
Submitted:
22 January 2023
Posted:
24 January 2023
You are already at the latest version
Abstract
In this review, we present several extracellular proteases, enzymes, membrane permeases, and transporters as essential accessories proteins for nutrient assimilation, conservation, and transportation as determined by nutrient repletion or depletion. As an obligate aerobic pathogen, it is crucial for invading Cryptococcus (C.) neoformans to negotiate its adaptation to human internal organs like the brain and spinal cord, where the oxygen level is low compared to peripheral organs. Besides, essential metals like copper and iron are important cofactors to functional proteins; however, these metals are not usually freely available to invading human pathogens. Again, the phagolysosome low pH with glucose paucity, internal temperature, immune response, and complex extracellular matrixes are challenging environments that must be circumvented by C. neoformans in the systemic tissues for survival, adaptation, and infection in humans. We review extensive works on several extracellular proteases, enzymes, membrane permeases and transporters orchestrated by different transcription factors and present these proteins as weapons needed to outwit systemic resistance to invading pathogens. Lastly, we examine the extracellular secretory vesicles of C. neoformans as “an exosomal virulence bag” that harbours urease, laccase, phosphatase, and capsular components as additional secretory weapons for tissue invasion and persistence.
Keywords:
Cryptococcus
; extracellular protease
; biofilm
; secretory vesicles
; hypoxia
; hydrolytic enzyme
; homeostasis
; transcription factors
; membrane permeases
; membrane transporters
1. Introduction
C. neoformans is an opportunistic yeast associated with humous soil contaminated with Aves dropping and around trees. As an encapsulated and obligate aerobe, it becomes pathogenic and able to infect, invade, and colonise various animal and human organ systems. Being an environmental saprophytic fungus, the internalisation of extracellular nutrients is necessary for survival of this fungus. With the help of numerous membrane proteins (permeases and transporters), extracellular enzymes, and cytoplasmic enzymes, digested nutrients can be assimilated.
The assimilation of extracellularly digested nutrients and micro-nutrients is a complex metabolic system. Besides, other membrane proteins are strategically made active for adaptation, survival, drug resistance, and human infection. The redox process uses membrane-bound proteins to obtain ions such as Cu2+, Fe2+, and Zn2+ for membrane potentials. Pumps, such as Na+/K+-ATPase and P-type Na+-ATPase (Ena1), are used to maintain membrane polarisation, while ABC (Afr1) and ABD-type multidrug homologue pumps (Pdr5, Yor1, and Snq1) are deployed for drug resistance. Several transporters and permeases like Ca2+ transporters (Cch1, Eca1, and Vcx1), Na+/H+ antiporter (Nha1), aquaporin (Aqp1), phosphate affinity permeases (Pho) are implemented for cellular homeostasis, tolerance, resistance, antioxidation, and coping with environmental stress (oxidative and osmotic). Altogether, these have been studied to be important in cryptococcal cell survival, invasion, colonisation, and infection [1,2].
One of the attributes of cryptococcal cells like C. albicans, A. fumigatus, and P. aeruginosa is the ability to form biofilm. This is an important phenotypic display of invasive pathogenic fungi and bacteria that involves a conglomerate of cells working together with common goals – colonisation, extracellular matrix modelling, quorum sensing, resistance, protection, adhesion, survival, adaptation, communication, sharing, maturation, filamentation, and invasion [3,4,5,6,7,8], Figure 1. Biofilm formation in C. neoformans is metabolically regulated, induced by unfavourable conditions, and capsular components [9]. Either in vivo or in vitro, C. neoformans can form a biofilm, even on surfaces and invasive clinical instruments [9]. Studies have shown that the redox process, anaerobic metabolism, substrate-level phosphorylation, proteolysis, and anti-osmotic processes are upregulated during biofilm formation, but general metabolic processes, replication, transcription, translation, transportation, permeases, pump systems are generally repressed [10].
C. neoformans, an opportunistic environmental pathogen, encounters abiotic factors with adequate cellular taxis to survive these conditions. It also encounters predatory association with surrounding biotic soil organisms. Amoeboid protozoans can “catch” and “kill” internalised cryptococcal cells similarly to mammalian macrophages that phagocytose invading pathogens. Surprisingly, though cryptococcal cells can be engulfed, the fungi can survive and replicate intracellularly within the soil amoeba. These survival mechanisms are insurmountable, especially in immunocompromised patients when infected. This invariably promotes gradual tissue intrusion, leading to a life-threatening condition, meningoencephalitis, due to fungemia.
Based on their immediate interaction, cryptococcal cells could be environmental or clinical. Both showed similar pathogenicity in animal models [11] because of the similarity in the virulence factors – capsule, melanin, extracellular hydrolytic enzymes, and vesicular virulence bags (secretory vesicles containing microbodies, laccase, urease, and phosphatase) [12]. C. neoformans reprograms its gene expression profile to survive inadequate nutrient supply by upregulating the membrane transporters and enzymes involved in the shuttling of carbon intermediates from alternative pathways such as β-oxidation (mitochondria function) and glyoxylate cycle (peroxisome function) when phagocytosed by amoeba/macrophage. These two pathways fetch cryptococcal cells acetyl-CoA [13], which is converted via gluconeogenesis into various sugars needed for the cell wall components [14]. In addition, C. neoformans resorts to anabolic pathways to generate resourceful intermediates for virulence expression, and this is observed with increased activity of fructose-2,6-bisphosphatase (encoded by Fbp1) when phagocytosed by amoeba and increased phosphoenolpyruvate carboxykinase activity (encoded by Pck1) when phagocytosed by macrophages [13,15].
Besides these, several other upregulated transcription genes during phagocytosis have been reported. These include genes for nutrient absorption (amino acids, copper, and iron), antioxidative stress genes, and genes involved in autophagy, apoptosis, and mating [16]. In extreme cases, the redox process, substrate-level phosphorylation, and anaerobic decarboxylation can be employed to generate energy that will maintain cellular function and growth [10]; however, the lack of glucose or other utilisable sugars generally impairs cryptococcal proliferation, virulence, persistence, and tissue invasion as displayed by Δpyk1, Δpyk1Δmig1, Δmig1, Δpck1, Δhxk1, Δhxk2, and Δhxk1Δhxk2 mutants in the presence of glucose, glycerol, and lactate [17]. These self-defence strategies are primarily enrooted in the signalling cascade events that interpret environmental factors into signalling sensors for effective metabolic and physiological responses [18]. For example, Fan et al. reported that genes in the Gpa1-cAMP-Pka pathway are upregulated during phagocytosis [16]. Contrarily, C. neoformans retrogressively limit the ribosome-related functional gene process and translation initiation or elongation [16], which tactically relates a central control response to phagocytosis and the initiation of spores for survival.
Though similarities occur in the responses of C. neoformans to soil amoeba and mammalian macrophages based on their intracellular attack, nevertheless, the ecological niches surrounding the amoeba are quite different from the extracellular environment. Interaction of C. neoformans with the soil amoeba is usually at the ambient temperature (28 – 30oC) in a slightly acidic environment, while the macrophage is at the physiological temperature of 37oC with a near-neutral pH range (7.2 – 7.4). By equilibrating the natural environment of Acanthamoeba castellanii and murine macrophage to C. neoformans, Derengowski et al. showed massive similarities in the modulation of several enzymes and membrane transporters involved in all significant metabolic systems with at least 2-fold increase as compared to the non-phagocytosed C. neoformans; however, only a few numbers of these proteins were differentially modulated [13].
So, the involvement of several membrane transporters, permeases, and extracellular proteins in the pathogenesis of C. neoformans is critical, as also their deployment in the nutrient inadequacy. Unfortunately, targeting many of these proteins for antifungal management may probably be a come by attempt as most of the proteins are common to human cells. We review several of these proteins and present a succinct analysis of their functional relevance to cryptococcal survival and adaptation in nutrient repletion or depletion, and in human infections. Careful consideration of their consequences to human cells is needed if any of these proteins is considered a target for antifungal formulation against invasive cryptococcal infections in humans.
4. Regulatory Transcription Factors in Cryptococcal Cells Control Membrane Transporters, Permeases and Enzymes Activities in Response to Nutrient Repletion/Depletion
The environmental transition of C. neoformans enables this fungus to cause infection in the susceptible hosts. The adaptation and survival of this pathogen in the nutrient-deprived macrophage milieu when phagocytosed are paramount to nutrient acquisition, assimilation, and metabolism. C. neoformans tactically switch on the important and inevitable transcription factors while some, considered unimportant and redundant, are temporarily switched off. When phagocytosed, the induced transcription factors include genes expressed for membrane transporters and secretion, cell wall formation, anti-oxidative and nitrosative stress, anti-autophagy and peroxisome functions, intracellular metabolism, mating, and genomic repair. However, the repressed genes are involved in ribosomal metabolisms and translational processes, most of which are repressed as the infection progresses [16]. This means that C. neoformans possesses a highly classical environmental sensor, fast enough to help decipher the micro-conditional changes and subsequently release the fort of transcription factors for nutrient acquisition that translate into cellular morphology for immune evasion and induction of infections.
4.1. Phospholipases
The secretion of extracellular phospholipases (PL) by C. neoformans was investigated by Chen et al., where a cluster of PL such as PLB, lysoPL hydrolase, and lysoPL transacylase/acyltransferase was detected and every Δplb1 mutant usually marked with a drastic reduction in the activity of each enzyme compared to the wt [19,20]. By popular demand, PLB is generally used to represent the three enzymes. By correlation, PL activity corroborated the mortality rate and mucosal invasion by Candida albicans in animal models [21]. Similarly, C. neoformans may use these hydrolytic enzymes to invade tissue and cross the BBB. The production of PL appeared similar in environmental and clinical strains of C. neoformans var. gattii, but the environmental isolates of C. neoformans var. neoformans produced more PL than the clinical isolates [19]. Various isolates of C. neoformans possess a different degree of PL secretion, which correlates with the fungal virulence and tissue burden in the lungs and brains of mice inoculated intravenously [19,20].
PLs, best secreted at 37oC [22], are a highly diverse group of hydrolytic enzymes targeting the ester linkages of glycerophospholipids carbonyl linkage and can also transfer acyl chain to the lysophospholipid to form diacylglycerophospholipid. These three oligomeric proteins are borne on the same coding gene, the Plb gene, just like in other yeasts such as Penicillium notatum [23], S. cerevisiae [24], and C. albicans [25]. Chen et al. showed that PLs are well secreted at about 6 – 10 hours of cell culture at 30oC, with optimal activity at pH 3.5 – 4.5 and stability at pH 3.8. The enzyme activity was not affected by exogenous serine protease inhibitors such as leupeptin, phenylmethylsulphonyl fluoride (PMSF), divalent cations (Ca2+, Mg2+, and Zn2+), and EDTA [19]. The stability of this enzyme at acidic pH vividly reveals one of the strategies put in place by C. neoformans to tolerate the micro-acidic environment of the phagosome, and, in so doing, the PLs may break down the phagosomal membrane leading to tissue dissemination.
The Ste12α gene has been implicated in the regulation of extracellular PL. Deletion of the Ste12α gene significantly reduced the production of PL in the egg yolk agar media [26]. As crucial as PLs are to the virulence of C. neoformans, the absence of these hydrolytic enzymes may not affect the masterminding virulence factors/phenotypes such as growth at 37oC, urease activity, capsule, and melanin formation. However, such a mutant is less virulent in the mouse inhalation and rabbit meningitis models when compared to the wt in so much that its growth is defective in a macrophage-like cell line [20]. The attenuated virulence observed in the Δplb1 mutant must have come from the lower density of the capsule; notwithstanding, the capsule and the cell wall dimensions are relatively retained compared to the wt [22].
The attenuated virulence observed in the Δplb1 mutant is likely comparable to Δure1 and Δlac1. PLB and their components are generally associated with C. neoformans virulence, yet an infection can still progress in their absence but at a slower rate. Increasing the temperature from 30 – 37oC promotes transmigration of the PLB from the cell membrane to the cell wall with a concomitant increase in the cytoplasmic translation, perhaps due to constitutive secretion from the Golgi apparatus. However, secretion of PLB under this heat stress is highly minimised to maintain cell wall integrity and promote membrane homeostasis [22].
The homologous Ssn3/Ume5 gene was serendipitously discovered adjacent to the Plb1 locus in C. neoformans. Thus, like S. cerevisiae, C. neoformans may be using the product of this low transcript to repress the transcription factor Hsp70 family. However, this repressing protein, known as Ume5p, is a cyclin-dependent protein kinase, which is sensitive to temperature- or oxidative-induced degradation via ubiquitin-mediated proteolysis [27], and may not be playing any significant role in the virulence factors [20].
Covalently, PLB is usually attached to the cell wall chitin protruding β-1,3-linked glucan via its β-1,6-linked glucan. This arrangement is anchored by the glycosylphosphatidylinositol (GPI) on the membrane lipid raft proteins. The release of PLB from the GPI-anchor is probably orchestrated by the phosphatidylinositol-specific phospholipase C (PI-PLC, putatively encoded by Plc1 and Plc2), and the inclusion of β-1,6-linked glucan to this release confirms the cell wall localisation of the PLB [22,28]. This shows that PLB constitutes part of the proteins involved in the cell wall integrity because Δplb1 exhibited morphological cell wall defect, which is sensitive to disruption by SDS and Congo red; however, the growth is not affected by caffeine [22]. The Δplc1 C. neoformans var. grubii strain H99 mutant, which failed to produce melanin and refused to grow at 37oC, exhibited less secretion of PLC with concomitant accumulation of cytoplasmic PLB because of the impaired secretion of PLB [28]. Furthermore, this mutant displayed high antifungal sensitivity (to 5-FC, azoles, and AmpB but not CpF), poor replication, and defective cell wall characterised by cell clumping and irregular morphology as confirmed by the diffusion of Calcofluor white staining and sensitivity to cell wall disruptors (Congo red, caffeine, and SDS – check Supplementary 1 endnote for details) due to impaired activation of Pkc/MAPK pathway [28].
Because the Δplc1 mutant had poor growth at 37oC, its virulence was attenuated in the mice study. Not only this, even at 25oC, this mutant failed to kill a significant number of Caenorhabditis elegans compared to the wt and reconstituted mutant [28]. Factually, the few numbers of C. elegans killed by this mutant means that attenuated virulence associated with this mutant is independent of temperature; after all, other virulence determinants are simultaneously attenuated as well. The actual killing of the worms at 25oC by the wt and reconstituted mutant indicates that the Plc1 virulence can be temperature-independent and that certain other factors may augment this virulence. In all, the Δplc2 mutant failed to exhibit any other major defect, and it is as active as the wt.
The IP3 produced by Plc1, besides the ER Ca2+ release, serves as a substrate for IP3 kinase (encoded by Arg1) to produce inositol polyphosphate. Expectedly, the Δplc1 mutant generally accumulates PIP2 but a reduced level of IP3, whereas in the Δarg1 mutant, IP3 will be accumulated [29]. Invariably, these two mutants shared similar phenotypic defects, including impaired thermotolerance, defective cell walls, impaired virulence factors, improper cell division, and defective mating and filamentation. Further analysis showed that Δarg1 accumulated larger intracellular vacuoles (excessive vacuolar fusion) than the wt and Δplc1 mutant [29]. Therefore, Arg1p inositol polyphosphate anabolism (IP4-8) is as important as the Plc1p catabolism for C. neoformans virulence, and that IP3 is a biological relay molecule for active Plc1p.
In another related work, efficient antioxidant control has been linked to PLB production. Mutation of Cu/Zn SOD encoded by the Sod1 gene has been characterised by a severe reduction in the activities of phospholipase B, urease, and laccase leading to attenuated virulence manifested in the low brain colonisation and persistence. However, this failed to affect the capsule formation under LIM, and the mutant culture media were characterised by high mannitol production that probably acted as an antioxidant to complement the loss of Cu/Zn SOD activity [30,31] (for details on the phenotypic display of different cryptococcal cell mutants, check Supplementary 1).
4.2. Phosphate Depletion
Ordinarily, C. neoformans prefers an acidic environment to neutral-alkaline or physiological pH, perhaps because of phosphate availability in the acidic environment. Intracellular inorganic phosphate (Pi) is highly essential for kinases, activation of membrane transporter, membrane biogenesis, coupling reactions, and genomic stabilisation. Very importantly, the production of second messengers such as cAMP, IPC, and DAG, which are essential for virulence (melanin and capsule), mating, filamentation, membrane biogenesis and integrity, are essential phosphate-related metabolic products. It is likely possible that the IPC formed from inositol-phosphoryl ceramide synthase 1 activity is salvaged in the Golgi apparatus to produce membrane sphingolipids. Furthermore, the IPC can readily form a sterol pocket [32], which may take ergosterol to be fused at the membrane bilipid layer for fluidity and rafting. In S. cerevisiae, IPC-mannosyl transferase (encoded by redundant Csg1 and Csh1 genes) can take mannose instead of sterol to form mannose-IPC in the presence of intracellular Ca2+ -binding protein (encoded by Csg2 gene) [33,34]. With more IP, mannose diinositolphosphoryl ceramide can be produced [35]. It may be possible that C. neoformans employed the same phosphate-dependent glycosphingolipid intermediates as membrane components to increase membrane thickness and thermal stability. Therefore, phosphate-containing intermediates are crucial to C. neoformans adaptation, survival, and pathogenesis.
C. neoformans deploys arrays of kinases, membrane transporters, and transcriptional regulatory factors to facilitate the phosphate-sensing and acquisition (PHO) pathway, a mechanism that seems to be absent in human cells. This enables the fungi to assimilate phosphate and perhaps store it as polyphosphate. These arrays of proteins and regulatory factors include:
- ❖
- plasma membrane high-affinity phosphate transporters (including Cdk encoded by Pho85, Pho85-cyclin encoded by Pho80, Cdk inhibitor of phosphate signalling pathway encoded by Pho81, and transcriptional regulatory factor encoded by Pho4),
- ❖
- low-affinity vacuolar phosphate transporter encoded by Pho90,
- ❖
- polyphosphate storage and processing (including vacuolar transport chaperone encoded by Vtc4, exopolyphosphatase encoded by Xpp1, endopolyphosphatase encoded by Epp1, and diacylglycerol trimethylhomoserine (DGTS – betaine lipid) synthase encoded by Bta1),
- ❖
- mitochondrial phosphate transporter encoded by putative Mip1 and Mip2 genes, and
- ❖
Generally, Pho4 gene expression is triggered whenever the level of Pi is critically low [36]. The high level of Pi increases the phosphorylation of Pho4p mediated by the Pho80-Pho85 complex because, under this condition, the Pho81 remained repressed. Contrarily, a low level of Pi activates the Pho81 expression to deactivate the Pho80-Pho85 complex and dephosphorylate the Pho4p regulatory factor, thereby increasing the induction and activation of Pho84, Pho89, and Pho840, which are high-affinity transmembrane Pi transporters and at the same time promoting the activities of Pho2 and constitutive Pho1 (both are orthologues of S. cerevisiae Pho5 encoding acid phosphatase) and Pho8 (encoding alkaline phosphatase).
This condition may probably promote the phosphate-scavenging activities of extracellular and vacuolar acid phosphatase (encoded by the Aph1 gene) secreted by C. neoformans. The phosphate-released activity of this enzyme, under phosphate starvation, has been demonstrated against expendable metabolites such as β-glycerol phosphate, α-D-glucose-1-phosphate, AMP, mannose-6-phosphate, O-phosphotyrosine. This enzyme, however, displayed less activity with ATP but no activity with phosphoserine and phosphothreonine [38]. In vitro assay with pNPP over an acidic pH range (5.0 – 6.9) showed that acidic phosphatase activity decreases as the pH approaches neutral value, which explains why C. neoformans survive phagolysosome (pH 4 – 5), cryptococcomas (pH 5.5), and inflammatory tissue (pH 6.5 – 6.9) [38]. The orthologues of S. cerevisiae Pho5 phosphate-repressed acid phosphatase gene have been identified in cryptococcal cells. These putative genes include Pho1 (constitutive acid phosphatase), Pho2 (secreted and vacuolar acid phosphatase), and Pho3 [36], while Lev et al. identified and designated these genes as Aph1 (similar to Pho2), Aph2, Aph3 (encoding phytases), and Aph4 (perhaps similar to Pho1) [38,39]. Furthermore, Aph2, Aph3, and Aph4 are suspected to be intracellular [39].
Experimentally, phosphate-starvation induces the expression of Aph1 and secretion of acid phosphatase when co-cultured with activated THP-1 monocytes and such Δaph1 mutants displayed attenuated virulence because infected mice survived longer than those infected with the wt serotype A H99 [38]. Mutation of Pho84, Pho89, and Pho840 genes (ΔΔΔpho) in C. neoformans interfered with growth in low-phosphate media; such mutants further displayed reduced survival and attenuated the virulence. However, the Δvtc4 mutant, which encoded the polyphosphate polymerase, behaved normally like the wt in terms of infection, but such a mutant lacked effective tissue invasion and dissemination from the lungs and failed to induce in vitro blood coagulation [37]. At 30 or 37oC, the impact of single/double mutations of Pho genes was not as conspicuous as the triple ΔΔΔpho mutant when the growth in the acidic-to-alkaline pH and non-to-low phosphate supplement media was examined. However, a higher phosphate supplement (100 mM) was able to restore the poor growth [37].
Regulatory evidence of high-affinity phosphate transporters is glaring. The ΔΔΔpho mutants inclined to accumulate intracellular sodium, iron, and zinc but failed to accumulate phosphate. This is tantamount to the poor and unrecovered formation of capsule and melanin compared to the mutants with either single or double gene disruption [37]. Again, contrary to Δvtc4 and Δepp-xpp1 mutants, viable ΔΔΔpho mutant failed to replicate in the macrophage [37].
The regulatory role of Pho4 in facilitating the acquisition of Pi is inevitable. This helix-loop-helix (Hlh3) transcription factor senses Pi starvation, and the cytoplasmic-nucleus translocation is initiated to induce the expression of phosphate sensing and acquisition genes. Thus, the growth defect, hypovirulence, inefficient tissue invasion/systemic dissemination, and other phenotypic traits of Δpho4 mutant can be restored with phosphate supplementation; however, these seem irreversible in an alkaline pH media and physiological pH [39]. Apparently, this alkaline pH intolerance creates phosphate starvation, which could have prevented the invasion and dissemination of cryptococcal infections, but the expression of the pho4 gene enables this fungus to subtly acquire phosphate in this ostensive environment through the activation of the PHO pathway.
4.3. Iron Depletion
Strategically, human physiological conditions never allow invading pathogens easy access to iron because iron is not usually found circulating without being locked in a storage protein like transferrin or transport protein like haemoglobin. Hence, invading pathogens must strive to obtain iron through the arsenal of high-affinity iron uptake and acquisition proteins. The arrays of membrane transporters (such as siderophores) and intracellularly expressed regulatory proteins (such as Cir1p) are released for iron acquisition and homeostasis (for details of cellular events that induced/repressed different membrane transporters and permeases in C. neoformans, check Supplementary 2). Interestingly, C. neoformans mainly released specific iron acquisition proteins based on the available iron sources in vivo or in vitro. Besides, C. neoformans can direct the expression of HapX or Hap3 to shut down other iron-dependent functional pathways in LIM to maximise the utilisation of the scarce iron for more growth while capsule production is being initiated. Generally, capsule production enlarges as iron decreases with concomitant slower growth.
The expression of the GATA iron regulatory factor, Cir1, is majorly responsible for iron uptake for growth at 37oC and the formation of virulence factors in the host tissue [40]. This expression is enhanced by the Hap genes such as HapX. Cryptococcus ferroxidase (Cfo1) expression enables this fungus to survive LIM; however, Δcfo1 mutant or as a background mutant to the deletion of Hap3, Hap5, or HapX cannot survive in the LIM unless feroxamine is supplied. Apart from the Δcfo1 mutant that can survive in the media supplied with FeCl3 and hemin, none of the doubly mutated iron gene mutants can survive in these two iron sources [40].
It appears that C. neoformans employed multiple alternative routes of acquiring iron from its environment for growth and proliferation. Though Δcfo1 mutant may have attenuated virulence notwithstanding deletion of HapX or Hap3 gene failed to extenuate further the virulence of this mutant [40]. Comparatively, the deletion of HapX seemed to have a greater influence on various transcriptional genes than deleting Hap3 or Cir1 under iron-limiting or iron-repleting conditions [40].
Paradoxically, though HapX seems to influence a greater percentage of other transcription factors under iron-limiting conditions, including siderophore transporters, yet deletion of Cir1 had a greater influence on the high-affinity iron transporters; however, this function is overlapping to some extent under low iron conditions. HapX and Hap3 cooperatively repressed most iron-dependent mitochondrial functional pathways under low iron conditions, but this overlapping function is not found in the iron-repleting conditions. While HapX majorly controls the subset genes for siderophore transporters in different iron sources, Hap3 mainly inhibits other iron-dependent regulatory pathways, especially in the mitochondria [40].
Though Cir1 and HapX positively regulate Sit1 expression for siderophore transporter, only Cir1 appeared to regulate virulence genes such as Lac1. Also, a report has shown that Cir1 positively affected the expression of Rim101 pH-responsive transcription factors, which further regulates the expression of various iron acquisition genes such as Sit1 and Cft1 [40]. Under low iron content, HapX influences the expression of Cir1 and Rim101, but under iron-repleting conditions, HapX apparently loses its influence while Cir1 influences the expression of rim101 but withdraws its influence on HapX to minimise redundancy of HapX perhaps. In a related study, Grx4, a 2Fe-2S cluster sensing and trafficking protein, was identified as an essential regulator of iron homeostasis capable of associating with Cir1p in the nucleus during iron depletion to induce multiple iron acquisition proteins [41,42]. This synergistic association may probably be under the influence of HapX during iron depletion. Reports showed that iron repletion promotes relocalisation of the Grx4p into the cytoplasm; besides, deletion of the Grx4 gene alone is sufficient to dysregulate iron-dependent metabolisms leading to oxidative stress, mitochondria dysfunction, and impairment of DNA repair [42].
The ability to utilise different sources of iron is peculiar to C. neoformans and other invasive pathogenic fungi like C. albicans. Iron is usually acquired in a reduced form from inorganic (hemin, feroxamine B, and FeCl3) or organic/biological form (transferrin and heme), and the effective absorption is centred on the interplay of Hap, Cfo1, and Cft1 genes. Acquisition of iron involves the reduction of Fe3+ by the cell surface reductases (Fre) to Fe2+ and then oxidised by the ferroxidase (Cfo1), which is coupled to Cryptococcus high-affinity iron permeases/transporters (Cft1 and Cft2). Besides, C. neoformans can acquire the non-reduced Fe3+ through the ferric-bound siderophores transporter (Sit1) presented by other closely associated microorganisms. This attribute is also common in other fungi, such as C. albicans and S. cerevisiae [43].
There are two putative orthologues of ferroxidase, Cfo1 and Cfo2, with the former being actively involved in the acquisition of iron while the latter plays a minor role. During infection, the austere iron withheld by the host tissues enables the fungus to express Cft1, which is trafficked to the plasma membrane under the influence of Pka1 to utilise transferrin for growth and virulence expression. The Δcfo1 and Δcfo1Δcfo2 mutants, irrespective of the temperature, are characterised by growth defect, attenuated virulence, copper toxicity, increased susceptibility to FCZ and AmpB due to impaired heme formation as a cofactor needed for ergosterol biosynthesis [44]. However, an exogenous supply of heme and feroxamine (siderophore) but not FeCl3 can restore some of these phenotypic deficiencies meaning that Δcfo1 mutant can still utilise iron through the low-affinity uptake proteins such as Cfo2 and perhaps Cft2. Iron assimilation mediated by the Cfo1 expression is independent of the Sit1 (feroxamine) and heme; nevertheless, when supplied at 100 μg/mL, these two iron sources can restore growth and relative resistance against FCZ in Δcfo1 mutants [44].
The Cft1-Cfo1 expression is needed to utilise iron from hemin and FeCl3 but not from heme or feroxamine, and any mutant with single/double deletion of these genes may not survive in the low-iron environment [44,45]. Thus, it seems the Cft1 and Cfo1 may be clustered on the same locus as observed in serotype D, and their transcripts are synergistically elevated with other iron regulatory genes in a low-iron condition [46]. Apart from these tandem iron-uptake-clustered genes, Lian et al. further suspected, in the LIM, other yeast orthologue genes involved in the intracellular iron transport and storage, and this includes:
- ❖
- a low copy Yfh1 transcript (encoding frataxin that mediates mitochondrial iron efflux cytosolic unbound iron),
- ❖
- Utr1p (augmenting ferric reductase activity),
- ❖
- Atm1p/ABC inner mitochondria iron transporter,
- ❖
- Mrs3p and Mrs4p (inner membrane protein suppressing mitochondrial RNA (mtRNA) splicing defect in the yeast, and also function as inner mitochondrial iron transporter),
- ❖
- fet5 (encoding Fet3-related integral membrane multicopper oxidase required for high-affinity iron uptake across the vacuolar membrane in conjunction with Cft1p to mobilise iron from the storage), and
- ❖
The evidence that Δhap3Δcfo1 and ΔhapXΔcfo1 failed to utilise hemin and yet are relatively virulence in mice experiment means that other additional iron acquisition transporters, apart from the Cfo1, may be orchestrating iron acquisition during infection [40].
The Cft1 and Cft2 are the two orthologues of high-affinity iron permease similar to the Ftr1 of S. cerevisiae, and their expression is influenced by Pka. The Cft1 expression is more effective and highly involved in the growth and iron uptake from FeCl3 and holo-transferrin than Cft2; however, the deletion of the two genes (Δcft1Δcft2) attenuated the virulence accompanied by low tissue burden (for details on the phenotypic display of different cryptococcal cell mutants, check Supplementary 1). In the presence of ≤1.0 mM FeCl3, the Δpka1 and Δpka2 mutants significantly failed to raise the expression level of cft1 compared to the wt, and even the Δpkr1 mutant further displayed reduced expression of cft1 by 2 – 3-fold. More significantly, the Δpka1 mutant displayed a more increased expression of cft2 compared to the Δpka2 mutant. In contrast, the Δpkr1 mutant failed to show any significant effect on the expression of cft2 when compared to the wt [45]. Though no significant contribution has been traced to Cft2 high-affinity iron uptake, the upregulation of this gene by Pka, however, might suggest the importance of this seemingly insignificant iron transporter. The reduced growth observed by Lian et al. with Δcft1 mutant in the low- and high-iron media [46] is redolent that Cft2 and perhaps Cfo expressions may be rescuing the mutant. This is like an SOS message received by the low-affinity iron uptake proteins to rescue the Δcft1 mutant.
In the absence of ferrous iron, C. neoformans may be compelled to use a non-reductive ferric-bound siderophore complex in vivo or in vitro as the iron source. Several pathogenic fungi have evolved to possess various specific siderophore transporters (Sitp) through which ferric-bound siderophores can be utilised by their competitors. These competitors may be other microorganisms capable of producing siderophore as deferoxamine, which can subtly steal iron from transferrin or heme and present it as siderophore feroxamine bearing Fe3+. A putative sit1 transcript identified in C. neoformans encodes a siderophore transporter, which is highly expressed in cells cultured in the LIM alone or with a ferrous iron chelator, bathophenanthrolinedisulfonic acid (BPDA), supplemented with deferoxamine. Experimental observation from Tangen et al. showed that Δsit1 mutants (either serotype A or D) could utilise ferrichrome but not feroxamine B as an iron source under iron-limiting conditions [53].
Furthermore, deletion of the Sit1 gene in the serotype D background enhances melanisation better than the serotype A Δsit1 mutant. The melanin production seemed better with increasing DOPA concentration in the medium. However, 1.0% glucose repressed melanin formation in the two serotype mutants. Additionally, the Sit1 expression appears to control copper availability to Cu2+-binding pocket of laccase. So, the loss of Sit1 gene apparently attenuates ionic homeostasis and reduces laccase activity more significantly in the wt than the mutants. This means that laccase in the mutants has more access to copper because of the absence of Sitp, which ordinarily could have distributed the copper among other copper-dependent proteins. In this case, Tangen et al. exogenously supplied copper and discovered an elevated laccase activity in the wt and reconstituted Δsit1 strain, which is comparable to the Δsit1 mutant [53]. Notwithstanding, an exogenous supply of copper or iron in the absence of glucose generally seems to favour melanisation in serotype D ΔSit1 mutant.
In serotype A, the lack of the Sit1 gene showed no significant effect on the Lac1 transcript, even if copper or iron was supplied in a non-glucose medium. However, in serotype D, this mutant showed a significantly high level of Lac1 transcript than the wt and reconstituted mutant – only when copper or iron is exogenously supplied in the absence of glucose [53]. With Calcofluor white, SDS, and Congo red in low-glucose or L-DOPA medium, serotype D Δsit1 mutant only showed a slight growth sensitivity to these compounds compared to the wt, but no significant sensitivity was observed in the serotype A Δsit1 mutant. Furthermore, serotype D Δsit1 mutant showed increasing growth defect from 30 – 37oC, but such temperature sensitivity is not conspicuous in the serotype A Δsit1 mutant [53]. This serotype differential phenotypic expression in response to copper and iron from Sit gene re-emphasises the existence of cellular morphotypes in C. neoformans.
To further emphasise the importance of Sit1 expression in cell wall integrity, Δsit1 mutants generally displayed a less dense and heterogenous cell wall thickness and polysaccharide fibril distribution. In addition, the serotype D Δsit1 mutant cell wall is easily distorted compared to the serotype A Δsit1 mutant, which reaffirms the phenotypical disadvantages earlier described in the serotype D Δsit1 mutant. Therefore, it is not surprising to observe that serotype D Δpka1 mutant could have a preponderant level of Sit1 transcript and that the serotype A Δpka1 and Δpka2 mutants equally showed an elevated level of sit1 transcript [53]. This observation is similar to the negative regulation of iron uptake by the cAMP-Pka event in S. cerevisiae [54]. To this effect, the Δsit1 mutant failed to improve defective capsule formation or induction in the Δpka mutant cultured in the low-iron or iron-supplemented medium. Also, the deletion of Sit1 did not initiate unilateral mating defect in serotype A MATα Δsit1 x MATa or serotype D MATα Δsit1 x MATa [53].
Expectedly, the hypermelanised serotype D Δsit1 mutant would have shown higher virulence than the wt or reconstituted mutant; however, Tangen et al. could not observe any significant difference in the virulence among these strains. Not only this, but the virulence from this serotype mutant is generally weak [53]. Still, serotype A Δsit1 mutant, alongside the wt and the reconstituted mutant, actually induced infection almost equally with no significant difference in the survival rate of experimental animals after the infection, tissue colonisation/burden within the brain and lungs, and extrapulmonary dissemination [53]. Thus, this outcome generally presents the Sit1 gene as not particularly required for virulence and infection in an animal model.
While Cir1 positively influences the expression of Cft1, Cft2 is negatively regulated. The transcription level of Cft1 in Δcft2 mutant decreases just like in the wt as FeCl3 concentration increases. However, the Cft2 transcripts level in Δcft1 mutant increases with a threshold at 10 mM FeCl3 but reduces in the wt as the iron level increases. The Sit1 transcript, on the other hand, is generally reduced in the wt but becomes better responsive to FeCl3 level ≤100 μM in Δcft1 than Δcft2; however, concentration beyond this makes Sit1 entirely unresponsive and drastically reduced in the Δcft1 [45]. This means that the loss of Cft1 potentially reduces the intracellular iron content, and this condition makes Sit1 transcription unresponsive even in the presence of 1.0 mM of FeCl3. This, therefore, means that Cft1 expression may come to rescue Δcfo1 background mutant during infection. Not only this, but evidence also showed that C. neoformans could secrete reductant metabolites such as α-ketoglutarate, 3-hydroxyanthranilate, and melanin to reduce Fe3+ to Fe2+ nonenzymatically for extracellular absorption [55]. This alternative reduction method might be responsible for late organ invasion observed with Δcft1 mutant in experimental animals [45]. The possibility also exists that though Δcft1 and Δcft1Δcft2 mutants cannot easily utilise transferrin in vitro, heme might be unavoidably used to supply iron in the late systemic infection. Nevertheless, the brain fungal burden of these mutants is constantly lower compared to other systemic organ invasions, perhaps due to transferrin availability in the brain parenchyma.
Evidence showed that the pH of cryptococcomas (mass lesions within the brain parenchyma caused by the dissemination of cryptococcal cells) is between 5.4 – 5.6, and this favoured the accumulation of pro- and anti-inflammatory metabolites such as polyols, acetoin, dihydroxyacetone, and GABA [56]. Accumulation of superoxide in this neutrophil micro-acidic milieu may have subtly deceived the host tissue into deploying iron into this environment. This condition favours iron acquisition through low-affinity/ferrous iron-specific systems in the C. neoformans. Convincingly, the growth of Δhap3Δcfo1, Δhap5Δcfo1, and ΔhapXΔcfo1 mutants are better rescued at pH 5.0 than 7.0 in media supplemented with FeCl3 or hemin. In addition, all the Δhap and Δcfo1 mutants failed to survive low iron irrespective of the pH, but all can utilise feroxamine at pH 5.0 and 7.0. The better growth of Δcfo1 mutants in the presence of FeCl3 and hemin, but the poorer growth of Δhap mutants, showed that Hap proteins function better to utilise different iron sources in a mutant lacking Cfo1 for high-affinity and reductive iron uptake system [40].
There are many reports on the connection of iron level with major virulence factors and cell surface-associated proteins in C. neoformans. With SAGE, iron-enriched media elevate various transcription factors connected to iron storage, glycolysis, mitochondria oxidative function, lipid and amino acid metabolisms, and calmodulin-calcineurin signalling events. However, other important transcription factors are well elevated in the LIM. Lian et al. reported an increased level of Cap60 gene, mannoprotein MP88, and GPI transamidase (which mediates cross-linking of cell wall mannoprotein to β-1,6-glucan) under iron-limiting conditions [46]. The transcript level of Cig1 encoding an extracellular glycoprotein (like mannoprotein) needed for capsule attachment to the cell, together with α-1,3-glucan (synthesised by α-1,3-glucan synthase), is usually found upregulated under the iron-limiting condition as well [46,57]. Though this gene responded to low-iron conditions where Δcig1 mutant is, the mutant is characterised by poor growth in a low-iron medium and failed to suppress capsule formation under an iron-supplemented medium. This means that Cig1p may be involved in the iron-sensing relay process or probably participates in maintaining membrane iron retention through the siderophore transporter. Still, its role in capsule formation is dispensable.
4.4. Copper Depletion
Copper is another micro-essential element needed for growth and virulence expression in C. neoformans. Copper acquisition and uptake are regulated by Ctr1 and Ctr4 (encoding copper transporters), copper transporter orthologue ccc2, atx1 (encoding copper chaperone), and copper-responsive transcription factor (encoded by cuf1). Mutation in these genes impaired melanin formation because copper is highly needed for laccase and also impaired growth under iron-limiting media [58]. This means an interplay of homeostatic regulation between the copper uptake and iron acquisition and regulation in C. neoformans during pathogenesis. The Δcuf1 mutants are characterised by impaired expression of Lac1, reduced dissemination, and tissue invasion coupled with growth defect in copper-limiting conditions because Ctr4 is poorly upregulated [59]. Thus, C. neoformans invasion and dissemination within CNS seems to be centred on the requirement for copper and iron because the same poor tissue burden was reported for both Δcft1 and Δcuf1 mutants. Additionally, Δcir1 mutant displayed positive upregulation of Ctr4 transcript in LIM or negatively downregulated in high-iron media [60].
One anti-cryptococcal strategy of the host cells is to channel excessive copper into the cytoplasm to initiate copper toxicity that can kill cryptococcal cells. However, with the expression of the Cmt gene, excessive copper influx can be detoxified by this obligate aerobic fungus (for details on the phenotypic display of different cryptococcal cell mutants, check Supplementary 1). Cryptococcal infection is characterised by increased serum Cu2+ due to the efflux of excessive Cu2+ from cryptococcal cells and presumptive excessive Cu2+ initially channelled to toxify the fungus. Evidence showed an increase in the expression of alveolar macrophage Ctr1 and a decreased expression of phagosomal Cu2+ transporter Atp7A during C. neoformans pulmonary infections [61]. Strategically, C. neoformans infections seem to promote nuclear factor-κB (NF-κB) that failed to coordinate macrophage growth, apoptosis, and inflammation. The preponderant level of NF-κB putatively binds the Ctr1 promoter to maintain the expression of macrophage Ctr1 while reducing the expression of phagosomal Atp7A ATPase to enhance fungal survival within the Cu2+ homeostatic phagocytes [61]. This, together with acidic phagocytic pH, probably keeps the infectious cryptococcal cell safe within the phagocyte until the condition is perturbed for non-lytic exocytosis, known as vomocytosis (phagosome extrusion/expulsion) [62].
Under Cu2+ limitation, Cuf1p acts as a transcriptional regulatory factor that enhances the expression of Ctr1 and Ctr4 as high-affinity Cu2+ transporters. Again, in the Cu2+-enrich medium, Cuf1p instead stimulates the expression of Cmt1 and Cmt2 against Cu2+ detoxification – a situation usually encountered during infection [63]. Though individual deletion of Cmt1 or Cmt2 appeared not to affect the fungal survival in the lungs, double deletion of these redundant genes, however, yielded mutants with severely attenuated virulence despite insignificant differences in the observable virulence expressions and phagocytosis rate when compared to the wt [61]. Also, the expression of Cmt1 and Cmt2 is very specific and can only be induced in response to Cu2+ but not to any other ions. Compared to the wt, the Δcmt1Δcmt2 mutant showed no growth defect in the presence of Fe2+, bathophenanthroline disulfonate (BPS – Fe2+ chelator), Cd2+, and menadione (MND), but the mutant was highly hypersensitive to 1 mM Cu2+ in the complete synthetic medium due to Cu2+ toxicity [61].
4.5. Magnesium Homeostasis
Three Mg2+/H+ antiporter encoding genes have been identified in C. neoformans (Mgt1 – 3). Apart from Mgt2, none of these genes is induced by exogenous Mg2+. Except for Mgt2, which seemed very important for intracellular Mg2+ homeostasis and proved difficult to be knocked out, Mgt1 and Mgt3 could not be associated with any phenotypical defect in this fungus [64]. Contrarily, different Mg2+ transporters have been identified in the yeasts, all of which are important to growth, mitochondrial functions, and spindle fibre formation during mitotic formation. Some of these transporters include plasma membrane Mg2+ transporters encoded by Alr1 and Alr2 [65], vacuolar Mg2+ transporter encoded by Mnr2 [66], and mitochondrial-associated Mg2+ importers encoded by lpe10 and Mrs2 [65] and exporter encoded by Mme1 [67].
The Mgt2 is a vacuolar membrane transporter and a homologue of Mnr2 of S. cerevisiae that influences the regulation of several other transcription factors, a few of which appeared to be Mg2+-independent. Suo et al. observed that Δmgt1, Pgal7-mgt2, Δmgt3, and Δmgt1Δmgt3 mutants showed normal growth in YPGal. However, only the Pgal7-mgt2 mutant exhibited a severe growth defect in YPD due to the significant downregulation of Mgt2 in the presence of a catabolite repressor (for details on the phenotypic responses of different mutants of Cryptococcus against various quantitative external factors, check Supplementary 3). This growth defect could be rescued precisely by exogenous Mg2+ supplementation (≥50 mmol/L) but not any other metallic salts. Among other downregulated genes in the transcriptome analysis of Pgal7-mgt2 mutant cultivated in YPD alone or supplied with 50 mmol/L MgSO4 include Aox1, Adh1, Tpo1, Fdh1, Hsp, Pdc1, Pcl12, Hxs1, Mup1, Mup3, Hxt1, Gal2, Aat, Pkp1, and Gal1 but Mf1α, Agn1, and many hypothetical proteins are upregulated mostly in the YPD alone [64]. The Pgal7-mgt2 mutant is completely albino in L-DOPA without Mg2+, but melanin is produced in the Δmgt1Δmgt3. Furthermore, adding Mg2+ restored melanin in the Pgal7-mgt2 mutant; however, this pigmentation appeared less when compared to Δmgt1Δmgt3 mutants [64]. On the other hand, capsule formation is highly induced in Pgal7-mgt2 mutant only in the absence of Mg2+, and this subsequently increased fungal proliferation and burden in the lungs of the infected animal model but with a wt tissue dissemination. Also, phagocytic inhibition was observed in murine macrophages infected with the Pgal7-mgt2 mutant [64].
4.6. Oxygen Depletion(Hypoxia)
C. neoformans is an encapsulated and environmental obligate aerobe (growing optimally under atmospheric oxygen at about 21% of the total air); therefore, adaptation to the relatively lower oxygen level of 1 – 5% in the human brain and lungs is inevitable to the survival of this fungus to cause infection. Oxygen availability is paramount to all the oxidative pathways, including mitochondria metabolism in this fungus and many other obligate aerobic basidiomycetes. These reactions are iron- and copper-dependent for electron transport leading to energy production. According to whole-genome microarray-based transcriptional profiling of C. neoformans under hypoxic conditions, about 347 transcripts are differentially regulated [68] and more than 100 expressed genes are statistically higher in the wt than the Δsre1 mutant under 1% O2 level in 3 hours. However, more than 414 genes are significantly expressed in the Δsre1 mutant than the wt under the same condition [69].
The major upregulated genes are involved in stress, iron and copper homeostasis, and the metabolism of sterol, heme, sphingolipid, sugar, and amino acids. In contrast, the downregulated genes are significantly involved in vesicular trafficking, cell wall formation, capsule synthesis, and ribosomal metabolism. Surprisingly, stress and antioxidant-related genes are highly upregulated in Δsre1 mutant via the cAMP/Pka or Hog1 pathway than the wt under the hypoxic condition. These genes include Tco2, Cat1, Cat3, Cas34, Hsp12, Hsp31, Gst1, Ecm4, and other genes involved in reductive pathways such as Gre2, Pkp1, and Ena1, some of which are evolutionarily conserved in the yeast.
The expression of Gst1, Gst2, and Gst3 encodes glutathione-S-transferase to protect the cells against reactive oxidants. Also, upregulation of Cat3 (encoding catalase A) in the Δsre1 mutant is highly significant in the hypoxic condition to degrade the H2O2 to water, and the speculation for this is that Δsre1 mutant may constantly be under more stress than the wt whenever the oxygen is limited [69]. In addition, Hem13 expression appeared unchanged under hypoxic conditions either in the wt or Δsre1 mutant; however, because of the strong connection between the Sre1p, ergosterol synthesis, and ionic homeostasis, the Sit1, Ctr4, Fre7, Erg12, and Erg25 transcripts are usually drastically reduced in Δsre1 mutants.
C. neoformans expresses the Sre1 gene to sense oxygen and regulate sterol synthesis. This expression is a combination of endoplasmic reticulum membrane-bound Sre1p (homologue of mammalian sterol-response element-binding protein, SREBP) and its associated sterol- and oxygen-sensor activator (Scp1p orthologue of SCAP) with other possible accessory proteolytic enzymes such as a metalloprotein site-2-protease (Stp) and Kex2-like protease. Hypoxia (<3% O2 level) or specifically low sterol level induces the proteolytic activation of Sre1p by Scp1p in the Golgi apparatus lumen and subsequently processed by Stp and Kex2 proteases to release a ≈75 kDa N-terminal transcription factor [69]. Any antifungal that blocks ergosterol biosynthesis will induce Sre1p activation; however, extremely low oxygen levels (0.2% O2 level) will drastically slow down C. neoformans growth due to the slow activation of Sre1p [69]. The nuclear-translocation of this basic helix-loop-helix (bHLH) leucine zipper DNA-binding domain of N-terminal activated Sre1p induces the plethora of transcription factors that enable the fungus to cope with oxygen-limiting conditions alongside sterol, iron, and copper depletions [69,70,71,72].
The interdependence of oxygen, copper, and iron to facilitate the environmental survival of C. neoformans and cause infection in the human hypoxic tissues cannot be overemphasised. Heme is a cofactor for sterol production in fungi. The production of heme is iron-dependent, which is induced by hypoxia. Hence, it is unsurprising that Δsre1 mutant, which failed to sense oxygen, is characterised by impaired ergosterol synthesis, iron, and copper uptake [69]. Under normal conditions, Δsre1 mutants tend to accumulate more sterol intermediates while the ergosterol level is reduced to basal need. These sterol intermediates become higher than in the wt in hypoxic conditions as the ergosterol level reduces further because of the absence of Sre1 expression. Thus, downstream accumulation of oxygen-dependent sterol intermediates under hypoxic conditions is necessitated in the Δsre1 mutant except for some upstream oxygen-independent intermediates such as 24-methylene lanosterol, which seemed to be unperturbed in the mutant and wt [69].
Apart from the Sre1 gene, Chun et al. established a hypoxia-sensitive trait in Δtco1 mutant, a surprisingly hypermelanised avirulent strain with normal capsule production [73]. All mutants generated, such as Δsre1, Δscp1, Δstp1, Δtco1, and Δsre1Δtco1, displayed normal growth in YPD at 37oC with no in vitro significant defect in the capsule formation but are sensitive to hypoxia condition, and the highest sensitivity is found in the double knockout mutant [68]. Furthermore, these mutants displayed a slight defect in melanin formation (except the Δtco1 mutant with hyper-melanisation), defective to moderate tissue proliferation vis-à-vis dissemination, antifungal susceptibility (except Δtco1 mutant), and impaired ergosterol biosynthesis (except Δtco1 mutant) [68]. Enzymatically, the Δsre1 mutant showed an insignificant defect in the activities of laccase, urease, and phospholipase, and the infection by this mutant failed to progress to terminal infection in an animal study due to limited brain lesions caused by poor growth in the brain though the mutant disseminated equally as the wt [69]. This in vivo weak infection may also be corroborated by the relatively smaller capsule size of this mutant in vivo [69].
Strictly, brain regions are endowed with self-secure barriers and nutrient stinginess to invading pathogens. Also, the outer regions of the brain, leptomeninges and cortex area are generally white with lower blood flow and iron content compared to the grey matter. Although Beard et al. observed a higher concentration of iron in the white matter, probably because of the preponderant transferrin circulation [74] yet the availability of this ferrous iron to the invading pathogen might not be corroborated. Furthermore, oxygen supply will be extremely low and stringently controlled in these brain regions because of the lower blood capillary network. Definitely, this self-care brain region might present a first-line defence against invading pathogens, including C. neoformans.
The phenotypic defects in the Δsre1 mutants are similar to the observed defects in the Δcir1, Δcuf1 and Δcft1 mutants, which further confirmed the interrelationship of the molecular oxygen, iron, and copper to facilitate the virulence traits of C. neoformans [43]. In fact, the Sre1 transcript was downregulated in the Δcir1 mutant, and the Δsre1 mutant will not grow under iron-limiting conditions or in the presence of an iron chelator. However, adding ferritin to the iron-chelating media could restore the growth but not as much as the wt or reconstituted mutant [60,69].
Unsaturated acyl-CoA is a component of the membrane bilipid layer. The synthesis of this fatty acid around the endoplasmic reticulum is controlled by the availability of molecular oxygen. There is evidence of upregulation of the Ole1 transcription factor due to hypoxia in C. neoformans, but this regulation seemed to be Sre1-independent [68]. The Ole1 encodes the endoplasmic reticulum membrane-bound Δ9-fatty acid desaturase in the yeast. This oxygen-dependent enzyme, in the presence of NADH-Cyt b5 reductase, incorporates a double-bond into the saturated fatty acyl-CoA through the diiron-oxo catalytic centre of the enzyme. Invariably, this enzymatic reaction is orchestrated by the ubiquitin/proteasome-mediated proteolytic activation of Spt23p and Mga2p and subsequent translocation of these dual-activators into the nucleus for the induction of Ole1 transcript formation [75,76]. This activation is influenced by the low level of oxygen, low level of unsaturated fatty acids (or high level of saturated fatty acid), metal chelators, and carbon source. Explicitly, any event leading to iron depletion, such as transition metals like cobalt and nickel that compete with iron for heme, and metal chelators, which withhold iron from binding the heme, will reduce the oxygen-binding capacity of the heme. These events will induce the Ole1 expression even under normoxic conditions.
C. neoformans can withstand excessive oxygen by expressing two loci, Oxy1 and Oxy2, presumably linking hyperoxia-sensitivity to melanin production. The Δoxy1Δoxy2 mutants are avirulent albino colonies with defective melanin production [77] and greater sensitivity to oxygen than Δoxy1 mutant [78]. Emery et al. observed a lack of correlation between the oxy phenotype and melanin production, and it was concluded that the degree of melanin production is a defective predictor of hyperoxia resistance [78]. The reason is that oxy2 defective loci in the Oxy1 genetic background were characterised by defective melanin production and hyperoxia sensitivity, which means that the Oxy2 locus is more important in conferring hyperoxia resistance. Though the antioxidant effect of melanin in C. neoformans might not be completely ruled out but Emery et al. predicted that the mechanism of defence against exogenous/endogenous neutral/reactive oxidant associated with hyperoxia stress in fungal aerobiosis solely relies on the interplay of Oxy1 and Oxy2 loci in the genetic make of C. neoformans [78].
Besides, Δoxy2 mutants are defective in ferric/cupric reductase and high-affinity copper uptake. Even after reconstitution of the mutant to restore melanin production, most of the mutants retained hyperoxia sensitivity, interpreted as non-allelic, non-interfered, and unlinked loci between the Mel (for melanin locus) and Oxy loci (for hyperoxia locus). Nonetheless, the importance of melanin to withstand reactive oxidants cannot be overestimated. The interconnection between the Oxy and Mel loci seems elusive. Melanin and polymeric dopamine are effective cellular anionic antioxidants produced by C. neoformans against hypochlorite (produced by myeloperoxidase as found in inflammatory cells such as neutrophils) [79], superoxide (O2‾) and other reactive oxidants. However, activity against neutral oxidants such as H2O2 is weak, especially in the presence of copper and oxygen [80], which by Fenton reaction generate reactive OH‾ (hydroxyl ion), OH• (hydroxyl radical), and HOO• (hydroperoxyl radical) nucleophile that predisposes phagocytosed C. neoformans to oxidative arrest. Sometimes, perhaps, neutral oxidants can cross the cell wall and bypass the effect of melanin, which is why C. neoformans are highly susceptible to peroxides such as H2O2, t-BOOH, and COOH (for details on the phenotypic responses of different mutants of Cryptococcus against various quantitative external factors, check Supplementary 3). Whatever the case, Oxy1 and Oxy2 expression may be needed to resist hyperoxia-based melanin oxidation imposed by the endogenous oxidant from the leucocytes and macrophages.
4.7. Carbon dioxide (CO2)
Via a bicarbonate (HCO3‾) production, C. neoformans employs CO2 signalling sensors such as calmodulin (Cam1) and carbonic anhydrase (Can) to detect the CO2-inducing cascade events leading to virulence factor formation, enhancing cell wall integrity, mating, resistance against excessive O2, CO2, pH, Ca2+, osmotic and oxidative stresses, toxigenic compounds, heavy metal toxicity, and antifungal drugs through direct activation of Cam1 or Can2-Cac1-cAMP/Pka cascade event. The Can gene encodes the families of four classes of carbonic anhydrase (α, β, γ, and δ), which are distributed differently in all living organisms [81], and the primary fungi transcript is Can2 encoding a zinc metalloenzyme β-carbonic anhydrase that maintains equilibrium between CO2/H2O and HCO3‾/H+. Surprisingly, though Can1 and Can2 are found and expressed in C. neoformans, only Can2 appeared to be the major transcript for β-carbonic anhydrase capable of expressing more of the enzyme at high CO2 (5%) than low CO2 (0.036%) [82].
The Δcan2 and Δcan1Δcan2 mutants of serotypes A and D displayed growth defects in a CO2-dependent manner. At the same time, the overexpression of Can2 impaired membrane integrity and made the cell susceptible to oxidative and genotoxic stress and antifungal drugs [82,83]. The mutant further displayed impaired decarboxylation reaction but exhibited an increased salvage pathway to produce pyrimidine via the upregulation of Fur1 expression. The lack of several Can2-dependent genes, including Fas1 (in lipogenesis), Meu1 (in methionine salvage pathway), and Gpb1 (in pheromone-response MAPK pathway for sexual differentiation) in this mutant warranted insufficient fatty acid production, impaired amino acid salvage pathway, and defective pheromone production for cell differentiation and filamentation, which are all responsible for defective growth of Δcan2 mutant incubated in ambient air with about 0.036% CO2. However, palmitate, among other fatty acids, appeared slightly effective at 2 mM to rescue some of these growth defects in the Δcan2 mutant [82]. This means that the ambient environment promotes the expression of the Can2 gene for growth and survival, but this enzyme is dispensable in a CO2-rich environment such as human tissue and can still proliferate with adequate virulence production.
Apart from Fas1, Meu1, and Gpb1, other Can2-dependent genes include Cfo2, Cas3, and Atf1. Most of the Can2-dependent genes are majorly ESR genes that will not respond to the overexpressed Can2 gene; this is very surprising. Kim et al. overexpressed the Can2 gene in C. neoformans mutant, which eventually displayed poor cell wall/membrane integrity coupled with hypersensitivity to oxidative, genotoxic, and antifungal (azole and polyene), probably because of the preponderant accumulation of HCO3‾/H+, which may be perceived as stress index to the cell [83].
Atf1p, which is involved in diverse stress responses together with Hog1p, is slightly upregulated when Can2 is repressed. Like Hog1p, Atf1p represses melanin and capsule formation to promote the expression of a group of genes involved in OSR, NSR, HTG, and OMSR (check Supplementary 4 for as many details as possible for the transcription factor, kinase, protein, enzyme, and gene acronyms used in this review). Again, Atf1p induces the expression of genes involved in the thioredoxin system [84]; perhaps this is synergistically enhanced by the Skn7 and Yap4 expression against oxidative and nitrosative stress, respectively (for details on the phenotypic responses of different mutants of Cryptococcus against various quantitative external factors, check Supplementary 3). However, deletion of Atf1 or Yap4 in a mutant has no significant effect on the macrophage survival and virulence [84], just as Can1 and Can2, with or without CO2, have no significant effect in modulating the C. neoformans virulence factors and response to osmotic shock, UV, temperature, oxidative and nitrosative stress [82]. Apparently, better survival was observed in the Δcan2 mutant than the wt in the CSF of the rabbit, and this mutant can only be recovered in the YPD supplied with a high level of CO2. Again, failure to observe any mating in the wt or Δcan2 mutant, in vivo, at a high CO2 level showed that other factors contributed to in vivo mating of C. neoformans [82].
High CO2 of about 5 – 6% in the human host induces capsule formation in cryptococcal cells, which is independent of Cas3 or Can2 expression but highly dependent on the Cac1 expression for CO2-mediated capsule production [83,85]. Surprisingly, due to its negative effect on cellular activity, C. neoformans cannot keep up with the continuous production of bicarbonate because overexpression of the Can2 gene under the Pctr4 aggravated sensitivity to several oxidative- and osmotic-inducing agents such as H2O2, CdSO4, MMS, SDS, AmpB, FCZ, KCZ, and ICZ. However, with a genotoxic-inducing agent such as hydroxycarbamide/hydroxyurea, only a wt resistance was displayed [83].
4.8. Sugar, Lipid, and Proteins (Amino Acids) Metabolisms
C. neoformans possess arrays of metabolic enzymes that help re-wire carbon metabolites to favour the sugar-conjugate products and metabolic intermediates such as acetyl-CoA used for capsule and melanin biosynthesis. In addition, the in vitro and in vivo analysis showed numerous membrane transporters specific for each nutrient within their environment. C. neoformans secretes hydrolytic enzymes such as peptidases, lipases, and glycosidases to release amino acids, fatty acids, and sugars, respectively. The plethora of enzymes involved in these metabolic activities are influenced by cAMP signalling pathways. The cAMP-deficient mutants are poor in sensing glucose (via Hxtp – a sugar/hexose transporter and G-protein coupled receptor, like Gpr1p) and amino acids (via the Gpr4p) because these metabolites provide precursors for growth, mating, cell differentiation, and virulence factors which are constantly under the regulation of cAMP-dependent signalling genes [86,87]. The Δgpr4 mutants are very poor in sensing certain amino acids, such as Met, which triggers receptor internalisation of Gpr4p and subsequent activation of the cAMP/Pka pathway [18]. However, their response to glucose remained unchanged, unlike the poor glucose response from Δhxt mutants. Despite a good response to glucose, the Δgpr4 mutants still displayed impaired capsule formation and mating defect, both of which are recoverable in the presence of exogenous cAMP [86].
In a DNA microarray analysis, nearly 20% of total gene expression is dedicated to the transport and metabolism of carbohydrates and amino acids metabolism [88]. The expression of Gpp2 encoding glycerol-3-phosphate phosphatase controls various metabolic systems as well as osmotic and cold shock responses. Deletion of this gene induced various permeases/transporters (Aapp, Mup1p, Mup3p, Ena1p, Nha1p, Plb1p, Hxt1p, Stl1p, Uga4p), significant facilitator transporter super-family genes, anti-oxidative enzymes (Cat2p, Ccp1p, and Gst1p), and membrane transporters (for sulphite, myo-inositol, and pantothenate). However, various genes involved in the redox process, the glycolytic pathway, and the TCA cycle are downregulated in this mutant [89]. More important is the induction of genes involved in sulphur-containing amino acid biosynthesis, such as Cys3, Sul1, Soa1, Bds1, Met17, and Jlp1 in this mutant. The expression of Gpp2 modulates the regulation of S-containing amino acids by facilitating the proteolytic degradation of Cys3p and subsequently reducing the sulphur assimilation [89,90].
Hitherto, the deletion of Aap4 and Aap5 allowed eugenol resistance in C. neoformans. However, deletion of Gpp2 alters the biosynthetic regulation of Met and Cys due to the perpetual activation of amino acid permease protein (Aapp), which probably explains the increased sensitivity of the Δgpp2 mutants to eugenol. This dire situation could not be rescued by exogenous proline; however, it is uncertain whether glycerol can suppress this defect or not [89]. Reasonably, Δhog1 mutants might as well be sensitive to eugenol because Aap is also induced in hog1 mutants [91]. Thus, osmotic stress response by glycerol in the yeast is essential, but Cys can feed the glutathione cycle to generate GSH for alternative osmotic stress response in a case where glycerol production is truncated in a mutant such as Δgpp2. This seemingly inversely proportional metabolic network was investigated further in the C. neoformans wt to be true because 1 M NaCl can downregulate Cys3 and Sul1 expression to some extent [89].
Furthermore, the activity of the Gpp2p is under the regulation of Ca2+ via the Cna1/Cnb1 cascade event. Just as in A. nidulans and N. crasssa, deletion of Cys3 in C. neoformans has been shown to reduce the expression of Sul1 and both expressions are grossly repressed in the presence of S-containing amino acids (e.g., Met or Cys) in SD (synthetic dextrose) media more than the YPD [90]. This shows that media with low S-containing amino acids will likely induce de novo sulphur amino acid synthesis. Again, Cys seems better as a sulphur source than Met because methionine could be formed from cysteine in a transulphurylation reaction [90]. Further investigation on Δcys3 mutants showed that the mutants displayed a wt level of urease activity and capsule size yet attenuated for virulence in Galleria mellonella. Bad still, the experimental condition with Cys or Met in the culture media failed to support the melanin and phospholipase assay of the fungus [90].
The proteomic analysis that showed a concurrent induction of Met3, Tef1, Gpd1, Ccp1, Rps0, Rps1, Tif1, Sub2, Tm8, Gpp2, Cna1, and Cnb1 has led to the proposal of a metabolic relationship between sulphur-containing amino acid biosynthesis, glycerol phosphatase (encoded by Gpp2), and calcineurin pathway (Cna1/Cnb1), which was further confirmed by protein-protein interactions [90]. Expression of Cna1/Cnb1 motivates and maintains Cys3p protein levels for nuclear localisation and subsequently increases the Cys3p-target genes such as Sul1, Met2, Met3, Met10, Str1, and Sam1 to promote sulphur, purine, Gly, Ser, Thr, Asp, Asn, Cys, and Met metabolism while Gpp2p orchestrates inactivation of Cys3p by proteolytic degradation. A proposed mechanism is that inorganic sulphur induces Cna1/Cnb1 activation to deactivate Gpp2p to promote Cys3p and activate downstream genes involved in S-containing amino acid biosynthesis. However, the presence of organic sulphur such as Met and Cys deactivates Cna1/Cnb1, and this activates Gpp2p to deactivate Cys3p and repress the downstream genes involved in S-containing amino acid biosynthesis [90].
Among the genes induced in Δcys3 mutant are putative Aro3 and Aro4 involved in aromatic amino acids biosynthesis; Jlp1 in sulfonate metabolism to provide SO42- for Met5/10p; sulphiredoxin in oxidative stress response; and Clr6 – encoding class I histone deacetylase (a transcriptional epigenetic repressor of Mat2 – Mat3 region and centromere for precise heterochromatin packaging during chromosomal segregation in fission yeast) [92], but the repression of Clr6 in the C. neoformans wt promotes capsule formation and biosynthesis of various amino acids. With Maier et al.,
- ❖
- Clr6, along with Hog1, Mbs1, Ste12α, and Tup1, is classified as a cluster of genes induced against reactive oxidants but repressor of respiratory activities and also a repressor of sugar, amino acid, and ionic transporters that are needed for growth,
- ❖
- the cluster of Ada2, Clr3, Clr4, Clr5, Ecm2201, Fkh101, Gat201, Hap3, and Rim101 induced ionic transporters but repressed chitin synthesis,
- ❖
- the cluster of Cac1, Cep3, Cir1, Clr2, Fap1, Fhl1, Fkh2, Gat1, Mcm1, Pdr802, Sp1, Swi6, and Usv101 are involved in the mitochondrial respiratory process and activation of proteins/enzymes involved in gluconeogenesis and ionic transporters for osmoregulation; however, cAMP signalling is repressed, and
- ❖
- the cluster of Clr1, Hap5, Nrg1, Pkr1, and Ssn801 enhanced ribosomal biogenesis, response to oxidative stress, amino acid biosynthesis, cAMP signalling but repressed respiratory activities, ionic transporters, gluconeogenesis, and osmoregulatory process [93].
Transcriptome analysis Δpka1 and Δpkr1 mutants have been characterised with higher-level transcripts of genes involved in glycolysis/carbohydrate metabolism, TCA, amino acid metabolism, iron uptake, cell wall glycan formation, translational ribosomal proteins, translational initiation and proteins elongation, heat shock, protein and vesicular trafficking, UPR, cytoskeletal proteins, membrane transporters/permeases, cell surface and extracellular transmembrane proteins, oxidative and nitrosative stress response proteins, mitochondria membrane proteins, virulence factors, and phospholipid metabolism [94].
In addition, there is a characteristic increase in transporters/permeases such as sugar transporter, hexose transporter, glucose transporter, and maltose permease (all for sugar assimilation); ammonium permease and amino acid transporter (to assimilate nitrogen and amino acids, respectively); peroxisomal fatty acid transporter and ATP carrier (to assimilate phospholipase products), and many other residential permeases and ion-affinity coupled transporters, which are used for importing nutrients and micro-needed ions from the extracellular digestion. Therefore, through the interplay of Pka1 and Pkr1 expressions, cAMP plays one of the major central roles in coupling environmental sensing to metabolism, virulence, cell differentiation, mating, and growth.
C. neoformans is sensitive to glucose limitation, and phagolysosome does this by constantly maintaining acidic pH to minimise glucose availability to pathogens. To circumvent this, C. neoformans embarked on gluconeogenesis to supply sugars for metabolisms leading to titanisation and capsule formation. The interdependence of peroxisome, glyoxysome and cytoplasm to break down assimilated fatty acids (β-oxidation) to generate acetyl-CoA is essential to drive the glyoxylate cycle to generate oxaloacetate used in gluconeogenesis. Contrary to C. albicans, which showed up-regulated genes for key enzymes of gluconeogenesis [95], phagocytosed C. neoformans showed no upregulation of such enzymes [16]. However, transcriptional response and analysis of C. neoformans ingested by macrophage and amoeba showed that key enzymes like phosphoenolpyruvate carboxykinase and fructose-1,6-bisphosphatase were respectively upregulated [13]. The Icl1 (encoding isocitrate lyase), Mls1 (encoding malate synthase in glyoxylate cycle), and genes for β-oxidation enzymes are upregulated in C. neoformans recovered from murine macrophage [16]; however, Mdh2 (encoding malate dehydrogenase) and genes for ergosterol biosynthesis were downregulated in the phagocytosed C. neoformans [13]. The findings of Rude et al. showed that though the Icl1p and the glyoxylate shunt are important for ATP production during fungal infection, still Δicl1 mutants failed to display any apparent virulent defect in animal studies and produced a wt level of capsule and melanin without any defect in macrophage survival or phagocytic index [96].
In all these reactions, reducing equivalents such as NADPH+ are highly required for redox processes. Hypervirulence-associated protein 1 (encoded by Hva1) has been identified to be associated with NADPH to regulate C. neoformans metabolisms [97]. Derengowski et al. have discovered a multiple-fold increase in the transcript level of Hva1 in amoeba-phagocytosed cryptococcal cells [13] and the absence of this Hva1p unexpectedly increased the level of phosphoenolpyruvate kinase but definitely impaired the TCA due to reduced activity of 2-ketoglutarate dehydrogenase complex activity. This condition probably attenuates mitochondria oxidative reaction but favours sudden energy increased from preponderant floating of cytoplasmic NADPH via alternative pathways to produce ATP. This metabolic energy production shift favours cell proliferation and growth, hence the hypervirulence trait of Δhva1 mutants in the murine model but not moths or worms [97]. This means the Δhva1 virulence is temperature dependent. The exogenous supply of NADPH+ further enhanced the higher tissue burden of this mutant in the spleen and brain than in the lungs [97]. Paradoxically, an extensive comparison of the virulence factor expression, capsule size and structure, melanin production, GXM content, phospholipase and urease activity, growth and doubling time under stressful conditions, survival and fungal burden in macrophage, animal model, moths, and worms revealed no significant difference between Δhva1 and Δhva1+Hva1 mutants [97]. This displayed the importance of regulated metabolic systems in C. neoformans virulence.
To further appreciate the involvement of metabolic systems and their regulatory enzymes in the virulence, cell wall integrity, and stress-resistance phenotypic traits in C. neoformans, the lack of Pgi1 expression (encoding phosphoglucose isomerase/glucose-6-phosphate isomerase), though showed no observable difference in the cell morphology but resulted in reduced capsule biosynthesis, impaired cell wall integrity, fragile cell membrane, osmotic stress hypersensitivity (due to impaired Hog1 pathway), and failure to utilise mannose and fructose [98]. Insertional mutation T-DNA at the promoter site of Pgi1 reduces the activity of Pgip, and in the presence of a 2% glucose supplement, this mutant tagged as LZM19 (Δpgi1) is able to produce melanin due to the derepression of Lac1. Further work showed that an exogenous supply of cAMP could restore capsule deficiency in the LZM19 strain, but this repressed the Lac1 expression [98].
The mammalian homologue of AMPK protein is Snf1p in the yeast, which is critical to cellular energy homeostasis through oxidative phosphorylation by facilitating sugar and fatty acid uptake. In a normal state, Snf1 is turned off by Reg1-Glc7 protein phosphatase 1 (PP1) [99]. However, high glucose concentration activates Cac1p to produce cAMP, which interacts to activate Pka to repress the H2C2 zinc-finger nuclear-translocated proteins Msn2p and Msn4p that are programmed to induce genes containing SRE-sequence promoters such as Hxt1, Hog1, Ras1 and others, which are generally regarded as NSR, OSR, OMSR, ESR, GSR, HMSR, and CSR. In addition, the cAMP/Pka also inhibits the redundant upstream Snf1p-activation kinases, Sak1, Tos3, and Elm1, to put Snf1 under repression in high glucose content.
High glucose deactivating Snf1 leads to the activation of Mig1p by dephosphorylation to enhance the association of Mig1p with Suc2p. The Mig1p is a universal gene repressor in the yeasts [100], which binds the promoters of target genes, including the Lac1 gene, to repress melanin biosynthesis as a response to glucose repression. In low glucose levels, however, the Cac1 activity is reduced to lower the cAMP level, which invariably elevates the Sak1 activity to activate Snf1p [101]. The activated Snf1p further inhibits the Cac1 to promote nuclear translocation of Msn2p to induce hxt7 and other similar stress response genes, including the Lac1 gene.
Canonically, despite glucose availability, mutation of Pgi1 expression may have limited glucose catabolism and subsequently altered the ATP homeostasis. This condition favours Snf1p activation leading to the deactivation of Mig1p and subsequently derepresses Lac1 for melanin production. Therefore, the involvement of the Snf1-Mig1 regulatory pathway through nutrient homeostasis cannot be ruled out in the pathogenesis and virulence of C. neoformans. Unprecedentedly, the involvement of Snf1 expression in antifungal, oxidative, and osmotic stress responses is less significant. Though Hu et al. claimed to have observed increasing sensitivity of Δsnf1 mutants to AmpB relative to the wt response; however, the Δsnf1 mutants apparently showed no significant growth defect to AmpB and FCZ, but a very slight growth defect in the presence of RPM [102].
Generally, phenotypic defects of Δsnf1 mutants become conspicuous with increasing temperature. For example, melanin formation decreases above 30oC and ultimately becomes a non-melanised mutant at 37oC [102]. Surprisingly, though the transcript level of Snf1 is practically increased in a glucose-limited or acetate medium, but there is no evidence of increased transcript in the cryptococcal cells recovered from the infected lungs. Besides, Δsnf1 mutants in a glucose-limited medium displayed an increased marginal level of Ady2, Acs1, Ato2, and Pck1 transcripts; a marginal decrease in the expression levels of Ctr4 and Cgl1; and a significant reduction in the Hxt1 and Cft1 transcripts [102]. This means the latter two transcription factors solely depend on the Snf1 expression, especially in the glucose-limited medium. In the acetate medium, however, deletion of Snf1 showed no effect on the expression of all these genes except the Hxt1, where an elevated transcript is observed. The additional report showed that Δsnf1 mutants displayed a wt Mls1 transcript irrespective of the media used [102]. The activation of Hxt1p in Δsnf1 mutants cultivated in the acetate medium showed the characteristic of glucose-deprived cryptococcal cells and readiness to assimilate sugars as soon as it is available.
In addition to Lac1 regulation, Snf1 expression also affects the SOD activity. A drastic reduction of Lac1 in Δsnf1 mutants cultivated in the glucose-limited medium indicates the Lac1 expression being regulated by Snf1p. Similarly, the Sod1 transcript is reduced in Δsnf1 mutants in a glucose-limited medium but not in the acetate medium. Irrespective of the medium, deletion of Snf1 failed to significantly affect the expression of Fhb1, Tps1, Glr1, and Skn7, but Lac2 and Ssa1 transcripts are significantly increased in acetate media [102]. This means that in the glucose-limiting medium, Snf1p orchestrates the expression of Lac1 to produce melanin and the expression of Sod1 against oxidative stress. Strategically, Lac2 expression may be preferable for melanin synthesis under alternative carbon sources coupled with oxidative and nitrosative stress, which might have been addressed by the concomitant expression of Tsa1 under the same nutrient-limiting medium; after all, Δlac1 mutants still cause infection in a murine model [102,103]. Unfortunately, Δlac1 and Δlac1Δlac2 mutants are totally albino. In contrast, Δlac2 mutants are melanised in catecholamine-enriched media [104]. However, the elevated Lac2 transcripts more than the Lac1 in the presence of alternative carbon sources such as acetate [102] is reminiscent that Lac2 may probably be involved in melanin production when unusual carbon sources weaken Lac1 involvement. Therefore, by inference, catecholamine (such as L-DOPA and norepinephrine) metabolism into eumelanin may seem more complex than already revealed, especially in the presence of alternative carbon sources.
The role of Acs1 expression encoding acetyl-CoA synthetase has also been investigated in the cryptococcal cells recovered from the lungs of infected animals. Evidence showed an elevated expression of the Acs1 gene in C. neoformans during infection, coupled with concurrent elevated expression of different membrane transporters and stress response genes, which is indicative of a nutrient-limited environment and the route for alternative carbon sources as facilitated by the expression of Snf1 [102]. Deleting Acs1 yielded moderately attenuated mutants that cannot utilise alternative carbon sources such as acetate.
Sometimes C. neoformans can utilise other carbon sources, including alcohol. Inexpediently, mutants lacking Hap3 and Hap5 showed weaker growth in sucrose, acetate, and ethanol and are more sensitive to 0.01% SDS. In contrast, the ΔhapX mutant and the wt showed similar growth in the glucose, sucrose, acetate, and ethanol, even in 0.01% SDS. Although Hap genes are iron-dependent regulatory genes, surprisingly, supplementing the media with hemin failed to improve the weak growth in hap mutants [40]. Because of this, iron-enrich media specifically improved gene expression encoding several iron-dependent enzymes in the glycolytic pathway, TCA cycle, functional oxidative pathways, and mitochondrial redox processes, but gene expression targeting the pentose phosphate pathway was scarcely tagged in the SAGE analysis [46]. Apart from these elevated key enzymes, other accessory proteins such as ubiquinone, prohibitin (a membrane-bound chaperone stabilising the mitochondria proteins), and ATPase are equally pre-eminent [46]. The expression of TCA-targeting enzymes such as aconitase and succinate dehydrogenase further confirmed the importance of the heme and Fe-S redox proteins to the upkeep of C. neoformans metabolic systems. In fact, aconitase identified in C. neoformans seems to have paralogous encoding genes to probably enhance its extracurricular activities such as mRNA translation/stability – a function known for Irp1p. It could probably be that C. neoformans regulates excessive iron by converting this Irp1p to cytosolic aconitase, which fails to stabilise mRNA but promotes Irp2 ubiquitination and proteasomal degradation [105].
Further, into the regulatory roles of Hap3p and HapXp, these two transcription factors negatively modulate the expression of the orthologue Lys4 in the LIM. This was confirmed by low detection of Lys4 in ΔhapX and Δhap3 mutants cultured in the presence of 100 μM FeCl3, but in the absence of FeCl3, the expression of Lys4 is well induced in either of these mutants [106]. Relative to the wt, the expression of Lys4 remained unchanged in Δcir1 mutants in either iron enrich media (IEM) or LIM. Like Lys4, Leu1 expression was also similarly regulated by Hap3p and HapXp, either in the LIM or IEM. Low iron supports the expression of Leu1 and Lys4 in Δhap3 and ΔhapX mutants, but only a high level of iron favours the expression of Leu1 in Δcir1 mutant [107]. This means that Cir1p positively regulates this cytoplasmic located Leu1p in Δhap3 and ΔhapX mutants cultured in LIM but regulates negatively in IEM. There is speculation, however, that Leu1 expression may be more under the regulation of Hap3p than HapXp expression because while ΔhapX mutant produced a comparable Leu1 transcript with the wt, Δhap3 mutant produced a significantly more transcript of Leu1 under the same IEM conditions [107]. Like Lys4 expression, Leu1, with a putative ―CCAAT―Hap binding element at the promoter site for repression during the iron-limiting condition, is also very important in Leu biosynthesis; however, the uptake of these amino acids is generally under the influence of NCR.
Being a eukaryote, C. neoformans compartmentalise and adopt differential regulation of transcription factors to facilitate catabolism (in the mitochondria, glyoxysome, and peroxisome) or anabolism (in the cytoplasm) depending on the inducing factor and the availability of specific nutrients. For example, IEM elevates enzymes (including fatty acid synthase and acetyl-CoA carboxylase) in the cytoplasmic lipogenesis and promotes extracellular digestion of lipids (lipolysis) which is usually commenced with the release of phospholipase B (encoded by Plb1). Analysis showed that C. neoformans possessed genes for lipase (encoded by Tgl2 among at least three encoding genes) and enoyl-CoA hydratase [46,108]. However, there is no specific evidence of the functional extracellular lipase expressed in this fungus due to the lack of a signal peptide in these proteins, unlike the lipase secreted by C. albicans [108,109].
The lack of trypsin, chymotrypsin, β-galactosidase, β-glucuronidase, and α-mannosidase by the species of this fungus showed that certain proteins and β-D-sugars might not be suitable in the culture media for C. neoformans [109]. However, many species of cryptococcal cells do possess esterase lipase (active against triacylglycerol with C4 – C8 acyl chain), α-galactosidase, α-glucosidase, β-glucosidase, N-acetyl-β-glucosaminidase, α-fucosidase, galactokinase, D-lactate dehydrogenase [46,109], which means that various α-D-sugars, amino sugars, and disaccharides are suitable carbon sources for cryptococcal cells. Furthermore, since IEM elevates sterol biosynthesis, it can be speculated that fatty acid/sterol-formulated media could motivate sterol and phospholipid biosynthesis because of the possibility of expressing lipolytic and sterol biosynthetic enzymes under the iron influence.
In addition, lipid biosynthesis is an essential biochemical pathway leading to cell membrane integrity. The Mga2 is an orthologue transcription activating factor for the component of fatty acid synthesis and regulates in vitro normal growth at a range of temperatures. Therefore, mutation of Mga2 makes C. neoformans thermosensitive and hypersensitive to FCZ with a concomitant reduction in the expression of Fas1, Acc1, Rpl11, Rps7, Leu1, Pho, Pdh, eEF3, and other orthologue proteins. However, expression of Lys2, Hxt, Cat, Pak1, Chs, Mdr1, ABC-family, Vps, Hsp78, Hsp104, calnexin, Rho-GAP domain protein, exo-1,3-β-glucanase, and other relevant proteins are induced in this mutant [110]. Ambiguously, it is unclear how the presence of Mga2 regulates polarisation and filamentation growth in C. neoformans from the work of Kraus et al. because Δmga2 mutants are characterised with elevated expression of the homologues of small GTPase effectors, which control the polarisation growth, and the Pak1, which controls the filamentation growth. Therefore, it is possible that during this morphogenesis, Mga2 expression is repressed at ≤30oC for GTPases and Pak1 expressions that will promote polarisation during growth, but this, however, will contradict the involvement of Mga2 in thermotolerance and morphogenesis as reported by Kraus et al. [110]. Besides, the dependency of fas1 and acc1 expressions on Mga1 for lipid and sterol biosynthesis will literally promote polarisation growth, mating, and haploid fruiting. Therefore, the background expression of Mga1 may have promoted polarisation and cell growth in Δmga2 mutant.
Still, under the influence of iron, secretory vesicles (such as exocyst complex encoded by Sec gene), enzymes (such as glutamate dehydrogenase, p-aminobutyrate aminotransferase, proteasomes, ubiquitin ligase, RNA-processing enzymes, and mRNA splicing proteins), and transporters/permeases (for allantoate, urea, ammonia, and purine-cytosine) involving in nitrogen and amino acid metabolism including protein trafficking have been observed to be elevated under the IEM [46]. Not only these, iron influences several of the amino acid metabolism (anabolism and catabolism). In S. cerevisiae, iron probably orchestrates the binding of α-isopropylmalate to homodimeric DNA-binding protein Leu3 (a repressor that, together with Gcn4, regulates amino acid biosynthesis) to regulate branched-chain amino acid biosynthesis, Aapp, nitrogen and carbohydrate metabolisms. The expression of Lys4p for Lys biosynthesis and Leu1p for Leu biosynthesis is iron-dependent because of the presence of conserved aconitase repeating motifs that bind Fe-S; and probably other amino acids biosynthesis proteins such as Met2p, Met3p, and Met6p for Met; Ilv2p for Ile and Val. Mutants generated from each of these transcription factors are avirulent in the MIMC.
Iron homeostasis in C. neoformans consists of arrays of gene clusters harmonising the acquisition and utilisation of iron, and this could be similar to the iron regulon of S. cerevisiae under the influence of Aft1 and Aft2 transcription factors. With NCR, certain nitrogen sources are not assimilated, perhaps for the reason of energy-costly catabolism. Deletion of Gat1, among other GATA transcription factors, promotes poor to no growth of C. neoformans in the majority of different nitrogen sources except in the presence of Pro and, to some extent, Arg, while single deletion of other GATA family genes (Δgat201, Δgat204, Δbwc2 or Δcir1) failed to shut down totally the fungal growth in all the popular nitrogen sources [111]. This means that various amino acid permeases may still be activated to assimilate other nitrogen sources under later conditions, contrary to Δgat1 mutants where the Amt and majority of the Aap gene expressions are impaired. However, Pro and Arg may probably not be under NCR control.
To affirm this, the Pro medium induces Put1 gene expression for proline oxidase in the wt under the influence of NCR as mediated by Gat1 expression in the presence of NH4+. In addition, the expression of Put5 (a paralogue of Put1) and Put2 genes, which are involved in the downstream catabolism of Pro to Glu, remained unchanged in Δgat1 mutants and wt [111]. This means that though Gat1 mediates NCR, Pro metabolism is Gat1-independent. The ability to form Glu from Arg may also explain the moderate growth of Δgat1 mutants in this medium. Gln and His can also form Glu directly. However, because the cytoplasmic content of Gln is always high as a form of nitrogen donor in most metabolic reactions and the metabolism of His is also more complex, this may explain why Δgat1 mutants had poor growth in a medium with each of these two amino acids as nitrogen sources.
The growth defect of Δgat1 strain H99 mutant in urea, urate, or creatinine-containing media with reduced expression of genes involved in nitrogen metabolism (such as Gdh1, Amt1, and Amt2) is a serendipitous observation that contradicts the ecological niche of C. neoformans. Similar growth survival observed with the wt and Δgat1 mutants cultivated in pigeon guano media is an indication that though this ecological niche is rich in urea, urate, and creatine, which may probably be selective by the NCR but at the same time, the survival of the Δgat1 mutant in the same environment poses an investigation into the unique nitrogen sources suitable for such a mutant. Perhaps, such media may be rich in Pro or Arg as inferred from Lee et al. observation [111]. Notwithstanding, limited nitrogen sources in the environmental pigeon guano may have repressed the Gat1 for the mating process that promotes propagule/basidiospore formation of the filamentous stage of this opportunistic human infectious fungus. Lee et al. had discovered that mating is greatly enhanced through the robust filament and basidia formation either in the unilateral or bilateral crossing of Δgat1 mutants [111]. In the same way, deletion of Gat1 drastically reduces capsule formation, enhances melanin formation at physiological temperature, and promotes extreme thermotolerance but shows no significant effect on urease activity. Unexpectedly, these conditions seem to moderately reduce the pathogenic efficiency of Δgat1 mutants compared to the wt [111]. This shows that the key virulence factor of C. neoformans is firmly under the influence of NCR.
Keeping NCR under repression by Gat1p is necessary to facilitate the utilisation of other nitrogen sources. Lee et al. discovered that capsule formation increased considerably in the strain H99 when cultivated in a medium containing Asn, urea, urate, or creatinine but not Gln, Pro or Ala-containing medium. This capsule formation is prosaically affected by the presence of NH4+ in each of the media [111]. This shows that NH4+ represses capsule formation by activating NCR even in capsule-induced nitrogen sources, but, on the contrary, the presence of NH4+ induces melanin formation. To buttress this, Lee et al. observed melanin formation in each of the aforementioned media when prepared along with NH4+ except in the Pro medium, where a slight melanisation was observed even when NH4+ is present [111]. Without NH4+, creatinine and urate failed to support melanin formation, and only minimal melanisation was observed in the Ala medium. However, melanin was significantly induced in urea, Asn, or Gln-containing media [111]. This shows that NCR differentially regulates virulence accessory factors and that certain amino acids can override the NCR effect in the dire need for virulence. So, while NH4+ represses capsule formation in the presence of different nitrogen sources, melanin is significantly induced.
It is shown that among the numerous amino acid permeases, Aap2, Aap4, Aap5, Aap6, and Aap8 are more differentially expressed in the amino acid fortified SD media compared to the rich medium (YPD) [112,113], which means that NCR is highly induced in YPD compared to the SD. Without NH4+ (a preferred nitrogen source), global amino acid control is triggered by nitrogen starvation leading to preponderant production of t-RNA for amino acid biosynthesis in the yeast. For example, nitrogen starvation induces Trp biosynthetic enzymes encoded by Trp2, Trp3, Trp4, and Trp5 [112]; also, under repressive NCR, Aap2 and Aap3 are induced in the presence of Lys-containing medium [106]. However, amino acid supplements in SD do repress the general amino acid control (GAAC) but promote the APC-superfamily membrane transporters/permeases (APC, AAAP, AGCS, CCC, HAAAP, BCCT, SSS, NSS, NCS1, NCS2, and SulP families [114,115,116]).
Trp supplement alone in the SD could increase Aap1 and Aap2 transcripts by 3.5 and 3.8-fold; similarly, adding Trp+Met+His could improve Aap4, Aap2, Aap8, and Aap5 transcripts by 3.3, 4.2, 7.5, and 21-fold respectively [112]. The transcription analysis further showed that NCR could repress Aap2, Aap5, and Aap8 in the presence of NH4+ but not Aap4, which means Aap4 is sensitive to any nitrogen sources; however, Aap1 and Aap7 remained minimally expressed [112]. Though the NCR selectively represses Aap, carbon catabolite repression (CCR) appears to repress all the Aap expression in the presence of glucose (preferred carbon source) compared to galactose (alternative carbon source) [112].
The Δaap1Δaap2 mutants are thermosensitive (at 37oC in synthetic media fortified with amino acids), hypocapsulated, and hypovirulent in the G. mellonella infection model [117]. Mutants such as Δaap1, Δaap2, Δaap6, Δaap8, and Δaap1Δaap2 failed to display any significant difference in growth when compared to the wt at 30 or 37oC in SD fortified with amino acids/NH4+ or in YPD; however, growth reduction was observed in liquid culture of Δaap1 containing Met or Pro at 30 or 37oC [117]. Also, a liquid culture of Δaap1Δaap2 mutant containing Gln or Arg displayed growth reduction at 30oC, and Δaap8 mutant displayed growth reduction in media containing Met, Glu, and Trp.
Generally, Δaap1Δaap2 mutants appeared to show defective growth in 60% of all culturable amino acid media (AAM), which means that these amino acid permeases are essential for growth at 37oC on amino acids media, just like Aap4 and Aap5. However, Aap1, Aap2, Aap6, and Aap8 appeared to be non-essential for virulence because mutants of these genes, including Δaap1Δaap2 mutant, failed to show any significant difference in the mating, filamentation, melanin expression, PLB and urease activities when compared to the wt [117]. Apart from being non-essential for virulence, none of the mutants seemed to be affected by oxidative stress agents, alkaline conditions, osmotic and salt stress agents, and cell wall stress agents at 30 or 37oC. Surprisingly, unlike the single mutants (Δaap1, Δaap2, Δaap4, Δaap5, Δmup1, and Δmup3), the double mutants Δaap1Δaap2, Δaap4Δaap5, and Δmup1Δmup3 displayed reduced capsule size at 37oC when compared to the wt. However, the capsules are similar at 30oC, and the virulence of Δaap1Δaap2 and Δaap8 mutants only is attenuated in G. mellonella [117].
From Martho et al., Δaap2, Δaap4, Δaap5, Δmup1, Δmup3, and Δmup1Δmup3 showed no sign of attenuated virulence in G. mellonella, but Δaap4Δaap5 mutants are hypovirulent at 30 or 37oC [118]. Furthermore, virulence investigation in the murine model showed that mice inoculated with the wt and Δaap4 and Δaap5 died within a month, while the mice inoculated with Δaap4Δaap5 mutants and PBS-treated strain survived with viable recovery colonies from their lungs and livers. In addition, Δaap2, Δaap4, Δaap5, and Δaap4Δaap5 mutants displayed wt phenotypic sensitivity to AmpB but are hypersensitive to FCZ, and each of the mutants is hyper-resistant to eugenol due to Aap inactivation. This invariably shows that Aap4 and Aap5 are important permeases for thermotolerance, antifungal resistance, and oxidative stress response for tissue invasion, survival, and virulence in the animal model. By speculation, they are a potential target for therapeutic application. Notwithstanding, the compensatory essence of permeases/transporters cannot be ruled out. By harmonising several works on amino acid permeases – Aap, Mup, and Dao:
- ❖
- all mutants of aap and mup are not affected in rich media at high temperatures except Δaap4Δaap5,
- ❖
- are not affected in AAM at 37oC and in capsule formation except Δaap1Δaap2, Δaap4Δaap5, and Δmup1Δmup3,
- ❖
- are not sensitive to any stress agent except Δaap4Δaap5,
- ❖
- are virulent in G. mellonella except for Δaap1Δaap2 and Δaap8 (hypovirulent), Δaap4Δaap5 (avirulent as well as in murine model),
- ❖
- the Aap1, Aap2, Aap3, Aap4, and Aap5 are very similar in the sequence homology, and each of the Aaps in the double mutants appeared redundant in their functions,
- ❖
- the Aap3 and Aap7 expressions are below detection in YPD or SD; however, Lys-containing medium induced Aap2 and Aap3 expressions under alleviated NCR,
- ❖
- irrespective of the nitrogen sources, the expression of Aap6 remained relatively unchanged,
- ❖
- the Aap8 responds to amino acid-supplemented media only, but the highest expression is usually found in Aap2, Aap4, and Aap5,
- ❖
- Mup1 and Mup3 expressions are under the NCR regulation as Aap2 and Aap5 but can be induced by His, Trp, and Met when NRC is shut down,
- ❖
- only Mup1 can be induced by S-containing amino acids,
- ❖
- galactose induces the expression of all the Aap and Mup genes,
- ❖
- expression of Aap6 and Aap8 is temperature-independent,
- ❖
- Aap4, Aap5, and Mup1 induction increased from 30 to 37oC in SD medium,
- ❖
- Aap2, Aap4, Aap5, and Mup3 expressions are repressed further from 30 to 37oC except for Mup1, which is induced further,
- ❖
- irrespective of the growing media, there is no significant change in the growth of Δaap2, Δaap4, Δaap5, Δmup1, Δmup3 and Δmup1Δmup3 mutants at 30 or 37oC,
- ❖
- the significant growth defect of Δaap4Δaap5 mutants at especially 37oC in YPD or SD showed that the two permeases (or at least one of them) are important for thermotolerance,
- ❖
- the use of amino acids as nitrogen sources impaired the growth in Δaap4Δaap5 mutants,
- ❖
- relative to NH4+ at 30oC, Val, Ile, and Met-containing SD media poorly support the growth of C. neoformans, but Leu, Ser, Lys, and Phe are better nitrogen sources,
- ❖
- Gly, Asp, Asn, Glu, Gln, Arg, Trp, and Pro are highly competitive with NH4+ in culturable AAM,
- ❖
- at 37oC, Val and Met poorly support the growth of C. neoformans in SD media, but Gly, Leu, Ile, Ser, Trp, and Phe are good nitrogen sources, while Asp, Asn, Glu, Gln, Arg, Lys, and Pro are better nitrogen sources,
- ❖
- stereospecifically, var. gattii metabolises D-amino acids because of the more active expression of Dao1, Dao2, and Dao3 genes (encoding D-amino acid oxidase) but less metabolisable for var. neoformans, which prefers L-amino acids as nitrogen sources due to the inefficient evolutionary expression of Dao gene,
- ❖
- in all, growth is denser in L-amino acids containing media compared to corresponding D-amino acids media,
- ❖
- pathologically, Δdao mutants of C. neoformans are virulent, but Δdao mutants of C. gattii are attenuated,
- ❖
- positive correlation of Gat1 expression that represses NCR is confirmed to aid the expression of Aap when the preferred nitrogen source is absent/limited but uncertain with Dao expression,
- ❖
- L-Tyr failed to dissolve at permissive pH for C. neoformans to grow,
- ❖
- Aap2, Aap3, Mup1, and Mup3 may be global amino acid permeases/transporters because of the most significant growth defect of their corresponding mutants, especially in their double mutant states, from 30 to 37oC and their ability to be induced by various amino acids,
- ❖
- in addition to being a global permease and redundant transcription factors, Aap4 and Aap5 promote thermotolerance and response to oxidative stress, and the growth of the double mutant is significantly impacted from 30 to 37oC in a single amino acid medium or in the presence of ≥5 mM H2O2,
- ❖
- unlike the Δaap4 and Δaap5 mutants, the growth defect of Δaap4Δaap5 mutants at 37oC appeared to be restored as pH increased gradually into the alkaline state or when supplemented with 0.75 M NaCl (this condition generates H+ via Na+/H+ antiporter that drives other amino acid permeases to compensate for the deletion of aap4 and aap5),
- ❖
It is very surprising, therefore, to have observed the relevance of the Ras1 pathway in the control of amino acid permeases leading to thermotolerance. The Δras1 mutants had shown growth reduction compared to the wt in various amino acid fortified cultures at 30 or 37oC except for media with Ala, Cys, His, Tyr, and Thr that failed to support the growth of Δras1 mutants [117]. The involvement of Ras1 in the assimilation of amino acids is a unique feature of this transcription factor apart from the popular thermotolerance that it is known for. This means that amino acid uptake may be part of the function of Ras1 to facilitate thermotolerance. Alternative carbon and nitrogen sources such as galactose and Pro may also induce aap expression. It is not surprising, thus, to observe growth reduction in Δras1 mutants cultured in either SD or synthetic galactose (SG) fortified with NH4+ or Pro at either 30 or 37oC [117]. This shows the importance of Ras1p in the nutritional balance of C. neoformans against thermal stress. The controlling effect of Ras1p on the expression of Aap was further investigated by quantifying the expression level of each Aap transcript in Δras1 mutants. All Aap and Mup transcripts were repressed in Δras1 mutants in SD fortified with Trp, His, and Met at 30 and 37oC except Aap5, which showed a wt transcript level at 37oC [117]. The exception of Aap5 may suggest a different regulatory mechanism in addition to or different from Ras1 expression.
5. Secretory Hydrolytic Enzymes and Extracellular Vesicles in Cryptococcal Cells
Several extracellular hydrolytic enzymes from C. neoformans have been studied with diverse functions and are freely secreted under the regulatory N-terminal signal peptide or cleavage from the cell wall located GTI-anchored proteins or remained covalently attached to the cell wall glycans. Though the specific functional proteins, such as those designated for antioxidative functions, may lack N-terminal signal peptide and GTI-anchor [120] yet the majority of the proteins involved in adhesion may require GTI-anchorage. For example, ectophosphatase is a membrane-surface hydrolytic enzyme that often releases phosphate from phosphothreonine substrate and is liable to be inhibited irreversibly by sodium orthovanadate. Irreversible inhibition of this enzyme in C. neoformans has been characterised by reduced adhesion to animal epithelial cells – an important step towards fungal pathogenesis and tissue invasion [121].
Various hydrolytic enzymes, including endoproteinases, exoproteinases, and glycosidases, have been implicated in the cryptococcal invasion and colonisation of the CNS (neurotropism). Vu et al. discovered that extracellular secreted metalloproteinase, which is an M36 protease class of fungalysins, is highly required for fungal colonisation of the CNS and brain endothelium but not dissemination, and the deletion of the Mpr1 gene incapacitated the fungus to penetrate the endothelial layer of BBB to establish infection [122]. This prolongs the survival of the infected host due to reduced brain pathology.
The nature of the media determines the type of secreted protease. Animal tissue culture induces metalloproteinase and serine endoprotease as major peptidases, but aspartyl endoprotease activity dominates in minimal media. Remarkably, the deletion of Qsp1 reduced aspartyl endoprotease in the minimal media but elevated the secretion of metalloproteinase and serine endoprotease in animal tissue culture more than the wt [123]. Likewise, Brueske discovered that media supplemented with NH4+ and glucose could inhibit the total secreted proteolytic enzymes of C. neoformans while growth in essential salts medium supplemented with bovine serum albumin only enhanced the extracellular protease profile [124].
The involvement of C. neoformans extracellular proteinases in the degradation of host immunological proteins has been demonstrated to be responsible for fungal invasion and dissemination. Convincingly, such proteinases could degrade β-casein, murine IgG1, bovine IgG, human C5, haemoglobin, and γ-globulin in media containing carbon and nitrogen sources [125,126]. Regarding the potential to cause infection, a relatively low proteinase profile of C. neoformans var. gattii may have reduced tissue invasion, dissemination, and systemic spread of this serotype compared to the C. neoformans var. neoformans. However, there is no difference in the total proteolytic activities of the environmental isolates compared to the clinical isolates of C. neoformans [127]. Proteases are released at permissive and non-permissive temperatures irrespective of their sources, but a positive correlation existed between the capsule size and the protease activity in the clinical isolates, which was not found in the environmental/bird-dropping isolates [128]. The majority of these proteinases are serine proteases having optimal activities between pH 7 – 8 at 37oC [125]
In another related study, aspartyl endopeptidase (encoded by putative May1) and carboxypeptidase D1 (encoded by putative Cxd1, Cxd2 or Cxd3) have been identified as the major extracellular endoprotease and carboxypeptidase, respectively, in C. neoformans that help cope with the acidic environment and significantly enhance virulence [129]. Specifically, the total proteolytic activity in the mutants lacking metalloendopeptidase/metalloproteinase (encoded by Mpr1), serine carboxypeptidase (encoded by Scx1), carboxypeptidase C (encoded by Prc1), serine endopeptidase (encoded by Prb1), carboxypeptidase D (encoded by Cxd2, Cxd3), and aspartyl endopeptidase (encoded by Pep4) was not significantly different from the wt in an enriched medium unlike the Δmay1 and Δcxd1 mutants, which showed significantly reduced activity of the extracellular proteases. Because it is a major secreted protease, it is tagged major pepsin-like aspartyl endopeptidase with optimal expression in an acidic medium.
This aspartyl endopeptidase, by mechanisms, can be inhibited by pepstatin A and antiviral protease inhibitors [129,130,131]. Mutation of these extracellular protease-encoding genes seemed not to affect melanin production, but with the fact that melanin synthesis is also a cell wall anchoring process enriched with mixed GTI-anchored proteins, chitin, chitosan, and glycan then, the deletion of Prb1 had shown hypomelanisation in such a mutant [129].
Targeting the proteolytic activities of pathogens is a significant way of therapeutic approach against infections. A few works have shown that antiretroviral drugs in AIDS patients could antagonise some key extracellularly released hydrolytic enzymes and virulence production in pathogenic fungi such as C. albicans and C. neoformans, and this invariably inhibits fungal growth. Unlike Saquinavir, Nelfinavir, Lopinavir, Darunavir, Tipranavir, and Atazanavir, Amprenavir, Brecanavir, Darunavir, Indinavir, Ritonavir, and Saquinavir as well as macrocycles have been shown to inhibit fungal growth, attenuate capsule production, and inhibit urease and proteases but not phospholipase and melanin production in C. neoformans [129,130,131].
The use of specific and effective antiretroviral drugs in a co-morbidity patient with cryptococcosis may be responsible for the higher clearance of the cryptococcal cells because of the increased susceptibility to the killing by innate effector cells such as monocytes, neutrophils, and macrophages [130]. Unlike C. albicans, which are easily cleared by antiretroviral drugs, C. neoformans may be relatively difficult. C. neoformans aspartyl protease is one of the proteases not inhibited by highly active antiretroviral therapy such as Indinavir, Lopinavir, and Ritonavir, unlike C. albicans, and again the transcriptional level of this enzyme is strategically independent of the media used [132].
Urease (encoded by Ure1) is another hydrolytic metalloenzyme that hydrolyses the urea to ammonia and carbamate. Apart from Ure1, other transcription factors like Ure4, Ure6, and Ure7 have been associated with the production of accessory proteins required for this nickel-dependent functional protein complex [133]. Via the nickel/cobalt transporter (encoded by Nic1), Ni2+ is shuttled into the cytoplasm to combine with the apo-urease (encoded by Ure1) via Ure7p to form an active metallocentric urease. The urease reaction may increase the media pH to create an alkali environment. While this may be deleterious to the infected host, C. neoformans deploys the pH-sensing signalling factors such as Rim21/Rim8/Rim20/Rim13 complex to counter the pH surge via the activation of Rim101 [18].
The Δure1 mutants displayed comparable phenotypic and virulence features with the wt; however, evidence existed that deleting this gene may attenuate the progression of infection caused by C. neoformans. This is corroborated by the outcome of Cox et al. that murine infected with Δure1 mutant, whether intranasally/intravenously, failed to show pulmonary distress, displayed delayed hydrocephalus and wasting, and significantly lived longer than the wt-infected mice [134]. Hydrocephalus and wasting are delayed manifestations of meningoencephalitis caused by Δure1 mutants after dissemination into the brain, but the infection caused by urease-positive strains proceeds rapidly in the lungs to result in acute pneumonitis capable of killing the mice before the brain encephalitis sets in [134]. Notwithstanding, wholistic urease activation is essential for the microcapillary sequestration, dissemination, and brain invasion of C. neoformans because Δure1, Δure7, and Δnic1 mutants, which lacked urease activities, significantly displayed defective growth in the presence of urea with poor blood-to-brain invasion in an animal model [133,135]. Though urease seems to be essential to the dissemination, invasion, and survival of C. neoformans in vivo yet, clinically infectious urease-negative isolates capable of dissemination and invasion have been reported in AIDs patients [136,137], which means this enzyme may be playing second fiddle in the pathogenesis of C. neoformans [138].
Cryptococcal cells can assimilate nitrogenous bases and nucleosides to produce nucleotides via the salvage pathway. By producing extracellular deoxyribonucleases (DNases), most infectious and parasitic fungi degrade the host and surrounding nucleic acids to salvage intermediates molecules needed to build nucleotides. Excessive nitrogenous compounds are degraded by ureases. The release of DNases tandemly correlated with the increased activity of ureases in most fungi except in a few isolates of Endomycopsis fibuligera and Lipomyces starkeyi [139].
Sod is another membrane residence anti-oxidative enzyme, which converts superoxide to H2O2 and O2 and complements the effort of melanin against reactive oxidants. The expression of this enzyme increases with temperature to facilitate growth and thermotolerance, thereby contributing to the virulence of C. neoformans [140].
Phospholipases (PLs) are also secretory enzymes that degrade membrane phospholipids in response to signal transduction. It is a complex extracellularly released enzyme that facilitates fungi-host cell attachment, tissue invasion, and hydrolysis of the epithelial plasma membrane for penetration. The activity of the PLs at the cell wall promotes the integrity for survival against temperature, antifungals, oxidative stress, and cell wall destabilisers [22]. Moreover, the involvement of PLC as a component of PLs in the virulence expression, antifungal resistance, homeostasis and IP3 kinase activity, and cytokinesis displays the incredible functions of this membrane-associated enzyme [28,29].
Special attention is needed on some extracellular lipases capable of inducing antibodies in infected animal studies. Incredibly, Chen et al. discovered that 92% of the tested cryptococcal strains secreted butyrate and caprylate esterase lipase. At the same time, some also produced acid phosphatase, naphthol-AS-BI-phosphohydrolase, and β-glucosidase, and the sera of infected rodents were able to precipitate several of these secreted proteins [141].
Phosphatases are enzymes released to remove the phosphate group (dephosphorylation) from transcription factors that are usually activated/deactivated by phosphorylation through serine/threonine/tyrosine kinase. This controls the on/off regulations of signalling pathways orchestrated by transcription factors such as Hog1 MAPK, Ste20/Pak kinase, Pkc1 MAPK, and cAMP/Pka. Such phosphatase could operate optimally in an acidic medium (acid phosphatase) or alkali medium (alkaline phosphatase). For example, C. neoformans can release extracellular acid phosphatase encoded by Aph1, which facilitates fungal adhesion to monocytes, but deletion of this gene attenuates the fungus virulence [38,121].
Extracellular vesicles described as “virulence bags” are exosomal secretory vesicles containing microbodies and proteins, some of which are associated with virulence because of the presence of laccase, urease, and phosphatase in these “virulence bags” [12]. The proteomic analysis of the C. neoformans secretomes showed diverse functional proteins and enzymes classified as antioxidative and heat-shock proteins (Hsp70 and Hsp90), signal transduction and nucleotide salvage enzymes, ribosomal proteins, metabolic enzymes (for sugar, lipid, protein, and amino acid), nucleoproteins, membrane proteins (transporters, carriers, pumps, channels, adhesins, inositol-3-phosphate synthase, Rab proteins), cytoskeleton proteins (actin and actin-binding proteins, tubulin, and annexin), and a few mitochondria membrane proteins [12]. In addition, the presence of UDP-glucose dehydrogenase (encoded by Ugd1) and UDP-glucuronate decarboxylase (encoded by Uxs1) in the extracellular vesicles of C. neoformans showed the presence of functional enzymes that not only promote capsule formation but also enhance cell wall integrity and growth at a temperature above 30oC [142].
6. Conclusions and Perspectives
The significant involvement of different hydrolytic enzymes, transcription factors, and membrane proteins like pumps, permeases, and transporters was discussed in detail in this article. While each of these proteins plays an important role in the survival, adaptation, and pathogenesis of C. neoformans in animals and humans, some are redundant with barely known function. These attributes are also common in other invasive fungi responsible for candidiasis and aspergillosis. The multiplicity and redundancy of these proteins may be critical in catabolite repression to rescuing the fungi when in environment with alternative carbon sources, low oxygen, fluctuating pH, high salt contents, high level of oxidant, low phosphate content, low ammonium content, and low micronutrient elements. Ability to produce exocytic vesicles of microbodies, enzymes, and virulence proteins appears as alternative way of inducing cellular morphology, biofilm formation, cellular communication, filamentation, fruiting, and mating. We described these proteins as weapons that facilitate tissue invasion, persistence, and infection in animals and humans. With reference to the supplementary files, we further described the morphological and functional impairments associated with the deletion or alteration in the encoding genes for these proteins. Because of the functional similarities of these proteins with human cells, targeting any of them for antifungal drug discovery needs careful consideration that will preclude the consequence in the human cells.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. File: Genes, Transcription Factors, Protein Kinases, Intracellular Proteins, and Other Abbreviations as Used in the Article.
Author Contributions
Conceptualization, O.S. and O.M.; methodology, O.S.; software, O.S.; validation, O.S. and O.M.; formal analysis, O.S.; investigation, O.S.; resources, O.S. and O.M.; data curation, O.S.; writing—original draft preparation, O.S.; writing—review and editing, O.S. and O.M.; visualization, O.S. and O.M.; supervision, O.M.; project administration, O.M.; funding acquisition, O.M. All authors have read and agreed to the published version of the manuscript.
Funding
The authors declared that no funding was obtained for this article.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors appreciated the conducive research environment and administrative procedures put in place by the Research Office of University of the Free State.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kmetzsch, L.; Staats, C.C.; Cupertino, J.B.; Fonseca, F.L.; Rodrigues, M.L.; Schrank, A.; Vainstein, M.H. The calcium transporter Pmc1 provides Ca2+ tolerance and influences the progression of murine cryptococcal infection. FEBS J 2013, 280, 4853–4864. [Google Scholar] [CrossRef]
- Kmetzsch, L.; Staats, C.C.; Simon, E.; Fonseca, F.L.; Oliveira, D.L.; Joffe, L.S.; Rodrigues, J.; Lourenco, R.F.; Gomes, S.L.; Nimrichter, L.; et al. The GATA-type transcriptional activator Gat1 regulates nitrogen uptake and metabolism in the human pathogen Cryptococcus neoformans. Fungal Genet Biol 2011, 48, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: a common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Mowat, E.; Jones, B.; Williams, C.; Lopez-Ribot, J. Our current understanding of fungal biofilms. Critical reviews in microbiology 2009, 35, 340–355. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annual review of microbiology 1995, 49, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.R.; Casadevall, A. Cryptococcus neoformans cells in biofilms are less susceptible than planktonic cells to antimicrobial molecules produced by the innate immune system. Infection and immunity 2006, 74, 6118–6123. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.R.; Casadevall, A. Susceptibility of Cryptococcus neoformans biofilms to antifungal agents in vitro. Antimicrobial agents and chemotherapy 2006, 50, 1021–1033. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.R.; Casadevall, A. Cryptococcus neoformans biofilm formation depends on surface support and carbon source and reduces fungal cell susceptibility to heat, cold, and UV light. Applied and environmental microbiology 2007, 73, 4592–4601. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.R.; Casadevall, A. Biofilm Formation by Cryptococcus neoformans. Microbiology Spectrum 2015, 3, 3.3.05. [Google Scholar] [CrossRef]
- Santi, L.; Beys-da-Silva, W.O.; Berger, M.; Calzolari, D.; Guimarães, J.A.; Moresco, J.J.; Yates, J.R., 3rd. Proteomic profile of Cryptococcus neoformans biofilm reveals changes in metabolic processes. J Proteome Res 2014, 13, 1545–1559. [Google Scholar] [CrossRef]
- Paul, C.; Emeka, N. Pathogenicity of Cryptococcus neoformans VNI (ST 32) recovered from environmental and clinical isolates in Nigeria. Comparative Clinical Pathology 2019, 28, 1013–1024. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Nakayasu, E.S.; Oliveira, D.L.; Nimrichter, L.; Nosanchuk, J.D.; Almeida, I.C.; Casadevall, A. Extracellular vesicles produced by Cryptococcus neoformans contain protein components associated with virulence. Eukaryot Cell 2008, 7, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Derengowski Lda, S.; Paes, H.C.; Albuquerque, P.; Tavares, A.H.; Fernandes, L.; Silva-Pereira, I.; Casadevall, A. The transcriptional response of Cryptococcus neoformans to ingestion by Acanthamoeba castellanii and macrophages provides insights into the evolutionary adaptation to the mammalian host. Eukaryot Cell 2013, 12, 761–774. [Google Scholar] [CrossRef]
- Wang, Z.A.; Li, L.X.; Doering, T.L. Unraveling synthesis of the cryptococcal cell wall and capsule. Glycobiology 2018, 28, 719–730. [Google Scholar] [CrossRef]
- Panepinto, J.; Liu, L.; Ramos, J.; Zhu, X.; Valyi-Nagy, T.; Eksi, S.; Fu, J.; Jaffe, H.A.; Wickes, B.; Williamson, P.R. The DEAD-box RNA helicase Vad1 regulates multiple virulence-associated genes in Cryptococcus neoformans. J Clin Invest 2005, 115, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Kraus, P.R.; Boily, M.J.; Heitman, J. Cryptococcus neoformans gene expression during murine macrophage infection. Eukaryot Cell 2005, 4, 1420–1433. [Google Scholar] [CrossRef] [PubMed]
- Price, M.S.; Betancourt-Quiroz, M.; Price, J.L.; Toffaletti, D.L.; Vora, H.; Hu, G.; Kronstad, J.W.; Perfect, J.R. Cryptococcus neoformans requires a functional glycolytic pathway for disease but not persistence in the host. mBio 2011, 2, e00103–e00111. [Google Scholar] [CrossRef] [PubMed]
- Maliehe, M.; Ntoi, M.A.; Lahiri, S.; Folorunso, O.S.; Ogundeji, A.O.; Pohl, C.H.; Sebolai, O.M. Environmental Factors That Contribute to the Maintenance of Cryptococcus neoformans Pathogenesis. Microorganisms 2020, 8, 180. [Google Scholar] [CrossRef]
- Chen, S.C.; Muller, M.; Zhou, J.Z.; Wright, L.C.; Sorrell, T.C. Phospholipase activity in Cryptococcus neoformans: a new virulence factor? J Infect Dis 1997, 175, 414–420. [Google Scholar] [CrossRef]
- Cox, G.M.; McDade, H.C.; Chen, S.C.; Tucker, S.C.; Gottfredsson, M.; Wright, L.C.; Sorrell, T.C.; Leidich, S.D.; Casadevall, A.; Ghannoum, M.A.; et al. Extracellular phospholipase activity is a virulence factor for Cryptococcus neoformans. Mol Microbiol 2001, 39, 166–175. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Mirbod, F.; Filler, S.G.; Banno, Y.; Cole, G.T.; Kitajima, Y.; Edwards, J.E., Jr.; Nozawa, Y.; Ghannoum, M.A. Evidence implicating phospholipase as a virulence factor of Candida albicans. Infect Immun 1995, 63, 1993–1998. [Google Scholar] [CrossRef]
- Siafakas, A.R.; Sorrell, T.C.; Wright, L.C.; Wilson, C.; Larsen, M.; Boadle, R.; Williamson, P.R.; Djordjevic, J.T. Cell wall-linked cryptococcal phospholipase B1 is a source of secreted enzyme and a determinant of cell wall integrity. J Biol Chem 2007, 282, 37508–37514. [Google Scholar] [CrossRef]
- Saito, K.; Sugatani, J.; Okumura, T. Phospholipase B from Penicillium notatum. In Methods in enzymology; Elsevier, 1991; Volume 197, pp. 446–456. [Google Scholar]
- Lee, K.S.; Patton, J.L.; Fido, M.; Hines, L.K.; Kohlwein, S.D.; Paltauf, F.; Henry, S.A.; Levin, D.E. The Saccharomyces cerevisiae PLB1 gene encodes a protein required for lysophospholipase and phospholipase B activity. J Biol Chem 1994, 269, 19725–19730. [Google Scholar] [CrossRef] [PubMed]
- Leidich, S.D.; Ibrahim, A.S.; Fu, Y.; Koul, A.; Jessup, C.; Vitullo, J.; Fonzi, W.; Mirbod, F.; Nakashima, S.; Nozawa, Y.; et al. Cloning and disruption of caPLB1, a phospholipase B gene involved in the pathogenicity of Candida albicans. J Biol Chem 1998, 273, 26078–26086. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Wickes, B.L.; Miller, G.F.; Penoyer, L.A.; Kwon-Chung, K.J. Cryptococcus neoformans STE12α regulates virulence but is not essential for mating. J Exp Med 2000, 191, 871–882. [Google Scholar] [CrossRef]
- Cooper, K.F.; Mallory, M.J.; Strich, R. Oxidative stress-induced destruction of the yeast C-type cyclin Ume3p requires phosphatidylinositol-specific phospholipase C and the 26S proteasome. Mol Cell Biol 1999, 19, 3338–3348. [Google Scholar] [CrossRef]
- Chayakulkeeree, M.; Sorrell, T.C.; Siafakas, A.R.; Wilson, C.F.; Pantarat, N.; Gerik, K.J.; Boadle, R.; Djordjevic, J.T. Role and mechanism of phosphatidylinositol-specific phospholipase C in survival and virulence of Cryptococcus neoformans. Mol Microbiol 2008, 69, 809–826. [Google Scholar] [CrossRef]
- Lev, S.; Desmarini, D.; Li, C.; Chayakulkeeree, M.; Traven, A.; Sorrell, T.C.; Djordjevic, J.T. Phospholipase C of Cryptococcus neoformans regulates homeostasis and virulence by providing inositol trisphosphate as a substrate for Arg1 kinase. Infect Immun 2013, 81, 1245–1255. [Google Scholar] [CrossRef]
- Narasipura, S.D.; Ault, J.G.; Behr, M.J.; Chaturvedi, V.; Chaturvedi, S. Characterization of Cu,Zn superoxide dismutase (SOD1) gene knock-out mutant of Cryptococcus neoformans var. gattii: role in biology and virulence. Mol Microbiol 2003, 47, 1681–1694. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Wong, B.; Newman, S.L. Oxidative killing of Cryptococcus neoformans by human neutrophils - Evidence that fungal mannitol protects by scavenging reactive oxygen intermediates. Journal of Immunology 1996, 156, 3836–3840. [Google Scholar] [CrossRef]
- Bjorkbom, A.; Ohvo-Rekila, H.; Kankaanpaa, P.; Nyholm, T.K.; Westerlund, B.; Slotte, J.P. Characterisation of membrane properties of inositol phosphorylceramide. Biochim Biophys Acta 2010, 1798, 453–460. [Google Scholar] [CrossRef]
- Beeler, T.; Gable, K.; Zhao, C.; Dunn, T. A Novel Protein, Csg2p, Is Required for Ca2+ Regulation in Saccharomyces-Cerevisiae. Journal of Biological Chemistry 1994, 269, 7279–7284. [Google Scholar] [CrossRef] [PubMed]
- Uemura, S.; Kihara, A.; Iwaki, S.; Inokuchi, J.; Igarashi, Y. Regulation of the transport and protein levels of the inositol phosphorylceramide mannosyltransferases Csg1 and Csh1 by the Ca2+-binding protein Csg2. J Biol Chem 2007, 282, 8613–8621. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.C.; Lester, R.L. Yeast sphingolipids. Biochimica et Biophysica Acta (BBA)-General Subjects 1999, 1426, 347–357. [Google Scholar] [CrossRef]
- Toh-e, A.; Ohkusu, M.; Li, H.M.; Shimizu, K.; Takahashi-Nakaguchi, A.; Gonoi, T.; Kawamoto, S.; Kanesaki, Y.; Yoshikawa, H.; Nishizawa, M. Identification of genes involved in the phosphate metabolism in Cryptococcus neoformans. Fungal Genet Biol 2015, 80, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, M.; Reiner, E.; Hu, G.; Tam, N.; Oliveira, D.L.; Caza, M.; Yeon, J.H.; Kim, J.; Kastrup, C.J.; Jung, W.H.; et al. Defects in phosphate acquisition and storage influence virulence of Cryptococcus neoformans. Infect Immun 2014, 82, 2697–2712. [Google Scholar] [CrossRef] [PubMed]
- Lev, S.; Crossett, B.; Cha, S.Y.; Desmarini, D.; Li, C.; Chayakulkeeree, M.; Wilson, C.F.; Williamson, P.R.; Sorrell, T.C.; Djordjevic, J.T. Identification of Aph1, a phosphate-regulated, secreted, and vacuolar acid phosphatase in Cryptococcus neoformans. mBio 2014, 5, e01649–e01614. [Google Scholar] [CrossRef] [PubMed]
- Lev, S.; Kaufman-Francis, K.; Desmarini, D.; Juillard, P.G.; Li, C.; Stifter, S.A.; Feng, C.G.; Sorrell, T.C.; Grau, G.E.; Bahn, Y.S.; et al. Pho4 Is Essential for Dissemination of Cryptococcus neoformans to the Host Brain by Promoting Phosphate Uptake and Growth at Alkaline pH. mSphere 2017, 2, e00381–e00316. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.H.; Saikia, S.; Hu, G.; Wang, J.; Fung, C.K.; D'Souza, C.; White, R.; Kronstad, J.W. HapX positively and negatively regulates the transcriptional response to iron deprivation in Cryptococcus neoformans. PLoS Pathog 2010, 6, e1001209. [Google Scholar] [CrossRef]
- Hu, G.; Horianopoulos, L.; Sanchez-Leon, E.; Caza, M.; Jung, W.; Kronstad, J.W. The monothiol glutaredoxin Grx4 influences thermotolerance, cell wall integrity, and Mpk1 signaling in Cryptococcus neoformans. G3 (Bethesda) 2021, 11. [Google Scholar] [CrossRef]
- Attarian, R.; Hu, G.; Sánchez-León, E.; Caza, M.; Croll, D.; Do, E.; Bach, H.; Missall, T.; Lodge, J.; Jung, W.H. The monothiol glutaredoxin Grx4 regulates iron homeostasis and virulence in Cryptococcus neoformans. MBio 2018, 9, e02377–e02318. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.H.; Kronstad, J.W. Iron and fungal pathogenesis: a case study with Cryptococcus neoformans. Cell Microbiol 2008, 10, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.H.; Hu, G.; Kuo, W.; Kronstad, J.W. Role of ferroxidases in iron uptake and virulence of Cryptococcus neoformans. Eukaryot Cell 2009, 8, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.H.; Sham, A.; Lian, T.; Singh, A.; Kosman, D.J.; Kronstad, J.W. Iron source preference and regulation of iron uptake in Cryptococcus neoformans. PLoS Pathog 2008, 4, e45. [Google Scholar] [CrossRef] [PubMed]
- Lian, T.; Simmer, M.I.; D'Souza, C.A.; Steen, B.R.; Zuyderduyn, S.D.; Jones, S.J.; Marra, M.A.; Kronstad, J.W. Iron-regulated transcription and capsule formation in the fungal pathogen Cryptococcus neoformans. Mol Microbiol 2005, 55, 1452–1472. [Google Scholar] [CrossRef] [PubMed]
- Radisky, D.C.; Babcock, M.C.; Kaplan, J. The yeast frataxin homologue mediates mitochondrial iron efflux. Evidence for a mitochondrial iron cycle. J Biol Chem 1999, 274, 4497–4499. [Google Scholar] [CrossRef] [PubMed]
- Spizzo, T.; Byersdorfer, C.; Duesterhoeft, S.; Eide, D. The yeast FET5 gene encodes a FET3-related multicopper oxidase implicated in iron transport. Mol Gen Genet 1997, 256, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Wiesenberger, G.; Link, T.A.; von Ahsen, U.; Waldherr, M.; Schweyen, R.J. MRS3 and MRS4, two suppressors of mtRNA splicing defects in yeast, are new members of the mitochondrial carrier family. J Mol Biol 1991, 217, 23–37. [Google Scholar] [CrossRef]
- Urbanowski, J.L.; Piper, R.C. The iron transporter Fth1p forms a complex with the Fet5 iron oxidase and resides on the vacuolar membrane. J Biol Chem 1999, 274, 38061–38070. [Google Scholar] [CrossRef]
- Li, L.; Chen, O.S.; McVey Ward, D.; Kaplan, J. CCC1 is a transporter that mediates vacuolar iron storage in yeast. J Biol Chem 2001, 276, 29515–29519. [Google Scholar] [CrossRef]
- Nyhus, K.J.; Ozaki, L.S.; Jacobson, E.S. Role of mitochondrial carrier protein Mrs3/4 in iron acquisition and oxidative stress resistance of Cryptococcus neoformans. Med Mycol 2002, 40, 581–591. [Google Scholar] [CrossRef]
- Tangen, K.L.; Jung, W.H.; Sham, A.P.; Lian, T.; Kronstad, J.W. The iron- and cAMP-regulated gene SIT1 influences ferrioxamine B utilization, melanisation and cell wall structure in Cryptococcus neoformans. Microbiology (Reading) 2007, 153, 29–41. [Google Scholar] [CrossRef]
- Robertson, L.S.; Causton, H.C.; Young, R.A.; Fink, G.R. The yeast A kinases differentially regulate iron uptake and respiratory function. Proc Natl Acad Sci U S A 2000, 97, 5984–5988. [Google Scholar] [CrossRef]
- Nyhus, K.J.; Wilborn, A.T.; Jacobson, E.S. Ferric iron reduction by Cryptococcus neoformans. Infect Immun 1997, 65, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Wright, L.; Bubb, W.; Davidson, J.; Santangelo, R.; Krockenberger, M.; Himmelreich, U.; Sorrell, T. Metabolites released by Cryptococcus neoformans var. neoformans and var. gattii differentially affect human neutrophil function. Microbes Infect 2002, 4, 1427–1438. [Google Scholar] [CrossRef]
- Reese, A.J.; Doering, T.L. Cell wall α-1,3-glucan is required to anchor the Cryptococcus neoformans capsule. Mol Microbiol 2003, 50, 1401–1409. [Google Scholar] [CrossRef]
- Walton, F.J.; Idnurm, A.; Heitman, J. Novel gene functions required for melanisation of the human pathogen Cryptococcus neoformans. Mol Microbiol 2005, 57, 1381–1396. [Google Scholar] [CrossRef] [PubMed]
- Waterman, S.R.; Hacham, M.; Hu, G.; Zhu, X.; Park, Y.D.; Shin, S.; Panepinto, J.; Valyi-Nagy, T.; Beam, C.; Husain, S.; et al. Role of a CUF1/CTR4 copper regulatory axis in the virulence of Cryptococcus neoformans. J Clin Invest 2007, 117, 794–802. [Google Scholar] [CrossRef]
- Jung, W.H.; Sham, A.; White, R.; Kronstad, J.W. Iron regulation of the major virulence factors in the AIDS-associated pathogen Cryptococcus neoformans. PLoS Biol 2006, 4, e410. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Festa, R.A.; Chen, Y.L.; Espart, A.; Palacios, O.; Espin, J.; Capdevila, M.; Atrian, S.; Heitman, J.; Thiele, D.J. Cryptococcus neoformans copper detoxification machinery is critical for fungal virulence. Cell Host Microbe 2013, 13, 265–276. [Google Scholar] [CrossRef]
- Nicola, A.M.; Robertson, E.J.; Albuquerque, P.; Derengowski Lda, S.; Casadevall, A. Nonlytic exocytosis of Cryptococcus neoformans from macrophages occurs in vivo and is influenced by phagosomal pH. mBio 2011, 2. [Google Scholar] [CrossRef]
- Ding, C.; Yin, J.; Tovar, E.M.; Fitzpatrick, D.A.; Higgins, D.G.; Thiele, D.J. The copper regulon of the human fungal pathogen Cryptococcus neoformans H99. Mol Microbiol 2011, 81, 1560–1576. [Google Scholar] [CrossRef] [PubMed]
- Suo, C.H.; Ma, L.J.; Li, H.L.; Sun, J.F.; Li, C.; Lin, M.H.; Sun, T.S.; Du, W.; Li, Y.J.; Gao, X.D.; et al. Investigation of Cryptococcus neoformans magnesium transporters reveals important role of vacuolar magnesium transporter in regulating fungal virulence factors. Microbiologyopen 2018, 7, e00564. [Google Scholar] [CrossRef]
- da Costa, B.M.; Cornish, K.; Keasling, J.D. Manipulation of intracellular magnesium levels in Saccharomyces cerevisiae with deletion of magnesium transporters. Appl Microbiol Biotechnol 2007, 77, 411–425. [Google Scholar] [CrossRef]
- Pisat, N.P.; Pandey, A.; Macdiarmid, C.W. MNR2 regulates intracellular magnesium storage in Saccharomyces cerevisiae. Genetics 2009, 183, 873–884. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, S.; Wang, J.; Wang, X.; Gao, B.; Fan, Q.; Sun, F.; Zhou, B. A novel mitochondrial carrier protein Mme1 acts as a yeast mitochondrial magnesium exporter. Biochim Biophys Acta 2015, 1853, 724–732. [Google Scholar] [CrossRef]
- Chun, C.D.; Liu, O.W.; Madhani, H.D. A link between virulence and homeostatic responses to hypoxia during infection by the human fungal pathogen Cryptococcus neoformans. PLoS Pathog 2007, 3, e22. [Google Scholar] [CrossRef]
- Chang, Y.C.; Bien, C.M.; Lee, H.; Espenshade, P.J.; Kwon-Chung, K.J. Sre1p, a regulator of oxygen sensing and sterol homeostasis, is required for virulence in Cryptococcus neoformans. Mol Microbiol 2007, 64, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Bien, C.M.; Chang, Y.C.; Nes, W.D.; Kwon-Chung, K.J.; Espenshade, P.J. Cryptococcus neoformans Site-2 protease is required for virulence and survival in the presence of azole drugs. Mol Microbiol 2009, 74, 672–690. [Google Scholar] [CrossRef]
- Todd, B.L.; Stewart, E.V.; Burg, J.S.; Hughes, A.L.; Espenshade, P.J. Sterol regulatory element binding protein is a principal regulator of anaerobic gene expression in fission yeast. Mol Cell Biol 2006, 26, 2817–2831. [Google Scholar] [CrossRef]
- Ingavale, S.S.; Chang, Y.C.; Lee, H.; McClelland, C.M.; Leong, M.L.; Kwon-Chung, K.J. Importance of mitochondria in survival of Cryptococcus neoformans under low oxygen conditions and tolerance to cobalt chloride. PLoS Pathog 2008, 4, e1000155. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Kojima, K.; Cox, G.M.; Heitman, J. A unique fungal two-component system regulates stress responses, drug sensitivity, sexual development, and virulence of Cryptococcus neoformans. Mol Biol Cell 2006, 17, 3122–3135. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.L.; Connor, J.R.; Jones, B.C. Iron in the brain. Nutr Rev 1993, 51, 157–170. [Google Scholar] [CrossRef]
- Martin, C.E.; Oh, C.S.; Jiang, Y. Regulation of long chain unsaturated fatty acid synthesis in yeast. Biochim Biophys Acta 2007, 1771, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Chellappa, R.; Kandasamy, P.; Oh, C.S.; Jiang, Y.; Vemula, M.; Martin, C.E. The membrane proteins, Spt23p and Mga2p, play distinct roles in the activation of Saccharomyces cerevisiae OLE1 gene expression. Fatty acid-mediated regulation of Mga2p activity is independent of its proteolytic processing into a soluble transcription activator. J Biol Chem 2001, 276, 43548–43556. [Google Scholar] [CrossRef]
- Jacobson, E.S.; Emery, H.S. Catecholamine uptake, melanization, and oxygen toxicity in Cryptococcus neoformans. J Bacteriol 1991, 173, 401–403. [Google Scholar] [CrossRef]
- Emery, H.S.; Shelburne, C.P.; Bowman, J.P.; Fallon, P.G.; Schulz, C.A.; Jacobson, E.S. Genetic study of oxygen resistance and melanization in Cryptococcus neoformans. Infect Immun 1994, 62, 5694–5697. [Google Scholar] [CrossRef]
- Test, S.T.; Weiss, S.J. The generation of utilisation of chlorinated oxidants by human neutrophils. Advances in Free Radical Biology & Medicine 1986, 2, 91–116. [Google Scholar] [CrossRef]
- Korytowski, W.; Sarna, T. Bleaching of melanin pigments. Role of copper ions and hydrogen peroxide in autooxidation and photooxidation of synthetic dopa-melanin. J Biol Chem 1990, 265, 12410–12416. [Google Scholar] [CrossRef]
- Tripp, B.C.; Smith, K.; Ferry, J.G. Carbonic anhydrase: new insights for an ancient enzyme. J Biol Chem 2001, 276, 48615–48618. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Cox, G.M.; Perfect, J.R.; Heitman, J. Carbonic anhydrase and CO2 sensing during Cryptococcus neoformans growth, differentiation, and virulence. Curr Biol 2005, 15, 2013–2020. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Ko, Y.J.; Maeng, S.; Floyd, A.; Heitman, J.; Bahn, Y.S. Comparative transcriptome analysis of the CO2 sensing pathway via differential expression of carbonic anhydrase in Cryptococcus neoformans. Genetics 2010, 185, 1207–1219. [Google Scholar] [CrossRef] [PubMed]
- Missall, T.A.; Lodge, J.K. Function of the thioredoxin proteins in Cryptococcus neoformans during stress or virulence and regulation by putative transcriptional modulators. Mol Microbiol 2005, 57, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Bahn, Y.S.; Muhlschlegel, F.A. CO2 sensing in fungi and beyond. Curr Opin Microbiol 2006, 9, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Bahn, Y.S.; Cox, G.M.; Heitman, J. G protein-coupled receptor Gpr4 senses amino acids and activates the cAMP-PKA pathway in Cryptococcus neoformans. Mol Biol Cell 2006, 17, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Bahn, Y.S.; Hicks, J.K.; Giles, S.S.; Cox, G.M.; Heitman, J. Adenylyl cyclase-associated protein Aca1 regulates virulence and differentiation of Cryptococcus neoformans via the cyclic AMP-protein kinase A cascade. Eukaryot Cell 2004, 3, 1476–1491. [Google Scholar] [CrossRef] [PubMed]
- Maeng, S.; Ko, Y.J.; Kim, G.B.; Jung, K.W.; Floyd, A.; Heitman, J.; Bahn, Y.S. Comparative transcriptome analysis reveals novel roles of the Ras and cyclic AMP signaling pathways in environmental stress response and antifungal drug sensitivity in Cryptococcus neoformans. Eukaryot Cell 2010, 9, 360–378. [Google Scholar] [CrossRef] [PubMed]
- Martho, K.F.; Brustolini, O.J.B.; Vasconcelos, A.T.; Vallim, M.A.; Pascon, R.C. The Glycerol Phosphatase Gpp2: A Link to Osmotic Stress, Sulfur Assimilation and Virulence in Cryptococcus neoformans. Front Microbiol 2019, 10, 2728. [Google Scholar] [CrossRef] [PubMed]
- de Melo, A.T.; Martho, K.F.; Roberto, T.N.; Nishiduka, E.S.; Machado, J., Jr.; Brustolini, O.J.B.; Tashima, A.K.; Vasconcelos, A.T.; Vallim, M.A.; Pascon, R.C. The regulation of the sulfur amino acid biosynthetic pathway in Cryptococcus neoformans: the relationship of Cys3, Calcineurin, and Gpp2 phosphatases. Sci Rep 2019, 9, 11923. [Google Scholar] [CrossRef]
- Ko, Y.J.; Yu, Y.M.; Kim, G.B.; Lee, G.W.; Maeng, P.J.; Kim, S.; Floyd, A.; Heitman, J.; Bahn, Y.S. Remodeling of global transcription patterns of Cryptococcus neoformans genes mediated by the stress-activated HOG signaling pathways. Eukaryot Cell 2009, 8, 1197–1217. [Google Scholar] [CrossRef]
- Grewal, S.I.; Bonaduce, M.J.; Klar, A.J. Histone deacetylase homologs regulate epigenetic inheritance of transcriptional silencing and chromosome segregation in fission yeast. Genetics 1998, 150, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Maier, E.J.; Haynes, B.C.; Gish, S.R.; Wang, Z.A.; Skowyra, M.L.; Marulli, A.L.; Doering, T.L.; Brent, M.R. Model-driven mapping of transcriptional networks reveals the circuitry and dynamics of virulence regulation. Genome Res 2015, 25, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Steen, B.R.; Lian, T.; Sham, A.P.; Tam, N.; Tangen, K.L.; Kronstad, J.W. Transcriptional regulation by protein kinase A in Cryptococcus neoformans. PLoS Pathog 2007, 3, e42. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.C.; Bender, J.A.; Fink, G.R. Transcriptional response of Candida albicans upon internalisation by macrophages. Eukaryot Cell 2004, 3, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Rude, T.H.; Toffaletti, D.L.; Cox, G.M.; Perfect, J.R. Relationship of the glyoxylate pathway to the pathogenesis of Cryptococcus neoformans. Infect Immun 2002, 70, 5684–5694. [Google Scholar] [CrossRef] [PubMed]
- McClelland, E.E.; Ramagopal, U.A.; Rivera, J.; Cox, J.; Nakouzi, A.; Prabu, M.M.; Almo, S.C.; Casadevall, A. A Small Protein Associated with Fungal Energy Metabolism Affects the Virulence of Cryptococcus neoformans in Mammals. PLoS Pathog 2016, 12, e1005849. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wei, D.; Li, Z.; Sun, Z.; Pan, J.; Zhu, X. Cryptococcal phosphoglucose isomerase is required for virulence factor production, cell wall integrity and stress resistance. FEMS Yeast Res 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.; Orlova, M.; Maziarz, M.; Kuchin, S. Protein kinase A contributes to the negative control of Snf1 protein kinase in Saccharomyces cerevisiae. Eukaryot Cell 2012, 11, 119–128. [Google Scholar] [CrossRef]
- Nehlin, J.O.; Ronne, H. Yeast MIG1 repressor is related to the mammalian early growth response and Wilms' tumour finger proteins. EMBO J 1990, 9, 2891–2898. [Google Scholar] [CrossRef]
- Nicastro, R.; Tripodi, F.; Gaggini, M.; Castoldi, A.; Reghellin, V.; Nonnis, S.; Tedeschi, G.; Coccetti, P. Snf1 Phosphorylates Adenylate Cyclase and Negatively Regulates Protein Kinase A-dependent Transcription in Saccharomyces cerevisiae. J Biol Chem 2015, 290, 24715–24726. [Google Scholar] [CrossRef]
- Hu, G.; Cheng, P.Y.; Sham, A.; Perfect, J.R.; Kronstad, J.W. Metabolic adaptation in Cryptococcus neoformans during early murine pulmonary infection. Mol Microbiol 2008, 69, 1456–1475. [Google Scholar] [CrossRef] [PubMed]
- Pukkila-Worley, R.; Gerrald, Q.D.; Kraus, P.R.; Boily, M.J.; Davis, M.J.; Giles, S.S.; Cox, G.M.; Heitman, J.; Alspaugh, J.A. Transcriptional network of multiple capsule and melanin genes governed by the Cryptococcus neoformans cyclic AMP cascade. Eukaryot Cell 2005, 4, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Williamson, P.R. Role of laccase in the biology and virulence of Cryptococcus neoformans. FEMS Yeast Res 2004, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, R.S. Iron regulatory proteins and the molecular control of mammalian iron metabolism. Annu Rev Nutr 2000, 20, 627–662. [Google Scholar] [CrossRef] [PubMed]
- Do, E.; Park, M.; Hu, G.; Caza, M.; Kronstad, J.W.; Jung, W.H. The lysine biosynthetic enzyme Lys4 influences iron metabolism, mitochondrial function and virulence in Cryptococcus neoformans. Biochem Biophys Res Commun 2016, 477, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Do, E.; Hu, G.; Caza, M.; Oliveira, D.; Kronstad, J.W.; Jung, W.H. Leu1 plays a role in iron metabolism and is required for virulence in Cryptococcus neoformans. Fungal Genet Biol 2015, 75, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Do, E.; Jung, W.H. Lipolytic enzymes involved in the virulence of human pathogenic fungi. Mycobiology 2013, 41, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martos, P.; Marin, P.; Hernandez-Molina, J.M.; Garcia-Agudo, L.; Aoufi, S.; Mira, J. Extracellular enzymatic activity in 11 Cryptococcus species. Mycopathologia 2001, 150, 1–4. [Google Scholar] [CrossRef]
- Kraus, P.R.; Boily, M.J.; Giles, S.S.; Stajich, J.E.; Allen, A.; Cox, G.M.; Dietrich, F.S.; Perfect, J.R.; Heitman, J. Identification of Cryptococcus neoformans temperature-regulated genes with a genomic-DNA microarray. Eukaryot Cell 2004, 3, 1249–1260. [Google Scholar] [CrossRef]
- Lee, I.R.; Chow, E.W.; Morrow, C.A.; Djordjevic, J.T.; Fraser, J.A. Nitrogen metabolite repression of metabolism and virulence in the human fungal pathogen Cryptococcus neoformans. Genetics 2011, 188, 309–323. [Google Scholar] [CrossRef]
- Fernandes, J.D.; Martho, K.; Tofik, V.; Vallim, M.A.; Pascon, R.C. The Role of Amino Acid Permeases and Tryptophan Biosynthesis in Cryptococcus neoformans Survival. PLoS One 2015, 10, e0132369. [Google Scholar] [CrossRef] [PubMed]
- Kingsbury, J.M.; McCusker, J.H. Threonine biosynthetic genes are essential in Cryptococcus neoformans. Microbiology (Reading) 2008, 154, 2767–2775. [Google Scholar] [CrossRef] [PubMed]
- Wong, F.H.; Chen, J.S.; Reddy, V.; Day, J.L.; Shlykov, M.A.; Wakabayashi, S.T.; Saier, M.H., Jr. The amino acid-polyamine-organocation superfamily. J Mol Microbiol Biotechnol 2012, 22, 105–113. [Google Scholar] [CrossRef]
- Mikros, E.; Diallinas, G. Tales of tails in transporters. Open Biol 2019, 9, 190083. [Google Scholar] [CrossRef] [PubMed]
- Wyman, C.L. Analysis of Membrane Transporter Systems Expressed during Symbiotic Nitrogen Fixation in the Model Legume Medicago truncatula; West Virginia University, 2018. [Google Scholar]
- Calvete, C.L.; Martho, K.F.; Felizardo, G.; Paes, A.; Nunes, J.M.; Ferreira, C.O.; Vallim, M.A.; Pascon, R.C. Amino acid permeases in Cryptococcus neoformans are required for high temperature growth and virulence; and are regulated by Ras signaling. PLoS One 2019, 14, e0211393. [Google Scholar] [CrossRef] [PubMed]
- Martho, K.F.; de Melo, A.T.; Takahashi, J.P.; Guerra, J.M.; Santos, D.C.; Purisco, S.U.; Melhem, M.S.; Fazioli, R.D.; Phanord, C.; Sartorelli, P.; et al. Amino Acid Permeases and Virulence in Cryptococcus neoformans. PLoS One 2016, 11, e0163919. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Khanal Lamichhane, A.; Bradley, J.; Rodgers, L.; Ngamskulrungroj, P.; Kwon-Chung, K.J. Differences between Cryptococcus neoformans and Cryptococcus gattii in the Molecular Mechanisms Governing Utilisation of D-Amino Acids as the Sole Nitrogen Source. PLoS One 2015, 10, e0131865. [Google Scholar] [CrossRef]
- Eigenheer, R.A.; Jin Lee, Y.; Blumwald, E.; Phinney, B.S.; Gelli, A. Extracellular glycosylphosphatidylinositol-anchored mannoproteins and proteases of Cryptococcus neoformans. FEMS Yeast Res 2007, 7, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Collopy-Junior, I.; Esteves, F.F.; Nimrichter, L.; Rodrigues, M.L.; Alviano, C.S.; Meyer-Fernandes, J.R. An ectophosphatase activity in Cryptococcus neoformans. FEMS Yeast Res 2006, 6, 1010–1017. [Google Scholar] [CrossRef]
- Vu, K.; Tham, R.; Uhrig, J.P.; Thompson, G.R., 3rd; Na Pombejra, S.; Jamklang, M.; Bautos, J.M.; Gelli, A. Invasion of the central nervous system by Cryptococcus neoformans requires a secreted fungal metalloprotease. mBio 2014, 5, e01101–e01114. [Google Scholar] [CrossRef]
- Homer, C.M.; Summers, D.K.; Goranov, A.I.; Clarke, S.C.; Wiesner, D.L.; Diedrich, J.K.; Moresco, J.J.; Toffaletti, D.; Upadhya, R.; Caradonna, I.; et al. Intracellular Action of a Secreted Peptide Required for Fungal Virulence. Cell Host Microbe 2016, 19, 849–864. [Google Scholar] [CrossRef] [PubMed]
- Brueske, C.H. Proteolytic activity of a clinical isolate of Cryptococcus neoformans. J Clin Microbiol 1986, 23, 631–633. [Google Scholar] [CrossRef]
- Yoo Ji, J.; Lee, Y.S.; Song, C.Y.; Kim, B.S. Purification and characterisation of a 43-kilodalton extracellular serine proteinase from Cryptococcus neoformans. J Clin Microbiol 2004, 42, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.C.; Blank, E.S.; Casadevall, A. Extracellular proteinase activity of Cryptococcus neoformans. Clin Diagn Lab Immunol 1996, 3, 570–574. [Google Scholar] [CrossRef]
- Ruma-Haynes, P.; Brownlee, A.G.; Sorrell, T.C. A rapid method for detecting extracellular proteinase activity in Cryptococcus neoformans and a survey of 63 isolates. J Med Microbiol 2000, 49, 733–737. [Google Scholar] [CrossRef]
- Leone, R.; Cabeli, P.; Sinicco, A.; Ito-Kuwa, S.; Aoki, S.; Vidotto, V. Relationship between protease production and capsule size in Cryptococcus neoformans. Journal De Mycologie Medicale 1999, 9, 42–44. [Google Scholar]
- Clarke, S.C.; Dumesic, P.A.; Homer, C.M.; O'Donoghue, A.J.; La Greca, F.; Pallova, L.; Majer, P.; Madhani, H.D.; Craik, C.S. Integrated Activity and Genetic Profiling of Secreted Peptidases in Cryptococcus neoformans Reveals an Aspartyl Peptidase Required for Low pH Survival and Virulence. PLoS Pathog 2016, 12, e1006051. [Google Scholar] [CrossRef]
- Monari, C.; Pericolini, E.; Bistoni, G.; Cenci, E.; Bistoni, F.; Vecchiarelli, A. Influence of indinavir on virulence and growth of Cryptococcus neoformans. J Infect Dis 2005, 191, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Sidrim, J.J.; Perdigao-Neto, L.V.; Cordeiro, R.A.; Brilhante, R.S.; Leite, J.J.; Teixeira, C.E.; Monteiro, A.J.; Freitas, R.M.; Ribeiro, J.F.; Mesquita, J.R.; et al. Viral protease inhibitors affect the production of virulence factors in Cryptococcus neoformans. Can J Microbiol 2012, 58, 932–936. [Google Scholar] [CrossRef]
- Pinti, M.; Orsi, C.F.; Gibellini, L.; Esposito, R.; Cossarizza, A.; Blasi, E.; Peppoloni, S.; Mussini, C. Identification and characterization of an aspartyl protease from Cryptococcus neoformans. FEBS Lett 2007, 581, 3882–3886. [Google Scholar] [CrossRef]
- Singh, A.; Panting, R.J.; Varma, A.; Saijo, T.; Waldron, K.J.; Jong, A.; Ngamskulrungroj, P.; Chang, Y.C.; Rutherford, J.C.; Kwon-Chung, K.J. Factors required for activation of urease as a virulence determinant in Cryptococcus neoformans. mBio 2013, 4, e00220–e00213. [Google Scholar] [CrossRef] [PubMed]
- Cox, G.M.; Mukherjee, J.; Cole, G.T.; Casadevall, A.; Perfect, J.R. Urease as a virulence factor in experimental cryptococcosis. Infect Immun 2000, 68, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, M.A.; Noverr, M.C.; Chen, G.H.; Toews, G.B.; Cox, G.M.; Perfect, J.R.; Huffnagle, G.B. Urease expression by Cryptococcus neoformans promotes microvascular sequestration, thereby enhancing central nervous system invasion. Am J Pathol 2004, 164, 1761–1771. [Google Scholar] [CrossRef] [PubMed]
- Bava, A.J.; Negroni, R.; Bianchi, M. Cryptococcosis produced by a urease negative strain of Cryptococcus neoformans. J Med Vet Mycol 1993, 31, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Ruane, P.J.; Walker, L.J.; George, W.L. Disseminated infection caused by urease-negative Cryptococcus neoformans. J Clin Microbiol 1988, 26, 2224–2225. [Google Scholar] [CrossRef]
- Almeida, F.; Wolf, J.M.; Casadevall, A. Virulence-Associated Enzymes of Cryptococcus neoformans. Eukaryot Cell 2015, 14, 1173–1185. [Google Scholar] [CrossRef] [PubMed]
- Cazin, J., Jr.; Kozel, T.R.; Lupan, D.M.; Burt, W.R. Extracellular deoxyribonuclease production by yeasts. J Bacteriol 1969, 100, 760–762. [Google Scholar] [CrossRef]
- Cox, G.M.; Harrison, T.S.; McDade, H.C.; Taborda, C.P.; Heinrich, G.; Casadevall, A.; Perfect, J.R. Superoxide dismutase influences the virulence of Cryptococcus neoformans by affecting growth within macrophages. Infect Immun 2003, 71, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.C.; Pirofski, L.A.; Casadevall, A. Extracellular proteins of Cryptococcus neoformans and host antibody response. Infect Immun 1997, 65, 2599–2605. [Google Scholar] [CrossRef]
- Moyrand, F.; Janbon, G. UGD1, encoding the Cryptococcus neoformans UDP-glucose dehydrogenase, is essential for growth at 37 degrees C and for capsule biosynthesis. Eukaryot Cell 2004, 3, 1601–1608. [Google Scholar] [CrossRef]
Figure 1.
An aggregate of cryptococcal cells forms a matured biofilm unit inside which are numerous fungi enveloped with a thin film coat of extracellular protective sheath. The sheath consists of polymeric molecules of protein, chitins, nucleic acids, β-1,3-glycans, capsular components, and lipids derivatives. Check Supplementary 4 for as many details as possible for the transcription factor, kinase, protein, enzyme, and gene acronyms used in this review.
Figure 1.
An aggregate of cryptococcal cells forms a matured biofilm unit inside which are numerous fungi enveloped with a thin film coat of extracellular protective sheath. The sheath consists of polymeric molecules of protein, chitins, nucleic acids, β-1,3-glycans, capsular components, and lipids derivatives. Check Supplementary 4 for as many details as possible for the transcription factor, kinase, protein, enzyme, and gene acronyms used in this review.
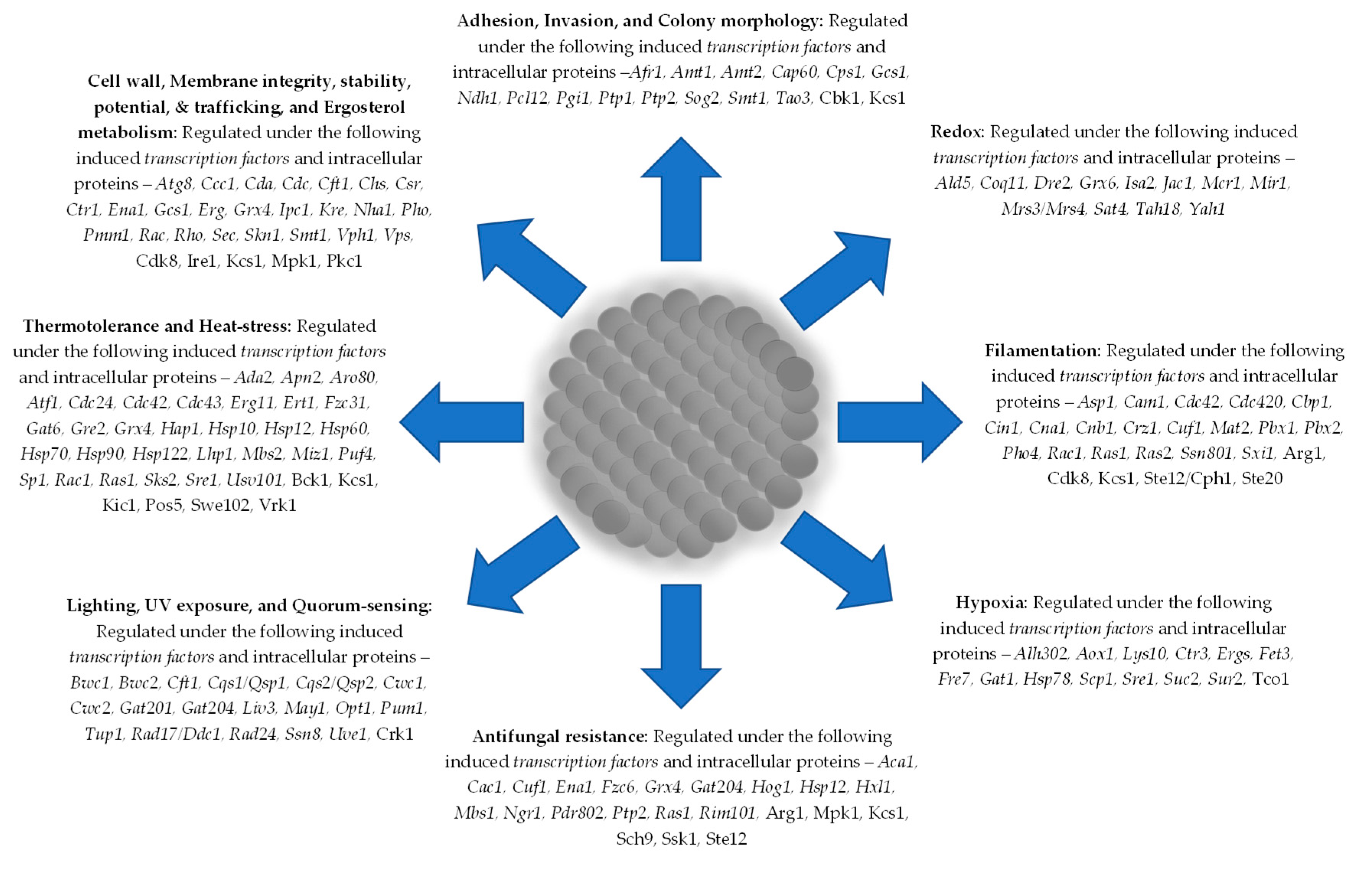
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



