
Submitted:
25 January 2023
Posted:
31 January 2023
You are already at the latest version
Abstract
The objective of this study was to evaluate the potential benefits of feeding spray-dried porcine plasma (SDPP) to pigs infected with African swine fever virus (ASFV). Two groups of twelve weaned pigs each were fed with CONVENTIONAL or 8% SDPP enriched diets. Two pigs (trojans)/group) were injected intramuscularly with the pandemic ASFV (Georgia 2007/01) and comingled with the rest of the pigs (1:5 trojan:naïve ratio) to simulate a natural route of transmission. Trojans developed ASF and died within the first week after inoculation but contact pigs did not develop ASF, viremia or seroconversion. Therefore, three more trojans per group were introduced to optimize the ASFV transmission (1:2 trojan:naïve ratio). Blood, nasal and rectal swabs were weekly harvested and at end of the study, ASFV-target organs collected. After the second exposure, rectal temperature of conventionally fed contact pigs increased >40.5˚C while fever was delayed in the SDPP contact pigs. Additionally, PCR Ct values in blood, secretions and tissue samples were significantly lower (P<0.05) for CONVENTIONAL compared to SDPP contact pigs. Under these study conditions, contact exposed pigs fed SDPP had delayed ASFV transmission and reduced virus load, likely by enhanced specific T-cell priming after the first ASFV-exposure.
Keywords:
African swine fever
; ASFV
; spray-dried porcine plasma
; challenge
; nutritional intervention
1. Introduction
African swine fever virus (ASFV) is an enveloped DNA virus of the Asfarviridae family that infects domestic pigs and wild boar of all ages causing African swine fever (ASF), a disease that must be reported to the World Organization for Animal Health (WOAH, formerly OIE).
Lithuania made in January 2014 the first notification of acute ASF cases in wild boar in the European Union (EU), and Poland followed in February 2014. In June and September 2014, Latvia and Estonia also reported ASFV. To date, Ukraine, Romania, Estonia, Hungary, Bulgaria, Czech Republic, Slovakia, Belgium, Greece, Germany and recently Italy, have reported cases of ASF in wild boar and some countries in domestic pigs. All epidemiological data indicate that EU has undergone repeated introduction of ASFV from the Eastern neighboring infected countries. The EU Reference Laboratory confirmed, through genetic studies that there was a 100% homology with the genotype II virus that entered Georgia in 2007, imported from Western Africa [1]. Furthermore, from August 2018, ASFV has spread in most pig producing countries in Asia, including China, Vietnam, South and North Korea, Philippines, Laos, Myanmar, Mongolia, Cambodia, Timor-Lester, Indonesia, Malaysia, India, Papua-New Guinea, and Thailand. In addition, genotype II ASFV has recently spread to the Caribbean and new cases in Haiti and Dominican Republic have been reported in wild and domestic pigs [2].
However, the pattern spread and severity of ASF outbreak in domestic pigs in different regions varies significantly, ranging from up to 70% reduction of total swine herd in China, Vietnam, Philippines, and other Asia countries to limited spot outbreaks in EU countries and South Korea. Biosecurity could be the main factor influencing the spread and outcome of the disease, as countries with less severe outbreaks had better biosecurity programs. The health and immune status of the herd may also play a key role. Pietschmann et al. [3] reported that immune compromised runt and gastrointestinal challenged unhealthy pigs are more susceptible for ASF infection. Radulovic et al. [4] reported that domestic pigs with a very high hygienic and health status experienced a milder and shorter form of the illness followed by complete recovery when inoculated with an attenuated field ASFV strain. Therefore, improving health and hygienic status of the herd may be a key for ASF prevention and control.
Spray-dried plasma (SDP) is a dry functional feed ingredient that contains a diverse mixture of many functional components such as immunoglobulins, albumin, growth factors, biologically active peptides, transferrin, and other molecules that have biological activity independent of their nutritional value. SDP either from porcine (SDPP) or bovine (SDBP) origin are extensively used in pig starter diets and consistently provide improvements in growth performance, feed efficiency, and animal survival, especially under stressful conditions like pathogen challenge [5]. The beneficial effects of SDP are related to its mode of action that supports an efficient immune system response [6]. In addition, feeding SDPP has been shown to promote a more favorable gut microflora [7,8,9]. Thus, SDPP helps improve the integrity of the intestinal barrier by modulating the functional and structural properties of the intestinal mucosa in pigs and other animal models [10]. In addition, dietary SDPP can favorably affect intestinal morphology and immune cell subsets of gut tissues and blood in weaned pigs [11]. Furthermore, SDPP exerts a systemic effect, as demonstrated by the reduction of lung inflammation and the presence of monocytes and cells involved in the inflammatory events under an acute pulmonary inflammation model with LPS in mice [12,13]. Moreover, in an epidemiologic study conducted in Manitoba, Canadian researchers found that farms with less pulmonary lesions associated with the presence of porcine reproductive and respiratory syndrome virus (PRRSV), porcine circovirus 2 (PCV-2) and Mycoplasma hyopneumoniae were well correlated with supplementation of SDPP in their feed [14].
Good intestinal health translates into a more efficient immune system and overall better growth performance [15,16]. Following this principle, an improved intestinal barrier function through gut microbiota has been demonstrated to influence ASFV susceptibility. Fecal transplantation from warthogs (African pigs resilient to ASF) to domestic pigs improved mucosal immunity and protected pigs against experimental challenge with a live attenuated ASFV strain [17]. Although mechanisms involved in ASFV protection are not fully understood, ASFV specific antibodies [18] and cytotoxic CD8 T-cell (CTL) responses [19], together with an appropriate innate immune response [20], have been demonstrated to play an important role. The activation of Th1 like responses, including specific CD8 T-cells is a strategy currently pursued to obtain efficient vaccines against ASFV [21]. The fact that dietary SDPP can protect mucosal integrity and also promote optimal immune responses, including Th1-like responses and CTL induction [22,23], prompted us to hypothesize that SDPP could be used as a potential nutritional intervention against ASFV transmission and progression of the infection.
Therefore, the objective of this study was to assess if supplementing SDPP in diets of healthy pigs reduced ASF virus load of infected animal and the rate of in pen ASFV transmission after an experimental ASFV pig-to-pig contact challenge.
2. Materials and Methods
Ethical statement
For this study 24 Landrace x Large White male pigs (5 weeks of age) were obtained from a commercial farm with high sanitary status.
The clinical state of the animals and the end-point criteria was evaluated by scoring the ASF-compatible clinical signs following a previously reported guide [24]. A score from 0 to 5 according to severity was applied as follows: 0: no clinical signs, 1: mild pyrexia (39.6–40.0 ºC), 2: mild pyrexia (39.6–40.0 ºC) and mild clinical signs (skin, digestive), 3: moderate pyrexia (40.0–40.5 ºC) and mild-moderate clinical signs (distal ear spots, mild limp, lying down, but remaining alert), 4: moderate-high pyrexia (40.5-41ºC) and moderate clinical signs (remains dormant, only stands up when touched, hesitant step, subcutaneous bleeding <10%, diarrhea, mild tremors), and 5: pyrexia higher than 41ºC and moderate-severe clinical signs (generalized subcutaneous bleeding, ataxia, spasticity, clouding, prostration, bloody diarrhea).
Study design
The 24 pigs were divided randomly into two groups of 12 pigs each and were placed in two separate rooms at the IRTA-CReSA biosecurity level 3 animal facility. Each group of 12 pigs represented an experimental treatment group: CONVENTIONAL: ASFV challenged pigs fed a conventional diet with soy protein concentrate, and SDPP: ASFV challenged pigs fed a diet with SDPP. The SDPP used in this study was randomly selected from a manufactured lot of AP920 produced by APC Europe S.L.U. (Granollers, Spain). The CONVENTIONAL group diet was formulated with soy protein concentrate (Soycomil ® P) at 100.9 g/kg and the SDPP group was fed a diet formulated with SDPP at 80 g/kg, which entirely replaced Soycomil. Both diets were iso-energy (3340 kcal ME/kg) and iso-nutrient balanced (Lysine SID 12.50 g/kg feed). The experimental diets (Table 1) were prepared in mash form and offered ad libitum during the adaptation period and the 31-day study.
After a brief adaptation period to the diets and facility, on day 4 of the experiment two randomly selected animals (for use as trojan pigs) per room were separated from the other pigs (10 contact pigs per room) by placing them in two subdivisions to avoid direct contact with the other pigs. Immediately thereafter, the two trojans per room were intramuscularly (IM) injected with a lethal dose of 103 genome equivalent copies (GEC) of the Georgia 2007/1-ASFV strain. Two days after virus inoculation, the trojan and contact pigs in each room were re-grouped to begin the contact exposure. The ratio of trojan to naïve contact pigs was 1:5, mimicking a potential field scenario of slow ASFV transmission. As expected, trojans developed ASF signs by day 4 after inoculation and were sacrificed for humane reasons by day 6 after inoculation when showing acute ASF clinical signs.
However, none of contact pigs in either group developed fever, ASF specific signs, or viremia after 19 days of observation. In view of these results, a modification of the protocol was obtained from the ethical committee to increase the infection pressure by providing a second exposure of ASFV infected trojan pigs to contact pigs. Accordingly, three pigs from each group were chosen randomly and inoculated IM with 103 GEC (acting as trojan seeders) while remaining inside the same pen with the 6 remaining contact pigs to have a 1:2 (trojan to contact pig) ratio. To maintain this optimal contact challenge ratio [25], one pig per each contact group was excluded from the study before the second ASFV-exposure. The study ended at day 12 post-second exposure (pse), following the originally approved protocol (Figure 1).
During the whole study animals were daily observed for ASF-compatible clinical signs (including rectal temperature) and samples of blood (EDTA 15 mL tubes) were taken from all animals at 0 days post-contact (dpc) immediately before IM injection to the two trojans, 4 dpc, 7 dpc, 14 dpc, 21 dpc (2d pse), and 28 dpc (9d pse). In addition, nasal and rectal swabs were collected at 4 dpc, 7 dpc, 11dpc 14 dpc, 18 dpc, 21 dpc, 25 dpc (6d pse) and 28 dpc (9d pse). Necropsies were done on day 31 dpc (12 d pse), following the schedule in Figure 1. All samples were stored at -75ºC until processed. Blood, nasal and rectal swabs were analyzed by real time PCR (qRT-PCR) to detect viremia. At necropsy, lesions were registered, and samples of spleen, tonsil, gastro-hepatic node, submaxillary node, and retropharyngeal node were taken and analyzed for virus detection by qRT-PCR.
ELISPOT analysis [26] was conducted in frozen cells where viability was >90% after thawing from blood obtained with EDTA at 11, 25 and 31 dpc.
Laboratory analyses
Samples of ASFV inoculum were analyzed by the qRT-PCR described by [27]. Blood samples were taken in tubes with EDTA at specified days. DNA extraction was done using the Indimag Pathogen Kit (Indical Biosciences, Leipzig, Germany). Viremia was determined by qRT-PCR analysis using the primers described by Fernández-Pinero et al. [28] and the probe ASF-VP72P1 described in the current OIE ASF chapter (Terrestrial Manual OIE, Section 3.9, Chapter 3.9.1 African Swine Fever Virus pages 1-18) with the following modification in the thermoprofile made by the Spanish National Reference Laboratory for ASF: 10 min at 95ºC, 5 cycles 1 min at 95ºC + 30 seg at 60ºC, 40 cycles 10 min at 95ºC + 30 seg at 60ºC with fluorescence acquisition in the FAM channel at the end of each PCR cycle. According to these amplification settings results were considered as positive when Ct values were ≤ 30, inconclusive Ct values between 30 to 35, and Ct >35 were considered negative.
Seroconversion was determined by ELISA (Ingezim PPA COMPAC, INGENASA; Madrid, Spain) with O.D. ≤ 0.061 as negative. Nasal and rectal swabs were analyzed for the presence of ASFV genome using the same procedures previously mentioned. Once the nasal and rectal swabs arrived at the laboratory, the end of the swab was cut and placed in a tube with 1 mL of PBS. The tubes were stored at -75ºC until DNA extraction and analysis by qRT-PCR.
At necropsy, samples of tonsil, spleen, and retropharyngeal, submaxillary, and gastro-hepatic lymph nodes were taken for subsequent qRT-PCR analysis. For each sample, 0.1 g of tissue was diluted 1:10 and homogenized using sterile PBS and TyssueLyser II (Qiagen, Hilden, Germany). DNA extraction and qRT-PCR were done as detailed above.
Correlation Ct vs Log10 HAD50/mL of the different analyzed tissues was carried out using ASFV Georgia 2007 viral stock at 6.99 Log10HAD50/mL as a standard. qRT-PCR were carried out as detailed above (Suppl. Table S1).
Peripheral blood mononuclear cells (PBMCs) were purified from EDTA blood samples by density-gradient centrifugation with Histopaque 1077 (Sigma-Aldrich, Missouri, USA). Cells were frozen at -80ºC but the viability was tested to be ≥ 90% after thawing before conducting the analysis. Commercial antibody Porcine IFN-γ P2G10 and biotin P2C11, from BD Biosciences (Pharmingen, California, USA) at 5 µg/mL was used to quantify by ELISPOT assay the number of IFNγ secreting cells. PBMC were stimulated against ASF Georgia/01 strain virus at a MOI between 0.1 and 0.01 to measure specific T-cell responses by ELISPOT in real time with fresh cells [28]. Plates were revealed using detection antibody from BD Pharmingen (BD Biosciences, California, USA), Strep-HRP from Life Technologies (California, USA) and TMB substrate for ELISPOT assay (MABTECH, Stockholm, Sweden). ELISPOTs were read under a magnifying glass.
Statistical analysis
Data were analyzed as a completely randomized design using the GLM procedures of SAS (SAS Inst., Inc., Cary, NC). An analysis of variance was conducted to detect differences among treatments. The independent variable was treatment. Dependent variables were body temperatures, blood, nasal and rectal swab and tissue Ct values. The LSMEANS procedure was used to calculate the mean values by treatment. If treatment effects were detected, least squares means were separated using the PDIFF option in SAS. Pig was considered the experimental unit. Means are considered significantly different if P < 0.05 while trends are reported as P = 0.05 to 0.10.
3. Results
3.1. First exposure
After the first exposure to ASFV-inoculated trojans (1:5 ratio of trojans:contact pigs), none of the contact pigs developed temperatures above 40.4ºC, had ASF compatible signs or viremia after 18 days of monitoring (Suppl. Table S2). One CONVENTIONAL contact pig on 4 dpc and 5 CONVENTIONAL and 3 SDPP contact pigs on 7 dpc displayed an ASFV qRT-PCR positive result in the nasal swab, but thereafter to 18 dpc all nasal swab results were negative. All blood and rectal swab samples for contact pigs were negative. Despite the nasal swab results for contact pigs, this did not result in a disseminated infection (Suppl. Table S3), even though trojan pigs in both groups had developed high fever and were euthanized 6 dpc per protocol requirements.
3.2. Second exposure
Second exposure: Conversely, after the second contact exposure (1:2 ratio of trojans:contact pigs), contact pigs from the CONVENTIONAL group became infected by ASFV, following the expected kinetics [29]. Thus, all six contact pigs from the CONVENTIONAL group developed acute ASF-compatible clinical signs, showing high fever (rectal temperature >41ºC) by day 12 pse with one contact pig (#15) having > 41ºC starting at day 9 pse (Figure 2A). Interestingly, all contact pigs from the SDPP group had a delay in the onset of ASF clinical signs with only two of the six pigs (# 4 and 7) having rectal temperatures >41ºC the last day of the study (Figure 2B; Suppl. Table S2).
Average body temperature was higher (P<0.05) for CONVENTIONAL contact pigs on day 6, and days 9 to 12 pse compared to SDPP contact pigs (Figure 3).
All three trojan pigs in the CONVENTIONAL group died between 4 to 7 days post-inoculation. However, two of the three trojan pigs in the SDPP group survived until the last day of the study with RT below 41ºC, even though these pigs displayed severe signs of disease. Therefore, the exposure time of contact pigs to trojans displaying fever was longer in the SDPP group than in the CONVENTIONAL group due to the longer survival rate of the trojan pigs in the SDPP group.
Similar to the rectal temperature results, viremia, ASFV positive rectal and nasal swabs were not detected in the SDPP group until 12d pse, while in the CONVENTIONAL group viremia and positive nasal and rectal samples were detected by 7d pse (Suppl. Table S3 and Suppl. Table S4). At necropsy, trojans from SDPP group showed lower load of virus genome in their tissues compared with the trojans in CONVENTIONAL group.
Despite the longer exposure time of contact pigs in the SDPP group to the trojans (2/3 survived), contact pigs in the SDPP group had delayed onset of clinical sign, lower viremia load, ASFV shedding, and ASFV genome copies or Log10HAD50/ml in every single tissue analyzed on day 12 pse, compared to contact pigs in the CONVENTIONAL group, which indicates reduced rate of in-pen transmission (Table 2; Figure 4; Suppl. Table S5).
Viremia was found on day 12 pse in all animals except for one pig from the SDPP group (#5) and the average Ct values in the SDPP group was numerically higher (indicating less ASFV genome copies) than in the CONVENTIONAL group. On day 12 pse, nasal swabs from all animals were qPCR positive for ASFV but again, the average Ct values in the SDPP group was numerically higher than the CONVENTIONAL group. The rectal swab average Ct values were low and almost identical for both groups with one and two pigs showing no detectable ASFV genome in the SDPP and CONVENTIONAL groups, respectively. Finally, submaxillary lymph nodes and tonsil tissue for all contact animals in both groups were qPCR positive, except for animal #3 of the SDPP group that was negative (Ct value >35). In gastro-hepatic and submaxillary lymph nodes there was a tendency (P<0.10) to have less average amount of virus genome copies (higher Ct values) for animals in the SDPP to the CONVENTIONAL group. In addition, pigs in the SDPP group had lower (P<0.05) ASFV load in tonsil and retropharyngeal lymph nodes.
To understand the mechanisms explaining the differential outcome of both ASF clinical sign evolution and virus transmission, the kinetics of both the specific antibody and T-cell responses were followed. None of the contact pigs from either group developed detectable anti-ASFV antibodies after the first ASFV contact challenge, which is consistent with the absence of detectable viremia. Even after the second exposure to ASFV inoculated trojans, contact pigs did not have time to develop significant amounts of anti-ASFV antibodies and by day 12 pse remained below the threshold limit of the technique (Suppl. Table S6). In clear contrast with the lack of antibody responses, ASFV-specific IFN-γ secreting T-cells were detectable by ELISPOT as soon as 11 days after first ASFV encounter (11 dpc); thus, 8 of 9 contact pigs in the SDPP group developed low, albeit detectable ASFV specific T-cells, while 2 of 7 contact pigs in the CONVENTIONAL group had detectable T-cells. On day 25 of the experiment (7 days pse) most pigs in both groups developed detectable specific T-cell response against ASFV (Figure 5. Suppl. Table S7).
4. Discussion
Despite the dramatic consequences that ASF represent for the swine industry and for global trading, ASFV cannot be considered as a highly transmissible virus, at least compared with other pig viruses of obligatory declaration to the WOAH [30], such as classical swine fever virus (CSFV) and foot and mouth disease virus (FMDV). In agreement with this observation, naïve pigs fed daily for 14 consecutive days with feed blended with liquid porcine plasma inoculated with 104.3 or 105.0 TCID50/g ASFV remained ASF-free [31]. The fact that ASFV recovered from the feed remained infectious when administered intra-muscularly or intra-gastric with a probe, confirmed that infection with ASFV through feed consumption is not as efficient as for other pathogens. Other studies feeding commercial inoculated ASFV contaminated feed not containing plasma resulted in infection of the naïve pigs [32,33,34]). These results suggest that unprocessed liquid porcine plasma or SDPP may contain inherent properties that diminish the infectious capacity of ASFV and may contribute to the survival of 2 of the 3 SDPP trojans until end of the present study.
This limited capability of ASFV transmission might explain the lack of evident viral transmission observed between trojan and contact pigs at a ratio of 1:5 (infected vs non-infected animal ratio). However, during the first exposure the contact pigs were probably exposed to the virus at low levels for a short time which was adequate to trigger the induction of a mild ASFV-specific T-cell response but was not sufficient to induce detectable ASFV-specific antibodies or viremia at the times tested. This finding was definitively confirmed after showing that some contact pigs in both groups had low amounts of ASFV genome detectable by qPCR in the nasal swab by 4 and 7 dpc.
The presence of low levels of ASFV in the nostrils of contact pigs without causing viremia or ASF clinical signs might be further explained by the induction of efficient innate immune responses capable of controlling ASFV infection [20] at the site of entry. The fact that positive nasal swab samples were detected in a lower number of contact pigs in SDPP versus the CONVENTIONAL group, seem to strengthen our original hypothesis because previous results in pigs and other animal species confirm the improvement of innate responses by SDPP feeding [11,22]. These results fit with the fact that at later time points (11 dpc), most contact pigs fed SDPP showed ASFV-specific IFN-γ secreting T-cells, compared with only two animals fed the CONVENTIONAL diet. The positive effect of SDPP on Th1-like memory responses was also confirmed by others [22,23], including the induction of cytotoxic CD8+ T cells [22,23,35]. Independently of this, as mentioned earlier, Th1-like responses and CD8+ T cells play a key role in ASFV protection [19,21] and dietary SDPP in other challenge studies has demonstrated increases in the percentage of CD8+ T cells [22,23,35].
The presence of ASFV specific CD8+ T-cells in contact pigs fed SDPP after the first exposure with the trojan pigs, may partially explain the delay of the ASF onset observed in this group of pigs after the second ASFV encounter [19,21,25], but also the unique feature observed in 2 of the trojan pigs used for the second challenge that survived 12 days after IM ASFV challenge. Despite that average viremia and nasal shedding genome values were similar between both groups of trojan by day six post challenge (Suppl. Table S3), these two pigs (# 8, #9) in the SDPP group continued secreting ASFV to the environment until the end of the experiment (day 12 pse).
SDPP is a functional ingredient that demonstrated to improve the integrity of the intestinal barrier, enrich the gut microbiota, and modulate the immune system of pigs and other animal species [6,7,8,9,10,15,16,36,37,38]. Improving integrity of intestinal barrier function may reduce the potential infectivity of ASFV and microbiota seems to have an important role for controlling ASFV infection [17,39]. In addition, SDPP supplemented in feed was shown to modulate the immune system and balance the ratio between activated and regulatory T cells [6,10], which are essential to protect against most pathogens, including ASFV [20]. As above mentioned, albeit little is known about the mechanisms involved in ASFV protection, innate and both memory B and T cells responses play a role, most probably in a regulated way to avoid excessive inflammation. Diets with SDPP may contribute to this regulation by providing an optimal cytokine environment [6,10,11,37]. Recently, a study conducted by Radulovic et al., [4] demonstrated that pigs can regulate immune activation more efficiently after vaccination with attenuated ASFV strains; these animals showed milder production of proinflammatory cytokines after virus inoculation, similar to the results in previous studies when animals are fed diets supplemented with SDPP [6,10,37].
In Asia-Pacific regions new less virulent strains and attenuated strains of ASFV compared with the original Georgia strain have appeared, probably due to genetic evolution the virus and the use of non-authorized vaccine prototypes [40,41]. Such scenarios complicate the epidemiological ASFV picture and control of ASF. Based on the results from this study using a virulent ASFV strain we speculate that healthier pigs fed diets with SDPP may improve resilience to ASFV strains because diets with SDPP improves the health status of pigs, especially under stressful conditions [5]. Our recent results showing SDPP improved the outcome of ASFV experimental vaccination [42] (back-to-back submitted manuscript) support the hypothesis that nutritional interventions may be useful in the future fight to control ASF in endemic regions.
5. Conclusions
In conclusion, feeding SDPP reduced virus load in the different target tissues and delayed in pen transmission in pigs exposed to ASFV inoculated trojans. Therefore, supplementation of SDPP in feed may be a strategic nutritional intervention to improve the degree of protection against ASFV.
Supplementary Materials
The following are available online at Preprints.org, Table S1: Correlation Ct vs Log10 HAD; Table S2: Body temperature throughout the duration of the study; Table S3: PCR results in blood, nasal and rectal swab; Table S4: Summary results of PCR in blood, nasal and rectal swab; Table S5: PCR results of different tissues; Table S6: ELISPOT results at different days during the study; Table S7: Antibodies against ASFV in serum at different days during the study.
Author Contributions
Conceptualization, E.B., J. Polo, J. Pujols, F.R. and J.S.; methodology, E.B., J. Pujols, F.R., R.R. and J.S.; software, J. Polo, J. Pujols and J.C.; validation, J. Pujols, F.R., J.S., R.R. and J. Polo; formal analysis, J.C, F.R., J. Pujols and J. Polo; investigation, J. Pujols, F.R. and J.S.; resources, J. Pujols and F.R.; data curation, J. Pujols, E.B., F.R. and J. Polo; writing—original draft preparation, J. Polo, E.B., J. Pujols, F.R. and J. S.; writing—review and editing, F.R., J. Polo, E.B. and J.S..; visualization, E.B. and J. Polo.; supervision, J. Pujols and J. Polo.; project administration, J. Pujols; funding acquisition, J. Polo. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
The study was approved by the committee of ethics and welfare “Comitè d’Experimentació Animal de la Generalitat de Catalunya” with the protocol approval number CEA-OH/11025/1.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data from this study is provided in the manuscript and supplementary tables.
Acknowledgments
We thank the technicians and PhD students of IRTA-CReSA helping in the experimental setup.
Conflicts of Interest
The authors have read the journal's policy and the authors of this manuscript have the following competing interests: EB, and J. Polo are employed by APC Europe, S.L.U. Granollers, Spain. JC and J. Polo are employed by APC LLC, Ankney, US. Both companies manufacture and sells spray-dried animal plasma; however, the companies had no role in the design of the study; in the collection, analysis, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results. J. Pujols, FR, RR and JS declare no conflict of interest. This does not alter the authors’ adherence to all journal policies on sharing data and materials.
References
- Śmietanka, K.; Woźniakowski, G.; Kozak, E.; Niemczuk, K.; Fraçzyk, M.; Bocian, Ł.; Kowalczyk, A.; Pejsak, Z. African Swine Fever Epidemic, Poland, 2014-2015. Emerg Infect Dis. 2016, 22, 1201–1207. [Google Scholar] [CrossRef]
- World organization for animal health (WOAH). African swine fever situation report. Available online: https://www.woah.org/app/uploads/2022/10/asf-report22.pdf (accessed on 10 January 2023).
- Pietschmann, J.; Guinat, C.; Beer, M.; Pronin, V.; Tauscher, K.; Petrov, A.; Keil, G.; Blome, S. Course and transmission characteristics of oral low-dose infection of domestic pigs and European wild boar with a Caucasian African swine fever virus isolate. Arch Virol. 2015, 160, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Radulovic, E.; Mehinagic, K.; Wuthrich, T.; Hilty, M.; Posthaus, H.; Summerfield, A.; Ruggli, N.; Benarafa, C. The baseline immunological and hygienic status of pigs impact disease severity of African swine fever. PLoS Pathog. 2022, 18, e1010522. [Google Scholar] [CrossRef] [PubMed]
- Torrallardona, D. Spray dried animal plasma as an alternative to antibiotics in weanling pigs. Asian-Australas J Anim Sci. 2010; 23, 131–148. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Polo, J.; Torrallardona, D. Spray dried plasma as an alternative to antibiotics in piglet feeds, mode of action and biosafety. Porcine Health Manag. 2016, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.; Anderson, C.L.; Bundy, J.W.; Fernando, S.C.; Miller, P.S.; Burkey, T.E. Effects of spray-dried porcine plasma on fecal microbiota in nursery pigs. J Anim Sci. 2018, 96, 1017–1031. [Google Scholar] [CrossRef]
- Moretó, M.; Miró, L.; Amat, C.; Polo, J.; Manichanh, C.; Pérez-Bosque, A. Dietary supplementation with spray-dried porcine plasma has prebiotic effects on gut microbiota in mice. Sci Rep. 2020, 10, 2926. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Paniagua, S.T.; Balebona, M. del C.; Firmino, J.P.; Rodríguez, C.; Polo, J.; Moriñigo, M.A.; Gisbert, E. The effect of spray-dried porcine plasma on gilthead seabream (Sparus aurata) intestinal microbiota. Aquac Nutr. 2020; 26, 801–811. [Google Scholar] [CrossRef]
- Moretó, M.; Pérez-Bosque, A. Dietary plasma proteins, the intestinal immune system, and the barrier functions of the intestinal mucosa. J Anim Sci. 2009, 87, 92–100. [Google Scholar] [CrossRef]
- Nofrarías, M.; Manzanilla, E.G.; Pujols, J.; Gibert, X.; Majó, N.; Segalés, J.; Gasa, J. Effects of spray-dried porcine plasma and plant extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J Anim Sci. 2006, 84, 2735–2742. [Google Scholar] [CrossRef]
- Maijó, M.; Miró, L.; Polo, J.; Campbell, J.; Russell, L.; Crenshaw, J.; Weaver, E.; Moretó, M.; Pérez-Bosque, A. Dietary Plasma Proteins Modulate the Adaptive Immune Response in Mice with Acute Lung Inflammation. J Nutr. 2102, 142, 264–270. [Google Scholar] [CrossRef]
- Maijó, M.; Miró, L.; Polo, J.; Campbell, J.; Russell, L.; Crenshaw, J.; Weaver, E.; Moretó, M.; Pérez-Bosque, A. Dietary plasma proteins attenuate the innate immunity response in a mouse model of acute lung injury. Br J Nutr. 2012, 107, 867–875. [Google Scholar] [CrossRef]
- Dewey, C.E.; Johnston, W.T.; Gould, L.; Whiting, T.L. Postweaning mortality in Manitoba swine. Can J Vet Res. 2006, 70, 161–167. [Google Scholar] [PubMed]
- Pérez-Bosque, A.; Amat, C.; Polo, J.; Campbell, J.M.; Crenshaw, J.; Russell, L.; Moretó, M. Spray-Dried Animal Plasma Prevents the Effects of Staphylococcus aureus Enterotoxin B on Intestinal Barrier Function in Weaned Rats. J Nutr. 2006, 136, 2838–2843. [Google Scholar] [CrossRef] [PubMed]
- Peace, R.M.; Campbell, J.; Polo, J.; Crenshaw, J.; Russell, L.; Moeser, A. Spray-Dried Porcine Plasma Influences Intestinal Barrier Function, Inflammation, and Diarrhea in Weaned Pigs. J Nutr. 2011, 141, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rodríguez, F.; Navas, M.J.; Costa-Hurtado, M.; Almagro, V.; Bosch-Camós, L.; López, E.; Cuadrado, R.; Accensi, F.; Pina-Pedrero, S.; Martínez, J.; Correa-Fiz, F. Fecal microbiota transplantation from warthog to pig confirms the influence of the gut microbiota on African swine fever susceptibility. Sci Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Onisk, D. v.; Borca, M. v.; Kutish, S.; Kramer, E.; Irusta, P.; Rock, D.L. Passively Transferred African Swine Fever Virus Antibodies Protect Swine against Lethal Infection. Virology 1994, 198, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Oura, C.A.L.; Denyer, M.S.; Takamatsu, H.; Parkhouse, R.M.E. In vivo depletion of CD8+ T lymphocytes abrogates protective immunity to African swine fever virus. J Gen Virol. 2005, 86, 2445–2450. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, H.H.; Denyer, M.S.; Lacasta, A.; Stirling, C.M.A.; Argilaguet, J.M.; Netherton, C.L.; Oura, C.A.L.; Martins, C.; Rodríguez, F. Cellular immunity in ASFV responses. Virus Res. 2013, 173, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Camós, L.; López, E.; Rodriguez, F. African swine fever vaccines: a promising work 1still in progress. Porcine Health Manag. 2020, 6. [Google Scholar] [CrossRef]
- Markowska-Daniel, I.; Szczotka, A.; Bednarek, D.; Pejsak, Z. Preliminary study of the influence of plasma proteins on immunological and production parameters in pigs. Pol J Vet Sci. 2003, 6, 275–277. [Google Scholar]
- Díaz, I.; Lorca, C.; Galindo, I.; Campbell, J.; Barranco, L.; Kuzemtseva, L.; Rodríguez-Gómez, I.-M.; Crenshaw, J.; Russell, L.; Polo, J.; Pujols, J. Potential positive effect of commercial spray-dried porcine plasma on pigs challenged with PRRS virus. Proceedings of 21st international pig veterinary society (IPVA), Vancouver, Canada, 2010. Pp. 560.
- Galindo-Cardiel, I.; Ballester, M.; Solanes, D.; Nofrarías, M.; López-Soria, S.; Argilaguet, J.M.; Lacasta, A.; Accensi, F.; Rodríguez, F.; Segalés, J. Standardization of pathological investigations in the framework of experimental ASFV infections. Virus Res. 2013, 173, 180–190. [Google Scholar] [CrossRef]
- Bosch-Camós,L.; Alonso, U.; Esteve-Codina, A.; Chang, C-Y.; Martín-Mur, B.; Accensi, F.; Muñoz, M.; Navas, M.J.; Dabad, M.; Vidal, E.; Pina-Pedrero, S.; Pleguezuelos, P.; Caratù, G.; Salas, M.L.; Liu, L.; Bataklieva, S.; Gavrilov, B.; Rodríguez, F.; Argilaguet, J. Cross-protection against African swine fever virus upon intranasal vaccination is associated with an adaptive-innate immune crosstalk. Plos Pathog. 2022; 18, e1010931. [CrossRef]
- Goatley, L.C.; Reis, A.L.; Portugal, R.; Goldswain, H.; Shimmon, G.L.; Hargreaves, Z.; Ho, C.S.; Montoya, M.; Sánchez-Cordón, P.J.; Taylor, G.; Dixon, L.K.; Netherton, C.L. A Pool of Eight Virally Vectored African Swine Fever Antigens Protect Pigs Against Fatal Disease. Vaccines (Basel) 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.A.; Darby, A.C.; Da Silva, M.; Upton, C.; Radford, A.D.; Dixon, L.K. Genomic analysis of highly virulent Georgia 2007/1 isolate of African swine fever virus. Emerg Infect Dis. 2011, 17, 599–605. [Google Scholar] [CrossRef]
- Fernández-Pinero, J.; Gallardo, C.; Elizalde, M.; Robles, A.; Go, C. Molecular Diagnosis of African Swine Fever by a New Real-Time PCR Using Universal Probe Library. Transbound Emerg Dis. 2013, 60, 48–58. [Google Scholar] [CrossRef]
- Díaz, I.; Darwich, L.; Pappaterra, G.; Pujols, J.; Mateu, E. Immune responses of pigs after experimental infection with a European strain of Porcine reproductive and respiratory syndrome virus. J Gen Virol. 2005, 86, 1943–1951. [Google Scholar] [CrossRef]
- Schulz, K.; Conraths, F.J.; Blome, S.; Staubach, C.; Sauter-Louis, C. African Swine Fever: Fast and Furious or Slow and Steady? Viruses 2019, 11. [Google Scholar] [CrossRef]
- Blázquez, E.; Pujols, J.; Segalés, J.; Rodríguez, F.; Crenshaw, J.; Rodríguez, C.; Ródenas, J.; Polo, J. Commercial feed containing porcine plasma spiked with African swine fever virus is not infective in pigs when administered for 14 consecutive days. Plos One 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Niederwerder, M.C.; Stoian, A.M.M.; Rowland, R.R.R.; Dritz, S.S.; Petrovan, V.; Constance, L.A.; Gebhardt, J.T.; Olcha, M.; Jones, C.K.; Woodworth, J.C.; Fang, Y.; Liang, J.; Hefley, T.J. Infectious dose of African swine fever virus when consumed naturally in liquid or feed. Emerg Infect Dis. 2019, 25, 891–897. [Google Scholar] [CrossRef]
- De Vos, M.; Tran, H.T.T.; Lannoo, K.; Bruggeman, G.; Dang, H.V. Medium chain fatty acids show potential to mitigate ASFv in feed. Proceedings of 26th international pig veterinary society congress (IPVS). Rio de Janeiro, Brazil. June 21st -24th, 2022. Pp. 704.
- Tran, H.T.T.; Truong, A.D.; Ly, D.V.; Hoang, T.V.; Chu, N.T.; Nguyen, H.T.; Dang, A.T.K.; De Vos, M.; Lannoo, K.; Bruggeman, G.; Dang, H.V. The potential anti-African swine fever virus effects of medium chain fatty acids on in vitro feed model: An evaluation study using epidemic ASFV strain circulating in Vietnam. Open Vet J. 2021, 11, 346–355. [Google Scholar] [CrossRef]
- Pujols, J.; Lorca-Oró, C.; Díaz, I.; Russell, L.E.; Campbell, J.M.; Crenshaw, J.D.; Polo, J.; Mateu, E.; Segalés, J. Commercial spray-dried porcine plasma does not transmit porcine circovirus type 2 in weaned pigs challenged with porcine reproductive and respiratory syndrome virus. Vet J. 2011; 190, e16–e20. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Pelegrí, C.; Vicario, M.; Castell, M.; Russell, L.; Campbell, J.M.; Quigley, J.D.; Polo, J.; Amat, C.; Moretó, M. Dietary plasma protein affects the immune response of weaned rats challenged with S. aureus Superantigen B. J Nutr. 2004, 134, 2667–2672. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Miró, L.; Amat, C.; Polo, J.; Moretó, M. The Anti-Inflammatory Effect of Spray-Dried Plasma Is Mediated by a Reduction in Mucosal Lymphocyte Activation and Infiltration in a Mouse Model of Intestinal Inflammation. Nutrients 2016, 8, 657. [Google Scholar] [CrossRef]
- Che, L.; Hu, L.; Zhou, Q.; Peng, X.; Liu, Y.; Luo, Y.; Fang, Z.; Lin, Y.; Xu, S.; Feng, B.; Li, J.; Tang, J.; Wu, D. Microbial insight into dietary protein source affects intestinal function of pigs with intrauterine growth retardation. Eur J Nutr. 2020, 59, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Lacasta, A.; Ballester, M.; Monteagudo, P.L.; Rodríguez, J.M.; Salas, M.L.; Accensi, F.; Pina-Pedrero, S.; Bensaid, A.; Argilaguet, J.; López-Soria, S.; Hutet, E.; le Potier, M.F.; Rodríguez, F. Expression library immunization can confer protection against lethal challenge with African swine fever virus. J Virol. 2014, 88, 13322–13332. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Huang, L.; Zhang, X.; Zhang, J.; Shen, D.; Zhang, Z.; Wang, Z.; Huo, H.; Wang, W.; Huangfu, H.; Wang, W.; Li, F.; Liu, R.; Sun, J.; Tian, Z.; Xia, W.; Guan, Y.; He, X.; Zhu, Y.; Zhao, D.; Bu, Z. Genotype I African swine fever viruses emerged in domestic pigs in China and caused chronic infection. Emerg Microbes Infect. 2021, 10, 2183–2193. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Bosch, J.; Martínez-Avilés, M.; Sánchez-Vizcaíno, J.M. The Evolution of African Swine Fever in China: A Global Threat? Front Vet Sci. 2022, 0, 248. [Google Scholar] [CrossRef]
- Pujols, J.; Blázquez, E.; Segalés, J.; Rodríguez, F.; Chang, C-Y; Argilaguet, J.; Bosch-Camós, L.; Rosell, R.; Pailler, L.; Gavrilov, B.; Campbell, J.; Polo, J. Feeding spray-dried porcine plasma to pigs improves the protection afforded by the African swine fever virus (ASFV) BA71ΔCD2 vaccine prototype against experimental challenge with the pandemic ASFV. Study 2. Vaccnes (under review).
Figure 1.
Schematic representation of the study design.
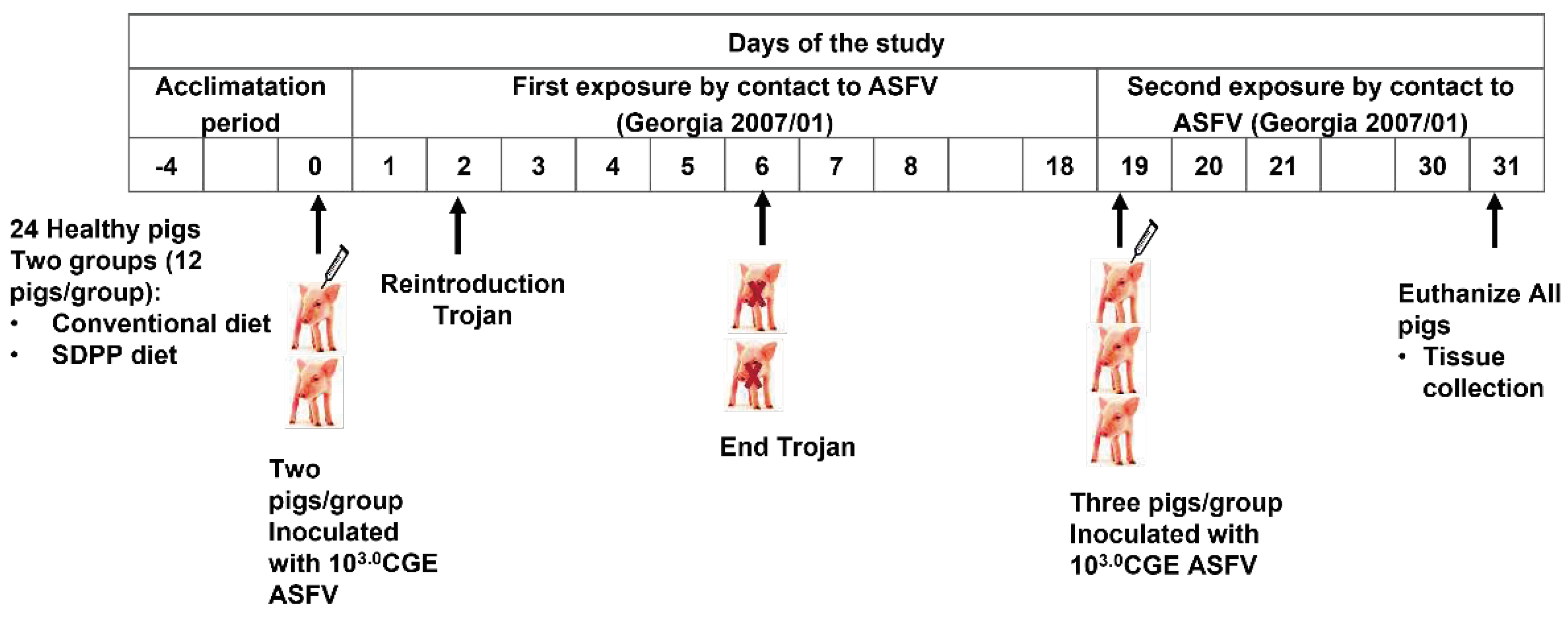
Figure 2.
Rectal temperature over time after second ASFV exposure. A: animals fed with CONVENTIONAL diet and B: animals fed with spray dried porcine plasma (SDPP) diet. In red color the trojans animals in each group.
Figure 2.
Rectal temperature over time after second ASFV exposure. A: animals fed with CONVENTIONAL diet and B: animals fed with spray dried porcine plasma (SDPP) diet. In red color the trojans animals in each group.
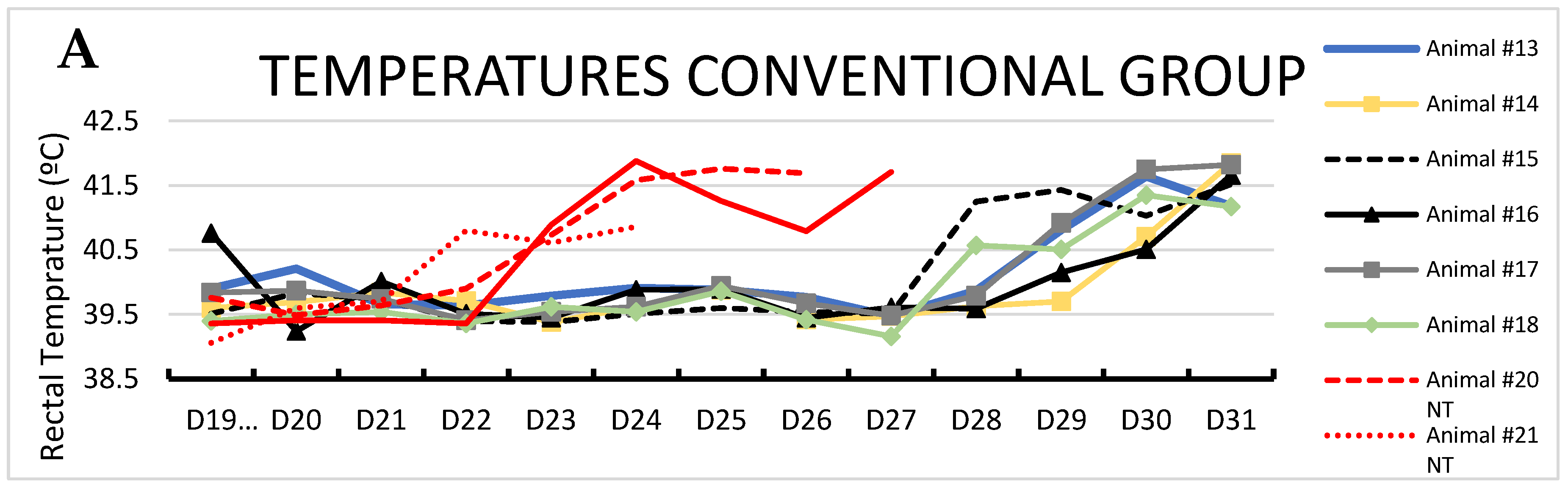
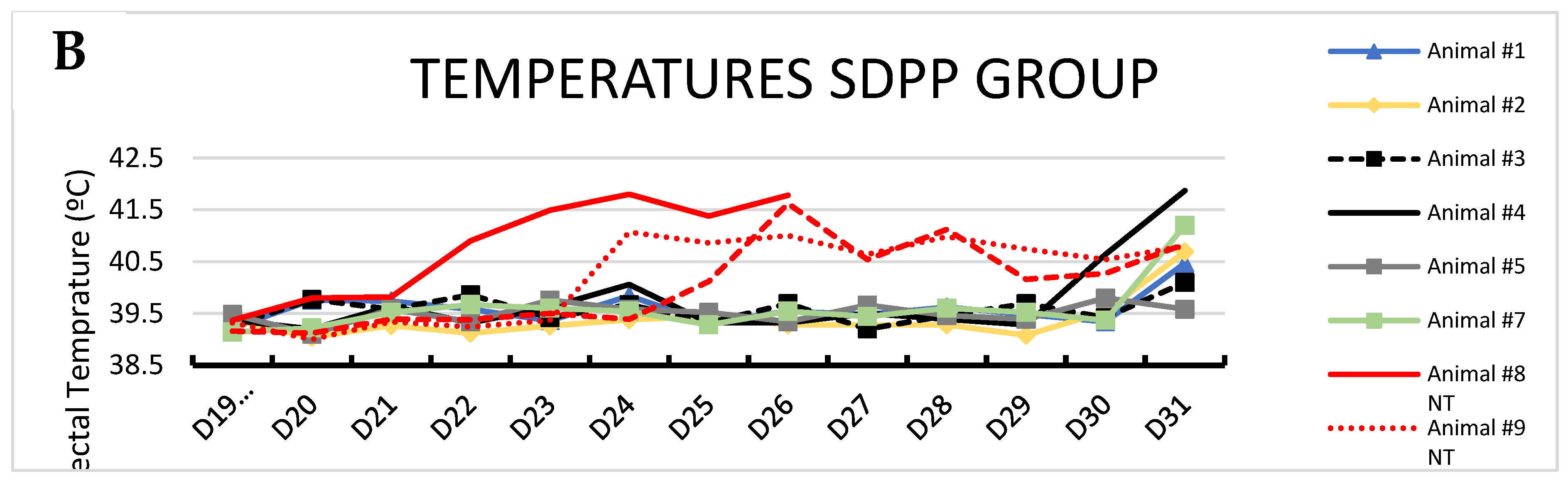
Figure 3.
Average rectal temperature over time between treatments after second ASFV exposure. # = P < 0.1; * = P < 0.05; ** = P < 0.01 SDPP = spray-dried porcine plasma.
Figure 3.
Average rectal temperature over time between treatments after second ASFV exposure. # = P < 0.1; * = P < 0.05; ** = P < 0.01 SDPP = spray-dried porcine plasma.
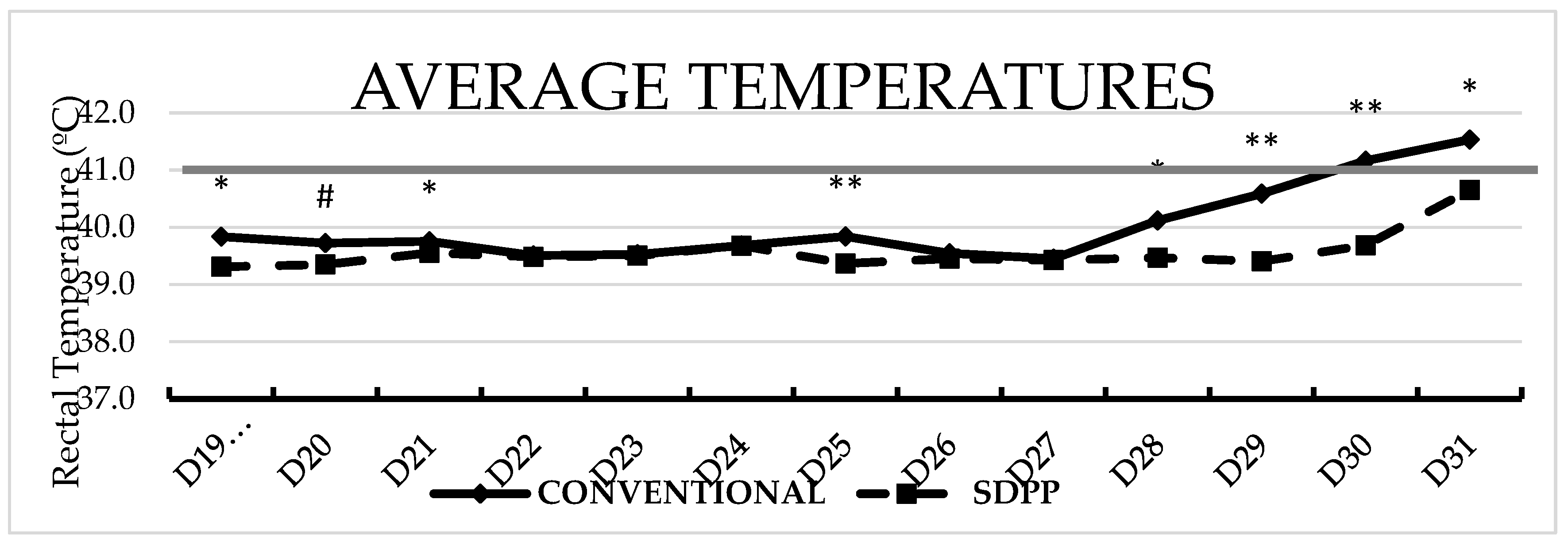
Figure 4.
Average viral load expressed as Log10 HAD/mL in different tissues analyzed at d31 of the study. # = P < 0.1; * = P < 0.05; ** = P < 0.01 SDPP = spray-dried porcine plasma.
Figure 4.
Average viral load expressed as Log10 HAD/mL in different tissues analyzed at d31 of the study. # = P < 0.1; * = P < 0.05; ** = P < 0.01 SDPP = spray-dried porcine plasma.
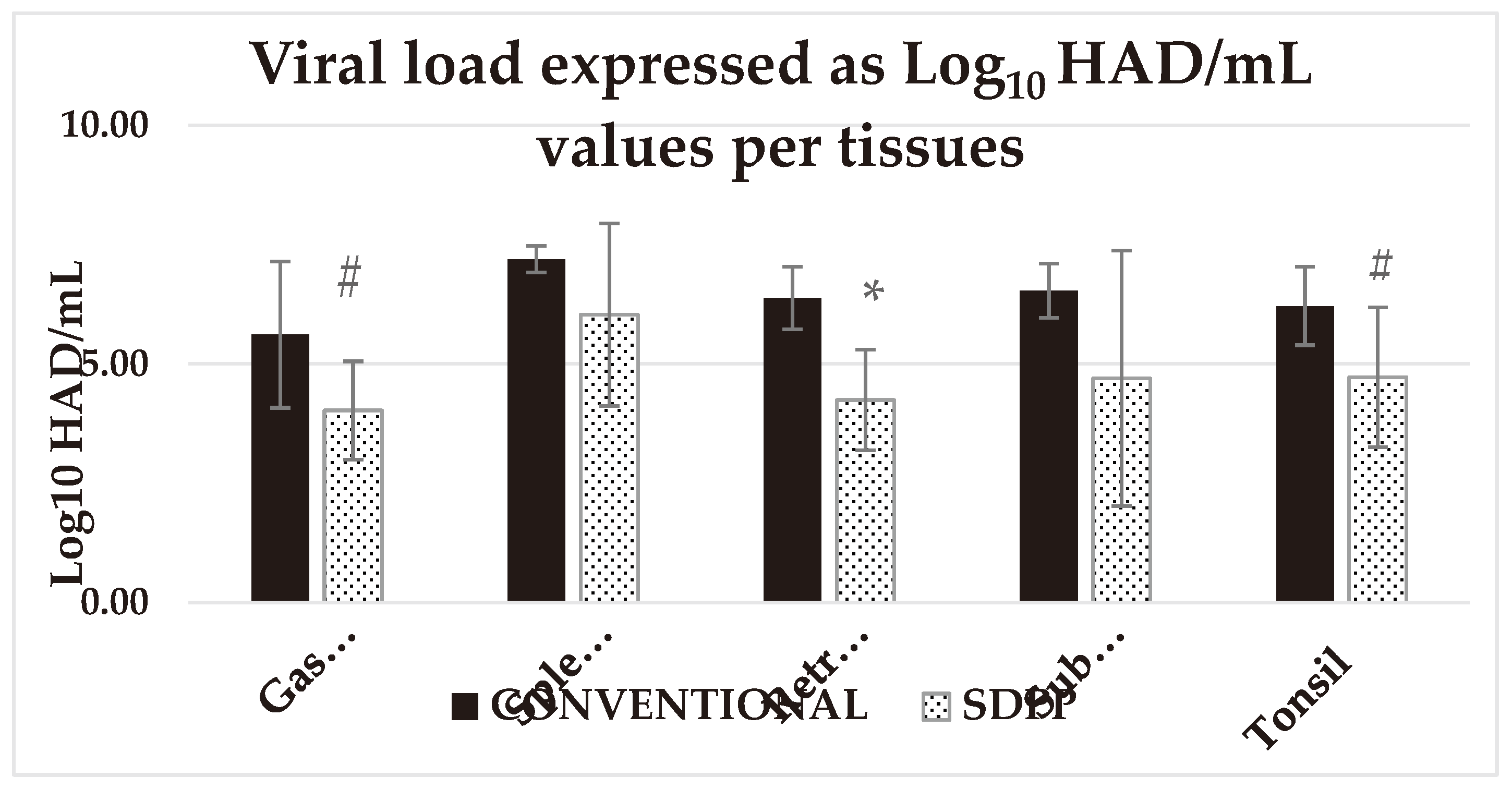
Figure 5.
ASFV-specific IFN-γ secreting T-cells analyzed by ELISPOT at 11 days after first ASFV encounter (11 dpc). PBMC from day 11pse were stimulated in vitro with Georgia 2007/1 and the number of ASFV-specific IFN-γ-secreting cells was quantified by ELISPOT. Values shown are individual values subtracting the corresponding values of mock-stimulated cells. C-#: Animals in CONVENTIONAL diet; SDPP-#: animals in spray-dried porcine plasma (SDPP) diet.
Figure 5.
ASFV-specific IFN-γ secreting T-cells analyzed by ELISPOT at 11 days after first ASFV encounter (11 dpc). PBMC from day 11pse were stimulated in vitro with Georgia 2007/1 and the number of ASFV-specific IFN-γ-secreting cells was quantified by ELISPOT. Values shown are individual values subtracting the corresponding values of mock-stimulated cells. C-#: Animals in CONVENTIONAL diet; SDPP-#: animals in spray-dried porcine plasma (SDPP) diet.
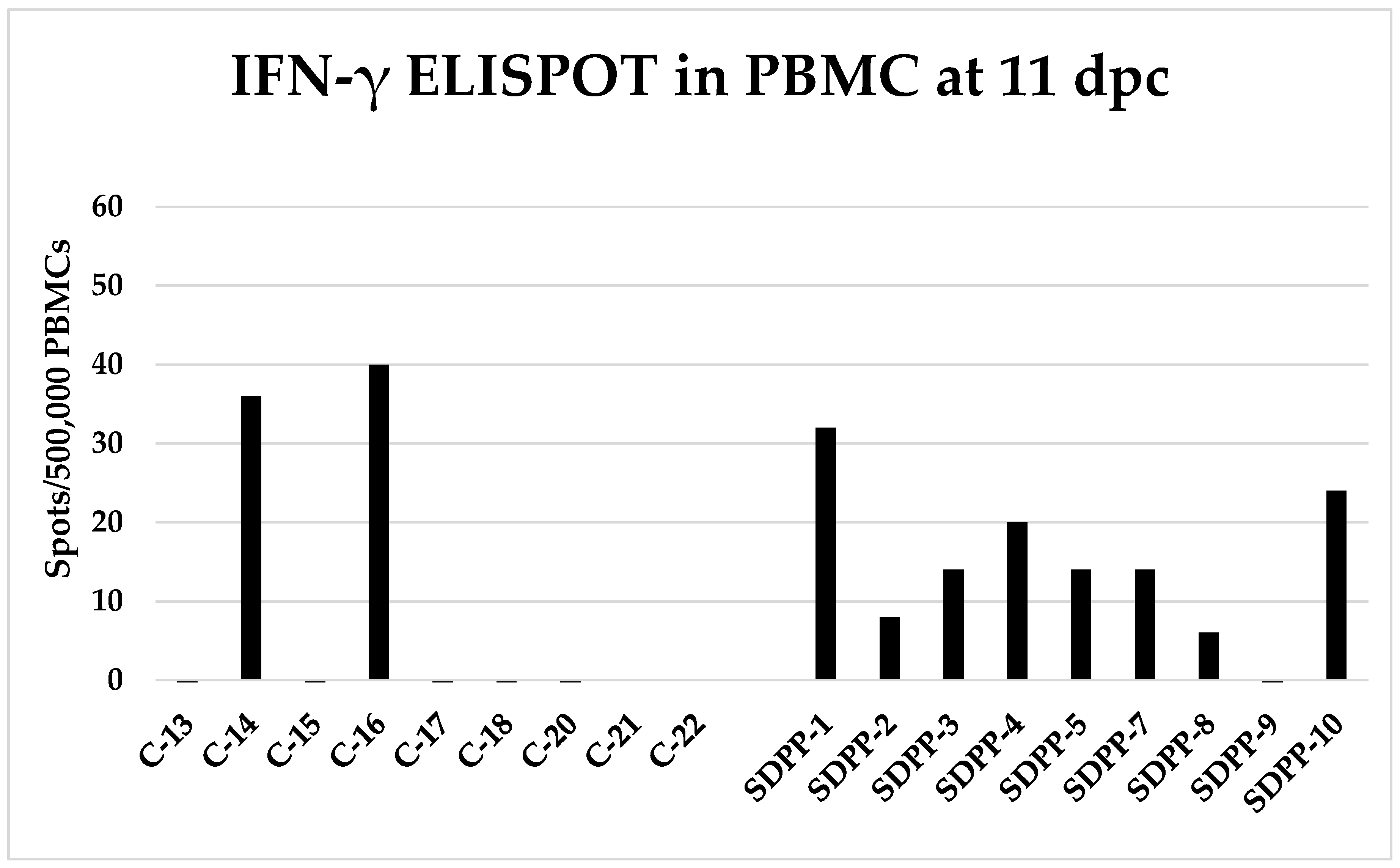

Table 1.
Composition and estimated nutritive composition of the basal experimental diet (g/kg feed).
Table 1.
Composition and estimated nutritive composition of the basal experimental diet (g/kg feed).
| Ingredients | CONVENTIONAL Diet | SDPP diet |
|---|---|---|
| Barley | 298.7 | 313.3 |
| Maize | 295.7 | 303.4 |
| Sweet milk whey | 137.2 | 137.2 |
| Soy protein concentrate | 100.9 | - |
| Spray-dried porcine plasma (SDPP) | - | 80.0 |
| Soybean meal (48% CP) | 107.4 | 110.6 |
| Animal fat | 26.7 | 30.0 |
| Dicalcium phosphate | 14.4 | 16.1 |
| Calcium carbonate | 1.2 | 0.5 |
| Salt | 4.7 | 0.2 |
| L-Lysine-HCl | 4.2 | 2.3 |
| L-Threonine | 1.9 | 0.5 |
| DL-Methionine | 2.1 | 1.4 |
| L-Tryptophan | 0.7 | 0.2 |
| L-Valine | 0.3 | - |
| Noxyfeed* | 0.2 | 0.2 |
| Vit-Min complex** | 4.0 | 4.0 |
| Nutritive composition | ||
| Crude Protein | 195.0 | 195.0 |
| Crude Fiber | 28.0 | 25.2 |
| Fat | 45.5 | 50.3 |
| Ash | 58.2 | 56.6 |
| Energy (kcal ME/kg) | 3340 | 3340 |
| Total sodium | 2.8 | 2.8 |
| Total chlorine | 6.7 | 4.5 |
| Total calcium | 7.5 | 7.5 |
| Total phosphorous | 6.7 | 7.4 |
| Digestible phosphorous | 3.8 | 3.8 |
| Lysine (SID) | 12.50 | 12.50 |
| Threonine (SID) | 8.13 | 8.13 |
| Methionine (SID) | 4.62 | 3.75 |
| Met+Cys (SID) | 7.38 | 7.78 |
| Tryptophan (SID) | 2.50 | 2.50 |
| Isoleucine (SID) | 7.36 | 6.75 |
| Valine (SID) | 8.50 | 9.26 |
| Leucine (SID) | 13.52 | 14.89 |
| Phenylalanine (SID) | 8.02 | 8.52 |
| Phe+Tyr (SID) | 13.41 | 14.60 |
| Histidine (SID) | 4.25 | 4.76 |
* ITPSA, Barcelona, Spain. Contains BHT+ propyl galate (56%) and citric acid (14%). ** Provides per kg feed: vitamin A (E-672) 10000 UI; vitamin D3 (E-671) 2000 UI; vitamin E (alfa-tocopherol) 25 mg; vitamin B1 1.5 mg; vitamin B2 3.5 mg; vitamin B6 2.4 mg; vitamin B12 20 µg; vitamin K3 1.5 mg; calcium panthotenate 14 mg; nicotinic acid 20 mg; folic acid 0.5 mg; biotin 50 µg; Fe (E-1) (from FeSO4·H2O) 120 mg; I (E-2) (from Ca(I2O3)2) 0.75 mg; Cu (E-4) (from CuSO4·5H2O) 6 mg; Mn (E-5) (from MnO) 60 mg; Zn (E-6) (from ZnO) 110 mg; Se (E-8) (from Na2SeO3) 0.37 mg.
Table 2.
qRT-PCR results of different tissues at 12 dpse. Values expressed as Ct values/mL.
| ANIMALS(Group + animal id) | Gastro-Hepatic Node | Spleen | Retropharyngeal Node | Submaxillary node | Tonsil |
|---|---|---|---|---|---|
| CONVENTIONAL #13 | 22 | 16 | 21 | 17 | 19 |
| CONVENTIONAL #14 | 20 | 15 | 20 | 20 | 19 |
| CONVENTIONAL #15 | 17 | 16 | 17 | 16 | 16 |
| CONVENTIONAL #16 | 26 | 17 | 15 | 17 | 20 |
| CONVENTIONAL #17 | 13 | 15 | 22 | 13 | 14 |
| CONVENTIONAL #18 | 23 | 15 | 19 | 17 | 20 |
| SDPP #1 | 30 | 25 | 30 | 30 | 30 |
| SDPP #2 | 25 | 14 | 22 | 16 | 17 |
| SDPP #3 | 31 | 20 | 32 | >35 | 33 |
| SDPP #4 | 24 | 15 | 25 | 18 | 21 |
| SDPP #5 | 35 | 34 | 32 | 20 | 26 |
| SDPP #7 | 25 | 15 | 25 | 27 | 26 |
| Average CONVENTIONAL | 20.1 ± 4.8 | 15.8 ± 1.1 | 19.4 ± 2.6 | 16.8 ± 2.3 | 17.9 ± 2.5 |
| Average SDPP | 28.6 ± 4.2 | 20.5 ± 7.8 | 27.7 ± 4.3 | 25.1 ± 9.2 | 25.8 ± 5.9 |
| P Value | 0.081 | 0.172 | 0.002 | 0.058 | 0.013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



