation and characterization of a mycosubtilin homologue antagonizing Verticillium dahliae produced by Bacillus subtilis strain Z15. PLoS One 2022, 17, e0269861. [Google Scholar] [CrossRef] [PubMed]
- Villa-Rodriguez, E.; Moreno-Ulloa, A.; Castro-Longoria, E.; Parra-Cota, F.I.; de Los Santos-Villalobos, S. Integrated omics approaches for deciphering antifungal metabolites produced by a novel Bacillus species, B. cabrialesii TE3(T), against the spot blotch disease of wheat (Triticum turgidum L. subsp. durum). Microbiol. Res. 2021, 251, 126826. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.K.; Joshi, N.; Singh, I.P.; Saini, H.S. Identification of cyclic lipopeptides produced by Bacillus vallismortis R2 and their antifungal activity against Alternaria alternata. J. Appl. Microbiol. 2017, 122, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Zhou, H.; Zou, J.; Wang, X.; Zhang, R.; Xiang, Y.; Chen, Z. Bacillomycin L and surfactin contribute synergistically to the phenotypic features of Bacillus subtilis 916 and the biocontrol of rice sheath blight induced by Rhizoctonia solani. Appl Microbiol Biotechnol 2015, 99, 1897–1910. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.; Tonelli, M.L.; Figueredo, M.S.; Ibáñez, F.; Fabra, A. The lipopeptide surfactin triggers induced systemic resistance and priming state responses in Arachis hypogaea L. Eur. J. Plant Pathol. 2018, 152, 845–851. [Google Scholar] [CrossRef]
- Hazarika, D.J.; Goswami, G.; Gautom, T.; Parveen, A.; Das, P.; Barooah, M.; Boro, R.C. Lipopeptide mediated biocontrol activity of endophytic Bacillus subtilis against fungal phytopathogens. BMC Microbiol 2019, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Jamali, H.; Sharma, A.; Roohi; Srivastava, A. K. Biocontrol potential of Bacillus subtilis RH5 against sheath blight of rice caused by Rhizoctonia solani. J. Basic Microbiol. 2020, 60, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiao, Q.; Yang, H.; Huang, J.; Li, Y. Characterization of a new Bacillus velezensis as a powerful biocontrol agent against tomato gray mold. Pestic Biochem Physiol 2022, 187, 105199. [Google Scholar] [CrossRef]
- Dryden, M. Reactive oxygen species: a novel antimicrobial. Int. J. Antimicrob. Agents 2018, 51, 299–303. [Google Scholar] [CrossRef]
- Wang, D.; Gong, N.; Liu, C.; Li, S.; Guo, Z.; Wang, G.; Shang, Q.; Wang, D.; Ji, X.; Xin, Y. MnASI1 Mediates Resistance to Botrytis cinerea in Mulberry (Morus notabilis). Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: mechanisms and applications. Scientifica (Cairo) 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Mageshwaran, V.; Gupta, R.; Singh, S.; Sahu, P.K.; Singh, U.B.; Chakdar, H.; Bagul, S.Y.; Paul, S.; Singh, H.V. Endophytic Bacillus subtilis antagonize soil-borne fungal pathogens and suppress wilt complex disease in chickpea plants (Cicer arietinum L.). Front Microbiol 2022, 13, 994847. [Google Scholar] [CrossRef]
- Cheng, X.; Ji, X.; Ge, Y.; Li, J.; Qi, W.; Qiao, K. Characterization of Antagonistic Bacillus methylotrophicus Isolated From Rhizosphere and Its Biocontrol Effects on Maize Stalk Rot. Phytopathology 2019, 109, 571–581. [Google Scholar] [CrossRef]
- Shan, H.; Zhao, M.; Chen, D.; Cheng, J.; Li, J.; Feng, Z.; Ma, Z.; An, D. Biocontrol of rice blast by the phenaminomethylacetic acid producer of Bacillus methylotrophicus strain BC79. Crop Protect. 2013, 44, 29–37. [Google Scholar] [CrossRef]
- Bergey, D.H. Bergey's manual of determinative bacteriology; Lippincott Williams & Wilkins, 1994. [Google Scholar]
- Alfiky, A.; L'Haridon, F.; Abou-Mansour, E.; Weisskopf, L. Disease Inhibiting Effect of Strain Bacillus subtilis EG21 and Its Metabolites Against Potato Pathogens Phytophthora infestans and Rhizoctonia solani. Phytopathology 2022, 112, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Heng, J.; Qin, S.; Bian, K. A comprehensive understanding of the biocontrol potential of Bacillus velezensis LM2303 against Fusarium head blight. PLoS One 2018, 13, e0198560. [Google Scholar] [CrossRef] [PubMed]
- Frank Leenders1, T.H.S. , Ba¨rbel Kablitz1, Peter Franke3 and Joachim Vater1*. Rapid Typing of Bacillus subtilis Strains by their Secondary Metabolites Using Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry of Intact Cells. 1999.
- Zhou, C.; Guo, R.; Ji, S.; Fan, H.; Wang, J.; Wang, Y.; Liu, Z. Isolation of Trichoderma from forestry model base and the antifungal properties of isolate TpsT17 toward Fusarium oxysporum. Microbiol. Res. 2020, 231, 126371. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Q.; Wei, X.; Xue, Q.; Lai, H. Biocontrol effects of Penicillium griseofulvum against monkshood (Aconitum carmichaelii Debx.) root diseases caused by Sclerotium rolfsiii and Fusarium spp. J. Appl. Microbiol. 2019, 127, 1532–1545. [Google Scholar] [CrossRef] [PubMed]
- Ben Khedher, S.; Mejdoub-Trabelsi, B.; Tounsi, S. Biological potential of Bacillus subtilis V26 for the control of Fusarium wilt and tuber dry rot on potato caused by Fusarium species and the promotion of plant growth. Biol. Control 2021, 152. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree constructed based on 16S rRNA sequences of JK-25. Analyze the comparison results with MAGA 7.0 software and build a tree using the Neighbor Joining method (1000 bootstrap values).
Figure 1.
Phylogenetic tree constructed based on 16S rRNA sequences of JK-25. Analyze the comparison results with MAGA 7.0 software and build a tree using the Neighbor Joining method (1000 bootstrap values).
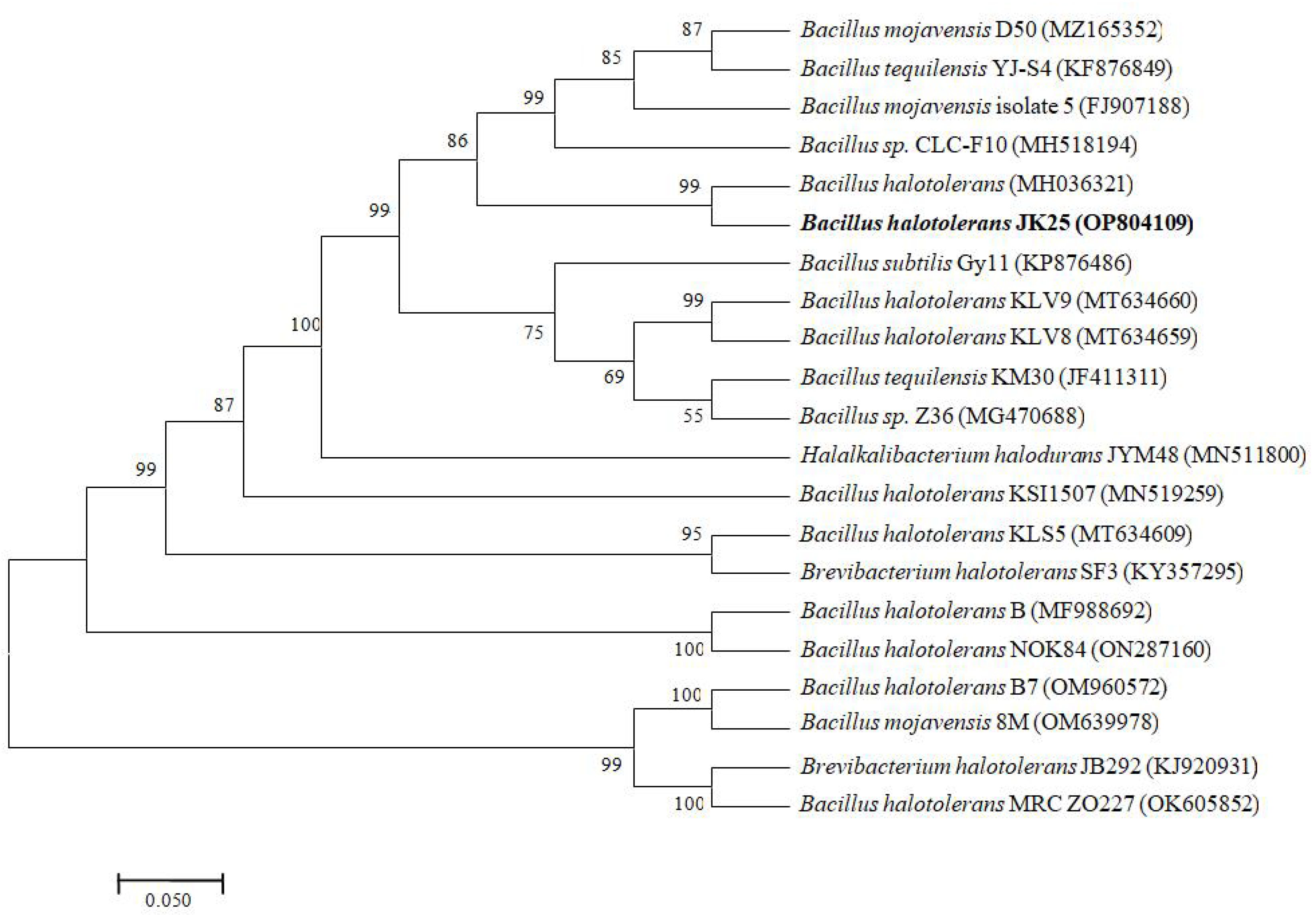
Figure 2.
Antagonistic effect of strain JK-25 on B. sorokiniana on PDA medium.
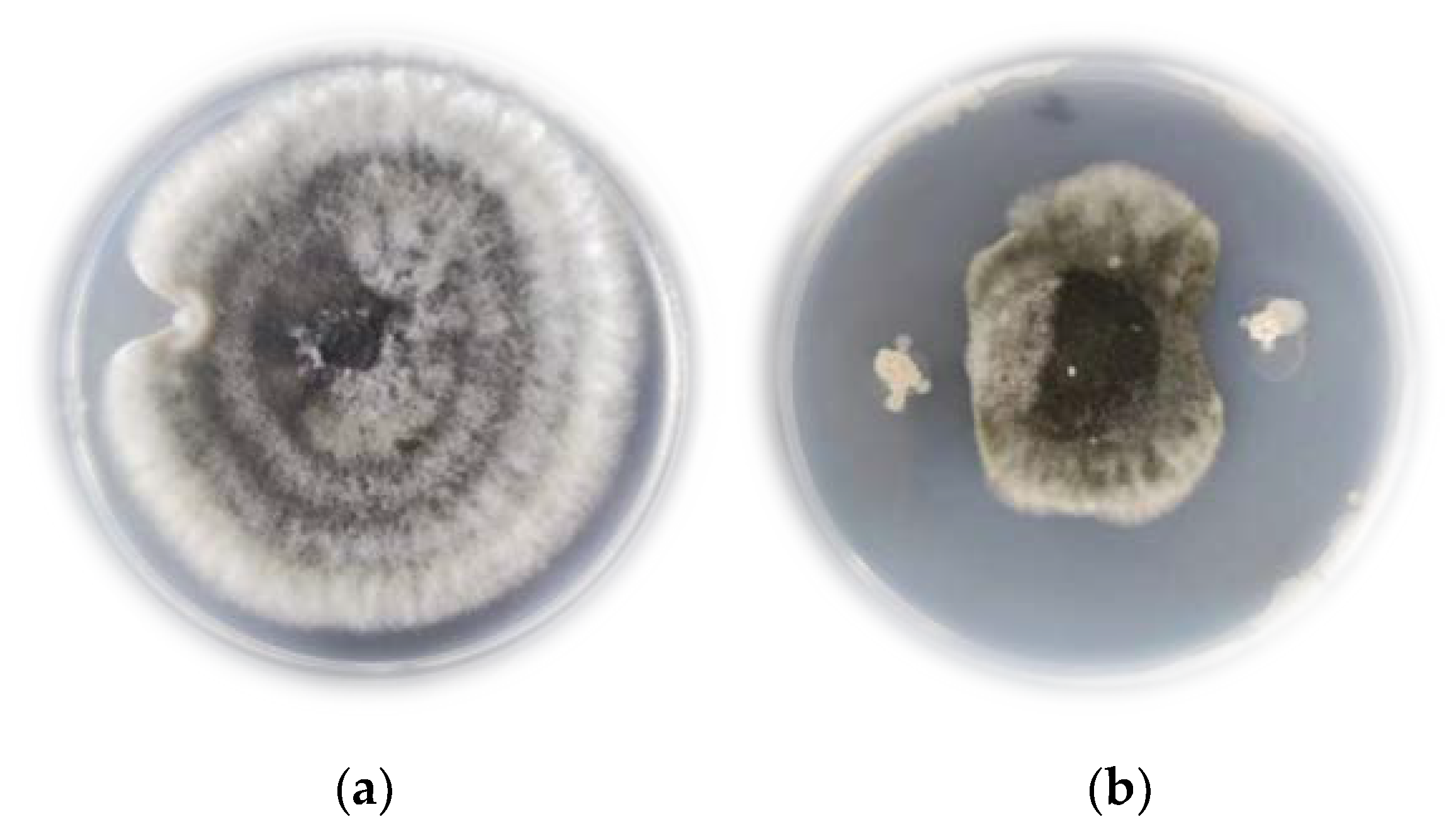
Figure 3.
JK-25 exerts an antagonistic effect on mycelial growth and spore germination of B.sorokiniana. Data are presented as the average ± the standard deviation, and * indicates a significant difference between treatment groups within a parameter (Duncan’s multiple range test at p ≤ 0.05.).
Figure 3.
JK-25 exerts an antagonistic effect on mycelial growth and spore germination of B.sorokiniana. Data are presented as the average ± the standard deviation, and * indicates a significant difference between treatment groups within a parameter (Duncan’s multiple range test at p ≤ 0.05.).
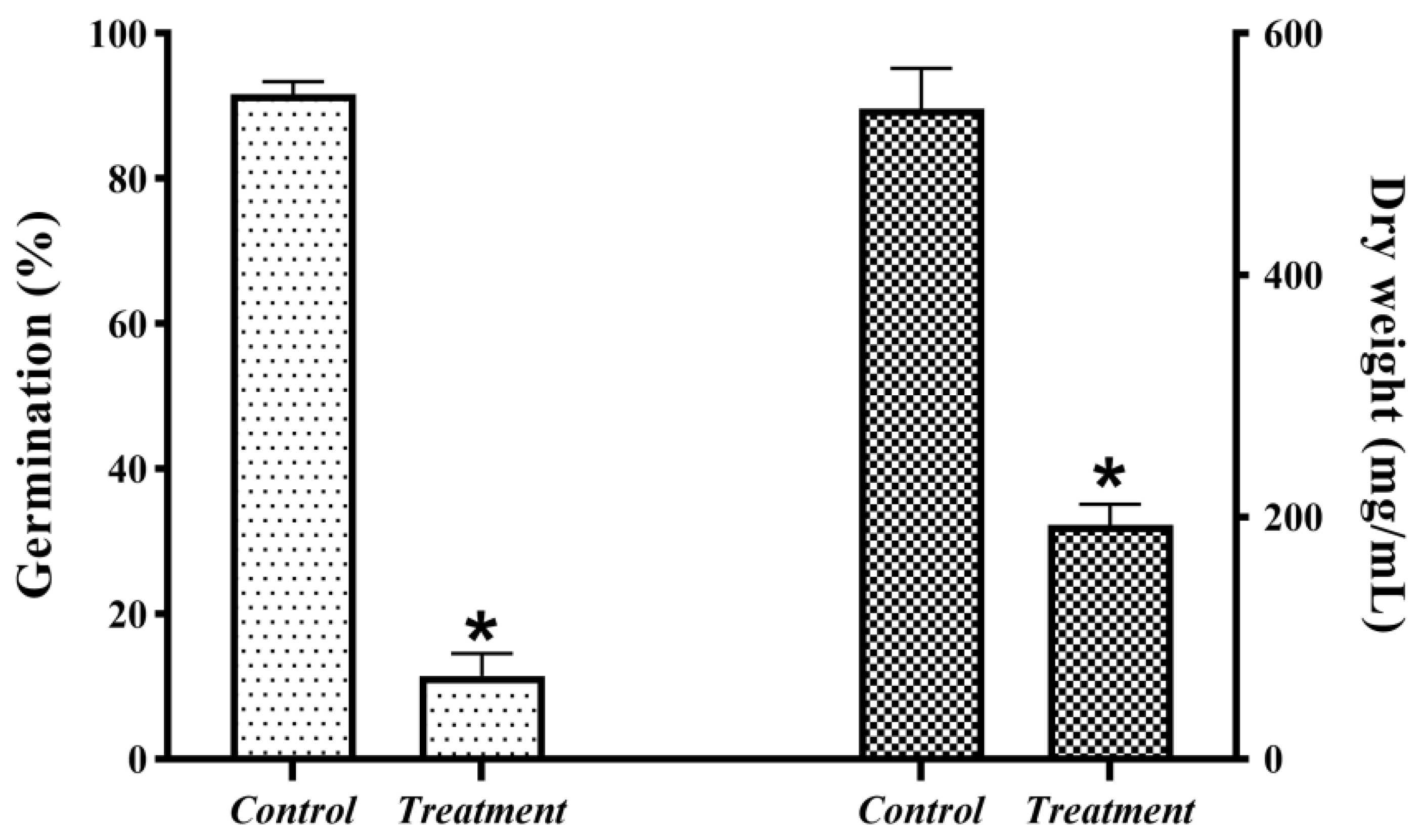
Figure 4.
MALDI-TOF mass spectra from Bacillus halotolerans JK-25 with the m/z range 1000–1140. m/z represent their corresponding lipopeptides.
Figure 4.
MALDI-TOF mass spectra from Bacillus halotolerans JK-25 with the m/z range 1000–1140. m/z represent their corresponding lipopeptides.
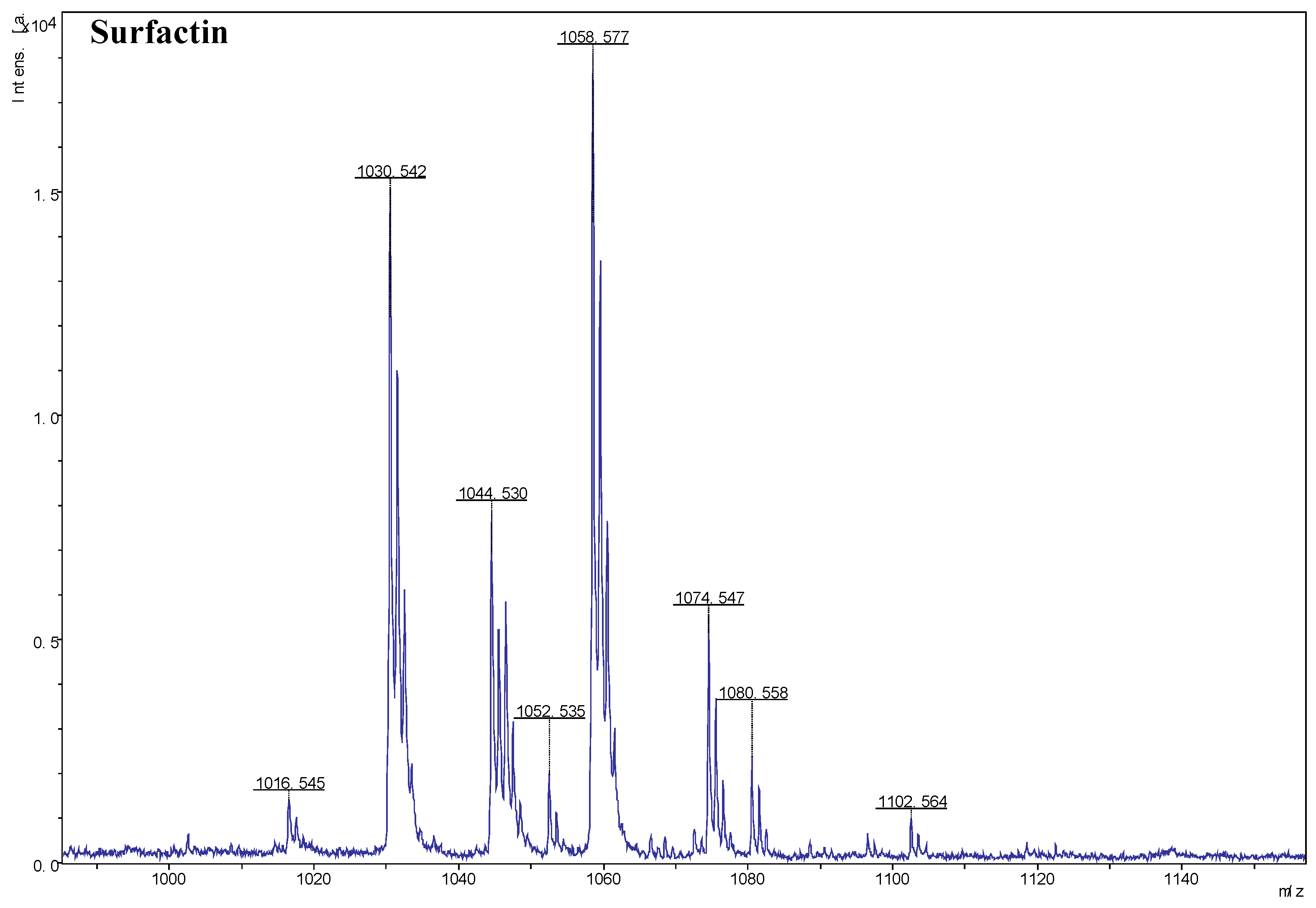
Figure 5.
Microscopy analyses of B.sorokiniana hyphae treated with JK-25 crude extract. The figure shows the control and treatment groups, respectively, and the images in each group include fluorescent field (Propidium iodide), bright field and merged mode conditions.
Figure 5.
Microscopy analyses of B.sorokiniana hyphae treated with JK-25 crude extract. The figure shows the control and treatment groups, respectively, and the images in each group include fluorescent field (Propidium iodide), bright field and merged mode conditions.
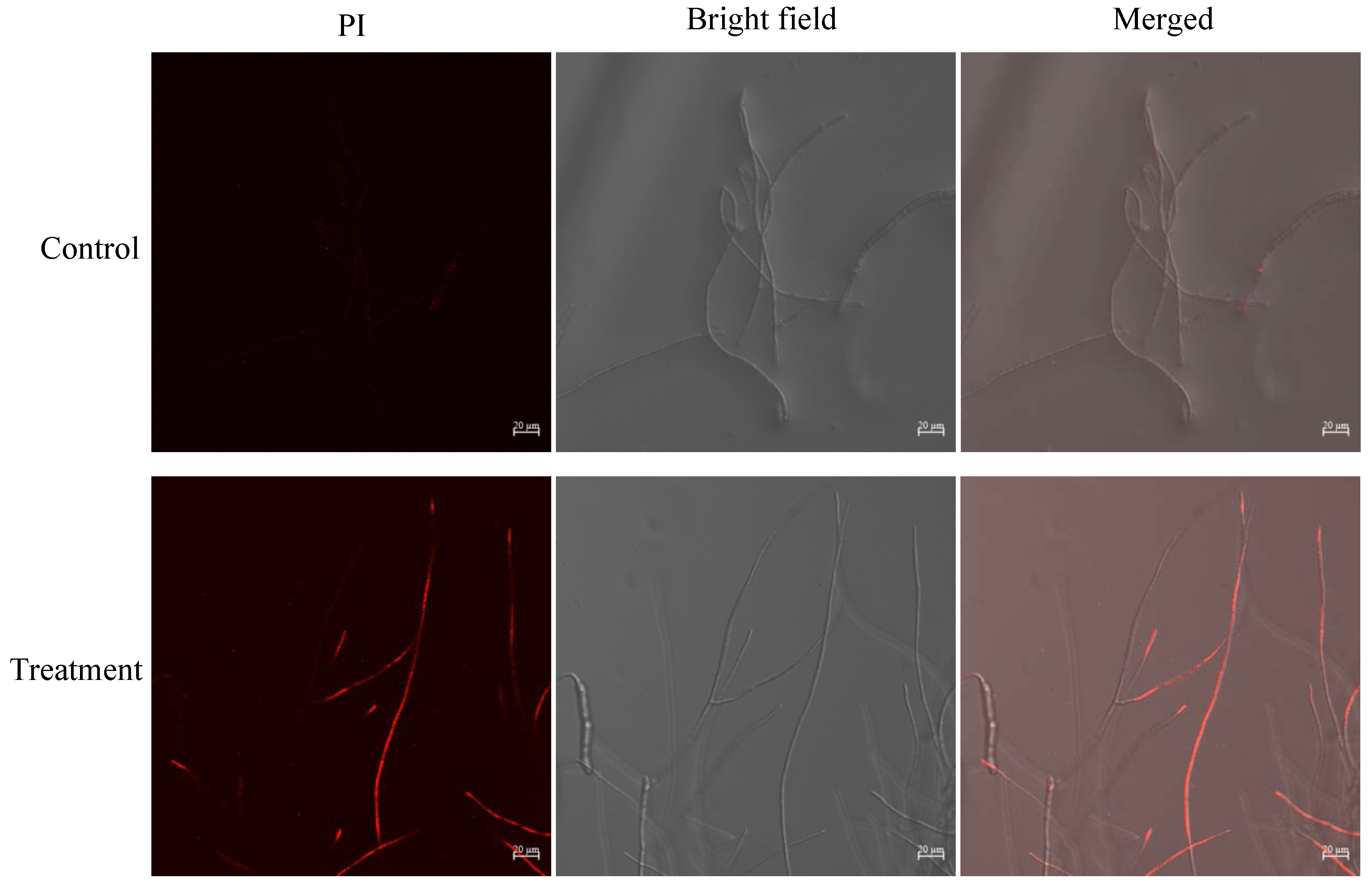
Figure 6.
Effect of lipopeptide crude extracts on the antioxidant activity of B.sorokiniana mycelium. Control (equal volume PBS treatment), T1 (1% crude extract), T2 (2% crude extract). (a): superoxide dismutase (SOD) activity, (b): peroxide (POD) activity, (c): catalase (CAT) activity, (d): malondialdehyde (MDA) content. Data are presented as the average ± the standard deviation, and * indicates a significant difference between treatment groups within a parameter (Duncan’s multiple range test at p ≤ 0.05.).
Figure 6.
Effect of lipopeptide crude extracts on the antioxidant activity of B.sorokiniana mycelium. Control (equal volume PBS treatment), T1 (1% crude extract), T2 (2% crude extract). (a): superoxide dismutase (SOD) activity, (b): peroxide (POD) activity, (c): catalase (CAT) activity, (d): malondialdehyde (MDA) content. Data are presented as the average ± the standard deviation, and * indicates a significant difference between treatment groups within a parameter (Duncan’s multiple range test at p ≤ 0.05.).

Figure 7.
Biocontrol efficiency of JK-25 CF on common root rot caused by B. sorokiniana. In all experimental groups, the wheat was pre-inoculated with pathogen during growth. (a1, a2) were sterile water treatment (control); (b1,b2) were 50% carbendazim treatment; (c1,c2) were JK-25 CF treatment.
Figure 7.
Biocontrol efficiency of JK-25 CF on common root rot caused by B. sorokiniana. In all experimental groups, the wheat was pre-inoculated with pathogen during growth. (a1, a2) were sterile water treatment (control); (b1,b2) were 50% carbendazim treatment; (c1,c2) were JK-25 CF treatment.


Table 1.
Inhibition rate of pathogens by five strains of bacteria.
| Strain | Inhibition Rate(%) | |||
|---|---|---|---|---|
|
Bipolaris sorokiniana |
Fusarium oxysporum |
Fusarium graminearum |
Rhizoctonia zeae |
|
| JK-5 | 78.38 ± 2.63ab | 76.97 ± 1.22a | 69.63 ± 1.46b | 25.73 ± 7.9c |
| JK-13 | 75.4 ± 2.79b | 71 ± 2.08b | 65.2 ± 2.75bc | 45.7 ± 2.21b |
| JK-25 | 82.63 ± 0.67a | 80.47 ± 1.4a | 75.33 ± 1.27a | 59.73 ± 2.45a |
| JK-44 | 66.06 ± 2.07c | 57.2 ± 1.97c | 62.47 ± 3.2c | 30 ± 8.94c |
| JK-58 | 67.2 ± 3.44c | 55.3 ± 3.22c | 64.87 ± 3.33bc | 32.47 ± 2.54c |
Values in the table are the means ± standard deviation of the results of at least nine replicates. Different letters indicate significant differences after Duncan′s multiple range test (p ≤ 0.05).
Table 2.
Physiological and biochemical characteristics of antagonistic strain JK-25.
| Biochemical tests | Reaction | Colony morphology | Description |
|---|---|---|---|
| Gram stain | + | Endospores | Present |
| Methyl red test | - | Morphology | Rounded |
| Voges-Proskauer test | + | Pigment | Creamy white |
| Indole test | - | Surface | Neat and smooth |
| Nitrate reduction | + | Margin | rough |
| Catalase | + | Opacity | Opaque |
| 10% salt tolerance test | + | ||
| Glucose fermentation | + | ||
| Starch hydrolusis | + | ||
| Citrate test | + | ||
| Gelatin liquefaction | + |
+”Positive; “–”negative..
Table 3.
Biocontrol efficacy of JK-25 on common root rot
| Treatments | Disease Incidence Rate (%) | Disease Index | Control Efficacy (%) |
|---|---|---|---|
| Sterile water control | 98.75±3.81 a | 71.59±7.24 a | - |
| Carbendazim control | 42.37±8.96 c | 15.85±2.41 b | 77.86±4.32 |
| CF treatment | 49.65±9.54 b | 20.20±3.58 b | 72.06±6.94 |
Values represent means ± SDs. The presence of different lowercase letters within the same column indicated a significant difference between treatments (P<0.05).
Table 4.
Identification of JK-25 growth-promoting conditions
| Bioboosters | Reaction |
|---|---|
| protease | + |
| pectinase | + |
| cellulase | + |
| chitinase | - |
| siderophores | + |
| indoleacetic acid (IAA) | - |
“+”Positive; “–”negative..
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



