Submitted:
01 February 2023
Posted:
03 February 2023
You are already at the latest version
Abstract
Vitamin K occupies a unique and often obscured place among its fellow fat-soluble vitamins. Evidence is mounting however, that vitamin K (VK) may play an important role in the visual system apart from the hepatic carboxylation of hemostatic related proteins. However, to our knowledge, no review covering the topic has appeared in the medical literature. Recent studies have confirmed that matrix Gla protein (MGP), a vitamin K-dependent protein (VKDP), is essential for the regulation of intraocular pressure in the mouse. The PREDIMED (Prevención con Dieta Mediterránea) study, a randomized trial involving 5860 adults at risk for cardiovascular disease, demonstrated a 29% reduction in the risk of cataract surgery in participants with the highest tertile of dietary vitamin K1 (PK) intake compared with the lowest tertile. However, the specific requirements of the eye and visual system (EVS) for VK, and what might constitute an optimized VK status, is currently unknown and largely unexplored. It is the intention therefore of this narrative review to provide an introduction concerning VK and the visual system, review ocular VK biology, and to provide some historical context to recent discoveries. Potential opportunities and gaps in current research efforts will be touched upon, in the hope of raising awareness, and encouraging continued VK related investigations in this important and highly specialized sensory system.
Keywords:
vitamin K
; vitamin K1
; vitamin K2
; eye
; visual function
; cognitive function
; vitamin K-dependent protein
; matrix Gla protein
; ferroptosis
1. Introduction
Losing one’s sight consistently ranks first among the most feared of conditions, with blindness seen as the worst possible affliction among a list that includes Alzheimer’s disease (AD), AIDS/HIV, cancer, loss of limb, heart disease, and deafness[1]. As early as 1525 B.C., the Ebers Papyrus highlights this long-standing human concern regarding vision loss, by chronicling numerous eye diseases and treatments, many of which were dietary based[2], and Hippocrates appreciated the importance of the eye as a biomarker and prognostic indicator circa 500 B.C.[3,4].
Blindness in the 21st Century remains an enormous challenge as the global population continues to grow and age. Age-related, as well as behavioral and lifestyle associated conditions, such as glaucoma, cataracts, age-related macular degeneration, and diabetic retinopathy, are assuming greater importance, as eye care services and treatments cannot keep up with demand despite numerous advances [5,6]. The best estimates of global blindness in the year 2020 suggest that 43.3 million people are blind and 295 million have moderate to severe vision loss, and by 2050, the numbers are predicted to increase to 61 million and 474 million respectively [5]. The most recent analysis of the burden of significant visual acuity loss and blindness (VL) in the US population indicates the problem may be much more significant than previously appreciated[7]. The mean prevalence of VL was 2.17% for the year 2017, which increased to 20.37% in persons aged 85 years and older, representing nearly 7.08 million people, 1.08 million of whom have blindness, a 68.7% increase over previous estimates [7]. The cost of VL is significant, with estimates of the yearly US total economic burden of VL in 2017 being $134.2 billion: $98.7 billion direct, and $35.5 billion in indirect costs [8] and the intangible price blindness exacts from those afflicted, and their families, is also substantial[9].
Experts acknowledge that nutrition and dietary interventions are the most powerful modifiable risk factors for disease across the life span, and correction of micronutrient deficiencies can represent a substantial return on investment for individuals, and society [10]. It is also recognized that it is imperative to understand the nutritional factors that can drive the prevention and treatment of age related conditions [11]. Though evidence exists that nutritional inputs can delay such diseases as macular degeneration and cataracts, public health measures related to functional losses are lacking, and fresh approaches to conduct both rapid and rigorous dietary interventions in diverse populations are needed [12]. The relative lack of research with respect to vitamin K in general [13], and with respect to the visual systems in particular, is striking, and deserving of particular attention.
Not a single trial in U.S National Institutes of Health ClinicTrial.gov database was found related to VK and the eye or visual system[14], and there is only one trial in the National Library of Medicine PubMed.gov database related to vitamin K1 and the reduction of cataract surgery risk [15]. Thus, the opportunity of furthering our understanding of the VK biology and specific VK requirements of the visual system is evident, with reports that for each dollar spent on vision loss prevention and treatment, there is at least a 5 dollar return on investment [16].
Vitamin K is a fat-soluble vitamin whose severe deficiency can lead to potentially fatal bleeding and blindness [17], the fact of which has dominated a great deal of its history, but whose long term deficiency has recently been linked with a number of clinical conditions associated with aging[18]. VK is one of four known fat-soluble vitamins, which also includes vitamins A, D, and E. In sharp contrast its fellow vitamins, VK is neither stored in the body to any great degree, nor produced de novo, with plasma concentrations of phylloquinone (PK or vitamin K1) around 0.5 nM, which is 1-4 orders of magnitude less than 25-hydroxyvitamin D, retinal (a form of vitamin A), and α-tocopherol (a form of vitamin E)[19]. Consequently, its physiologic function is predicated upon ongoing and critical intracellular regeneration of its active form, vitamin K hydroquinone (VKH2), dietary intake, gut microbiome synthesis, or therapeutic supplementation[19]. Pool size of PK in healthy volunteers may be as low as 17-194 μg (0.28-2.17 μg/kg) with a mean and (SD) of 87.6 μg (60), and a body pool turnover of approximately 1.5 days [20], highlighting the potentially precarious nature of VK metabolism within the human body.
VK is a general term for 2-methyl-1,4-naphthoquinone or menadione (MD, K3) and any of its derivatives with antihemorrhagic effects [21]. MD a synthetic form of VK used in early work, and its water-soluble derivatives remain an important addition to animal feed, particularly in chickens (and thus to the human food chain), who are acutely vulnerable to VK related deficiency and bleeding. MD is now known however, to be an important endogenously produced intermediary in human VK metabolism as a source of menaquinone-4 (MK4) [21]. As isoprenoid quinones, these compounds are among the most important in living organisms as a result of their ability to share electrons and participate in redox-cycles [22], with menaquinones (MK-n) functioning as part of the respiratory electron transport chain in bacteria, and phylloquinone (PK) participating in electron transport for photosynthesis in plants [23].
The VK isoprenoid quinones are highly conserved evolutionally and their metabolism is complex compared to its fellow fat-soluble vitamins in important ways which remain under intense investigation. The reader is referred to the excellent reviews by Shearer et al. 2014,2018 [19,24]. It has long been held that the only firmly established function of VK is its action as a cofactor for a single membrane bound endoplasmic reticular enzyme, γ-carboxyglutamyl carboxylase (GGCX), and its central role in maintaining hemostasis via the liver is its only health role supported by incontrovertible evidence[19]. However, in addition to supporting hemostasis, extrahepatic vitamin K dependent proteins (VKDP), are now known to effect a broad range of processes of potential importance to the visual system including regulation of tissue calcification, apoptosis, growth control, signal transduction and angiogenesis [25]. In addition, non-canonical VK pathways, unrelated to VKDP, such as the nonenzymatic, antioxidant mediated, inhibition of ferroptotic programmed cell death [26], and activation of the steroid and xenobiotic (SXR) nuclear hormone receptor [27], are being shown to have potentially significant roles in ocular health[28].
Synthesis of VKDPs by GGCX appears to be a unique biochemical process that generates a previously unknown amino acid, γ-carboxyglutamic acid (Gla), with two negative charges, from VKDP glutamic acid (Glu) residues, first reported by Stenflo and Nelsestuen in 1974 [29], as part of their effort to discover the structure and function of prothrombin [30]. This conversion adds an additional negative charge on the Glu residues which allows for a unique calcium ion(Ca++)-protein interaction that had not been previously described [30].
Calcium is the third most abundant metal in nature and Ca++ are the most tightly regulated ions in membrane-bound organisms and organelles, with a 20,000-fold gradient between the intracellular (~100nM) and extracellular (mM) concentrations [31]. Cells, and organelles within them, such as mitochondria and endoplasmic reticulum (ER), must chelate, compartmentalize, or extrude these ions, by binding them to proteins to effect function, localization, and association [31]. Seventeen currently known VKDPs represent such highly specialized proteins in humans [32,33]. Protein function is determined by shape and charge, and Ca++ binding, as well as phosphorylation, trigger these changes in proteins, and thus regulate their function [31]. In nature, six to seven oxygen atoms of carboxyl groups surround and coordinate a single Ca++, forming a pentagonal bipyramid [31,34], changing the VKDP conformation, and thus effecting it’s activation.
A detailed examination of the VK metabolism, VK-related molecular pathways, and VK-dependent protein activation is beyond the scope of this review, and the reader is referred again to the reviews by Shear et al. 2014,2018 [19,24], and a comprehensive review of VK metabolism and VK metabolic pathways by Beato et al. 2020 [35], along with the recent detailed review of VKDP activation via GGCX by Berkner and Runge 2022 [36].
2. Vitamin K biology and the visual system
Limited data exists from which to begin to understand VK metabolism, transport, and pathways within the eye and visual system (EVS). As visual processing is estimated to occupy 55% of the primate neocortex[37], and one of the higher cortical functions of the human brain[38],brain related VK data will be included as important relevant information within this review.
Some of the fundamental questions to be answered are 1) what is the nature of VK action in the eye, and by extension, how might this compare with the brain and central nervous system in general, 2) how is VK trafficked in the EVS, 3) what constitutes optimized VK levels in the EVS, and how is this best achieved and maintained over the life span, and 4) what new biomarkers related to VK status might be present and readily accessible in the EVS? None of the answers to these questions are definitively known.
2.1. Vitamin K content and deliver to ocular and related tissue
Only one study reports the tissue concentration of PK and MK4 in the eyes of female and male mice [39]. The remainder of the data must be extrapolated from reports of PK and MK content and distribution in human brains, as well as animal brain studies, or gleaned from analyses of associated factors in human and bovine vitreous and aqueous humor. Studies related to the delivery of other nutrients to the eye and brain via LDL and HDL also exist.
In their 2008 foundational work, Okano and co-works [39] using deuterium labeling, provided unequivocal evidence that MK4-d7 in the mouse cerebrum originates from dietary PK-d7 and/or MD-d8 by one of two routes, either by the conversion of PK to MD in the intestines followed by MD uptake and conversion to MK4 in the cerebrum, or uniquely, the first evidence of tissue specific, PK>MD>MK4 conversion within the brain. They conclude that their data suggests that the cerebrum is particularly unique as it contained relatively large amounts of MK4 but little to no PK, likely the result of the blood brain barrier (BBB) or other unknown mechanisms [39]. They also provided valuable oral PK>MD>MK4 kinetic data showing in female mice, PK-d7 and MK-d8 were converted into cerebral MK4-d7 over 24 hours in a dose-dependent fashion from a physiological dose of 0.1 μmol/kg body weight to what was considered a pharmacological dose of 10 μmol/kg body weight. Additional they showed that a single oral administration of PK- d7 at 10 μmol/kg body weight resulted in the accumulation of MK-4- d7 in the cerebra at a concentration of 82.9 ± 6.1 pmol/g tissue, nearly 30% of the concentrations (252.5 ± 10.3) of MK-4 in the cerebra of age-matched female mice within 24 hours [39].This conversion of PK to MD had long been understood to occur but was not previously proven. Many aspects of this pathway remain important open questions.
Table 1 of their work is reproduced below and appears to be the only source of detailed information with respect to VK levels in the eyes and 27 other tissues of mice. It can be readily seen that there is marked variability in PK and MK4 concentrations among tissue types and even within different regions of the brain, which is in concert with MK/PK concentration data reported in nine separate areas of the rat brain but without eye associated information reported previously by Ferland in 2004 [40].
Figure 1 is a graph of the tissue distribution of vitamin K (pmol/g) in mice fed a normal diet sorted with respect to female MK4 concentrations.

Table 1.
Tissue distribution of vitamin K (pmol/g) in mice fed a normal diet.
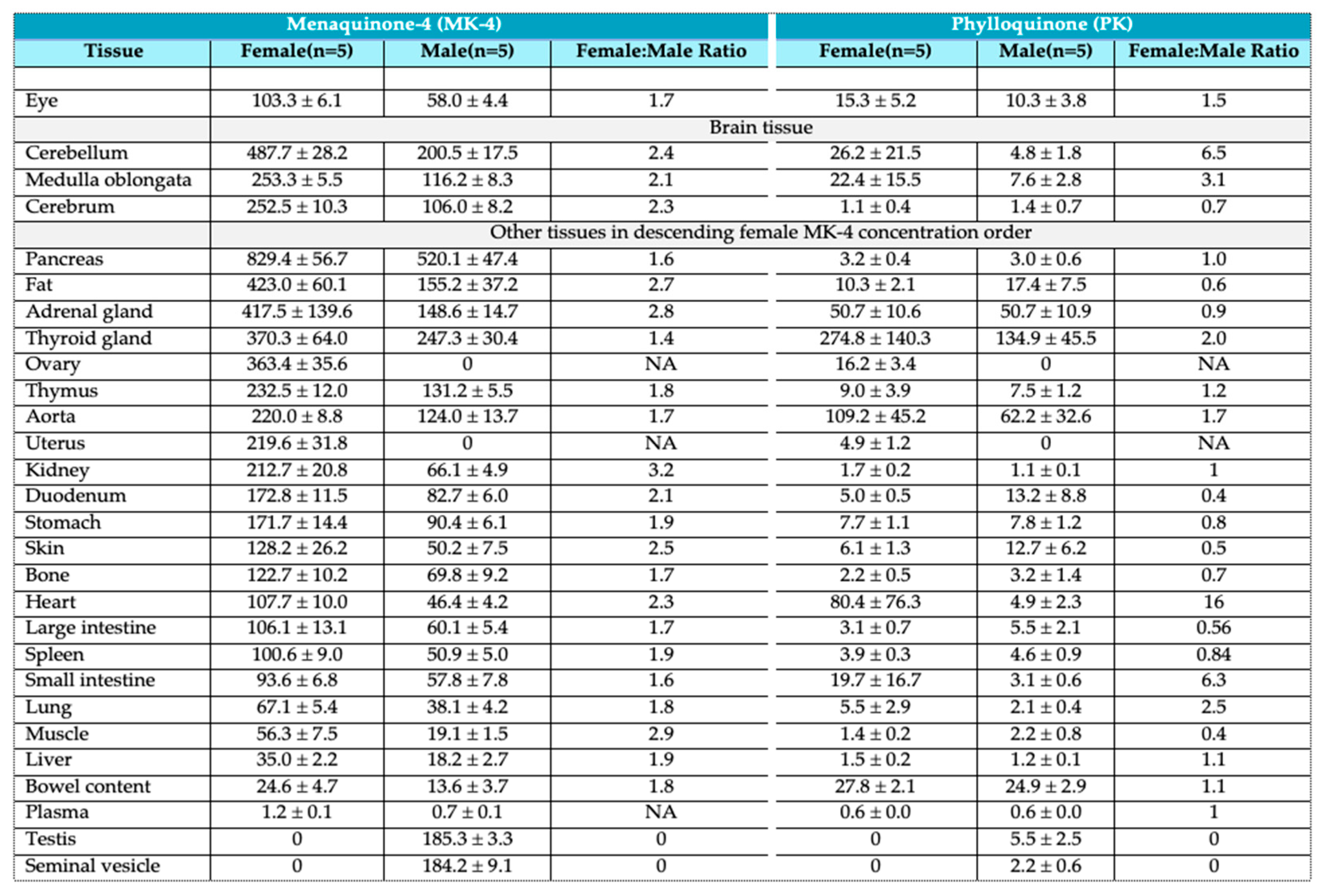
Adapted from: Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of Phylloquinone (Vitamin K1) into Menaquinone-4 (Vitamin K2) in Mice: Two Possible Routes for Menaquinone-4 Accumulation in Cerebra of Mice. J Biol Chem 2008, 283, 11270–11279, doi:10.1074/jbc.M702971200. [39] CC BY license.
Figure 1.
Mean (n=5) tissue MK4 and PK concentration (pmol/g) in mice fed a normal diet sorted by female MK4 concentration. Red color denotes female MK4, Green-Female PK, Orange-male MK4, and Blue-male PK. Note tissue type, gender, female:male MK and PK ratios as well as tissue type and gender MK:PK ratios. (Adapted from: Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of Phylloquinone (Vitamin K1) into Menaquinone-4 (Vitamin K2) in Mice: Two Possible Routes for Menaquinone-4 Accumulation in Cerebra of Mice. J Biol Chem 2008, 283, 11270–11279, doi:10.1074/jbc.M702971200. [39] CC BY license).
Figure 1.
Mean (n=5) tissue MK4 and PK concentration (pmol/g) in mice fed a normal diet sorted by female MK4 concentration. Red color denotes female MK4, Green-Female PK, Orange-male MK4, and Blue-male PK. Note tissue type, gender, female:male MK and PK ratios as well as tissue type and gender MK:PK ratios. (Adapted from: Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of Phylloquinone (Vitamin K1) into Menaquinone-4 (Vitamin K2) in Mice: Two Possible Routes for Menaquinone-4 Accumulation in Cerebra of Mice. J Biol Chem 2008, 283, 11270–11279, doi:10.1074/jbc.M702971200. [39] CC BY license).
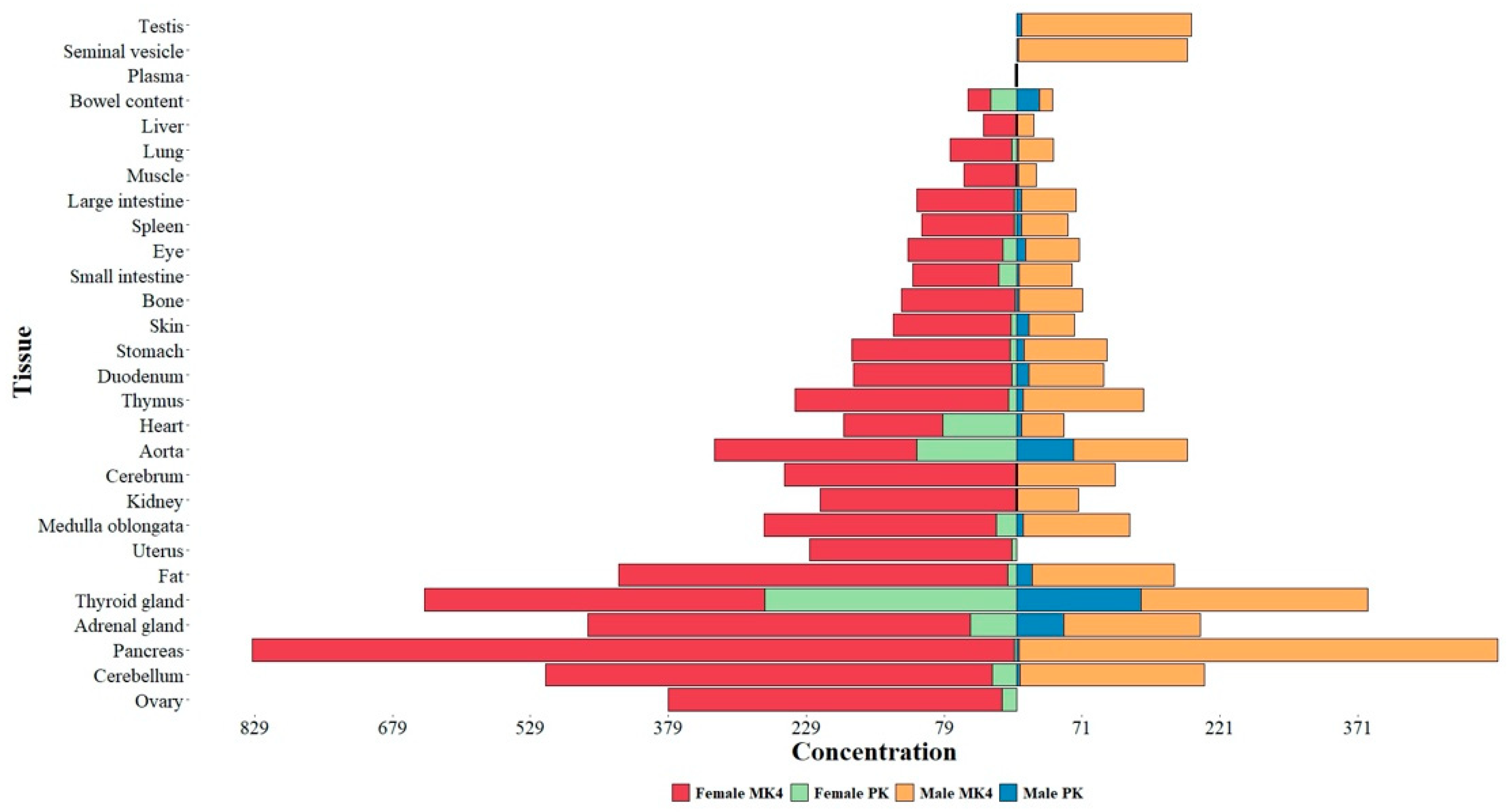
Importantly, the eye was found to contain both PK and MK4 with a MK4/PK ratio of 6.75 and 5.63 in females and males respectively. PK and MK4 values are consistently higher in females with a ratio of 1.49 for PK and 1.78 for MK4 in the eye. The eye appears to be enriched with respect to PK compared to the cerebrum and to be reduced in MK-4 with ratios of 0.41 and 0.55 in females and males respectively. PK values would appear less certain given smaller concentrations and larger SE values. Caution should be taken in drawing too many conclusions from this preliminary ocular data as the primary purpose of the study was to understand the generation of MK4 within the mouse cerebrum. The eye, in part an embryologic extension of the brain, is however much less homogeneous than the brain, containing many different tissue types, including a large volume of vitreous humor which is 99% water, thus giving rational for the undertaking of an aggressive program to map tissue specific MK4 (and possible other MKn) and PK concentration of the human eye.
Building upon their work, Nakagawa, Hirota, and colleagues in 2013 [41] demonstrated unequivocally using deuterium-labeled PK (PK-d7), that tissue MK4 in rats is primarily derived from dietary PK that is converted to MD in the intestines and released into the general circulation by way of the lymphatic and circulatory systems. They further demonstrated a similar time course change in PK, MK4, and MD concentrations in healthy adult volunteers that they believed provided evidence of similar action in humans. Their kinetic data suggested that the bulk of the PK was absorbed in the intestine and passed unaltered via the mesenteric circulation to the liver, but that a smaller, yet critical amount of PK, was converted to both MD and MK4 in the intestines and entered the mesenteric lymphatics. The lymph concentrations of PK-d7 were approximately 50-fold higher than those of MD-d7 and 1,000-fold higher than those of MK4-d7. They maintain that their tissue concentration data, along with their previous work [39], indicates that PK is mainly stored in the liver, and that the cerebrum actively accumulates MK4 in rats and mice, with mice accumulations being higher than rats. Unfortunately, no ocular data was provided in this work.
The authors comment that their results are in concert with Thijssen et al. 2006 [42] who found that approximately 5-20% of an oral dose of PK in healthy male volunteers was catabolized to MD, and that unsupplemented subjects had a daily excretion of MD in the urine of 1.6-9.1 μg. The implication for the eye and brain are clear, as both Nakagawa and Thijssen note, that the smaller lipophilic MD can more easily pass the blood-brain barrier (BBB), and by extension, the blood-retinal barrier (BRB), to supply intracellular MK4 upon conversion. Partridge 1998 [43] and Vita et al. 2011[44] detail that lipid-soluble molecules with a mass under 400-600 Da are transported readily by lipid mediated free diffusion through the vascular endothelial plasma membranes and are desirable for central nervous system drug deliver, which may indicate that MK is able to rapidly and passively enter the circulation after conversion in the enterocytes. The molecular weight of MD is 172.2 Da, and by contrast-MK4 is 444.6, PK is 450.07, MK7 is 649.0, MK9 is 785.23, and MK13 1057.7. It is important to note that this property of MD may be a two-edged sword, in that the use of MD as a pharmacologic agent to treat VK deficiency was stopped long ago, as it caused hemolytic anemia, and liver damage [21], but it is now being used in small titrated doses along with high dose vitamin C to treat glioblastoma [44]. Thus, there appears to be a very elegant sorting mechanism by which abundant PK, and possibly all MKn dietary forms, as recently demonstrated by Ellis et al. 2021[45], are converted to the smaller more lipid soluble MD in a titrated manner to facilitate delivery to the brain and eye, while larger forms are packaged into chylomicrons, LDL, and HDL, for export via the vascular circulation to the liver, pancreas, kidney, bones, eye, brain, and other sites.
With respect to human studies, Thijssen et al. 1996 [46] report on six autopsy cases of which three contributed brain tissue. PK was found in all tissues type with relatively small amounts in the brains with a median and (SD) of 1.5 (0.7) pmol/g. MK4 levels were relatively high in the brain, with a median of 6.3 (2.8) pmol/g. MK4 exceeded PK levels only in the brain and kidney. The MK4/PK ratios in the brains were 2.4, 3.4, and 13.3. with a mean of 6.4 (6.0) (not unlike the MK4/PK ratios of 5.63-6.75 reported by Okano in 2008 in mice eyes). In contrast, the livers were found to have total VK stores ranging between 40-125 pmol/g wet weight with PK ranging from 4-45% of the total. Thus, both PK and MK were consistently found in the human brain.
Tanprasertsu et al. in 2021 [47] reported on the MK4 and PK levels in the frontal and temporal cortex of 45 decedent centenarians in the Georgia Centenarian Study with and without dementia. The overall mean and (SD) concentrations for MK4 were 4.96 (2.32) and 0.40 (0.39) pmol/g for PK, which were not significantly different between those with and without dementia. Of interest and potential significance, all brains were found to contain MK4, but in 38% no PK was detected. The MK4/PK ratio was 12.4 among all subjects which is within the range reported by Thijssen. Interestingly, in a companion paper reporting on the same individuals [48], brain retinol mean pmol/g and (SD) levels were 691.97 (305.56) and α-tocopherol mean pmol/g and (SD) levels were 66,917 (13,676). The ratios of retinol to MK4 and PK were thus 140 and 1730 respectfully, and that to α-tocopherol were 13,491 and 167,293, both highlighting and reaffirming the low relative tissue levels of VK compared to other fat-soluble vitamins[19].
Further brain VK data was found in a retrospective analysis of frozen brain tissue. Fu et al. 2021[49] report on 499 participants in the Rush Memory and Aging Project (mean age 92 y, 72% female) and found the MK4 levels in brain tissue stored from 0.4 to 13.0 years was 1.2 pmol/g, which they note is much lower than others have found. PK levels were reported as non-detectable, though small amounts were seen below levels able to be accurately reported.
In a recent major advance, Booth et al. in 2022 [50], reporting on a subset of the Rush Memory and Aging Project, presented data linking VK content in 325 human brains, for the first time, to post-mortem neuropathologic histology and cognitive function prior to death. Brain MK4, mean pmol/g and (SD), ranged from 1.51 (5.5), 0.73 (4.6), 1.61 (7.2) in the mid-frontal/mid-temporal cortex, anterior watershed area, and cerebellum respectfully. Brain MK4 concentrations were inversely correlated with Alzheimer’s disease (AD) global pathology, neurofibrillary tangle density, Braak stage, and Lewy body presence, all well-established markers of neurodegeneration, as well as positively correlated with higher cognitive function and a slower rate of cognitive decline prior to death. Both the neurodegenerative findings and cognitive decline have clear connections to visual function and clinically relevant anatomic correlates to ocular findings, which will be addressed subsequently. Of interest, average brain MK4 concentrations, which were very carefully assayed and validated, stood on the order of three to five times less than those reported by Tanprasertsu et al., and Thijssen et al., which may reflect differences in the way brains were handled and stored, as well as differences in assay protocols, or possibly, actual tissues variances among the populations.
Thus, the available data would suggest that MK and PK are consistently found in human, mouse, and rat brain tissue with MK4/PK ratios that vary, but that consistently favors MK4. Mouse evidence documents MK4 and PK in the eye with MK4/PK ratios of 6.75-5.63 in females and males respectfully. MK4 and PK levels in human eyes are currently unreported but should be able to be quickly confirmed. Further, a portion (possibly 5-20%) of dietary PK, MK4, and likely MKn forms as well, are converted to MD in the intestines, passively absorbed into the circulation because of its small M.W (172.2 Da) and lipophilicity; key physicochemical properties determining absorption, distribution, metabolism, excretion, and toxicity (ADMET) of a compound [51]. MD may then rapidly cross the BBB and BRB to be taken up by the cells in the brain and eye and quickly converted to MK4. In addition to providing an elegant mechanism to overcome the important BBB and BRB, and likely optimize brain and ocular VK levels, this PK>MD>MK4 conversion has long suggested to experts[19,21,41,46,52], that MK4 likely has important intracellular functions in addition to carboxylation of VKDP’s.
Corollaries to the above discussion are the facts that 75% or more of the dietary PK and MK is not converted to MD, and that the conversion PK> MD>MK4 does not explain the mechanism for the accumulation of PK in the eye and brain. PK and MK, as lipophilic vitamins, along with cholesterol and carotenoids such as lutein and zeaxanthin , are insoluble non-swelling amphiphilic polar lipids, absorbed along with other dietary fats as has been expertly reviewed by Ji et al. in 2009 [53] and again by Shearer et al in 2012 [54], and more recently by Lai et al in 2022 [55]. Important with respect to the ocular system is the fact LDL and HDL each carry approximately 7% of serum associated PK [56], with LDL containing both MK4 and MK9 forms as well, while HDL was found to carry only MK4 [57], which may be particularly important with respect to MK4 preferential concentration in the eye and brain.
In the early 1980’s it was pointed out that lipids were present in human aqueous humor (AH) at a mean concentration of 16.4 mg/dl, with HDL felt likely to be the sole lipid transporting lipoprotein, at concentrations of 4 micrograms/ml. This suggested to the authors that HDL was transported into the anterior chamber via AH, and that HDL may be an important source of lipids to the lens and other anterior chamber tissues [58,59]. The near exclusive and relatively abundant presence of HDL related apoproteins in the AH, and only rare or absent LDL related ApoB lipoprotein, has been confirmed using state of the art liquid-chromatography mass spectrometry (LC-MS/MS) by two groups in 2009 and 2020 [60,61]. Further, Conner et al. 2007 [62] and Li et al. [63] have demonstrated the unique role HDL plays in delivery of carotenoids to the retinal pigment epithelium and retina, and likely by extension, MK4 as demonstrated by Schurgers and Vermeer 2002 [57]. Thus, the importance of HDL transporters in ocular VK biology are implicated.
Yamanashi et al. 2017 [64], have recently linked the intestinal absorption and export of dietary PK by enterocytes to membrane proteins previously known to be essential in cholesterol metabolism, including Niemann-Pick C1 like 1 (NPC1L1), ATP-binding cassette transporter A1 (ABCDA1), scavenger receptor class B type 1 (SR-B1), and cluster of differentiation 36 (CD36). These proteins are expressed in the retina [65] , suggesting the retina maintains internal lipid processing machinery that involves HDL-like particles of which PK and MK4 are likely components [66]. Genome-wide associations studies have linked these HDL transporters and their receptors to carotenoids levels in the human retina [63], and the transporter ABCDA1 has also been linked to primary open angle glaucoma risk [67]; indicating these transporters and their participation in VK biology need to be considered in further research, if a more complete understanding of the role of VK in the EVS is to be realized.
Thus, a fundamental gap exists with respect to essentially all aspects of the VK forms, tissue distribution, concentrations, and trafficking within the human eye, and to a lesser extent the human brain. These studies are required and urgently needed prior to any well designed clinical trials of therapeutic interventions and solid recommendations with respect to optimized VK dietary intake or supplementation throughout the life cycle.
2.2. Vitamin K dependent proteins in the ocular/visual system.
Having established that VK is found in the mammalian eye, as well as in human, mouse, and rat brains, and that MK4 levels in human decedent brains, are correlated with recognized dementia-related neuropathologies and cognitive function prior to death; it is appropriate to begin to address the nature of VK action in the EVS.
Broadly, VK function can be categorized as canonical, primarily referring to the gamma carboxylation of hepatic coagulation related VKDPs, and secondarily to the gamma carboxylation of non-hepatic VKDPs; and non-canonical, referring largely to VK quinones ability to share electrons in biologic systems.
It has been nearly fifty years since the mode of action of VK, γ-carboxylation of glutamic acid, has been unambiguously demonstrated in relationship to its activation of VKDPs by Nelsestuen et al. in 1974 [68], and noted by Suttie in his excellent review of vitamin K-dependent protein synthesis in 1993[30]. There are currently felt to be 17 VKDPs [33]. Interestingly, γ-carboxyglutamyl carboxylase (GGCX), which is abundantly found in brain tissue [69], as well as in the retina [70], has also been shown to be a VKDP [71], though it is essentially never mentioned in a list of VKDPs. Importantly, very few works regarding VKDPs in general mention these proteins in the brain, and even fewer, with respect to the EVS.
Ferland in 1998 [32] reviewed the 13 then known VKDPs and classified seven as related to blood coagulation, three related to bone metabolism, and four as “other”, with largely unknown functions, apart from Gas6, as a regulator of cellular growth. No mention of VKDPs in the eye or brain were noted.
Having conducted ground breaking work in brain sphingolipids, as well as the relationship of VK intake and cognitive function in rats and humans, Ferland subsequently published a comprehensive review of the action of VK in the nervous system in 2012 [69], in which the roles of Gas6 in cell survival, growth, and myelinations, and Protein S in signal-mediated neuroprotection, and local antithrombotic activity in the brain, were discussed. Again, no mention of VK or VKDP action in the EVS was noted.
In an expansive effort, McCann and Ames 2009 [18], systematically reviewed VK and VKDP within the framework of the “Triage Theory” of micronutrient distribution and allocation, which posits that “when the availability of a micronutrient is inadequate, nature insures that micronutrient-dependent functions required for short-term survival are protected at the expense of functions whose lack has only long-term consequences, such as the diseases associated with aging” [18]. This work is the most detailed coverage of the major known VKDPs, and though limited, the only review to mention extrahepatic VKDP and their action in the eye, referencing prothrombin (Factor II) in the cornea, and the role of Tgfbi in corneal dystrophy (which recent research calls into question with respect to its activation by VK [72]).
Before addressing the VKDPs specifically found in the EVS, brief mention should be made of the importance of VK and the seven hepatic VKDP clotting factors with respect to hemorrhagic visual complications. A properly functioning clotting system is essential for preservation of both life and visual function. Vitamin K deficiency in the newborn period can present with blindness in survivors [73]. Children treated with vitamin K antagonists (VKA such as warfarin) and not properly monitored, have been blinded due to orbital hemorrhages [74]. Numerous cases in the literature report major ocular complications associated with the use of VKA in adults as well [75,76]. Thus, disruption of hepatic VK metabolism from any number of causes, has long been known to result in potentially blinding complications, and we can now turn our attention to the lesser known, extrahepatic VKDPs and their localization, and potential importance to the EVS.
To date, four VKDPs along with GGCX have been shown to have expression and activity in ocular tissues. Using technology they largely pioneered and developed to identify and isolate differentially expressed genes from very small human tissue samples [77], Borrás et al in 2000 [78] reported for the first time, that the gene coding for matrix Gla protein (MGP) was among the most highly expressed in human trabecular meshwork tissue. Later in 2000, Canfield et al, after having studied the osteogenic potential of bovine retinal vascular pericytes (BRVP) for more than a decade, reported MGP was differentially expressed by BRVP during deposition of a calcified matrix [79]. Borrás et al. went on to carefully prove over the next two decades that MGP was essential to maintain physiologic intraocular pressure [80], as well as to show the presence of MGP gene expression in the trabecular meshwork (TM), sclera above the TM, in the ciliary muscle, adjacent to the optic nerve in peripapillary sclera [81], and in the vascular smooth muscle cells of the retinal vasculature [82]. Most recently in 2021, Sarosiak et al. [83] demonstrated the presence of MGP, and active VK metabolic pathways, in the corneas of normal controls, and patients with Schnyder corneal dystrophy.
Collett et al. in 2003 reported that bovine retinal capillary pericytes (BRCP) produced Gas6 [84], and Valverde et al. 2004 found Gas6 in bovine vitreous humor [85], while in a series of papers between 2001-2005, Hall et al. demonstrated that both Gas6 and Protein S were essential for the phagocytosis of retinal outer segments (ROS) by the retinal pigment epithelium (RPE) [86–88].
Finally, prothrombin (Factor II), has been shown to be produced locally at all levels in human corneas, and participates in corneal wound healing [89]. Table 2 below summarizes the currently known ocular associated VKDP, their location, number of Gla residues, and proposed functions.
Table 2.
Summary of ocular related VKDP and their function.
| Author, Year Country [Ref] |
Location | Protein [Number of Gla Residues] |
Expression | Function |
|---|---|---|---|---|
| Sarosiak et al. 2021 Poland [83] |
Human Cornea | Matrix Gla Protein (MGP) [5 Gla] |
Epithelium, Stromal Fibroblasts and Keratinocytes |
Essentially unknown, possible maintenance of calcium homeostasis and anti-calcification role. Possible role in calcium signaling of epithelial proliferation and differentiation [53] |
| Borrás et al. 2000-2021 USA [78,80,81,82] |
Human & mouse trabecular meshwork (TM), Peripapillary Sclera (SC), Retinal vascular smooth muscle cells (VSMC) |
MGP [5 Gla] |
TM, SC, Capillaries, and pericytes |
Essential to maintain physiological IOP |
| Ayala et al. 2007 USA [89] |
Human Cornea | Prothrombin [10 Gla] | All layers of the cornea, intracellularly and in extracellular matrix | Production of thrombin, corneal wound healing, regulation of growth factors, and other signaling molecules via protease-activated receptors. |
| Hall al. 2001-2005 USA [86,87,88] |
Rat Photoreceptor Outer Segments (POS), Retinal Pigment Epithelium (RPE) |
Growth Arrest Specific 6 (Gas6) [11 Gla], Protein S (PROS1) [11 Gla] |
RPE, POS | Phagocytosis of POS by the overlying RPE via vitamin K-dependent gamma-carboxylation of Gas6 and PS in a Ca2+ mediated linking of POS to RPE Mer receptors for phagocytosis |
| Valverde et al. 2004 USA [85] |
Vitreous humor |
Gas6 [11 Gla] |
Bovine vitreous humor | Gas6/Axl signaling in lens epithelial cells support cell growth and survival. |
| Collett et al. 2003 UK [84] |
Bovine retinal capillary pericytes |
Gas6 [11 Gla] |
Bovine retinal capillary pericytes | Interactions in regulating osteogenesis and diseases involving ectopic calcification. |
| Canfield et al. 2000 UK [79] |
Bovine retinal capillary pericytes | MGP [5 Gla] |
Bovine retinal capillary pericytes | Regulation cell differentiation and calcification |
3. Vitamin K and Specific Ocular Conditions
3.1. Glaucoma
Glaucoma is the leading cause of irreversible blindness, with a calculated 3.6 million blind in 2020, and an estimated age adjusted prevalence of 2.04 (per 1000) worldwide[90]. It is a multi-factorial ocular condition characterized by optic neuropathy and generally associated clinically with increased intraocular pressure[91].The ongoing reduction of intraocular pressure remains the only proven treatment for the condition, and currently there is no cure[92,93]. The intraocular pressure is determined by the resistance to aqueous humor flow generated by the trabecular meshwork (TM), located in the anterior chamber of the eye at the juncture of the iris and cornea [94],with most of the resistance believed to be a function of the modulation and turnover of the extracellular matrix (ECM) in the region [95,96].
The VKDP MGP has been conclusively demonstrated to play a central role in glaucoma. Borrás et al. 2020 [80] have shown that the mouse Mgp gene is both responsible and sufficient to maintain normal pressure in the eye, though the exact mechanisms remain to be determined. This discovery was the result of meticulous work beginning more than two decades ago [77,78], demonstrating that the gene encoding MGP, is among the 10 most expressed genes in the human TM, and that MGP activity correlated with the calcification process [97]. They also demonstrated that MGP is heavily expressed in the TM, regionally in the sclera overlying the trabecular meshwork, and surrounding the optic nerve [81].
A detailed recounting of the twenty-year effort and cardinal milestones in this research achievement from the Borrás lab is beyond the scope of this review, but is a remarkable accomplishment and worthy of recognition-one that may well usher in a new era of glaucoma gene therapy, a welcome option for possible glaucoma treatment in the future.
No studies in humans have yet been done to directly investigate VK with respect to glaucoma. However, Wei et al 2018 [98] demonstrated circulating immature nonphosphorylated and uncarboxylated MGP (dp-ucMGP), a marker of reduced VK nutritional status, is a long-term predictor of reduced retinal arteriolar diameter, and referenced a number of studies linking retinal microvascular traits and generalized arteriolar narrowing with glaucoma [98]. A single in vivo study exploring the effect of dietary PK in a surgically induced ocular hypertension rat model was found [99]. Deng et al. found that rats fed a high PK diet for a total of seven weeks were protected from retinal ganglion cell (RGC) loss and TM injury. MGP levels were supported, and there was a transient reduction in intraocular pressure (IOP) at two weeks. Serum PK levels were 0.4 ± 0.52 ng/mL vs. 7.62 ± 5.98 ng/mL; P = 0.054 in controls vs. the experimental group, which was a 19x elevation, yet no significant adverse effects were found on coagulation parameters or body weight.
In 2018 a systematic review and meta-analysis of the effects of vitamins on glaucoma, encompassing 629 publications, found only dietary vitamin A to have a significant association with primary open-angle glaucoma (POAG), having a pooled OR [95% CI] of 0.45 [0.30–0.68] (I2 = 0%) [100]. The authors noted that no studies specifically related to VK were found, but importantly, four studies related to green leafy vegetables intake, (recently shown in a prospective randomized cross over clinical trial to effect statistically significant improvement in VKDP parameters [68]), demonstrated significant negative associations with POAG. Increased dietary green leafy vegetables intake was associated with a 20-69% lower risk of glaucoma [101–104]. Though the quality of the studies varied, the results suggest that even a relatively modest increase in green leafy vegetables, which are the main source of VK in the diet, may have a significant impact on the risk of glaucoma. Table 3 below provides a summary of the studies.
Table 3.
Glaucoma and dietary intake studies.
| Author, Year, Country [Ref] |
Subjects | Design | Intervention | Findings |
|---|---|---|---|---|
| Coleman et al. 2008 USA [101] |
1,155 women with glaucoma in at least one eye |
Observational Cross-sectional cohort | Block Food Frequency Questionnaire |
Decreased glaucoma risk by 69% (odds ratio [OR], 0.31; 95% confidence interval [CI], 0.11 to 0.91) in women who consumed ≥ one serving per month of green collards and kale compared with those who consumed < than one serving per month |
| Moïse et al. 2012 Africa [102] |
500 African type II diabetics |
Cross-sectional design |
Mediterranean-style dietary score (MSDPS) using the Harvard semi quantitative FFQ adapted for Africa. |
Regular intake of the Mediterranean Diet and the consumption of locally grown vegetables including Brassica Rapa, dry beans, Abelmoschus esculentus, and Musa acuminata reduced significantly the absolute risk of glaucoma. |
| Giaconi et al. 2012 USA [103] |
584 African-American women participants in the Study of Osteoporotic Fractures |
Observational Cross-sectional cohort |
The Block Food Frequency Questionnaire | >1 serving/week compared to ≤1 serving/month of collard greens/kale decreased the odds of glaucoma by 57% (OR = 0.43; 95% CI: 0.21–0.85) |
| Kang et al. 2016 USA [104] |
Prospective cohorts of the Nurses’ Health Study (63 893 women; 1984-2012) and the Health Professionals Follow-up Study (41 094 men; 1986-2012) | Prospective cross-sectional observational study | Primary exposure was dietary nitrate intake. Information on diet and potential confounders was updated with validated questionnaires |
Very robust study demonstrating greater intake of dietary nitrate and green leafy vegetables was associated with a 20% to 30% lower POAG risk; and a 40%-50% lower risk for POAG with early paracentral VF loss at diagnosis, in 1483 incident cases identified from 63 893 women and 41 094 men followed for more than 25 years |
Thus, recent work confirms a role of VK with respect to glaucoma and support the need for further research. A mouse model confirms the essential function of the VKDP MGP in glaucoma which may serve as a powerful platform for future VK ocular research. In vivo studies in the rat have shown that PK may be effective in preserving RGC cells and addressing TM injury. Several observational studies have found that increased VK intake may reduce the risk of glaucoma, and that systemic VK status is associated with retinal arteriolar narrowing, a risk factor associated with glaucoma.
3.2. Cataracts
Cataracts are the leading cause of reversable blindness worldwide [105], with a global age-standardized pooled prevalence estimate (ASPPE) of 17.20% (95% CI 13.39–21.01) [106], indicating that for 1000 people randomly sampled anywhere, 133-210 would be estimated to have cataracts. Age was found to correlate with cataract type directly, and age was the source of variance for cataract prevalence, and regional differences [106]. This is not surprising given the fact that fetal lens cells persist for the lifetime of an individual [107], and are felt to serve as an excellent model of the aging process.
Despite advancements, unoperated cataracts remain ophthalmology’s major unsolved problem [108], and efforts to train and support additional surgeons cannot keep pace with aging demographics [105]. Measures to prevent or treat cataracts have met with limited success, but antioxidants and free radical scavengers have shown potential in the lab, and thus further research and clinical trials are needed [109].
In a secondary analysis, Camacho-Barcia et al. 2017[15] analyzed prospectively collected PK intake using baseline and yearly food frequency questionnaires in a group of 5860 individuals in the Prevención con Dieta Mediterránea Study (PREDIMED) [110]with a the mean (SD) age of 66.3 (6.1) years, followed for a median of 5.6 years. After adjusting for multiple potential confounders, individuals in the highest tertile of mean energy-adjusted dietary vitamin K1 intake, had a lower risk of cataract surgery than those in the lowest tertile (HR, 0.71; 95% CI, 0.58-0.88; P = .002). A total of 768 individuals underwent cataract surgery. Of note, the European Food Safety Authority has set an adequate intake (AI) of 1 μg phylloquinone/kg body weight per day for all age and sex groups [111], and though the PREDIMED study did not report body weights, the average weight for adults in Spain are 82 Kg and 66 Kg for males and females [112], the United States and Canada AI’s are 120 and 90 μg/day[113] . Thus, on average, the estimated daily intake of PK in this cohort was 2.75-5.5 times the recommended AIs on the low and high ends respectively, yet a near doubling of the PK intake from 249.4 to 496.7 µg/d was associated with a 29% reduction in the hazard ratio (HR) for cataract surgeries comparing the lowest to highest tertiles. A review of the supplemental data reveals that daily average green leafy vegetable intake was 48.2 g/day and 130.2 g/day in the two groups, representing a difference of approximately 1/2 cup of cooked kale per day [113].
Though experts have pointed out a number of limitations in the study methodology with respect to the estimation of dietary PK intake [114]; the study population size, control for multiple confounding dietary and other variables, the firm end point of a large number of surgeries, along with the 29% relative risk reduction in such a clinically relevant indicator that occurred in close to 1 in 8 participants, may well serve as a strong rational for the undertaking of well-designed interventional studies with respect to VK and cataract prevention. Of interest, and possibly strengthening this assertion, is a companion study by the authors [115] reporting on these same individuals, showing that the incidence of cataract surgery was similar in the interventional MedDiet groups supplemented with olive oil or nuts, compared with the low-fat diet group, suggesting that possibly a doubling of PK intake itself may possibly be protective, while other more general dietary factors are not. Certainly, well designed interventional studies will be needed to define possible causality.
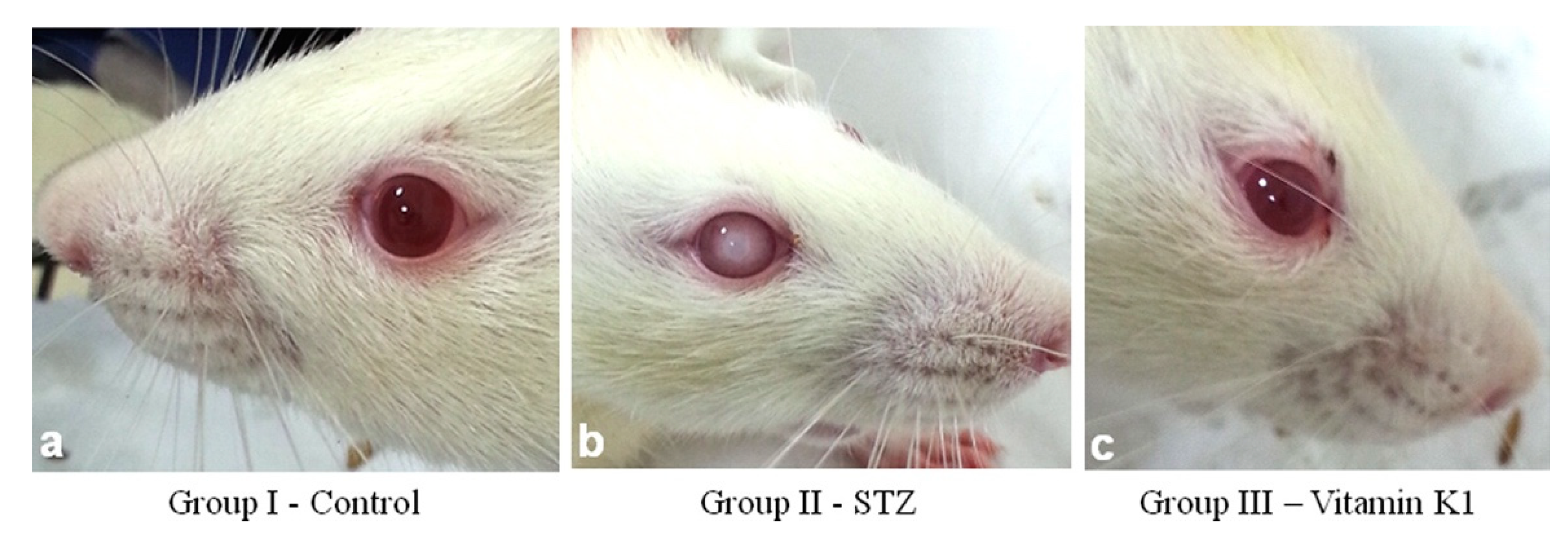
In a series of animal studies beginning in 2014, Varsha and Thiagarajan et al. [116–118], demonstrated in a streptozotocin (STZ)-induced diabetic model in Wistar rats, that PK was able to prevent the formation of lens opacity by modulating lens Ca++ homeostasis and hyperglycemic effects through direct action on the pancreas. In a second experiment they found that PK was able to reduce rat lens sorbitol via direct competitive inhibition of DL-glyceraldehyde binding to lens aldose reductase 2 (ALR2). Lens calcium in PK treated rats was lower than STZ treated rats and even slightly lower than control. Likewise, Ca++-ATPase activity was higher in PK treated rats compared to the STZ group and controls. This suggested to the authors that PK has potential to inhibit diabetic cataracts by supporting pancreatic function, inhibiting lens oxidative stress, and directly modulating lens Ca++ levels. Figure 2, shows PK inhibition of STZ-induced diabetic-cataracts in Group III from Sai Varsha et al. 2014[118].
Figure 2.
Effect of vitamin K1 on diabetic cataractogenesis. Control group I (2a), cortical cataract observed in Group II STZ induced diabetes (2b), treatment of diabetic rat with vitamin K1 prevented appearance of cataract in group III rats (3c). Adapted from: Sai Varsha, M.K.N.; Raman, T.; Manikandan, R. Inhibition of Diabetic-Cataract by Vitamin K1 Involves Modulation of Hyperglycemia-Induced Alterations to Lens Calcium Homeostasis. Experimental Eye Research 2014, 128, 73–82, doi: 10.1016/j.exer.2014.09.007 [118]. Copyright Elsevier-used with permission.
Figure 2.
Effect of vitamin K1 on diabetic cataractogenesis. Control group I (2a), cortical cataract observed in Group II STZ induced diabetes (2b), treatment of diabetic rat with vitamin K1 prevented appearance of cataract in group III rats (3c). Adapted from: Sai Varsha, M.K.N.; Raman, T.; Manikandan, R. Inhibition of Diabetic-Cataract by Vitamin K1 Involves Modulation of Hyperglycemia-Induced Alterations to Lens Calcium Homeostasis. Experimental Eye Research 2014, 128, 73–82, doi: 10.1016/j.exer.2014.09.007 [118]. Copyright Elsevier-used with permission.
The human lens is the only transparent, avascular, and nerveless organ in the body [119], and Ca++ ion concentrations, which are 10,000-fold reduced across the lens plasma membrane compared to the surrounding aqueous humor, have long been known to be a key factor in lens physiology and pathophysiology, largely serving as secondary messengers to regulate numerous essential cellular processes[120]. Receptor tyrosine kinases (RTK), and G-protein-coupled receptors are the cell plasma membrane receptors that detect extracellular signals and modulate free Ca++, thus controlling cell processes such as gene expression, cell proliferation, apoptosis, and cell death [120]. Gap junctions between lens cells serve as channels for the flow of metabolites, small molecules, and ions in the avascular tissue [121]. Mutations in the connexin proteins making up theses junctions disrupts the calcium ion flux, and in the extreme, leads to intracellular calcium precipitation and cataract formation [121].
Tyros3, Axl, and Mertk are RTK’s that make up the TAM family of tyrosine kinase receptors that have Gas6 and PS, two VKDPs as their ligands, with important function in the nervous system [122], as well as the retina as demonstrated by Hall et al. [88]. Valverde et al. 2004 [85] have demonstrated Axl expression in rat and bovine lens epithelial cells, as well as in human lens epithelial. They also demonstrated the presence of Gas6 in bovine aqueous humor. Using a human lens epithelial cell (HLEC) culture model, they show that Gas6 through Axl activation, supports lens epithelial cell growth, survival, and homeostasis. Interestingly, Carnes et al. 2018, report elevated Gas6 mRNA expression in human ciliary bodies, compared to human trabecular meshwork and corneal tissue, suggesting that this important VKDP ligand, may be exported into the aqueous humor, in part to support lens physiology [123]. PS has been found free in aqueous humor in the 7-8 ng/ml range and varies in disease states such as diabetes[124], indicating an additional potential role for VK in homeostatic mechanisms related to diabetic cataracts.
3.3. Schnyder corneal dystrophy
It is undeniable that the eye, and particularly the cornea, have played a pivotal role in discovering both the existence, and function, of fat soluble vitamins, beginning with vitamin A, and its role in xerophthalmia and keratomalacia[125,126], and extending to the essential role of vitamin A in night blindness and visual function[127]. Some of the earliest references to night blindness being treated with vitamin A rich liver appear in ancient Egypt and Greece, with the bulk of the scientific work with regard to vitamin A being completed in the late 19th and early 20th Century [127].
In the 21st Century, the cornea is providing a window into the physiology of VK with respect to the visual system, and more broadly, into complex intracellular molecular interactions in general[128]. This work is largely the result of the completion of the Human Genome Project[129], and genomic advances driving basic and translational research, resulting in an enhanced ability to identify and study genetic variations underlying diseases [130–132].
UBIAD1 is the gene responsible for Schnyder corneal dystrophy (SCD), a rare autosomal dominant disorder that manifests as a progressive bilateral corneal opacification due to widespread deposition of intercellular and extracellular cholesterol and phospholipids in the epithelium, Bowman’s layer, and stroma of the cornea[133–135]. It is variably associated as well with systemic abnormalities in serum lipids, lipoproteins, cholesterol levels, and most recently, higher serum immature dp-ucMGP, a mark of VK deficiency[83].
The modern era of vitamin K research with respect to the eye, can be said to have begun in July 1987 with the referral of a 56-year-old “Swede-Finn” women to Jane S. Weiss, M.D., at the University of Massachusetts Medical Center, for consultation regarding her cloudy corneas[136]. She was diagnosed with what was known at the time as Schnyder’s crystalline corneal dystrophy (SCCD), and within 18 months, two other individuals, possibly related to the same family, were also identified. A “unique opportunity to study a large number of patients with Schnyder’s crystalline dystrophy” was identified, and in what was otherwise considered a rare disease, a systematic effort was launched in January 1989 to do so [137]. Between 1990 and 1992 a series of papers were published detailing the clinical and laboratory findings, epidemiology, genetics, lipid profile, histology, histochemistry, and ultrastructure, of one hundred seventy-three living members of four families, all of whom originated from a 100-k2 area in southwest Finland near the Bay of Bothnia[134,136,138,139].
In 2007, this work culminated in the independent identification by three groups employing various techniques, of the UbiA prenyltranferase domain-containing protein 1 (UBIAD1) gene on chromosome 1p36, as the cause of the disease [140–142]. However, the function of UBIAD1, and its relationship to corneal cholesterol deposition remained unknown.
Using the human genome database to screen for relevant prenylation enzymes, Nakagawa et al. 2010, demonstrated that UBIAD1 was a novel ubiquitously expressed human enzyme with activity in most tissues, responsible for the biosynthesis of MK4 within cells[143]. UBIAD1 was shown to have enzymatic activity to both cleave the prenyl side chains from PK to generate MD, and also to possess prenylation activity to convert MD to MK4 in proportion to the concentration of the prenyl side group, geranylgeranyl pyrophosphate (GGPP)[143]. The authors concluded that both MK4 and UBIAD1 were likely physiologically essential cellular factors, and that the MK4 biosynthesis by UBIAD1 may be related to the pathophysiology of SCD, though the mechanism was unknown[143]. In 2013, Nickerson et al, demonstrated MK4 synthesis was significantly reduced by 22 to 39% in individuals with three different UBIAD1 mutations compared to wt UBIAD1, and that UBIAD1 interacts with the enzymes that catalyze cholesterol synthesis and storage, 3-hydroxy-3methylglutaryl-CoA reductase (HMGCR), and sterol O-acyltransferase (SOAT1)[144]. This suggested to the authors that impaired MK4 synthesis is the biomolecular defect in SDC patients, and that UBIAD1 links VK and cholesterol metabolism, with endogenous MK4 playing a role in sustaining corneal health and visual acuity [145].
In 2015, DeBose-Boyd et al. began to explore how UBIAD1 may contribute to excess cholesterol accumulation in patients with SCD and the novel link between synthesis of MK4 and cholesterol. This has resulted in the discovery that SCD UBIAD1 mutants are sequestered in the endoplasmic reticulum (ER) and bind to HMGCR, which is the rate limiting enzyme for cholesterol synthesis, preventing its normal endoplasmic reticulum-associated degradation (ERAD) and leading to an increased and dysregulated cholesterol and nonsterol isoprenoid production[128,146–151]. This work, stimulated in part by SCD, has led to a much deeper understanding of the regulation of the mevalonate pathway with implication not just for cholesterol metabolism, but also for a number of other essential molecules including steroid hormones, vitamins D and K, bile acids, Heme A, ubiquinone, and other nonsterol isoprenoid, as well as in cardiovascular diseases and cancer[128].
Dong et al. 2018 [152], reported the creation of a CRISPR/Cas 9 based Ubiand1 mutant mouse with a N100s mis-sense mutation which corresponds to the most common N102S mutation reported in humans, and noted that mitochondrial dysfunction is a prominent component of the disease, and suggested that this could result in dysregulated cholesterol synthesis and abnormal accumulation of cholesterol in SCD corneas, but found no differences in corneal cholesterol levels, and the absence of cholesterol crystals among the control and mutant genotypes mice. Thus, the authors conclude that the implications of VK deficiency may be of particular importance in the cornea, stemming from unique conditions present there, and possible differences between humans and mice with respect to cholesterol metabolism[152].
Sarosiak et al. in 2018 [153,154] began reporting important clinical and molecular findings on four previously unpublished Polish families with SCD that were followed prospectively over at least seven years, one of which represented a novel UBIAD1 variant, UBIAD1 p.Thr120Arg. In a 2021 follow-up study designed to explore the corneal and vascular VK status in SCD patients by focusing on matrix MGP, Sarosiak et al. [83] reported for the first time, “quite unexpected and remarkable findings” related to VKDP expression in human corneas, along with differential perturbations by SCD UBIAD1 variants, of VK metabolic pathways.
They showed that MGP is found abundantly in its fully activated form in SCD and normal control corneas, noting that this was quite unexpected given the known reduction in MK4 synthesis associated with UBIAD1 pathologic variants. They document high levels of post-translationally modified phosphorylated and carboxylated, active mature MGP (pcMGP), throughout the corneal epithelium. This pcMGP was localized in both the cytoplasm, and nuclear compartments, which was stratified and most highly concentrated in the epithelial basal layers of both SCD and normal corneas. They further hypothesize, as others have done [56], that external and internal corneal specific factors such as mechanical forces, inflammation, and pH, may influence calcium levels differentially, and thus explain the stratification of the MGP. They note however, that there have been no reported calcium deposits in SCD corneas, and that they found no calcium staining in these cases when specifically searched for. They concede that given the multifunctional nature of other VKDPs such as PS, and Gas6 [155,156], MGP may have some yet unknown specific function associated with corneal intracellular calcium homeostasis, as suggested by Proudfoot et al. [157]
MGP was also found in the corneal stromal keratocytes with no detection in the extracellular matrix. Using light microscopy, no immature ucMGP and dpMGP was detected within the corneal epithelium in either SCD or normal controls. The BGLAP gene encoding the VKDP osteocalcin was not found in SCD patients or controls, and no calcium deposits were detected.
UBIAD1 variants were found to differentially increase MGP mRNA by 12.31-fold in p.Asn112Asp patients and 3.25-fold in p.Asp102Ser. GGCX mRNA was increased 2.38 and 1.48-fold respectively, while Vitamin K Epoxide Reductase Complex Subunit 1 (VKORC1) mRNA was increased 1.90-fold in p.Asn112Asp UBIAD1, yet in the p.Asp102Ser UBIAD1 patient, no difference in VKORC1 levels were seen compared to controls. Thus, UBIAD1 variants were shown to influence VK related gene expression and do so uniquely. In this case, MGP mRNA differed by nearly a factor of 4x. The authors speculate these VK metabolism related genes are up regulated to compensate for reduced GGCX carboxylation of MGP due to lowered MK4 cofactor generation, though acknowledge the mechanism remains unclear, and how this might relate to SCD phenotypic expression is unknown.
Lending support to these findings and possibly supplying corroboratory data, Chen et al. [158], report a HEK293 cell-based model in which they selectively insert and test UBIAD1 mutant variants and independently measure MK4 biosynthesis and VKD protein carboxylation. Comparing p.Asn112Asp to p.Asp102Ser variants, they found an approximate 4.7x difference in VKD carboxylation which is in line with the nearly 4x MGP mRNA induction differences produced by these variants found by Sarosiak et al., thus showing some proportionality in the observed variant effects. Chen et al., commenting on their findings, note that SCD patients do not in general exhibit typical phenotypic expressions associated with defects in MK4 or VKDP under carboxylation, and that their findings suggest that UBIAD1’s MK4 biosynthetic activity does not correlate directly with SCD patient corneal phenotypic expressions.
Sarosiak et al. also measured serum dp-ucMGP, a biomarker of functional vitamin K status, in SCD patients, unaffected family members, and unrelated controls, and found for the first time, that the levels were significantly higher in SCD individuals, reflective of a systemic low vitamin K activity.
The authors conclude VK metabolic machinery is active in SCD and normal corneas, and that MGP appears to have a vital role in maintaining corneal health and to possibly protect against corneal calcification. Sadly, despite these major advances, they acknowledge as others have, that the “molecular mechanism by which UBIAD1 pathogenic variants affect the cornea leading to lipid deposition in SCD patients has yet to be determined”[154].
3.4. Retinal disease
Unlike glaucoma, cataracts, and SCD, specific research with regard to VK and retinal disease are more limited, though this is likely to change quite rapidly with recent discoveries with regard to VK and ferroptosis [26,159] addressed below. Age related macular degeneration (AMD) is the most common cause of irreversible blindness, accounting for 8.7% of global cases [160], with an estimated prevalence of 196 million in 2020, increasing to 288 million by 2040 [161]. While AMD represents the largest well-phenotyped cohort of people who have provided dietary intake data examining the relationship of diet and nutrition with visual function [162,163],the association with VK has not been addressed, and these lack of findings regarding VK and the retina are not surprising, given that a search of the 433,444 trials listed in U.S. National Library of Medicine Clinical Trials database returned 83 trials with the search terms “vitamin” and “retina” [164], and zero with respect to “vitamin K” and “retina”[165]. A few human and basic research studies are however available.
Wei et al. 2018 [98,166] reported in a large prospective longitudinal study that a doubling of serum inactive VKDP dp-ucMGP, an inverse serum marker of systemic VK status [167], was associated with a 1.40 μm narrowing of retinal arteriolar diameter, and concluded that their findings underscore the pivotal role of activated MGP in ocular homeostasis, and that VK supplementation may promote retinal health [98]. The observations were further thought to be clinically relevant because smaller retinal arterioles and lower arteriole-to-venule diameters predict cardiovascular mortality [168], coronary heart disease [169], and may be reflective of similar changes in the cerebral microvasculature[170], thus linking MGP activity in the eye to the broader topic of VK in aging and age-associated diseases [171–173]. Borrás et al likewise confirmed the expression MGP in the mouse retinal vasculature, including capillaries and pericytes [81,82].
Years earlier, beginning in 1990, Canfield[174] and others at Manchester University, began to report on the spontaneous transition of bovine retinal microvascular pericyte cells (BMPC) into multicellular nodules that produced hydroxyapatite needle-like crystals (deposition of calcium phosphate salts in the form of hydroxyapatite is the hallmark of vascular calcification [175]) , and ultimately differentiated along osteogenic lines. In 1998 [176], they reported for the first time, that microvascular retinal pericytes have osteogenic potential in vitro and in vivo, and are able to differentiate into bone, cartilage, adipose, and fibrous connective tissue-concluding that pericytes may give rise to cells of multiple lineages and that microvascular pericytes may be playing an important role in calcification of vascular tissue. In 2000[79],they showed that the VKDP MGP had a key role in regulating the differentiation of these multipotent cells as well in the calcification process itself.
Three years later in 2003 [84], they were the first to show that the VKDP Gas6 and its ligan Axl, were highly expressed in cultured BMPC, and suggested Gas6/Axl signaling was important in the regulation of retinal physiology and pathophysiology. Then again in 2007 [177], the group was the first to show in cultured vascular smooth muscle cells (VSMC) from bovine aortas that Gas6/Axl signaling inhibited mineral deposition by VSMCs, and thus is potentially an important regulator of vascular calcification.
The work of Canfield et al. demonstrates the important and broad-based involvement of vitamin K with retinal vasculature and provides a strong mechanistic rational to explain the findings of Wei et al. [98,166] with respect to the association of dietary vitamin K intake, dp-ucMGP, and reduced retinal arteriolar diameters. So as not to be remiss, it is important to point out as Sweeney et al. 2016 [178], and Simó et al. 2018 [179]have shown in excellent reviews, that the microvascular retinal pericytes[179], as well as pericytes of the brain and central nervous system[178] do much more than serve to support the vascular system, but have a critical role at the center of the neurovascular unit-which is made up of the vascular cells, glial cells, and the neurons. Thus, these findings link VK through MGP, and Gas6, not only to the retinal vasculature but also to the neuroretina as well.
Regulation of the phagocytic function of the retinal pigment epithelium (RPE) is critical to photoreceptor function and thus to vision, with disruption of RPE phagocytosis leading to diseases such as retinitis pigmentosa, and AMD [120]. Two VKDPs, Gas6 and PS are critical ligands in this Mer receptor mediated process [122] . Beginning in 2001, Hall et al. [86–88] demonstrated for the first time that both Gas6, along with the coagulation related molecule PS, had a specific function in the eye. It was shown that Gas6 and PS require VD dependent gamma-carboxylation before linking shedding photoreceptor (PR) outer segments (OS) in a Ca++ mediated manner, to the RPE Mer receptors for phagocytosis. Seven percent of the photoreceptor mass is said to turn over daily, making the RPE the most active, and among the most specialized phagocytes in the human body [180,181]. Both Gas6 and PS are widely expressed in the nervous system and are involved in the regulation of phagocytosis in several important processes [87], in addition to their function with respect to the RPE. After having demonstrated common and redundant roles of Gas6 and PS for the first time in any biological process, Hall et al. emphasize the critical role of VK and VKDPs in retinal OS phagocytosis, and thus global retinal function [88]
4. Non-canonical Vitamin K functions and ferroptosis
In what may prove to be the most significant advance with respect to VK and EVS in the last decade, a non-VKDP vitamin K cycle has been shown to be a potent suppressor of a programmed cell death process known as ferroptosis [26,182].
VK’s ability to act as an antioxidant has been reported in the past [183,184], though it has long been held[19,45],that the only firmly established physiologic function of VK is its action as a cofactor for γ-carboxyglutamyl carboxylase (GGCX), to effect the posttranslational modification of VKDPs. Recent discoveries however, with respect to VK and its relationship to the ferroptosis suppressor protein 1 (FSP1)[26,159], may warrant reevaluations of this view.
330 billion human cells are estimated to turnover daily (4 million per second)[185], likely the result of numerously evolved programs of regulated cell death (RCD) to control cancer[186], and promote cooperation for survival of the organism[187]. Corneal epithelial cells for example are estimated to turnover every seven days [188], yet uniquely, and by way of contrast, the adult lens nucleus is comprised of cells generated in utero at approximately the sixth week of gestation[189], with no turnover of the cells or their membrane lipids [190,191], and are present for a lifetime. This may well be the result in part to adaptations of human lens cell membranes to contain high levels of sphingolipids (for which VK has been shown to have a direct role in synthesis [192]) which are more resistant to oxidation and might confer unique chemical and physical stability to the lens fiber cells [191,193].
Our understanding of the orchestration of cellular turnover, and how cells live and die together [194] has undergone a profound transformation over the past one hundred or so years, as the mechanisms of RCD (death programs controlled by dedicated molecular machinery and thus potentially modifiable through environmental, nutritional, pharmacologic, or genetic inputs, among others [195,196]), as opposed to accidental cell death (ACD-sudden and catastrophic death of cells as a result of exposure to severe physical, chemical, or mechanical insults resulting in the disassembly of the cells plasma membrane[195]), have been discovered and elucidated. Cell death once taken for granted as an inevitable and natural consequence of cell life, was first recognized as an essential function in directing metamorphosis of reptiles in the mid 19th century, and was further appreciated in the early 20th century as a genetically determined process in insects and amphibians[194].
Ferroptosis, was first described by Dixon and Stockwell et al. in 2012 [197] as a nonapoptotic RCD dependent on intracellular iron and which was genetically, biochemically, and morphologically distinct from apoptosis, necrosis, and autophagy. In a 2022 comprehensive review, Stockwell [198] defined ferroptosis as a form of cell death characterized by iron-dependent accumulation of lethal membrane-localized lipid peroxides, that sits at the intersection of reactive oxygen species (ROS) biology, metabolism, and iron regulation, with highly relevant biologic and therapeutic significance [198].
In 2019, Doll et al. [199] and Bersuker et al.[200], independently discovered that reduced ubiquinol (CoQ10), a related lipoquinone to VK menaquinones[201], mediated the suppression of ferroptosis in conjunction with its NADH dependent oxidoreductive enzyme, FSP1. The myristoylation of FSP1 was shown to recruit the enzyme to plasma membranes, where it serves to reduce CoQ10 [200], which then acts as a lipophilic radical-trapping antioxidant (RTA) to stop the production of lipid peroxides, thus demonstrating the relay NADH/FSP1/CoQ10 as a powerful suppressor of ferroptosis[199].
In a major advance, both with respect to VK and to ferroptosis research, Mishima et al 2022 [26], extending their earlier work[199], discovered that the reduced form of VK (VKH2), acting also as a potent RTA to inhibit phospholipid peroxidation, possessed powerful anti-ferroptotic activity as well, and importantly that FSP1 was the warfarin insensitive VK reductase sustaining the reaction[26]. They thus established that FSP1 acts to reduce both CoQ10 and VK to RTA’s and that FSP1 is the long sought after antidotal enzyme overcoming warfarin poisoning[202–204]. Independently, Jin et al. 2022 [159], using genome-wide CRISPR-Cas9 knockout screening, likewise demonstrated FSP1 to be the elusive warfarin-resistant vitamin K reductase responsible for the reduction of VK, establishing reduced VK not only as a cofactor for VK dependent carboxylation, but also as a scavenger of phospholipid radicals to suppress ferroptosis[159].
The implications of these findings with respect to VK and the EVS are significant. Zhang et al. 2022[205] provide a comprehensive review of ferroptotic biology and its involvement in major ocular diseases, and conclude that the effects of ferroptosis cannot be underestimated, deeming ferroptosis-related cell death to be a critical emerging field of study [205]. Yang et al. 2022 [206] provide evidence in human primary RPE cells and in mice that the FSP1-CoQ10/VK-NADH pathways inhibit retinal ferroptotic pathology and may have a potential therapeutic role, providing supporting biochemical, histologic, and electroretinogram data [206]. Likewise, Wei et al. 2021[207] provide experimental evidence that human lens epithelial cells (LEC) and mouse lens epithelium are highly susceptible ferroptosis, and that cataractous and aging human lenses may exhibit more hallmarks of ferroptosis than any other human organ [207].
Though researchers are rightfully expressing caution and the need for clinical trials and focused research [205], these and related findings, have lead Hirschhorn and Stockwell, 2022 [182] to conclude that VK status may serve as a biomarker for ferroptotic sensitivity and propose that VK supplementation may be able to reduce symptoms of neurodegenerative and other ferroptosis related conditions [182], which by extension likely include ocular and visual processing pathologies.
5. Vitamin K, higher cortical visual processing function, and ocular correlates
Dietary VK intake, variations in gut MK isoform biosynthesis, and MK-4 concentrations in the brain, are increasingly associated with cognitive function and neurodegenerative histopathology [47,50,208–212]. Though the domain of memory is often associated with, and evaluated as a measure of cognition[213], it is important to recognize that visual perception, processing, and hallucinations are higher cortical functions as well, and can be used as diagnostic criteria in neurologic degeneration [214].
Though memory tends to dominate cognitive neurology, it should be noted that visual function can be selectively impaired, and prominent cortical visual dysfunction, with relative memory sparing, can occur in various neurodegenerative conditions [215,216]. Increasingly, measurements beyond simple visual acuity assessment, such as perception of biologic motion in mild cognitive impairment [217,218], as well as color vision and formed hallucinations in dementia with Lewy bodies (DLB) [219,220], are being shown to be possible biomarkers of early disease. There is also a growing awareness of possible cross talk between decreased visual acuity and cognitive decline as well [221].
The retina likewise, by virtue of its common embryogenesis, as well as anatomic and physiologic similarities to the brain, provides a unique “window” to observe correlates and potential consequences or brain pathology from the earliest to advanced stages, with increasing sensitivity, noninvasiveness, precision, and speed; largely the result of advances in optical coherence tomography (OCT)[170,222,223].
Thus, not only does an increasing understanding of VK biology have the potential to support and preserve vision, by addressing cataracts and retinal pathology for example, it also has the potential to have similar effects on cognition, and thus higher cortical visual processing-likely providing synergistic and compounding benefits to patients on multiple fronts. Targeting basic and transitional VK ocular research, as well as incorporating ocular and visual data and biomarkers in neurodegenerative research more broadly, can have significant impact, and should be considered a high priority given recent advances in the field[10].
6. Discussion
The eye plays a central role in the health and wellbeing of humanity and has long contributed significantly to unlocking important scientific discoveries [130,132].Vision and its loss has a particular immanency in the human psyche [1], and visual processing occupies a large percentage of the neocortex in terms of neural connections [37] and energy expenditure [224,225], largely a result of the high cost of moving ions across neuronal membranes, and the fact that the neuronal systems are a constant energy sink regardless of whether they are active or at rest [226]. Vision loss is an increasing problem worldwide as populations age, with significant economic and social impact [8] despite numerous advances in medical and surgical interventions [6]; therefore, it is imperative to identify and address the nutritional factors associated with maintaining and optimizing visual function [10,11]. Recent advances in VK research seem poised to provide much needed actionable insights.
It is well established that the fat-soluble vitamins (FSV) A, D, and E play essential roles, and in some cases can have toxic effects, in the eye and visual system [227–229]. Vitamin A in the form of 11-cis-retinal is indeed the singular light sensitive molecule setting in motion the visual cycle resulting in the transduction of photons into nervous impulses[228], and its deficiency has been heralded by night blindness and xerophthalmia for millennia-the direct result of which lead to its discovery as the first FSV[125]. Vitamins D [227] and E [229], likewise have been shown to have pleotropic benefits as well as rare toxicity for the EVS, with toxic levels of A, D, and E, unlike VK, able to accumulate within the body [230].
In contrast, VK tissue concentrations in the mouse eye and human brains are in the pmol/g range [39,46,47,50] , several orders of magnitude less than its fellow FSV [19,48], and the human total body pool of VK is estimated to be as low as 17-194 μg (0.28-2.17 μg/kg), with a mean and (SD) of 87.6 μg (60), and a total body pool turnover of approximately 1.5 days. This is a unique feature in the realm of FSV biology and an important point to grasp when approaching problems and developing solutions related to this nutrient. These small concentrations and rapid tissue loss, lead directly to the discovery of VK in chicks, who are uniquely sensitive to the withdrawal of VK from the diet, showing clinical evidence of deficiency within hours. Similar vulnerabilities are seen in human infants, resulting in nearly universal worldwide parenteral administration of VK shortly after birth to prevent vitamin K deficiency bleeding (VKDB) of early infancy[231], which is sometimes associated with vision loss. Unique also to VK is the fact no known toxicity to naturally occurring VK isoforms have been shown, and natural forms of VK “appear to be essentially innocuous”[232]. Noteworthy also with respect to its fellow FSV, is the presence of naturally occurring VK antagonists in the form of fungal conversion of plant derived coumarins into dicoumarol, the prototypic anticoagulant [233].
Ample evidence now exists of the presence and importance of VK in the eye and visual system, primarily in the form of the menaquinone MK4, and to a lesser extent PK in the eye and human brain. The VKDP mouse gene Mgp has been shown to be both responsible and sufficient to control physiologic mouse intraocular pressure [80]. Human brain MK4 concentrations are now known to be inversely associated with neurodegenerative histologic pathology and antemortem cognitive function loss [50], and a clinical trial has demonstrated a beneficial association of increased daily dietary VK intake and a reduced risk of cataract surgery [15]. Yet, there is a profound lack of human research and clinical trials with respect to VK and the EVS, which represents a significant opportunity.
To date, there appears to be no published data regarding the concentrations and tissue distribution of VK in the human eye. Neither have age, gender, genetic, nutritional, microbiome, environmental, or regional factors affecting human ocular VK biology, been reported in any detail.
Despite the prominent importance vision plays in the human experience, and growing evidence of VK involvement in the EVS, not a single trial with respect to VK and the eye has been registered in the U.S. National Institutes of Health ClinicalTrial.gov database, and only one trial has been reported regarding VK and cataracts in PubMed [15]. The AREDS and AREDS2 studies, the largest and most well characterized trials related to macular degeneration, analyzed 44 nutrients, identifying 9, which were significantly (P< 0.0005) associated with decreased risk, and 3 with increased risk, of various forms of AMD[163]. Associations with VK were not reported however, though it is hoped that a secondary analysis of the existing data may provide insights regarding VK and AMD.
Several factors may explain this absence of clinical trials. First, there has been a general lack of awareness and appreciation for the potential of broad-based pleiotrophic VK action in the eye. The dramatic and immediate clinical consequences of VK deficiency, and the utility of using VKA to treat diseases of blood clotting, have dominated its nearly 100 year history [18].This lead directly to the establishment early on of VK requirements based solely on the minimal amounts of PK needed to provide the liver with adequate PK to support functioning clotting factors [233]. Though PK and MK forms of the vitamin were discovered roughly in tandem, the abundance and importance of PK from plants in the human diet, and the pressing need to prevent bleeding disorders, may have initially eclipsed any imperatives to explore extrahepatic requirements for VK. Ames and coworkers have advanced the “triage theory” [18,234] to highlight the tendency to focus on the life-threatening aspects of micronutrient deficiencies, at the expense of exploring the long-term debilitating consequences of critical nutrient inadequacy on age-related diseases, as exemplified, for example, by cataracts, glaucoma, and AMD.
Secondly, a lack of critical biomarkers, broad-based nutritional databases with respect to both PK and MK, and a comprehensive accounting of VK content and flux in the eye and brain, have all conspired to greatly hamper research interest and efforts. Further, the assessment of VK status is made more difficult by the fact that serum VK levels have large interindividual variability, especially compared to other FSV, showing significant age, gender, racial and ethnic differences not solely related to intake, that remain largely unaccounted for [235–237]. Not to mention the fact, that as many as 25% of participants in studies may have PK serum levels <0.1 nmol/L (the current lower limit of detection)[237].
Finally, funding issues also have significant impact on VK ocular and visual system research. Though the categories are not mutually exclusive, The National Institutes of Health 2023 funding estimates for the category of Eye Diseases and Disorders of Vision are $1,120 million, which ranks 54th in a list of 309 Research/Disease areas, contrasting with Alzheimer’s and Alzheimer’s Disease Related Dementias at $ 3,324 million ranking 24th [238]. Likewise, the 2022 funding levels for the National Eye Institute was one-third of that of the National Institute of Neurological Disorders And Stroke for example, at $833,012 million vs. $2,503,517 million [239]
Despite a recognition that nutrition is the most powerful intervention to reduce disease across the life span, and that correction of micronutrient deficiencies yields substantial returns on investment [10], vitamin research in general, and VK research specifically, has lagged. Chambers et al. reported in 2017 that U.S. federal funding for vitamin research declined between 1992-2015 as a percentage of total federal research grant spending [13], from 0.6% to 0.2%, or to $95 million (2016 US dollars). Between the years 2000-2015, VK research had the fewest average projects funded at 8/yr., compared to 115/yr. for vitamin A, with VK garnering the lowest average yearly project value of $2.4 million, versus $34 million for vitamin D, both of which represent a 14:1 ratio in favor of other FSV. Increasing these funding levels, especially considering recent findings, has the potential to deliver needed breakthroughs to advance EVS nutritional recommendations and health.
Great strides have been made in the last 25 years to increase our understanding of VK biology, particularly in the areas leveraging the Human Genome Project, CRISPR/Cas9 technology, radioisotope studies regarding PK and MKn, and the discovery of UBIAD1, to name a few. Most recently the role of VK in ferroptosis, is a major advance which is likely to rapidly drive VR research in the EVS for years into the future, and possible serve as a much need biomarker of VK status[182]. So also is the prospectively gathered data that increased dietary VK is associated with a decreased risk of cataract surgery, and that higher MK4 levels in decedent brains are inversely associated with a broad range of neurohistopatholgy, and better overall cognitive function, all of which strengthen the nutritional epidemiologic evidence necessary to justify VK interventional trials [240,241].
7. Clinical considerations
Clinical considerations with respect to VK and the EVS for those seeking consultation with an eye care professional might be reasonably drawn from the human studies cited. First to consider is, the most recent analysis of the National Health and Nutrition Examination Survey (NHANES) data indicates that VK intake is declining and greater than half of adults over 70 years of age are not meeting minimal dietary recommendations, which are 90 μg/d for females and 120 μg/d males, [242], and secondly, there are no known toxicities associated with VK intake in healthy individuals [232,243]. Therefore, it would seem reasonable to encourage most patients to increase their intake of green leafy vegetables unless there are potential medical contraindications such as the use of a VKA (warfarin for example) or other dietary limitations. Patient with glaucoma, and those wishing to postpone cataract surgery as long as possible, should be encouraged, if they are able, to consume a cup of nutrient dense green leafy vegetables such as kale, or broccoli, as well as foods rich in MKn daily, as even modest increases in intakes have been shown in observational studies [101,102,103,104] to significantly reduce the risk of glaucoma progression. Similarly, the difference between one-half and one cup of green leafy vegetables a day in the PREDIMED study was associated with a 29 percent risk reduction of cataract surgery over less than a six time year period[15]. Patients with neurodegenerative diseases such as mild cognitive impairment, Lewy body dementia, AD, or Parkinson disease, should be asked about subtle visual hallucinations, or assessed for disruption of color vision, and in the absence of contraindications, encouraged with the same dietary recommendations as those with glaucoma and cataract concerns. Longitudinal ocular imaging such as OCT and fundus photos, as well as markers of higher cortical visual processing, should be considered in an effort to promote detection of neurodegeneration in the earliest possible stages, and to inform clinical decision making and therapeutic interventions, as well as to promote interdisciplinary and coordinated patient care decision making and research[244,245].
Health care professionals should be encouraged to recognize the pervasive nature of nutritional and micronutrient deficiency and its impact on multiple individual and global health and economic indices, particularly for certain vulnerable groups such as the economically disadvantaged, pregnant women, young children, and the elderly [18,246]. Widespread deficiencies are particularly acute for the FSV A,D,E,K [246,247],with half of adults over 70 years not meeting VK adequate intake recommendations in US NHANES surveys as noted above [243].
8. Conclusions
VK has been shown to be unique among it fellow VSF with respect to its relatively small tissue concentrations, limited and precarious total body stores, and lack of any known significant toxicity. Evidence has been presented from basic and clinical sciences that VK is found in the eye and brain, that MGP a VKDP, is responsible for controlling intraocular pressure, and that for the first time, prospectively obtained VK levels in human brains have been associated with histologic neuropathology and a slower decline in cognitive function, while increased dietary intake of VK has been demonstrated prospectively, to be associated with a significant reduction in the risk of cataract surgery.
Taken together, the relevance of VK biology with respect to the eye and visual system can confidently be asserted. The virtual lack of human VK tissue concentration and distribution data, along with no firm understanding of the trafficking of VK to and within the eye, reveals a significant gap in our knowledge, as well as a tremendous opportunity-inviting scientists, clinicians, funding partners, advocacy groups, along with patients, to rise to the occasion and shape a healthier, clearer, and brighter future.
Author Contributions
M.A.M. Conceptualized the review design, searched, and reviewed the literature, selected referenced papers, wrote the manuscript, and directed table and figure design. Author has read and agreed to the published version of the manuscript.
Funding
This article received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
Dedication
Dr. Stuart Paul Richer, OD. PhD. FAAO. Mentor, friend, and visionary.
References
- Scott, A.W.; Bressler, N.M.; Ffolkes, S.; Wittenborn, J.S.; Jorkasky, J. Public Attitudes About Eye and Vision Health. JAMA Ophthalmol. 2016, 134, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.R. The Eye and Its Diseases in Ancient Egypt. Acta Ophthalmol. Scand. 1997, 75, 338–344. [Google Scholar] [CrossRef] [PubMed]
- The Writings of Hippocrates and Galen | Online Library of Liberty Available online:. Available online: https://oll.libertyfund.org/title/coxe-the-writings-of-hippocrates-and-galen (accessed on 3 October 2021).
- Papadopoulos, G.; Liarmakopoulou, A.; Zisoulis, E.; Tzimas, P.; Lena, A.; Kitsos, G. Treatment of Eye Diseases in the Hippocratic Era. Hell. J. Surg. 2018, 90, 143–145. [Google Scholar] [CrossRef]
- GBD 2019 Blindness and Vision Impairment, C. Trends in Prevalence of Blindness and Distance and near Vision Impairment over 30 Years: An Analysis for the Global Burden of Disease Study. Lancet Glob. Health 2021, 9, e130. [Google Scholar] [CrossRef]
- Keel, S.; Cieza, A. Rising to the Challenge: Estimates of the Magnitude and Causes of Vision Impairment and Blindness. Lancet Glob. Health 2021, 9, e100. [Google Scholar] [CrossRef] [PubMed]
- Flaxman, A.D.; Wittenborn, J.S.; Robalik, T.; Gulia, R.; Gerzoff, R.B.; Lundeen, E.A.; Saaddine, J.; Rein, D.B. ; Vision and Eye Health Surveillance System study group Prevalence of Visual Acuity Loss or Blindness in the US: A Bayesian Meta-Analysis. JAMA Ophthalmol. 2021, 139, 717–723. [Google Scholar] [CrossRef]
- Rein, D.B.; Wittenborn, J.S.; Zhang, P.; Sublett, F.; Lamuda, P.A.; Lundeen, E.A.; Saaddine, J. The Economic Burden of Vision Loss and Blindness in the United States. Ophthalmology, 2021; S0161-6420(21)00710-7. [Google Scholar] [CrossRef]
- Assi, L.; Chamseddine, F.; Ibrahim, P.; Sabbagh, H.; Rosman, L.; Congdon, N.; Evans, J.; Ramke, J.; Kuper, H.; Burton, M.J.; et al. A Global Assessment of Eye Health and Quality of Life: A Systematic Review of Systematic Reviews. JAMA Ophthalmol. 2021, 139, 526–541. [Google Scholar] [CrossRef]
- Bailey, R.L.; Jr, K.P.W.; Black, R.E. The Epidemiology of Global Micronutrient Deficiencies. Ann. Nutr. Metab. 2015, 66, 22–33. [Google Scholar] [CrossRef]
- Harshman, S.G.; Shea, M.K. The Role of Vitamin K in Chronic Aging Diseases: Inflammation, Cardiovascular Disease, and Osteoarthritis. Curr. Nutr. Rep. 2016, 5, 90. [Google Scholar] [CrossRef]
- Roberts, S.B.; Silver, R.E.; Das, S.K.; Fielding, R.A.; Gilhooly, C.H.; Jacques, P.F.; Kelly, J.M.; Mason, J.B.; McKeown, N.M.; Reardon, M.A.; et al. Healthy Aging—Nutrition Matters: Start Early and Screen Often. Adv. Nutr. 2021, 12, 1438–1448. [Google Scholar] [CrossRef]
- Chambers, J.D.; Anderson, J.E.; Salem, M.N.; Bügel, S.G.; Fenech, M.; Mason, J.B.; Weber, P.; West, K.P.; Wilde, P.; Eggersdorfer, M.; et al. The Decline in Vitamin Research Funding: A Missed Opportunity? Curr. Dev. Nutr. 2017, 1, e000430. [Google Scholar] [CrossRef] [PubMed]
- Search of: VITAMIN | Eye Diseases - Search Details - ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results/details?cond=Eye+Diseases&term=VITAMIN&cntry=&state=&city=&dist=&Search=Search (accessed on 15 January 2022).
- Camacho-Barcia, M.L.; Bulló, M.; Garcia-Gavilán, J.F.; Ruiz-Canela, M.; Corella, D.; Estruch, R.; Fitó, M.; García-Layana, A.; Arós, F.; Fiol, M.; et al. Association of Dietary Vitamin K1 Intake With the Incidence of Cataract Surgery in an Adult Mediterranean Population: A Secondary Analysis of a Randomized Clinical Trial. JAMA Ophthalmol. 2017, 135, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.R. LXIII Edward Jackson Memorial Lecture: Eye Care: Dollars and Sense. Am. J. Ophthalmol. 2007, 143, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Andreatta, W.; Boukouvala, S.; Bansal, A. Combined Acute Haemolytic and Secondary Angle Closure Glaucoma Following Spontaneous Intraocular Haemorrhages in a Patient on Warfarin. Case Rep. Ophthalmol. 2016, 7, 233–238. [Google Scholar] [CrossRef] [PubMed]
- McCann, J.C.; Ames, B.N. Vitamin K, an Example of Triage Theory: Is Micronutrient Inadequacy Linked to Diseases of Aging? Am. J. Clin. Nutr. 2009, 90, 889–907. [Google Scholar] [CrossRef] [PubMed]
- Shearer, M.J.; Newman, P. Recent Trends in the Metabolism and Cell Biology of Vitamin K with Special Reference to Vitamin K Cycling and MK-4 Biosynthesis. J. Lipid Res. 2014, 55, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Olson, R.E.; Chao, J.; Graham, D.; Bates, M.W.; Lewis, J.H. Total Body Phylloquinone and Its Turnover in Human Subjects at Two Levels of Vitamin K Intake. Br. J. Nutr. 2002, 87, 543–553. [Google Scholar] [CrossRef]
- Suttie, J.W. VITAMIN K in Health and Disease, 1st ed.; CRC Press Kindle edition, 2009; ISBN 978-0-429-11978-1. [Google Scholar]
- Ivanova, D.; Zhelev, Z.; Getsov, P.; Nikolova, B.; Aoki, I.; Higashi, T.; Bakalova, R. Vitamin K: Redox-Modulation, Prevention of Mitochondrial Dysfunction and Anticancer Effect. Redox Biol. 2018, 16, 352–358. [Google Scholar] [CrossRef]
- Nowicka, B.; Kruk, J. Occurrence, Biosynthesis and Function of Isoprenoid Quinones. Biochim. Biophys. Acta BBA - Bioenerg. 2010, 1797, 1587–1605. [Google Scholar] [CrossRef]
- Shearer, M.J.; Okano, T. Key Pathways and Regulators of Vitamin K Function and Intermediary Metabolism. Annu. Rev. Nutr. 2018, 38, 127–151. [Google Scholar] [CrossRef]
- Berkner, K.L.; Runge, K.W. Vitamin K-Dependent Protein Activation: Normal Gamma-Glutamyl Carboxylation and Disruption in Disease. Int. J. Mol. Sci. 2022, 23, 5759. [Google Scholar] [CrossRef]
- Mishima, E.; Ito, J.; Wu, Z.; Nakamura, T.; Wahida, A.; Doll, S.; Tonnus, W.; Nepachalovich, P.; Eggenhofer, E.; Aldrovandi, M.; et al. A Non-Canonical Vitamin K Cycle Is a Potent Ferroptosis Suppressor. Nature 2022. [Google Scholar] [CrossRef] [PubMed]
- Tabb, M.M.; Sun, A.; Zhou, C.; Grün, F.; Errandi, J.; Romero, K.; Pham, H.; Inoue, S.; Mallick, S.; Lin, M.; et al. Vitamin K2 Regulation of Bone Homeostasis Is Mediated by the Steroid and Xenobiotic Receptor SXR *. J. Biol. Chem. 2003, 278, 43919–43927. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, J.; Wang, Y.; Liu, Z.; Wu, Y. Ferroptosis Drives Photoreceptor Degeneration in Mice with Defects in All-Trans-Retinal Clearance. J. Biol. Chem. 2021, 296, 100187. [Google Scholar] [CrossRef] [PubMed]
- Stenflo, J.; Fernlund, P.; Egan, W.; Roepstorff, P. Vitamin K Dependent Modifications of Glutamic Acid Residues in Prothrombin. Proc. Natl. Acad. Sci. USA 1974, 71, 2730–2733. [Google Scholar] [CrossRef]
- Suttie, J.W. Synthesis of Vitamin K-Dependent Proteins. FASEB J. 1993, 7, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Ferland, G. The Vitamin K-Dependent Proteins: An Update. Nutr. Rev. 1998, 56, 223–230. [Google Scholar] [CrossRef]
- H, X.; J, C.; L, D.; S, L. Role of Emerging Vitamin K-dependent Proteins: Growth Arrest-specific Protein 6, Gla-rich Protein and Periostin (Review). Int. J. Mol. Med. 2021, 47. [Google Scholar] [CrossRef]
- Carafoli, E.; Krebs, J. Why Calcium? How Calcium Became the Best Communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef]
- Beato, S.; Toledo-Solís, F.J.; Fernández, I. Vitamin K in Vertebrates’ Reproduction: Further Puzzling Pieces of Evidence from Teleost Fish Species. Biomolecules 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Berkner, K.L.; Runge, K.W. Vitamin K-Dependent Protein Activation: Normal Gamma-Glutamyl Carboxylation and Disruption in Disease. Int. J. Mol. Sci. 2022, 23, 5759. [Google Scholar] [CrossRef]
- Felleman, D.J.; Van Essen, D.C. Distributed Hierarchical Processing in the Primate Cerebral Cortex. Cereb. Cortex N. Y. N 1991 1991, 1, 1–47. [Google Scholar] [CrossRef]
- Moini, J.; Piran, P. Chapter 14 - Visual System. In Functional and Clinical Neuroanatomy; Moini, J., Piran, P., Eds.; Academic Press, 2020; pp. 417–466 ISBN 978-0-12-817424-1.
- Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of Phylloquinone (Vitamin K1) into Menaquinone-4 (Vitamin K2) in Mice: Two Possible Routes for Menaquinone-4 Accumulation in Cerebra of Mice. J. Biol. Chem. 2008, 283, 11270–11279. [Google Scholar] [CrossRef] [PubMed]
- Carrié, I.; Portoukalian, J.; Vicaretti, R.; Rochford, J.; Potvin, S.; Ferland, G. Menaquinone-4 Concentration Is Correlated with Sphingolipid Concentrations in Rat Brain. J. Nutr. 2004, 134, 167–172. [Google Scholar] [CrossRef]
- Hirota, Y.; Tsugawa, N.; Nakagawa, K.; Suhara, Y.; Tanaka, K.; Uchino, Y.; Takeuchi, A.; Sawada, N.; Kamao, M.; Wada, A.; et al. Menadione (Vitamin K3) Is a Catabolic Product of Oral Phylloquinone (Vitamin K1) in the Intestine and a Circulating Precursor of Tissue Menaquinone-4 (Vitamin K2) in Rats. J. Biol. Chem. 2013, 288, 33071. [Google Scholar] [CrossRef]
- Thijssen, H.H.W.; Vervoort, L.M.T.; Schurgers, L.J.; Shearer, M.J. Menadione Is a Metabolite of Oral Vitamin K. Br. J. Nutr. 2006, 95, 260–266. [Google Scholar] [CrossRef]
- Pardridge, W.M. CNS Drug Design Based on Principles of Blood-Brain Barrier Transport. J. Neurochem. 1998, 70, 1781–1792. [Google Scholar] [CrossRef] [PubMed]
- Vita, M.F.; Nagachar, N.; Avramidis, D.; Delwar, Z.M.; Cruz, M.H.; Siden, Å.; Paulsson, K.M.; Yakisich, J.S. Pankiller Effect of Prolonged Exposure to Menadione on Glioma Cells: Potentiation by Vitamin C. Invest. New Drugs 2011, 29, 1314–1320. [Google Scholar] [CrossRef]
- Ellis, J.L.; Fu, X.; Karl, J.P.; Hernandez, C.J.; Mason, J.B.; DeBose-Boyd, R.A.; Booth, S.L. Multiple Dietary Vitamin K Forms Are Converted to Tissue Menaquinone-4 in Mice. J. Nutr. 2021. [Google Scholar] [CrossRef]
- Thijssen, H.H.; Drittij-Reijnders, M.J. Vitamin K Status in Human Tissues: Tissue-Specific Accumulation of Phylloquinone and Menaquinone-4. Br. J. Nutr. 1996, 75, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Tanprasertsuk, J.; Ferland, G.; Johnson, M.A.; Poon, L.W.; Scott, T.M.; Barbey, A.K.; Barger, K.; Wang, X.-D.; Johnson, E.J. Concentrations of Circulating Phylloquinone, but Not Cerebral Menaquinone-4, Are Positively Correlated with a Wide Range of Cognitive Measures: Exploratory Findings in Centenarians. J. Nutr. 2020, 150, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Tanprasertsuk, J.; Scott, T.M.; Barbey, A.K.; Barger, K.; Wang, X.-D.; Johnson, M.A.; Poon, L.W.; Vishwanathan, R.; Matthan, N.R.; Lichtenstein, A.H.; et al. Carotenoid-Rich Brain Nutrient Pattern Is Positively Correlated With Higher Cognition and Lower Depression in the Oldest Old With No Dementia. Front. Nutr. 2021, 8, 704691. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Shea, M.K.; Dolnikowski, G.G.; Patterson, W.B.; Dawson-Hughes, B.; Holland, T.M.; Schneider, J.A.; Booth, S.L. Vitamin D and Vitamin K Concentrations in Human Brain Tissue Are Influenced by Freezer Storage Time: The Memory and Aging Project. J. Nutr. 2021, 151, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.L.; Shea, M.K.; Barger, K.; Leurgans, S.E.; James, B.D.; Holland, T.M.; Agarwal, P.; Fu, X.; Wang, J.; Matuszek, G.; et al. Association of Vitamin K with Cognitive Decline and Neuropathology in Community-dwelling Older Persons. Alzheimers Dement. Transl. Res. Clin. Interv. 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Arnott, J.A.; Planey, S.L. The Influence of Lipophilicity in Drug Discovery and Design. Expert Opin. Drug Discov. 2012, 7, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.L.; Fu, X.; Karl, J.P.; Hernandez, C.J.; Mason, J.B.; DeBose-Boyd, R.A.; Booth, S.L. Multiple Dietary Vitamin K Forms Are Converted to Tissue Menaquinone-4 in Mice. J. Nutr. [CrossRef] [PubMed]
- Ji, Y.; Li, X.; Tso, P. Intestinal Fatty Acid Absorption. Immunol. Endocr. Metab. Agents Med. Chem. 2009, 9, 60–73. [Google Scholar] [CrossRef]
- Shearer, M.J.; Fu, X.; Booth, S.L. Vitamin K Nutrition, Metabolism, and Requirements: Current Concepts and Future Research. Adv. Nutr. 2012, 3, 182. [Google Scholar] [CrossRef]
- Lai, Y.; Masatoshi, H.; Ma, Y.; Guo, Y.; Zhang, B. Role of Vitamin K in Intestinal Health. Front. Immunol. 2022, 12, 791565. [Google Scholar] [CrossRef]
- Lamon-Fava, S.; Sadowski, J.A.; Davidson, K.W.; O’Brien, M.E.; McNamara, J.R.; Schaefer, E.J. Plasma Lipoproteins as Carriers of Phylloquinone (Vitamin K1) in Humans. Am. J. Clin. Nutr. 1998, 67, 1226–1231. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Vermeer, C. Differential Lipoprotein Transport Pathways of K-Vitamins in Healthy Subjects. Biochim. Biophys. Acta 2002, 1570, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Jahn, C.E.; Leiss, O.; von Bergmann, K. Lipid Composition of Human Aqueous Humor. Ophthalmic Res. 1983, 15, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Cenedella, R.J. Lipoproteins and Lipids in Cow and Human Aqueous Humor. Biochim. Biophys. Acta 1984, 793, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.R.; Price, M.O.; Price, F.W.; Pardo, J.C.; Grandin, J.C.; You, J.; Wang, M.; Yoder, M.C. Proteomic Analysis of Human Aqueous Humor Using Multidimensional Protein Identification Technology. Mol. Vis. 2009, 15, 2740. [Google Scholar] [PubMed]
- Kodeboyina, S.K.; Lee, T.J.; Churchwell, L.; Ulrich, L.; Bollinger, K.; Bogorad, D.; Estes, A.; Zhi, W.; Sharma, S.; Sharma, A. The Constitutive Proteome of Human Aqueous Humor and Race Specific Alterations. Proteomes 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Connor, W.E.; Duell, P.B.; Kean, R.; Wang, Y. The Prime Role of HDL to Transport Lutein into the Retina: Evidence from HDL-Deficient WHAM Chicks Having a Mutant ABCA1 Transporter. Invest. Ophthalmol. Vis. Sci. 2007, 48, 4226–4231. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Vachali, P.; Chang, F.-Y.; Gorusupudi, A.; Arunkumar, R.; Shi, L.; Rognon, G.T.; Frederick, J.M.; Bernstein, P.S. HDL Is the Primary Transporter for Carotenoids from Liver to Retinal Pigment Epithelium in Transgenic ApoA-I-/-/Bco2-/- Mice. Arch. Biochem. Biophys. 2022, 716, 109111. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Takada, T.; Kurauchi, R.; Tanaka, Y.; Komine, T.; Suzuki, H. Transporters for the Intestinal Absorption of Cholesterol, Vitamin E, and Vitamin K. J. Atheroscler. Thromb. 2017, 24, 347–359. [Google Scholar] [CrossRef]
- Storti, F.; Raphael, G.; Griesser, V.; Klee, K.; Drawnel, F.; Willburger, C.; Scholz, R.; Langmann, T.; von Eckardstein, A.; Fingerle, J.; et al. Regulated Efflux of Photoreceptor Outer Segment-Derived Cholesterol by Human RPE Cells. Exp. Eye Res. 2017, 165, 65–77. [Google Scholar] [CrossRef]
- Tserentsoodol, N.; Gordiyenko, N.V.; Pascual, I.; Lee, J.W.; Fliesler, S.J.; Rodriguez, I.R. Intraretinal Lipid Transport Is Dependent on High Density Lipoprotein-like Particles and Class B Scavenger Receptors. Mol. Vis. 2006, 12, 1319–1333. [Google Scholar] [PubMed]
- Gharahkhani, P.; Burdon, K.P.; Fogarty, R.; Sharma, S.; Hewitt, A.W.; Martin, S.; Law, M.H.; Cremin, K.; Bailey, J.N.C.; Loomis, S.J.; et al. Common Variants near ABCA1, AFAP1 and GMDS Confer Risk of Primary Open-Angle Glaucoma. Nat. Genet. 2014, 46, 1120–1125. [Google Scholar] [CrossRef]
- Nelsestuen, G.L.; Zytkovicz, T.H.; Howard, J.B. The Mode of Action of Vitamin K: IDENTIFICATION OF γ-CARBOXYGLUTAMIC ACID AS A COMPONENT OF PROTHROMBIN. J. Biol. Chem. 1974, 249, 6347–6350. [Google Scholar] [CrossRef]
- Ferland, G. Vitamin K and the Nervous System: An Overview of Its Actions. Adv. Nutr. 2012, 3, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Tissue Expression of GGCX - Summary - The Human Protein Atlas. Available online: https://www.proteinatlas.org/ENSG00000115486-GGCX/tissue (accessed on 30 October 2022).
- Berkner, K.L.; Pudota, B.N. Vitamin K-Dependent Carboxylation of the Carboxylase. Proc. Natl. Acad. Sci. USA 1998, 95, 466. [Google Scholar] [CrossRef] [PubMed]
- Annis, D.S.; Ma, H.; Balas, D.M.; Kumfer, K.T.; Sandbo, N.; Potts, G.K.; Coon, J.J.; Mosher, D.F. Absence of Vitamin K-Dependent γ-Carboxylation in Human Periostin Extracted from Fibrotic Lung or Secreted from a Cell Line Engineered to Optimize γ-Carboxylation. PLoS ONE 2015, 10, e0135374. [Google Scholar] [CrossRef]
- Demirören, K.; Yavuz, H.; Cam, L. Intracranial Hemorrhage Due to Vitamin K Deficiency after the Newborn Period. Pediatr. Hematol. Oncol. 2004, 21, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Bui Quoc, E.; Bonnet, D.; Bajolle, F. [Vitamin K antagonist overdose induced blindness in an infant: An argument for a therapeutic educational program]. Arch. Pediatr. Organe Off. Soc. Francaise Pediatr. 2012, 19, 22–26. [Google Scholar] [CrossRef]
- Holden, R. Spontaneous Hyphaema as a Result of Systemic Anticoagulation in Previously Abnormal Eyes. Postgrad. Med. J. 1991, 67, 1008. [Google Scholar] [CrossRef]
- Masri, I.; Smith, J.; Wride, N.; Ghosh, S. A Rare Case of Acute Angle Closure Due to Spontaneous Suprachoroidal Haemorrhage Secondary to Loss of Anti-Coagulation Control: A Case Report. BMC Ophthalmol. 2018, 18. [Google Scholar] [CrossRef]
- Gonzalez, P.; Zigler, J.S.; Epstein, D.L.; Borrás, T. Identification and Isolation of Differentially Expressed Genes from Very Small Tissue Samples. BioTechniques 1999, 26, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, P.; Epstein, D.L.; Borrás, T. Characterization of Gene Expression in Human Trabecular Meshwork Using Single-Pass Sequencing of 1060 Clones. Invest. Ophthalmol. Vis. Sci. 2000, 41, 3678–3693. [Google Scholar] [PubMed]
- Canfield, A.E.; Doherty, M.J.; Kelly, V.; Newman, B.; Farrington, C.; Grant, M.E.; Boot-Handford, R.P. Matrix Gla Protein Is Differentially Expressed During the Deposition of a Calcified Matrix by Vascular Pericytes. FEBS Lett. 2000, 487, 267–271. [Google Scholar] [CrossRef]
- Borrás, T.; Cowley, D.O.; Asokan, P.; Pandya, K. Generation of a Matrix Gla (Mgp) Floxed Mouse, Followed by Conditional Knockout, Uncovers a New Mgp Function in the Eye. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Borrás, T.; Smith, M.H.; Buie, L.K. A Novel Mgp-Cre Knock-in Mouse Reveals an Anticalcification/Antistiffness Candidate Gene in the Trabecular Meshwork and Peripapillary Scleral Region. Invest. Ophthalmol. Vis. Sci. 2015, 56, 2203. [Google Scholar] [CrossRef] [PubMed]
- Asokan, P.; Mitra, R.N.; Periasamy, R.; Han, Z.; Borrás, T. A Naturally Fluorescent Mgp Transgenic Mouse for Angiogenesis and Glaucoma Longitudinal Studies. Invest. Ophthalmol. Vis. Sci. 2018, 59, 746. [Google Scholar] [CrossRef] [PubMed]
- Sarosiak, A.; Oziębło, D.; Udziela, M.; Vermeer, C.; Malejczyk, J.; Szaflik, J.P.; Ołdak, M. High Expression of Matrix Gla Protein in Schnyder Corneal Dystrophy Patients Points to an Active Role of Vitamin K in Corneal Health. Acta Ophthalmol. (Copenh.) 2021, 99, e171–e177. [Google Scholar] [CrossRef] [PubMed]
- Collett, G.; Wood, A.; Alexander, M.Y.; Varnum, B.C.; Boot-Handford, R.P.; Ohanian, V.; Ohanian, J.; Fridell, Y.-W.; Canfield, A.E. Receptor Tyrosine Kinase Axl Modulates the Osteogenic Differentiation of Pericytes. Circ. Res. 2003, 92, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Valverde, P.; Obin, M.S.; Taylor, A. Role of Gas6/Axl Signaling in Lens Epithelial Cell Proliferation and Survival. Exp. Eye Res. 2004, 78, 27–37. [Google Scholar] [CrossRef]
- Hall, M.O.; Prieto, A.L.; Obin, M.S.; Abrams, T.A.; Burgess, B.L.; Heeb, M.J.; Agnew, B.J. Outer Segment Phagocytosis by Cultured Retinal Pigment Epithelial Cells Requires Gas6. Exp. Eye Res. 2001, 73, 509–520. [Google Scholar] [CrossRef]
- Hall, M.O.; Obin, M.S.; Prieto, A.L.; Burgess, B.L.; Abrams, T.A. Gas6 Binding to Photoreceptor Outer Segments Requires Gamma-Carboxyglutamic Acid (Gla) and Ca(2+) and Is Required for OS Phagocytosis by RPE Cells in Vitro. Exp. Eye Res. 2002, 75, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.O.; Obin, M.S.; Heeb, M.J.; Burgess, B.L.; Abrams, T.A. Both Protein S and Gas6 Stimulate Outer Segment Phagocytosis by Cultured Rat Retinal Pigment Epithelial Cells. Exp. Eye Res. 2005, 81, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Warejcka, D.J.; Olague-Marchan, M.; Twining, S.S. Corneal Activation of Prothrombin to Form Thrombin, Independent of Vascular Injury. Invest. Ophthalmol. Vis. Sci. 2007, 48, 134–143. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Blindness and Vision Impairment, C. Causes of Blindness and Vision Impairment in 2020 and Trends over 30 Years, and Prevalence of Avoidable Blindness in Relation to VISION 2020: The Right to Sight: An Analysis for the Global Burden of Disease Study. Lancet Glob. Health 2021, 9, e144. [Google Scholar] [CrossRef]
- Casson, R.J.; Chidlow, G.; Wood, J.P.; Crowston, J.G.; Goldberg, I. Definition of Glaucoma: Clinical and Experimental Concepts. Clin. Experiment. Ophthalmol. 2012, 40, 341–349. [Google Scholar] [CrossRef]
- Kass, M.A.; Heuer, D.K.; Higginbotham, E.J.; Johnson, C.A.; Keltner, J.L.; Miller, J.P.; Parrish, R.K.; Wilson, M.R.; Gordon, M.O. The Ocular Hypertension Treatment Study: A Randomized Trial Determines That Topical Ocular Hypotensive Medication Delays or Prevents the Onset of Primary Open-Angle Glaucoma. Arch. Ophthalmol. Chic. Ill 1960 2002, 120, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Heijl, A.; Leske, M.C.; Bengtsson, B.; Hyman, L.; Bengtsson, B.; Hussein, M. ; Early Manifest Glaucoma Trial Group Reduction of Intraocular Pressure and Glaucoma Progression: Results from the Early Manifest Glaucoma Trial. Arch. Ophthalmol. Chic. Ill 1960 2002, 120, 1268–1279. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M. “What Controls Aqueous Humour Outflow Resistance? ” Exp. Eye Res. 2006, 82, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Acott, T.S.; Kelley, M.J. Extracellular Matrix in the Trabecular Meshwork. Exp. Eye Res. 2008, 86, 543. [Google Scholar] [CrossRef]
- Vranka, J.A.; Kelley, M.J.; Acott, T.S.; Keller, K.E. Extracellular Matrix in the Trabecular Meshwork: Intraocular Pressure Regulation and Dysregulation in Glaucoma. Exp. Eye Res. 2015, 133, 112–125. [Google Scholar] [CrossRef]
- Xue, W.; Wallin, R.; Olmsted-Davis, E.A.; Borrás, T. Matrix Gla Protein Function in Human Trabecular Meshwork Cells: Inhibition of Bmp2-Induced Calcification Process. Invest. Ophthalmol. Vis. Sci. 2006, 47, 997–1007. [Google Scholar] [CrossRef]
- Wei, F.-F.; Huang, Q.-F.; Zhang, Z.-Y.; Van Keer, K.; Thijs, L.; Trenson, S.; Yang, W.-Y.; Cauwenberghs, N.; Mujaj, B.; Kuznetsova, T.; et al. Inactive Matrix Gla Protein Is a Novel Circulating Biomarker Predicting Retinal Arteriolar Narrowing in Humans. Sci. Rep. 2018, 8, 15088. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Yao, K.; Peng, F.; Zhao, B.; Chen, Z.; Chen, W.; Zhao, Y.; Zhang, H.; Wang, J. The Effect of Dietary Vitamin K1 Supplementation on Trabecular Meshwork and Retina in a Chronic Ocular Hypertensive Rat Model. Invest. Ophthalmol. Vis. Sci. 2020, 61, 40. [Google Scholar] [CrossRef] [PubMed]
- Ramdas, W.D.; Schouten, J.S.A.G.; Webers, C.A.B. The Effect of Vitamins on Glaucoma: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 359. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.L.; Stone, K.L.; Kodjebacheva, G.; Yu, F.; Pedula, K.L.; Ensrud, K.E.; Cauley, J.A.; Hochberg, M.C.; Topouzis, F.; Badala, F.; et al. Glaucoma Risk and the Consumption of Fruits and Vegetables Among Older Women in the Study of Osteoporotic Fractures. Am. J. Ophthalmol. 2008, 145, 1081–1089. [Google Scholar] [CrossRef]
- Moïse, M.M.; Benjamin, L.-M.; Doris, T.M.; Dalida, K.N.; Augustin, N.O. Role of Mediterranean Diet, Tropical Vegetables Rich in Antioxidants, and Sunlight Exposure in Blindness, Cataract and Glaucoma among African Type 2 Diabetics. Int. J. Ophthalmol. 2012, 5, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Giaconi, J.A.; Yu, F.; Stone, K.L.; Pedula, K.L.; Ensrud, K.E.; Cauley, J.A.; Hochberg, M.C.; Coleman, A.L. The Association of Consumption of Fruits/Vegetables With Decreased Risk of Glaucoma Among Older African-American Women in the Study of Osteoporotic Fractures. Am. J. Ophthalmol. 2012, 154, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Willett, W.C.; Rosner, B.A.; Buys, E.; Wiggs, J.L.; Pasquale, L.R. Association of Dietary Nitrate Intake With Primary Open-Angle Glaucoma: A Prospective Analysis From the Nurses’ Health Study and Health Professionals Follow-up Study. JAMA Ophthalmol. 2016, 134, 294–303. [Google Scholar] [CrossRef]
- Lam, D.; Rao, S.K.; Ratra, V.; Liu, Y.; Mitchell, P.; King, J.; Tassignon, M.-J.; Jonas, J.; Pang, C.P.; Chang, D.F. Cataract. Nat. Rev. Dis. Primer 2015, 1, 1–15. [Google Scholar] [CrossRef]
- Hashemi, H.; Pakzad, R.; Yekta, A.; Aghamirsalim, M.; Pakbin, M.; Ramin, S.; Khabazkhoob, M. Global and Regional Prevalence of Age-Related Cataract: A Comprehensive Systematic Review and Meta-Analysis. Eye 2020, 34, 1357. [Google Scholar] [CrossRef]
- Hejtmancik, J.F.; Shiels, A. Overview of the Lens. Prog. Mol. Biol. Transl. Sci. 2015, 134, 119. [Google Scholar] [CrossRef]
- Gillies, M.; Brian, G.; La Nauze, J.; Le Mesurier, R.; Moran, D.; Taylor, H.; Ruit, S. Modern Surgery for Global Cataract Blindness: Preliminary Considerations. Arch. Ophthalmol. Chic. Ill 1960 1998, 116, 90–92. [Google Scholar] [CrossRef]
- Heruye, S.H.; Maffofou Nkenyi, L.N.; Singh, N.U.; Yalzadeh, D.; Ngele, K.K.; Njie-Mbye, Y.-F.; Ohia, S.E.; Opere, C.A. Current Trends in the Pharmacotherapy of Cataracts. Pharmaceuticals 2020, 13, 15. [Google Scholar] [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef] [PubMed]
- Turck, D.; Bresson, J.-L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Dietary Reference Values for Vitamin K. EFSA J. 2017, 15, e04780. [Google Scholar] [CrossRef]
- López-Sobaler, A.M.; Aparicio, A.; Aranceta-Bartrina, J.; Gil, Á.; González-Gross, M.; Serra-Majem, L.; Varela-Moreiras, G.; Ortega, R.M. Overweight and General and Abdominal Obesity in a Representative Sample of Spanish Adults: Findings from the ANIBES Study. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef]
- Online Food Calculator. Food Weight to Volume Conversions. Available online: https://www.aqua-calc.com/calculate/food-weight-to-volume (accessed on 12 January 2022).
- Shea, M.K.; Berkner, K.L.; Ferland, G.; Fu, X.; Holden, R.M.; Booth, S.L. Perspective: Evidence before Enthusiasm—A Critical Review of the Potential Cardiovascular Benefits of Vitamin K. Adv. Nutr. 2021, 12, 632–646. [Google Scholar] [CrossRef]
- García-Layana, A.; Ciufo, G.; Toledo, E.; Martínez-González, M.; Corella, D.; Fitó, M.; Estruch, R.; Gómez-Gracia, E.; Fiol, M.; Lapetra, J.; et al. The Effect of a Mediterranean Diet on the Incidence of Cataract Surgery. Nutrients 2017, 9. [Google Scholar] [CrossRef]
- Sai Varsha, M.K.N.; Raman, T.; Manikandan, R. Inhibition of Diabetic-Cataract by Vitamin K1 Involves Modulation of Hyperglycemia-Induced Alterations to Lens Calcium Homeostasis. Exp. Eye Res. 2014, 128, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Varsha, M.K.N.S.; Thiagarajan, R.; Manikandan, R.; Dhanasekaran, G. Vitamin K1 Alleviates Streptozotocin-Induced Type 1 Diabetes by Mitigating Free Radical Stress, as Well as Inhibiting NF-ΚB Activation and INOS Expression in Rat Pancreas. Nutrition 2015, 31, 214–222. [Google Scholar] [CrossRef]
- Thiagarajan, R.; Varsha, M.K.N.S.; Srinivasan, V.; Ravichandran, R.; Saraboji, K. Vitamin K1 Prevents Diabetic Cataract by Inhibiting Lens Aldose Reductase 2 (ALR2) Activity. Sci. Rep. 2019, 9, 14684. [Google Scholar] [CrossRef]
- Wang, Z.; Su, D.; Liu, S.; Zheng, G.; Zhang, G.; Cui, T.; Ma, X.; Sun, Z.; Hu, S. RNA Sequencing and Bioinformatics Analysis of Human Lens Epithelial Cells in Age-Related Cataract. BMC Ophthalmol. 2021, 21. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.D.; Sanderson, J. The Mechanisms of Calcium Homeostasis and Signalling in the Lens. Exp. Eye Res. 2009, 88, 226–234. [Google Scholar] [CrossRef]
- Berthoud, V.M.; Gao, J.; Minogue, P.J.; Jara, O.; Mathias, R.T.; Beyer, E.C. Connexin Mutants Compromise the Lens Circulation and Cause Cataracts through Biomineralization. Int. J. Mol. Sci. 2020, 21, E5822. [Google Scholar] [CrossRef] [PubMed]
- Burstyn-Cohen, T.; Hochberg, A. TAM Signaling in the Nervous System. Brain Plast. Amst. Neth. 2021, 7, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Carnes, M.U.; Allingham, R.R.; Ashley-Koch, A.; Hauser, M.A. Transcriptome Analysis of Adult and Fetal Trabecular Meshwork, Cornea, and Ciliary Body Tissues by RNA Sequencing. Exp. Eye Res. 2018, 167, 91–99. [Google Scholar] [CrossRef]
- Sugimoto, M.; Kondo, M.; Yasuma, T.; D’Alessandro-Gabazza, C.N.; Toda, M.; Imai, H.; Nakamura, M.; Gabazza, E.C. Increased Expression of Protein S in Eyes with Diabetic Retinopathy and Diabetic Macular Edema. Sci. Rep. 2021, 11, 10449. [Google Scholar] [CrossRef]
- Sommer, A. Vitamin A Deficiency and Clinical Disease: An Historical Overview. J. Nutr. 2008, 138, 1835–1839. [Google Scholar] [CrossRef]
- Semba, R.D. On the ‘Discovery’ of Vitamin A. Ann. Nutr. Metab. 2012, 61, 192–198. [Google Scholar] [CrossRef]
- Wolf, G. The Discovery of the Visual Function of Vitamin A. J. Nutr. 2001, 131, 1647–1650. [Google Scholar] [CrossRef]
- Faulkner, R.; Jo, Y. Synthesis, Function, and Regulation of Sterol and Nonsterol Isoprenoids. Front. Mol. Biosci. 2022, 9, 1006822. [Google Scholar] [CrossRef] [PubMed]
- The Human Genome Project Available online:. Available online: https://www.genome.gov/human-genome-project (accessed on 12 November 2022).
- Stahl, A.; Smith, L.E.H. An Eye for Discovery. J. Clin. Invest. 2010, 120, 3008–3011. [Google Scholar] [CrossRef] [PubMed]
- Duggal, P.; Ibay, G.; Klein, A.P. Current Gene Discovery Strategies for Ocular Conditions. Invest. Ophthalmol. Vis. Sci. 2011, 52, 7761–7770. [Google Scholar] [CrossRef] [PubMed]
- Porter, L.; Black, G. Personalized Ophthalmology. Clin. Genet. 2014, 86. [Google Scholar] [CrossRef] [PubMed]
- Garner, A.; Tripathi, R.C. Hereditary Crystalline Stromal Dystrophy of Schnyder. II. Histopathology and Ultrastructure. Br. J. Ophthalmol. 1972, 56, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.S.; Rodrigues, M.M.; Kruth, H.S.; Rajagopalan, S.; Rader, D.J.; Kachadoorian, H. Panstromal Schnyder’s Corneal Dystrophy: Ultrastructural and Histochemical Studies. Ophthalmology 1992, 99, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Kruth, H.S. Accumulation of Unesterified Cholesterol in Limbal Cornea and Conjunctiva of Rabbits Fed a High-Cholesterol Diet. Detection with Filipin. Atherosclerosis 1987, 63, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.S. *Schnyder’s Dystrophy of the Cornea. A Swede-Finn Connection. Cornea 1992, 11, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.S. VISUAL MORBIDITY IN THIRTY-FOUR FAMILIES WITH SCHNYDER CRYSTALLINE CORNEAL DYSTROPHY (AN AMERICAN OPHTHALMOLOGICAL SOCIETY THESIS). Trans. Am. Ophthalmol. Soc. 2007, 105, 616–648. [Google Scholar]
- Weiss, J.S.; Rodrigues, M.M.; Rajagopalan, S.; Kruth, H. Atypical Schnyder’s Crystalline Dystrophy of the Cornea: A Light and Electron Micoscopic Study [Abstract]. Proc Int Soc Eye Res 1990, 6, 198. [Google Scholar]
- Weiss, J.S.; Rodrigues, M.M.; Rajagopalan, S.; Kruth, H. Schnyder’s Corneal Dystrophy: Clinical, Ultrastructural, and Histo-Chemical Studies. Ophthalmology 1990, 97, 141. [Google Scholar]
- Orr, A.; Dubé, M.-P.; Marcadier, J.; Jiang, H.; Federico, A.; George, S.; Seamone, C.; Andrews, D.; Dubord, P.; Holland, S.; et al. Mutations in the UBIAD1 Gene, Encoding a Potential Prenyltransferase, Are Causal for Schnyder Crystalline Corneal Dystrophy. PLoS ONE 2007, 2, e685. [Google Scholar] [CrossRef] [PubMed]
- Yellore, V.S.; Khan, M.A.; Bourla, N.; Rayner, S.A.; Chen, M.C.; Sonmez, B.; Momi, R.S.; Sampat, K.M.; Gorin, M.B.; Aldave, A.J. Identification of Mutations in UBIAD1 Following Exclusion of Coding Mutations in the Chromosome 1p36 Locus for Schnyder Crystalline Corneal Dystrophy. Mol. Vis. 2007, 13, 1777–1782. [Google Scholar] [PubMed]
- Weiss, J.S.; Kruth, H.S.; Kuivaniemi, H.; Tromp, G.; White, P.S.; Winters, R.S.; Lisch, W.; Henn, W.; Denninger, E.; Krause, M.; et al. Mutations in the UBIAD1 Gene on Chromosome Short Arm 1, Region 36, Cause Schnyder Crystalline Corneal Dystrophy. Invest. Ophthalmol. Vis. Sci. 2007, 48, 5007–5012. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a Novel Human Menaquinone-4 Biosynthetic Enzyme. Nature 2010, 468, 117–121. [Google Scholar] [CrossRef]
- Nickerson, M.L.; Bosley, A.D.; Weiss, J.S.; Kostiha, B.N.; Hirota, Y.; Brandt, W.; Esposito, D.; Kinoshita, S.; Wessjohann, L.; Morham, S.G.; et al. The UBIAD1 Prenyltransferase Links Menaquione-4 Synthesis to Cholesterol Metabolic Enzymes. Hum. Mutat. 2013, 34, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, M.L.; Bosley, A.D.; Weiss, J.S.; Kostiha, B.N.; Hirota, Y.; Brandt, W.; Esposito, D.; Kinoshita, S.; Wessjohann, L.; Morham, S.G.; et al. The UBIAD1 Prenyltransferase Links Menaquinone-4 [Corrected] Synthesis to Cholesterol Metabolic Enzymes. Hum. Mutat. 2013, 34, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.M.; Elsabrouty, R.; Seemann, J.; Jo, Y.; DeBose-Boyd, R.A. The Prenyltransferase UBIAD1 Is the Target of Geranylgeraniol in Degradation of HMG CoA Reductase. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.M.; Jun, D.-J.; Jo, Y.; Seemann, J.; DeBose-Boyd, R.A. Geranylgeranyl-Regulated Transport of the Prenyltransferase UBIAD1 between Membranes of the ER and Golgi. J. Lipid Res. 2016, 57, 1286–1299. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.M.; Jun, D.-J.; Johnson, B.M.; DeBose-Boyd, R.A. UbiA Prenyltransferase Domain-Containing Protein-1 Modulates HMG-CoA Reductase Degradation to Coordinate Synthesis of Sterol and Nonsterol Isoprenoids. J. Biol. Chem. 2018, 293, 312–323. [Google Scholar] [CrossRef]
- Jo, Y.; Hamilton, J.S.; Hwang, S.; Garland, K.; Smith, G.A.; Su, S.; Fuentes, I.; Neelam, S.; Thompson, B.M.; McDonald, J.G.; et al. Schnyder Corneal Dystrophy-Associated UBIAD1 Inhibits ER-Associated Degradation of HMG CoA Reductase in Mice. eLife 2019, 8, e44396. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.-Y.; Tang, J.-J.; Xiao, X.; Qi, W.; Wu, S.; Jiang, C.; Hong, J.; Xu, J.; Song, B.-L.; Luo, J. Schnyder Corneal Dystrophy-Associated UBIAD1 Mutations Cause Corneal Cholesterol Accumulation by Stabilizing HMG-CoA Reductase. PLOS Genet. 2019, 15, e1008289. [Google Scholar] [CrossRef] [PubMed]
- Jun, D.-J.; Schumacher, M.M.; Hwang, S.; Kinch, L.N.; Grishin, N.V.; DeBose-Boyd, R.A. Schnyder Corneal Dystrophy-Associated UBIAD1 Is Defective in MK-4 Synthesis and Resists Autophagy-Mediated Degradation. J. Lipid Res. 2020, 61, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Jin, X.; Boettler, M.; Sciulli, H.; Abu-Asab, M.; Greco, C.; Wang, S.; Hu, Y.-C.; Campos, M.; Jackson, S.; et al. A Mouse Model of Schnyder Corneal Dystrophy with the N100S Point Mutation. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Sarosiak, A.; Ołdak, M. Functional Characterization of the UBIAD1 Protein: The Nodal Point for Vitamin K and Cholesterol Synthesis. From Corneal Dystrophies to Lifestyle Diseases. Postepy Hig. Med. Dosw. 2018, 72, 116–130. [Google Scholar] [CrossRef]
- Sarosiak, A.; Udziela, M.; Ścieżyńska, A.; Oziębło, D.; Wawrzynowska, A.; Szaflik, J.P.; Ołdak, M. Clinical Diversity in Patients with Schnyder Corneal Dystrophy-a Novel and Known UBIAD1 Pathogenic Variants. Graefes Arch. Clin. Exp. Ophthalmol. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 2018, 256, 2127–2134. [Google Scholar] [CrossRef]
- Suleiman, L.; Négrier, C.; Boukerche, H. Protein S: A Multifunctional Anticoagulant Vitamin K-Dependent Protein at the Crossroads of Coagulation, Inflammation, Angiogenesis, and Cancer. Crit. Rev. Oncol. Hematol. 2013, 88, 637–654. [Google Scholar] [CrossRef]
- Ireland, L.; Luckett, T.; Schmid, M.C.; Mielgo, A. Blockade of Stromal Gas6 Alters Cancer Cell Plasticity, Activates NK Cells, and Inhibits Pancreatic Cancer Metastasis. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Proudfoot, D.; Shanahan, C.M. Molecular Mechanisms Mediating Vascular Calcification: Role of Matrix Gla Protein (Review Article). Nephrology 2006, 11, 455–461. [Google Scholar] [CrossRef]
- Chen, X.; Furukawa, N.; Jin, D.-Y.; Liu, Y.; Stafford, D.W.; Williams, C.M.; Suhara, Y.; Tie, J.-K. Naturally Occurring UBIAD1 Mutations Differentially Affect Menaquinone Biosynthesis and Vitamin K-Dependent Carboxylation. FEBS J. 2021. [Google Scholar] [CrossRef]
- Jin, D.-Y.; Chen, X.; Liu, Y.; Williams, C.; Pedersen, L.; Stafford, D.; Tie, J.-K. A Genome-Wide CRISPR-Cas9 Knockout Screen Reveals FSP1 as Warfarin-Resistant Vitamin K Reductase 2022.
- Jonas, J.B.; Cheung, C.M.G.; Panda-Jonas, S. Updates on the Epidemiology of Age-Related Macular Degeneration. Asia-Pac. J. Ophthalmol. 2017, 6, 493–497. [Google Scholar] [CrossRef]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.-Y.; Wong, T.Y. Global Prevalence of Age-Related Macular Degeneration and Disease Burden Projection for 2020 and 2040: A Systematic Review and Meta-Analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef] [PubMed]
- Age-Related Eye Disease Study Research Group The Relationship of Dietary Carotenoid and Vitamin A, E, and C Intake With Age-Related Macular Degeneration in a Case-Control Study: AREDS Report No. 22. Arch. Ophthalmol. 2007, 125, 1225–1232. [Google Scholar] [CrossRef]
- Agrón, E.; Mares, J.; Clemons, T.E.; Swaroop, A.; Chew, E.Y.; Keenan, T.D.L. Dietary Nutrient Intake and Progression to Late Age-Related Macular Degeneration in the Age-Related Eye Disease Studies 1 and 2. Ophthalmology 2021, 128, 425–442. [Google Scholar] [CrossRef] [PubMed]
- Search of: Vitamin | Retina - List Results - ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results?cond=retina&term=vitamin&cntry=&state=&city=&dist=&Search=Search (accessed on 14 November 2022).
- Search of: Vitamin k | Retina - List Results - ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/results?cond=retina&term=vitamin+k&cntry=&state=&city=&dist= (accessed on 14 November 2022).
- Wei, F.-F.; Trenson, S.; Verhamme, P.; Vermeer, C.; Staessen, J.A. Vitamin K–Dependent Matrix Gla Protein as Multifaceted Protector of Vascular and Tissue Integrity. Hypertension 2019, 73, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Dalmeijer, G.W.; van der Schouw, Y.T.; Magdeleyns, E.; Ahmed, N.; Vermeer, C.; Beulens, J.W.J. The Effect of Menaquinone-7 Supplementation on Circulating Species of Matrix Gla Protein. Atherosclerosis 2012, 225, 397–402. [Google Scholar] [CrossRef]
- Wong, T.Y.; Klein, R.; Nieto, F.J.; Klein, B.E.K.; Sharrett, A.R.; Meuer, S.M.; Hubbard, L.D.; Tielsch, J.M. Retinal Microvascular Abnormalities and 10-Year Cardiovascular Mortality: A Population-Based Case-Control Study. Ophthalmology 2003, 110, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.Y.; Klein, R.; Sharrett, A.R.; Duncan, B.B.; Couper, D.J.; Tielsch, J.M.; Klein, B.E.K.; Hubbard, L.D. Retinal Arteriolar Narrowing and Risk of Coronary Heart Disease in Men and Women. The Atherosclerosis Risk in Communities Study. JAMA 2002, 287, 1153–1159. [Google Scholar] [CrossRef]
- Cheung, C.Y.; Ikram, M.K.; Chen, C.; Wong, T.Y. Imaging Retina to Study Dementia and Stroke. Prog. Retin. Eye Res. 2017, 57, 89–107. [Google Scholar] [CrossRef]
- Simes, D.C.; Viegas, C.S.B.; Araújo, N.; Marreiros, C. Vitamin K as a Diet Supplement with Impact in Human Health: Current Evidence in Age-Related Diseases. Nutrients 2020, 12, 138. [Google Scholar] [CrossRef]
- Popa, D.-S.; Bigman, G.; Rusu, M.E. The Role of Vitamin K in Humans: Implication in Aging and Age-Associated Diseases. Antioxidants 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Khalil, Z.; Alam, B.; Akbari, A.R.; Sharma, H. The Medical Benefits of Vitamin K2 on Calcium-Related Disorders. Nutrients 2021, 13, 691. [Google Scholar] [CrossRef] [PubMed]
- Schor, A.M.; Allen, T.D.; Canfield, A.E.; Sloan, P.; Schor, S.L. Pericytes Derived from the Retinal Microvasculature Undergo Calcification in Vitro. J. Cell Sci. 1990, 97, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Villa-Bellosta, R. New Insights into Endogenous Mechanisms of Protection against Arterial Calcification. Atherosclerosis 2020, 306, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M.J.; Ashton, B.A.; Walsh, S.; Beresford, J.N.; Grant, M.E.; Canfield, A.E. Vascular Pericytes Express Osteogenic Potential In Vitro and In Vivo. J. Bone Miner. Res. 1998, 13, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Collett, G.D.M.; Sage, A.P.; Kirton, J.P.; Alexander, M.Y.; Gilmore, A.P.; Canfield, A.E. Axl/Phosphatidylinositol 3-Kinase Signaling Inhibits Mineral Deposition by Vascular Smooth Muscle Cells. Circ. Res. 2007, 100, 502–509. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Ayyadurai, S.; Zlokovic, B.V. Pericytes of the Neurovascular Unit: Key Functions and Signaling Pathways. Nat. Neurosci. 2016, 19, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Simó, R.; Stitt, A.W.; Gardner, T.W. Neurodegeneration in Diabetic Retinopathy: Does It Really Matter? Diabetologia 2018, 61, 1902–1912. [Google Scholar] [CrossRef]
- Nandrot, E.F.; Dufour, E.M. Mertk in Daily Retinal Phagocytosis: A History in the Making. In Retinal Degenerative Diseases: Laboratory and Therapeutic Investigations; Anderson, R.E., Hollyfield, J.G., LaVail, M.M., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, 2010; pp. 133–140. ISBN 978-1-4419-1399-9. [Google Scholar]
- Penberthy, K.K.; Lysiak, J.J.; Ravichandran, K.S. Re-Thinking Phagocytes: Clues from the Retina and Testes. Trends Cell Biol. 2018, 28, 317. [Google Scholar] [CrossRef]
- Hirschhorn, T.; Stockwell, B.R. Vitamin K: A New Guardian against Ferroptosis. Mol. Cell 2022, 82, 3760–3762. [Google Scholar] [CrossRef]
- Mukai, K.; Itoh, S.; Morimoto, H. Stopped-Flow Kinetic Study of Vitamin E Regeneration Reaction with Biological Hydroquinones (Reduced Forms of Ubiquinone, Vitamin K, and Tocopherolquinone) in Solution. J. Biol. Chem. 1992, 267, 22277–22281. [Google Scholar] [CrossRef] [PubMed]
- Vervoort, L.; Ronden, J.; Thijssen, H. The Potent Antioxidant Activity of the Vitamin K Cycle in Microsomal Lipid Peroxidation. Biochem. Pharmacol. 1997, 54. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Milo, R. The Distribution of Cellular Turnover in the Human Body. Nat. Med. 2021, 27, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Aktipis, C.A.; Boddy, A.M.; Jansen, G.; Hibner, U.; Hochberg, M.E.; Maley, C.C.; Wilkinson, G.S. Cancer across the Tree of Life: Cooperation and Cheating in Multicellularity. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140219. [Google Scholar] [CrossRef]
- West, S.A.; Griffin, A.S.; Gardner, A. Evolutionary Explanations for Cooperation. Curr. Biol. 2007, 17, R661–R672. [Google Scholar] [CrossRef] [PubMed]
- Hanna, C.; Bicknell, D.S.; O’brien, J.E. Cell Turnover in the Adult Human Eye. Arch. Ophthalmol. 1961, 65, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Barishak, Y.R. Embryology of the Eye and Its Adnexae. Dev. Ophthalmol. 1992, 24, 1–142. [Google Scholar] [PubMed]
- Lynnerup, N.; Kjeldsen, H.; Heegaard, S.; Jacobsen, C.; Heinemeier, J. Radiocarbon Dating of the Human Eye Lens Crystallines Reveal Proteins without Carbon Turnover throughout Life. PLoS ONE 2008, 3, e1529. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.R.; Levchenko, V.A.; Blanksby, S.J.; Mitchell, T.W.; Williams, A.; Truscott, R.J. No Turnover in Lens Lipids for the Entire Human Lifespan. eLife 4 e06003. [CrossRef]
- Sundaram, K.S.; Fan, J.H.; Engelke, J.A.; Foley, A.L.; Suttie, J.W.; Lev, M. Vitamin K Status Influences Brain Sulfatide Metabolism in Young Mice and Rats. J. Nutr. 1996, 126, 2746–2751. [Google Scholar] [CrossRef]
- Borchman, D.; Yappert, M.C.; Afzal, M. Lens Lipids and Maximum Lifespan. Exp. Eye Res. 2004, 79, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, Y.; Steller, H. Live to Die Another Way: Modes of Programmed Cell Death and the Signals Emanating from Dying Cells. Nat. Rev. Mol. Cell Biol. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.; Abrams, J.; Adam, D.; P, A.; Es, A.; L, A.; I, A.; Dw, A.; et al. Molecular Mechanisms of Cell Death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25. [Google Scholar] [CrossRef]
- Mishima, E.; Conrad, M. Nutritional and Metabolic Control of Ferroptosis Available online:. Available online: https://www.annualreviews.org/doi/abs/10.1146/annurev-nutr-062320-114541 (accessed on 7 August 2022).
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Non-Apoptotic Cell Death. Cell 2012, 149, 1060. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.R. Ferroptosis Turns 10: Emerging Mechanisms, Physiological Functions, and Therapeutic Applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef] [PubMed]
- Doll, S.; Freitas, F.P.; Shah, R.; Aldrovandi, M.; da Silva, M.C.; Ingold, I.; Goya Grocin, A.; Xavier da Silva, T.N.; Panzilius, E.; Scheel, C.H.; et al. FSP1 Is a Glutathione-Independent Ferroptosis Suppressor. Nature 2019, 575, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Bersuker, K.; Hendricks, J.M.; Li, Z.; Magtanong, L.; Ford, B.; Tang, P.H.; Roberts, M.A.; Tong, B.; Maimone, T.J.; Zoncu, R.; et al. The CoQ Oxidoreductase FSP1 Acts Parallel to GPX4 to Inhibit Ferroptosis. Nature 2019, 575, 688–692. [Google Scholar] [CrossRef]
- Braasch-Turi, M.M.; Koehn, J.T.; Crans, D.C. Chemistry of Lipoquinones: Properties, Synthesis, and Membrane Location of Ubiquinones, Plastoquinones, and Menaquinones. Int. J. Mol. Sci. 2022, 23, 12856. [Google Scholar] [CrossRef] [PubMed]
- Lowenthal, J.; MacFarlane, J.A. THE NATURE OF THE ANTAGONISM BETWEEN VITAMIN K AND INDIRECT ANTICOAGULANTS. J. Pharmacol. Exp. Ther. 1964, 143, 273–277. [Google Scholar] [PubMed]
- Wallin, R.; Hutson, S. Vitamin K-Dependent Carboxylation. Evidence That at Least Two Microsomal Dehydrogenases Reduce Vitamin K1 to Support Carboxylation. J. Biol. Chem. 1982, 257, 1583–1586. [Google Scholar] [CrossRef]
- Tie, J.-K.; Stafford, D.W. Structural and Functional Insights into Enzymes of the Vitamin K Cycle. J. Thromb. Haemost. JTH 2016, 14, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sheng, S.; Wang, W.; Dai, J.; Zhong, Y.; Ren, J.; Jiang, K.; Li, S.; Bian, X.; Liu, L. Molecular Mechanisms of Iron Mediated Programmed Cell Death and Its Roles in Eye Diseases. Front. Nutr. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Tsui, M.G.; Tsang, J.K.W.; Goit, R.K.; Yao, K.-M.; So, K.-F.; Lam, W.-C.; Lo, A.C.Y. Involvement of FSP1-CoQ10-NADH and GSH-GPx-4 Pathways in Retinal Pigment Epithelium Ferroptosis. Cell Death Dis. 2022, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Hao, C.; Huangfu, J.; Srinivasagan, R.; Zhang, X.; Fan, X. Aging Lens Epithelium Is Susceptible to Ferroptosis. Free Radic. Biol. Med. 2021, 167, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Wang, Y.; Barnes, L.L.; Bennett, D.A.; Dawson-Hughes, B.; Booth, S.L. Nutrients and Bioactives in Green Leafy Vegetables and Cognitive Decline: Prospective Study. Neurology 2018, 90, e214. [Google Scholar] [CrossRef] [PubMed]
- Kiely, A.; Ferland, G.; Ouliass, B.; O’Toole, P.W.; Purtill, H.; O’Connor, E.M. Vitamin K Status and Inflammation Are Associated with Cognition in Older Irish Adults. Nutr. Neurosci. 2020, 23, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Bigman, G.; Rusu, M.E. Low Dietary Intakes of Vitamin K and Leafy Green Vegetables Are Individually Associated With Low Cognitive Functioning in A National Sample of U.S. Older Adults. Curr. Dev. Nutr. 2021, 5, 1306. [Google Scholar] [CrossRef]
- McCann, A.; Jeffery, I.B.; Ouliass, B.; Ferland, G.; Fu, X.; Booth, S.L.; Tran, T.T.T.; O’Toole, P.W.; O’Connor, E.M. Exploratory Analysis of Covariation of Microbiota-Derived Vitamin K and Cognition in Older Adults. Am. J. Clin. Nutr. 2019, 110, 1404–1415. [Google Scholar] [CrossRef] [PubMed]
- Alisi, L.; Cao, R.; De Angelis, C.; Cafolla, A.; Caramia, F.; Cartocci, G.; Librando, A.; Fiorelli, M. The Relationships Between Vitamin K and Cognition: A Review of Current Evidence. Front. Neurol. 2019, 10, 239. [Google Scholar] [CrossRef]
- Wilson, R.; Boyle, P.; Yu, L.; Barnes, L.; Sytsma, J.; As, B.; Bennett, D.; Schneider, J. Temporal Course and Pathologic Basis of Unawareness of Memory Loss in Dementia. Neurology 2015, 85. [Google Scholar] [CrossRef]
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E. Clinical Diagnosis of Alzheimer’s Disease: Report of the NINCDS-ADRDA Work Group under the Auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34. [Google Scholar] [CrossRef]
- Greene, J.D.W. Apraxia, Agnosias, and Higher Visual Function Abnormalities. J. Neurol. Neurosurg. Psychiatry 2005, 76, v25–v34. [Google Scholar] [CrossRef]
- Liu, Y.; Pelak, V.; van Stavern, G.; Moss, H. Higher Cortical Dysfunction Presenting as Visual Symptoms in Neurodegenerative Diseases. Front. Neurol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Garobbio, S.; Pilz, K.S.; Kunchulia, M.; Herzog, M.H. No Common Factor Underlying Decline of Visual Abilities in Mild Cognitive Impairment. Exp. Aging Res. 2022, 1–18. [Google Scholar] [CrossRef]
- Pratt, J.; Radulescu, P.; Guo, R.; Al-Aidroos, N.; Abrams, R. Biological Motion Captures Attention. J. Vis. 2010, 10, 120. [Google Scholar] [CrossRef]
- Harding, A.J.; Broe, G.A.; Halliday, G.M. Visual Hallucinations in Lewy Body Disease Relate to Lewy Bodies in the Temporal Lobe. Brain J. Neurol. 2002, 125, 391–403. [Google Scholar] [CrossRef]
- Matar, E.; Phillips, J.R.; Martens, K.A.E.; Halliday, G.M.; Lewis, S.J.G. Impaired Color Discrimination-A Specific Marker of Hallucinations in Lewy Body Disorders. J. Geriatr. Psychiatry Neurol. 2019, 32, 257–264. [Google Scholar] [CrossRef]
- Fuller-Thomson, E.; Nowaczynski, A.; MacNeil, A. The Association Between Hearing Impairment, Vision Impairment, Dual Sensory Impairment, and Serious Cognitive Impairment: Findings from a Population-Based Study of 5.4 Million Older Adults. J. Alzheimers Dis. Rep. 2022, 6, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Y.; Shi, C.; Shen, M.; Lu, F. Advances in Retina Imaging as Potential Biomarkers for Early Diagnosis of Alzheimer’s Disease. Transl. Neurodegener. 2021, 10, 6. [Google Scholar] [CrossRef]
- Jeevakumar, V.; Sefton, R.; Chan, J.; Gopinath, B.; Liew, G.; Shah, T.M.; Siette, J. Association between Retinal Markers and Cognition in Older Adults: A Systematic Review. BMJ Open 2022, 12, e054657. [Google Scholar] [CrossRef]
- Hofmeijer, J.; van Putten, M.J.A.M. Ischemic Cerebral Damage: An Appraisal of Synaptic Failure. Stroke 2012, 43, 607–615. [Google Scholar] [CrossRef]
- Laughlin, S.B.; de Ruyter van Steveninck, R.R.; Anderson, J.C. The Metabolic Cost of Neural Information. Nat. Neurosci. 1998, 1, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Niven, J.E.; Laughlin, S.B. *Energy Limitation as a Selective Pressure on the Evolution of Sensory Systems. J. Exp. Biol. 2008, 211, 1792–1804. [Google Scholar] [CrossRef]
- Chan, H.; Zhang, X.; Ling, X.; Bui, C.; Wang, Y.; Ip, P.; Chu, W.; Chen, L.; Tham, C.; Yam, J.; et al. Vitamin D and Ocular Diseases: A Systematic Review. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Sajovic, J.; Meglič, A.; Glavač, D.; Markelj, Š.; Hawlina, M.; Fakin, A. The Role of Vitamin A in Retinal Diseases. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Edwards, G.; Olson, C.; Euritt, C.; Koulen, P. Molecular Mechanisms Underlying the Therapeutic Role of Vitamin E in Age-Related Macular Degeneration. Front. Neurosci. 2022, 16. [Google Scholar] [CrossRef]
- Reddy, P.; Jialal, I. Biochemistry, Fat Soluble Vitamins; StatPearls Publishing, 2022. [Google Scholar]
- Clarke, P.; Shearer, M.; Card, D.; Nichols, A.; Ponnusamy, V.; Mahaveer, A.; Voong, K.; Dockery, K.; Holland, N.; Mulla, S.; et al. Exclusively Breastmilk-Fed Preterm Infants Are at High Risk of Developing Subclinical Vitamin K Deficiency despite Intramuscular Prophylaxis at Birth. J. Thromb. Haemost. JTH 2022, 20. [Google Scholar] [CrossRef]
- National Research Council. Vitamin Tolerance of Animals; The National Academies Press.: Washington, DC,, 1987; ISBN 978-0-309-03728-0. [Google Scholar]
- Handbook of Vitamins; Suttie, J. W., Zempleni, J., Gregory III, J.F.G., Stover, P.J., Eds.; 5th ed.; CRC Press: Boca Raton, 2013; ISBN 978-0-429-18955-5. [Google Scholar]
- Ames, B.N. Prolonging Healthy Aging: Longevity Vitamins and Proteins. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 10836. [Google Scholar] [CrossRef]
- Shea, M.K.; Booth, S.L. Concepts and Controversies in Evaluating Vitamin K Status in Population-Based Studies. Nutrients 2016, 8. [Google Scholar] [CrossRef]
- Westerman, K.; Kelly, J.M.; Ordovás, J.M.; Booth, S.L.; DeMeo, D.L. Epigenome-Wide Association Study Reveals a Molecular Signature of Response to Phylloquinone (Vitamin K1) Supplementation. Epigenetics 2020, 15, 859–870. [Google Scholar] [CrossRef]
- Shea, M.; Benjamin, E.; Dupuis, J.; Massaro, J.; Jacques, P.; D’Agostino, R.; Ordovas, J.; O’Donnell, C.; Dawson-Hughes, B.; Vasan, R.; et al. Genetic and Non-Genetic Correlates of Vitamins K and D. Eur. J. Clin. Nutr. 2009, 63. [Google Scholar] [CrossRef] [PubMed]
- Research Portfolio Online Reporting Tools (RePORT) Estimates of NIH Funding for Various Research, Condition, and Disease Categories (RCDC) Available online:. Available online: https://report.nih.gov/funding/categorical-spending#/ (accessed on 22 January 2023).
- NIH Office of Budget Supplemental Tables NIH — Office of Budget —Budget Request FY 2023 Available online:. Available online: https://officeofbudget.od.nih.gov/pdfs/FY23/br/Overview%20of%20FY%202023%20Supplementary%20Tables.pdf (accessed on 27 January 2023).
- Booth, S.L. RePORT-Vitamins D and K and Neuropathologically-Defined Alzheimer and Other Dementias in Older Persons Available online:. Available online: https://reporter.nih.gov/search/kehAwJ-zEE24u-RBTtLAcw/project-details/9193206 (accessed on 27 January 2023).
- Satija, A.; Yu, E.; Willett, W.; Hu, F. Understanding Nutritional Epidemiology and Its Role in Policy. Adv. Nutr. Bethesda Md 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Medicine, I. of Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; 2000; ISBN 978-0-309-07279-3. [Google Scholar]
- Shea, M.K.; Booth, S.L. Vitamin K. Adv. Nutr. 2022, 13, 350. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, E.; Lengyel, I.; Parravano, M.; Biagini, I.; Veldsman, M.; Badhwar, A.; Betts, M.; Cherubini, A.; Llewellyn, D.J.; Lourida, I.; et al. Ocular Biomarkers for Alzheimer Disease Dementia: An Umbrella Review of Systematic Reviews and Meta-Analyses. JAMA Ophthalmol. 2023, 141, 84. [Google Scholar] [CrossRef] [PubMed]
- Barrett-Young, A. An Eye on Clinical Translation-Considerations for Research Into Ocular Biomarkers of Alzheimer Disease and Related Dementias. JAMA Ophthalmol. 2023, 141. [Google Scholar] [CrossRef] [PubMed]
- Hoeft, B.; Weber, P.; Eggersdorfer, M. Micronutrients - a Global Perspective on Intake, Health Benefits and Economics. Int. J. Vitam. Nutr. Res. Int. Z. Vitam.- Ernahrungsforschung J. Int. Vitaminol. Nutr. 2012, 82, 316–320. [Google Scholar] [CrossRef]
- Victor, L. Fulgoni, I.I.I. Foods, Fortificants, and Supplements: Where Do Americans Get Their Nutrients? J. Nutr. 2011, 141, 1847. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



