Submitted:
07 February 2023
Posted:
07 February 2023
You are already at the latest version
Abstract
Based on functional magnetic resonance imaging and multilayer dynamic network model, the quantified temporal stability of brain network has shown potentials in predicting altered brain functions. The present review focuses on summarizing current knowledge on the commonly-used measures of brain network’s temporal stability and the clinical research progress on them. There are a variety of widely used measures of temporal stability such as the variance/standard deviation of dynamic functional connectivity strengths, the temporal variability, the flexibility (switching rate), and the temporal clustering coefficient, while there is no consensus to date that which measure is the best. The temporal stability of human brain networks may be associated with several factors such as sex, age, cognitive functions, head motion, and data preprocessing/analyzing strategies, which should be considered in the clinical studies. Multiple common psychiatric diseases such as schizophrenia, major depressive disorder, and bipolar disorder have been found to be related to altered temporal stability, especially during the resting state; generally, both excessively decreased and increased temporal stabilities were thought to reflect disease-related brain dysfunctions. However, the measures of temporal stability are still far from applications in clinical diagnoses for neuropsychiatric diseases partly because of the divergent results, and further studies are warranted.
Keywords:
connectome
; dynamic functional connectivity
; dynamic brain network
; schizophrenia
; major depressive disorder
; bipolar disorder
1. Introduction
Functional magnetic resonance imaging (fMRI) is currently one of the best methods to study the functional activity of the human brain under non-invasive conditions [1,2]. Using fMRI technology, many common psychiatric diseases such as schizophrenia [2,3], bipolar disorder [4,5] and major depressive disorder [6,7] have been found to be accompanied by abnormal functional connectivity (FC) between brain areas and changes in brain network topological properties on large scales, which provide potential markers for the auxiliary diagnosis of these diseases.
In traditional fMRI research, computational analyses are generally based on the assumption that functional connections between different areas of the brain remain constant during the whole fMRI scan. However, it has been suggested that the patterns of brain FC actually fluctuate dynamically over time even at the resting state, which are ignored by traditional fMRI data analysis methods [8,9]. Therefore, research on the dynamic fluctuations of the FC patterns of the brain during fMRI scan so-called dynamic FC (dFC) has emerged in recent years [10,11,12]. Several studies have shown that after excluding disturbing factors such as head movement, dFC can still accurately reflect the inherent individual differences between subjects [13,14]. Based on multiple repeated fMRI scans on the same subjects, it was also shown that multiple dFC measures have acceptable test-retest reliability [14,15], which demonstrates the feasibility and reliability of dFC measures.
In dFC studies, a commonly used approach is to model the human brain network as a multilayer “dynamic network” and then evaluate the temporal stability/variability of such dynamic networks in a graph theory-based framework. Such an approach has shown potential for predicting alterations in brain functions in both normal [14,16] and pathological [17,18] conditions in clinical studies. However, compared with traditional static brain network measures, the dynamic brain network model remains a relatively new area and its clinical applications are still in the exploration phase. This review briefly introduces the basic concepts, commonly-used measures, and possible influencing factors of the temporal stability of brain networks, and narratively summarizes its clinical research progresses in common psychiatric diseases such as schizophrenia, bipolar disorder, and depression, in order to provide new highlights for understanding the relationship between common psychiatric diseases and brain functional stability based on the dynamic brain network model.
2. The Basic Concepts and Commonly Used Measures of the Temporal Stability of a Brain Network
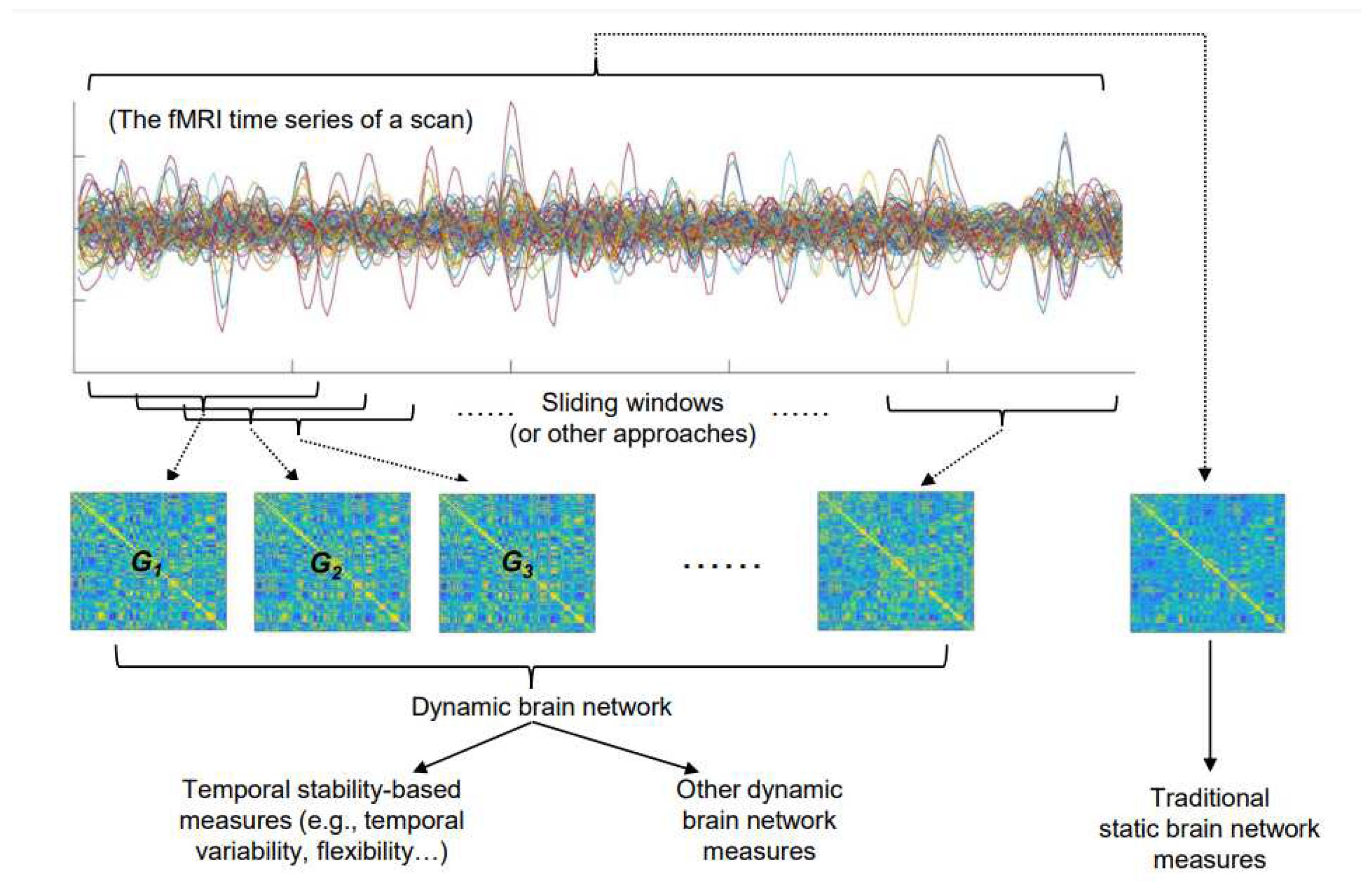
In a framework of the graph theory, the functional brain network can be modeled as a complex network, which abstracts the brain into a series of “nodes” and “edges” connecting those “nodes” [19]. Each of these “nodes” typically represents a brain region defined based on an anatomical or functional brain map, while the “edge” between each of the two “nodes” represents the strength of the functional connection between them [19,20]. In traditional static brain network models, the strengths of these “edges” are constant; in contrast, in dynamic network models, the strengths of each “edge” change dynamically over time [21]. Such changes are typically estimated by dividing the fMRI signal of the entire scan into several time periods, and then calculating the connection strengths of each “edge” in the brain network in different time periods. To achieve this, the often-used method is dividing the entire fMRI signal into several non-overlapped or partially overlapped “time windows” by the “sliding-window” approach [17,22,23,24], while there are also other methods [25]. As a result, a dynamic brain network can be constructed as G = (Gt)t = 1, 2, 3…, where Gt is the layer representing brain dFC within the tth time period (the tth time window) (Figure 1). Compared with the static brain network model, the dynamic brain network model thus incorporates an additional time dimension to track the dynamic changes of the brain network.
After constructing a dynamic brain network, the temporal stability of the brain network can be then estimated from both global (at the whole-brain level) and regional levels. The higher temporal stability indicates that a brain network is more stable (fluctuates less) over time. It is noteworthy that there are various measures of temporal stability proposed by different researchers, which will be introduced later. Moreover, there are measures to quantitatively analyze the other characteristics of a dynamic brain network beyond simply estimating its temporal stability (Figure 1). For instance, another dynamic brain network-based analysis technique is to use clustering, where the layers (time windows) in a dynamic brain network are portioned into a set of clusters or “states” using the K-means clustering [26] or principal components analysis (PCA) [27]; this family of technique summarizes the dFC patterns into a number of “states” and may provide a more comprehensive overview of dynamic changes in the brain network [12]. Nevertheless, there is no consensus to date that which measure is the best to characterize the temporal fluctuations of dFC patterns. Below we introduce several most commonly used measures of temporal stability for a dynamic brain network:
(1) The variance/the standard deviation [16,23,28,29,30]: the possibly most straightforward and simple method to estimate the temporal stability of “edges” in a dynamic brain network is to calculate the variance or the standard deviation of dFC strengths of each edge across different layers (time windows). Intuitively, a higher variance/a higher standard deviation suggests lower temporal stability of dFC between brain nodes. The advantages of this measure include the simple analyzing steps and short calculating time. However, there are studies reporting that such measures may be less reliable when compared to other measures [28,31].
(2) The temporal variability [17,18,32,33,34]: the “temporal variability” of a dynamic brain network is estimated by averaging the dissimilarity of its network structures between different layers (time windows). The exact formula for such calculations can be found in the previous related publications [17,18,32,33,34]. This measure can be computed at both the global (temporal variability) and regional (nodal temporal variability) levels. Besides, it can be also computed at the subnetwork level (within-subnetwork and between-subnetwork temporal variability) to reflect the temporal stability of dFC patterns within/between particular large-scale brain subsystems. A higher value of temporal variability indicates lower temporal stability of a dynamic brain network (more variable over time).
(3) The flexibility [22,35,36,37,38]: this measure is also called “switching rate”; it estimates the temporal stability of a dynamic brain network according to its modularity structures. The flexibility or “switching rate” of a given brain node quantifies the rate at which it transits between different functional modules. Higher flexibility or “switching rate” suggests that the brain network transit between different configurations at a higher rate, and thus with lower temporal stability. The flexibility can be also computed at the global and subnetwork levels, by averaging all nodes within the whole brain network or those nodes belonging to a particular subnetwork.
(4) The temporal clustering coefficient [14,17,21,39,40]: this measure is also called “the temporal correlation coefficient”; it is derived from the definition of analogous metric (clustering coefficient) for conventional static networks. The temporal clustering coefficient estimates the tendency of a dynamic brain network to keep stable over time (its “temporal clustering”) by quantifying the average topological overlap of the network connections between any two consecutive layers (i.e., neighboring time windows). A higher temporal clustering coefficient suggests that the brain network is more likely to keep stable over time (showing higher temporal stability). Similar to the brain network flexibility, the temporal clustering coefficient can be computed at all the nodal, subnetwork, and global levels.
Besides the above four measures, there are also other methods to measure the temporal stability of dynamic brain networks. One example is independently calculating the traditional “static” topological properties of the brain network within each layer (time window) and then estimating their changing ranges by the area under the curve (AUC) [41] or the temporal deviation of each network property (“temporal grading index”) [42]. Based on the earlier-mentioned state-clustering algorithm, the stability of dynamic brain networks can be also measured by switching frequencies between different “states” [43]. Nevertheless, in this review, we will focus on the above-mentioned four kinds of measures that have been commonly used in published neuroimaging studies.
3. Possible Influencing Factors when Analyzing the Temporal Stability of a Brain Network
Prior studies of dynamic human brain networks have shown that the temporal stability of brain networks may be associated with factors such as sex, age, and cognitive functions. Furthermore, it was suggested that the results of dFC studies might be influenced by some factors such as the participants’ head motion during the fMRI scanning, as well as several data preprocessing and analyzing strategies. These reports suggest that attention should be paid to controlling the potential influences of these factors in the clinical applications of dynamic brain network models.
(1) Sex: in studies using the traditional static brain network models, sex and age were typically controlled as covariates since they are thought to have significant influences on many important brain network properties [44,45]. For dFC studies, there have been also several studies to investigate the possible sex effects on the temporal stability of brain networks. Interestingly, although conflicting results exist [46], we notice many of these studies found that the females’ brain networks seem to be more stable over time compared to the males’ brain networks [14,43,47]. For instance, one research using a state-clustering algorithm suggested that the females’ brain networks switch connectivity states less frequently than the males’ and can thus be considered more stable over time [43]; in another study using the temporal clustering coefficient, it was found that the females’ brain networks are more stable than the males’ and such differences are significant at both the global and regional (particularly within the default-mode and subcortical regions) levels [14]. These findings may not only highlight the necessity to control for sex effects in dFC studies, but also provide valuable insight into the sex differences in clinical characteristics of many mental problems and diseases (e.g., females are more likely to be affected by depression) [48,49,50].
(2) Age: there have been a number of studies investigating the possible age-related effects in the temporal stability of dynamic brain networks at the resting state [16,51,52,53], many of which used relatively large samples. For instance, using data from 902 healthy controls, Tang et al. [54] found a significant aging-related increase in temporal variability of dFC within the default-mode network. Similarly, Qin et al. [53] examined age-related differences in dFC patterns with data from 183 subjects aged 7–30, and found that higher age is associated with higher temporal variability of the connections among the visual network, default mode network, and cerebellum. In addition, Marusak et al. [52] analyzed the dynamic brain network characteristics of 146 children and adolescents aged 7 to 16 years old, and found that the temporal variabilities of dFC among multiple neurocognitive networks are positively associated with age. Overall, these results suggest the process of maturation/aging of the brain may be accompanied by increased temporal variability (decreased temporal stability) of the brain network. However, there were also opposite findings: e.g., Xia et al. [51] reported that age is negatively associated with the variability of dynamic functional networks by analyzing the resting scan data of a total of 434 subjects. Nevertheless, the changes in temporal stability-based dFC measures may be an important characteristic of the maturation/aging process but further investigations are still needed to provide a deep understanding of such relationships.
(3) Cognitive functions: several studies have shown that the temporal stability of dynamic brain networks may be associated with multiple dimensions of cognitive function. For instance, Hilger et al. [55] found that higher intelligence is associated with higher temporal stability (lower temporal variability) of brain network modularity. The other cognitive factors which may be associated with the temporal stability of brain networks include executive function [56], learning ability [35], working memory [36], cognitive flexibility [57], verbal creativity [33], and the need for cognition [58]. Therefore, it might be better to consider the possible influences of different levels of cognition in clinical studies on dFC, especially when performing direct comparisons between the groups of healthy controls and psychiatric populations. An alternative way to achieve this could be to include education level as a covariate in the analyzes, as done by many researchers [14,17,59,60].
(4) Head motion: there have been multiple studies pointing out that the subjects’ head motion would considerably affect the obtained dFC patterns [61,62]. Therefore, to minimize the possible influences of head motion, it is necessary to perform motion artifact removal steps during the data pre-processing stage [61]. Moreover, we propose that in clinical studies, it would be better to match the levels of head motion between different groups; it would also be better to include the head motion parameters (e.g., mean framewise-displacement) as an additional covariate in the analyzes, as done by many researchers [14,17,59,60].
(5) Data preprocessing and analyzing strategies: in dFC studies, it is possible that the values of dynamic brain network measures (including temporal stability) could be affected by different choices in some data preprocessing and analyzing strategies. For example, the dynamic brain network measures may be different when performing and not performing the global signal regression (GSR) during data preprocessing; the dynamic brain network measures also could be different when choosing different window lengths and step lengths, two key parameters of the sliding-window approach [61]. Actually, there are still debates about the necessity of GSR [63] and about the optimal window lengths/step lengths in the sliding-window approach [31,64]. Nevertheless, in a number of studies on the temporal stability of brain networks, the analyzes were repeated with different strategies and it was found that consistent results can be obtained when performing/not performing GSR, and when using a set of different window lengths/step lengths in the sliding-window step [14,17,24]; therefore, the main conclusions in these studies are unlikely to be largely driven by the choices in data preprocessing and analyzing strategies.
4. Research Progress on Possible Relationships between the Temporal Stability of Resting-State Brain Networks and Common Psychiatric Diseases
In recent years, researchers have explored abnormal changes in the temporal stability of brain networks in patients with multiple psychiatric diseases, especially at the resting state and in three of the most common psychiatric diseases worldwide: schizophrenia, major depressive disorder, and bipolar disorder. Here, we briefly summarized the relevant research progress as follows (all mentioned studies are listed in Table 1, Table 2 and Table 3):
(1) Schizophrenia: schizophrenia is perhaps one of the most well-studied psychiatric diseases as for its relationship with the brain’s temporal stability based on the dynamic brain network model. For instance, in 2016, Zhang et al. [18] firstly studied two separate samples of 69 schizophrenic patients/62 healthy controls and 53 schizophrenic patients/67 healthy controls, respectively. They found that the patients with schizophrenia showed significantly increased temporal variability (decreased temporal stability) of dFCs in the subcortical regions such as the thalamus, pallidum, and putamen, as well as in the visual cortex at the resting state; meanwhile, the schizophrenia patients showed significantly decreased temporal variability (increased temporal stability) in the relevant regions of the default-mode network. After that, Dong et al. [34] and Long et al. [32] also investigated the changes in temporal variability of brain dFC patterns in schizophrenia patients and they found similar results: both of them observed that the schizophrenia patients showed significantly increased temporal variability (decreased temporal stability) in sensory and perceptual systems (including the visual, sensorimotor, and attention networks, as well as thalamus) but decreased temporal variability (increased temporal stability) in higher-order networks (e.g., the default-mode and frontal–parietal networks). Using the measure of flexibility, Gifford et al. [65] also found that the temporal stabilities of dFC in multiple brain regions including the thalamus are significantly decreased in schizophrenia patients.
Overall, all the above findings point to a significant widespread decrease in temporal stability of dFC in most of the brain systems, especially the sensory and perceptual systems (e.g., visual cortex, sensorimotor cortex, and thalamus), and an increase in certain higher systems (e.g., frontal-parietal and default-mode networks) in schizophrenia patients. Actually, there are several other studies (beyond the above ones) that reported similar conclusions [66,67], although inconsistent reports also exist [68]. These findings may support the opinion that such widespread aberrant dynamic brain network reconfigurations may be a potential biomarker for schizophrenia, suggestive of impaired abilities in effectively filter inputs in sensory/perceptual systems and integrating information in high-order networks, which may underlie the perceptual and cognitive deficits in schizophrenia [32,34].

Table 1.
Summary of main findings of changes in the temporal stability of brain network in patients with schizophrenia (SZ) as mentioned in this review. HCs, healthy controls.
Table 1.
Summary of main findings of changes in the temporal stability of brain network in patients with schizophrenia (SZ) as mentioned in this review. HCs, healthy controls.
| Reference | Sample | The measure of temporal stability | Main findings on the temporal stability of dFC in SZ patients |
| Zhang et al. [18] | Two independent datasets: 69 SZ patients/62 HCs and 53 SZ patients/67 HCs | Temporal variability | Decreased temporal stability in the subcortical and visual regions; increased temporal stability in the default-mode regions |
| Dong et al. [34] | 102 SZ patients/124 HCs | Temporal variability | Decreased temporal stability in visual, sensorimotor, and attention networks, as well as thalamus; increased temporal stability in high-order networks (default-mode and frontal–parietal networks) |
| Long et al. [32] | 66 SZ patients/66 HCs | Temporal variability | Decreased temporal stability involving wide areas including the sensorimotor, visual, attention, limbic, and subcortical areas; increased temporal stability in the default-mode areas |
| Gifford et al. [65] | 55 SZ patients/72 HCs | Flexibility | Decreased temporal stability in cerebellar, subcortical (including the thalamus), and fronto-parietal task control resting state networks |
| Guo et al. [68] | 22 SZ patients/60 HCs | Temporal variability | Decreased temporal stability of dFC anchored on the precuneus |
| Wang et al. [66] | 42 SZ patients/35 HCs | Standard deviation | Decreased temporal stability in dFCs among multiple networks |
| Sheng et al. [67] | Two independent datasets: 51 SZ patients/63 HCs and 36 SZ patients/60 HCs | Temporal variability | Decreased temporal stability in the prefrontal cortex, anterior cingulate cortex, temporal cortex, visual cortex, and hippocampus |
(2) Major depressive disorder: major depressive disorder is also one of the most focused diseases in clinical studies on the temporal stability of human brain networks. Among these studies, Long et al. [17] reported that the level of temporal variabilities of dFCs are significantly increased (indicating decreased temporal stability) in patients with major depressive disorder at both the whole-brain level and sub-network level within the default-mode, sensorimotor and subcortical networks. Using the measure of temporal clustering coefficient, Zhao et al. [39] found that patients with major depressive disorder showed decreased temporal stability at the global level and in regions of sensory perception systems. Hou et al. [69] and Ouyang et al. [40] found that patients with major depressive disorder showed decreased temporal stability within the striatum and default-mode regions, respectively. Overall, these findings suggest that major depressive disorder may be associated with excessive fluctuation (decreased temporal stability) of functional brain network organizations, and the default-mode areas are most prominently involved. The default-mode network is thought to be primarily involved in processing one’s self-referential and internally directed information [70]. Therefore, the instability of the default-mode network dFCs was hypothesized to be related to uncontrollable, too-frequent thinking activities about negative emotions and event, known as “rumination” in patients with depression [17]. Actually, another study by Wise at al. [71] has also strongly supported such an opinion: they proved that temporal stabilities of dFCs within several key default-mode regions are significantly decreased in major depressive disorder, which was replicated in two independent samples.
However, we notice that the findings in a number of other published studies are not consistent with the above reports. For example, Demirtaş et al. [72] compared the temporal variability of dFCs quantified as the index of dispersion (variance/mean) between patients with major depressive disorder and healthy controls; and they found that the patients showed significantly decreased variability (increased temporal stability) of dFCs between the default-mode and fronto-parietal networks. In another study using the measure of brain network flexibility, Tian et al. [73] found that the patients with major depressive disorder showed increased temporal stability within the default-mode and cognitive control networks. Han et al. [74] also reported decreased switching rates (increased temporal stability) within several default-mode regions such as the precuneus and dorsal medial prefrontal cortex in patients with major depressive disorder. Furthermore, Zhou at al. [30] also reported that major depressive disorder is characterized by excessive stable (increased temporal stability) dFCs within default-mode areas such as the precuneus. These results are not consistent and even conflict with those studies mentioned in the last paragraph.
Here, we propose that the inconsistencies in previous studies on major depressive disorder may be partly due to several reasons. First, we notice that the sample sizes in some studies are relatively small (Table 2), which may lead to less reliable results [75]. Second, it is possible that major depressive disorder is a disease with significant clinical and biological heterogeneity; for example, it has been suggested that major depressive disorder-related brain dysfunctions may differ in different age groups (e.g., early adulthood vs late-life) of patients [76]. Therefore, subtypes with distinct dFC profiles (e.g., hyper- and hypo-stability) may exist in major depressive disorder, which can be investigated in future studies.
Table 2.
Summary of main findings of changes in the temporal stability of brain network in patients with major depressive disorder (MDD) as mentioned in this review. HCs, healthy controls.
Table 2.
Summary of main findings of changes in the temporal stability of brain network in patients with major depressive disorder (MDD) as mentioned in this review. HCs, healthy controls.
| Reference | Sample | The measure of temporal stability | Main findings in MDD patients |
| Demirtaş et al. [72] | 27 MDD patients/ 27 HCs |
Variance/mean | Increased temporal stability of dFCs between the default-mode and fronto-parietal networks |
| Long et al. [17] | 460 MDD patients/ 473 HCs |
Temporal variability and temporal clustering coefficient | Decreased temporal stability which were mainly found in the default-mode, sensorimotor and subcortical areas |
| Wise et al. [71] | Two independent datasets: 20 MDD patients/19 HCs and 19 MDD patients/19 HCs | The standard deviation | Decreased temporal stability within several key default-mode regions |
| Zhao et al. [39] | 55 MDD patients/ 62 HCs |
Temporal clustering coefficient | Decreased temporal stability at the global level and in regions of sensory perception systems |
| Hou et al. [69] | 77 MDD patients/ 40 HCs |
Temporal variability | Decreased temporal stability in the inferior occipital gyrus and pallidum |
| Ouyang et al. [40] | 55 MDD patients/ 21 HCs |
Temporal clustering coefficient | Decreased temporal stability at the global level, and within the default-mode and subcortical networks |
| Zhou et al. [30] | 19 MDD patients/ 22 HCs |
The variance | Increased temporal stability in the dorsolateral prefrontal cortex and precuneus connectivity |
| Tian et al. [73] | 35 MDD patients/ 35 HCs |
Flexibility | Increased temporal stability within the default-mode and cognitive control networks |
| Han et al. [74] | 61 MDD patients/61 HCs | Flexibility (switching rate) | Increased temporal stability in the precuneus, parahippocampal gyrus, dorsal medial prefrontal cortex, anterior insula, amygdala, and striatum |
(3) Bipolar disorder: similar to schizophrenia and major depressive disorder, many researchers have explored possible changes in the temporal stability of brain networks in patients with bipolar disorder. For example, Nguyen et al. [77] found that compared with healthy controls, patients with euthymic bipolar disorder showed significantly reduced dFC variability (increased temporal stability) between the medial prefrontal lobe and the posterior cingulate gyrus in the resting-state. Han et al. [74] found that patients with bipolar disorder showed decreased network switching rates (increased temporal stability) of regions including the left precuneus, bilateral dorsal medial prefrontal cortex, and bilateral parahippocampal gyrus. Liang et al. [78] found that compared to healthy participants, patients with bipolar disorder showed decreased variance (increased temporal stability) of dFC between the posterior cingulate cortex and medial prefrontal cortex. Using the measure of temporal variability, Long et al. [32] found increased temporal variability (decreased temporal stability) of dFCs profiles within subcortical areas, as well as between the thalamus and sensorimotor areas. Wang et al. [79] reported that depressed bipolar disorder patients showed increased temporal stability of dFC between the default-mode network and central executive network when compared to healthy controls. Furthermore, Luo et al. [80] reported the group of depressed bipolar disorder patients had reduced dFC variance (increased temporal stability) between the bilateral posterior cingulate cortex/ precuneus and the left inferior parietal lobule than the healthy group.
In summary, most of the published studies mentioned above suggest that bipolar disorder is associated with an excessively increased temporal stability of the brain network. Moreover, at the local level, certain brain areas have been repeatedly reported to be involved in two or more studies (e.g., the posterior cingulate gyrus) (Table 3). However, generally, there is no significant convergence of the regional dFC abnormalities in bipolar disorder across these published literature. Such inconsistencies in previous studies may be firstly due to their relatively small sample sizes (Table 3), which may lead to relatively low statistical power and relatively low reliability [75]. Beyond that, it is possible that bipolar disorder-related brain dysfunctions may be characterized by different changes in different clinical states (e.g., in depression, mania, and euthymic states [81]. However, the direct comparisons on brain network stability between different clinical states of bipolar disorder are still relatively limited to our knowledge, which merits further investigations in the future.
Table 3.
Summary of main findings of changes in the temporal stability of brain network in patients with bipolar disorder (BD) as mentioned in this review. HCs, healthy controls.
Table 3.
Summary of main findings of changes in the temporal stability of brain network in patients with bipolar disorder (BD) as mentioned in this review. HCs, healthy controls.
| Reference | Sample | The measure of temporal stability | Main findings in BD patients |
| Nguyen et al. [77] | 15 euthymic BD patients/19 HCs | Standard deviation | Increased temporal stability between the medial prefrontal lobe and posterior cingulate gyrus |
| Han et al. [74] | 40 BD patients/61 HCs | Flexibility (switching rate) | Increased temporal stability in the precuneus, parahippocampal gyrus, and dorsal medial prefrontal cortex |
| Wang et al. [79] | 51 depressed BD patients/52 HCs | Standard deviation | Increased temporal stability between default-mode network and central executive network |
| Long et al. [32] | 53 BD patients/66 HCs | Temporal variability | Decreased temporal stability of regional dFCs within subcortical areas as well as between the thalamus and sensorimotor areas |
| Liang et al. [78] | 18 BD patients/19 HC | Standard deviation | Increased temporal stability in dFC between the posterior cingulate cortex and medial prefrontal cortex |
| Luo et al. [80] | 106 depressed BD patients/130 HCs | Standard deviation | Decreased temporal stability between the posterior cingulate cortex/precuneus and the inferior parietal lobule |
(4) Other psychiatric disorders: although we focus on schizophrenia, major depressive disorder, and bipolar disorder in this review, alterations in temporal stabilities of dynamic brain networks have been also reported in multiple other neuropsychiatric disorders. For example, Harlalka et al. [82] found that patients with autism spectrum disorder showed a significant increase in dynamic variability (decreased temporal stability) in a wide range of brain network connections. Besides, significantly decreased temporal stabilities of brain networks have been associated with both substance [83] and non-substance [59] addictions, suggesting that brain network instability may play an important role in the onset of these disorders.
5. Discussion: Summary and Prospects
In summary, based on the fMRI technical and the dynamic brain network model, researchers have proposed a number of measures to quantify the temporal stability of the human brain network, and their associations with multiple common psychiatric diseases (especially in patients with schizophrenia, major depressive disorder, and bipolar disorder) have been explored. Generally, both excessively decreased and increased temporal stabilities have been reported in psychiatric populations, and both of them were thought to be reflective of disease-related brain dysfunctions. For example, in most of the published relevant studies, schizophrenia was often associated with decreased temporal stability (Table 1), while bipolar disorder was often associated with increased temporal stability of brain networks (Table 3). These findings might provide a unique perspective for deepening our understanding of these diseases.
However, the measures of temporal stability are still far from applications in clinical diagnoses for neuropsychiatric diseases, partly because of the divergent and even conflicting results on its associations with common psychiatric diseases (Table 1, Table 2 and Table 3). As discussed earlier, the inconsistent results in previous studies may be due to the relatively small sample sizes in many of them, as well as the possibility that there is biological heterogeneity in these diseases (e.g., major depressive disorder-related brain dysfunctions may differ in different age groups of patients). Therefore, future studies with larger samples may be warranted to further investigate the relationships between brain network’s temporal stability and common psychiatric diseases, as well as the potential biological heterogeneity in these diseases as reflected by temporal stability.
It is also noteworthy that while most current clinical studies on the temporal stability of brain networks were performed during a resting state, it has been suggested that altered temporal stability during a certain task (e.g., working memory task [84]) can be reflective of brain dysfunctions, too. Nevertheless, the number of task-based studies on temporal stability is much more limited than that of resting-state ones; for such a reason, we chose to focus on the temporal stability of brain networks during rest in this review. More future studies may be needed to investigate the possible associations between common psychiatric diseases and the temporal stability of brain networks under certain cognitive/emotional tasks.
Another potentially valuable direction in future studies is to investigate the possible transdiagnostic alterations in the temporal stability of brain networks (those alterations shared by different diseases). It is well-known that many psychiatric disorders have some overlapping clinical features (e.g., psychotic symptoms can happen in both schizophrenia and bipolar disorder), and it is thus hypothesized that there may be some common pathogenesis between them [32]. Therefore, investigating the common and different features of the brain network’s temporal stability across different psychiatric diseases may help to strengthen our understanding of their shared and distinct pathogenesis. Actually, several studies have evaluated the differences in temporal stability of brain networks between schizophrenia and bipolar disorder [32], as well as between major depressive disorder and bipolar disorder [74]. Nevertheless, the number of studies simultaneously including multiple diseases is still limited to our knowledge, and more studies are needed in the future.
Finally, many psychiatric disorders (e.g., schizophrenia) have been proved to be accompanied by structural changes in the brain, such as a significant decline in the white matter integrity [85]. However, it is poorly known whether there is a link between these changes and altered temporal stability of the brain network. All these questions mentioned above may deserve further exploration in future studies.
Author Contributions
XL: YL, and ZL conceived the idea and structured the manuscript. XL and YL drafted the manuscript. YL designed the figures and tables. ZL reviewed and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (82201692 to YL, 82071506 to ZL), and the Natural Science Foundation of Hunan Province, China (2021JJ40851 to YL).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Canario, E.; Chen, D.; Biswal, B. A Review of Resting-State FMRI and Its Use to Examine Psychiatric Disorders. Psychoradiology 2021, 1, 42–53. [Google Scholar] [CrossRef]
- Lin, Z.; Long, Y.; Wu, Z.; Xiang, Z.; Ju, Y.; Liu, Z. Associations between Brain Abnormalities and Common Genetic Variants for Schizophrenia: A Narrative Review of Structural and Functional Neuroimaging Findings. Ann Palliat Med 2021, 10, 10031–10052. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Zhou, H.; Cannon, T.D. Functional Connectome-Wide Associations of Schizophrenia Polygenic Risk. Mol Psychiatry 2021, 26, 2553–2561. [Google Scholar] [CrossRef]
- Du, M.; Zhang, L.; Li, L.; Ji, E.; Han, X.; Huang, G.; Liang, Z.; Shi, L.; Yang, H.; Zhang, Z. Abnormal Transitions of Dynamic Functional Connectivity States in Bipolar Disorder: A Whole-Brain Resting-State FMRI Study. J Affect Disord 2021, 289, 7–15. [Google Scholar] [CrossRef]
- Macoveanu, J.; Stougaard, M.E.; Kjærstad, H.L.; Knudsen, G.M.; Vinberg, M.; Kessing, L. v; Miskowiak, K.W. Trajectory of Aberrant Reward Processing in Patients with Bipolar Disorder – A Longitudinal FMRI Study. J Affect Disord 2022, 312, 235–244. [Google Scholar] [CrossRef]
- Yang, H.; Chen, X.; Chen, Z.B.; Li, L.; Li, X.Y.; Castellanos, F.X.; Bai, T.J.; Bo, Q.J.; Cao, J.; Chang, Z.K.; et al. Disrupted Intrinsic Functional Brain Topology in Patients with Major Depressive Disorder. Mol Psychiatry 2021, 26, 7363–7371. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.G.; Chen, X.; Li, L.; Castellanos, F.X.; Bai, T.J.; Bo, Q.J.; Cao, J.; Chen, G.M.; Chen, N.X.; Chen, W.; et al. Reduced Default Mode Network Functional Connectivity in Patients with Recurrent Major Depressive Disorder. Proc Natl Acad Sci U S A 2019, 116, 9078–9083. [Google Scholar] [CrossRef]
- Hutchison, R.M.; Womelsdorf, T.; Gati, J.S.; Everling, S.; Menon, R.S. Resting-State Networks Show Dynamic Functional Connectivity in Awake Humans and Anesthetized Macaques. Hum Brain Mapp 2013, 34, 2154–2177. [Google Scholar] [CrossRef]
- Chang, C.; Glover, G.H. Time-Frequency Dynamics of Resting-State Brain Connectivity Measured with FMRI. Neuroimage 2010, 50, 81–98. [Google Scholar] [CrossRef]
- Cavanna, F.; Vilas, M.G.; Palmucci, M.; Tagliazucchi, E. Dynamic Functional Connectivity and Brain Metastability during Altered States of Consciousness. Neuroimage 2018, 180. [Google Scholar] [CrossRef]
- Shunkai, L.; Chen, P.; Zhong, S.; Chen, G.; Zhang, Y.; Zhao, H.; He, J.; Su, T.; Yan, S.; Luo, Y.; et al. Alterations of Insular Dynamic Functional Connectivity and Psychological Characteristics in Unmedicated Bipolar Depression Patients with a Recent Suicide Attempt. Psychol Med 2022. [Google Scholar] [CrossRef] [PubMed]
- Preti, M.G.; Bolton, T.A.; van de Ville, D. The Dynamic Functional Connectome: State-of-the-Art and Perspectives. Neuroimage 2017, 160, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liao, X.; Xia, M.; He, Y. Chronnectome Fingerprinting: Identifying Individuals and Predicting Higher Cognitive Functions Using Dynamic Brain Connectivity Patterns. Hum Brain Mapp 2018, 39. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Ouyang, X.; Yan, C.; Wu, Z.; Huang, X.; Pu, W.; Cao, H.; Liu, Z.; Palaniyappan, L. Evaluating Test–Retest Reliability and Sex-/Age-Related Effects on Temporal Clustering Coefficient of Dynamic Functional Brain Networks. Hum Brain Mapp 2023, n/a. [Google Scholar] [CrossRef]
- Omidvarnia, A.; Zalesky, A.; Mansour, L.S.; van de Ville, D.; Jackson, G.D.; Pedersen, M. Temporal Complexity of FMRI Is Reproducible and Correlates with Higher Order Cognition. Neuroimage 2021, 230. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, W.; Zhao, X.; Sha, M.; Liu, Y.; Zhang, X.; Ma, J.; Ni, H.; Ming, D. Age-Related Decline in the Variation of Dynamic Functional Connectivity: A Resting State Analysis. Front Aging Neurosci 2017, 9. [Google Scholar] [CrossRef]
- Long, Y.; Cao, H.; Yan, C.; Chen, X.; Li, L.; Castellanos, F.X.; Bai, T.; Bo, Q.; Chen, G.; Chen, N.; et al. Altered Resting-State Dynamic Functional Brain Networks in Major Depressive Disorder: Findings from the REST-Meta-MDD Consortium. Neuroimage Clin 2020, 26, 102163. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, W.; Liu, Z.; Zhang, K.; Lei, X.; Yao, Y.; Becker, B.; Liu, Y.; Kendrick, K.M.; Lu, G.; et al. Neural, Electrophysiological and Anatomical Basis of Brain-Network Variability and Its Characteristic Changes in Mental Disorders. Brain 2016, 139, 2307–2321. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, M.; Sporns, O. Complex Network Measures of Brain Connectivity: Uses and Interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Liu, D.; Tang, S.; Wu, Z.; Yang, J.; Liu, Z.; Wu, G.; Sariah, A.; Ouyang, X.; Long, Y. Changes in Brain Network Properties in Major Depressive Disorder Following Electroconvulsive Therapy: A Combined Static and Dynamic Functional Magnetic Resonance Imaging Study. Ann Palliat Med 2022, 11, 1969–1980. [Google Scholar] [CrossRef]
- Sizemore, A.E.; Bassett, D.S. Dynamic Graph Metrics: Tutorial, Toolbox, and Tale. Neuroimage 2018, 180, 417–427. [Google Scholar] [CrossRef]
- Long, Y.; Chen, C.; Deng, M.; Huang, X.; Tan, W.; Zhang, L.; Fan, Z.; Liu, Z. Psychological Resilience Negatively Correlates with Resting-State Brain Network Flexibility in Young Healthy Adults: A Dynamic Functional Magnetic Resonance Imaging Study. Ann Transl Med 2019, 7, 809–809. [Google Scholar] [CrossRef]
- Chen, Y.; Cui, Q.; Xie, A.; Pang, Y.; Sheng, W.; Tang, Q.; Li, D.; Huang, J.; He, Z.; Wang, Y.; et al. Abnormal Dynamic Functional Connectivity Density in Patients with Generalized Anxiety Disorder. J Affect Disord 2020, 261. [Google Scholar] [CrossRef]
- Sun, Y.; Collinson, S.L.; Suckling, J.; Sim, K. Dynamic Reorganization of Functional Connectivity Reveals Abnormal Temporal Efficiency in Schizophrenia. Schizophr Bull 2019, 45, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Zheng, C.Y.; Handwerker, D.A.; Bandettini, P.A.; Calhoun, V.D.; Mitra, S.; Gonzalez-Castillo, J. Efficacy of Different Dynamic Functional Connectivity Methods to Capture Cognitively Relevant Information. Neuroimage 2019, 188. [Google Scholar] [CrossRef]
- Allen, E.A.; Damaraju, E.; Plis, S.M.; Erhardt, E.B.; Eichele, T.; Calhoun, V.D. Tracking Whole-Brain Connectivity Dynamics in the Resting State. Cerebral Cortex 2014, 24, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, N.; Richiardi, J.; Gschwind, M.; Simioni, S.; Annoni, J.M.; Schluep, M.; Vuilleumier, P.; van de Ville, D. Principal Components of Functional Connectivity: A New Approach to Study Dynamic Brain Connectivity during Rest. Neuroimage 2013, 83, 937–950. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, J.; Yang, Y.; Zhao, S.; Guo, L.; Han, J.; Hu, X. Test–Retest Reliability of Dynamic Functional Connectivity in Naturalistic Paradigm Functional Magnetic Resonance Imaging. Hum Brain Mapp 2022, 43. [Google Scholar] [CrossRef] [PubMed]
- Choe, A.S.; Nebel, M.B.; Barber, A.D.; Cohen, J.R.; Xu, Y.; Pekar, J.J.; Caffo, B.; Lindquist, M.A. Comparing Test-Retest Reliability of Dynamic Functional Connectivity Methods. Neuroimage 2017, 158, 155–175. [Google Scholar] [CrossRef]
- Zhou, W.; Yuan, Z.; Yingliang, D.; Chaoyong, X.; Ning, Z.; Chun, W. Differential Patterns of Dynamic Functional Connectivity Variability in Major Depressive Disorder Treated with Cognitive Behavioral Therapy. J Affect Disord 2021, 291. [Google Scholar] [CrossRef]
- Zhang, C.; Baum, S.A.; Adduru, V.R.; Biswal, B.B.; Michael, A.M. Test-Retest Reliability of Dynamic Functional Connectivity in Resting State FMRI. Neuroimage 2018, 183, 907–918. [Google Scholar] [CrossRef]
- Long, Y.; Liu, Z.; Chan, C.K.Y.; Wu, G.; Xue, Z.; Pan, Y.; Chen, X.; Huang, X.; Li, D.; Pu, W. Altered Temporal Variability of Local and Large-Scale Resting-State Brain Functional Connectivity Patterns in Schizophrenia and Bipolar Disorder. Front Psychiatry 2020, 11, 422. [Google Scholar] [CrossRef]
- Sun, J.; Liu, Z.; Rolls, E.T.; Chen, Q.; Yao, Y.; Yang, W.; Wei, D.; Zhang, Q.; Zhang, J.; Feng, J.; et al. Verbal Creativity Correlates with the Temporal Variability of Brain Networks during the Resting State. Cerebral Cortex 2019, 29, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Duan, M.; Wang, Y.; Zhang, X.; Jia, X.; Li, Y.; Xin, F.; Yao, D.; Luo, C. Reconfiguration of Dynamic Functional Connectivity in Sensory and Perceptual System in Schizophrenia. Cerebral Cortex 2019, 29, 3577–3589. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Wymbs, N.F.; Porter, M.A.; Mucha, P.J.; Carlson, J.M.; Grafton, S.T. Dynamic Reconfiguration of Human Brain Networks during Learning. Proc Natl Acad Sci U S A 2011, 108, 7641–7646. [Google Scholar] [CrossRef]
- Pedersen, M.; Zalesky, A.; Omidvarnia, A.; Jackson, G.D. Multilayer Network Switching Rate Predicts Brain Performance. Proc Natl Acad Sci U S A 2018, 115, 13376–13381. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Liu, Z.; Cao, H.; Yang, J.; Wu, Z.; Long, Y. Childhood Trauma Is Linked to Decreased Temporal Stability of Functional Brain Networks in Young Adults. J Affect Disord 2021, 290, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Betzel, R.F.; Satterthwaite, T.D.; Gold, J.I.; Bassett, D.S. Positive Affect, Surprise, and Fatigue Are Correlates of Network Flexibility. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Chen, N.; Li, Y.; Guo, H.; Guo, M.; Yao, Z.; Hu, B. Altered Temporal Reachability Highlights the Role of Sensory Perception Systems in Major Depressive Disorder. Prog Neuropsychopharmacol Biol Psychiatry 2021, 112. [Google Scholar] [CrossRef]
- Ouyang, X.; Long, Y.; Wu, Z.; Liu, D.; Liu, Z.; Huang, X. Temporal Stability of Dynamic Default Mode Network Connectivity Negatively Correlates with Suicidality in Major Depressive Disorder. Brain Sci 2022, 12. [Google Scholar] [CrossRef]
- Liao, W.; Li, J.; Duan, X.; Cui, Q.; Chen, H.; Chen, H. Static and Dynamic Connectomics Differentiate between Depressed Patients with and without Suicidal Ideation. Hum Brain Mapp 2018, 39. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, L.; Ji, J.; Ma, B.; Zhao, Z.; Wu, M.; Zheng, W.; Zhang, Z. Temporal Grading Index of Functional Network Topology Predicts Pain Perception of Patients With Chronic Back Pain. Front Neurol 2022, 13. [Google Scholar] [CrossRef]
- de Lacy, N.; McCauley, E.; Kutz, J.N.; Calhoun, V.D. Sex-Related Differences in Intrinsic Brain Dynamism and Their Neurocognitive Correlates. Neuroimage 2019, 202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cahill, N.D.; Arbabshirani, M.R.; White, T.; Baum, S.A.; Michael, A.M. Sex and Age Effects of Functional Connectivity in Early Adulthood. Brain Connect 2016, 6, 700–713. [Google Scholar] [CrossRef]
- Tian, L.; Wang, J.; Yan, C.; He, Y. Hemisphere- and Gender-Related Differences in Small-World Brain Networks: A Resting-State Functional MRI Study. Neuroimage 2011, 54, 191–202. [Google Scholar] [CrossRef]
- Mao, N.; Zheng, H.; Long, Z.; Yao, L.; Wu, X. Gender Differences in Dynamic Functional Connectivity Based on Resting-State FMRI. In Proceedings of the Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society, EMBS; IEEE, 2017; pp. 2940–2943. [Google Scholar]
- Cai, B.; Zhang, G.; Zhang, A.; Hu, W.; Stephen, J.M.; Wilson, T.W.; Calhoun, V.D.; Wang, Y.P. A GICA-TVGL Framework to Study Sex Differences in Resting State FMRI Dynamic Connectivity. J Neurosci Methods 2020, 332. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, D.; Zhang, J.; Zhang, W.; Tao, H.; Ouyang, X.; Wu, G.; Chen, M.; Yu, M.; Zhou, L.; et al. Sex Difference in the Prevalence of Psychotic-like Experiences in Adolescents: Results from a Pooled Study of 21,248 Chinese Participants. Psychiatry Res 2022, 317, 114894. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Wang, D.; Tian, Y.; Du, Y.; Chen, J.; Zhou, H.; Chen, D.; Wang, L.; Alonzo, B.A.; Emily Wu, H.; et al. Sex Difference in Association between Insomnia and Cognitive Impairment in Patients with Chronic Schizophrenia. Schizophr Res 2022, 240. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wang, B.; Xiang, Z.; Zou, Z.; Liu, Z.; Long, Y.; Chen, X. Increasing Trends in Mental Health Problems Among Urban Chinese Adolescents: Results From Repeated Cross-Sectional Data in Changsha 2016–2020. Front Public Health 2022, 10, 1–7. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, Q.; Shi, L.; Li, M.Z.; Gong, W.; Chen, H.; Qiu, J. Tracking the Dynamic Functional Connectivity Structure of the Human Brain across the Adult Lifespan. Hum Brain Mapp 2019, 40, 717–728. [Google Scholar] [CrossRef]
- Marusak, H.A.; Calhoun, V.D.; Brown, S.; Crespo, L.M.; Sala-Hamrick, K.; Gotlib, I.H.; Thomason, M.E. Dynamic Functional Connectivity of Neurocognitive Networks in Children. Hum Brain Mapp 2017, 38, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Chen, S.G.; Hu, D.; Zeng, L.L.; Fan, Y.M.; Chen, X.P.; Shen, H. Predicting Individual Brain Maturity Using Dynamic Functional Connectivity. Front Hum Neurosci 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Wu, Z.; Cao, H.; Chen, X.; Wu, G.; Tan, W.; Liu, D.; Yang, J.; Long, Y.; Liu, Z. Age-Related Decrease in Default-Mode Network Functional Connectivity Is Accelerated in Patients With Major Depressive Disorder. Front Aging Neurosci 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Hilger, K.; Fukushima, M.; Sporns, O.; Fiebach, C.J. Temporal Stability of Functional Brain Modules Associated with Human Intelligence. Hum Brain Mapp 2020, 41, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Schäfer, A.; Walter, H.; Erk, S.; Romanczuk-Seiferth, N.; Haddad, L.; Schweiger, J.I.; Grimm, O.; Heinz, A.; Tost, H.; et al. Dynamic Reconfiguration of Frontal Brain Networks during Executive Cognition in Humans. Proc Natl Acad Sci U S A 2015, 112, 11678–11683. [Google Scholar] [CrossRef] [PubMed]
- Douw, L.; Wakeman, D.G.; Tanaka, N.; Liu, H.; Stufflebeam, S.M. State-Dependent Variability of Dynamic Functional Connectivity between Frontoparietal and Default Networks Relates to Cognitive Flexibility. Neuroscience 2016, 339, 12–21. [Google Scholar] [CrossRef]
- He, L.; Zhuang, K.; Li, Y.; Sun, J.; Meng, J.; Zhu, W.; Mao, Y.; Chen, Q.; Chen, X.; Qiu, J. Brain Flexibility Associated with Need for Cognition Contributes to Creative Achievement. Psychophysiology 2019, 56. [Google Scholar] [CrossRef]
- Liu, D.; Liu, X.; Long, Y.; Xiang, Z.; Wu, Z.; Liu, Z.; Bian, D.; Tang, S. Problematic Smartphone Use Is Associated with Differences in Static and Dynamic Brain Functional Connectivity in Young Adults. Front Neurosci 2022, 16. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, L.; Li, K.; Li, P.; Xie, W.; Zeng, Y.; Kong, L.; Long, T.; Huang, L.; Liu, X.; et al. Abnormal Dynamic Functional Network Connectivity in Male Obstructive Sleep Apnea with Mild Cognitive Impairment: A Data-Driven Functional Magnetic Resonance Imaging Study. Front Aging Neurosci 2022, 14. [Google Scholar] [CrossRef]
- Savva, A.D.; Kassinopoulos, M.; Smyrnis, N.; Matsopoulos, G.K.; Mitsis, G.D. Effects of Motion Related Outliers in Dynamic Functional Connectivity Using the Sliding Window Method. J Neurosci Methods 2020, 330. [Google Scholar] [CrossRef]
- Hutchison, R.M.; Womelsdorf, T.; Allen, E.A.; Bandettini, P.A.; Calhoun, V.D.; Corbetta, M.; della Penna, S.; Duyn, J.H.; Glover, G.H.; Gonzalez-Castillo, J.; et al. Dynamic Functional Connectivity: Promise, Issues, and Interpretations. Neuroimage 2013. [Google Scholar] [CrossRef]
- Murphy, K.; Fox, M.D. Towards a Consensus Regarding Global Signal Regression for Resting State Functional Connectivity MRI. Neuroimage 2017, 154, 169–173. [Google Scholar] [CrossRef]
- Leonardi, N.; van de Ville, D. On Spurious and Real Fluctuations of Dynamic Functional Connectivity during Rest. Neuroimage 2015, 104, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Gifford, G.; Crossley, N.; Kempton, M.J.; Morgan, S.; Dazzan, P.; Young, J.; McGuire, P. Resting State FMRI Based Multilayer Network Configuration in Patients with Schizophrenia. Neuroimage Clin 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, W.; Sun, Y.; Hu, M.; Chen, A. Aberrant Intra-Salience Network Dynamic Functional Connectivity Impairs Large-Scale Network Interactions in Schizophrenia. Neuropsychologia 2016, 93. [Google Scholar] [CrossRef] [PubMed]
- Sheng, D.; Pu, W.; Linli, Z.; Tian, G.L.; Guo, S.; Fei, Y. Aberrant Global and Local Dynamic Properties in Schizophrenia with Instantaneous Phase Method Based on Hilbert Transform. Psychol Med 2021. [Google Scholar] [CrossRef]
- Guo, S.; Zhao, W.; Tao, H.; Liu, Z.; Palaniyappan, L. The Instability of Functional Connectivity in Patients with Schizophrenia and Their Siblings: A Dynamic Connectivity Study. Schizophr Res 2018, 195, 183–189. [Google Scholar] [CrossRef]
- Hou, Z.; Kong, Y.; He, X.; Yin, Y.; Zhang, Y.; Yuan, Y. Increased Temporal Variability of Striatum Region Facilitating the Early Antidepressant Response in Patients with Major Depressive Disorder. Prog Neuropsychopharmacol Biol Psychiatry 2018, 85, 39–45. [Google Scholar] [CrossRef]
- Whitfield-Gabrieli, S.; Ford, J.M. Default Mode Network Activity and Connectivity in Psychopathology. Annu Rev Clin Psychol 2012, 8. [Google Scholar] [CrossRef]
- Wise, T.; Marwood, L.; Perkins, A.M.; Herane-Vives, A.; Joules, R.; Lythgoe, D.J.; Luh, W.M.; Williams, S.C.R.; Young, A.H.; Cleare, A.J.; et al. Instability of Default Mode Network Connectivity in Major Depression: A Two-Sample Confirmation Study. Transl Psychiatry 2017, 7. [Google Scholar] [CrossRef]
- Demirtaş, M.; Tornador, C.; Falcón, C.; López-Solà, M.; Hernández-Ribas, R.; Pujol, J.; Menchón, J.M.; Ritter, P.; Cardoner, N.; Soriano-Mas, C.; et al. Dynamic Functional Connectivity Reveals Altered Variability in Functional Connectivity among Patients with Major Depressive Disorder. Hum Brain Mapp 2016, 37, 2918–2930. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Zhang, S.; Mo, Z.; Chattun, M.R.; Wang, Q.; Wang, L.; Zhu, R.; Shao, J.; Wang, X.; Yao, Z.; et al. Antidepressants Normalize Brain Flexibility Associated with Multi-Dimensional Symptoms in Major Depressive Patients. Prog Neuropsychopharmacol Biol Psychiatry 2020, 100. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Cui, Q.; Wang, X.; Li, L.; Li, D.; He, Z.; Guo, X.; Fan, Y.S.; Guo, J.; Sheng, W.; et al. Resting State Functional Network Switching Rate Is Differently Altered in Bipolar Disorder and Major Depressive Disorder. Hum Brain Mapp 2020, 41, 3295–3304. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, B.; Yan, C.G. Reproducibility of R-FMRI Metrics on the Impact of Different Strategies for Multiple Comparison Correction and Sample Sizes. Hum Brain Mapp 2018, 39, 300–318. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Ouyang, X.; Huang, D.; Wu, Z.; Liu, Z.; He, Z.; Long, Y. Disrupted Intrinsic Functional Brain Network in Patients with Late-Life Depression: Evidence from a Multi-Site Dataset. J Affect Disord 2023, 323, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Kovacevic, S.; Dev, S.I.; Lu, K.; Liu, T.T.; Eyler, L.T. Dynamic Functional Connectivity in Bipolar Disorder Is Associated with Executive Function and Processing Speed: A Preliminary Study. Neuropsychology 2017, 31, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Jiang, X.; Zhu, W.; Shen, Y.; Xue, F.; Li, Y.; Chen, Z. Disturbances of Dynamic Function in Patients With Bipolar Disorder I and Its Relationship With Executive-Function Deficit. Front Psychiatry 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Huang, H.; Jia, Y.; Zheng, S.; Zhong, S.; Chen, G.; Huang, L.; Huang, R. Abnormal Dynamic Functional Network Connectivity in Unmedicated Bipolar and Major Depressive Disorders Based on the Triple-Network Model. Psychol Med 2020, 50. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Chen, G.; Jia, Y.; Zhong, S.; Gong, J.; Chen, F.; Wang, J.; Qi, Z.; Liu, X.; Huang, L.; et al. Shared and Specific Dynamics of Brain Segregation and Integration in Bipolar Disorder and Major Depressive Disorder: A Resting-State Functional Magnetic Resonance Imaging Study. J Affect Disord 2021, 280, 279–286. [Google Scholar] [CrossRef]
- Yang, J.; Ouyang, X.; Tao, H.; Pu, W.; Fan, Z.; Zeng, C.; Huang, X.; Chen, X.; Liu, J.; Liu, Z.; et al. Connectomic Signatures of Working Memory Deficits in Depression, Mania, and Euthymic States of Bipolar Disorder. J Affect Disord 2020, 274. [Google Scholar] [CrossRef]
- Harlalka, V.; Bapi, R.S.; Vinod, P.K.; Roy, D. Atypical Flexibility in Dynamic Functional Connectivity Quantifies the Severity in Autism Spectrum Disorder. Front Hum Neurosci 2019, 13. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wu, Z.; Liu, Z.; Liu, D.; Huang, D.; Long, Y. Acute Effect of Betel Quid Chewing on Brain Network Dynamics: A Resting-State Functional Magnetic Resonance Imaging Study. Front Psychiatry 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Schäfer, A.; Bassett, D.S.; Rausch, F.; Schweiger, J.I.; Bilek, E.; Erk, S.; Romanczuk-Seiferth, N.; Grimm, O.; Geiger, L.S.; et al. Dynamic Brain Network Reconfiguration as a Potential Schizophrenia Genetic Risk Mechanism Modulated by NMDA Receptor Function. Proc Natl Acad Sci U S A 2016, 113, 12568–12573. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Ouyang, X.; Liu, Z.; Chen, X.; Hu, X.; Lee, E.; Chen, E.Y.H.; Pu, W.; Shan, B.; Rohrbaugh, R.M. Associations among Suicidal Ideation, White Matter Integrity and Cognitive Deficit in First-Episode Schizophrenia. Front Psychiatry 2018, 9. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A summary of the steps for constructing dynamic networks and estimating the temporal stability of brain networks, as well as the relationships between temporal stability and other brain network measures.
Figure 1.
A summary of the steps for constructing dynamic networks and estimating the temporal stability of brain networks, as well as the relationships between temporal stability and other brain network measures.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



