
Submitted:
02 February 2023
Posted:
07 February 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) that caused the COVID-19 pandemic still able to infect the population in many countries. The Omicron strain is the most mutated variant of SARS-CoV-2. The high transmissibility of the strain and the ability to evade immunity require a priority study of its properties in order to quickly create effective means of preventing it. The present work is devoted to the study of in silico interaction of piRNAs with the genome of the SARS-CoV-2 (gRNA) in order to identify endogenous piRNAs and propose synthetic piRNAs with high antiviral activity for drug development. The studies were carried out using proven bioinformatic methods of interaction of the entire SARS-CoV-2 genome with more than eight million piRNAs. Binding sites (BSs) of piRNAs in the 5'UTR were located with overlapping nucleotide sequences called clusters of BSs. Several clusters of BSs were found in the nsp3, nsp7, RNA-dependent RNA polymerase, endoRNAse, S surface glycoprotein, ORF7a and nucleocapsid. 16 synthetic piRNAs have been proposed that interact with gRNA with free binding energy from -170 kJ/mol to -175 kJ/mol, which can be used to create drugs that suppress the reproduction of SARS-CoV-2.
Keywords:
Omicron
; SARS-CoV-2
; piRNAs
; coronavirus genome
; translation
; replication
1. Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) that caused the COVID-19 pandemic still able to infect the population in many countries. SARS-CoV-2 causes disease in humans and many vertebrates [1], which facilitates the study of the pathogenic effect of SARS-CoV-2 and the testing of drugs intended for humans. In addition to typical fever and respiratory symptoms, many patients with COVID-19 experience a variety of neurological complications and cancer [2,3,4,5]. SARS-CoV-2 is distributed in many strains: B.1.1.7 (Alpha), B.1.351 (Beta), B.1.525 (Eta), B.1.427/B.1.429 (Epsilon), B.1.526 (Iota), B.1.617.1 (Kappa), B.1.617.2 (Delta), C.37 (Lambda), P.1 (Gamma), P.2 (Zeta), P.3 (Theta) and the recently discovered B.1.1.529 (Omicron) [6], which forces the establishment of their features that require specific treatment methods. As an object of research, we chose the Omicron strain (B.1.1.529), which is the most mutated variant of SARS-CoV-2, and its high transmissibility and ability to evade immunity are of major concern.
Due to the absence of effective treatments for coronavirus strains, medications must be created that can both treat and stop the spread of infection. One of the properties of the coronavirus (gRNA) genome that contributes to its survival in animals lies in the structure of gRNA similar to the mRNA structure of protein-coding genes. Figure 1 shows that the gRNA nucleotide sequence contains 5’UTR, CDS, and 3’UTR, which allows it to be similar to the mRNA structure of many genes, including tens of thousands of nucleotides in length. A number of studies have examined the features of all functional regions of gRNA that could serve as the basis for the development of methods to prevent SARS-CoV-2. Since gRNA contains the 5’UTR, there have been attempts to elucidate the role of this region of SARS-CoV-2 in the pathogenicity of the coronavirus. The viral 5’UTRs are believed to provide reliable expression of viral mRNAs. These features can be used as potential therapeutic targets [7]. The 5’UTR of coronavirus genomes plays key roles in the viral replication cycle and translation of viral gRNA [8]. Several studies have shown that miRNA binding sites (BSs) are located in the 5’UTR, which can interfere with protein synthesis [9,10,11]. The effect of compounds on the 5’UTR of SARS-CoV-2 acting on the 5’UTR mRNAs of human protein-coding genes was studied [12,13,14,15]. The authors conclude that the combined NSP1-mediated degradation of spliced mRNAs and translation inhibition of single-exon genes, along with the unique features present in the viral 5’UTRs, ensure robust expression of viral mRNAs. These features can be exploited as potential therapeutic targets. The combined NSP1-mediated degradation of spliced mRNAs and inhibition of translation of single exon genes, along with unique features present in the viral 5’UTRs, mediate the expression of viral mRNAs. These features can be used as potential therapeutic targets [15]. The effect of the 5’UTR gRNA structure as a potential therapeutic target against SARS-CoV-2 has been studied [16,17,18,19]. The spike protein (S protein) encoded in gRNA has attracted the interest of researchers since it is thought to play a vital role in viral penetration into the recipient cell [6,20,21,22,23,24]. A number of publications have elucidated the role of S protein as a target for viral suppression [1,22,25,26,27]. Based on a number of studies, individual protein-coding regions of gRNA can be utilized autonomously to examine the effect of the piRNAs we identified that act within the NSP3 gene [28,29,30]. Regions encoding Nsp2, Nsp3, S1, and ORF8 [31], omicron S protein [32] have been studied. S protein, receptor-binding domain (RBD), nucleocapsid (N), orf3a, orf8, nsp3, nsp13, and membrane (M) antigenic specificities have been examined [33]. The expression and subcellular localization of 11 SARS-CoV-2 nonstructural replicase proteins: nsp1, nsp2, nsp3, nsp5, nsp7, nsp8, nsp9, nsp10, nsp13, nsp14, and nsp15 were systematically analyzed [34]. Understanding the process of replication and transcription of SARS-CoV-2 is essential for antiviral strategy development. The replicase polyprotein is indispensable for viral replication [35]. Papain-Like Proteases is a cysteine protease, essential for viral replication and proliferation, as a highly conserved enzyme it cleaves peptide linkage between Nsp1, Nsp2, Nsp3, and Nsp4. As valid therapeutic targets, it stops viral reproduction and boosts host [36]. Surprisingly, free Hb at 1mM inhibited viral replication (99%), and its interaction with SARS-CoV-2 was localized into the RBD region of the S protein. The authors in this study identified that five proteins (S, N, M, Nsp3, and Nsp7) of SARS-CoV-2 recruit Hb/metabolites [37]. The above analysis of the properties of gRNA regions shows that the 5’UTR and each of the protein-coding regions can be modified by various agents. However, for the most part, these agents are difficult to use to suppress the viability of the coronavirus in vivo.
More than 20 years ago, piRNAs (PIWI-interacting RNAs) were discovered that, due to a number of misconceptions, were not used as inhibitors of the protein translation process on mRNA [38]. A recently created piRNAs database provides data on over than eight million of these molecules [39]. The interaction of piRNAs with mRNA of human genes has been established in recent years [40,41]. It has been shown that piRNAs can interact with the gRNA of the Delta strain [42]. This information allowed us to suggest that piRNAs can influence the expression of protein-coding genes and participate in the suppression of SARS-CoV-2 reproduction in the human body. It must be borne in mind that synthetic piRNAs (spiRNAs) created at random, in addition to suppressing the reproduction of the virus, can suppress the expression of human genes, causing side effects. For this reason, numerous successful experiments on the suppression of coronavirus in cell culture and in experimental animals have not been brought to clinical use. This work is devoted to the in silico study of the interaction of piRNAs with the genome of the SARS-CoV-2 Omicron strain in order to identify endogenous piRNAs and propose spiRNAs for drug development. To elucidate this problem, we identified endogenous piRNAs capable of suppressing viral replication and, based on these piRNAs, propose spiRNAs.
Figure 1.
Localization of piRNA BSs clusters in gRNA of Omicron variant of SARS-CoV-2.

2. Materials and Methods
The nucleotide (nt) sequence of Omicron strain (B.1.1.529) of the SARS-COV-2 gRNA was downloaded from National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/). The nucleotide sequences of 8.426 million piRNAs were taken from Wang et al. [43]. The piRNA BSs in gRNA were predicted using the MirTarget program [44]. This program predicts the following features of piRNA binding to mRNA: (a) the initiation of piRNA binding to the gRNA from the first nucleotide of the gRNA; (b) the localization of the piRNA BSs in the 5′-untranslated region (5′UTR), coding domain sequence (CDS), and 3′-untranslated region (3′UTR) of the mRNAs; (c) the schemes of nucleotide interactions between piRNAs and gRNA; (d) the free energy of the interaction between piRNAs and the gRNA (ΔG, kJ/mol); and the ratio ΔG/ΔGm (%) is determined for each site (ΔGm equals the free energy of piRNA binding with its fully complementary canonical nucleotide sequence). Only piRNAs whose nucleotides interacted with gRNA using canonical (G-C and A-U) and non-canonical (G-U and A-C) nucleotides with a given ΔG value were selected from the calculated data. The MirTarget program finds hydrogen bonds between adenine (A) and uracil (U), guanine (G), and cytosine (C), G and U, and A and C. The free energy of interactions (ΔG) of a pair of G and C is equal to 6.37 kJ/mol, a pair of A and U is equal to 4.25 kJ/mol, G and U, A and C equal to 2.12 kJ/mol. The distances between bound A and C (1.04 nm) and G and U (1.02 nm) are similar to the distances between bound G and C, A and U, which are equal to 1.03 nm [45,46,47,48]. The numbers of hydrogen bonds in the G-C, A-U, G-U, and A-C interactions were 3, 2, 1, and 1, respectively. MirTarget differs from other programs in terms of finding piRNA BSs on gRNA in the following: it takes into account the interaction of piRNA with gRNA over the entire piRNA sequence; it considers non-canonical pairs G–U and A–C; and it calculates the free energy of the interaction of the piRNAs with gRNA.
3. Results
The first region of the gRNA nucleotide sequence that interacted with piRNAs was located at the 5’UTR (Figure 1). The BSs of four piRNAs form a cluster with partial overlap of nucleotides (Figure 2).
The overlap of the nucleotide sequences of piRNA BSs in gRNA leads to the competition of different piRNAs while suppressing the start of the process of translation of coronavirus proteins. Given that the genes encoding these piRNAs can be located in different parts of the human genome, in the case of mutations that disrupt the synthesis of one of these piRNAs, the body will retain the ability to defend itself against coronavirus. In addition to the above four piRNAs interacting with gRNA with a free energy from -130 kJ/mol and higher, in the 5’UTR BSs cluster at position 156 nt, piR-2388809 also bound with a ΔG value of -132 kJ/mol (Table S1). Therefore, by synthesizing endogenous piRNAs, a human has acquired the ability to protect himself from a deadly pathogen.
Figure 2.
Nucleotide sequences of piRNA BSs cluster in the 5’UTR of gRNA from 40 nt to 84 nt and nucleotide sequences of four piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.
Figure 2.
Nucleotide sequences of piRNA BSs cluster in the 5’UTR of gRNA from 40 nt to 84 nt and nucleotide sequences of four piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.

In the nsp3 protein-coding part of gRNA, we have identified several clusters of BSs for four or more piRNAs (Figure 1). Consider the properties of these clusters of BSs in the order of their location in gRNA. In the CDS region of gRNA from 4207 nt to 4241 nt, the BSs of four piRNAs were located (Figure 3). The BSs of these piRNAs were located with a partial overlap of nucleotides, that is, they formed a cluster of BSs. piR-3942773 and piR-4093935 interacted with gRNA with ΔG value of -138 kJ/mol and -136 kJ/mol (Table S1). Such free energy of interaction allows these piRNAs to be strongly attached to nsp3 gRNA and inhibit protein synthesis. The BSs cluster contained the ACUU tetranucleotide, which ensures the binding of the UGAA tetranucleotide of piRNAs using canonical base pairs. In the center of piR-3942773 and piR-4093935 was the CCCAGUCCCAAA oligonucleotide, which also bound to gRNA via canonical GGGUCAGGGUUU base pairs. In piR-1758885 and piR-962633, the UGAAUAGGUCCAAUUCUAGAUUU polynucleotide provided complementary interaction with gRNA through canonical and non-canonical base pairs almost along their entire length.
The third cluster of piRNA BSs from 7137 nt to 7161 nt was also located in the nsp3 protein-coding region of gRNA (Figure 4). A feature of this cluster is the content of a large number of piRNA BSs, all nucleotides of which fully complementary interacted with gRNA. Such a set of piRNAs is guaranteed to provide high human protection against coronavirus. Only a significant mutation in the cluster of gRNA BSs could increase the protection of the coronavirus against these endogenous piRNAs. With the exception of piR-193777, the remaining 13 piRNAs had the same oligonucleotide UUGACAUGUUUGAUGAUGGAG, which provides strong interaction with gRNA mainly due to canonical base pairs. The first four GGAG nucleotides from the 5’-end of piRNAs provided strong binding to gRNA through G-C pairs with three hydrogen bonds. The example of this BSs cluster shows that the human genome in the process of evolution created protection against SARS-CoV-2. It seems that it is better for the human genome to maintain a constant set of piRNAs than to frequently adapt to coronavirus mutations, although both ways of protecting against coronavirus take place. Further research on different strains of SARS-CoV-2 and other coronaviruses can be conducted to see whether this claim is true.
The BSs cluster of four piRNAs with gRNA is shown in Figure 5. All piRNAs interacted with gRNA through canonical and non-canonical base pairs. The nucleotide sequence of piR-65720 CGUGUUGUCUUGUUUUUUAGUU was identical to part of the nucleotide sequences of other piRNAs, except for the replacement of the U nucleotide with the C nucleotide at position 7478 of piR-8100036 (Figure 5). As a result, the identified four piRNAs can significantly suppress the synthesis of the protein encoded by the nsp3 region.
The largest number of piRNAs bound in the BSs cluster located from 12029 nt to 12055 nt in the nsp7 protein-coding region of gRNA (Figure 1). The nucleotide sequences of the BSs of 29 piRNAs were arranged so that the 5’ end of the piRNAs binds at the same gRNA position (12055 nt) despite their different binding positions from the 3’ end (Figure 6). This localization of BSs for all 29 piRNAs is not random and was due to the identity of the UGGAGU oligonucleotide of the 5’-end of piRNAs, which bound to gRNA only through canonical base pairs. Other conserved nucleotides in all piRNAs were GAUG and AUGUU, which also bound to gRNA with the formation of canonical base pairs.
All nucleotides of four piRNAs bound to gRNA to form canonical and non-canonical pairs, as shown in Figure 7. The BSs of these piRNAs formed a cluster from 13919 nt to 13646 nt in the gRNA encoding RNA-dependent RNA polymerase (Figure 1). The AGAGACUGACAGUUGUACCUUU oligonucleotide site was shared by four piRNAs, and termination of synthesis of any of the four piRNAs could be replaced by the remaining three piRNAs.
In the region from 20598 nt to 20624 nt of gRNA, there was a BSs cluster of eighteen piRNAs that inhibit endoRNAase synthesis (Figure 8). All these piRNAs contained the GAUCAGUUCGC oligonucleotide, which interacted with gRNA only through canonical base pairs. All piRNAs had the same UCGUUC hexanucleotide flanked by nucleotides that do not bind gRNA (Figure 8).
The next piRNA BSs cluster from 24308 nt to 24338 nt was located in the gRNA region encoding the S surface glycoprotein (Figure 1). All piRNAs, starting from the 5’-end, contained tetranucleotide CACGU, trinucleotide CGU, pentanucleotide AGGUG, and nanonucleotide GAGUGAAAG interacting to form only canonical base pairs (Figure 9). These seven piRNAs, ranging in length from 28 nt to 31 nt, bound to gRNA with a free energy from -130 kJ/mol to -138 kJ/mol (Table S1).
By replacing nucleotides forming non-canonical pairs with nucleotides forming canonical pairs in piRNAs and replacing non-interacting nucleotides, the resulting spiRNAs bound to gRNA with a free energy from -152 kJ/mol to -165 kJ/mol. Such a large value of ΔG is also associated with the length of piRNAs from 28 nt to 31 nt. These spiRNAs, fully complementary to gRNAs, are identical in function to widely used siRNAs that cause inhibition of viral RNA protein synthesis [49,50,51,52,53]. These examples demonstrate the possibility of creating drugs that suppress the reproduction of coronavirus in human cells.
The gRNA region from 27375 nt to 27399 nt encodes the ORF7a protein (Figure 1). Most of the nucleotides of the identified piRNAs formed canonical pairs, and in general, all nucleotides of these piRNAs formed hydrogen bonds with gRNA nucleotides (Figure 13). All piRNAs had an identical oligonucleotide ACACUCAAGAUGGUAACGGUUU from the 5’ end, which indicates almost complete interchangeability of piRNAs in the putative function of inhibition of ORF7a protein synthesis. piRNAs associated with gRNA in the cluster will interfere with the replication of the coronavirus genome. The localization of the cluster of piRNA BSs from 27375 nt to 27399 nt, that is close to the 3’UTR, will terminate erly gRNA replication that starts from the 3’UTR.
Figure 10.
Nucleotide sequences of six piRNA BSs cluster in the CDS of gRNA from 27375 nt to 27399 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. After the piRNAs name, the beginning of their BSs is indicated.
Figure 10.
Nucleotide sequences of six piRNA BSs cluster in the CDS of gRNA from 27375 nt to 27399 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. After the piRNAs name, the beginning of their BSs is indicated.

In the gRNA region encoding the N protein, a cluster of piRNA BSs from 28447 nt to 28471 nt was detected (Figure 1). All piRNAs from the 5’ end, starting from piR-252298, had an identical oligonucleotide CCGGUAAUAGCUUCU, and then the hexanucleotide GGUUUA was located, which bound only through canonical nucleotide pairs (Figure 11). The BSs of piR-806264 and piR-1125646 were located at positions 28360 nt and 28636 nt, respectively, from the 5’ end of the cluster and from the 3’ end of the cluster (Table S1). The interaction energy of these 33 nt piRNAs was -142 kJ/mol and -134 kJ/mol, respectively, indicating the importance of these piRNAs in the inhibition of N protein synthesis.
Figure 11.
Nucleotide sequences of 10 piRNA BSs cluster in the CDS of gRNA from 28447 nt to 28471 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.
Figure 11.
Nucleotide sequences of 10 piRNA BSs cluster in the CDS of gRNA from 28447 nt to 28471 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.

The last cluster of piRNA BSs was located in the region from 29280 nt to 29308 nt of gRNA, which also encodes the N protein (Figure 12). Before this BSs cluster at position 29030 nt was the piR-1134823 BS and after position 29377 nt was the piR-3158024 BS, both with an interaction energy of -130 kJ/mol (Table S1). That is, as in the previous cluster of BSs, this cluster of four piRNAs BSs “insured” by the binding of piRNAs before and after the BSs cluster. Four piRNAs contained identical 3’-GUGUAGGUUUAGUAAGGUCGGUUGU-5’ nucleotides, which indicates their almost complete interchangeability in the suppression of N protein synthesis.
Figure 12.
Nucleotide sequences of 4 piRNA BSs cluster of gRNA from 29280 nt to 29308 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.
Figure 12.
Nucleotide sequences of 4 piRNA BSs cluster of gRNA from 29280 nt to 29308 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.
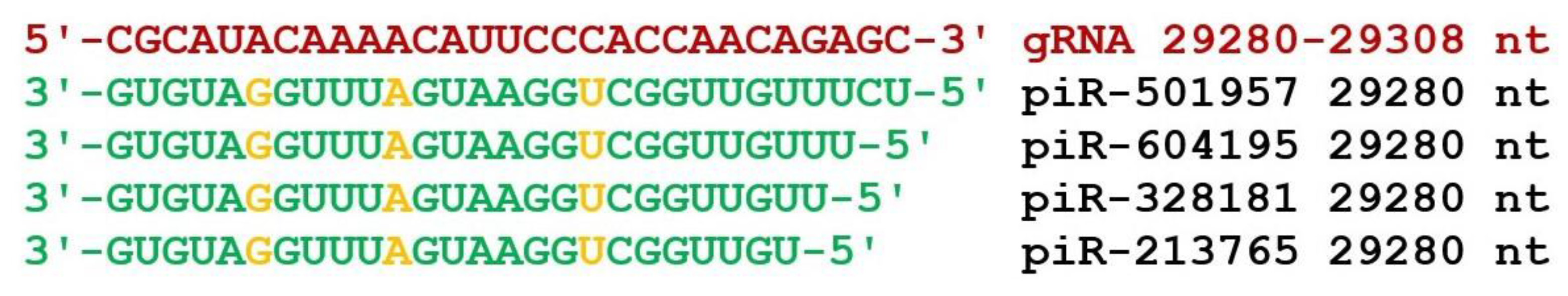
The clusters of piRNA BSs in gRNA that we have identified form the basis for selecting the most effective piRNAs that can be used to suppress SARS-COV-2 proliferation. From the list of piRNAs (Table S1) that interacted with gRNA with a free energy of -130 kJ/mol or more, we selected 16 piRNAs on the basis of which, by replacing nucleotides, we made the spiRNA nucleotide sequences that interact with gRNA completely complementary by forming canonical nucleotide pairs. Interaction patterns of spiRNAs and gRNA BSs are shown in Figure 13. Each of these spiRNAs will reliably block both synthesis of the protein encoded by the spiRNAs target and gRNA replication. The effectiveness of spiRNAs is expected to be the same as in many experiments using synthetic interfering siRNA molecules [49,50,51,52,53].
Figure 13.
Nucleotide sequences of spiRNAs and characteristics of their interaction in gRNA regions. Note: The gRNA nucleotides of the cluster of spiRNA BSs are highlighted in red. Nucleotides of spiRNAs forming canonical base pairs with gRNA are highlighted in blue.
Figure 13.
Nucleotide sequences of spiRNAs and characteristics of their interaction in gRNA regions. Note: The gRNA nucleotides of the cluster of spiRNA BSs are highlighted in red. Nucleotides of spiRNAs forming canonical base pairs with gRNA are highlighted in blue.

4. Discussion
Coronavirus has been found in many animal organisms, which indicates the early occurrence of their protective methods to fight this pathogen [54,55,56]. It is obvious that the low mortality of animal organisms is due to the creation in the process of evolution of endogenous substances that prevent high lethality from coronavirus. At present, there is information about the involvement of piRNAs in the regulation of the expression of protein-coding genes [42,57]. Since gRNA has features of the mRNA structure, that is, 5’UTR, CDS and 3’UTR, it is logical to assume that the synthesis of proteins based on gRNA as a template can also be regulated by piRNAs. The possibility of the influence of piRNAs on the synthesis of coronavirus proteins was previously shown [42]. In this work, we show the possibility of piRNAs affecting protein synthesis by binding piRNAs to the gRNA of Omicron strain of SARS-CoV-2 genome.
Of the 8 426 000 piRNAs, 92 piRNAs could bind to gRNA with a value of -130 kJ/mol or more (Table S1). These piRNAs bound fully complementary through the interaction of canonical and non-canonical base pairs. The length of these piRNAs varied from 28 nt to 34 nt, indicating a strong interaction of piRNAs with gRNA. The chosen selection criteria for piRNAs strongly interacting with gRNA made it possible to identify the gRNA regions to which two or more piRNAs bound with overlapping nucleotide sequences of BSs. Such regions, which we called clusters of miRNA and piRNA BSs, indicate that the corresponding piRNAs can more effectively suppress both protein synthesis on mRNA and gRNA and gRNA replication [58,59,60,61,62,63]. The strong interaction of several piRNAs in the gRNA BSs cluster makes it possible to more reliably suppress the reproduction of the coronavirus, since a decrease in the concentration of one of the piRNAs will not significantly affect the inhibitory effect of the piRNAs group. Some piRNAs bound to mRNAs of human protein-coding genes, and diversion of such piRNAs would facilitate the replication of the coronavirus. Based on this, it is required to control the concentration of those piRNAs in cells and the body that inhibit the reproduction of coronavirus. It is logical that the human body produces many piRNAs that can suppress the coronavirus in order to reliably protect the body from this pathogen. It is logical that human produces many piRNAs that can suppress the coronavirus in order to reliably protect the body from this pathogen. Let us consider the features of the interaction of piRNAs with gRNA to understand why BSs clusters are located in different regions of gRNA. The gRNA of the coronavirus, like protein-coding genes, contains 5’UTR, CDS and 3’UTR (Figure 1). The first cluster of piRNA BSs with gRNA was located in the 5’UTR. Figure 1 shows how piRNAs can interact with gRNA. This localization of the piRNA BSs cluster in the 5’UTR immediately prevents the binding of ribosomes to gRNA and blocks the synthesis of coronavirus proteins. The location of the piRNA BSs cluster allows the use of several spiRNAs based on these piRNAs that will fully complementary bind in this cluster and will reliably suppress the synthesis of encoded gRNA proteins.
Single nucleotide mutations in BS clusters cannot significantly protect the coronavirus from piRNAs. However, substitutions of three or four nucleotides in the piRNA BSs cluster with a length of 30-34 nucleotides can significantly reduce their impact on coronavirus reproduction. Therefore, during the evolution of animals, they selected longer piRNAs to protect against coronavirus. For this reason, miRNAs, which are much shorter than piRNAs, are less used by animals to protect against coronavirus. The clusters of long piRNA BSs in gRNA that we found are an indicator of the protective function of the body.
In addition to BSs clusters of long piRNAs in gRNA, we found BSs clusters for 24–28 nt long piRNAs. Dozens of piRNAs of this length can bind in these clusters. The best example of this is the cluster of piRNA BSs in the gRNA region from 12029 nt to 12055 nt (Figure 6). Such a number of piRNAs is guaranteed to be able to suppress the synthesis of the protein that is involved in the formation of the coronavirus envelope. A similar cluster of piRNA BSs was found in the region from 7137 nt to 7161 nt (Figure 4). This region is included in the nucleotides encoding the nsp3 protein (Figure 1). In this cluster, all nucleotides of 14 piRNAs were involved in the formation of hydrogen bonds with gRNA. In addition to this BSs cluster, two more BSs clusters were found in the nsp3 protein gene (Figure 3 and Figure 5). In the cluster of BSs from 7467 nt to 7492 nt, all piRNA nucleotides also interact with gRNA (Figure 5). In the second half of the gRNA, nucleotide sequence from 20598 nt to 20624 nt there was a large cluster of 18 piRNAs BSs (Figure 6), which can be an effective target for piRNAs. In addition to the clusters of piRNA BSs described above, several more clusters were identified for a smaller number of piRNAs (Figure 1). The results obtained indicate a developed system for protecting the human body from coronavirus using piRNAs. However, not all body cells synthesize the entire set of eight million piRNAs. Therefore, coronaviruses manage to multiply in body cells in which the set of antiviral piRNAs is small or absent. Information about antiviral piRNAs in such cells is very important because such cells can be targeted to protect them. Exosomes and vesicles in which miRNAs and piRNA are carried in the body are 30–200 nm in size [64,65]. In this regard, it is necessary to know in which tissues and cells a low level of expression of antiviral piRNAs is observed, at least among the most effective piRNAs.
5. Conclusions
The data obtained in this work on the effect of piRNAs on coronavirus can be used to predict the likelihood of infection with coronavirus and in the development of drugs based on endogenous and spiRNAs. It is logical to create spiRNAs based on piRNAs 30–34 nt long that interact with gRNAs with high free energy and bind in the gRNA regions encoding the most important protein components of the coronavirus. At the same time, these highly binding piRNAs will also interfere with gRNA replication.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Characteristics of human piRNA interaction with gRNA SARS-CoV-2 omicron stain.
Author Contributions
Conceptualization, A.I. and A.R.; methodology, A.I., A.P., P.Z.; software, A.P., A.I.; validation, A.R.; investigation, T.N., A.R., A.A.; resources, A.R.; data curation, A.P., S.K., A.K., A.-G.R.; writing—original draft preparation, A.R., A.I., P.Z.; writing—review and editing, A.A., T.N; visualization, A.R., T.N.; supervision, A.I., P.Z.; project administration, P.Z., A.I.; funding acquisition, P.Z. All authors have read and agreed to the published version of the manuscript.
Funding
Polish Ministry of Science and Higher Education, under the project: DIR/WK/2018/06.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wei, C.; Shan, K.J.; Wang, W.; Zhang, S.; Huan, Q.; Qian, W. Evidence for a mouse origin of the SARS-CoV-2 Omicron variant. J Genet Genomics. 2021, 48, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.W.; Xu, H.S.; Liu, S.J. COVID-19 and neurodegenerative diseases. Eur Rev Med Pharmacol Sci. 2022, 26, 4535–4544. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Bhushan, B.; Maurya, A.; Mishra, G.; Singh, S.K.; Awasthi, R. Novel coronavirus disease 2019 (COVID-19) and neurodegenerative disorders. Dermatol. Ther. 2020, 33, e13591. [Google Scholar] [CrossRef]
- Angelini, M.; Teglia, F.; Astolfi, L.; Casolari, G.; Boffetta, P. Decrease of cancer diagnosis during COVID-19 pandemic: a systematic review and meta-analysis. Eur. J. Epidemiol. 2023, 2023, 1–8. [Google Scholar] [CrossRef]
- Alrahawy, M.; Johnson, C.; Aker, M.; Eltyeb, H.A.; Green, S. Impact of COVID-19 on the Mode of Presentation and Stage at Diagnosis of Colorectal Cancer. Cureus. 2022, 14, e32037. [Google Scholar] [CrossRef]
- Ghosh, N.; Nandi, S.; Saha, I. A review on evolution of emerging SARS-CoV-2 variants based on spike glycoprotein. Int Immunopharmacol. 2022, 105, 108565. [Google Scholar] [CrossRef]
- Melidis, L.; Hill, H.J.; Coltman, N.J.; Davies, S.P.; Winczura, K.; Chauhan, T.; Craig, J.S.; Garai, A.; Hooper, C.A.J.; Egan, R.T.; McKeating, J.A.; Hodges, N.J.; Stamataki, Z.; Grzechnik, P.; Hannon, M.J. Supramolecular Cylinders Target Bulge Structures in the 5′ UTR of the RNA Genome of SARS-CoV-2 and Inhibit Viral Replication. Angew. Chem. Int. Ed. 2021, 60, 18144–18151. [Google Scholar] [CrossRef]
- Sosnowski, P.; Tidu, A.; Eriani, G.; Westhof, E.; Martin, F. Correlated sequence signatures are present within the genomic 5’UTR RNA and NSP1 protein in coronaviruses. RNA. 2022, 28, 729–741. [Google Scholar] [CrossRef]
- Xu, T.; Li, L.X.; Jia, Y.; Wu, Q.; Zhu, W.; Xu, Z.; Zheng, B.; Lu, X. One microRNA has the potential to target whole viral mRNAs in a given human coronavirus. Front Microbiol. 2022, 13, 1035044. [Google Scholar] [CrossRef]
- Li, C.; Wang, R.; Wu, A.; Yuan, T.; Song, K.; Bai, Y.; Liu, X. SARS-COV-2 as potential microRNA sponge in COVID-19 patients. BMC Med Genomics. 2022, 15, 94. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.F.; Ackerson, B.K.; Luo, Y.; Sy, L.S.; Talarico, C.A.; Tian, Y.; Bruxvoort, K.J.; Tubert, J.E.; Florea, A.; Ku, J.H.; Lee, G.S.; Choi, S.K.; Takhar, H.S.; Aragones, M.; Qian, L. Effectiveness of mRNA-1273 against SARS-CoV-2 Omicron and Delta variants. Nat Med. 2022, 28, 1063–1071. [Google Scholar] [CrossRef]
- Yao, Y.; Sun, H.; Chen, Y.; Tian, L.; Huang, D.; Liu, C.; Zhou, Y.; Wang, Y.; Wen, Z.; Yang, B.; Chen, X.; Pei, R. RBM24 inhibits the translation of SARS-CoV-2 polyproteins by targeting the 5’-untranslated region. Antiviral Res. 2022, 209, 105478. [Google Scholar] [CrossRef]
- Dhorne-Pollet, S.; Fitzpatrick, C.; Da Costa, B.; Bourgon, C.; Eléouët, J.F.; Meunier, N.; Burzio, V.A.; Delmas, B.; Barrey, E. Antisense oligonucleotides targeting ORF1b block replication of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Front Microbiol. 2022, 13, 915202. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.; Scott, G.; Calderon, M.; Haghighi, A.P. N6-Adenosine Methylation of SARS-CoV-2 5’-UTR Regulates Translation. bioRxiv. 2022, 2022.10.17.512569. [Google Scholar] [CrossRef]
- Condé, L.; Allatif, O.; Ohlmann, T.; de Breyne, S. Translation of SARS-CoV-2 gRNA Is Extremely Efficient and Competitive despite a High Degree of Secondary Structures and the Presence of an uORF. Viruses. 2022, 14, 1505. [Google Scholar] [CrossRef]
- Bignon, E.; Miclot, T.; Terenzi, A.; Barone, G.; Monari, A. Structure of the 5’ untranslated region in SARS-CoV-2 genome and its specific recognition by innate immune system via the human oligoadenylate synthase 1. Chem Commun (Camb). 2022, 58, 2176–2179. [Google Scholar] [CrossRef]
- Vora, S.M.; Fontana, P.; Mao, T.; Leger, V.; Zhang, Y.; Fu, T.M.; Lieberman, J.; Gehrke, L.; Shi, M.; Wang, L.; Iwasaki, A.; Wu, H. Targeting stem-loop 1 of the SARS-CoV-2 5’ UTR to suppress viral translation and Nsp1 evasion. Proc Natl Acad Sci U S A. 2022, 119, e2117198119. [Google Scholar] [CrossRef]
- Garcia-Moran, E.; Hernández, M.; Abad, D.; Eiros, J.M. Putative Secondary Structure at 5’UTR as a Potential Antiviral Target against SARS-CoV-2. Rev Esp Quimioter. 2022, 35, 204–212. [Google Scholar] [CrossRef]
- Slobodin, B.; Sehrawat, U.; Lev, A.; Hayat, D.; Zuckerman, B.; Fraticelli, D.; Ogran, A.; Ben-Shmuel, A.; Bar-David, E.; Levy, H.; Ulitsky, I.; Dikstein, R. Cap-independent translation and a precisely located RNA sequence enable SARS-CoV-2 to control host translation and escape anti-viral response. Nucleic Acids Res. 2022, 50, 8080–8092. [Google Scholar] [CrossRef]
- Thakur, V.; Ratho, R.K. OMICRON (B.1.1.529): A new SARS-CoV-2 variant of concern mounting worldwide fear. J Med Virol. 2022, 94, 1821–1824. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Thambiraja, T.S.; Karuppanan, K.; Subramaniam, G. Omicron and Delta variant of SARS-CoV-2: A comparative computational study of spike protein. J Med Virol. 2022, 94, 1641–1649. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Q.; Liang, Z.; Li, T.; Liu, S.; Cui, Q.; Nie, J.; Wu, Q.; Qu, X.; Huang, W.; Wang, Y. The significant immune escape of pseudotyped SARS-CoV-2 variant Omicron. Emerg Microbes Infect. 2022, 11, 1–5. [Google Scholar] [CrossRef]
- Saxena, S.K.; Kumar, S.; Ansari, S.; Paweska, J.T.; Maurya, V.K.; Tripathi, A.K.; Abdel-Moneim, A.S. Characterization of the novel SARS-CoV-2 Omicron (B.1.1.529) variant of concern and its global perspective. J Med Virol. 2022, 94, 1738–1744. [Google Scholar] [CrossRef]
- Papanikolaou, V.; Chrysovergis, A.; Ragos, V.; Tsiambas, E.; Katsinis, S.; Manoli, A.; Papouliakos, S.; Roukas, D.; Mastronikolis, S.; Peschos, D.; Batistatou, A.; Kyrodimos, E.; Mastronikolis, N. From delta to Omicron: S1-RBD/S2 mutation/deletion equilibrium in SARS-CoV-2 defined variants. Gene. 2022, 814, 146134. [Google Scholar] [CrossRef]
- Tian, D.; Sun, Y.; Xu, H.; Ye, Q. The emergence and epidemic characteristics of the highly mutated SARS-CoV-2 Omicron variant. J Med Virol. 2022, 94, 2376–2383. [Google Scholar] [CrossRef]
- Ma, W.; Yang, J.; Fu, H.; Su, C.; Yu, C.; Wang, Q.; de Vasconcelos, A.T.R.; Bazykin, G.A.; Bao, Y.; Li, M. Genomic Perspectives on the Emerging SARS-CoV-2 Omicron Variant. Genomics Proteomics Bioinformatics. 2022, 20, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Liu, B.; Li, F. Cryo-EM structure of a SARS-CoV-2 omicron spike protein ectodomain. Nat Commun. 2022, 13, 1214. [Google Scholar] [CrossRef] [PubMed]
- Rainville, C.; Sterner, D.E.; Suresh, K. A Method to Monitor Activity of SARS-CoV-2 Nsp3 from Cells. Methods Mol Biol. 2023, 2591, 269–282. [Google Scholar] [CrossRef]
- Gahbauer, S.; Correy, G.J.; Schuller, M.; Ferla, M.P.; Doruk, Y.U.; Rachman, M.; Wu, T.; Diolaiti, M.; Wang, S.; Neitz, R.J.; Fearon, D.; Radchenko, D.; Moroz, Y.; Irwin, J.J.; Renslo, A.R.; Taylor, J.C.; Gestwicki, J.E.; von Delft, F.; Ashworth, A.; Ahel, I.; Shoichet, B.K.; Fraser, J.S. Structure-based inhibitor optimization for the Nsp3 Macrodomain of SARS-CoV-2. bioRxiv 2022, 06.27.497816. [Google Scholar] [CrossRef]
- Diebold, O.; Gonzalez, V.; Venditti, L.; Sharp, C.; Blake, R.A.; Tan, W.S.; Stevens, J.; Caddy, S.; Digard, P.; Borodavka, A.; Gaunt, E. Using Species a Rotavirus Reverse Genetics to Engineer Chimeric Viruses Expressing SARS-CoV-2 Spike Epitopes. J Virol. 2022, 96, e0048822. [Google Scholar] [CrossRef]
- Akaishi, T.; Fujiwara, K.; Ishii, T. Insertion/deletion hotspots in the Nsp2, Nsp3, S1, and ORF8 genes of SARS-related coronaviruses. BMC Ecol Evol. 2022, 22, 123. [Google Scholar] [CrossRef]
- Zhao, L.P.; Lybrand, T.P.; Gilbert, P.B.; Payne, T.H.; Pyo, C.W.; Geraghty, D.E.; Jerome, K.R. Rapidly identifying new coronavirus mutations of potential concern in the Omicron variant using an unsupervised learning strategy. Sci Rep. 2022, 12, 19089. [Google Scholar] [CrossRef]
- Peddireddy, S.P.; Rahman, S.A.; Cillo, A.R.; Vijay, G.M.; Somasundaram, A.; Workman, C.J.; Bain, W.; McVerry, B.J.; Methe, B.; Lee, J.S.; Ray, P.; Ray, A.; Bruno, T.C.; Vignali, D.A.A.; Kitsios, G.D.; Morris, A.; Singh, H.; Sarkar, A.; Das, J. Antibodies targeting conserved non-canonical antigens and endemic coronaviruses associate with favorable outcomes in severe COVID-19. Cell Rep. 2022, 39, 111020. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.S.; Yu, Y.; Li, Y.L.; Cui, L.; Zhao, Z.; Wang, M.; Wang, B.; Zhang, R.; Huang, Y.W. Expression Profile and Localization of SARS-CoV-2 Nonstructural Replicase Proteins in Infected Cells. Microbiol Spectr. 2022, 10, e0074422. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Ouyang, M.; Yu, T.; Zhuang, J.; Wang, W.; Liu, X.; Duan, F.; Guo, D.; Peng, X.; Pan, J.A. Genome-Wide Analysis of the Indispensable Role of Non-structural Proteins in the Replication of SARS-CoV-2. Front Microbiol. 2022, 13, 907422. [Google Scholar] [CrossRef] [PubMed]
- Jupudi, S.; Rajagopal, K.; Murugesan, S.; Kumar, B.K.; Raman, K.; Byran, G.; Chennaiah, J.; Muthiah, V.P.; Dasan, P.B.; Sankaran, S.S. Identification of Papain-Like Protease inhibitors of SARS CoV-2 through HTVS, Molecular docking, MMGBSA and Molecular dynamics approach. Afr J Bot. 2021, 151, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Lechuga, G.C.; Souza-Silva, F.; Sacramento, C.Q.; Trugilho, M.R.O.; Valente, R.H.; Napoleão-Pêgo, P.; Dias, S.S.G.; Fintelman-Rodrigues, N.; Temerozo, J.R.; Carels, N.; Alves, C.R.; Pereira, M.C.S.; Provance, D.W., Jr.; Souza, T.M.L.; De-Simone, S.G. SARS-CoV-2 Proteins Bind to Hemoglobin and Its Metabolites. Int J Mol Sci. 2021, 22, 9035. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.N. Small RNAs just got bigger: Piwi-interacting RNAs (piRNAs) in mammalian testes. Genes Dev. 2006, 20, 1993–1997. [Google Scholar] [CrossRef]
- Wang, J.; Shi, Y.; Zhou, H.; Zhang, P.; Song, T.; Ying, Z.; Yu, H.; Li, Y.; Zhao, Y.; Zeng, X.; He, S.; Chen, R. piRBase: integrating piRNA annotation in all aspects. Nucleic Acids Res, 2021, 50, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Belkozhayev, A.; Niyazova, R.; Wilson, C.; Jainakbayev, N.; Pyrkova, A.; Ashirbekov, Y.; Akimniyazova, A.; Sharipov, K.; Ivashchenko, A. Bioinformatics Analysis of the Interaction of miRNAs and piRNAs with Human mRNA Genes. Having di- and Trinucleotide Repeats. Genes, 2022, 13, 800. [Google Scholar] [CrossRef]
- Akimniyazova, A.N.; Niyazova, T.K.; Yurikova, O.Y.; Pyrkova, A.Y.; Zhanuzakov, M.A.; Ivashchenko, A.T. piRNAs may regulate expression of candidate genes of esophageal adenocarcinoma. Front. Genet. 2022, 13, 1069637. [Google Scholar] [CrossRef]
- Akimniyazova, A.; Yurikova, O.; Pyrkova, A.; Rakhmetullina, A.; Niyazova, T.; Ryskulova, A.; Ivashchenko, A. In Silico Study of piRNA Interactions with the SARS-CoV-2 Genome. Int J Mol Sci. 2022, 23, 9919. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, P.; Lu, Y.; Li, Y.; Zheng, Y.; Kan, Y.; Chen, R.; He, S. piRBase: A comprehensive database of piRNA sequences. Nucleic Acids Res. 2018, 47, 175–180. [Google Scholar] [CrossRef]
- Ivashchenko, A.; Berillo, O.; Pyrkova, A.; Niyazova, R.; and Atambayeva, S. MiR-3960 binding sites with mRNA of human genes. Bioinformation 2014, 10, 423–427. [Google Scholar] [CrossRef]
- Friedman, R.A.; Honig, B.A. A free energy analysis of nucleic acid base stacking in aqueous solution. Biophys. J. 1995, 69, 1528–1535. [Google Scholar] [CrossRef] [PubMed]
- Kool, E.T. Hydrogen Bonding, Base Stacking, and Steric Effects in DNA Replication. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Heinemann, U.A. A novel form of RNA double helix based on G·U and C·A+ wobble base pairing. RNA 2018, 24, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Leontis, N.B.; Stombaugh, J.; Westhof, E. The non-watson-crick base pairs and their associated isostericity matrices. Nucleic Acids Res. 2002, 30, 3497–3531. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.D. A Brief History of the Discovery of RNA-Mediated Antiviral Immune Defenses in Vector Mosquitos. Microbiol Mol Biol Rev. 2022, e0019121. [Google Scholar] [CrossRef]
- Yang, Q.; Tang, J.; Cao, J.; Liu, F.; Fu, M.; Xue, B.; Zhou, A.; Chen, S.; Liu, J.; Zhou, Y.; Shi, Y.; Peng, W.; Chen, X. SARS-CoV-2 infection activates CREB/CBP in cellular cyclic AMP-dependent pathways. J Med Virol. 2022. [Google Scholar] [CrossRef]
- Lin, M.H.; Li, D.; Tang, B.; Li, L.; Suhrbier, A.; Harrich, D. Defective Interfering Particles with Broad-Acting Antiviral Activity for Dengue, Zika, Yellow Fever, Respiratory Syncytial and SARS-CoV-2 Virus Infection. Microbiol Spectr. 2022, e0394922. [Google Scholar] [CrossRef]
- Casseb, S.M.M.; Khayat, A.S.; de Souza, J.E.S.; de Oliveira, E.H.C.; Dos Santos, S.E.B.; da Costa Vasconcelos, P.F.; de Assumpção, P.P. Anticipating the Next Chess Move: Blocking SARS-CoV-2 Replication and Simultaneously Disarming Viral Escape Mechanisms. Genes, 2022, 13, 2147. [Google Scholar] [CrossRef]
- Sarangi, M.K.; Padhi, S.; Rath, G.; Nanda, S.S.; Yi, D.K. Success of nano-vaccines against COVID-19: a transformation in nanomedicine. Expert Rev Vaccines 2022, 21, 1739–1761. [Google Scholar] [CrossRef]
- Lee, C.Y.; Lowen, A.C. Animal models for SARS-CoV-2. Curr Opin Virol 2021, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Cleary, S.J.; Pitchford, S.C.; Amison, R.T.; Carrington, R.; Robaina Cabrera, C.L.; Magnen, M.; Looney, M.R.; Gray, E.; Page, C.P. Animal models of mechanisms of SARS-CoV-2 infection and COVID-19 pathology. Br J Pharmacol. 2020, 177, 4851–4865. [Google Scholar] [CrossRef] [PubMed]
- Frazzini, S.; Amadori, M.; Turin, L.; Riva, F. SARS CoV-2 infections in animals, two years into the pandemic. Arch Virol. 2022, 167, 2503–2517. [Google Scholar] [CrossRef] [PubMed]
- Kamenova, S.; Sharapkhanova, A.; Akimniyazova, A.; Kuzhybayeva, K.; Kondybayeva, A.; Rakhmetullina, A.; Pyrkova, A.; Ivashchenko, A. piRNA and miRNA can suppress the expression of multiple sclerosis candidate genes. Nanomaterials 2022, 13, 22. [Google Scholar] [CrossRef] [PubMed]
- Kondybayeva, A.; Akimniyazova, A.N.; Kamenova, S.U.; Ivashchenko, A. The characteristics of miRNA binding sites in mRNA of ZFHX3 gene And its orthologs. Vavilov J. Genet. Breed. 2018, 22, 438–444. [Google Scholar] [CrossRef]
- Aisina, D.; Niyazova, R.; Atambayeva, S.; Ivashchenko, A. Prediction of clusters of miRNA binding sites in mRNA candidate genes of breast cancer subtypes. PeerJ 2019, 7, e8049. [Google Scholar] [CrossRef] [PubMed]
- Kondybayeva, A.; Akimniyazova, A.; Kamenova, S.; Duchshanova, G.; Aisina, D.; Goncharova, A.; Ivashchenko, A. Prediction of miRNA interaction with mRNA of stroke candidate genes. Neurol. Sci. 2019, 41, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Mukushkina, D.; Aisina, D.; Pyrkova, A.; Ryskulova, A.; Labeit, S.; Ivashchenko, A. In silico Prediction of miRNA Interactions With Candidate Atherosclerosis Gene mRNAs. Front. Genet. 2020, 11, 605054. [Google Scholar] [CrossRef] [PubMed]
- Kamenova, S.; Aralbayeva, A.; Kondybayeva, A.; Akimniyazova, A.; Pyrkova, A.; Ivashchenko, A. Evolutionary Changes in the Interaction of miRNA With mRNA of Candidate Genes for Parkinson’s Disease. Front. Genet. 2021, 12, 647288. [Google Scholar] [CrossRef] [PubMed]
- Akimniyazova, A.; Pyrkova, A.; Uversky, V.; Ivashchenko, A. Predicting Associations of miRNAs and Candidate Gastric Cancer Genes for Nanomedicine. Nanomaterials 2021, 11, 691. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory Mechanisms and Intercellular Transfer of MicroRNAs in Living Cells. JBC. 2010, 285, 17442–17452. [Google Scholar] [CrossRef]
- Urbanelli, L.; Magini, A.; Buratta, S.; Brozzi, A.; Sagini, K.; Polchi, A.; Tancini, B.; Emiliani, C. Signaling Pathways in Exosomes Biogenesis, Secretion and Fate. Genes 2013, 4, 152–170. [Google Scholar] [CrossRef]
Figure 3.
Nucleotide sequences of piRNA BSs cluster in the CDS of gRNA from 4207 nt to4241 nt and nucleotide sequences of four piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.
Figure 3.
Nucleotide sequences of piRNA BSs cluster in the CDS of gRNA from 4207 nt to4241 nt and nucleotide sequences of four piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.

Figure 4.
Nucleotide sequences of piRNA BSs cluster in the CDS of gRNA from 7137 nt to 7161 nt and nucleotide sequences of four piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. After the piRNAs name, the beginning of their BSs is indicated.
Figure 4.
Nucleotide sequences of piRNA BSs cluster in the CDS of gRNA from 7137 nt to 7161 nt and nucleotide sequences of four piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. After the piRNAs name, the beginning of their BSs is indicated.

Figure 5.
Nucleotide sequences of four piRNA BSs cluster in the region from 7467 nt to 7492 nt of gRNA and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. After the piRNAs name, the beginning of their BSs is indicated.
Figure 5.
Nucleotide sequences of four piRNA BSs cluster in the region from 7467 nt to 7492 nt of gRNA and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. After the piRNAs name, the beginning of their BSs is indicated.

Figure 6.
Nucleotide sequences of 29 piRNA BSs cluster in the CDS of gRNA from 12029 nt to 12055 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.
Figure 6.
Nucleotide sequences of 29 piRNA BSs cluster in the CDS of gRNA from 12029 nt to 12055 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.

Figure 7.
Nucleotide sequences of four piRNA BSs cluster in the region of gRNA from 13919 nt to 13646 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. After the piRNAs name, the beginning of their BSs is indicated.
Figure 7.
Nucleotide sequences of four piRNA BSs cluster in the region of gRNA from 13919 nt to 13646 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. After the piRNAs name, the beginning of their BSs is indicated.

Figure 8.
Nucleotide sequences of piRNA BSs cluster in the region of gRNA from 20598 nt to 20624 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNA names, the beginning of their BSs is indicated.
Figure 8.
Nucleotide sequences of piRNA BSs cluster in the region of gRNA from 20598 nt to 20624 nt and nucleotide sequences of piRNAs. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNA names, the beginning of their BSs is indicated.

Figure 9.
Nucleotide sequences of seven piRNA BSs cluster in the CDS of gRNA from 24308 nt to 24338 nt and piRNA nucleotide sequences. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.
Figure 9.
Nucleotide sequences of seven piRNA BSs cluster in the CDS of gRNA from 24308 nt to 24338 nt and piRNA nucleotide sequences. Note: The gRNA nucleotides of the piRNA BSs cluster are highlighted in red. Nucleotides of piRNAs forming canonical and non-canonical pairs with gRNA are highlighted in green. piRNA nucleotides that do not interact with gRNA are highlighted in orange. After the piRNAs name, the beginning of their BSs is indicated.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



