
Submitted:
26 January 2023
Posted:
30 January 2023
You are already at the latest version
Abstract
The extracellular matrix (ECM) is a ubiquitous member of the body and is key to the maintenance of tissues and organs integrity. Initially thought to be a by-stander in many cellular processes, the extracellular matrix has been shown to have diverse components that regulate and activate many cellular processes and ultimately influence cell phenotype. Importantly, the ECM composition, architecture, and stiffness/elasticity influence cellular phenotypes. Under normal conditions and during development, the synthesized ECM is constantly undergoing degradation and remodeling processes via the action of matrix proteases that maintain tissue homeostasis. In many pathological conditions including fibrosis and cancer, the ECM synthesis, remodeling, and degradation is dysregulated causing its integrity to be altered. Both physical and chemical cues from the ECM are sensed via receptors including integrins and play key roles in driving cellular proliferation and differentiation and in various disease progression such as cancers. Advances in ‘omics’ technologies have seen an increase in studies focussing on bi-directional cell-matrix interactions and here we highlight emerging knowledge on the role played by the ECM during normal development and in pathological conditions. This review summarizes current ECM-targeted-therapies that can modify tumor ECM to overcome drug resistance and better cancer treatment.
Keywords:
Extracellular matrix
; Tissues
; Organs
; Development
; Tumor progression
; Collagens
; Fibronectin
; Integrins
; Metastasis
; Matrix metalloproteases
; Cell Adhesion
; Signaling
Introduction
Tissues and organs in the human body are composed of cells, biomolecules as well as the extracellular matrix [1]. The extracellular matrix (ECM) is key in many developmental stages from embryogenesis to adult development, tissue repair as well as the maintenance of tissue and organ homeostasis [1,2]. Once synthesized in the cytoplasm, ECM components are secreted into the extracellular space where they are then modified further into final molecules [1,2]. The main recognized function of the ECM is provision of physical support for cells within tissues and organs as well as availing biomolecules such as growth factors and cytokines to cells. Recent reports indicate that the ECM is involved in activation of several mechano-sensitive signaling cascades and therefore impact several cellular processes [3,4,5,6,7]. The two forms of the ECM are the interstitial ECM and the basement membrane. This review mainly focusses on the interstitial matrix.
The ECM is made up of several components that bond to form a complex network of different size molecules in a 3-D unit. These ECM molecules are of different sizes, shapes, and spatial organisation. Most tissues and organs have a specific type of ECM produced because of differential expression of ECM genes as well as post-transcriptional splicing and -translational modifications [8,9,10,11]. The ECM is in most cases in a state of flux, changing over time because of tissue development and disease [12,13,14]. Recent reports indicate the ECM play major roles in disease progression and development of chemoresistance [15,16,17,18]. Whilst cells synthesize the ECM, the ECM has been referred to as the ‘theatre’ within which cells interact with each other and biomolecules to effectively determine how cells behave [17,19,20]. Thus, cellular functions and phenotypes rely not just on gene expression but also on cues from the ECM.
ECM remodeling under normal physiological conditions is a tightly controlled complex process, with many proteins playing different roles to maintain homeostasis. The ECM also undergoes remodeling during tumorigenesis, with several reports indicating that it can promote tumorigenesis as well as being anti-tumorigenic [21,22,23,24]. In the early stages of tumor formation, stromal cells synthesize large amounts of ECM proteins in a bid to protect normal tissue from tumor cells [17,25,26,27]. This results in stiffening of the tissue around the newly formed tumor. The stiffening of the ECM is due to enhanced collagen as well as hyaluronic acid deposition [28,29,30,31]. Chronic insult to tissue results in enhanced synthesis of ECM proteins leading to a ‘fibrotic’ condition. Reports indicate that enhanced ECM deposition is positively correlated with tumor initiation and growth [32,33,34,35]. Circulating tumor cells have been shown to hone and colonize tissues and organs displaying increased ECM synthesis [36,37,38,39]. Both ECM proteins and the biomolecules found within it have been identified as valuable markers for diagnostic analysis of tumors [40,41,42]. This review discusses ECM composition, function and remodeling processes and present evidence of several ECM components suggested as novel therapeutic targets and currently being investigated or undergoing validation [43,44,45,46,47].
The Extracellular Matrix Macromolecules
The macromolecules found within tissues as well as organs that surround cells and provide tensile strength and other cues is what is termed the extracellular matrix. Various ‘omics’ studies have comprehensively identified ECM components, referred to as the ‘matrisome’ and above 200 genes have been assigned in humans [48,49]. The macromolecules form a fibrillar network that interact with cells and biomolecules to influence cell behaviour in tissues and organs. The exact number of extracellular matrix macromolecules in the human body is unknown. Two major classes of the ECMs are known, tissue-specific ECM and interstitial ECM. The type and composition of the ECM varies depending on several factors including the tissue and organ of the body. There are several classes of the ECM macromolecules including fibrillar collagens, filament-forming collagens to glycoproteins. Other classes include elastic proteins as well as proteoglycans. Important classes include the collagens that constitute the connective tissue. Under normal physiological conditions, the ECM is highly organized into sheets that confer tensile strength to tissues and organs. However, the ECM composition may differ under conditions such as stress and diseases. The ECM also provides cues to cells via tethered biomolecules and ligands to effectively influence cell behaviour [50,51].
Collagens
The collagen family of proteins is the major component of the ECM and provides both mechanical strength and cues to cells and tissues. Reports indicate that collagens constitute around 90% of the ECM in humans [41,49]. Thus, collagens influence many cellular processes in the body including proliferation, migration and adhesion [52]. Currently, about 28 proteins have been identified to belong to the collagen family [53]. Being the major proteins in the ECM, collagens undergo multiple changes and remodeling throughout an animal’s growth and development and in pathological conditions such as wound healing and cancers [54,55,56,57]. In addition, synthesis of collagens require modifications through addition of di-sulfide bonds and other post-translational changes (Figure 1) [58,59]. Other ECM molecules also play a role in collagen synthesis and deposition. For example, the glycoprotein fibronectin is known to play a part and influence the deposition and attachment of collagens in the extracellular space [60,61]. The overall structure and organization of the ECM is therefore a result of the interaction between its constituents including collagens, glycoproteins and other molecules [17,18,60,62,63,64]. Seven collagens have been grouped in the fibrillar class with type I collagen (or collagen type I) being a major component of this class. The other members include type II, type III, type V, type XI, type XXIV and type XXVII collagens [10,19,52,53]. Most collagens that form part of the basement membrane are grouped in the network-forming collagen class and these include type IV, type VIII, type X, type XV and type XVIII collagens [53]. Type VI and type XXVI collagens form the filament-forming class. The triple helical structure of some fibril-linked collagens can be interrupted and these include type IX, type XII, type XIV, type XVI, type XIX, and type XXII collagens [53]. Other collagens family members are found within or bound to membranes and these include type XIII, type XVII, type XXII, type XXIII and type XXV collagens [53].
Many studies have shown a link between changes in deposition and amounts of collagens including type I collagen with impaired development and development of cancers [32,65,66]. Collagens found within the ECM in normal tissues can be highly uniform in orientation whilst in pathological conditions the orientation is varied [67,68]. Overall, the amounts of the different collagens in the ECM influence its properties from elasticity to availability of biomolecules such as growth factors and chemokines [69,70]. Collagens within the ECM also play other important roles within the body. For example, collagens are important within basement membranes where they contribute towards separation of different layers of tissues. Increased collagen deposition within basement membranes can lead to membrane hardening disrupting normal exchange of biomolecules and movement of cells [71,72,73]. In many pathological conditions such as cancer, basement membranes are thinner compared to normal tissues. This has been attributed to less deposition of collagens including type IV, type XV and type XIX collagens [74,75,76]. Indeed, several in vitro studies have also shown that collagen knockdown can enhance migration of cancer cells [17,77,78,79].
Other Extracellular Matrix macromolecules
A combination of proteins and carbohydrates make up glycoproteins and proteoglycans, with about 30 genes encoding for these ECM components. The carbohydrates form repeating chains that are connected to a core made up of proteins. Proteoglycans are part of the glycoprotein family but different from other glycoproteins in terms of their synthesis and structure. This ultimately influence their function in the body. Whilst glycoproteins have short and branched carbohydrate chains covalently linked to a protein core, the carbohydrate chains in proteoglycans are long and unbranched glycosaminoglycan chains also attached to a protein core [80,81]. Glycoproteins side chains create enough buffer to allow the ECM to resist stress and forces applied onto the ECM [82,83]. In addition, glycoproteins are actively involved in regulating processes including proliferation and adhesion [83,84,85]. The glycosaminoglycan chains of proteoglycans also negatively charged allowing proteoglycans to impact on the organization of other ECM constituents [84,85]. The negative charge on proteoglycans also allow the ECM as a whole to sequester growth factors and other biomolecules [86,87,88]. Due to their size and structure, proteoglycans can also participate in the binding of ligands to receptors, allowing cells to respond to various changes in extracellular cues. Several signaling pathways including the AKT-MEK and PI3-Akt cascades are activated through the participation of proteoglycans in bonding to various receptors [89,90]. Most well-known glycoproteins include fibronectin, fibrinogen, vitronectin, laminin, thrombospondins, periostin and osteopontin. Among the well-known proteoglycans are decorin, aggrecan and perlecan.
Laminin
A glycoprotein consisting of α, β and γ chains that come together to form trimeric proteins, laminin or laminins is/are found within the basal lamina and contribute towards cell-specific functions including differentiation, adhesion, and migration [91,92]. Laminins as ECM glycoproteins play major roles in creating a link between the ECM and cells via binding to cellular receptors such as integrins. Thus, laminins are key to cellular migration and cancer cell invasive behavior. Currently, twelve mammalian chains (α, β and γ) have been identified and these can combine in different amounts to form about sixty known laminins [93,94]. The α chains (200 to 400 kDa) are bigger than the β and γ chains (120 to 200 kDa) with the trimer formed ranging from 400 to 800 kDa in size. Referred to as the ‘god molecule’ in some reports, the trimeric laminin has a ‘cross’ shape formed as a result of its α-helical coiled coil structure [95,96]. Laminins also bind to other ECM components including collagen type IV. In this case laminins acts as intermediary or ‘glue’ between various ECM molecules within the basement membrane. Laminin polymerization is thought to be the main initiator of basement membrane assembly, placing laminin polymerization at the ‘center’ of cell function and tissue structure.
A well-known laminin molecule is Laminin-332 (LN-332), formed from β3, α3 and γ2 chains play key roles in cellular migration, adhesion and contributes towards tumor cell metastasis [91,92,97,98]. Laminin molecules are also implicated in maintaining stem cell self-renewal capabilities. For example, laminin-332 maintains CSCs self-renewal abilities and contribute towards drug resistance [98]. Several reports show that the presence of laminins is closely linked to significantly lower patients survival in cancers such as colorectal and pancreatic cancer [97,99]. Laminins bind to other ECM proteins and this promotes cell migration and adhesion as well as enhancing drug resistance [100,101]. For example, the binding of laminin 332 to integrin α3β1 receptor increases resistance to gefitinib in hepatocellular carcinoma [102]. Various signaling cascades are also known to be activated through laminin-integrins interactions. For example, laminins interactions with integrins cause activation of the mTOR cell surviving signaling pathway [98,103].
Fibronectin
Structurally, fibronectin (FN) has several domains and is involved in the interactions between the ECM and cells. Fibronectin forms a fibrillar network and is key to cell differentiation, adhesion and migration [104]. Fibronectin exists as a dimer of two molecules joined together via cysteine disulfide bonds. Assembly of fibronectin into the ECM occurs when it binds to α5β1 integrins via the RGD motif. Furthermore, the binding of fibronectin to integrins causes clustering of integrin molecules leading to increased levels of fibronectin molecules on cell surface. Fibronectin-focal adhesion interactions alters the conformation of fibronectin resulting in binding sites for other ECM molecules to be revealed. Fibronectin is therefore able to bind to collagens, laminins and other proteins, allowing cells to adhere to the ECM and migrate [104]. Whilst it is a single gene encoded protein, it has several isoforms resulting in proteins that form ECM fibrillar structures. Fibronectin binds to cell surface receptors and other ECM proteins such as collagens causing the alterations of the cells’ actin filaments and this allows cells to migrate. Various reports show that fibronectin is key in cellular processes such as wound healing as well as in tumor growth [105]. Importantly, the adhesion of tumor cells to ECM proteins including fibronectin enhances the tumorigenic capacity of cancer cells as well as drug resistance [106,107]. Various studies have also associated increased fibronectin expression to tumor progression in various cancers [108,109,110]. Furthermore, clinical data associated enhanced fibronectin expression in tumors versus normal tissues with lower patients’ survival [105,111,112]. FN-induced migration was shown to be mediated via αvβ6 and α9β1 integrins in various cancers [105,113,114]. The binding of cancer cells to ECM proteins including fibronectin can protect cells from drug-induced apoptosis compared to cells attached to plastic [106]. Fibronectin-mediated reduction in apoptosis occurs via inducing the cyclooxygenase-2 (COX-2) as well as the activation of integrin α5β1 [115,116]. In addition, various signaling cascades are activated when fibronectin binds to other ECM proteins [117]. The binding of cells to fibronectin also protects cells against many drug-induced [98,117,118].
Periostin
Periostin is an adhesion-linked protein expressed as an ECM protein and produced within the periosteum as well as the periodontal ligaments [119,120]. It is a cell adhesion and non-structural protein that function to maintain tissue homeostasis especially that of the tooth and bone tissues. It is mostly involved in many processes during development including cardiac development and healing but is expressed in low amounts in adult tissues [91,119,120]. Periostin mediate most of its effects via interacting with surface receptors such as integrins. The enhanced expression of periostin is associated with various pathological conditions including inflammatory disease, many types of cancer including colon, lung, breast and head and neck carcinomas [119,120]. Periostin is involved in regulating ECM-cell interactions via attachment to other ECM molecules including collagens, tenascin C and fibronectin [120]. Periostin can bind to various integrins such as αvβ3, αvβ5 and α6β4, and thus influence the activation of many signaling cascades [121]. Some of the signaling cascades are the Notch 1 and B-catenin signaling, that are important in cell differentiation and tissue specification. Various reports show that periostin is aberrantly expressed in pathological conditions such as arthritis, cancers, and fibrosis [120,121]. In various cancers, periostin has been shown to induces signaling cascades including PI3K-Akt through attaching to αvβ3 and αvβ5 integrins [122]. The presence of periostin enhances cancer cell proliferation and the process of EMT cancers such as gastric cancer [122,123]. Cancer cells showing resistance to various drugs including cisplatin and 5-fluorouracil (5-FU) also show increased periostin expression [120]. Thus, evidence suggest that increased periostin levels correlated with drug resistance, tumor relapse and tumor angiogenesis [124]. Periostin activates the Akt phosphorylation in cancers including epithelial ovarian carcinoma and this results in resistance, especially to paclitaxel [125]. Recent data suggest that periostin can be used as a prognostic marker in various cancers including pancreatic, ovarian and esophageal cancers [126,127,128].
Hyaluronic acid
Discovered almost a century ago by Karl Meyer and John Palmer whilst working on vitreous of bovine eyes, hyaluronic acid is a glycosaminoglycan made up of N-acetylglucosamine and glucuronic acid repeats and is a common component of the ECM [129,130]. Hyaluronic acid is a long high molecular weight polymer with many hydroxyl moieties, allowing it to mix well in water [131,132]. Indeed, one of the main functions of hyaluronic acid is to retain water in various tissues [133,134]. Due to its size and the ability to form coils in water, hyaluronic acid can control the movement of biomolecules and ions within the ECM, allowing small molecules to pass whilst blocking the free movement or transport of larger biomolecules and substances [135]. Hyaluronic acid display various unique properties including being biodegradability and great viscoelasticity, and has been utilized in various application such as hydrogel formation and drug delivery systems [136,137].Various studies have shown that hyaluronic acid play crucial roles in cell migration and invasion through its interaction with receptors including CD44 and hyaluronan binding protein 4 [138,139,140].
Extracellular Matrix Function
The most important function of the ECM is providing an anchorage stage to cells as this is key to the maintenance of cell division and polarity. No longer is the ECM seen as only a scaffold necessary for cell structure, but it provides both biophysical and biochemical cues to cells. In addition, the ECM can regulate cellular attachment and migration [141]. Several pieces of evidence also show that the ECM can sequester growth factors and other biomolecules, and these are released at specific stages of development and disease progression to influence cell behaviour and phenotype [18,142,143]. During development sequestered factors can cause gradients in biomolecules concentrations and this is important during change in tissue form and structure [143,144]. Furthermore, secreted factors are involved in the activation of various signaling cascades and influence focal adhesion formations [145,146].
The development of an organism from an embryo to an adult involves a lot of ECM changes, both in terms of quantity and type [147,148]. These changes must be controlled tightly at each stage of development to avoid over- and down-regulation which can have deleterious effects. ECM physical properties including topography, elasticity and rigidity influence cell proliferation and differentiation and ultimately influence tissue structure and integrity [7,62,63].
An important function of the ECM is aiding cell migration. For cells to migrate, binding to the ECM via integrins and cadherins must occur first. Integrins allows cells to attach to various ECM molecules including collagens, fibronectin, and laminins. Integrins can then influence intracellular actin cytoskeleton via the focal adhesion proteins including talins and vinculins. The ECM alignment and topography have been shown to influence both the speed of cell migration and the direction of migration [149,150,151]. To influence cell migration in a specific direction, scientists have utilized specific ECM molecules as well as ECM gradients [148,152]. Studies have shown that cells tend to migrate from low ECM concentration areas to areas of high ECM concentration, but this is not always the case [149,153,154]. Migration of cells is characterized by repeated adhesion to the ECM as well as de-adhesion from the ECM [155]. Therefore, the rate of cellular migration is dependent on various ECM properties including composition, alignment and elasticity [156,157,158]. Our earlier publication demonstrated that cells migrate slower on ECMs lacking collagens compared to those on collagen containing-ECMs [17]. ECM stiffness for example has been shown to influence cellular migration [159,160]. Investigations are under way to identify specific ECM components required for cell differentiation and migration during development as the ECM is constantly being remodeled. Importantly, for cells to migrate and invade surrounding tissues is the action of matrix metalloproteases and other proteases that can degrade the ECM [161,162,163].
The development of organs and tissues in the body is dependent on the presence of ECM proteins. For example, for the process of branching to occur, ECM proteins such as collagens and laminins are required to provide the anchor on which the formation of tubes can take place whilst ECM molecules such as hyaluronic acid allow epithelial cells to continue to migrate at the end bud [164,165]. The ECM’s alignment and architecture help in controlling tissue formation and branching patterns [164,166]. When a bud continues to grow, the ECM at the end is degraded, allowing various biomolecules including growth factors, cytokines and chemokines to be released [161,167,168]. The released factors in turn influence the rate of branching and the direction of the bud [161,169]. The presence of the ECM at the end bud also means that various growth factors and signaling molecules can be sequestered and released at specific times when needed [83,170]. Whilst ECM proteins are important during budding, specific ECM are required to stop the growth of the bud. For example, deposition of type I collagen leads to termination of growth of the bud [164]. In summary, the ECM plays major roles during tissue and organ formation as well as formation of tubular structures. Importantly, beside the provision of a scaffolding on which tissues and organs can form, the ECM act as a reservoir of various biomolecules needed by cells at specific times for differentiation and proliferation.
Various studies have shown that the ECM cellular differentiation through release of tethered factors as well as its physical structure and composition [16,171,172,173]. To study the effect of ECM composition on cell fate, we cultured fibroblasts on a fibroblast-derived ECM (fd-ECM) and showed that these fibroblasts downregulate type I collagen synthesis compared to controls [62]. Via the use of function-blocking antibodies, our study demonstrated that blockage of type I collagen gene expression in the presence of the ECM is mediated via integrins including α2β1 [62]. In addition, the same study revealed that ECM-mediated reduction in collagen was activated through the Ras-MEK/ERK signaling pathway [62]. Importantly, through deletion analysis of the COL1A2 promoter, our study showed the presence of a ECM-responsive element within the -375 and -107 region [62]. This study and other published reports demonstrate that the ECM composition play pivotal roles in determining cellular gene expression and function [174,175]. Using a different approach, we also investigated adipose-derived MSCs (ad-MSCs) fate when cultured on a cell-derived ECM (cd-ECM). The use of the cd ECM in this case was done so as to model the in vivo physiological microenvironment. Our data showed that ad-MSCs cultured on the ECM lost their multipotency and differentiated into the chondrogenic cellular lineage compared to controls [7]. Elaborate studies including loss of function studies showed that cells are able to sense the mechanical environment and activate the Notch1 and β-catenin signaling cascades needed for the ECM-mediated ad-MSCs chondrogenesis [7]. Thus, ad-MSCs must sense the mechanical environment of the ECM in order to undergo chondrogenesis, proving that the ECM has the capability to induce differentiation of cells.
The process of maintaining tissue homeostasis require that the ECM be constantly altered as cells undergo proliferation and differentiation [176,177]. Once differentiated, cells must maintain their phenotypes whilst changes to tissues occur including formation of new blood vessels [178,179]. A bi-directional interaction exists between cells and the ECM. In a classic study of the interaction between cells and the ECM, we demonstrated that cells can ‘feel’ the presence of ECM components and other ECM properties through integrins including α2β1 and adjust ECM proteins’ gene expression based on cues from the ECM [16,63]. Thus, a feedback process allows communication between the ECM and cells and is important to maintaining tissue homeostasis. Once this feedback mechanism is altered and unable to maintain ECM degradation and deposition in check, conditions such as cancer can develop. Other ECM properties including elasticity also influence cell behaviour. For example, Zhang and colleagues showed that the ECM elasticity impact on osteocyte gap junction elongation and also demonstrated the involvement of paxillin in signal transduction [180].
Various techniques have been utilized to study the role of the ECM during development and in tumorigenesis. The most common technique involved ECM gene knockout, downregulation, and upregulation, leading to alteration of ECM composition [181,182]. Addition of enzymes that can degrade specific ECM proteins can also be used to alter ECM composition and reveal the role os specific proteins in various cellular processes [183,184,185]. Antibodies against ECM proteins and their respective receptors including integrins can be used to investigate the role of ECM proteins in development and maintenance of homeostasis. In our earlier publication, we showed that function-blocking antibodies to downregulate collagen gene expression can reveal that type I collagen interact with α2β1 integrin [57,62]. Other techniques that can be used to study ECM proteins’ role in development and tumorigenesis include the use of 3-dimensional culture, atomic force microscopy and utilizing ECM proteins crosslinking [7,186,187,188].
Extracellular Matrix Modifications
The composition of the ECM in a specific location or tissue is influenced by several parameters such as the synthesis and degradation of its components. In addition, the biomolecules found within the ECM is location or tissue-specific and is influenced to a large extent by resident stromal cells such as fibroblasts and immune cells. Further changes in ECM composition and the biomolecules found within it are brought about by the action of enzymes including matrix metalloproteases and hydroxylases [189,190,191]. Studies in various cancers including breast and bladder cancers have shown that post-translational hydroxylation of collagens leads to increased crosslinking and is linked to low patients survival [192,193]. Crosslinking of the ECM components, sulfation as well as glycosylation are some of the further changes that occur to ECM components after synthesis [189,190,191]. These post-translational changes impact ECM components interactions with other members of the ECM as well as with receptors on cell surfaces [7,62,63]. Whilst changes in ECM composition and sequestered biomolecules are necessary for tissue homeostasis, the state of lax is also necessary for development and growth. Altered synthesis and accumulation of any one component of the ECM can alter the existing delicate homeostasis and lead to conditions including fibrosis and promotion of cancer growth [194,195,196,197]. The contribution of stromal cells and immune cells has been recognized as key to the maintenance of homeostasis and the development of several pathological conditions [17,18,198,199,200].
Reports indicate that when there is enhanced crosslinking of the ECM, a dense meshwork of ECM components is formed leading to fibrosis and other pathological conditions [33,178,201,202]. Importantly, accumulation of ECM proteins leads to stiffening which influence ECM-receptor interactions and cellular signaling [33,203,204]. Excessive crosslinked ECM proteins also lead to reduced ECM turnover, allowing some ECM proteins to prolong their presence within certain tissues. For example, ECM proteins known to promote wound healing via their participation in certain stages of the process may prolong their presence around the wound leading to aberrant process [205,206,207]. Various enzymes are known to take part in ECM crosslinking, and these include lysyl oxidases, transglutaminases [208,209]. The lysyl oxidase family of enzymes allow the deposition and accumulation of collagens and elastins in the ECM and this has significant implications for cell morphology and movement [210,211]. By influencing the deposition of collagens in the ECM, lysyl oxidases also affect cellular signaling and response to therapy [212]. Transglutaminases are enzymes involved in glutamine deamination during ECM proteins and glycoproteins synthesis [213,214,215]. Transglutaminases are also involved in transamidatig glutamine residues during ECM synthesis and the proper alignment of fibres during ECM synthesis. Alignment of fibres by transglutaminases leads to ECM stiffening and reduction of proteolytic degradation. A stiff ECM influence ECM-receptor interactions including integrin-mediated signaling [216]. Another form of spontaneous ECM crosslinking is glycation, a process including Amadori rearrangement and Schiff base adduct formation. This process does not involve enzymes.
Glycosylation of ECM molecules have been linked to various processes of tumorigenesis. For example, enhanced glycosylation of fibronectin leads to increased EMT and high levels of invasive cell behaviour in prostate cancer cells and carcinomas, respectively [217,218,219]. Inhibition of integrins glycosylation including αvβ6 integrin leads to enhanced invasive behaviour of cells involved in metastasis [220,221]. Demonstrating the importance of fibronectin as a component of the ECM, the phosphorylation of fibronectin leads to increased mechanical forces needed for cell adhesion in various cancers [222,223,224]. Glycosaminoglycans can undergo sulphation in various cancers and this impact cell-matrix signaling [225,226,227].
Proteolytic Degradation of the Extracellular Matrix
Homeostasis involves the constant synthesis and degradation of ECM components over time [178,228]. Enhanced or reduced synthesis and degradation beyond what is normal can lead to several pathological states including fibrosis. Many enzymes are involved in both the synthesis and degradation of the ECM. Importantly, ECM degradation is controlled by zinc-containing endopeptidases including matrix metalloproteinases and a disintegrin and metalloproteinase proteins with thrombospondin motifs (ADAMTSs) [229,230]. Excessive ECM degradation can lead to an abnormal ECM characterized by unbalanced ECM components or ECM components not crosslinked properly [231,232]. More degradation than ECM synthesis can result in removal of whole tissue components such as basement membranes and the vasculature, allowing cells to migrate in an uncontrolled manner [233,234,235]. Growth factors and cytokines previously bound to ECM proteins can increase locally due to excessive ECM degradation, leading to unregulated activation of signaling cascades [17,51,236]. For example, TGF-b and VEGF have been shown to be released from degraded ECM proteins through the action of MMPs, leading to activation of various signaling cascades and angiogenesis [18,237,238,239]. Inorganic ions including calcium ions can also be released from the degraded ECM leading to the activation of calcium-dependent MMPs [240,241].
One major class of enzymes involved in ECM degradation is the matrix metalloproteinases (MMPs). As reviewed by Kesenbrock and colleagues, human MMPs are a total of 23 enzymes with a Zn-containing domain as well as four haemopexin-like domains [242]. Four MMP members namely MMP14, MMP15, MMP16 and MMP24 contain both a transmembrane and cytoplasmic domain. Through degrading the ECM, MMPs impact cellular process via the release of sequestered growth factors and cytokines [243]. Whilst most MMPs have a specific substrate leading to their being grouped as collagenases and gelatinases for example, many other MMPs cannot be grouped this way [199]. Due to their actions in the body, MMPs are highly regulated to ensure maintenance of homeostasis as well as allow growth and development. Unregulated expression and the eventual action of MMPs has been associated with many pathological conditions [244,245,246]. For example, MMP2 and MMP11 are associated with poor survival in ovarian cancer [247]. However, some MMPs including MMP8 have been associated with increased survival in oral squamous cell carcinoma patients [248]. The varied and sometimes opposing actions of MMPs in the body has derailed efforts to develop inhibitors for these enzymes.
There are 21 a disintegrin and metalloproteinase proteins (ADAMs) and about 19 ADAMs with thrombospondin motifs (ADAMTSs) in humans that play a role in degrading the basement membrane of vascular system vessels [249,250]. Many ECM proteins and proteoglycans including collagens, fibronectin and vitronectin are degraded by ADAMs [250]. ADAMTSs are secreted enzymes and involved in degrading proteoglycans and collagens [161]. Several signaling cascades have been shown to be influenced by Adamalysins through removal of ligands from the cell surface [251,252,253].
Cathepsins are a family of ECM degrading enzymes made up of 11 serine, cysteine and aspartic peptidases [254,255,256]. Cathepsins are mostly found in lysosomes and involved in degradation of proteins precursors such as pro-collagens within the cell [200,257,258]. Cathepsins have been implicated in altered homeostasis and many pathological conditions including scar formation and cancer development and metastasis [259,260,261,262]. On the contrary, cathepsin B has been shown to be important during tissue regeneration in wounded human epidermal keratinocytes [263].
Originally referred to as procollagen C-proteinases, bone morphogenetic protein I and tolloid-like proteinases are enzymes that play key roles in the maturation of procollagen molecules and have no known role in ECM degradation [264]. These enzymes are able to cleave the carboxy terminus of procollagens leading to maturation of the procollagen molecules into collagens [264,265]. These proteinases work in cahoots with growth factors in promoting ECM deposition during growth and development [266,267,268]. Bone Morphogenetic Protein I and Tolloid-like proteinases have been shown to be essential in skin wound healing but their upregulation is linked to corneal scarring [269,270].
Hyaluronidases are involved in the degradation of hyaluronan in the body to maintain tissue homeostasis. Hyaluronan is rapidly degraded after synthesis compared to other ECM components, and this is essential for tissue homeostasis [271,272]. Accumulation of hyaluronan as well as its increased degradation is often associated with several pathological conditions including cancers [272]. Additional enzymes, CEMIP and transmembrane protein 2, are also referred to as hyaluronidases and the increased activity of hyaluronidases can lead to the formation of hyaluronan fragments that have been linked with increased formation of blood vessels [273,274]. Hyaluronan fragments, resulting from enhanced hyaluronan degradation have also been linked to increased synthesis and release of chemokines and cytokines, leading to activation of various signaling cascades [275,276,277]. Reports indicate that hyaluronan fragments accumulate during injury and lead to increased inflammatory factors being synthesised by immune cells around a wound [278,279,280]. The persistent inflammation as a result of lack of removal of hyaluronan and its fragments can lead to promotion of tumorigenesis [281]. Dysregulated hyaluronan production disrupts normal ECM structure as well as formation of blood vessels [282]. The main hyaluronan receptor is CD44 and is involved in the removal of hyaluronan and its fragments in case of injury.
Heparin sulfate glycosaminoglycans (HSGs) are cleaved by heparinase from proteoglycans core proteins and eventually degraded into oligosaccharides [283]. HSGs are involved in ECM organization and activation of cell signaling via their binding to other ECM components as well as receptors. Heparanase has been associated with enhanced wound healing and angiogenesis in various animal models [284,285]. However, its overexpression has been linked to other pathological conditions including cancers [286,287]. Disruption of normal heparinase expression causes ECM-heparan sulfate interactions to be altered leading to weak ECM structures. Cell movement can be increased under these conditions as ‘pores’ are present in the ECM and these allow cells to migrate easily.
To maintain normal ECM structure and amount, ECM degrading enzymes are tightly regulated by the action of their respective inhibitors. Inhibitors of ECM degrading enzymes are secreted by various cells and they act in both autocrine and paracrine modes. Some of the well-known inhibitors of ECM-degrading enzymes are the tissue inhibitors of metalloproteinases and cystanins. One of the well-known endogenous inhibitors of MMPs is the tissue inhibitors of metalloproteinases (TIMPs) family. In mammals this family has four members namely TIMP1, TIMP2, TIMP3, and TIMP4 [288,289,290]. TIMPs have recently been shown to inhibit the Adamalysins family of ECM proteinases, expanding their role in maintaining ECM homeostasis [290,291,292]. Disruption in TIMPs synthesis and secretion has been noted in many pathological conditions including cancers and aberrant wound healing [293,294].
The action of cathepsins is inhibited by cystatins in a reversible manner. Cystatins have both intracellular and extracellular activities and therefore influence both ECM synthesis and remodeling of mature ECM. Cystatins are also known to show inhibitory activities against papain and legumains [295]. Serine and cysteine proteases involved in the degradation of various ECM proteins and proteoglycans can be irreversible inhibited by serpins [296]. Serpins play a significant role in maintaining tissue and vascular homeostasis as well as in fibrinolysis [297]. Serpins have also been shown to play key roles in thrombosis [298]. Serpins are a large family and therefore have many contrasting roles in the mammals. Several ECM molecules contain structures called cryptic domains that release fragments called ‘matricryptins’ when the molecules are cleaved [299,300]. These ‘matricryptins’ fragments are important for cell adhesion and differentiation [161]. Due to their many functions, some ‘matricryptins’ may serve as enzyme inhibitors and abrogate proteolytic activities, thus influencing ECM synthesis and degradation [300]. One ‘matricryptin’ derived from the collagen XVIII molecule is Endostatin which has been shown to cripple the function of androgen receptor [301]. Included in the matricryptins derived from collagen are arresten, tumstatin and canstatin and these have been associated with many pathological conditions [302,303,304]. Several matricryptins have been shown to have anti-growth properties and can cause senescence in cells, but evidence also points to ‘matricryptins’ fragment from ECM molecules such as laminin 111 can promote cellular growth [305].
Fibroblasts and Extracellular Matrix Remodeling
The unregulated synthesis and deposition of extracellular matrix associated with many cancers results in fibrosis or the so-called ‘hardening’ associated with mostly advanced tumors (Figure 2) [33,306]. Whilst all stromal cells contribute towards the synthesis and deposition of the ECM, cancer associated fibroblasts are the main cells doing this job. In addition, recent reports indicate that cancer cells also synthesise and deposit ECM proteins and proteoglycans [17,18,87]. Similarities have been noted between the cancer associated fibroblasts and fibroblasts found during wound healing [307,308]. Cancer associated fibroblasts display great heterogeneity and different cells have been suggested as sources of CAFs [309,310,311]. Both local and recruited cells are potential sources of CAFs. For example, local fibroblasts can easily be recruited to growing tumors where they undergo activation to CAFs. Mesenchymal stem cells from the bone marrow as well as adipose tissue have been shown to be transformed into CAFs [18,312]. Activation of local fibroblasts and transformation of other cells into CAFs is driven by growth factors and chemokines release by both cancer cells and stromal cells [18]. Such growth factors and chemokines can be released within the vicinity of the cells or can be transported via exosomes from distant environments [313,314].
Studies show that CAFs presence and increased synthesis and deposition of ECM is linked to recurrence of disease and decreased patients’ survival [315,316,317,318]. Research into which subset of CAFs drive tumorigenesis and recurrence of disease is needed as great heterogeneity is displayed by CAFs, with some subsets of CAFs known to be involved in inflammation whilst others are myofibroblasts-like, for example [319,320]. Attempts have been made to characterize and classify CAFs, but the duly is out on the utility of such endeavours. Besides phenotypical differences displayed by CAFs, their precise location in the body and in tumors also determine the role they play in various processes. Thus, CAFs subsets release different growth factors and chemokines depending on their spatial location. Data is lacking on the changeability of the different CAFs subsets and the contribution of the different subsets to tumorigenesis and disease outcomes. Since CAFs, other stromal cells and cancer cells contribute towards the ECM and biomolecules found within tumors, it is currently difficult to assign a specific role of CAF-derived, stromal cell- and cancer-derived ECM molecules to disease progression and outcomes. Cancer cells within tumors for example have also been shown to synthesis unusual ECM proteins and proteoglycans that may play a role in fibrosis and disease progression [321,322,323].
Hyperactivation of the sympathetic nervous system due to cellular stress has been shown to be associated with remodeling of the stroma [324,325,326]. Furthermore, cellular stress has been shown to promote the synthesis and release of ECM molecules such as collagens [327]. Increased ECM synthesis is linked to development of chemoresistance in various cancers [17,328,329]. In order for tumor cells to migrate and metastasise, there is need for space and secretion of factors involved in invasion. ECM deposition has been shown to be downregulated by tumor cells via the release of colony-stimulating factor 1 [330,331,332]. Reduced ECM synthesis leads to enhanced tumor cell invasion and migration. The specific contribution of ECM molecules, from either tumor cells and stromal cells, to the progression of tumors is still a subject of intense research. In addition, the specific point when ECM molecules change from anti-tumorigenic to pro-tumorigenic and vice versa is not yet known. Another approach is to ‘normalize’ stromal cells involved in ECM synthesis, so that there is normal production of ECM [17,18].
Extracellular Matrix Signaling
Tumorigenesis leads to alterations of the ECM in terms of structure and composition, with such alterations being pro-tumorigenic [333,334,335]. Furthermore, such ECM alterations can also result in development of chemoresistance [336,337]. Cues from the ECM can be physical as well as sequestered biomolecules and these causes immediate as well as long term changes in gene expression [142,338,339]. The turnover of most ECM proteins and glycoproteins can be hours, days and weeks and thus their continued presence provides a continuous stimulus to cells, leading to activation of cellular cascades over a long time.
Whilst stromal cells and tumor cells both can release growth factors and chemokines and thus influence signaling, the ECM can enhance or decrease the resulting signaling via release of sequestered growth factors and chemokines and sequestering synthesised factors, respectively [142,334,335,336,337,338]. In addition, the stiffness of the ECM can influence integrin-based signaling during normal development and in diseases [340,341]. Reports indicate that signaling cascades including the MEK-ERK and the JNK signaling cascades can be activated by ECM stiffening in various conditions [342,343,344]. Other signaling cascades also respond to ECM composition and stiffness, thus targeting these signaling cascades together with ECM composition and stiffness are plausible strategies to control tumorigenesis and metastasis.
One of the major receptors for ECM-cell interactions is integrins. Integrins are heterodimers involved in transmitting extracellular cues into cellular signalling [16,345,346]. During development and in some pathological conditions, specific integrins are expressed and these influence specific cellular activities such as migration, proliferation and adhesion [347,348,349]. Importantly, the binding of various integrins including αvβ1, αvβ3 and α4β1 to ECM molecules has been linked with tumor cell invasion during tumorigenesis [350]. The precise expression of certain integrins may be linked to promotion of tumorigenesis, drug resistance and metastasis [351]. The enhanced expression of α5β1 and its binding to ECM molecule fibronectin has been linked to reduced drug efficacy in models of cancers [352,353]. Overall, the involvement of integrins in development and in pathological conditions such as cancers depends on the type of integrin, ECM molecules and cell type [354,355,356]. The inter-conversion of integrins has also been associated with ‘cadherin switching’ during epithelial to mesenchymal transition [357,358]. The switching or converting of integrins from one heterodimer to another is thought to be linked to their binding to different ECM ligands as well as different cells [359].
Several non-integrin receptors are known to bind to ECM components and these include heparan sulfate proteoglycans bound to the surface of cells, discoidin domain containing receptor 1 as well as leukocyte-associated immunoglobin-like receptor 1 [360,361]. Together with integrins, these receptors relay extracellular cues and signals to activate various signaling cascades. Activation of these non-integrin ECM receptors in stromal and cancer cells can enhance ECM synthesis via a feedback loop, thereby decreasing access of drugs to cancer cells [362,363,364]. For example, syndecan 4 is highly expressed in various cancers and is known to activate signaling cascades associated with cancer cell survival [365,366]. A major receptor for collagen, and hyaluronic acid is CD44. Structurally, CD44 traverse the cell membrane and has both an extracellular domain and a cytoplasmic component. The interaction of CD44 and hyaluronic acid leads to the activation of various other receptors including EGFR and c-MET [367,368]. Through activation of signalling cascades, hyaluronic acid-CD44 play a key role in tumorigenesis [369,370]. Infiltration of lymphocytes into tumors is partly mediated by the interaction of CD44 and fibronectin, which allows lymphocytes to bind to fibronectin and to other ECM components [371]. CD44 has been reported to play key roles in tumorigenesis and its overexpression is linked to poor patients survival and drug resistance [372,373]. CD44 is one of the important cancer stem cell markers in many cancers including colon cancer, breast cancer, prostate cancer and lung cancer [374,375,376,377].
External cues are sensed by receptors on the cell membrane and converted into cellular signalling to cause specific cellular behaviour. This process known as mechanotransduction is important in various cellular processes, from normal development to diseases. Mechanotransduction influences all cells within tumors, ultimately determining the progression of tumor development [378,379]. Whilst cells influence the deposition and accumulation of the extracellular matrix, the extracellular matrix properties such as mechanical and elasticity modulate cell behaviour [380,381,382]. Various signaling cascades are known to be activated by extracellular matrix-derived cues and these include the MEK-ERK, PI3K-Akt and the YAP-TAZ signaling [383,384].
Cellular behaviour is modulated not just by the amount of ECM components, but also by the biochemical properties of the ECM such as tensile strength, mechano-resistance and elasticity. These biomechanical properties affect cellular processes such as cellular migration and metabolism [385,386]. Adhesion of cells on ECM of different elasticity show that ECM elasticity influence gene expression and integrin levels on cell surfaces [387]. In addition, cues and signaling molecules released during adhesion of cells to ECM influence the organisation of the cytoskeleton and therefore affect cellular migration and invasive behaviour [388,389]. Various signaling cascades including the FAK, PI3K-Akt have been shown to modulate ECM-cellular interactions, influencing normal cell growth and movement [390,391,392]. Increase in ECM proteins including collagens have been associated with migration of cells and development of chemoresistance [393,394].
Extracellular Matrix and Cell Invasion and Metastasis
Four major features of metastasis are the migration of cancer cells from their origin, honing to new sites and the regulation of secondary sites in preparation for tumor growth, heterogeneity of cancer cells and lastly the colonization of the new sites and growth of secondary tumors (Figure 3) [19,395,396]. At each stage of the metastatic process, the ECM plays a central role and its remodeling influence the progression of the process. The alignment of ECM proteins including collagens and fibronectin have been shown to influence tumor cell metastasis in various cancers [397,398,399,400,401,402]. Linear collagen molecules appear to promote tumor cell migration as the spaces between fibres allow cancer cells to move in a certain direction [397,398,399,400]. Furthermore, the action of both tumor- and stromal cell-derived MMPs in the degradation of ECM molecules allows spaces to be created for tumor cell migration and invasion into surrounding areas [403,404,405]. ECM molecule post-translational modification processes including hydroxylation also promote metastasis by promoting enhanced synthesis of ECM molecules [192,406,407]. Due to their size and physical structures, ECM molecules can shield invading and migrating tumor cells from effects of shear stress during the tortuous journey to secondary sites [406,407,408]. One major feature of successful metastasis is the preparation of ‘new sites’ for tumor cells to colonize and grow. Various theories have been given to explain this process. For example, it is thought that tumor-derived MMPs degrade and remodel the existing ECM of the ‘new sites’ prior to tumor cells colonizing these sites [409,410]. Following the ‘seed and soil’ hypothesis, this remodeling of existing ECM in new sites is important to create the right pro-tumorigenic environments for colonization by the metastatic tumor cells [411,412]. Overall, it is accepted that the ECM play an important role in allowing metastatic tumor cells colonize new sites and be able to grow. In other instances, tumor cells do not grow into secondary tumors but remain quiescent for some time. These tumor cells can survive for a long time, re-awaken and grow into tumors after a long time [413,414,415,416,417]. For example, an upregulation of ECM molecule periostin has been shown to promote the re-awakening of breast cancer cells from dormancy [418,419].
Extracellular Matrix in Drug Resistance: The Extracellular Matrix shield tumor cells from anti-cancer drugs
Studies reveal that the ECM is a major player in tumorigenesis and treatment outcomes [187,420]. Furthermore, reports indicate that therapy itself can induce ECM remodeling and can result in molecules deposition within tumors [421,422,423]. TGF-B levels have been associated with increased ECM remodeling induced by drugs [424,425]. Increased levels of endogenous tissue inhibitors of metalloproteinases (TIMPs) are linked to positive clinical outcomes in many cancers [426,427]. On the other hand, increased levels of receptors including integrins is linked to poor outcomes and disease recurrence [350,428,429]. ECM stiffness impacts adhesion of cells, movement of cells, and response to therapy [430,431,432,433,434]. Increased matrix stiffness within tumors is linked to less responsive tumors and drug resistance (Figure 4) [408,435,436]. Generally, stiffer ECMs are found surrounding tumors compared to ECM in normal tissues [433,437,438,439,440,441,442,443]. ECM stiffness is linked to fibrosis in many cancers including breast cancer where it is observed that many signaling cascades are also activated [335,342,444,445,446,447]. Furthermore, various reports indicate that a stiff TME promote tumor progression via activation of integrin signaling [448,449,450,451]. Tumor metastasis has been shown to be promoted by ECM stiffening via the action of lysyl oxidase and deposition of collagens [452,453,454]. ECM stiffening can also induce microRNAs involved in downregulation of the tumor suppressor protein PTEN [455,456]. Drug delivery rely on diffusion and pressure within the interstitial spaces [457]. The remodeling of the ECM can create a barrier to drug diffusion and either impede drug movement altogether or limit its movement and therefore reduce its effectiveness [458,459,460].
The question on every scientist mind is whether the ECM has relevance to disease initiation and progression beyond what is already known. Analysis of tumor biopsy samples has so far revealed normal and disease-specific ECM signatures in various cancers [461,462]. Low levels of specific ECM molecules including decorin has been associated with poor patients’ survival in various cancers [322,463,464,465]. The understanding of ECM composition and amounts at different stages of tumorigenesis is valuable in disease targeting. Drugs can be designed to target or have great adherence for specific ECM molecules in order to deliver the drugs to specific tumor sites [466]. Studies in various cancers have shown that specific cancers have a specific ECM signature that is both predictive of patients’ survival [322,462,467], and the question is whether specific ECM proteins are pro-tumorigenic or anti-tumorigenic. Various in vitro studies have shown that knockdown or removal of certain ECM proteins can sensitize cancer cells to drugs [17,468,469]. Currently, the use of cancer-specific ECM signatures in treatment strategies is limited and require further detailed analysis of the ECM proteins at specific stages of tumorigenesis. Data from Senthebane and colleagues suggest that targeting fibrillar collagen and fibronectin in tumors may allow drugs to access tumor cells and therefore improve therapeutic efficiency [17].
Many approaches have been utilized to block the pro-tumorigenic properties of the ECM. For example, stopping the expression of collagens and fibronectin leading to reduced drug resistance of cancer cells [142,470]. Several ECM members are known to accumulate in various cancers, and their degradation in combination with chemotherapy can result in better outcomes for patients [471,472]. Furthermore, since most ECM proteins require post-translational modifications for stability and to have the right conformation, disruption of modifications can also result in instable ECM proteins. This has a two-fold effect: decrease in ECM components leads to less fibrosis and can lead to increased drug efficacy and reduced ECM components means less MMPs are required and present within the TME. Less MMPs can reduce the aggressiveness of many cancers [473,474,475]. It is important to note that inhibitors of MMPs activities have given very disappointing results in many clinical trials [476,477,478]. Part of the reason why MMPs inhibitors did not succeed as expected in the many members of the MMPs family and their overlapping activities [479,480].
In addition to targeting ECM components, many signaling cascades that are activated or downregulated by ECM proteins can be regulated. Various survival pathways are activated in various cancers [481,482,483]. Many developed small molecule inhibitors can abrogate cancer cell signaling, leading to induce drug sensitivity in cancer cells [484,485]. Whilst studies using inhibitors of ECM synthesis and signaling are giving promising data, it important to note that ECM components can have both pro- and anti-tumorigenic behavior. This complex behavior requires deep analysis, and many hurdles are still to be overcome.
Therapeutic strategies targeting the Extracellular Matrix
ECM composition consideration as well as genetic mutations has allowed the administration of drugs that target specific ECM molecules and specific organs leading to improved patient outcomes. Important in this regard are new drugs that target specific ECM components, either to upregulate or downregulate their expression, as these can be used in combination with existing drugs. Total removal of specific ECM components may not be the best way forward as this can actually promote tumor progression and impact normal tissue function. In tumors, collagen levels can be regulated at different stages of their synthesis and degradation. Collagen levels can be controlled via targeting various signalling cascades involved in its synthesis such as TGF-B signalling. Antibodies including fresolimumab is currently under clinical trials in cancers where its used to reduce collagen levels [486]. The inhibition of TGF-B signalling through the use of halofuginone has been shown to be effective at reducing collagen levels in various cancers [487,488]. Other drugs used to target the TGF-B cascade include pirfenidone, metformin, tranilast and Ki26894 [489,490]. Caution is needed when inhibiting collagen levels via blocking the TGF-B signalling cascade as TGF-B is involved in other body processes such as inflammation [491,492]. Another way to reduce collagen levels is to use collagenases. In normal tissues, collagenases can easily be made available to degrade collagen [493]. In solid tumors that are compact, collagenases cannot be transported easily due to their large size [494]. A major issue regarding degrading collagen within solid tumors is the potential release of sequestered growth factors, resulting in unintended effects [495,496]. In addition, degradation of any ECM component may create ‘highways’ for cancer cell migration and metastasis [29,497]. MMPs can also be used to degrade ECM components such as collagen. The effectiveness of MMPs use and their inhibitors in cancers has been disappointing with many clinical trials yielding no good results [498].
Various cancer therapies have been used to target fibronectin and these include its potential as a drug delivery molecule. Reports indicate that fibronectin or its isoforms are upregulated in many cancer tissues as well as in normal tissues [499,500,501]. Antibodies against fibronectin domains including L19 have been used to inhibit cancer cell growth [502,503]. In addition, peptides that bind to the fibronectin domain EDB (extra domain B) can be used to deliver drugs and drug-containing exosomes to tumors resulting in better shrinkage of tumors than just drugs alone [504]. Another ECM protein that has been targeted during cancer therapy is hyaluronic acid. Hyaluronic acid synthesis can be inhibited by 4-methylumbelliferone. The inhibition of hyaluronic acid synthesis leads to loss of tissue integrity, thus cause tumors to be leaky with no proper structure. Hyaluronic acid synthesis inhibition thus leads to more drugs reaching tumor cells compared to tumors with normal hyaluronic acid levels [505,506]. Hyaluronic acid can also be degraded via the use of hyaluronidase. Various reports and clinical trials are underway to evaluate the usefulness of hyaluronidase in combination with drugs [471,507]. Various integrins, expressed by cancer cells, can also be targeted in various cancers. The use of antibodies against integrins has shown great results in cancers including breast and colon cancers [508,509]. Such anti-integrin antibodies include volociximab and vitaxin. Small molecule integrin antagonists can target specific integrin-ECM interactions can block integrin-mediated cancer cell migration and therefore prevents tumor cell invasion and metastasis. Such integrin antagonists include cilengitide [510,511]. Antibodies against CD44 including bivatuzumab and RO5429083 have shown anti-tumor activity in patients with advanced cancers [512,513].
Conclusions
The ECM exist in normal and tumor tissues. During development the ECM performs functions including providing structural support for cells and directing cell differentiation. In tumors, the ECM is needed for continual support to the growing tumor mass as well as to promote tumor cell migration and metastasis. Over the years, new technologies and bioinformatic softwares have been developed to delve deeper into the composition of both normal and tumor ECM, revealing that the ECM can be used in directing cellular function and in diagnosis and predictive manner, for example. A better understanding of the ECM led to efforts to interfere with its synthesis and degradation in an effort to improve patient outcomes. In its simplistic nature, targeting individual ECM proteins inadvertently affect other physiological processes and must be done investigated further. Disruptions in ECM synthesis and degradation is likely going to impact tissue homeostasis, a complex state maintained by many interlinked processes. Importantly, just merely disrupting ECM synthesis and degradation is not going to stop pathological conditions such as cancer but require being combined with therapeutic strategies such as chemotherapy, immunotherapy and radiotherapy. This calls for further deeper investigations of how multiple anti-tumor strategies can be combined to have synergistic or additive effects.
References
- Bissell, M.J.; Hall, H.G.; Parry, G. How does the extracellular matrix direct gene expression? Journal of theoretical biology 1982, 99, 31–68. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.M.; Bissell, M.J. Of extracellular matrix, scaffolds, and signaling: tissue architecture regulates development, homeostasis, and cancer. Annu. Rev. Cell Dev. Biol. 2006, 22, 287–309. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.; Mojares, E.; del Río Hernández, A. Role of extracellular matrix in development and cancer progression. International journal of molecular sciences 2018, 19, 3028. [Google Scholar] [CrossRef] [PubMed]
- Pupa, S.M.; Ménard, S.; Forti, S.; Tagliabue, E. New insights into the role of extracellular matrix during tumor onset and progression. Journal of cellular physiology 2002, 192, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Clause, K.C.; Barker, T.H. Extracellular matrix signaling in morphogenesis and repair. Current opinion in biotechnology 2013, 24, 830–833. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K.; Vogelsang, M.; Parker, M.I. Wnt/β-catenin and MEK-ERK signaling are required for fibroblast-derived extracellular matrix-mediated endoderm differentiation of embryonic stem cells. Stem Cell Reviews and Reports 2015, 11, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K.; Turnley, T.; Wishart, A.; Rowe, A.; Kallmeyer, K.; Van Vollenstee, F.A.; Thomford, N.E.; Dandara, C.; Chopera, D.; Pepper, M.S. Fibroblast-derived extracellular matrix induces chondrogenic differentiation in human adipose-derived mesenchymal stromal/stem cells in vitro. International journal of molecular sciences 2016, 17, 1259. [Google Scholar] [CrossRef]
- Yin, H.; Wang, J.; Li, H.; Yu, Y.; Wang, X.; Lu, L.; Lv, C.; Chang, B.; Jin, W.; Guo, W.; et al. Extracellular matrix protein-1 secretory isoform promotes ovarian cancer through increasing alternative mRNA splicing and stemness. Nature Communications 2021, 12, 4230. [Google Scholar] [CrossRef]
- Dussoyer, M.; Page, A.; Delolme, F.; Rousselle, P.; Nyström, A.; Moali, C. Comparison of extracellular matrix enrichment protocols for the improved characterization of the skin matrisome by mass spectrometry. J Proteomics 2022, 251, 104397. [Google Scholar] [CrossRef]
- Mienaltowski, M.J.; Gonzales, N.L.; Beall, J.M.; Pechanec, M.Y. Basic Structure, Physiology, and Biochemistry of Connective Tissues and Extracellular Matrix Collagens. Adv Exp Med Biol 2021, 1348, 5–43. [Google Scholar] [CrossRef]
- Giblin, S.P.; Schwenzer, A.; Midwood, K.S. Alternative splicing controls cell lineage-specific responses to endogenous innate immune triggers within the extracellular matrix. Matrix Biol 2020, 93, 95–114. [Google Scholar] [CrossRef]
- Chen, W.; Yuan, Y.; Li, C.; Mao, H.; Liu, B.; Jiang, X. Modulating Tumor Extracellular Matrix by Simultaneous Inhibition of Two Cancer Cell Receptors. Adv Mater 2021, e2109376. [Google Scholar] [CrossRef]
- Gu, X.; Ge, L.; Ren, B.; Fang, Y.; Li, Y.; Wang, Y.; Xu, H. Glucocorticoids Promote Extracellular Matrix Component Remodeling by Activating YAP in Human Retinal Capillary Endothelial Cells. Front Cell Dev Biol 2021, 9, 738341. [Google Scholar] [CrossRef] [PubMed]
- Laurito, T.L.; França, F.T.; Vieira-Damiani, G.; Pelegati, V.B.; Baratti, M.O.; de Carvalho, H.F.; Cesar, C.L.; de Moraes, A.M.; Cintra, M.L.; Teixeira, F. The texture of collagen in the microenvironments of Merkel cell carcinoma. Medicine (Baltimore) 2021, 100, e27925. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K. Taking a Full Snapshot of Cancer Biology: Deciphering the Tumor Microenvironment for Effective Cancer Therapy in the Oncology Clinic. Omics 2020, 24, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K.; Leaner, V.D.; Parker, M.I. Feedback regulation of the α2(1) collagen gene via the Mek-Erk signaling pathway. IUBMB Life 2012, 64, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Senthebane, D.A.; Jonker, T.; Rowe, A.; Thomford, N.E.; Munro, D.; Dandara, C.; Wonkam, A.; Govender, D.; Calder, B.; Soares, N.C.; et al. The Role of Tumor Microenvironment in Chemoresistance: 3D Extracellular Matrices as Accomplices. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Senthebane, D.A.; Rowe, A.; Thomford, N.E.; Shipanga, H.; Munro, D.; Mazeedi, M.; Almazyadi, H.A.M.; Kallmeyer, K.; Dandara, C.; Pepper, M.S.; et al. The Role of Tumor Microenvironment in Chemoresistance: To Survive, Keep Your Enemies Closer. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- Cox, T.R. The matrix in cancer. Nat Rev Cancer 2021, 21, 217–238. [Google Scholar] [CrossRef]
- Bissell, M.J.; Hall, H.G.; Parry, G. How does the extracellular matrix direct gene expression? J Theor Biol 1982, 99, 31–68. [Google Scholar] [CrossRef]
- Seo, B.R.; Bhardwaj, P.; Choi, S.; Gonzalez, J.; Eguiluz, R.C.A.; Wang, K.; Mohanan, S.; Morris, P.G.; Du, B.; Zhou, X.K. Obesity-dependent changes in interstitial ECM mechanics promote breast tumorigenesis. Science translational medicine 2015, 7, 301ra130–301ra130. [Google Scholar] [CrossRef]
- Lukashev, M.E.; Werb, Z. ECM signalling: orchestrating cell behaviour and misbehaviour. Trends in cell biology 1998, 8, 437–441. [Google Scholar] [CrossRef]
- Ghajar, C.M.; Bissell, M.J. Extracellular matrix control of mammary gland morphogenesis and tumorigenesis: insights from imaging. Histochemistry and cell biology 2008, 130, 1105. [Google Scholar] [CrossRef]
- Dzobo, K.; Vogelsang, M.; Thomford, N.E.; Dandara, C.; Kallmeyer, K.; Pepper, M.S.; Parker, M.I. Wharton’s Jelly-Derived Mesenchymal Stromal Cells and Fibroblast-Derived Extracellular Matrix Synergistically Activate Apoptosis in a p21-Dependent Mechanism in WHCO1 and MDA MB 231 Cancer Cells<i> In Vitro</i>. Stem Cells International 2016, 2016, 4842134. [Google Scholar] [CrossRef]
- Sarrazy, V.; Billet, F.; Micallef, L.; Coulomb, B.; Desmoulière, A. Mechanisms of pathological scarring: role of myofibroblasts and current developments. Wound Repair and Regeneration 2011, 19, s10–s15. [Google Scholar] [CrossRef]
- Vitale, I.; Manic, G.; Galassi, C.; Galluzzi, L. Stress responses in stromal cells and tumor homeostasis. Pharmacology & therapeutics 2019, 200, 55–68. [Google Scholar]
- Nallanthighal, S.; Heiserman, J.P.; Cheon, D.-J. The role of the extracellular matrix in cancer stemness. Frontiers in cell and developmental biology 2019, 7, 86. [Google Scholar] [CrossRef]
- Dzobo, K.; Senthebane, D.A.; Dandara, C. The tumor microenvironment in tumorigenesis and therapy resistance revisited. Cancers 2023, 15, 376. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Yuan, J.; Peng, C.; Li, Y. Collagen as a double-edged sword in tumor progression. Tumor Biology 2014, 35, 2871–2882. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Rasch, M.G.; Weaver, V.M. Dynamic interplay between the collagen scaffold and tumor evolution. Current opinion in cell biology 2010, 22, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Deegan, D.B.; Zimmerman, C.; Skardal, A.; Atala, A.; Shupe, T.D. Stiffness of hyaluronic acid gels containing liver extracellular matrix supports human hepatocyte function and alters cell morphology. Journal of the mechanical behavior of biomedical materials 2016, 55, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R.; Erler, J.T. Molecular pathways: connecting fibrosis and solid tumor metastasis. Clin Cancer Res 2014, 20, 3637–3643. [Google Scholar] [CrossRef] [PubMed]
- Piersma, B.; Hayward, M.K.; Weaver, V.M. Fibrosis and cancer: A strained relationship. Biochim Biophys Acta Rev Cancer 2020, 1873, 188356. [Google Scholar] [CrossRef]
- Houghton, A.M. Mechanistic links between COPD and lung cancer. Nat Rev Cancer 2013, 13, 233–245. [Google Scholar] [CrossRef]
- Michelotti, G.A.; Machado, M.V.; Diehl, A.M. NAFLD, NASH and liver cancer. Nat Rev Gastroenterol Hepatol 2013, 10, 656–665. [Google Scholar] [CrossRef]
- Cox, T.R.; Bird, D.; Baker, A.M.; Barker, H.E.; Ho, M.W.; Lang, G.; Erler, J.T. LOX-mediated collagen crosslinking is responsible for fibrosis-enhanced metastasis. Cancer Res 2013, 73, 1721–1732. [Google Scholar] [CrossRef]
- Joo, Y.N.; Jin, H.; Eun, S.Y.; Park, S.W.; Chang, K.C.; Kim, H.J. P2Y2R activation by nucleotides released from the highly metastatic breast cancer cell MDA-MB-231 contributes to pre-metastatic niche formation by mediating lysyl oxidase secretion, collagen crosslinking, and monocyte recruitment. Oncotarget 2014, 5, 9322–9334. [Google Scholar] [CrossRef]
- Choi, S.K.; Kim, H.S.; Jin, T.; Moon, W.K. LOXL4 knockdown enhances tumor growth and lung metastasis through collagen-dependent extracellular matrix changes in triple-negative breast cancer. Oncotarget 2017, 8, 11977–11989. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.H.; Hsia, S.M.; Shieh, T.M. Lysyl Oxidase and the Tumor Microenvironment. Int J Mol Sci 2016, 18. [Google Scholar] [CrossRef] [PubMed]
- Giussani, M.; Landoni, E.; Merlino, G.; Turdo, F.; Veneroni, S.; Paolini, B.; Cappelletti, V.; Miceli, R.; Orlandi, R.; Triulzi, T. Extracellular matrix proteins as diagnostic markers of breast carcinoma. Journal of cellular physiology 2018, 233, 6280–6290. [Google Scholar] [CrossRef]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The extracellular matrix: Tools and insights for the “omics” era. Matrix Biology 2016, 49, 10–24. [Google Scholar] [CrossRef]
- Giussani, M.; Triulzi, T.; Sozzi, G.; Tagliabue, E. Tumor extracellular matrix remodeling: new perspectives as a circulating tool in the diagnosis and prognosis of solid tumors. Cells 2019, 8, 81. [Google Scholar] [CrossRef]
- Yang, J.D.; Nakamura, I.; Roberts, L.R. The tumor microenvironment in hepatocellular carcinoma: current status and therapeutic targets. in Seminars in cancer biology. 2011. Elsevier. [CrossRef]
- Jun, J.-I.; Lau, L.F. Taking aim at the extracellular matrix: CCN proteins as emerging therapeutic targets. Nature reviews Drug discovery 2011, 10, 945–963. [Google Scholar] [CrossRef]
- Wood, S.L.; Pernemalm, M.; Crosbie, P.A.; Whetton, A.D. The role of the tumor-microenvironment in lung cancer-metastasis and its relationship to potential therapeutic targets. Cancer treatment reviews 2014, 40, 558–566. [Google Scholar] [CrossRef]
- Zanconato, F.; Battilana, G.; Cordenonsi, M.; Piccolo, S. YAP/TAZ as therapeutic targets in cancer. Current opinion in pharmacology 2016, 29, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Fuster, M.M.; Esko, J.D. The sweet and sour of cancer: glycans as novel therapeutic targets. Nature Reviews Cancer 2005, 5, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, L.; Wan, D.; Zhou, L.; Zheng, S.; Lin, S.; Qiao, Y. Extracellular matrix and its therapeutic potential for cancer treatment. Signal Transduction and Targeted Therapy 2021, 6, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O.; Naba, A. Overview of the matrisome—an inventory of extracellular matrix constituents and functions. Cold Spring Harbor perspectives in biology 2012, 4, a004903. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.T.; Horwitz, A.R.; Schwartz, M.A. Cell adhesion: integrating cytoskeletal dynamics and cellular tension. Nat Rev Mol Cell Biol 2010, 11, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. The extracellular matrix: not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef]
- Fang, M.; Yuan, J.; Peng, C.; Li, Y. Collagen as a double-edged sword in tumor progression. Tumour Biol 2014, 35, 2871–2882. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S. The collagen family. Cold Spring Harb Perspect Biol 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed]
- Gay, S.; Vijanto, J.; Raekallio, J.; Penttinen, R. Collagen types in early phases of wound healing in children. Acta Chirurgica Scandinavica 1978, 144, 205–211. [Google Scholar] [PubMed]
- Nyström, A.; Velati, D.; Mittapalli, V.R.; Fritsch, A.; Kern, J.S.; Bruckner-Tuderman, L. Collagen VII plays a dual role in wound healing. The Journal of clinical investigation 2013, 123, 3498–3509. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Young, A.; McNaught, C.-E. The physiology of wound healing. Surgery (Oxford) 2017, 35, 473–477. [Google Scholar] [CrossRef]
- Dzobo, K. Matrix-mediated regulation of type 1 collagen synthesis and degradation in cultured fibroblasts. 2009. [Google Scholar]
- Scott, I.; Yamauchi, M.; Sricholpech, M. Lysine post-translational modifications of collagen. Essays in biochemistry 2012, 52, 113–133. [Google Scholar] [CrossRef]
- Garnero, P.; Borel, O.; Gineyts, E.; Duboeuf, F.; Solberg, H.; Bouxsein, M.L.; Christiansen, C.; Delmas, P.D. Extracellular post-translational modifications of collagen are major determinants of biomechanical properties of fetal bovine cortical bone. Bone 2006, 38, 300–309. [Google Scholar] [CrossRef]
- Sottile, J.; Shi, F.; Rublyevska, I.; Chiang, H.-Y.; Lust, J.; Chandler, J. Fibronectin-dependent collagen I deposition modulates the cell response to fibronectin. American journal of physiology-Cell Physiology 2007, 293, C1934–C1946. [Google Scholar] [CrossRef]
- Kubow, K.E.; Vukmirovic, R.; Zhe, L.; Klotzsch, E.; Smith, M.L.; Gourdon, D.; Luna, S.; Vogel, V. Mechanical forces regulate the interactions of fibronectin and collagen I in extracellular matrix. Nature communications 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Dzobo, K.; Leaner, V.D.; Parker, M.I. Feedback regulation of the alpha2(1) collagen gene via the Mek-Erk signaling pathway. IUBMB Life 2012, 64, 87–98. [Google Scholar] [CrossRef]
- Dzobo, K.; Leaner, V.D.; Parker, M.I. Absence of feedback regulation in the synthesis of COL1A1. Life Sci 2014, 103, 25–33. [Google Scholar] [CrossRef]
- Chelyshev, Y.A.; Kabdesh, I.M.; Mukhamedshina, Y.O. Extracellular Matrix in Neural Plasticity and Regeneration. Cell Mol Neurobiol 2020. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R.; Erler, J.T. Fibrosis, cancer and the premetastatic niche. Breast Cancer Management 2014, 3, 453–455. [Google Scholar] [CrossRef]
- Hastings, J.F.; Skhinas, J.N.; Fey, D.; Croucher, D.R.; Cox, T.R. The extracellular matrix as a key regulator of intracellular signalling networks. British journal of pharmacology 2019, 176, 82–92. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Eliceiri, K.W.; Campbell, J.M.; Inman, D.R.; White, J.G.; Keely, P.J. Collagen reorganization at the tumor-stromal interface facilitates local invasion. BMC Med 2006, 4, 38. [Google Scholar] [CrossRef]
- Amatangelo, M.D.; Bassi, D.E.; Klein-Szanto, A.J.; Cukierman, E. Stroma-derived three-dimensional matrices are necessary and sufficient to promote desmoplastic differentiation of normal fibroblasts. Am J Pathol 2005, 167, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. Journal of cell science 2010, 123, 4195–4200. [Google Scholar] [CrossRef]
- Hay, E.D. Cell biology of extracellular matrix; Springer Science & Business Media, 2013. [Google Scholar]
- Sonbol, H.S. Extracellular matrix remodeling in human disease. Journal of microscopy and ultrastructure 2018, 6, 123. [Google Scholar] [CrossRef] [PubMed]
- Halfter, W.; Moes, S.; Asgeirsson, D.O.; Halfter, K.; Oertle, P.; Melo Herraiz, E.; Plodinec, M.; Jenoe, P.; Henrich, P.B. Diabetes-related changes in the protein composition and the biomechanical properties of human retinal vascular basement membranes. PLoS One 2017, 12, e0189857. [Google Scholar] [CrossRef]
- Candiello, J.; Balasubramani, M.; Schreiber, E.M.; Cole, G.J.; Mayer, U.; Halfter, W.; Lin, H. Biomechanical properties of native basement membranes. The FEBS journal 2007, 274, 2897–2908. [Google Scholar] [CrossRef]
- Amenta, P.S.; Briggs, K.; Xu, K.; Gamboa, E.; Jukkola, A.F.; Li, D.; Myers, J.C. Type XV collagen in human colonic adenocarcinomas has a different distribution than other basement membrane zone proteins. Hum Pathol 2000, 31, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Amenta, P.S.; Hadad, S.; Lee, M.T.; Barnard, N.; Li, D.; Myers, J.C. Loss of types XV and XIX collagen precedes basement membrane invasion in ductal carcinoma of the female breast. J Pathol 2003, 199, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Tosios, K.; Kapranos, N.; Papanicolaou, S. Loss of basement membrane components laminin and type IV collagen parallels the progression of oral epithelial neoplasia. Histopathology 1998, 33, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Jolly, L.A.; Novitskiy, S.; Owens, P.; Massoll, N.; Cheng, N.; Fang, W.; Moses, H.L.; Franco, A.T. Fibroblast-mediated collagen remodeling within the tumor microenvironment facilitates progression of thyroid cancers driven by BrafV600E and Pten loss. Cancer research 2016, 76, 1804–1813. [Google Scholar] [CrossRef]
- Spivey, K.A.; Chung, I.; Banyard, J.; Adini, I.; Feldman, H.A.; Zetter, B.R. A role for collagen XXIII in cancer cell adhesion, anchorage-independence and metastasis. Oncogene 2012, 31, 2362–2372. [Google Scholar] [CrossRef]
- Madsen, C.D. Pancreatic cancer is suppressed by fibroblast-derived collagen I. Cancer Cell 2021, 39, 451–453. [Google Scholar] [CrossRef]
- Kresse, H.; Schönherr, E. Proteoglycans of the extracellular matrix and growth control. Journal of cellular physiology 2001, 189, 266–274. [Google Scholar] [CrossRef]
- Lee, K.; Loganathan, D.; Merchant, Z.; Linhardt, R. Carbohydrate analysis of glycoproteins A review. Applied biochemistry and biotechnology 1990, 23, 53–80. [Google Scholar] [CrossRef]
- Hughes, R.C. Membrane glycoproteins: a review of structure and function. 2014. [Google Scholar]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol 2015, 42, 11–55. [Google Scholar] [CrossRef]
- Iozzo, R.V. Matrix proteoglycans: from molecular design to cellular function. Annu Rev Biochem 1998, 67, 609–652. [Google Scholar] [CrossRef]
- Hardingham, T.E.; Fosang, A.J. Proteoglycans: many forms and many functions. Faseb j 1992, 6, 861–870. [Google Scholar] [CrossRef]
- Ishihara, J.; Ishihara, A.; Fukunaga, K.; Sasaki, K.; White, M.J.V.; Briquez, P.S.; Hubbell, J.A. Laminin heparin-binding peptides bind to several growth factors and enhance diabetic wound healing. Nature Communications 2018, 9, 2163. [Google Scholar] [CrossRef]
- Schönherr, E.; Hausser, H.-J. Extracellular Matrix and Cytokines: A Functional Unit. Developmental Immunology 2000, 7, 031748. [Google Scholar] [CrossRef]
- Broekelmann, T.J.; Bodmer, N.K.; Mecham, R.P. Identification of the growth factor–binding sequence in the extracellular matrix protein MAGP-1. Journal of Biological Chemistry 2020, 295, 2687–2697. [Google Scholar] [CrossRef] [PubMed]
- Bohaumilitzky, L.; Huber, A.K.; Stork, E.M.; Wengert, S.; Woelfl, F.; Boehm, H. A Trickster in Disguise: Hyaluronan's Ambivalent Roles in the Matrix. Front Oncol 2017, 7, 242. [Google Scholar] [CrossRef] [PubMed]
- Price, Z.K.; Lokman, N.A.; Ricciardelli, C. Differing roles of hyaluronan molecular weight on cancer cell behavior and chemotherapy resistance. Cancers 2018, 10, 482. [Google Scholar] [CrossRef] [PubMed]
- Januchowski, R.; Zawierucha, P.; Ruciński, M.; Nowicki, M.; Zabel, M. Extracellular Matrix Proteins Expression Profiling in Chemoresistant Variants of the A2780 Ovarian Cancer Cell Line. BioMed Research International 2014, 2014, 365867. [Google Scholar] [CrossRef] [PubMed]
- Timpl, R.; Rohde, H.; Robey, P.G.; Rennard, S.I.; Foidart, J.M.; Martin, G.R. Laminin--a glycoprotein from basement membranes. The Journal of biological chemistry 1979, 254, 9933–9937. [Google Scholar] [CrossRef] [PubMed]
- Hohenester, E.; Yurchenco, P.D. Laminins in basement membrane assembly. Cell adhesion & migration 2013, 7, 56–63. [Google Scholar]
- Domogatskaya, A.; Rodin, S.; Tryggvason, K. Functional diversity of laminins. Annual review of cell and developmental biology 2012, 28, 523–553. [Google Scholar] [CrossRef]
- Hohenester, E. Structural biology of laminins. Essays Biochem 2019, 63, 285–295. [Google Scholar] [CrossRef]
- Aumailley, M. The laminin family. Cell Adh Migr 2013, 7, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, S.; Shinto, E.; Tsuda, H.; Ueno, H.; Shikina, A.; Kajiwara, Y.; Yamamoto, J.; Hase, K. Laminin β3 expression as a prognostic factor and a predictive marker of chemoresistance in colorectal cancer. Japanese Journal of Clinical Oncology 2015, 45, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Govaere, O.; Wouters, J.; Petz, M.; Vandewynckel, Y.P.; Van den Eynde, K.; Van den Broeck, A.; Verhulst, S.; Dolle, L.; Gremeaux, L.; Ceulemans, A.; et al. Laminin-332 sustains chemoresistance and quiescence as part of the human hepatic cancer stem cell niche. Journal of hepatology 2016, 64, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Hasebe, T.; Oda, T.; Sasaki, S.; Kinoshita, T.; Konishi, M.; Ochiai, T.; Ochiai, A. Cytoplasmic expression of laminin γ2 chain correlates with postoperative hepatic metastasis and poor prognosis in patients with pancreatic ductal adenocarcinoma. Cancer 2002, 94, 1894–1901. [Google Scholar] [CrossRef] [PubMed]
- Shang, M.; Koshikawa, N.; Schenk, S.; Quaranta, V. The LG3 module of laminin-5 harbors a binding site for integrin α3β1 that promotes cell adhesion, spreading, and migration. Journal of Biological Chemistry 2001, 276, 33045–33053. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.-C.; Ziober, B.L.; Squillace, R.M.; Kramer, R.H. α7 integrin mediates cell adhesion and migration on specific laminin isoforms. Journal of Biological Chemistry 1996, 271, 25598–25603. [Google Scholar] [CrossRef] [PubMed]
- Giannelli, G.; Azzariti, A.; Fransvea, E.; Porcelli, L.; Antonaci, S.; Paradiso, A. Laminin-5 offsets the efficacy of gefitinib (‘Iressa’) in hepatocellular carcinoma cells. British Journal of Cancer 2004, 91, 1964–1969. [Google Scholar] [CrossRef] [PubMed]
- Tsurutani, J.; West, K.A.; Sayyah, J.; Gills, J.J.; Dennis, P.A. Inhibition of the phosphatidylinositol 3-kinase/Akt/mammalian target of rapamycin pathway but not the MEK/ERK pathway attenuates laminin-mediated small cell lung cancer cellular survival and resistance to imatinib mesylate or chemotherapy. Cancer research 2005, 65, 8423–8432. [Google Scholar] [CrossRef]
- Pankov, R.; Yamada, K.M. Fibronectin at a glance. Journal of Cell Science 2002, 115, 3861–3863. [Google Scholar] [CrossRef]
- Gopal, S.; Veracini, L.; Grall, D.; Butori, C.; Schaub, S.; Audebert, S.; Camoin, L.; Baudelet, E.; Radwanska, A.; Beghelli-de la Forest Divonne, S.; et al. Fibronectin-guided migration of carcinoma collectives. Nature communications 2017, 8, 14105. [Google Scholar] [CrossRef]
- Rintoul, R.C.; Sethi, T. Extracellular matrix regulation of drug resistance in small-cell lung cancer. Clinical Science 2002, 102, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Hazlehurst, L.A.; Argilagos, R.F.; Emmons, M.; Boulware, D.; Beam, C.A.; Sullivan, D.M.; Dalton, W.S. Cell adhesion to fibronectin (CAM-DR) influences acquired mitoxantrone resistance in U937 cells. Cancer research 2006, 66, 2338–2345. [Google Scholar] [CrossRef] [PubMed]
- Kosmehl, H.; Berndt, A.; Strassburger, S.; Borsi, L.; Rousselle, P.; Mandel, U.; Hyckel, P.; Zardi, L.; Katenkamp, D. Distribution of laminin and fibronectin isoforms in oral mucosa and oral squamous cell carcinoma. British Journal of Cancer 1999, 81, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, M.; Zardi, L.; Neri, D. Fibronectin as target for tumor therapy. International journal of cancer 2006, 118, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Cheng, S.; Asa, S.L.; Ezzat, S. The melanoma-associated antigen A3 mediates fibronectin-controlled cancer progression and metastasis. Cancer research 2008, 68, 8104–8112. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.K.; Kim, A.; Kim, M.K.; Choi, J.E.; Kang, S.H.; Lee, S.J. Fibronectin expression in carcinoma cells correlates with tumor aggressiveness and poor clinical outcome in patients with invasive breast cancer. Human pathology 2013, 44, 2028–2037. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Ansari, D.; Zhou, Q.; Sasor, A.; Said Hilmersson, K.; Andersson, R. Stromal fibronectin expression in patients with resected pancreatic ductal adenocarcinoma. World journal of surgical oncology 2019, 17, 1–8. [Google Scholar] [CrossRef]
- Thomas, G.; Nyström, M.; Marshall, J. αvβ6 integrin in wound healing and cancer of the oral cavity. Journal of oral pathology & medicine 2006, 35, 1–10. [Google Scholar]
- Singh, P.; Reimer, C.L.; Peters, J.H.; Stepp, M.A.; Hynes, R.O.; Van De Water, L. The spatial and temporal expression patterns of integrin α9β1 and one of its ligands, the EIIIA segment of fibronectin, in cutaneous wound healing. Journal of investigative dermatology 2004, 123, 1176–1181. [Google Scholar] [CrossRef]
- Han, S.; Sidell, N.; Roser-Page, S.; Roman, J. Fibronectin stimulates human lung carcinoma cell growth by inducing cyclooxygenase-2 (COX-2) expression. International Journal of Cancer 2004, 111, 322–331. [Google Scholar] [CrossRef]
- Han, S.; Roman, J. Fibronectin induces cell proliferation and inhibits apoptosis in human bronchial epithelial cells: pro-oncogenic effects mediated by PI3-kinase and NF-κB. Oncogene 2006, 25, 4341–4349. [Google Scholar] [CrossRef]
- Han, S.; Sidell, N.; Roman, J. Fibronectin stimulates human lung carcinoma cell proliferation by suppressing p21 gene expression via signals involving Erk and Rho kinase. Cancer letters 2005, 219, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Weng, D.; Chen, G.; Tao, W.; Zhu, T.; Yang, X.; Meng, L.; Wang, S.; Lu, Y.; Ma, D. Activation of fibronectin/PI-3K/Akt2 leads to chemoresistance to docetaxel by regulating survivin protein expression in ovarian and breast cancer cells. Cancer letters 2008, 261, 108–119. [Google Scholar] [CrossRef]
- Horiuchi, K.; Amizuka, N.; Takeshita, S.; Takamatsu, H.; Katsuura, M.; Ozawa, H.; Toyama, Y.; Bonewald, L.F.; Kudo, A. Identification and characterization of a novel protein, periostin, with restricted expression to periosteum and periodontal ligament and increased expression by transforming growth factor beta. Journal of bone and mineral research : the official journal of the American Society for Bone and Mineral Research 1999, 14, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Moniuszko, T.; Wincewicz, A.; Koda, M.; Domysławska, I.; Sulkowski, S. Role of periostin in esophageal, gastric and colon cancer. Oncology letters 2016, 12, 783–787. [Google Scholar] [CrossRef]
- Gillan, L.; Matei, D.; Fishman, D.A.; Gerbin, C.S.; Karlan, B.Y.; Chang, D.D. Periostin secreted by epithelial ovarian carcinoma is a ligand for alpha(V)beta(3) and alpha(V)beta(5) integrins and promotes cell motility. Cancer research 2002, 62, 5358–5364. [Google Scholar]
- Underwood, T.J.; Hayden, A.L.; Derouet, M.; Garcia, E.; Noble, F.; White, M.J.; Thirdborough, S.; Mead, A.; Clemons, N.; Mellone, M.; et al. Cancer-associated fibroblasts predict poor outcome and promote periostin-dependent invasion in oesophageal adenocarcinoma. The Journal of pathology 2015, 235, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, T.; Tamai, K.; Shibuya, R.; Nakamura, M.; Mochizuki, M.; Yamaguchi, K.; Abe, J.; Takahashi, S.; Sato, I.; Kudo, A. Periostin is a negative prognostic factor and promotes cancer cell proliferation in non-small cell lung cancer. Oncotarget 2018, 9, 31187. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Fejzo, M.S.; Anderson, L.; Dering, J.; Ginther, C.; Ramos, L.; Gasson, J.C.; Karlan, B.Y.; Slamon, D.J. Periostin promotes ovarian cancer angiogenesis and metastasis. Gynecologic oncology 2010, 119, 337–344. [Google Scholar] [CrossRef]
- Tumbarello, D.A.; Temple, J.; Brenton, J.D. ß3 integrin modulates transforming growth factor beta induced (TGFBI) function and paclitaxel response in ovarian cancer cells. Molecular cancer 2012, 11, 36–36. [Google Scholar] [CrossRef]
- Sung, P.-L.; Jan, Y.-H.; Lin, S.-C.; Huang, C.-C.; Lin, H.; Wen, K.-C.; Chao, K.-C.; Lai, C.-R.; Wang, P.-H.; Chuang, C.-M.; et al. Periostin in tumor microenvironment is associated with poor prognosis and platinum resistance in epithelial ovarian carcinoma. Oncotarget 2016, 7, 4036–4047. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Du, L. Role of pancreatic stellate cells and periostin in pancreatic cancer progression. Tumor Biology 2015, 36, 3171–3177. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Fejzo, M.S.; Anderson, L.; Dering, J.; Ginther, C.; Ramos, L.; Gasson, J.C.; Karlan, B.Y.; Slamon, D.J. Periostin promotes ovarian cancer angiogenesis and metastasis. Gynecologic oncology 2010, 119, 337–344. [Google Scholar] [CrossRef]
- Tammi, M.I.; Day, A.J.; Turley, E.A. Hyaluronan and homeostasis: a balancing act. Journal of Biological Chemistry 2002, 277, 4581–4584. [Google Scholar] [CrossRef] [PubMed]
- Fallacara, A.; Baldini, E.; Manfredini, S.; Vertuani, S. Hyaluronic acid in the third millennium. Polymers 2018, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Kakehi, K.; Kinoshita, M.; Yasueda, S.-i. Hyaluronic acid: separation and biological implications. Journal of Chromatography B 2003, 797, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.E. Supramolecular organization of extracellular matrix glycosaminoglycans, in vitro and in the tissues. The FASEB journal 1992, 6, 2639–2645. [Google Scholar] [CrossRef]
- Kupper, S.; Kłosowska-Chomiczewska, I.; Szumała, P. Collagen and hyaluronic acid hydrogel in water-in-oil microemulsion delivery systems. Carbohydrate polymers 2017, 175, 347–354. [Google Scholar] [CrossRef]
- Pereira, H.; Sousa, D.A.; Cunha, A.; Andrade, R.; Espregueira-Mendes, J.; Oliveira, J.M.; Reis, R.L. Hyaluronic acid. Osteochondral Tissue Engineering 2018, 137–153. [Google Scholar]
- Henry, C.B.; Duling, B.R. Permeation of the luminal capillary glycocalyx is determined by hyaluronan. American Journal of Physiology-Heart and Circulatory Physiology 1999. [Google Scholar] [CrossRef] [PubMed]
- Barbucci, R.; Lamponi, S.; Borzacchiello, A.; Ambrosio, L.; Fini, M.; Torricelli, P.; Giardino, R. Hyaluronic acid hydrogel in the treatment of osteoarthritis. Biomaterials 2002, 23, 4503–4513. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Kirker, K.R.; Prestwich, G.D. Cross-linked hyaluronic acid hydrogel films: new biomaterials for drug delivery. Journal of controlled release 2000, 69, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Mattheolabakis, G.; Milane, L.; Singh, A.; Amiji, M.M. Hyaluronic acid targeting of CD44 for cancer therapy: from receptor biology to nanomedicine. Journal of drug targeting 2015, 23, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.S.; Svechkarev, D.; Souchek, J.; Hill, T.K.; Taylor, M.; Natarajan, A.; Mohs, A.M. Impact of structurally modifying hyaluronic acid on CD44 interaction. Journal of Materials Chemistry B 2017, 5, 8183–8192. [Google Scholar] [CrossRef]
- Necas, J.; Bartosikova, L.; Brauner, P.; Kolar, J. Hyaluronic acid (hyaluronan): a review. Veterinarni medicina 2008, 53, 397–411. [Google Scholar] [CrossRef]
- Brown, N.H. Extracellular matrix in development: insights from mechanisms conserved between invertebrates and vertebrates. Cold Spring Harb Perspect Biol 2011, 3. [Google Scholar] [CrossRef]
- Senthebane, D.A.; Jonker, T.; Rowe, A.; Thomford, N.E.; Munro, D.; Dandara, C.; Wonkam, A.; Govender, D.; Calder, B.; Soares, N.C. The role of tumor microenvironment in chemoresistance: 3D extracellular matrices as accomplices. International journal of molecular sciences 2018, 19, 2861. [Google Scholar] [CrossRef]
- Entchev, E.V.; González-Gaitán, M.A. Morphogen gradient formation and vesicular trafficking. Traffic 2002, 3, 98–109. [Google Scholar] [CrossRef]
- Marois, E.; Mahmoud, A.; Eaton, S. The endocytic pathway and formation of the Wingless morphogen gradient. Development 2006, 133, 307–317. [Google Scholar] [CrossRef]
- Uhler, C.; Shivashankar, G.V. Regulation of genome organization and gene expression by nuclear mechanotransduction. Nat Rev Mol Cell Biol 2017, 18, 717–727. [Google Scholar] [CrossRef]
- Ilic, D.; Damsky, C.H.; Yamamoto, T. Focal adhesion kinase: at the crossroads of signal transduction. Journal of cell science 1997, 110, 401–407. [Google Scholar] [CrossRef]
- Kai, F.; Laklai, H.; Weaver, V.M. Force Matters: Biomechanical Regulation of Cell Invasion and Migration in Disease. Trends Cell Biol 2016, 26, 486–497. [Google Scholar] [CrossRef]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: a dynamic view. Dev Biol 2010, 341, 126–140. [Google Scholar] [CrossRef]
- Park, J.; Kim, D.-H.; Levchenko, A. Topotaxis: a new mechanism of directed cell migration in topographic ECM gradients. Biophysical journal 2018, 114, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Janson, I.A.; Putnam, A.J. Extracellular matrix elasticity and topography: Material-based cues that affect cell function via conserved mechanisms. Journal of biomedical materials research Part A 2015, 103, 1246–1258. [Google Scholar] [CrossRef]
- Doyle, A.D.; Petrie, R.J.; Kutys, M.L.; Yamada, K.M. Dimensions in cell migration. Current opinion in cell biology 2013, 25, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Gunawan, R.C.; Silvestre, J.; Gaskins, H.R.; Kenis, P.J.; Leckband, D.E. Cell migration and polarity on microfabricated gradients of extracellular matrix proteins. Langmuir 2006, 22, 4250–4258. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Mao, Z.; Tan, H.; Han, L.; Ren, T.; Gao, C. Gradient biomaterials and their influences on cell migration. Interface focus 2012, 2, 337–355. [Google Scholar] [CrossRef] [PubMed]
- Shellard, A.; Mayor, R. All roads lead to directional cell migration. Trends in cell biology 2020, 30, 852–868. [Google Scholar] [CrossRef]
- Palecek, S.P.; Loftus, J.C.; Ginsberg, M.H.; Lauffenburger, D.A.; Horwitz, A.F. Integrin-ligand binding properties govern cell migration speed through cell-substratum adhesiveness. Nature 1997, 385, 537–540. [Google Scholar] [CrossRef]
- Hartman, C.D.; Isenberg, B.C.; Chua, S.G.; Wong, J.Y. Extracellular matrix type modulates cell migration on mechanical gradients. Exp Cell Res 2017, 359, 361–366. [Google Scholar] [CrossRef]
- Hartman, C.D.; Isenberg, B.C.; Chua, S.G.; Wong, J.Y. Vascular smooth muscle cell durotaxis depends on extracellular matrix composition. Proc Natl Acad Sci U S A 2016, 113, 11190–11195. [Google Scholar] [CrossRef]
- Plotnikov, S.V.; Waterman, C.M. Guiding cell migration by tugging. Curr Opin Cell Biol 2013, 25, 619–626. [Google Scholar] [CrossRef]
- Pathak, A.; Kumar, S. Independent regulation of tumor cell migration by matrix stiffness and confinement. Proceedings of the National Academy of Sciences 2012, 109, 10334–10339. [Google Scholar] [CrossRef]
- Charras, G.; Sahai, E. Physical influences of the extracellular environment on cell migration. Nature reviews Molecular cell biology 2014, 15, 813–824. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat Rev Mol Cell Biol 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Pytliak, M.; Vargová, V.; Mechírová, V. Matrix metalloproteinases and their role in oncogenesis: a review. Onkologie 2012, 35, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Rainero, E. Extracellular matrix endocytosis in controlling matrix turnover and beyond: emerging roles in cancer. Biochem Soc Trans 2016, 44, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Hinck, L.; Silberstein, G.B. Key stages in mammary gland development: the mammary end bud as a motile organ. Breast Cancer Res 2005, 7, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Alford, D.; Baeckström, D.; Geyp, M.; Pitha, P.; Taylor-Papadimitriou, J. Integrin-matrix interactions affect the form of the structures developing from human mammary epithelial cells in collagen or fibrin gels. J Cell Sci 1998, 111 Pt 4, 521–532. [Google Scholar] [CrossRef]
- Walker, C.; Mojares, E.; Del Río Hernández, A. Role of Extracellular Matrix in Development and Cancer Progression. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Nelson, C.M. Extracellular matrix and cytoskeletal dynamics during branching morphogenesis. Organogenesis 2012, 8, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Larsen, M.; Yamada, K.M. Fibronectin requirement in branching morphogenesis. Nature 2003, 423, 876–881. [Google Scholar] [CrossRef]
- Ortega, N.; Werb, Z. New functional roles for non-collagenous domains of basement membrane collagens. J Cell Sci 2002, 115, 4201–4214. [Google Scholar] [CrossRef] [PubMed]
- Sternlicht, M.D.; Kouros-Mehr, H.; Lu, P.; Werb, Z. Hormonal and local control of mammary branching morphogenesis. Differentiation 2006, 74, 365–381. [Google Scholar] [CrossRef]
- Yue, B. Biology of the extracellular matrix: an overview. Journal of glaucoma 2014, S20. [Google Scholar] [CrossRef] [PubMed]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: a dynamic view. Developmental biology 2010, 341, 126–140. [Google Scholar] [CrossRef]
- Walma, D.A.C.; Yamada, K.M. The extracellular matrix in development. Development 2020, 147, dev175596. [Google Scholar] [CrossRef]
- Streuli, C.H.; Schmidhauser, C.; Bailey, N.; Yurchenco, P.; Skubitz, A.P.; Roskelley, C.; Bissell, M.J. Laminin mediates tissue-specific gene expression in mammary epithelia. J Cell Biol 1995, 129, 591–603. [Google Scholar] [CrossRef]
- Muncie, J.M.; Weaver, V.M. The physical and biochemical properties of the extracellular matrix regulate cell fate. Current topics in developmental biology 2018, 130, 1–37. [Google Scholar] [PubMed]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: a dynamic microenvironment for stem cell niche. Biochim Biophys Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Dityatev, A.; Schachner, M.; Sonderegger, P. The dual role of the extracellular matrix in synaptic plasticity and homeostasis. Nature Reviews Neuroscience 2010, 11, 735–746. [Google Scholar] [CrossRef]
- Cox, T.R.; Erler, J.T. Remodeling and homeostasis of the extracellular matrix: implications for fibrotic diseases and cancer. Disease models & mechanisms 2011, 4, 165–178. [Google Scholar]
- Mongiat, M.; Andreuzzi, E.; Tarticchio, G.; Paulitti, A. Extracellular matrix, a hard player in angiogenesis. International journal of molecular sciences 2016, 17, 1822. [Google Scholar] [CrossRef]
- Zhang, D.; Zhou, C.; Wang, Q.; Cai, L.; Du, W.; Li, X.; Zhou, X.; Xie, J. Extracellular matrix elasticity regulates osteocyte gap junction elongation: involvement of paxillin in intracellular signal transduction. Cellular Physiology and Biochemistry 2018, 51, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-J.; Kang, M.H.; Ooi, Y.H.; Choi, K.R.; Sage, E.H.; Rhee, D.J. Overexpression of SPARC in human trabecular meshwork increases intraocular pressure and alters extracellular matrix. Investigative ophthalmology & visual science 2013, 54, 3309–3319. [Google Scholar]
- Terajima, M.; Taga, Y.; Cabral, W.A.; Liu, Y.; Nagasawa, M.; Sumida, N.; Kayashima, Y.; Chandrasekaran, P.; Han, L.; Maeda, N. Cyclophilin B control of lysine post-translational modifications of skin type I collagen. PLoS genetics 2019, 15, e1008196. [Google Scholar] [CrossRef]
- Reynders, M.; Matsuura, B.S.; Bérouti, M.; Simoneschi, D.; Marzio, A.; Pagano, M.; Trauner, D. PHOTACs enable optical control of protein degradation. Science advances 2020, 6, eaay5064. [Google Scholar] [CrossRef]
- Liu, J.; Chen, H.; Ma, L.; He, Z.; Wang, D.; Liu, Y.; Lin, Q.; Zhang, T.; Gray, N.; Kaniskan, H.Ü. Light-induced control of protein destruction by opto-PROTAC. Science advances 2020, 6, eaay5154. [Google Scholar] [CrossRef]
- Ryan, A.; Liu, J.; Deiters, A. Targeted protein degradation through fast optogenetic activation and its application to the control of cell signaling. Journal of the American Chemical Society 2021, 143, 9222–9229. [Google Scholar] [CrossRef] [PubMed]
- Petrie, R.J.; Gavara, N.; Chadwick, R.S.; Yamada, K.M. Nonpolarized signaling reveals two distinct modes of 3D cell migration. Journal of Cell Biology 2012, 197, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Piersma, B.; Hayward, M.-K.; Weaver, V.M. Fibrosis and cancer: A strained relationship. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer 2020, 1873, 188356. [Google Scholar] [CrossRef] [PubMed]
- Viji Babu, P.K.; Rianna, C.; Mirastschijski, U.; Radmacher, M. Nano-mechanical mapping of interdependent cell and ECM mechanics by AFM force spectroscopy. Scientific reports 2019, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Roycik, M.D.; Fang, X.; Sang, Q.X. A fresh prospect of extracellular matrix hydrolytic enzymes and their substrates. Curr Pharm Des 2009, 15, 1295–1308. [Google Scholar] [CrossRef] [PubMed]
- Myllyharju, J. Prolyl 4-hydroxylases, the key enzymes of collagen biosynthesis. Matrix Biol 2003, 22, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Xu, R. Roles of PLODs in Collagen Synthesis and Cancer Progression. Front Cell Dev Biol 2018, 6, 66. [Google Scholar] [CrossRef]
- Qi, Y.; Xu, R. Roles of PLODs in collagen synthesis and cancer progression. Frontiers in cell and developmental biology 2018, 6, 66. [Google Scholar] [CrossRef]
- Gilkes, D.M.; Bajpai, S.; Wong, C.C.; Chaturvedi, P.; Hubbi, M.E.; Wirtz, D.; Semenza, G.L. Procollagen Lysyl Hydroxylase 2 Is Essential for Hypoxia-Induced Breast Cancer MetastasisPLOD2 Is Essential for Hypoxia-Induced Metastasis. Molecular cancer research 2013, 11, 456–466. [Google Scholar] [CrossRef]
- Eddy, A.A. Molecular basis of renal fibrosis. Pediatric nephrology 2000, 15, 290–301. [Google Scholar] [CrossRef]
- Alcolado, R.; Arthur, M.; Iredale, J. Pathogenesis of liver fibrosis. Clinical science (London, England: 1979) 1997, 92, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Libring, S.; Shinde, A.; Chanda, M.K.; Nuru, M.; George, H.; Saleh, A.M.; Abdullah, A.; Kinzer-Ursem, T.L.; Calve, S.; Wendt, M.K. The dynamic relationship of breast cancer cells and fibroblasts in fibronectin accumulation at primary and metastatic tumor sites. Cancers 2020, 12, 1270. [Google Scholar] [CrossRef] [PubMed]
- Comoglio, P.M.; Trusolino, L. Cancer: the matrix is now in control. Nature medicine 2005, 11, 1156–1158. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.; Greenshields-Watson, A.; Jones, E.; Smart, K.; Lauder, S.N.; Somerville, M.; Milutinovic, S.; Kendrick, H.; Hindley, J.P.; French, R.; et al. Immune Remodeling of the Extracellular Matrix Drives Loss of Cancer Stem Cells and Tumor Rejection. Cancer Immunol Res 2020, 8, 1520–1531. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Wang, C.Y.; Werb, Z. Matrix metalloproteinases in stem cell regulation and cancer. Matrix Biol 2015, 44-46, 184–190. [Google Scholar] [CrossRef]
- Fonović, M.; Turk, B. Cysteine cathepsins and extracellular matrix degradation. Biochim Biophys Acta 2014, 1840, 2560–2570. [Google Scholar] [CrossRef]
- Cox, T.R.; Erler, J.T. Fibrosis and Cancer: Partners in Crime or Opposing Forces? Trends Cancer 2016, 2, 279–282. [Google Scholar] [CrossRef]
- Filipe, E.C.; Chitty, J.L.; Cox, T.R. Charting the unexplored extracellular matrix in cancer. Int J Exp Pathol 2018, 99, 58–76. [Google Scholar] [CrossRef]
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.T.; Fong, S.F.; Csiszar, K.; Giaccia, A.; Weninger, W.; et al. Matrix crosslinking forces tumor progression by enhancing integrin signaling. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef]
- Nicolas-Boluda, A.; Vaquero, J.; Vimeux, L.; Guilbert, T.; Barrin, S.; Kantari-Mimoun, C.; Ponzo, M.; Renault, G.; Deptula, P.; Pogoda, K.; et al. Tumor stiffening reversion through collagen crosslinking inhibition improves T cell migration and anti-PD-1 treatment. Elife 2021, 10. [Google Scholar] [CrossRef]
- Kench, J.A.; Russell, D.M.; Fadok, V.A.; Young, S.K.; Worthen, G.S.; Jones-Carson, J.; Henson, J.E.; Henson, P.M.; Nemazee, D. Aberrant wound healing and TGF-β production in the autoimmune-prone MRL/+ mouse. Clinical immunology 1999, 92, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bao, L.; Chan, L.S.; DiPietro, L.A.; Chen, L. Aberrant wound healing in an epidermal interleukin-4 transgenic mouse model of atopic dermatitis. PLoS One 2016, 11, e0146451. [Google Scholar] [CrossRef] [PubMed]
- Hertle, M.D.; Kubler, M.-D.; Leigh, I.M.; Watt, F. Aberrant integrin expression during epidermal wound healing and in psoriatic epidermis. The Journal of clinical investigation 1992, 89, 1892–1901. [Google Scholar] [CrossRef] [PubMed]
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.T.; Fong, S.F.; Csiszar, K.; Giaccia, A.; Weninger, W. Matrix crosslinking forces tumor progression by enhancing integrin signaling. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef]
- Stephens, P.; Grenard, P.; Aeschlimann, P.; Langley, M.; Blain, E.; Errington, R.; Kipling, D.; Thomas, D.; Aeschlimann, D. Crosslinking and G-protein functions of transglutaminase 2 contribute differentially to fibroblast wound healing responses. Journal of cell science 2004, 117, 3389–3403. [Google Scholar] [CrossRef] [PubMed]
- Chitty, J.L.; Setargew, Y.F.I.; Cox, T.R. Targeting the lysyl oxidases in tumour desmoplasia. Biochem Soc Trans 2019, 47, 1661–1678. [Google Scholar] [CrossRef] [PubMed]
- Ponce, I.; Garrido, N.; Tobar, N.; Melo, F.; Smith, P.C.; Martínez, J. Matrix stiffness modulates metabolic interaction between human stromal and breast cancer cells to stimulate epithelial motility. Metabolites 2021, 11, 432. [Google Scholar] [CrossRef]
- DuFort, C.C.; DelGiorno, K.E.; Hingorani, S.R. Mounting Pressure in the Microenvironment: Fluids, Solids, and Cells in Pancreatic Ductal Adenocarcinoma. Gastroenterology 2016, 150, 1545–1557. [Google Scholar] [CrossRef]
- Scarpellini, A.; Germack, R.; Lortat-Jacob, H.; Muramatsu, T.; Billett, E.; Johnson, T.; Verderio, E.A. Heparan sulfate proteoglycans are receptors for the cell-surface trafficking and biological activity of transglutaminase-2. J Biol Chem 2009, 284, 18411–18423. [Google Scholar] [CrossRef]
- Barsigian, C.; Fellin, F.M.; Jain, A.; Martinez, J. Dissociation of fibrinogen and fibronectin binding from transglutaminase-mediated cross-linking at the hepatocyte surface. J Biol Chem 1988, 263, 14015–14022. [Google Scholar] [CrossRef]
- Cardoso, I.; Stamnaes, J.; Andersen, J.T.; Melino, G.; Iversen, R.; Sollid, L.M. Transglutaminase 2 interactions with extracellular matrix proteins as probed with celiac disease autoantibodies. Febs j 2015, 282, 2063–2075. [Google Scholar] [CrossRef] [PubMed]
- Akimov, S.S.; Krylov, D.; Fleischman, L.F.; Belkin, A.M. Tissue transglutaminase is an integrin-binding adhesion coreceptor for fibronectin. J Cell Biol 2000, 148, 825–838. [Google Scholar] [CrossRef]
- Richter, P.; Junker, K.; Franz, M.; Berndt, A.; Geyer, C.; Gajda, M.; Kosmehl, H.; Berndt, A.; Wunderlich, H. IIICS de novo glycosylated fibronectin as a marker for invasiveness in urothelial carcinoma of the urinary bladder (UBC). Journal of cancer research and clinical oncology 2008, 134, 1059–1065. [Google Scholar] [CrossRef]
- Freire-de-Lima, L.; Gelfenbeyn, K.; Ding, Y.; Mandel, U.; Clausen, H.; Handa, K.; Hakomori, S.-i. Involvement of O-glycosylation defining oncofetal fibronectin in epithelial-mesenchymal transition process. Proceedings of the National Academy of Sciences 2011, 108, 17690–17695. [Google Scholar] [CrossRef] [PubMed]
- SUZUkI, O.; Abe, M.; Hashimoto, Y. Sialylation and glycosylation modulate cell adhesion and invasion to extracellular matrix in human malignant lymphoma: Dependency on integrin and the Rho GTPase family. International Journal of Oncology 2015, 47, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.; Shyanti, R.K.; Singh, V.; Kale, R.K.; Mishra, J.P.; Singh, R.P. Integrin expression and glycosylation patterns regulate cell-matrix adhesion and alter with breast cancer progression. Biochemical and biophysical research communications 2018, 499, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, R.; Alahari, S.K. Important role of integrins in the cancer biology. Cancer and Metastasis Reviews 2010, 29, 223–237. [Google Scholar] [CrossRef]
- Yalak, G.; Vogel, V. Ectokinases as novel cancer markers and drug targets in cancer therapy. Cancer medicine 2015, 4, 404–414. [Google Scholar] [CrossRef]
- Yalak, G.; Shiu, J.-Y.; Schoen, I.; Mitsi, M.; Vogel, V. Phosphorylated fibronectin enhances cell attachment and upregulates mechanical cell functions. PloS one 2019, 14, e0218893. [Google Scholar] [CrossRef]
- Wolanska, K.I.; Morgan, M.R. Fibronectin remodelling: cell-mediated regulation of the microenvironment; Portland Press Ltd., 2015. [Google Scholar] [CrossRef]
- Soares Da Costa, D.; Reis, R.L.; Pashkuleva, I. Sulfation of glycosaminoglycans and its implications in human health and disorders. Annual review of biomedical engineering 2017, 19, 1–26. [Google Scholar] [CrossRef]
- Escobar Galvis, M.L.; Jia, J.; Zhang, X.; Jastrebova, N.; Spillmann, D.; Gottfridsson, E.; Van Kuppevelt, T.H.; Zcharia, E.; Vlodavsky, I.; Lindahl, U. Transgenic or tumor-induced expression of heparanase upregulates sulfation of heparan sulfate. Nature chemical biology 2007, 3, 773–778. [Google Scholar] [CrossRef]
- Vlodavsky, I.; Gross-Cohen, M.; Weissmann, M.; Ilan, N.; Sanderson, R.D. Opposing functions of heparanase-1 and heparanase-2 in cancer progression. Trends in biochemical sciences 2018, 43, 18–31. [Google Scholar] [CrossRef]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nature reviews Molecular cell biology 2014, 15, 802–812. [Google Scholar] [CrossRef]
- Hamacher, S.; Matern, S.; Roeb, E. Extracellular matrix--from basic research to clinical significance. An overview with special consideration of matrix metalloproteinases. Deutsche Medizinische Wochenschrift (1946) 2004, 129, 1976–1980. [Google Scholar] [CrossRef]
- Toth, M.; Fridman, R. Assessment of gelatinases (MMP-2 and MMP-9 by gelatin zymography. In Metastasis research protocols; Springer, 2001; pp. 163–174. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Y.; Gao, S.; Zhou, M.; Qi, F.; Ding, N.; Zhang, J.; Li, R.; Wang, J.; Shi, J. Excessive DNA damage mediates ECM degradation via the RBBP8/NOTCH1 pathway in sporadic aortic dissection. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2022, 1868, 166303. [Google Scholar] [CrossRef]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harbor perspectives in biology 2011, 3, a005058. [Google Scholar] [CrossRef]
- Stetler-Stevenson, W.G.; Liotta, L.A.; Kleiner Jr, D.E. Extracellular matrix 6: role of matrix metalloproteinases in tumor invasion and metastasis. The FASEB Journal 1993, 7, 1434–1441. [Google Scholar] [CrossRef]
- Yuzhalin, A.E.; Lim, S.Y.; Kutikhin, A.G.; Gordon-Weeks, A.N. Dynamic matrisome: ECM remodeling factors licensing cancer progression and metastasis. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer 2018, 1870, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.; Cena, J.; Schulz, R. Acute actions and novel targets of matrix metalloproteinases in the heart and vasculature. British journal of pharmacology 2007, 152, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Mott, J.D.; Werb, Z. Regulation of matrix biology by matrix metalloproteinases. Curr Opin Cell Biol 2004, 16, 558–564. [Google Scholar] [CrossRef]
- Bergers, G.; Brekken, R.; McMahon, G.; Vu, T.H.; Itoh, T.; Tamaki, K.; Tanzawa, K.; Thorpe, P.; Itohara, S.; Werb, Z.; et al. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat Cell Biol 2000, 2, 737–744. [Google Scholar] [CrossRef]
- Siqueira, A.S.; Carvalho, M.R.; Monteiro, A.C.; Freitas, V.M.; Jaeger, R.G.; Pinheiro, J.J. Matrix metalloproteinases, TIMPs and growth factors regulating ameloblastoma behaviour. Histopathology 2010, 57, 128–137. [Google Scholar] [CrossRef]
- Rodríguez, D.; Morrison, C.J.; Overall, C.M. Matrix metalloproteinases: What do they not do? New substrates and biological roles identified by murine models and proteomics. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 2010, 1803, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc Res 2006, 69, 562–573. [Google Scholar] [CrossRef]
- Hamacher, S.; Matern, S.; Roeb, E. [Extracellular matrix -- from basic research to clinical significance. An overview with special consideration of matrix metalloproteinases]. Dtsch Med Wochenschr 2004, 129, 1976–1980. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Ohtsuka, T.; Okada, Y.; Kanai, Y. Stromal metalloproteinases: Crucial contributors to the tumor microenvironment. Pathol Int 2021, 71, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat Commun 2020, 11, 5120. [Google Scholar] [CrossRef]
- Mittal, R.; Patel, A.P.; Debs, L.H.; Nguyen, D.; Patel, K.; Grati, M.h.; Mittal, J.; Yan, D.; Chapagain, P.; Liu, X.Z. Intricate functions of matrix metalloproteinases in physiological and pathological conditions. Journal of cellular physiology 2016, 231, 2599–2621. [Google Scholar] [CrossRef]
- Jacob, M.P. Extracellular matrix remodeling and matrix metalloproteinases in the vascular wall during aging and in pathological conditions. Biomedicine & pharmacotherapy 2003, 57, 195–202. [Google Scholar]
- Stadlmann, S.; Pollheimer, J.; Moser, P.L.; Raggi, A.; Amberger, A.; Margreiter, R.; Offner, F.A.; Mikuz, G.; Dirnhofer, S.; Moch, H. Cytokine-regulated expression of collagenase-2 (MMP-8) is involved in the progression of ovarian cancer. Eur J Cancer 2003, 39, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Åström, P.; Juurikka, K.; Hadler-Olsen, E.S.; Svineng, G.; Cervigne, N.K.; Coletta, R.D.; Risteli, J.; Kauppila, J.H.; Skarp, S.; Kuttner, S.; et al. The interplay of matrix metalloproteinase-8, transforming growth factor-β1 and vascular endothelial growth factor-C cooperatively contributes to the aggressiveness of oral tongue squamous cell carcinoma. Br J Cancer 2017, 117, 1007–1016. [Google Scholar] [CrossRef]
- Raeeszadeh-Sarmazdeh, M.; Do, L.D.; Hritz, B.G. Metalloproteinases and their inhibitors: potential for the development of new therapeutics. Cells 2020, 9, 1313. [Google Scholar] [CrossRef]
- Markland, F.S. Snake venoms and the hemostatic system. Toxicon 1998, 36, 1749–1800. [Google Scholar] [CrossRef]
- Rossello, A.; Nuti, E.; Ferrini, S.; Fabbi, M. Targeting ADAM17 Sheddase Activity in Cancer. Curr Drug Targets 2016, 17, 1908–1927. [Google Scholar] [CrossRef]
- Van Goor, H.; Melenhorst, W.B.; Turner, A.J.; Holgate, S.T. Adamalysins in biology and disease. The Journal of Pathology: A Journal of the Pathological Society of Great Britain and Ireland 2009, 219, 277–286. [Google Scholar] [CrossRef]
- Killar, L.; White, J.; Black, R.; Peschon, J. Adamalysins: a family of metzincins including TNF-α converting enzyme (TACE). Annals of the New York Academy of Sciences 1999, 878, 442–452. [Google Scholar] [CrossRef]
- Turk, V.; Stoka, V.; Vasiljeva, O.; Renko, M.; Sun, T.; Turk, B.; Turk, D. Cysteine cathepsins: from structure, function and regulation to new frontiers. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics 2012, 1824, 68–88. [Google Scholar] [CrossRef]
- Conus, S. Cathepsins and their involvement in immune responses. Swiss medical weekly 2010. [Google Scholar] [CrossRef]
- Yadati, T.; Houben, T.; Bitorina, A.; Shiri-Sverdlov, R. The ins and outs of cathepsins: physiological function and role in disease management. Cells 2020, 9, 1679. [Google Scholar] [CrossRef]
- Ishidoh, K.; Kominami, E. Procathepsin L degrades extracellular matrix proteins in the presence of glycosaminoglycans in vitro. Biochem Biophys Res Commun 1995, 217, 624–631. [Google Scholar] [CrossRef]
- Taleb, S.; Cancello, R.; Clément, K.; Lacasa, D. Cathepsin s promotes human preadipocyte differentiation: possible involvement of fibronectin degradation. Endocrinology 2006, 147, 4950–4959. [Google Scholar] [CrossRef]
- Nomura, T.; Katunuma, N. Involvement of cathepsins in the invasion, metastasis and proliferation of cancer cells. The journal of medical investigation 2005, 52, 1–9. [Google Scholar] [CrossRef]
- Tan, G.-J.; Peng, Z.-K.; Lu, J.-P.; Tang, F.-Q. Cathepsins mediate tumor metastasis. World journal of biological chemistry 2013, 4, 91. [Google Scholar] [CrossRef]
- Rudzińska, M.; Parodi, A.; Soond, S.M.; Vinarov, A.Z.; Korolev, D.O.; Morozov, A.O.; Daglioglu, C.; Tutar, Y.; Zamyatnin, A.A. The role of cysteine cathepsins in cancer progression and drug resistance. International journal of molecular sciences 2019, 20, 3602. [Google Scholar] [CrossRef]
- Llorens, F.; Thüne, K.; Sikorska, B.; Schmitz, M.; Tahir, W.; Fernández-Borges, N.; Cramm, M.; Gotzmann, N.; Carmona, M.; Streichenberger, N. Altered Ca2+ homeostasis induces Calpain-Cathepsin axis activation in sporadic Creutzfeldt-Jakob disease. Acta neuropathologica communications 2017, 5, 1–20. [Google Scholar] [CrossRef]
- Büth, H.; Buttigieg, P.L.; Ostafe, R.; Rehders, M.; Dannenmann, S.R.; Schaschke, N.; Stark, H.-J.; Boukamp, P.; Brix, K. Cathepsin B is essential for regeneration of scratch-wounded normal human epidermal keratinocytes. European journal of cell biology 2007, 86, 747–761. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.R.; Keles, S.; Greenspan, D.S. The bone morphogenetic protein 1/Tolloid-like metalloproteinases. Matrix Biology 2007, 26, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Steiglitz, B.M.; Ayala, M.; Narayanan, K.; George, A.; Greenspan, D.S. Bone morphogenetic protein-1/Tolloid-like proteinases process dentin matrix protein-1. Journal of Biological Chemistry 2004, 279, 980–986. [Google Scholar] [CrossRef]
- Vadon-Le Goff, S.; Hulmes, D.J.; Moali, C. BMP-1/tolloid-like proteinases synchronize matrix assembly with growth factor activation to promote morphogenesis and tissue remodeling. Matrix Biol 2015, 44-46, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Ge, G.; Greenspan, D.S. Developmental roles of the BMP1/TLD metalloproteinases. Birth Defects Res C Embryo Today 2006, 78, 47–68. [Google Scholar] [CrossRef]
- Vadon-Le Goff, S.; Hulmes, D.J.; Moali, C. BMP-1/tolloid-like proteinases synchronize matrix assembly with growth factor activation to promote morphogenesis and tissue remodeling. Matrix Biology 2015, 44, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Malecaze, F.; Massoudi, D.; Fournié, P.; Tricoire, C.; Cassagne, M.; Malbouyres, M.; Hulmes, D.J.; Moali, C.; Galiacy, S.D. Upregulation of bone morphogenetic protein-1/mammalian tolloid and procollagen C-proteinase enhancer-1 in corneal scarring. Investigative Ophthalmology & Visual Science 2014, 55, 6712–6721. [Google Scholar]
- Muir, A.M.; Massoudi, D.; Nguyen, N.; Keene, D.R.; Lee, S.-J.; Birk, D.E.; Davidson, J.M.; Marinkovich, M.P.; Greenspan, D.S. BMP1-like proteinases are essential to the structure and wound healing of skin. Matrix Biology 2016, 56, 114–131. [Google Scholar] [CrossRef]
- Stern, R. Hyaluronidases in cancer biology. Semin Cancer Biol 2008, 18, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Tolg, C.; Turley, E. Dissecting the Dual Nature of Hyaluronan in the Tumor Microenvironment. Front Immunol 2019, 10, 947. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Yamamoto, H.; Tobisawa, Y.; Irie, F. TMEM2: A missing link in hyaluronan catabolism identified? Matrix Biol 2019, 78-79, 139–146. [Google Scholar] [CrossRef]
- Tammi, M.I.; Oikari, S.; Pasonen-Seppänen, S.; Rilla, K.; Auvinen, P.; Tammi, R.H. Activated hyaluronan metabolism in the tumor matrix - Causes and consequences. Matrix Biol 2019, 78-79, 147–164. [Google Scholar] [CrossRef]
- Stern, R. Hyaluronan metabolism: a major paradox in cancer biology. Pathologie Biologie 2005, 53, 372–382. [Google Scholar] [CrossRef]
- Girish, K.; Kemparaju, K. The magic glue hyaluronan and its eraser hyaluronidase: a biological overview. Life sciences 2007, 80, 1921–1943. [Google Scholar] [CrossRef]
- Stern, R.; Asari, A.A.; Sugahara, K.N. Hyaluronan fragments: an information-rich system. European journal of cell biology 2006, 85, 699–715. [Google Scholar] [CrossRef]
- Jiang, D.; Liang, J.; Noble, P.W. Hyaluronan in Tissue Injury and Repair. Annual Review of Cell and Developmental Biology 2007, 23, 435–461. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Chanmee, T.; Itano, N. Hyaluronan: Metabolism and function. Biomolecules 2020, 10, 1525. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Liang, J.; Noble, P.W. Hyaluronan as an immune regulator in human diseases. Physiological reviews 2011, 91, 221–264. [Google Scholar] [CrossRef] [PubMed]
- Noble, P.W. Hyaluronan and its catabolic products in tissue injury and repair. Matrix biology 2002, 21, 25–29. [Google Scholar] [CrossRef]
- Heldin, P.; Basu, K.; Olofsson, B.; Porsch, H.; Kozlova, I.; Kahata, K. Deregulation of hyaluronan synthesis, degradation and binding promotes breast cancer. The Journal of Biochemistry 2013, 154, 395–408. [Google Scholar] [CrossRef]
- Bame, K.J. Heparanases: endoglycosidases that degrade heparan sulfate proteoglycans. Glycobiology 2001, 11, 91R–98R. [Google Scholar] [CrossRef]
- Zcharia, E.; Zilka, R.; Yaar, A.; Yacoby-Zeevi, O.; Zetser, A.; Metzger, S.; Sarid, R.; Naggi, A.; Casu, B.; Ilan, N. Heparanase accelerates wound angiogenesis and wound healing in mouse and rat models. The FASEB journal 2005, 19, 211–221. [Google Scholar] [CrossRef]
- Crispel, Y.; Ghanem, S.; Attias, J.; Kogan, I.; Brenner, B.; Nadir, Y. Involvement of the heparanase procoagulant domain in bleeding and wound healing. Journal of Thrombosis and Haemostasis 2017, 15, 1463–1472. [Google Scholar] [CrossRef]
- Sanderson, R.D.; Elkin, M.; Rapraeger, A.C.; Ilan, N.; Vlodavsky, I. Heparanase regulation of cancer, autophagy and inflammation: new mechanisms and targets for therapy. Febs j 2017, 284, 42–55. [Google Scholar] [CrossRef]
- Khanna, M.; Parish, C.R. Heparanase: Historical Aspects and Future Perspectives. Adv Exp Med Biol 2020, 1221, 71–96. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Wang, C.-Y.; Werb, Z. Matrix metalloproteinases in stem cell regulation and cancer. Matrix biology 2015, 44, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y. Membrane-type matrix metalloproteinases: Their functions and regulations. Matrix Biology 2015, 44, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovascular research 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: structure, function, and biochemistry. Circulation research 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Brew, K.; Nagase, H. The tissue inhibitors of metalloproteinases (TIMPs): an ancient family with structural and functional diversity. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 2010, 1803, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.A.; Leaper, D.J. Profiles of matrix metalloproteinases and their tissue inhibitors in intraperitoneal drainage fluid: relationship to wound healing. Wound Repair and Regeneration 2003, 11, 268–274. [Google Scholar] [CrossRef]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv Wound Care (New Rochelle) 2015, 4, 119–136. [Google Scholar] [CrossRef]
- Breznik, B.; Mitrović, A.; Lah, T.T.; Kos, J. Cystatins in cancer progression: More than just cathepsin inhibitors. Biochimie 2019, 166, 233–250. [Google Scholar] [CrossRef]
- van Gent, D.; Sharp, P.; Morgan, K.; Kalsheker, N. Serpins: structure, function and molecular evolution. The international journal of biochemistry & cell biology 2003, 35, 1536–1547. [Google Scholar]
- Rau, J.; Beaulieu, L.; Huntington, J.; CHURCH, F.C. Serpins in thrombosis, hemostasis and fibrinolysis. Journal of thrombosis and haemostasis 2007, 5, 102–115. [Google Scholar] [CrossRef]
- Carrell, R.W.; Evans, D.L.; Stein, P.E. Mobile reactive centre of serpins and the control of thrombosis. Nature 1991, 353, 576–578. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, L. The impact of the extracellular matrix on inflammation. Nat Rev Immunol 2010, 10, 712–723. [Google Scholar] [CrossRef]
- Ricard-Blum, S.; Vallet, S.D. Proteases decode the extracellular matrix cryptome. Biochimie 2016, 122, 300–313. [Google Scholar] [CrossRef]
- Lee, J.H.; Isayeva, T.; Larson, M.R.; Sawant, A.; Cha, H.R.; Chanda, D.; Chesnokov, I.N.; Ponnazhagan, S. Endostatin: A novel inhibitor of androgen receptor function in prostate cancer. Proc Natl Acad Sci U S A 2015, 112, 1392–1397. [Google Scholar] [CrossRef]
- Magnon, C.; Galaup, A.; Mullan, B.; Rouffiac, V.; Bouquet, C.; Bidart, J.M.; Griscelli, F.; Opolon, P.; Perricaudet, M. Canstatin acts on endothelial and tumor cells via mitochondrial damage initiated through interaction with alphavbeta3 and alphavbeta5 integrins. Cancer Res 2005, 65, 4353–4361. [Google Scholar] [CrossRef]
- Wang, S.; Lu, X.A.; Liu, P.; Fu, Y.; Jia, L.; Zhan, S.; Luo, Y. Endostatin has ATPase activity, which mediates its antiangiogenic and antitumor activities. Mol Cancer Ther 2015, 14, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Colorado, P.C.; Torre, A.; Kamphaus, G.; Maeshima, Y.; Hopfer, H.; Takahashi, K.; Volk, R.; Zamborsky, E.D.; Herman, S.; Sarkar, P.K.; et al. Anti-angiogenic cues from vascular basement membrane collagen. Cancer Res 2000, 60, 2520–2526. [Google Scholar] [PubMed]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361. [Google Scholar] [CrossRef]
- Chandler, C.; Liu, T.; Buckanovich, R.; Coffman, L.G. The double edge sword of fibrosis in cancer. Transl Res 2019, 209, 55–67. [Google Scholar] [CrossRef]
- Foster, D.S.; Jones, R.E.; Ransom, R.C.; Longaker, M.T.; Norton, J.A. The evolving relationship of wound healing and tumor stroma. JCI insight 2018, 3. [Google Scholar] [CrossRef]
- Dvorak, H.F. Tumors: wounds that do not heal—a historical perspective with a focus on the fundamental roles of increased vascular permeability and clotting. in Seminars in thrombosis and hemostasis. 2019. Thieme Medical Publishers. [CrossRef]
- Dzobo, K. Taking a Full Snapshot of Cancer Biology: Deciphering the Tumor Microenvironment for Effective Cancer Therapy in the Oncology Clinic. Omics 2020. [Google Scholar] [CrossRef]
- Dzobo, K.; Dandara, C. Architecture of Cancer-Associated Fibroblasts in Tumor Microenvironment: Mapping Their Origins, Heterogeneity, and Role in Cancer Therapy Resistance. Omics 2020, 24, 314–339. [Google Scholar] [CrossRef]
- Dzobo, K.; Dandara, C. Broadening Drug Design and Targets to Tumor Microenvironment? Cancer-Associated Fibroblast Marker Expression in Cancers and Relevance for Survival Outcomes. Omics 2020, 24, 340–351. [Google Scholar] [CrossRef]
- Bochet, L.; Lehuédé, C.; Dauvillier, S.; Wang, Y.Y.; Dirat, B.; Laurent, V.; Dray, C.; Guiet, R.; Maridonneau-Parini, I.; Le Gonidec, S.; et al. Adipocyte-derived fibroblasts promote tumor progression and contribute to the desmoplastic reaction in breast cancer. Cancer Res 2013, 73, 5657–5668. [Google Scholar] [CrossRef] [PubMed]
- Mu, W.; Rana, S.; Zöller, M. Host matrix modulation by tumor exosomes promotes motility and invasiveness. Neoplasia 2013, 15, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Webber, J.P.; Spary, L.K.; Sanders, A.J.; Chowdhury, R.; Jiang, W.G.; Steadman, R.; Wymant, J.; Jones, A.T.; Kynaston, H.; Mason, M.D.; et al. Differentiation of tumour-promoting stromal myofibroblasts by cancer exosomes. Oncogene 2015, 34, 290–302. [Google Scholar] [CrossRef]
- Liu, L.; Liu, L.; Yao, H.H.; Zhu, Z.Q.; Ning, Z.L.; Huang, Q. Stromal Myofibroblasts Are Associated with Poor Prognosis in Solid Cancers: A Meta-Analysis of Published Studies. PLoS One 2016, 11, e0159947. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Ogawa, T.; Zhang, X.; Hanamura, N.; Kashikura, Y.; Takamura, M.; Yoneda, M.; Shiraishi, T. Role of stromal myofibroblasts in invasive breast cancer: stromal expression of alpha-smooth muscle actin correlates with worse clinical outcome. Breast Cancer 2012, 19, 170–176. [Google Scholar] [CrossRef]
- Tsujino, T.; Seshimo, I.; Yamamoto, H.; Ngan, C.Y.; Ezumi, K.; Takemasa, I.; Ikeda, M.; Sekimoto, M.; Matsuura, N.; Monden, M. Stromal myofibroblasts predict disease recurrence for colorectal cancer. Clin Cancer Res 2007, 13, 2082–2090. [Google Scholar] [CrossRef]
- Liu, C.; Mak, M. Fibroblast-mediated uncaging of cancer cells and dynamic evolution of the physical microenvironment. Sci Rep 2022, 12, 791. [Google Scholar] [CrossRef] [PubMed]
- Öhlund, D.; Handly-Santana, A.; Biffi, G.; Elyada, E.; Almeida, A.S.; Ponz-Sarvise, M.; Corbo, V.; Oni, T.E.; Hearn, S.A.; Lee, E.J.; et al. Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. J Exp Med 2017, 214, 579–596. [Google Scholar] [CrossRef] [PubMed]
- Elyada, E.; Bolisetty, M.; Laise, P.; Flynn, W.F.; Courtois, E.T.; Burkhart, R.A.; Teinor, J.A.; Belleau, P.; Biffi, G.; Lucito, M.S.; et al. Cross-Species Single-Cell Analysis of Pancreatic Ductal Adenocarcinoma Reveals Antigen-Presenting Cancer-Associated Fibroblasts. Cancer Discov 2019, 9, 1102–1123. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Öhlund, D.; Rickelt, S.; Lidström, T.; Huang, Y.; Hao, L.; Zhao, R.T.; Franklin, O.; Bhatia, S.N.; Tuveson, D.A.; et al. Cancer Cell–Derived Matrisome Proteins Promote Metastasis in Pancreatic Ductal Adenocarcinoma. Cancer Research 2020, 80, 1461–1474. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.B.; Chua, M.L.K.; Yeong, J.P.S.; Tan, S.J.; Lim, W.-T.; Lim, C.T. Pan-cancer analysis connects tumor matrisome to immune response. NPJ precision oncology 2019, 3, 1–9. [Google Scholar] [CrossRef]
- Rafaeva, M.; Erler, J.T. Framing cancer progression: influence of the organ-and tumour-specific matrisome. The FEBS journal 2020, 287, 1454–1477. [Google Scholar] [CrossRef]
- Le, C.P.; Nowell, C.J.; Kim-Fuchs, C.; Botteri, E.; Hiller, J.G.; Ismail, H.; Pimentel, M.A.; Chai, M.G.; Karnezis, T.; Rotmensz, N.; et al. Chronic stress in mice remodels lymph vasculature to promote tumour cell dissemination. Nat Commun 2016, 7, 10634. [Google Scholar] [CrossRef]
- Tacconi, C.; Correale, C.; Gandelli, A.; Spinelli, A.; Dejana, E.; D'Alessio, S.; Danese, S. Vascular endothelial growth factor C disrupts the endothelial lymphatic barrier to promote colorectal cancer invasion. Gastroenterology 2015, 148, 1438–1451. [Google Scholar] [CrossRef]
- Le, C.P.; Karnezis, T.; Achen, M.G.; Stacker, S.A.; Sloan, E.K. Lymphovascular and neural regulation of metastasis: shared tumour signalling pathways and novel therapeutic approaches. Best Pract Res Clin Anaesthesiol 2013, 27, 409–425. [Google Scholar] [CrossRef]
- Nagaraja, A.S.; Dood, R.L.; Armaiz-Pena, G.; Kang, Y.; Wu, S.Y.; Allen, J.K.; Jennings, N.B.; Mangala, L.S.; Pradeep, S.; Lyons, Y.; et al. Adrenergic-mediated increases in INHBA drive CAF phenotype and collagens. JCI Insight 2017, 2. [Google Scholar] [CrossRef]
- Insua-Rodríguez, J.; Pein, M.; Hongu, T.; Meier, J.; Descot, A.; Lowy, C.M.; De Braekeleer, E.; Sinn, H.P.; Spaich, S.; Sütterlin, M.; et al. Stress signaling in breast cancer cells induces matrix components that promote chemoresistant metastasis. EMBO Mol Med 2018, 10. [Google Scholar] [CrossRef]
- Afasizheva, A.; Devine, A.; Tillman, H.; Fung, K.L.; Vieira, W.D.; Blehm, B.H.; Kotobuki, Y.; Busby, B.; Chen, E.I.; Tanner, K. Mitogen-activated protein kinase signaling causes malignant melanoma cells to differentially alter extracellular matrix biosynthesis to promote cell survival. BMC Cancer 2016, 16, 186. [Google Scholar] [CrossRef]
- Steins, A.; van Mackelenbergh, M.G.; van der Zalm, A.P.; Klaassen, R.; Serrels, B.; Goris, S.G.; Kocher, H.M.; Waasdorp, C.; de Jong, J.H.; Tekin, C.; et al. High-grade mesenchymal pancreatic ductal adenocarcinoma drives stromal deactivation through CSF-1. EMBO Rep 2020, 21, e48780. [Google Scholar] [CrossRef]
- Vera, R.E.; Fernandez-Zapico, M.E. Stromal deactivation by CSF1: a new feature of the aggressive pancreatic cancer microenvironment. EMBO Rep 2020, 21, e50468. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, S.; Zeng, S.; Shen, H. The critical roles of activated stellate cells-mediated paracrine signaling, metabolism and onco-immunology in pancreatic ductal adenocarcinoma. Mol Cancer 2018, 17, 62. [Google Scholar] [CrossRef]
- Wei, L.; Ye, H.; Li, G.; Lu, Y.; Zhou, Q.; Zheng, S.; Lin, Q.; Liu, Y.; Li, Z.; Chen, R. Cancer-associated fibroblasts promote progression and gemcitabine resistance via the SDF-1/SATB-1 pathway in pancreatic cancer. Cell Death Dis 2018, 9, 1065. [Google Scholar] [CrossRef] [PubMed]
- Palumbo Jr, A.; Ferreira, L.B.; de Souza, P.A.R.; de Oliveira, F.L.; Pontes, B.; Viana, N.B.; Machado, D.E.; Palmero, C.Y.; Alves, L.M.; Gimba, E.R. Extracellular matrix secreted by reactive stroma is a main inducer of pro-tumorigenic features on LNCaP prostate cancer cells. Cancer letters 2012, 321, 55–64. [Google Scholar] [CrossRef]
- Barcus, C.E.; Holt, E.C.; Keely, P.J.; Eliceiri, K.W.; Schuler, L.A. Dense collagen-I matrices enhance pro-tumorigenic estrogen-prolactin crosstalk in MCF-7 and T47D breast cancer cells. PloS one 2015, 10, e0116891. [Google Scholar] [CrossRef]
- Brown, Y.; Hua, S.; Tanwar, P.S. Extracellular matrix-mediated regulation of cancer stem cells and chemoresistance. The international journal of biochemistry & cell biology 2019, 109, 90–104. [Google Scholar]
- Yeldag, G.; Rice, A.; del Río Hernández, A. Chemoresistance and the self-maintaining tumor microenvironment. Cancers 2018, 10, 471. [Google Scholar] [CrossRef]
- Pietilä, E.A.; Gonzalez-Molina, J.; Moyano-Galceran, L.; Jamalzadeh, S.; Zhang, K.; Lehtinen, L.; Turunen, S.P.; Martins, T.A.; Gultekin, O.; Lamminen, T. Co-evolution of matrisome and adaptive adhesion dynamics drives ovarian cancer chemoresistance. Nature Communications 2021, 12, 1–19. [Google Scholar] [CrossRef]
- Keeratichamroen, S.; Lirdprapamongkol, K.; Svasti, J. Mechanism of ECM-induced dormancy and chemoresistance in A549 human lung carcinoma cells. Oncology reports 2018, 39, 1765–1774. [Google Scholar] [CrossRef]
- Mocanu, M.M.; Fazekas, Z.; Petrás, M.; Nagy, P.; Sebestyén, Z.; Isola, J.; Tímár, J.; Park, J.W.; Vereb, G.; Szöllosi, J. Associations of ErbB2, beta1-integrin and lipid rafts on Herceptin (Trastuzumab) resistant and sensitive tumor cell lines. Cancer Lett 2005, 227, 201–212. [Google Scholar] [CrossRef]
- Guo, W.; Pylayeva, Y.; Pepe, A.; Yoshioka, T.; Muller, W.J.; Inghirami, G.; Giancotti, F.G. Beta 4 integrin amplifies ErbB2 signaling to promote mammary tumorigenesis. Cell 2006, 126, 489–502. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Sleiman, M.; Moriarty, T.; Herrick, W.G.; Peyton, S.R. Sorafenib resistance and JNK signaling in carcinoma during extracellular matrix stiffening. Biomaterials 2014, 35, 5749–5759. [Google Scholar] [CrossRef]
- Keely, P.J. Mechanisms by which the extracellular matrix and integrin signaling act to regulate the switch between tumor suppression and tumor promotion. Journal of mammary gland biology and neoplasia 2011, 16, 205–219. [Google Scholar] [CrossRef]
- Weigelt, B.; Lo, A.T.; Park, C.C.; Gray, J.W.; Bissell, M.J. HER2 signaling pathway activation and response of breast cancer cells to HER2-targeting agents is dependent strongly on the 3D microenvironment. Breast Cancer Res Treat 2010, 122, 35–43. [Google Scholar] [CrossRef]
- Dzobo, K. Integrins Within the Tumor Microenvironment: Biological Functions, Importance for Molecular Targeting, and Cancer Therapeutics Innovation. Omics 2021, 25, 417–430. [Google Scholar] [CrossRef]
- Dzobo, K.; Vogelsang, M.; Parker, M.I. Wnt/β-Catenin and MEK-ERK Signaling are Required for Fibroblast-Derived Extracellular Matrix-Mediated Endoderm Differentiation of Embryonic Stem Cells. Stem Cell Rev Rep 2015, 11, 761–773. [Google Scholar] [CrossRef]
- Guerrero, P.A.; McCarty, J.H. Integrins in Vascular Development and Pathology. Adv Pharmacol 2018, 81, 129–153. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.R.W.; Jacquemet, G. Cell matrix adhesion in cell migration. Essays Biochem 2019, 63, 535–551. [Google Scholar] [CrossRef]
- Zhang, Y.; Reif, G.; Wallace, D.P. Extracellular matrix, integrins, and focal adhesion signaling in polycystic kidney disease. Cell Signal 2020, 72, 109646. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, H.; Ivaska, J. Every step of the way: integrins in cancer progression and metastasis. Nature Reviews Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Samaržija, I.; Dekanić, A.; Humphries, J.D.; Paradžik, M.; Stojanović, N.; Humphries, M.J.; Ambriović-Ristov, A. Integrin crosstalk contributes to the complexity of signalling and unpredictable cancer cell fates. Cancers 2020, 12, 1910. [Google Scholar] [CrossRef] [PubMed]
- Varner, J.A.; Emerson, D.A.; Juliano, R.L. Integrin alpha 5 beta 1 expression negatively regulates cell growth: reversal by attachment to fibronectin. Molecular biology of the cell 1995, 6, 725–740. [Google Scholar] [CrossRef] [PubMed]
- Stoeltzing, O.; Liu, W.; Reinmuth, N.; Fan, F.; Parry, G.C.; Parikh, A.A.; McCarty, M.F.; Bucana, C.D.; Mazar, A.P.; Ellis, L.M. Inhibition of integrin α5β1 function with a small peptide (ATN-161) plus continuous 5-FU infusion reduces colorectal liver metastases and improves survival in mice. International journal of cancer 2003, 104, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Varner, J. Integrins: roles in cancer development and as treatment targets. British journal of cancer 2004, 90, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Varner, J.A.; Cheresh, D.A. Integrins and cancer. Current opinion in cell biology 1996, 8, 724–730. [Google Scholar] [CrossRef]
- Weis, S.M.; Cheresh, D.A. αV integrins in angiogenesis and cancer. Cold Spring Harbor perspectives in medicine 2011, 1, a006478. [Google Scholar] [CrossRef]
- Janiszewska, M.; Primi, M.C.; Izard, T. Cell adhesion in cancer: Beyond the migration of single cells. Journal of Biological Chemistry 2020, 295, 2495–2505. [Google Scholar] [CrossRef]
- Truong, H.; Danen, E.H. Integrin switching modulates adhesion dynamics and cell migration. Cell adhesion & migration 2009, 3, 179–181. [Google Scholar]
- Zuidema, A.; Wang, W.; Sonnenberg, A. Crosstalk between cell adhesion complexes in regulation of mechanotransduction. Bioessays 2020, 42, 2000119. [Google Scholar] [CrossRef]
- Kerrisk, M.E.; Cingolani, L.A.; Koleske, A.J. ECM receptors in neuronal structure, synaptic plasticity, and behavior. Progress in brain research 2014, 214, 101–131. [Google Scholar]
- Boraschi-Diaz, I.; Wang, J.; Mort, J.S.; Komarova, S.V. Collagen type I as a ligand for receptor-mediated signaling. Frontiers in Physics 2017, 5, 12. [Google Scholar] [CrossRef]
- Takai, K.; Drain, A.P.; Lawson, D.A.; Littlepage, L.E.; Karpuj, M.; Kessenbrock, K.; Le, A.; Inoue, K.; Weaver, V.M.; Werb, Z. Discoidin domain receptor 1 (DDR1) ablation promotes tissue fibrosis and hypoxia to induce aggressive basal-like breast cancers. Genes & development 2018, 32, 244–257. [Google Scholar]
- Gonzalez, M.E.; Martin, E.E.; Anwar, T.; Arellano-Garcia, C.; Medhora, N.; Lama, A.; Chen, Y.-C.; Tanager, K.S.; Yoon, E.; Kidwell, K.M. Mesenchymal stem cell-induced DDR2 mediates stromal-breast cancer interactions and metastasis growth. Cell reports 2017, 18, 1215–1228. [Google Scholar] [CrossRef]
- Bayer, S.V.; Grither, W.R.; Brenot, A.; Hwang, P.Y.; Barcus, C.E.; Ernst, M.; Pence, P.; Walter, C.; Pathak, A.; Longmore, G.D. DDR2 controls breast tumor stiffness and metastasis by regulating integrin mediated mechanotransduction in CAFs. Elife 2019, 8, e45508. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.-C.; Yen, H.-R.; Chen, C.-H.; Chu, Y.-H.; Song, Y.-C.; Tseng, T.-J.; Liu, C.-H. CHPF promotes malignancy of breast cancer cells by modifying syndecan-4 and the tumor microenvironment. American journal of cancer research 2021, 11, 812. [Google Scholar] [PubMed]
- Leblanc, R.; Sahay, D.; Houssin, A.; Machuca-Gayet, I.; Peyruchaud, O. Autotaxin-β interaction with the cell surface via syndecan-4 impacts on cancer cell proliferation and metastasis. Oncotarget 2018, 9, 33170. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, T.; Li, R.; Sy, M.-S. Mechanisms regulating the binding activity of CD44 to hyaluronic acid. Frontiers in Bioscience-Landmark 1998, 3, 631–636. [Google Scholar]
- Naor, D. interaction between hyaluronic acid and its receptors (CD44, RHAMM) regulates the activity of inflammation and cancer. 2016, Frontiers Media SA. p. 39. [CrossRef]
- Misra, S.; Heldin, P.; Hascall, V.C.; Karamanos, N.K.; Skandalis, S.S.; Markwald, R.R.; Ghatak, S. Hyaluronan–CD44 interactions as potential targets for cancer therapy. The FEBS journal 2011, 278, 1429–1443. [Google Scholar] [CrossRef]
- Toole, B.P. Hyaluronan-CD44 interactions in cancer: paradoxes and possibilities. Clinical Cancer Research 2009, 15, 7462–7468. [Google Scholar] [CrossRef]
- Jalkanen, S.; Jalkanen, M. Lymphocyte CD44 binds the COOH-terminal heparin-binding domain of fibronectin. The Journal of cell biology 1992, 116, 817–825. [Google Scholar] [CrossRef]
- Dzobo, K.; Sinkala, M. Cancer Stem Cell Marker CD44 Plays Multiple Key Roles in Human Cancers: Immune Suppression/Evasion, Drug Resistance, Epithelial–Mesenchymal Transition, and Metastasis. OMICS: A Journal of Integrative Biology 2021, 25, 313–332. [Google Scholar] [CrossRef]
- Thapa, R.; Wilson, G.D. The importance of CD44 as a stem cell biomarker and therapeutic target in cancer. Stem cells international 2016, 2016. [Google Scholar] [CrossRef]
- Hirata, K.; Suzuki, H.; Imaeda, H.; Matsuzaki, J.; Tsugawa, H.; Nagano, O.; Asakura, K.; Saya, H.; Hibi, T. CD44 variant 9 expression in primary early gastric cancer as a predictive marker for recurrence. British journal of cancer 2013, 109, 379–386. [Google Scholar] [CrossRef]
- Palapattu, G.S.; Wu, C.; Silvers, C.R.; Martin, H.B.; Williams, K.; Salamone, L.; Bushnell, T.; Huang, L.S.; Yang, Q.; Huang, J. Selective expression of CD44, a putative prostate cancer stem cell marker, in neuroendocrine tumor cells of human prostate cancer. The Prostate 2009, 69, 787–798. [Google Scholar] [CrossRef]
- Yae, T.; Tsuchihashi, K.; Ishimoto, T.; Motohara, T.; Yoshikawa, M.; Yoshida, G.J.; Wada, T.; Masuko, T.; Mogushi, K.; Tanaka, H. Alternative splicing of CD44 mRNA by ESRP1 enhances lung colonization of metastatic cancer cell. Nature communications 2012, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Leung, E.L.-H.; Fiscus, R.R.; Tung, J.W.; Tin, V.P.-C.; Cheng, L.C.; Sihoe, A.D.-L.; Fink, L.M.; Ma, Y.; Wong, M.P. Non-small cell lung cancer cells expressing CD44 are enriched for stem cell-like properties. PloS one 2010, 5, e14062. [Google Scholar] [CrossRef]
- Smith, L.R.; Cho, S.; Discher, D.E. Stem cell differentiation is regulated by extracellular matrix mechanics. Physiology 2018, 33, 16–25. [Google Scholar] [CrossRef]
- Hoffmann, E.J.; Ponik, S.M. Biomechanical contributions to macrophage activation in the tumor microenvironment. Frontiers in Oncology 2020, 10, 787. [Google Scholar] [CrossRef]
- Chaudhuri, O.; Cooper-White, J.; Janmey, P.A.; Mooney, D.J.; Shenoy, V.B. Effects of extracellular matrix viscoelasticity on cellular behaviour. Nature 2020, 584, 535–546. [Google Scholar] [CrossRef]
- Charrier, E.E.; Pogoda, K.; Wells, R.G.; Janmey, P.A. Control of cell morphology and differentiation by substrates with independently tunable elasticity and viscous dissipation. Nature communications 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Elosegui-Artola, A. The extracellular matrix viscoelasticity as a regulator of cell and tissue dynamics. Current Opinion in Cell Biology 2021, 72, 10–18. [Google Scholar] [CrossRef]
- Tschumperlin, D.J.; Lagares, D. Mechano-therapeutics: targeting mechanical signaling in fibrosis and tumor stroma. Pharmacology & therapeutics 2020, 212, 107575. [Google Scholar]
- Pratt, S.J.; Lee, R.M.; Martin, S.S. The mechanical microenvironment in breast cancer. Cancers 2020, 12, 1452. [Google Scholar] [CrossRef]
- Nazemi, M.; Rainero, E. Cross-talk between the tumor microenvironment, extracellular matrix, and cell metabolism in cancer. Frontiers in Oncology 2020, 10, 239. [Google Scholar] [CrossRef]
- Pizzo, A.M.; Kokini, K.; Vaughn, L.C.; Waisner, B.Z.; Voytik-Harbin, S.L. Extracellular matrix (ECM) microstructural composition regulates local cell-ECM biomechanics and fundamental fibroblast behavior: a multidimensional perspective. Journal of applied physiology 2005, 98, 1909–1921. [Google Scholar] [CrossRef]
- Wala, J.; Das, S. Mapping of biomechanical properties of cell lines on altered matrix stiffness using atomic force microscopy. Biomechanics and Modeling in Mechanobiology 2020, 19, 1523–1536. [Google Scholar] [CrossRef]
- Souza, S.T.; Agra, L.C.; Santos, C.E.; Barreto, E.; Hickmann, J.M.; Fonseca, E.J. Macrophage adhesion on fibronectin evokes an increase in the elastic property of the cell membrane and cytoskeleton: an atomic force microscopy study. European Biophysics Journal 2014, 43, 573–579. [Google Scholar] [CrossRef]
- Park, J.S.; Burckhardt, C.J.; Lazcano, R.; Solis, L.M.; Isogai, T.; Li, L.; Chen, C.S.; Gao, B.; Minna, J.D.; Bachoo, R. Mechanical regulation of glycolysis via cytoskeleton architecture. Nature 2020, 578, 621–626. [Google Scholar] [CrossRef]
- Vitillo, L.; Kimber, S.J. Integrin and FAK regulation of human pluripotent stem cells. Current stem cell reports 2017, 3, 358–365. [Google Scholar] [CrossRef]
- Dedhar, S. Cell–substrate interactions and signaling through ILK. Current opinion in cell biology 2000, 12, 250–256. [Google Scholar] [CrossRef]
- Schlie-Wolter, S.; Ngezahayo, A.; Chichkov, B.N. The selective role of ECM components on cell adhesion, morphology, proliferation and communication in vitro. Experimental cell research 2013, 319, 1553–1561. [Google Scholar] [CrossRef]
- Nallanthighal, S.; Rada, M.; Heiserman, J.P.; Cha, J.; Sage, J.; Zhou, B.; Yang, W.; Hu, Y.; Korgaonkar, C.; Hanos, C.T. Inhibition of collagen XI alpha 1-induced fatty acid oxidation triggers apoptotic cell death in cisplatin-resistant ovarian cancer. Cell death & disease 2020, 11, 1–12. [Google Scholar]
- Even-Ram, S.; Yamada, K.M. Cell migration in 3D matrix. Current opinion in cell biology 2005, 17, 524–532. [Google Scholar] [CrossRef]
- Welch, D.R.; Hurst, D.R. Defining the hallmarks of metastasis. Cancer research 2019, 79, 3011–3027. [Google Scholar] [CrossRef]
- Yilmaz, M.; Christofori, G.; Lehembre, F. Distinct mechanisms of tumor invasion and metastasis. Trends in molecular medicine 2007, 13, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Conklin, M.W.; Gangnon, R.E.; Sprague, B.L.; Van Gemert, L.; Hampton, J.M.; Eliceiri, K.W.; Bredfeldt, J.S.; Liu, Y.; Surachaicharn, N.; Newcomb, P.A. Collagen Alignment as a Predictor of Recurrence after Ductal Carcinoma In SituCollagen Alignment and Recurrence of DCIS. Cancer Epidemiology, Biomarkers & Prevention 2018, 27, 138–145. [Google Scholar]
- Conklin, M.W.; Eickhoff, J.C.; Riching, K.M.; Pehlke, C.A.; Eliceiri, K.W.; Provenzano, P.P.; Friedl, A.; Keely, P.J. Aligned collagen is a prognostic signature for survival in human breast carcinoma. The American journal of pathology 2011, 178, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Drifka, C.R.; Loeffler, A.G.; Mathewson, K.; Keikhosravi, A.; Eickhoff, J.C.; Liu, Y.; Weber, S.M.; Kao, W.J.; Eliceiri, K.W. Highly aligned stromal collagen is a negative prognostic factor following pancreatic ductal adenocarcinoma resection. Oncotarget 2016, 7, 76197. [Google Scholar] [CrossRef]
- Yang, N.; Friedl, A. Syndecan-1-induced ECM fiber alignment requires integrin αvβ3 and syndecan-1 ectodomain and heparan sulfate chains. PloS one 2016, 11, e0150132. [Google Scholar] [CrossRef]
- Erdogan, B.; Ao, M.; White, L.M.; Means, A.L.; Brewer, B.M.; Yang, L.; Washington, M.K.; Shi, C.; Franco, O.E.; Weaver, A.M. Cancer-associated fibroblasts promote directional cancer cell migration by aligning fibronectin. Journal of Cell Biology 2017, 216, 3799–3816. [Google Scholar] [CrossRef]
- Ao, M.; Brewer, B.M.; Yang, L.; Franco Coronel, O.E.; Hayward, S.W.; Webb, D.J.; Li, D. Stretching fibroblasts remodels fibronectin and alters cancer cell migration. Scientific reports 2015, 5, 1–9. [Google Scholar] [CrossRef]
- Feinberg, T.Y.; Zheng, H.; Liu, R.; Wicha, M.S.; Yu, S.M.; Weiss, S.J. Divergent matrix-remodeling strategies distinguish developmental from neoplastic mammary epithelial cell invasion programs. Developmental cell 2018, 47, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. Journal of enzyme inhibition and medicinal chemistry 2016, 31, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Werb, Z. The many faces of metalloproteases: cell growth, invasion, angiogenesis and metastasis. Trends in cell biology 2001, 11, S37–S43. [Google Scholar] [CrossRef]
- Elia, I.; Rossi, M.; Stegen, S.; Broekaert, D.; Doglioni, G.; van Gorsel, M.; Boon, R.; Escalona-Noguero, C.; Torrekens, S.; Verfaillie, C. Breast cancer cells rely on environmental pyruvate to shape the metastatic niche. Nature 2019, 568, 117–121. [Google Scholar] [CrossRef]
- Ge, J.; Cui, H.; Xie, N.; Banerjee, S.; Guo, S.; Dubey, S.; Barnes, S.; Liu, G. Glutaminolysis promotes collagen translation and stability via α-ketoglutarate–mediated mTOR activation and proline hydroxylation. American journal of respiratory cell and molecular biology 2018, 58, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Micalizzi, D.S.; Maheswaran, S.; Haber, D.A. A conduit to metastasis: circulating tumor cell biology. Genes & development 2017, 31, 1827–1840. [Google Scholar]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y. Pre-metastatic niches: organ-specific homes for metastases. Nature Reviews Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef]
- Gao, Y.; Bado, I.; Wang, H.; Zhang, W.; Rosen, J.M.; Zhang, X.H.-F. Metastasis organotropism: redefining the congenial soil. Developmental cell 2019, 49, 375–391. [Google Scholar] [CrossRef] [PubMed]
- Haemmerle, M.; Taylor, M.L.; Gutschner, T.; Pradeep, S.; Cho, M.S.; Sheng, J.; Lyons, Y.M.; Nagaraja, A.S.; Dood, R.L.; Wen, Y. Platelets reduce anoikis and promote metastasis by activating YAP1 signaling. Nature communications 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Yu, M.; Ting, D.T.; Stott, S.L.; Wittner, B.S.; Ozsolak, F.; Paul, S.; Ciciliano, J.C.; Smas, M.E.; Winokur, D.; Gilman, A.J. RNA sequencing of pancreatic circulating tumour cells implicates WNT signalling in metastasis. Nature 2012, 487, 510–513. [Google Scholar] [CrossRef]
- Pantel, K.; Alix-Panabières, C.; Riethdorf, S. Cancer micrometastases. Nature reviews Clinical oncology 2009, 6, 339–351. [Google Scholar] [CrossRef]
- Goddard, E.T.; Bozic, I.; Riddell, S.R.; Ghajar, C.M. Dormant tumour cells, their niches and the influence of immunity. Nature cell biology 2018, 20, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Yeh, A.C.; Ramaswamy, S. Mechanisms of cancer cell dormancy—another hallmark of cancer? Cancer research 2015, 75, 5014–5022. [Google Scholar] [CrossRef]
- Boire, A.; Coffelt, S.B.; Quezada, S.A.; Vander Heiden, M.G.; Weeraratna, A.T. Tumour dormancy and reawakening: Opportunities and challenges. Trends in Cancer 2019, 5, 762–765. [Google Scholar] [CrossRef]
- Phan, T.G.; Croucher, P.I. The dormant cancer cell life cycle. Nature Reviews Cancer 2020, 20, 398–411. [Google Scholar] [CrossRef]
- Gay, L.J.; Malanchi, I. The sleeping ugly: tumour microenvironment's act to make or break the spell of dormancy. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer 2017, 1868, 231–238. [Google Scholar] [CrossRef]
- Ghajar, C.M.; Peinado, H.; Mori, H.; Matei, I.R.; Evason, K.J.; Brazier, H.; Almeida, D.; Koller, A.; Hajjar, K.A.; Stainier, D.Y. The perivascular niche regulates breast tumour dormancy. Nature cell biology 2013, 15, 807–817. [Google Scholar] [CrossRef]
- Mancini, M.L.; Sonis, S.T. Mechanisms of cellular fibrosis associated with cancer regimen-related toxicities. Frontiers in pharmacology 2014, 5, 51. [Google Scholar] [CrossRef]
- Cox, T.R.; Erler, J.T. Fibrosis and cancer: partners in crime or opposing forces? Trends in cancer 2016, 2, 279–282. [Google Scholar] [CrossRef]
- Chandler, C.; Liu, T.; Buckanovich, R.; Coffman, L.G. The double edge sword of fibrosis in cancer. Translational Research 2019, 209, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R.; Erler, J.T. Molecular pathways: connecting fibrosis and solid tumor metastasis. Clinical Cancer Research 2014, 20, 3637–3643. [Google Scholar] [CrossRef] [PubMed]
- Principe, D.R.; Narbutis, M.; Kumar, S.; Park, A.; Viswakarma, N.; Dorman, M.J.; Kamath, S.D.; Grippo, P.J.; Fishel, M.L.; Hwang, R.F. Long-Term Gemcitabine Treatment Reshapes the Pancreatic Tumor Microenvironment and Sensitizes Murine Carcinoma to Combination ImmunotherapyGemcitabine Primes Pancreatic Cancer for Immunotherapy. Cancer research 2020, 80, 3101–3115. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.J.; Sharma, A.; Vuong, D.-V.; Erler, J.T.; Pruschy, M.; Broggini-Tenzer, A. Ionizing radiation induces tumor cell lysyl oxidase secretion. BMC cancer 2014, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.M.; Walsh, L.A. Candidate prognostic markers in breast cancer: focus on extracellular proteases and their inhibitors. Breast Cancer: Targets and Therapy 2014, 6, 81. [Google Scholar]
- Kanayama, H.o.; Yokota, K.y.; Kurokawa, Y.; Murakami, Y.; Nishitani, M.; Kagawa, S. Prognostic values of matrix metalloproteinase-2 and tissue inhibitor of metalloproteinase-2 expression in bladder cancer. Cancer: Interdisciplinary International Journal of the American Cancer Society 1998, 82, 1359–1366. [Google Scholar] [CrossRef]
- Bergamaschi, A.; Tagliabue, E.; Sørlie, T.; Naume, B.; Triulzi, T.; Orlandi, R.; Russnes, H.; Nesland, J.; Tammi, R.; Auvinen, P. Extracellular matrix signature identifies breast cancer subgroups with different clinical outcome. The Journal of pathology 2008, 214, 357–367. [Google Scholar] [CrossRef]
- Valdembri, D.; Serini, G. The roles of integrins in cancer. Faculty reviews 2021, 10. [Google Scholar] [CrossRef]
- Acerbi, I.; Cassereau, L.; Dean, I.; Shi, Q.; Au, A.; Park, C.; Chen, Y.Y.; Liphardt, J.; Hwang, E.S.; Weaver, V.M. Human breast cancer invasion and aggression correlates with ECM stiffening and immune cell infiltration. Integr Biol (Camb) 2015, 7, 1120–1134. [Google Scholar] [CrossRef]
- Bordeleau, F.; Mason, B.N.; Lollis, E.M.; Mazzola, M.; Zanotelli, M.R.; Somasegar, S.; Califano, J.P.; Montague, C.; LaValley, D.J.; Huynh, J.; et al. Matrix stiffening promotes a tumor vasculature phenotype. Proceedings of the National Academy of Sciences of the United States of America 2017, 114, 492–497. [Google Scholar] [CrossRef]
- Hui, L.; Zhang, J.; Ding, X.; Guo, X.; Jiang, X. Matrix stiffness regulates the proliferation, stemness and chemoresistance of laryngeal squamous cancer cells. Int J Oncol 2017. [Google Scholar] [CrossRef]
- McLane, J.S.; Ligon, L.A. Stiffened Extracellular Matrix and Signaling from Stromal Fibroblasts via Osteoprotegerin Regulate Tumor Cell Invasion in a 3-D Tumor in Situ Model. Cancer Microenviron 2016. [Google Scholar] [CrossRef]
- Tadeo, I.; Berbegall, A.P.; Navarro, S.; Castel, V.; Noguera, R. A stiff extracellular matrix is associated with malignancy in peripheral neuroblastic tumors. Pediatr Blood Cancer 2017. [Google Scholar] [CrossRef]
- Qin, X.; Lv, X.; Li, P.; Yang, R.; Xia, Q.; Chen, Y.; Peng, Y.; Li, L.; Li, S.; Li, T. Matrix stiffness modulates ILK-mediated YAP activation to control the drug resistance of breast cancer cells. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2020, 1866, 165625. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Ge, H.; Ghadban, T.; Reeh, M.; Güngör, C. The Extracellular Matrix: A Key Accomplice of Cancer Stem Cell Migration, Metastasis Formation, and Drug Resistance in PDAC. Cancers 2022, 14, 3998. [Google Scholar] [CrossRef]
- Grantab, R.H.; Tannock, I.F. Penetration of anticancer drugs through tumour tissue as a function of cellular packing density and interstitial fluid pressure and its modification by bortezomib. BMC cancer 2012, 12, 214. [Google Scholar] [CrossRef]
- Harisi, R.; Jeney, A. Extracellular matrix as target for antitumor therapy. Onco Targets Ther 2015, 8, 1387–1398. [Google Scholar] [CrossRef]
- Holle, A.W.; Young, J.L.; Spatz, J.P. In vitro cancer cell-ECM interactions inform in vivo cancer treatment. Advanced drug delivery reviews 2016, 97, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Miroshnikova, Y.A.; Mouw, J.K.; Barnes, J.M.; Pickup, M.W.; Lakins, J.N.; Kim, Y.; Lobo, K.; Persson, A.I.; Reis, G.F.; McKnight, T.R.; et al. Tissue mechanics promote IDH1-dependent HIF1alpha-tenascin C feedback to regulate glioblastoma aggression. Nat Cell Biol 2016, 18, 1336–1345. [Google Scholar] [CrossRef]
- Mittal, V.; El Rayes, T.; Narula, N.; McGraw, T.E.; Altorki, N.K.; Barcellos-Hoff, M.H. The Microenvironment of Lung Cancer and Therapeutic Implications. Adv Exp Med Biol 2016, 890, 75–110. [Google Scholar] [CrossRef] [PubMed]
- Mumenthaler, S.M.; Foo, J.; Choi, N.C.; Heise, N.; Leder, K.; Agus, D.B.; Pao, W.; Michor, F.; Mallick, P. The Impact of Microenvironmental Heterogeneity on the Evolution of Drug Resistance in Cancer Cells. Cancer Inform 2015, 14, 19–31. [Google Scholar] [CrossRef]
- Nieponice, A.; McGrath, K.; Qureshi, I.; Beckman, E.J.; Luketich, J.D.; Gilbert, T.W.; Badylak, S.F. An extracellular matrix scaffold for esophageal stricture prevention after circumferential EMR. Gastrointest Endosc 2009, 69, 289–296. [Google Scholar] [CrossRef]
- Barcus, C.E.; O'Leary, K.A.; Brockman, J.L.; Rugowski, D.E.; Liu, Y.; Garcia, N.; Yu, M.; Keely, P.J.; Eliceiri, K.W.; Schuler, L.A. Elevated collagen-I augments tumor progressive signals, intravasation and metastasis of prolactin-induced estrogen receptor alpha positive mammary tumor cells. Breast cancer research : BCR 2017, 19, 9. [Google Scholar] [CrossRef]
- Brechbuhl, H.M.; Finlay-Schultz, J.; Yamamoto, T.; Gillen, A.; Cittelly, D.M.; Tan, A.C.; Sams, S.B.; Pillai, M.; Elias, A.; Robinson, W.A.; et al. Fibroblast subtypes regulate responsiveness of luminal breast cancer to estrogen. Clinical cancer research : an official journal of the American Association for Cancer Research 2016. [Google Scholar] [CrossRef]
- Cui, L.; Tse, K.; Zahedi, P.; Harding, S.M.; Zafarana, G.; Jaffray, D.A.; Bristow, R.G.; Allen, C. Hypoxia and cellular localization influence the radiosensitizing effect of gold nanoparticles (AuNPs) in breast cancer cells. Radiat Res 2014, 182, 475–488. [Google Scholar] [CrossRef]
- Cun, X.; Ruan, S.; Chen, J.; Zhang, L.; Li, J.; He, Q.; Gao, H. A dual strategy to improve the penetration and treatment of breast cancer by combining shrinking nanoparticles with collagen depletion by losartan. Acta Biomater 2016, 31, 186–196. [Google Scholar] [CrossRef]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: a dynamic niche in cancer progression. J Cell Biol 2012, 196, 395–406. [Google Scholar] [CrossRef]
- Sung, S.Y.; Hsieh, C.L.; Wu, D.; Chung, L.W.; Johnstone, P.A. Tumor microenvironment promotes cancer progression, metastasis, and therapeutic resistance. Curr Probl Cancer 2007, 31, 36–100. [Google Scholar] [CrossRef]
- Whatcott, C.J.; Han, H.; Posner, R.G.; Hostetter, G.; Von Hoff, D.D. Targeting the tumor microenvironment in cancer: why hyaluronidase deserves a second look. Cancer Discov 2011, 1, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Mbeunkui, F.; Johann, D.J. Cancer and the tumor microenvironment: a review of an essential relationship. Cancer chemotherapy and pharmacology 2009, 63, 571–582. [Google Scholar] [CrossRef]
- Xiao, Q.; Ge, G. Lysyl oxidase, extracellular matrix remodeling and cancer metastasis. Cancer microenvironment 2012, 5, 261–273. [Google Scholar] [CrossRef]
- Gao, Y.; Xiao, Q.; Ma, H.; Li, L.; Liu, J.; Feng, Y.; Fang, Z.; Wu, J.; Han, X.; Zhang, J. LKB1 inhibits lung cancer progression through lysyl oxidase and extracellular matrix remodeling. Proceedings of the National Academy of Sciences 2010, 107, 18892–18897. [Google Scholar] [CrossRef] [PubMed]
- Vallet, S.D.; Ricard-Blum, S. Lysyl oxidases: from enzyme activity to extracellular matrix cross-links. Essays in biochemistry 2019, 63, 349–364. [Google Scholar]
- Seewaldt, V. ECM stiffness paves the way for tumor cells. Nature medicine 2014, 20, 332–333. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Chong, S.G.; Upagupta, C.; Yanagihara, T.; Saito, T.; Shimbori, C.; Bellaye, P.-S.; Nishioka, Y.; Kolb, M.R. Fibrotic extracellular matrix induces release of extracellular vesicles with pro-fibrotic miRNA from fibrocytes. Thorax 2021, 76, 895–906. [Google Scholar] [CrossRef]
- Netti, P.A.; Baxter, L.T.; Boucher, Y.; Skalak, R.; Jain, R.K. Time-dependent behavior of interstitial fluid pressure in solid tumors: implications for drug delivery. Cancer research 1995, 55, 5451–5458. [Google Scholar]
- Morin, P.J. Drug resistance and the microenvironment: nature and nurture. Drug Resist Updat 2003, 6, 169–172. [Google Scholar] [CrossRef]
- Gouarderes, S.; Mingotaud, A.-F.; Vicendo, P.; Gibot, L. Vascular and extracellular matrix remodeling by physical approaches to improve drug delivery at the tumor site. Expert opinion on drug delivery 2020, 17, 1703–1726. [Google Scholar] [CrossRef]
- Stylianopoulos, T.; Munn, L.L.; Jain, R.K. Reengineering the physical microenvironment of tumors to improve drug delivery and efficacy: from mathematical modeling to bench to bedside. Trends in cancer 2018, 4, 292–319. [Google Scholar] [CrossRef]
- Sercu, S.; Zhang, L.; Merregaert, e.J. The extracellular matrix protein 1: its molecular interaction and implication in tumor progression. Cancer investigation 2008, 26, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Chakravarthy, A.; Khan, L.; Bensler, N.P.; Bose, P.; De Carvalho, D.D. TGF-β-associated extracellular matrix genes link cancer-associated fibroblasts to immune evasion and immunotherapy failure. Nature communications 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Troup, S.; Njue, C.; Kliewer, E.V.; Parisien, M.; Roskelley, C.; Chakravarti, S.; Roughley, P.J.; Murphy, L.C.; Watson, P.H. Reduced expression of the small leucine-rich proteoglycans, lumican, and decorin is associated with poor outcome in node-negative invasive breast cancer. Clinical cancer research 2003, 9, 207–214. [Google Scholar] [PubMed]
- Biaoxue, R.; Xiguang, C.; Hua, L.; Hui, M.; Shuanying, Y.; Wei, Z.; Wenli, S.; Jie, D. Decreased expression of decorin and p57 (KIP2) correlates with poor survival and lymphatic metastasis in lung cancer patients. The International journal of biological markers 2011, 26, 9–21. [Google Scholar] [CrossRef]
- Matsumine, A.; Shintani, K.; Kusuzaki, K.; Matsubara, T.; Satonaka, H.; Wakabayashi, T.; Iino, T.; Uchida, A. Expression of decorin, a small leucine-rich proteoglycan, as a prognostic factor in soft tissue tumors. Journal of surgical oncology 2007, 96, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Järveläinen, H.; Sainio, A.; Koulu, M.; Wight, T.N.; Penttinen, R. Extracellular matrix molecules: potential targets in pharmacotherapy. Pharmacological reviews 2009, 61, 198–223. [Google Scholar] [CrossRef]
- Erkan, M.; Michalski, C.W.; Rieder, S.; Reiser–Erkan, C.; Abiatari, I.; Kolb, A.; Giese, N.A.; Esposito, I.; Friess, H.; Kleeff, J. The activated stroma index is a novel and independent prognostic marker in pancreatic ductal adenocarcinoma. Clinical gastroenterology and hepatology 2008, 6, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Niland, S.; Eble, J.A. Hold on or cut? Integrin-and MMP-mediated cell–matrix interactions in the tumor microenvironment. International Journal of Molecular Sciences 2020, 22, 238. [Google Scholar] [CrossRef]
- Henke, E.; Nandigama, R.; Ergün, S. Extracellular matrix in the tumor microenvironment and its impact on cancer therapy. Frontiers in molecular biosciences 2020, 6, 160. [Google Scholar] [CrossRef]
- Hanker, A.B.; Estrada, M.V.; Bianchini, G.; Moore, P.D.; Zhao, J.; Cheng, F.; Koch, J.P.; Gianni, L.; Tyson, D.R.; Sánchez, V. Extracellular Matrix/Integrin Signaling Promotes Resistance to Combined Inhibition of HER2 and PI3K in HER2+ Breast CancerECM Promotes Resistance to HER2/PI3K Inhibition. Cancer research 2017, 77, 3280–3292. [Google Scholar] [CrossRef] [PubMed]
- Hingorani, S.R.; Zheng, L.; Bullock, A.J.; Seery, T.E.; Harris, W.P.; Sigal, D.S.; Braiteh, F.; Ritch, P.S.; Zalupski, M.M.; Bahary, N. HALO 202: randomized phase II study of PEGPH20 plus nab-paclitaxel/gemcitabine versus nab-paclitaxel/gemcitabine in patients with untreated, metastatic pancreatic ductal adenocarcinoma. Journal of Clinical Oncology 2018, 36, 359–366. [Google Scholar] [CrossRef]
- Ramanathan, R.K.; McDonough, S.L.; Philip, P.A.; Hingorani, S.R.; Lacy, J.; Kortmansky, J.S.; Thumar, J.; Chiorean, E.G.; Shields, A.F.; Behl, D. Phase IB/II randomized study of FOLFIRINOX plus pegylated recombinant human hyaluronidase versus FOLFIRINOX alone in patients with metastatic pancreatic adenocarcinoma: SWOG S1313. Journal of Clinical Oncology 2019, 37, 1062. [Google Scholar] [CrossRef] [PubMed]
- Lokeshwar, B.L. MMP inhibition in prostate cancer. Annals of the New York Academy of Sciences 1999, 878, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Turpeenniemi-Hujanen, T. Gelatinases (MMP-2 and-9) and their natural inhibitors as prognostic indicators in solid cancers. Biochimie 2005, 87, 287–297. [Google Scholar] [CrossRef]
- Gomes, L.R.; Terra, L.F.; Wailemann, R.A.; Labriola, L.; Sogayar, M.C. TGF-β1 modulates the homeostasis between MMPs and MMP inhibitors through p38 MAPK and ERK1/2 in highly invasive breast cancer cells. BMC cancer 2012, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Matter, H.; Schudok, M. Recent advances in the design of matrix metalloprotease inhibitors. Current Opinion in Drug Discovery & Development 2004, 7, 513–535. [Google Scholar]
- Nuti, E.; Tuccinardi, T.; Rossello, A. Matrix metalloproteinase inhibitors: new challenges in the era of post broad-spectrum inhibitors. Current pharmaceutical design 2007, 13, 2087–2100. [Google Scholar] [CrossRef]
- Mannello, F.; Tonti, G.; Papa, S. Matrix metalloproteinase inhibitors as anticancer therapeutics. Current cancer drug targets 2005, 5, 285–298. [Google Scholar] [CrossRef]
- Radisky, E.S.; Raeeszadeh-Sarmazdeh, M.; Radisky, D.C. Therapeutic potential of matrix metalloproteinase inhibition in breast cancer. Journal of cellular biochemistry 2017, 118, 3531–3548. [Google Scholar] [CrossRef] [PubMed]
- Devel, L.; Czarny, B.; Beau, F.; Georgiadis, D.; Stura, E.; Dive, V. Third generation of matrix metalloprotease inhibitors: Gain in selectivity by targeting the depth of the S1′ cavity. Biochimie 2010, 92, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Broekgaarden, M.; Weijer, R.; van Gulik, T.M.; Hamblin, M.R.; Heger, M. Tumor cell survival pathways activated by photodynamic therapy: a molecular basis for pharmacological inhibition strategies. Cancer and Metastasis Reviews 2015, 34, 643–690. [Google Scholar] [CrossRef] [PubMed]
- Grant, S. Cotargeting survival signaling pathways in cancer. The Journal of clinical investigation 2008, 118, 3003–3006. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, J.-Y.; Wei, W.-Z.; Wu, G.S. Activation of the Akt survival pathway contributes to TRAIL resistance in cancer cells. PloS one 2010, 5, e10226. [Google Scholar] [CrossRef]
- Roy-Luzarraga, M.; Hodivala-Dilke, K. Molecular Pathways: Endothelial Cell FAK—A Target for Cancer TreatmentEndothelial Cell FAK in Cancer Treatment. Clinical Cancer Research 2016, 22, 3718–3724. [Google Scholar] [CrossRef]
- McCormick, F. Small-molecule inhibitors of cell signaling. Current Opinion in Biotechnology 2000, 11, 593–597. [Google Scholar] [CrossRef]
- Morris, J.C.; Tan, A.R.; Olencki, T.E.; Shapiro, G.I.; Dezube, B.J.; Reiss, M.; Hsu, F.J.; Berzofsky, J.A.; Lawrence, D.P. Phase I study of GC1008 (fresolimumab): a human anti-transforming growth factor-beta (TGFβ) monoclonal antibody in patients with advanced malignant melanoma or renal cell carcinoma. PloS one 2014, 9, e90353. [Google Scholar] [CrossRef]
- Lin, R.; Yi, S.; Gong, L.; Liu, W.; Wang, P.; Liu, N.; Zhao, L.; Wang, P. Inhibition of TGF-β signaling with halofuginone can enhance the antitumor effect of irradiation in Lewis lung cancer. OncoTargets and therapy 2015, 8, 3549. [Google Scholar] [CrossRef]
- Juárez, P.; Mohammad, K.S.; Yin, J.J.; Fournier, P.G.; McKenna, R.C.; Davis, H.W.; Peng, X.H.; Niewolna, M.; Javelaud, D.; Chirgwin, J.M. Halofuginone Inhibits the Establishment and Progression of Melanoma Bone MetastasesHalofuginone Decreases Melanoma Bone Metastases. Cancer research 2012, 72, 6247–6256. [Google Scholar] [CrossRef]
- Miyazono, K. Transforming growth factor-β signaling in epithelial-mesenchymal transition and progression of cancer. Proceedings of the Japan Academy, Series B 2009, 85, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Shaty, M.H.; Arif, I.S.; Al-Ezzi, M.I.; Hanna, D.B. Metformin attenuate fibrosis in both acute and chronic doxorubicin cardiotoxicity in rabbits. Journal of Pharmaceutical Sciences and Research 2018, 10, 1559–1565. [Google Scholar]
- Lampi, M.C.; Reinhart-King, C.A. Targeting extracellular matrix stiffness to attenuate disease: From molecular mechanisms to clinical trials. Science translational medicine 2018, 10, eaao0475. [Google Scholar] [CrossRef] [PubMed]
- Sporn, M.B.; Roberts, A.B.; Wakefield, L.M.; Assoian, R.K. Transforming growth factor-β: biological function and chemical structure. Science 1986, 233, 532–534. [Google Scholar] [CrossRef] [PubMed]
- Krane, S.M. Collagenases and collagen degradation. Journal of Investigative Dermatology 1982, 79, 83–86. [Google Scholar] [CrossRef]
- McKee, T.D.; Grandi, P.; Mok, W.; Alexandrakis, G.; Insin, N.; Zimmer, J.P.; Bawendi, M.G.; Boucher, Y.; Breakefield, X.O.; Jain, R.K. Degradation of fibrillar collagen in a human melanoma xenograft improves the efficacy of an oncolytic herpes simplex virus vector. Cancer research 2006, 66, 2509–2513. [Google Scholar] [CrossRef] [PubMed]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nature Reviews Immunology 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Khokha, R.; Murthy, A.; Weiss, A. Metalloproteinases and their natural inhibitors in inflammation and immunity. Nature Reviews Immunology 2013, 13, 649–665. [Google Scholar] [CrossRef]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nature reviews Molecular cell biology 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Coussens, L.M.; Fingleton, B.; Matrisian, L.M. Matrix metalloproteinase inhibitors and cancer—trials and tribulations. Science 2002, 295, 2387–2392. [Google Scholar] [CrossRef]
- Castellani, P.; Viale, G.; Dorcaratto, A.; Nicolo, G.; Kaczmarek, J.; Querze, G.; Zardi, L. The fibronectin isoform containing the ED-B oncofetal domain: a marker of angiogenesis. International journal of cancer 1994, 59, 612–618. [Google Scholar] [CrossRef]
- Glukhova, M.A.; Frid, M.G.; Shekhonin, B.V.; Balabanov, Y.V.; Koteliansky, V.E. Expression of fibronectin variants in vascular and visceral smooth muscle cells in development. Developmental biology 1990, 141, 193–202. [Google Scholar] [CrossRef]
- Han, Z.; Lu, Z.-R. Targeting fibronectin for cancer imaging and therapy. Journal of Materials Chemistry B 2017, 5, 639–654. [Google Scholar] [CrossRef] [PubMed]
- Carnemolla, B.; Leprini, A.; Allemanni, G.; Saginati, M.; Zardi, L. The inclusion of the type III repeat ED-B in the fibronectin molecule generates conformational modifications that unmask a cryptic sequence. Journal of Biological Chemistry 1992, 267, 24689–24692. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.-M.; Lan, Y.; Lauder, S.; Zhang, J.; Brunkhorst, B.; Qin, G.; Verma, R.; Courtenay-Luck, N.; Gillies, S.D. huBC1-IL12, an immunocytokine which targets EDB-containing oncofetal fibronectin in tumors and tumor vasculature, shows potent anti-tumor activity in human tumor models. Cancer Immunology, Immunotherapy 2007, 56, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Saw, P.E.; Kim, S.; Lee, I.-h.; Park, J.; Yu, M.; Lee, J.; Kim, J.-I.; Jon, S. Aptide-conjugated liposome targeting tumor-associated fibronectin for glioma therapy. Journal of Materials Chemistry B 2013, 1, 4723–4726. [Google Scholar] [CrossRef]
- Lokman, N.A.; Price, Z.K.; Hawkins, E.K.; Macpherson, A.M.; Oehler, M.K.; Ricciardelli, C. 4-Methylumbelliferone inhibits cancer stem cell activation and overcomes chemoresistance in ovarian cancer. Cancers 2019, 11, 1187. [Google Scholar] [CrossRef] [PubMed]
- Kohli, A.G.; Kivimäe, S.; Tiffany, M.R.; Szoka, F.C. Improving the distribution of Doxil® in the tumor matrix by depletion of tumor hyaluronan. Journal of controlled release 2014, 191, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Doherty, G.J.; Tempero, M.; Corrie, P.G. HALO-109–301: a Phase III trial of PEGPH20 (with gemcitabine and nab-paclitaxel) in hyaluronic acid-high stage IV pancreatic cancer. Future oncology 2018, 14, 13–22. [Google Scholar] [CrossRef]
- Schnittert, J.; Bansal, R.; Storm, G.; Prakash, J. Integrins in wound healing, fibrosis and tumor stroma: High potential targets for therapeutics and drug delivery. Advanced drug delivery reviews 2018, 129, 37–53. [Google Scholar] [CrossRef]
- Gutheil, J.C.; Campbell, T.N.; Pierce, P.R.; Watkins, J.D.; Huse, W.D.; Bodkin, D.J.; Cheresh, D.A. Targeted antiangiogenic therapy for cancer using Vitaxin: a humanized monoclonal antibody to the integrin ανβ3. Clinical Cancer Research 2000, 6, 3056–3061. [Google Scholar] [PubMed]
- Raguse, J.-D.; Gath, H.J.; Bier, J.; Riess, H.; Oettle, H. Cilengitide (EMD 121974) arrests the growth of a heavily pretreated highly vascularised head and neck tumour. Oral oncology 2004, 40, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Mas-Moruno, C.; Rechenmacher, F.; Kessler, H. Cilengitide: the first anti-angiogenic small molecule drug candidate. Design, synthesis and clinical evaluation. Anti-Cancer Agents in Medicinal Chemistry (Formerly Current Medicinal Chemistry-Anti-Cancer Agents) 2010, 10, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Tijink, B.M.; Buter, J.; De Bree, R.; Giaccone, G.; Lang, M.S.; Staab, A.; Leemans, C.R.; Van Dongen, G.A. A phase I dose escalation study with anti-CD44v6 bivatuzumab mertansine in patients with incurable squamous cell carcinoma of the head and neck or esophagus. Clinical Cancer Research 2006, 12, 6064–6072. [Google Scholar] [CrossRef]
- Riechelmann, H.; Sauter, A.; Golze, W.; Hanft, G.; Schroen, C.; Hoermann, K.; Erhardt, T.; Gronau, S. Phase I trial with the CD44v6-targeting immunoconjugate bivatuzumab mertansine in head and neck squamous cell carcinoma. Oral oncology 2008, 44, 823–829. [Google Scholar] [CrossRef]
Figure 1.
Collagen structure.
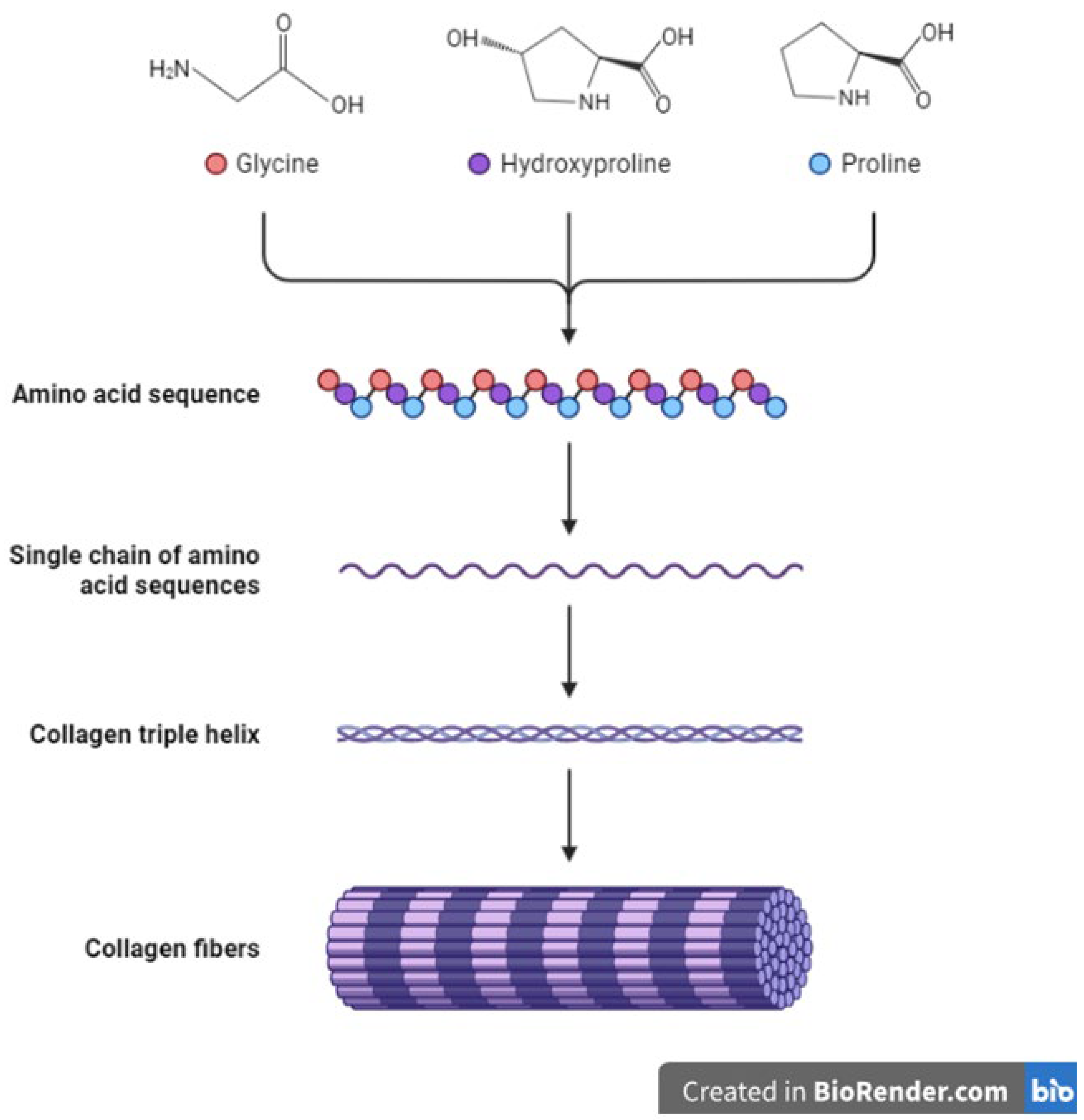
Figure 2.
Extracellular matrix remodeling in tumors.

Figure 3.
The role of the ECM in tumor metastasis.
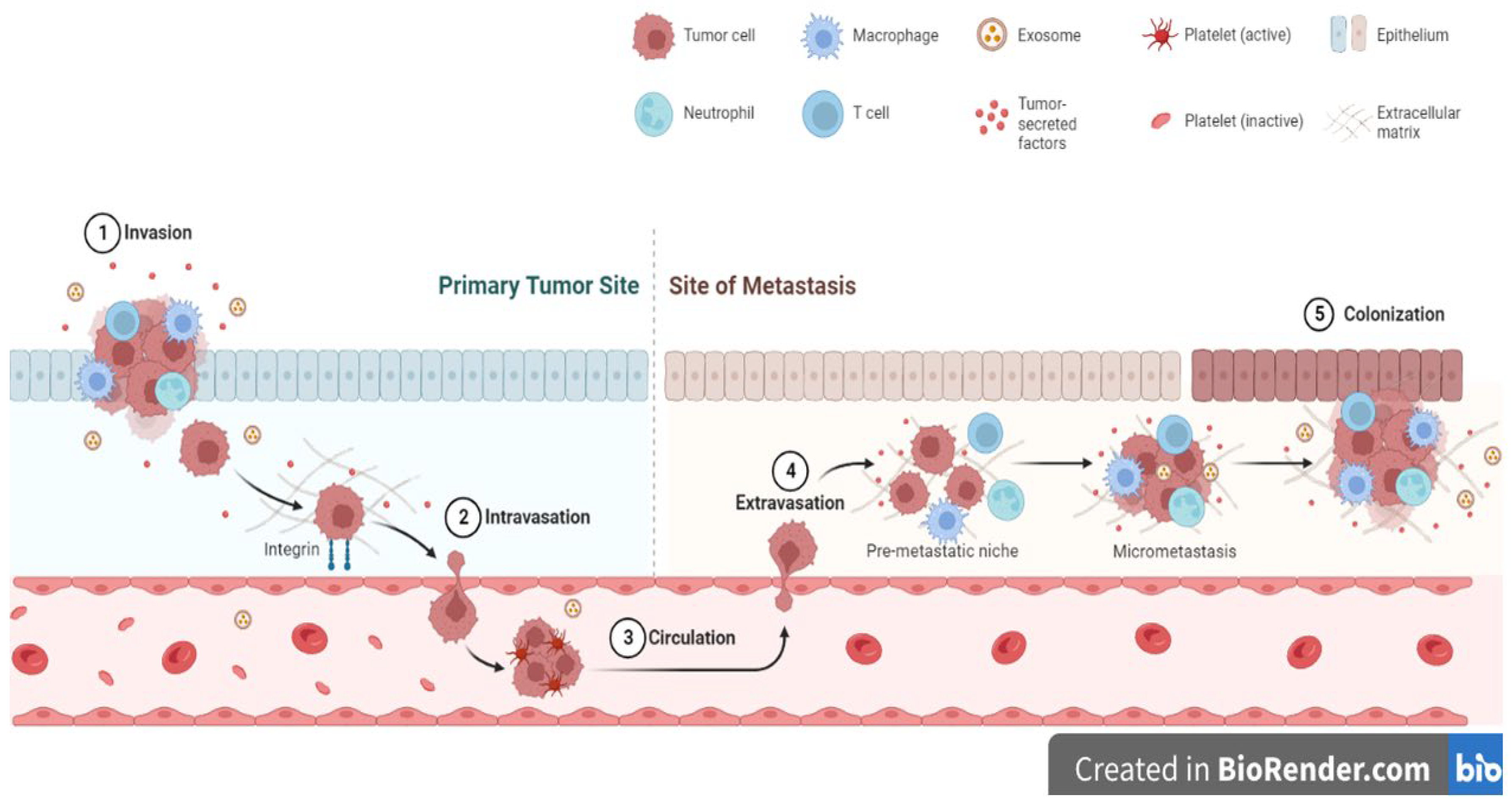
Figure 4.
Tumor ECM reduces therapeutic efficiency in solid tumors.
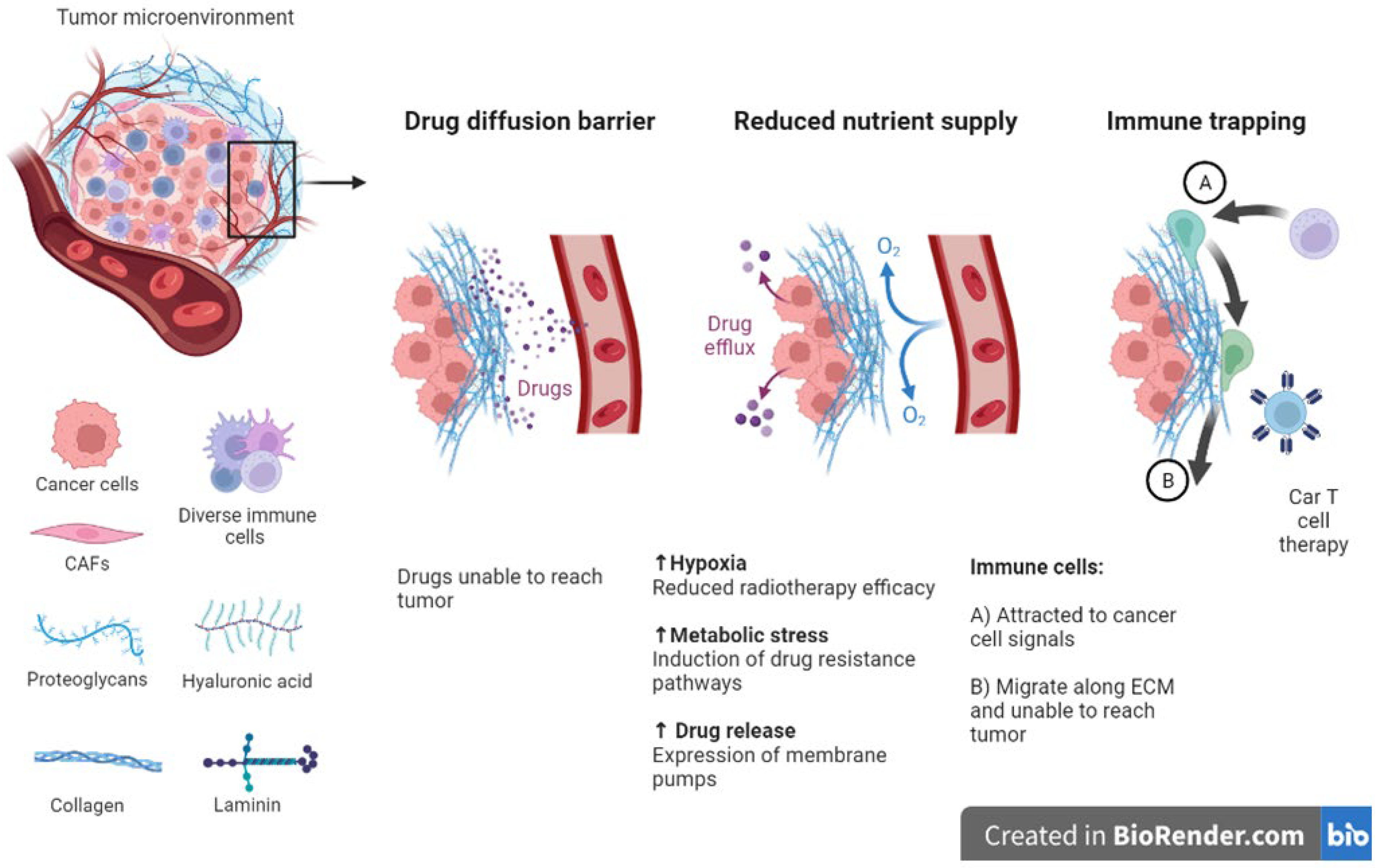
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



