
Submitted:
07 February 2023
Posted:
08 February 2023
You are already at the latest version
Abstract
French Guiana mosquito fauna diversity encompasses 242 species, of which nearly half of them belong to Culex (Diptera: Culicidae) genus. Whereas several species from Culex genus are important vectors of arbovirus, scarce studies focus on Culex genus, notably due to the inaccurate morphological identification of field-caught females gender. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) has been reported as a promising method for the identification of mosquitoes. Culex females from French Guiana field were dissected based on morphological classification. Abdomens were used for molecular identification using COI (cytochrome oxydase 1) gene. Legs and thorax of 169 specimens belonging to 13 Culex species,( i.e. Cx. declarator, Cx. nigripalpus, Cx. quinquefasciatus, Cx. usquatus, Cx. adamesi, Cx. dunni, Cx. eastor, Cx. idottus, Cx. pedroi, Cx. phlogistus, Cx. portesi, Cx. rabanicolus and Cx. spissipes) were then submitted to MALDI-TOF MS analysis. A high intra-species reproducibility and inter-species specificity of MS spectra for each mosquito body part tested were noticed. A corroboration of the specimen identification was revealed between MALDI-TOF MS, morphological and molecular results. MALDI-TOF MS protein profiling proves to be a suitable tool for identification of neotropical Culex species and will permit to enhance knowledge on this highly diverse Culex mosquitoes genus.
Keywords:
Culex species
; neotropical mosquito
; French Guiana
; MALDI-TOF MS
; proteomic tool
; arbovirus
; monitoring
Introduction
The neotropical region is inventoried as a hotspot of mosquito (Diptera: Culicidae) species richness. French Guiana (hereafter FG) is a department of France situated in South America which harbors one of the highest relative species density of mosquitoes in the world [1,2]. Currently, 242 mosquito species, classified into 22 genera are known in this French territory [3]. Some species are medically important because they are proven vectors of human pathogens. For example, several Anopheles species are vectors of malaria parasitic infection in FG [4,5,6,7]. Yellow Fever (YFV), dengue (DENV), chikungunya (CHIKV) and Zika (ZIKV) viruses are transmitted by the day-biting Aedes (Stegomyia) aegypti (Linnaeus) in urban area [8,9,10].
Containing 788 species classified into 26 subgenera worldwide, the genus Culex (abbreviated Cx.) is one of the largest groups of the Culicidae family [11]. In FG, 104 species belong to the genus Culex [3] with a majority of species linked to two subgenus named Culex (abbreviated Cux.) and Melanoconion (abbreviated Mel.). In the Culex subgenus many species are vectors of arboviruses and parasites. Culex (Cux.) quinquefasciatus Say was identified as vector of the nematode Wuchereria bancrofti, agent of lymphatic filariasis in FG [12] and as primary vector in northeast Brazil [13]. In a larger scale, this anthropophilic mosquito is also known to be vector of Western equine encephalitis virus (WEEV, Alphavirus) and many viruses from the Flavivirus genus, as Saint-Louis encephalitis virus (SLEV) and West Nile virus (WNV); besides, is also vector of nematode Dirofilaria immitis, that could infect both humans and dogs [14,15,16,17]. In South America, Cx. (Cux.) declarator Dyar & Knab is a vector of SLEV and Bussuquara virus (BSQV, Flavivirus genus) [18]. Culex (Cux.) nigripalpus Theobald allowed the isolation of Cabassou virus (CABV, Alphavirus genus) in FG [19] and is a recognized vector for members of Flavivirus genus with SLEV, WNV and ZIKV and for members of Alphavirus genus with Venezuelan equine encephalitis virus (VEEV) and Eastern equine encephalitis virus (EEEV) in America [20,21]. Culex (Cux.) usquatus Dyar belonging to the Coronator Complex is not a recognized as vector, but the females are indistinguishable from Cx. (Cux.) coronator Dyar & Knab, which is known as vector of WNV [22] and may participate in the transmission of SLEV, VEEV and Mucambo Virus (MUCV, Alphavirus genus) [23,24].
In the Melanoconion subgenus, Cx. (Mel.) portesi Senevet & Abonnenc and Cx. (Mel.) spissipes Theobald are highly suspected to be natural vectors of Tonate virus (TONV) that belongs to the Alphavirus genus in FG [25,26,27], but are also found infected by Bunyavirus (Caru, group Guama, Caraparu and group C) [19]. Culex (Mel.) portesi is a recognized natural vector for MUCV in Trinidad [28] and is a natural vector for the disease caused by El Huayo virus (a group C Orthobunyavirus) in Peru [29]. Ecological studies and the isolation of many arboviral species in Cx. (Mel.) portesi rank it as high potential invertebrate host of numerous viruses [19]. Culex (Mel.) adamesi Sirivanakarn & Galindo, Cx. (Mel.) dunni Dyar, and Cx. (Mel.) pedroi Sirivanakarn & Belkin are natural enzootic vectors of VEEV in Columbia, Peru and Panama [30,31,32]. Culex (Mel.) dunni was also found infected with Pacora virus (PCAV, Bunyavirus-like) in Panama [33]. A last example is Cx. (Mel.) idottus Dyar which is a caiman-biting mosquito involved in the transmission of Hepatozoon caimani and suggested as potential vector of WNV [34].
With global warming, deforestation and urbanization, the distribution areas of mosquitoes change, colonizing new territories. Such changes could promote the emergence of new mosquito vectors due to their exposure to disease agents and hosts, inducing potential outbreaks. To manage and to prevent epidemic emergence, the monitoring and accurate identification of Culex mosquitoes at the species level remains essential. Presently, morphological and molecular methods are the main strategies available for identification and classification of Culex species.
Morphological identification at species level of Culex mosquitoes requires the careful dissection and mounting of male genitalia by an entomological expert because of the presence of slight morphologic characters that allows the identification of sibling species of the genera [35]. Females identification is even harder because morphological characters may be either polymorphic or isomorphic among distinct species [36]. This approach is labor-intensive and time-consuming and, therefore, might not be adapted for routine identification of Culex females. Furthermore, field-caught specimens are rarely in perfect condition, which preclude any reliable morphological identification. For damaged mosquitoes or for immature stages, molecular tools can be an interesting alternative, especially to distinguish morphologically close species [37]. Barcoding and metabarcoding markers which combine DNA barcoding with high-throughput sequencing are sometimes used for delimitation and identification of species. However, species-level identification depends heavily on the choice of marker and the selected primer pair, often with a trade-off between successful species amplification and taxonomic resolution. Variation of a partial sequence of the cytochrome c oxidase subunit I (COI) gene is often used for the identification of mosquito species and contribute to discover cryptic diversity [38]. If on the one side, the COI barcode can be successfully used for delimiting and identifying numerous mosquito species, on the other hand recent studies on the Culex subgenus identification with this fragment indicate a poor resolution in separating species among complexes [36]. To improve species identification, multiple marker/primer pairs are often recommended and remains an expensive method for screening of large samples [39].
In the last decade, an innovative and cheaper proteomic tool named MALDI-TOF MS profiling, emerged for rapid mosquito species identification [40]. The principle is based on the matching of species-specific protein signatures of a specimen with reference spectra database. This promising tool has been reported to distinguish cryptic species of Anopheles genus with high efficiency [41]. Already used successfully for different taxonomic groups of insects, including culicids [42] and phlebotomids [43], standardization of protocols and optimized procedures could enable sharing all MS spectra reference and lead to the creation of an international MS database [44].
Thus, the aim of the present study was to assess whether this proteomic tool could distinct species from the neotropical Culex genus. In this way, field-caught mosquitoes from FG of the Culex genus were selected, classified by morphological criteria and confirmed by molecular barcoding. The legs and thoraxes of the selected Culex specimens were then submitted independently to MALDI-TOF MS to assess intra-species reproducibility and inter-species specificity of MS spectra. The establishment of such innovative tool for Culex mosquito identification should improve studies on this genus which are often hampered by the complexity of current methods for accurate classification.
Materials and Methods
2.Mosquito Collection
Adult mosquitoes were collected from different sites of the coastal floodplain of FG. These mosquito collections occurred during entomological surveys conducted from November 2018 to January 2020, using backpack aspirators and CDC light traps baited (or not) with dry ice. Female specimens of Culex were sorted out and morphologically identified (whenever possible) using taxonomic keys [45,46,47,48,49]under a Leica M165 C stereomicroscope at a magnification up to ×120 (Leica Microsystems, Wetzlar, Germany). When specimens could not be identified until species level, they were grouped according to morphospecies using the genus name followed by the suffix ‘sp.’ combined with a number. Abdomen, thorax and legs of each mosquito specimen were dissected and kept individually at -20°C for subsequent molecular and MALDI-TOF MS analyses. A total of 169 selected females were used in this study.
2.Molecular Identification of Mosquitoes
DNA was individually extracted from the abdomen of each mosquito specimen using the QIAamp DNA tissue extraction kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. Sequences of a fragment of the cytochrome c oxidase subunit I (COI) gene from extracted DNA of each mosquito were amplified using DNA-barcode primers forward LCO1490 (5’-GGT CAA CAA ATC ATA AAG ATA TTG G-3’) and reverse HC02198 (5’-TAA ACT TCA GGG TGA CCA AAA AAT CA-3’), according to protocols previously described [50]. After expected amplified DNA fragment size (658pb) verified by electrophoresis (1.5% agarose gel), samples were sent for sequencing. Sequences were edited using the Molecular Evolutionary Genetics Analysis (MEGA) X software, assembled and aligned in Clustal ω2 algorithm using the default parameters.
A reference database of Culex COI sequences obtained from sequences accessible in the Barcode of Life Data Systems (BOLD) [51] and in GenBank® [52] was used for identification and to construct a Maximum Likelihood tree using a General Time Reversible (GTR) model +G+I (Gamma distribution ; evolutionarily Invariable). Support for internal nodes was estimated using the nonparametric bootstrap method with 1000 replicates. By using the Nucleotide Basic Local Alignment Search Tool (BLAST) [53], the identity percentage and query coverage parameter was also extracted for each sample. Only specimens with identity >97% were selected for this study. Each specimen was then assigned to a mosquito species name. Species identification through DNA-barcoding was used for all specimens including undetermined Culex female specimens but also for confirmation of specimens already identified by morphology.
2.Phylogenetic tree analyses based on COI sequences for MALDI-TOF comparison
COI sequences from specimens included in the MALDI-TOF reference database, were used for the creation of phylogenetic trees. Sequences were aligned through Clustal ω2 algorithm in MEGA X and applied to construct a Maximum Likelihood tree for thoraxes with similar analyses used for identification of the COI sequences to species level.
2.Sample homogenization and MALDI-TOF MS analysis
Thorax and legs of each mosquito specimen were prepared for MALDI-TOF MS as previously described [54,55]. After homogenization of each sample with mix buffer (1:1 solution of 70% (v/v) formic acid and 50% (v/v) acetonitrile), 1 µL of the supernatant was spotted on the MALDI-TOF steel target plate (Bruker Daltonics, Wissembourg, France) in quadruplicate and air-dried. The spots were covered with 1 µL of matrix solution containing saturated α-cyano-4-hydroxycinnamic acid, accordingly to previous work [56]. To control matrix quality (i.e. absence of MS peaks due to matrix buffer impurities) and MALDI-TOF apparatus performance, matrix solution was loaded in duplicate onto each MALDI-TOF plate alone. Likewise, thorax and legs from one fresh specimen of Ae. (Stg.) aegypti Bora-Bora strain was loaded on each plate as quality control of sample preparation and MS spectra acquisition.
2.MALDI-TOF MS parameters
Protein mass profiles were obtained using a MicroFlex LT MALDI-TOF Mass Spectrometer (Bruker Daltonics, Germany), with detection in the linear positive-ion mode at a laser frequency of 50 Hz within a mass range of 2‒20 kDa. The setting parameters of the MALDI-TOF MS apparatus were identical to those previously used [57]. MS spectra were acquired automatically using the AutoXecute of the Flex Control v.2.4 software (Bruker Daltonics).
2.MS spectra analysis
MS spectra profiles were controlled visually first with flexAnalysis v.3.3 software (Bruker Daltonics), then exported to ClinProTools v.2.2 and MALDI-Biotyper v.3.0 software (Bruker Daltonics) for data processing (smoothing, baseline subtraction, peak picking). MS spectra reproducibility was assessed by the comparison of the average spectral profiles (MSP, Main Spectrum Profile) obtained from the four spots for each specimen according to body part with MALDI-Biotyper (Bruker Daltonics). Additionally, mosquito body part MS spectra reproducibility and specificity were assessed using cluster analyses (MSP dendrogram) and the composite correlation index (CCI) tool. The MSP dendrograms were performed from clusters of the specimens, based on comparison of their protein mass profile (i.e. their mass signals and intensities).The CCI tool from MALDI-Biotyper was also used to assess the spectral variations within and between each sample group, as previously described [58]. Higher correlation values (expressed as the mean ± standard deviation, SD) reflecting higher reproducibility for the MS spectra, were used to estimate MS spectra distance between species for each body part. In addition, ClinProTools was used to identify discriminatory peaks among the 13 mosquito species for each body part. The top five of the most intense MS peaks, per mosquito species and per body part, were analyzed with ClinProTools to estimate their performance to discriminate the Culex species. The default parameter settings in ClinProTools for spectrum preparation were applied as previously described [54]. Based on the peak list obtained per species for each body part, the top five of the most intense m/z peaks were selected to include them into the genetic algorithm (GA) model. The selected peaks by the operator provided a recognition capability (RC) value bind to the highest cross-validation (CV) value. The presence or absence of all discriminating peak masses generated by the GA model was controlled by comparing the average spectra from each species per body part.
2.Database creation and blind tests
The MS reference database (DB) was created using spectra from paired legs and thorax of one to four specimens per species using MALDI-Biotyper (Bruker Daltonics). MS spectra were created with an unbiased algorithm using information on the peak position, intensity and frequency. A total of 58 MSP from thoraxes or legs, were included in the reference MS spectra DB (Table 1). The raw MS spectra from legs and thoraxes of Culex mosquitoes are provided for free use (Supplementary file S1). MS spectra from mosquito legs and thoraxes of the 111 remaining specimens were tested against this in-house MS reference spectra DB. The reliability of species identification was estimated using the log score values (LSVs) obtained from the MALDI Biotyper software v.3.0, which ranged from 0 to According to previous studies [42,56,57], LSVs greater than 1.8 were considered reliable for species identification. Data were analyzed with Prism software v.7.00 (GraphPad, San Diego, CA, USA).

Table.
Overview of Culex mosquito origins and identification by COI molecular typing.
| Collection | ||||||
|---|---|---|---|---|---|---|
| Species | Site | Year | Number of specimens | BOLD# accession number (number of associated sequences) | COI gene sequence coverage (%) / identity (%) | Number of specimens included in the reference MS DB per body part (Thoraxes/Legs)§ |
| Cx. (Mel.) adamesi | Mac. | 2018 | 1 | FGMOS2220-20 (1) | 99% / 97% | 1 / 1 |
| Cx. (Cux.) declarator | Mac. | 2018, 2019 | 7 | FGMOS2272-20 (7) | 100% / 99‒100% | 2 / 1 |
| Cx. (Mel.) dunni | Mac. | 2018, 2019 | 30 | FGMOS2748-20 (20); FGMOS2750-20 (10) | 100% / 98‒100% | 4 / 3 |
| Cx. (Mel.) eastor | Mac. | 2018, 2019 | 3 | FGMOS2695-20 (1); FGMOS2743-20 (1); FGMOS2752-20 (1) | 100% / 99‒100% | 1 / 1 |
| Cx. (Mel.) idottus | Mac. | 2018 | 2 | FGMOS3098-23 (2) | 100% / 100% | 1 / 0 |
| Cx. (Cux.) nigripalpus | Mac. | 2018, 2019 | 12 | FGMOS225-16 (12) | 100% / 100% | 3 / 2 |
| Cx. (Mel.) pedroi | Mac. | 2018, 2019 | 15 | FGMOS2700-20 (12); FGMOS2758-20 (3) | 100% / 100% | 4 / 2 |
| Cx. (Mel.) phlogistus | Mac. | 2019 | 1 | FGMOS1542-20 (1) | 100% / 99% | 1 / 1 |
| Cx. (Mel.) portesi | Mac. | 2018 | 28 | FGMOS2416-20 (28) | 100% / 100% | 4 / 1 |
| Cx. (Cux.) quinquefasciatus | Cay. | 2019, 2020 | 34 | FGMOS2275-20 (34) | 100% / 100% | 4 / 4 |
| Cx. (Mel.) rabanicolus | Mac. | 2018 | 5 | FGMOS2744-20 (5) | 100% / 100% | 2 / 2 |
| Cx. (Mel.) spissipes | Mac. | 2018, 2019 | 9 | FGMOS2701-20 (9) | 100% / 97‒99% | 3 / 2 |
| Cx. (Cux.) usquatus | Mac., Rem. | 2018, 2019 | 22 | FGMOS046-16 (9); FGMOS049-16 (1); FGMOS2284-20 (12) | 100% / 99‒100% | 4 / 4 |
| Total | 169 | 34 / 24 | ||||
#BOLD: Barcode of Life Data Systems; COI: cytochrome oxidase subunit 1; §Number of specimens used to create the reference MS database per body part; Cx.: Culex; Cux.: Culex; Mel.: Melanoconion; Mac.: Macouria; Cay.: Cayenne; Rem.: Remire-Montjoly.
Results
3.Morphological and molecular identification of Culex specimens
Field mosquitoes catching were performed during 2018 to 2020 in three distinct areas from FG (Table 1). Among the mosquitoes captured, only Culex female specimens were selected for the present study. The morphological identification was applied to 205 females. However, 29% (60/205) of the specimens were unsuccessfully classified to species level. All Culex specimens were then subjected to molecular analysis by DNA barcoding, using COI gene marker, and each COI sequence was queried against the BOLD system. Reliable species identifications were obtained for 169 females with identity and coverage ranges of 97‒100% and 100%, respectively (Table 1). Culex females belonged to 13 distinct species classified in two subgenera. Four species were categorized in the Culex subgenus (Cx. declarator, Cx. nigripalpus, Cx. quinquefasciatus and Cx. usquatus), and the nine remaining species were classified in the Melanoconion subgenus (Cx. adamesi, Cx. dunni, Cx. eastor, Cx. idottus, Cx. pedroi, Cx. phlogistus, Cx. portesi, Cx. rabanicolus and Cx. spissipes). According to our data set availability, 1 to 34 specimens per species were acquired. Thirty four Culex specimens encompassing the 13 species and used for MALDI-TOF MS reference database were selected for the phylogenetic analysis based on COI gene sequences (Supplementary file S2). Specimens are successfully assembled and classified by species.
3.Reproducible and specific MS spectra from two Culex body parts
Among the 1352 MS spectra (169 specimens × two body parts × four replicates) acquired, MS profiles of high intensity (>3000 a.u.) were reached for the vast majority of the samples (>95%). The samples presenting MS spectra with low intensity were not selected for their introduction as reference spectra in the MS DB. For example, MS spectra of legs from the two Cx. idottus specimens were excluded of the analysis due to the low intensity and inter-sample heterogeneity of MS profiles (Supplementary file S3). Then, in total, MS spectra of 12 and 13 Culex species were analyzed for legs and thoraxes, respectively. The visual comparison of MS profiles acquired for legs (Figure 1A) and thoraxes (Figure 2A) differed importantly among the Culex species. To assess the reproducibility and specificity of MS spectra per body part according to species, CCI and cluster analyses were carried out. MS spectra of high intensity (>3000 a.u.) from paired legs (n=24) and thorax (n=34) of one to four specimens per species were selected for these analyses. The mean CCI-values obtained among MS spectra per body part and per species were elevated, ranging from 0.59 to 0.84 for legs (Figure 1B) and from 0.66 to 0.93 for thoraxes (Figure 2B). Conversely, the mean CCI-values obtained between species for legs and thoraxes were very low (<0.20), supporting the high MS spectra species-specificity. Nevertheless, it is interesting to notice, that elevate CCI-values were observed for thoraxes MS spectra between Cx. idottus and Cx. rabanicolus (mean ± SD: 0.77 ± 0.01).
The MSP dendrograms created per body part with the same MS spectra, revealed the clustering of specimens from the same species on the same branch for each Culex species for legs (Figure 1C) and thoraxes (Figure 2C). The proximity and low distance of branches between Cx. idottus and Cx. rabanicolus for thorax body part, confirmed the proximity of their MS spectra. However, the absence of species interlacing emphasized the reproducibility and specificity of the protein profiles for both body parts. Although, a clustering of Culex species from the same subgenus was obtained on both MSP dendrograms, the separation was clearer for MS spectra from legs than from thoraxes. Interestingly, the ordination of the species on MSP dendrogram was similar to those obtained in the phylogenetic tree from COI gene sequences of the 34 Culex specimens (Supplementary file S2).
3.Leg and Thoraxe Biomarkers Distinguishing Culex Species
To identify discriminatory MS peaks among the 13 Culex species for legs and thoraxes, MS spectra from the 169 specimens were analyzed using the GA tool from ClinProTools 2.2 software. As the accuracy of identification is directly linked to the intensity of MS spectra, the analysis was carried out on the most intense mass peaks from legs and thoraxes per mosquito species. After verification of the peak report in the average spectrum, the selection of the top-five mass peak lists per species conducted to a total of 29 and 26 MS peaks for legs and thoraxes, respectively (Supplementary files S4 and S5). The inclusion of these MS peak lists in the GA model displayed respective recognition capability (RC) and cross-validation (CV) values of 93.2% and 91.4% for legs and 94.5% and 86.5% for thoraxes. When the top 10 mass peak lists per species was selected, a total of 37 and 39 MS peaks were found for legs and thoraxes, respectively. The application of the GA model on the top 10 mass peak lists did not improve RC and CV values for legs, 92.8% and 91.4%, respectively, whereas, for thoraxes the CV reached 90.1% and the RC remained at 93.7%. These results underlined that the top five mass peak list per species appeared as the main MS peaks to discriminate these Culex species for both body parts. The comparison of the top five mass peak list between legs and thoraxes indicated that three mass-to-charge ratio were similar (at about 5206, 5405 and 6426 m/z), confirming the high specificity of MS spectra per body part (Supplementary files S4 and S5).
3.Creation of the MS reference spectra database and validation steps
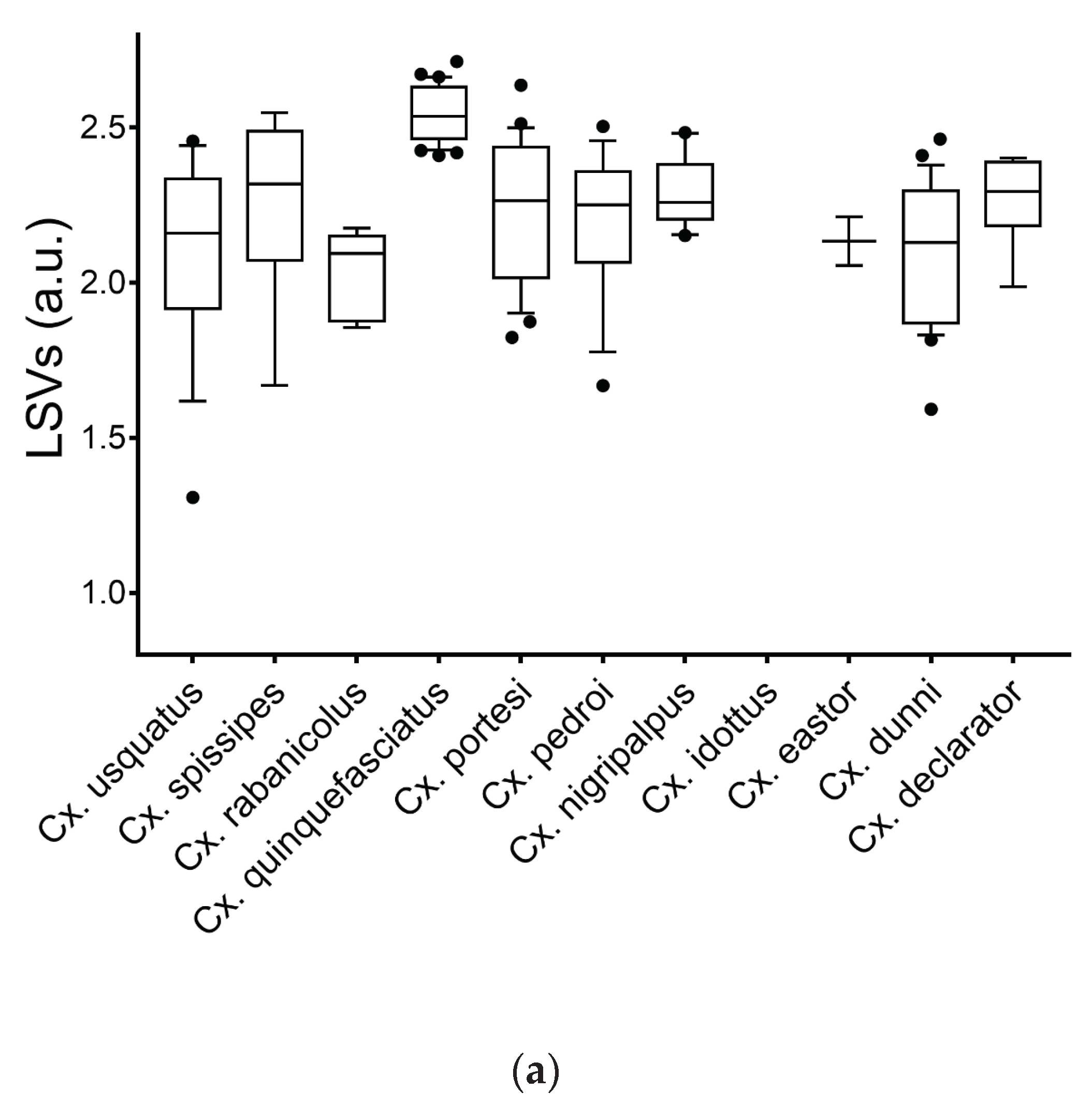
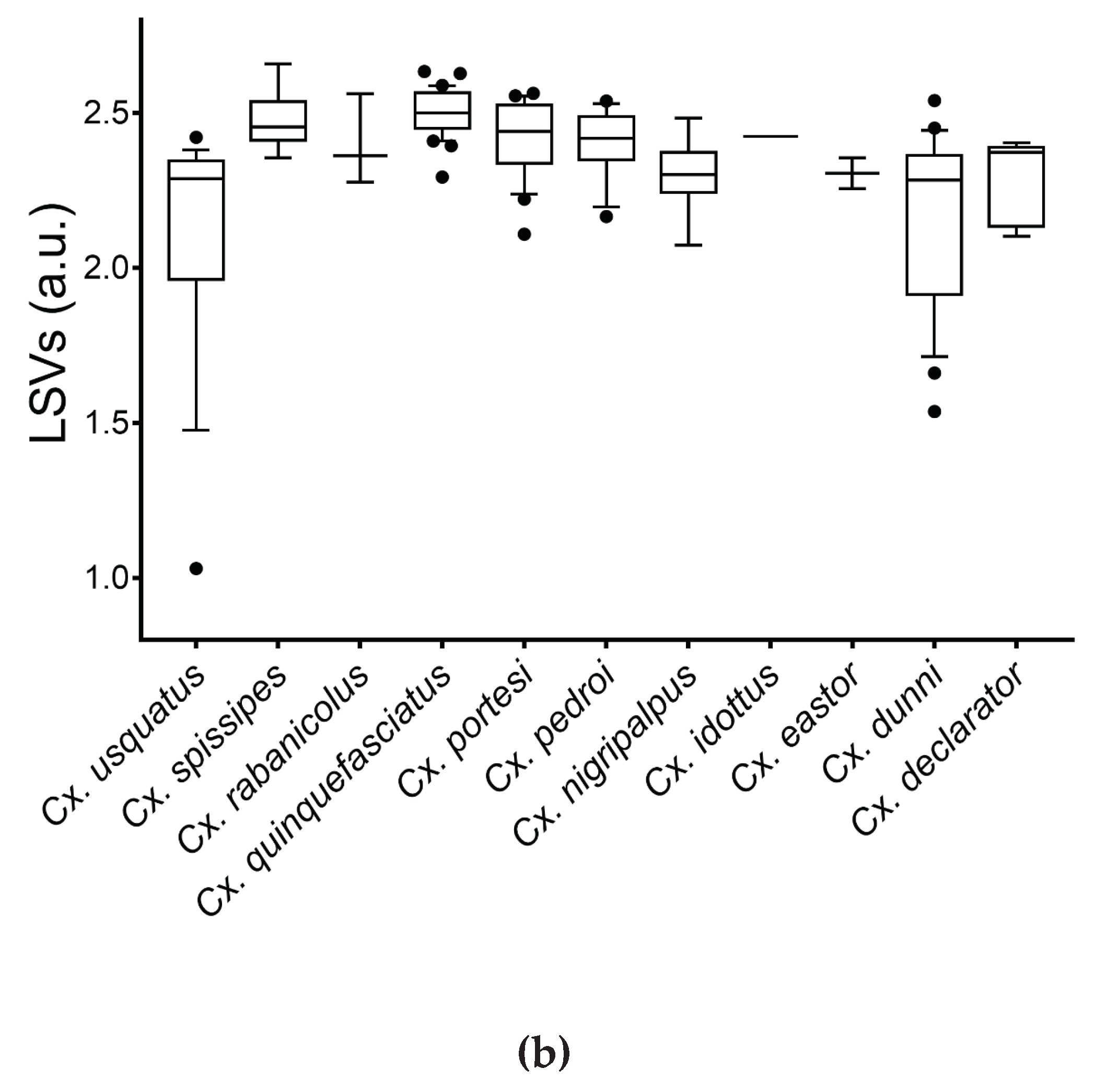
MS spectra from 12 and 13 Culex species for legs (n=24) and thoraxes (n=34), respectively, including one to four specimens per species, were selected for database creation (Table 1). MS spectra from the remaining specimens (legs, n=145; thoraxes, n=135) were queried against the DB. The LSVs ranged from 1.31 to 2.71 for legs (Figure 3A) and from 1.03 to 2.66 for thoraxes (Figure 3B). For a reliable identification, a LSV upper than the 1.8 threshold is required [42,56]. In these conditions, a correct identification (LSVs>1.8) could be considered for 95.9% (n=139/145) of legs MS spectra and 94.8% (n=128/135) for thoraxes.
Among these MS spectra reaching the LSVs threshold, 100% of the identification results were concordant with the molecular results. Interestingly, if paired samples per specimen were considered (n=133), solely two specimens of Cx. usquatus failed to obtain a LSV upper than 1.8 for at least one body part (Supplementary file S6). If the LSV cut-off is increased at 2.0 to improve identification confidence, the rate of the specimens which reached this threshold reduces to 78.2% (n=104/133) for legs and 90.2% (n=120/133) for thoraxes. However, 91.0% (n=121/133) of paired samples per specimen achieved this higher threshold in at least one body part.
Discussion
MALDI-TOF MS is a method routinely used in microbiological diagnostic laboratory for identification of bacteria and archaea [59], but the relevance of the technique remains largely underestimated for identification of multicellular organisms. The success of the MALDI-TOF MS for arthropod identification in this last decade, highlights this emerging tool as a relevant alternative for mosquitoes species identification [43,60,61,62]. As many mosquitoes species from the Culex genus have been proven to be vectors of pathogenic agents of human and veterinary importance [63,64,65], an accurate classification of the specimens is primordial. The present work demonstrated that MALDI-TOF MS can be used as an alternative to current methods for Culex female identification at the adult stage, using legs and thorax.
Prior MS submission, a relevant classification of the Culex mosquitoes per species was compulsory. Firstly, the morphological identification was performed allowing to classify mosquitoes from only five distinct species, two from the Culex subgenus (Cx. quinquefasciatus and Cx. usquatus) and three from the Melanoconion subgenus (Cx. portesi, Cx. pedroi and Cx. spissipes). The accuracy of this morphological classification was confirmed by molecular DNA barcoding of the COI gene. Morphological method failed to identify at the species level 29% (n=60) of Culex mosquitoes. The limitation of classification of mosquitoes by morphological approach was also reported for others genera [36,66,67]. For instance, in the Anopheles genus, the members of the Gambiae Complex and Funestus Group could not be distinguished uniquely by examining external features of their anatomy [68,69]. Moreover, the skill and regular training of entomological experts is also another factor playing a role in the capacity of correct specimen identification [70]. The genus Culex is recognized as a highly diverse group of mosquitoes for which delimitation and identification of species is particularly difficult. Complementary methods are therefore required for improving species identification.
DNA barcoding using the mitochondrial COI gene is widely used for species identification in molecular taxonomy [71]. A first COI barcoding database, available in the BOLD system, established based on morphological classification of male Culex specimens of FG [3] was used to validate morphological identification and to classify the unidentified females of this study. The COI gene has been recorded as an effective and accessible DNA barcode that provides a high performance in delimiting species within the subgenus Melanoconion of Culex species, a subgenus displaying the highest species diversity in tropical regions [66]. However, others studies also applied COI barcode fragment for identification of species within the subgenus Culex and concluded that this barcode do not contain enough information to distinguish species within this subgenus [36]. Based on COI sequences from Brazil and Argentina, Laurito et al. [36] highlighted that Cx. declarator cannot be differentiated from Cx. bidens Dyar and Cx. tatoi Casal & García. However, these last two species have never been detected in FG [3], and only Cx. declarator was considered here. Similarly, COI sequences of Cx. usquatus can be confused with Cx. camposi Dyar, Cx. coronator, Cx. maxi Dyar and Cx. surinamensis [36]. In FG, only Cx. coronator, Cx. surinamensis and Cx. usquatus have already been reported. Nevertheless, a recent review of Culex species from FG revealed that specimens identified as Cx. coronator in historical literature were closer to the lectotype of Cx. usquatus described from Suriname [3]. Culex usquatus is morphologically distinguishable from Cx. surinamensis, thus confirming that COI sequences classified as Cx. usquatus could be attributed to this species in this study.
To improve the identification, molecular tools often develop new algorithms of analysis or add others molecular markers. For Anopheles species, COI is not enough to discriminate species of the Gambiae Complex. The addition of ITS2 analysis increases greatly the potential of identification of these species [72]. Indeed, the understanding lack on COI sequences analysis highlight the need to explore complementary or innovative tools.
The proteomic MALDI-TOF MS approach is increasingly used for rapid arthropod identification [42,43,60,61,73]. Indeed, to improve the intra-species reproducibility of MS spectra and to share reference protein profiles, protocols were standardized [56,74]. In order to develop an accessible mosquito spectra database of Culex species, the present study employed MALDI-TOF MS double spectra biotyping strategy. Here, for each specimen tested and for DB creation, legs and thorax from the same specimen were independently submitted to MS analysis, whenever possible. Specific MS spectra from the 13 Culex species per body part were obtained. The cluster analysis on MSP dendrogram confirmed this species specificity. Besides, similar ordination of the specimens for both body parts with the COI phylogenetic tree, revealed a relationship between molecular taxonomic classification and MS spectra profiles. However, as MS spectra from closely-related species were grouped in the same branch of MS dendrogram, a mismatching could occur. To prevent misidentification, the legs and thorax were independently query against the DB. This double query allowed to corroborate identification of each specimen using both body parts, enhancing the identification confidence [55]. In this study, 100% of the samples were correctly identified at the species level on both body parts and the rate of relevant identification (LSVs>1.8) was about 95% for each ones. Interestingly, a higher reproducibility of MS spectra per species was obtained for thorax compared to legs. The lower reproducibility of legs could be attributed to the number of legs available for each specimen. Effectively, legs are breakable and during the catching or storing period some could be lost, which could induced heterogeneity of MS profile intensities among specimens from the same species [75]. In the cases of all the legs were lost, the specimen could be always identified using thorax compartment. Then, the thorax appears as the more appropriate compartment for mosquito identification, followed by legs accordingly to previous work [44]. Furthermore, these two body parts does not prevent screening for viruses, parasites, or source of blood feeding which can be researched in the head or abdomen by molecular [76,77] or MALDI-TOF MS methods [78,79]. These complementary data are primordial in the frame of surveillance programs.
Quality of MS spectra can be altered by various factors such as sample homogenization, quantity of mix buffer, engorgement status, storage conditions or even geographical origin of the collection [42,55,75]. The storage is too critical for field caught mosquitoes. The best method for long storage of arthropods for MALDI-TOF MS analysis is frozen or to maintain them at RT with silica gel [80], when immediate freezing is not possible. In this study, all the specimens were field-derived, dried and stored frozen at -20°C in tubes or plates from one to three years. The elevate intra-species reproducibility and inter-species specificity of MS spectra for both body parts, suggested that specimens were appropriately preserved. Among the 169 mosquitoes tested, only two Cx. usquatus failed, on both body parts, to reach the relevant threshold value (LSV>1.8). These low LSVs could be attributed to improper storing of the mosquitoes. It is possible that protein degradation occurred in these specimens conducting to lower quality of MS spectra, as reported in previous studies [58,81,82].
It is noteworthy that all the Culex species selected in this study were correctly classified using MALDI-TOF MS profiling, with concordant results between thorax and legs, and in agreement with molecular identification. Three species from the Melanoconion subgenus, Cx. idottus, Cx. phlogistus and Cx. rabanicolus, presenting isomorphic traits among females and which could not be accurately distinguished morphologically, were unambiguously identified by this proteomic tool.
The present spectral database, created using thoraxes and legs from females of 13 Culex species from FG, represents the first attempt to create a MALDI-TOF MS database for identification of neotropical Culex species. However, the database encompass only 12.5% (n=13/104) of the total number of Culex species known in FG and solely two out of the eight subgenera from the Culex genus of the mosquitoes actually inventoried in the territory [3]. The widening of this MALDI-TOF MS spectra reference DB with missing Culex species, become compulsory for application of this tool in the monitoring of Culex genus vectors in FG. An upgrading of this DB including others mosquito species occurring in FG (n=242) will improve its usage and may be helpful to discriminate cryptic or close-related species. MS spectra for the identification of eight distinct Anopheles species from FG, among which four are malaria vectors, are already available [54].
Several works reported the efficiency of the MALDI-TOF MS for identification of mosquitoes at immature stages [83,84] and demonstrated it interest for management of Culicidae larval habitats [85]. As larva can be confusing to distinguish morphologically, it could be interesting to assess MALDI-TOF MS for identification of immature stages. MALDI-TOF MS appears then highly promising for discriminating mosquito fauna [41,56,82].
The present work demonstrated that MALDI-TOF MS could be an alternative to current methods for identification of Culex female, using dissected legs and thorax. The double body part protein signature of each specimen improved the identification quality. Raw MS spectra from legs and thorax of these 13 Culex species included in the DB are freely available (Supplementary file S1). The sharing of reference MS spectra is essential in the frame of the creation of an international DB. Except for the expensive cost of the MALDI-TOF MS instrument, this approach is highly competitive economically compared to current molecular methods. It does not require particular skills and could be used for “live” monitoring of vectors due to its short time of handling and obtaining results. This approach appears suitable for the identification of mosquitoes from the Culex genus, for which morphological and molecular methods remains time consuming and with a potential risk of misidentification. In the near future, the widening of this reference MS spectra DB with specimens from others Culex species, at both adult and immature stages, could improve knowledge of these species for adapted surveillance and control measures to reduce the risks of pathogen transmission not exclusively to French Guiana.
Supplementary Materials
The following supporting information is available and can be downloaded: Supplementary File S1; Supplementary File S2; Supplementary File S3; Supplementary File S4; Supplementary File S5 and Supplementary File S6.
Author Contributions
Conceptualization and methodology of the experiments: L.A.; experiments investigation and tools contribution: M.M.C. and A.G.; data analysis: L.A., A.G., M.M.C. and S.B.; field collections preparation: R.C., P.G. and A.G.; field collections dissection: A.G., J.I. and H.N.; writing—original draft preparation: L.A., S.B., M.M.C. and A.G.; funding acquisition: L.A., J.B.D., I.D. and A.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been supported by the Délégation Générale pour l’Armement (DGA, MSProfileR project, Grant no PDH-2-NBC 2-B-2201) and by Regional Health Agency of French Guiana (Conventions n° FIR/ARS/2014/91, n°FIR/ARS/2015/68, and Convention ARS/IPG 2021–2023 - Nov 26th 2021). This work was also partially supported by the European Union FEDER controle Programme under Grant number GY0010695.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets of MS reference spectra added to the MS DB in the current study are freely available and downloadable from the Additional File S1.
Acknowledgments
We would like to acknowledge Denis Blanchet for the fruitful discussions and all people involve in the creation of an international open source databank.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Foley, D.H.; Rueda, L.M.; Wilkerson, R.C. Insight into Global Mosquito Biogeography from Country Species Records. J. Med. Entomol. 2007, 44, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Talaga, S.; Dejean, A.; Carinci, R.; Gaborit, P.; Dusfour, I.; Girod, R. Updated Checklist of the Mosquitoes (Diptera: Culicidae) of French Guiana. J. Med. Entomol. 2015, 52, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Talaga, S.; Duchemin, J.-B.; Girod, R.; Dusfour, I. The Culex Mosquitoes (Diptera: Culicidae) of French Guiana: A Comprehensive Review With the Description of Three New Species. J. Med. Entomol. 2021, tjaa205. [Google Scholar] [CrossRef]
- Pommier de Santi, V.P.; Girod, R.; Mura, M.; Dia, A.; Briolant, S.; Djossou, F.; Dusfour, I.; Mendibil, A.; Simon, F.; Deparis, X.; et al. Epidemiological and Entomological Studies of a Malaria Outbreak among French Armed Forces Deployed at Illegal Gold Mining Sites Reveal New Aspects of the Disease’s Transmission in French Guiana. Malar. J. 2016, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Dusfour, I.; Issaly, J.; Carinci, R.; Gaborit, P.; Girod, R. Incrimination of Anopheles (Anopheles) Intermedius Peryassú, An. (Nyssorhynchus) Nuneztovari Gabaldón, An. (Nys.) Oswaldoi Peryassú as Natural Vectors of Plasmodium Falciparum in French Guiana. Mem. Inst. Oswaldo Cruz 2012, 107, 429–432. [Google Scholar] [CrossRef]
- Pommier de Santi, V.; Dia, A.; Adde, A.; Hyvert, G.; Galant, J.; Mazevet, M.; Nguyen, C.; Vezenegho, S.B.; Dusfour, I.; Girod, R.; et al. Malaria in French Guiana Linked to Illegal Gold Mining. Emerg. Infect. Dis. 2016, 22, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Vezenegho, S.B.; Adde, A.; de Santi, V.P.; Issaly, J.; Carinci, R.; Gaborit, P.; Dusfour, I.; Girod, R.; Briolant, S. High Malaria Transmission in a Forested Malaria Focus in French Guiana: How Can Exophagic Anopheles Darlingi Thwart Vector Control and Prevention Measures? Mem. Inst. Oswaldo Cruz 2016, 111, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenço-de-Oliveira, R.; Failloux, A.-B. Differential Susceptibilities of Aedes Aegypti and Aedes Albopictus from the Americas to Zika Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004543. [Google Scholar] [CrossRef] [PubMed]
- Fouque, F.; Vazeille, M.; Mousson, L.; Gaborit, P.; Carinci, R.; Issaly, J.; Rodhain, F.; Failloux, A.-B. Aedes Aegypti in French Guiana: Susceptibility to a Dengue Virus. Trop. Med. Int. Health 2001, 6, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Girod, R.; Guidez, A.; Carinci, R.; Issaly, J.; Gaborit, P.; Ferrero, E.; Ardillon, V.; Fontaine, A.; Dusfour, I.; Briolant, S. Detection of Chikungunya Virus Circulation Using Sugar-Baited Traps during a Major Outbreak in French Guiana. PLoS Negl. Trop. Dis. 2016, 10, e0004876. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E. Classification within the Cosmopolitan Genus Culex (Diptera: Culicidae): The Foundation for Molecular Systematics and Phylogenetic Research. Acta Trop. 2011, 120, 1–14. [Google Scholar] [CrossRef]
- Floch, H.; De Lajudie, P. Streptococci, Dick reaction and endemic lymphangitis in French Guiana. Bull. Soc. Pathol. Exot. Filiales 1945, 38, 127–132. [Google Scholar]
- Fontes, G.; Leite, A.B.; Vasconcelos de Lima, A.R.; Freitas, H.; Ehrenberg, J.P.; da Rocha, E.M.M. Lymphatic Filariasis in Brazil: Epidemiological Situation and Outlook for Elimination. Parasit. Vectors 2012, 5, 272. [Google Scholar] [CrossRef] [PubMed]
- Gorris, M.E.; Bartlow, A.W.; Temple, S.D.; Romero-Alvarez, D.; Shutt, D.P.; Fair, J.M.; Kaufeld, K.A.; Del Valle, S.Y.; Manore, C.A. Updated Distribution Maps of Predominant Culex Mosquitoes across the Americas. Parasit. Vectors 2021, 14, 547. [Google Scholar] [CrossRef]
- Labarthe, N.; Guerrero, J. Epidemiology of Heartworm: What Is Happening in South America and Mexico? Vet. Parasitol. 2005, 133, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Rochlin, I.; Faraji, A.; Healy, K.; Andreadis, T.G. West Nile Virus Mosquito Vectors in North America. J. Med. Entomol. 2019, 56, 1475–1490. [Google Scholar] [CrossRef] [PubMed]
- Stenn, T.; Peck, K.J.; Rocha Pereira, G.; Burkett-Cadena, N.D. Vertebrate Hosts of Aedes Aegypti, Aedes Albopictus, and Culex Quinquefasciatus (Diptera: Culicidae) as Potential Vectors of Zika Virus in Florida. J. Med. Entomol. 2019, 56, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.J. da S.P.; Thies, S.F.; da Silva, D.J.F.; Kubiszeski, J.R.; Barreto, E.S.; Monteiro, H.A. de O.; Mondini, A.; São Bernardo, C.S.; Bronzoni, R.V. de M. Ecological Aspects of Potential Arbovirus Vectors (Diptera: Culicidae) in an Urban Landscape of Southern Amazon, Brazil. Acta Trop. 2020, 202, 105276. [Google Scholar] [CrossRef] [PubMed]
- Degallier, N. Les arbovirirus selvatiques en Guyane Francaise et leurs vecteurs, 1982.
- Day, J.F.; Curtis, G.A. Annual Emergence Patterns of Culex Nigripalpus Females before, during and after a Widespread St. Louis Encephalitis Epidemic in South Florida. J. Am. Mosq. Control Assoc. 1993, 9, 249–255. [Google Scholar] [PubMed]
- Sardelis, M.R.; Turell, M.J.; Dohm, D.J.; O’Guinn, M.L. Vector Competence of Selected North American Culex and Coquillettidia Mosquitoes for West Nile Virus. Emerg. Infect. Dis. 2001, 7, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Connelly, C.R.; O’Meara, G.F.; Hickman, D.; Karr, N. Reproductive Biology and Susceptibility of Florida Culex Coronator to Infection with West Nile Virus. Vector Borne Zoonotic Dis. 2014, 14, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Consoli, R.A.G.B.; Lourenço de Oliveira, R. Principais Mosquitos de Importância Sanitária No Brasil Available online:. Available online: https://www.arca.fiocruz.br/handle/icict/2708 (accessed on 20 September 2022).
- Forattini, O.P. Produto | Detalhes | Culicidologia Médica Vol. Available online: https://www.edusp.com.br/loja/produto/370/culicidologia-medica-vol--2--identificacao,-biologia,-epidemiologia (accessed on 20 September 2022).
- Digoutte, J.P. Ecologie des Arbovirus et Leur Rôle Pathogène chez L’Homme en Guyane Française; Institut Pasteur de la Guyane Française—Groupe I.N.S.E.R.M. U79, 1975.
- Fischer, C.; Pontier, D.; Filippi-Codaccioni, O.; Pons, J.-B.; Postigo-Hidalgo, I.; Duhayer, J.; Brünink, S.; Drexler, J.F. Venezuelan Equine Encephalitis Complex Alphavirus in Bats, French Guiana. Emerg. Infect. Dis. 2021, 27, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Mutricy, R.; Djossou, F.; Matheus, S.; Lorenzi-Martinez, E.; De Laval, F.; Demar, M.; Nacher, M.; Rousset, D.; Epelboin, L. Discriminating Tonate Virus from Dengue Virus Infection: A Matched Case–Control Study in French Guiana, 2003–Am. J. Trop. Med. Hyg. 2020, 102, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Auguste, A.J.; Volk, S.M.; Arrigo, N.C.; Martinez, R.; Ramkissoon, V.; Adams, A.P.; Thompson, N.N.; Adesiyun, A.A.; Chadee, D.D.; Foster, J.E.; et al. Isolation and Phylogenetic Analysis of Mucambo Virus (Venezuelan Equine Encephalitis Complex Subtype IIIA) in Trinidad. Virology 2009, 392, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Treangen, T.J.; Schoeler, G.; Phillippy, A.M.; Bergman, N.H.; Turell, M.J. Identification and Genomic Analysis of a Novel Group C Orthobunyavirus Isolated from a Mosquito Captured near Iquitos, Peru. PLoS Negl. Trop. Dis. 2016, 10, e0004440. [Google Scholar] [CrossRef]
- Ferro, C.; Boshell, J.; Moncayo, A.C.; Gonzalez, M.; Ahumada, M.L.; Kang, W.; Weaver, S.C. Natural Enzootic Vectors of Venezuelan Equine Encephalitis Virus in the Magdalena Valley, Colombia. Emerg. Infect. Dis. 2003, 9, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Padilla, S.L.; Prieto, K.; Dohm, D.J.; Turell, M.J.; Klein, T.A.; Fernández, R.; Watts, D.M.; Lowen, R.G.; Palacios, G.F.; Pitt, M.L.; et al. Complete Genomic Sequences of Venezuelan Equine Encephalitis Virus Subtype IIID Isolates from Mosquitoes. Arch. Virol. 2020, 165, 1715–1717. [Google Scholar] [CrossRef]
- Torres, R.; Samudio, R.; Carrera, J.-P.; Young, J.; Márquez, R.; Hurtado, L.; Weaver, S.; Chaves, L.F.; Tesh, R.; Cáceres, L. Enzootic Mosquito Vector Species at Equine Encephalitis Transmission Foci in the República de Panamá. PLoS ONE 2017, 12, e0185491. [Google Scholar] [CrossRef]
- Galindo, P. Los Arbovírus de Panama. Rev Med Panama 1978, 1–41. [Google Scholar]
- Viana, L.A.; Soares, P.; Paiva, F.; Lourenço-De-Oliveira, R. Caiman-Biting Mosquitoes and the Natural Vectors of Hepatozoon Caimani in Brazil. J. Med. Entomol. 2010, 47, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Sallum, M.A.; Forattini, O.P. Revision of the Spissipes Section of Culex (Melanoconion) (Diptera:Culicidae). J. Am. Mosq. Control Assoc. 1996, 12, 517–600. [Google Scholar] [PubMed]
- Laurito, M.; Oliveira, T.M. de; Almiron, W.R.; Sallum, M.A.M. COI Barcode versus Morphological Identification of Culex ( Culex ) (Diptera: Culicidae) Species: A Case Study Using Samples from Argentina and Brazil. Mem. Inst. Oswaldo Cruz 2013, 108, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Andrić, A.; Šikoparija, B.; Obreht, D.; Dan, M.; Preradović, J.; Radenković, S.; Pérez-Banon, C.; Vujić, A. DNA Barcoding Applied: Identifying the Larva of Merodon Avidus (Diptera: Syrphidae). Acta Entomol. Musei Natl. Pragae 2014, 54, 741–757. [Google Scholar]
- Hernández-Triana, M.; Brugman, A.; Nikolova, I.; Ruiz-Arrondo, I.; Barrero, E.; Thorne, L.; Fernández de Marco, M.; Krüger, A.; Lumley, S.; Johnson, N.; et al. DNA Barcoding of British Mosquitoes (Diptera, Culicidae) to Support Species Identification, Discovery of Cryptic Genetic Diversity and Monitoring Invasive Species. ZooKeys 2019, 832, 57–76. [Google Scholar] [CrossRef] [PubMed]
- Bourke, B.P.; Oliveira, T.P.; Suesdek, L.; Bergo, E.S.; Sallum, M.A.M. A Multi-Locus Approach to Barcoding in the Anopheles Strodei Subgroup (Diptera: Culicidae). Parasit. Vectors 2013, 6, 111. [Google Scholar] [CrossRef] [PubMed]
- Raharimalala, F.N.; Andrianinarivomanana, T.M.; Rakotondrasoa, A.; Collard, J.M.; Boyer, S. Usefulness and Accuracy of MALDI-TOF Mass Spectrometry as a Supplementary Tool to Identify Mosquito Vector Species and to Invest in Development of International Database. Med. Vet. Entomol. 2017, 31, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Pflüger, V.; Wittwer, M.; Ziegler, D.; Chandre, F.; Simard, F.; Lengeler, C. Identification of Cryptic Anopheles Mosquito Species by Molecular Protein Profiling. PLoS ONE 2013, 8, e57486. [Google Scholar] [CrossRef] [PubMed]
- Yssouf, A.; Almeras, L.; Raoult, D.; Parola, P. Emerging Tools for Identification of Arthropod Vectors. Future Microbiol. 2016, 11, 549–566. [Google Scholar] [CrossRef] [PubMed]
- Chavy, A.; Nabet, C.; Normand, A.C.; Kocher, A.; Ginouves, M.; Prévot, G.; Vasconcelos dos Santos, T.; Demar, M.; Piarroux, R.; de Thoisy, B. Identification of French Guiana Sand Flies Using MALDI-TOF Mass Spectrometry with a New Mass Spectra Library. PLoS Negl. Trop. Dis. 2019, 13, e0007031. [Google Scholar] [CrossRef]
- Bamou, R.; Costa, M.M.; Diarra, A.Z.; Martins, A.J.; Parola, P.; Almeras, L. Enhanced Procedures for Mosquito Identification by MALDI-TOF MS. Parasit. Vectors 2022, 15, 240. [Google Scholar] [CrossRef] [PubMed]
- Gaffigan, T.V.; Ward, R.A. Index to the Second Supplement to “A Catalog of the Mosquitoes of the World”, with Corrections and Additions (Diptera: Culicidae). Mosq. Syst. 1985, 52–63. [Google Scholar]
- Harbach, R.E. Mosquito Taxonomic Inventory Available online: http://mosquitotaxonomic- inventory.info/.
- Knight, K.L.; Stone, A. A Catalog of the Mosquitoes of the World (Diptera: Culicidae); 2nd ed.; Entomological Society of America: College Park, Maryland, 1977; Volume 6. [Google Scholar]
- Pecor, J.E.; Mallampalli, V.L.; Harbach, R.E.; Peyton, E.L. Catalog and Illustrated Review of the Subgenus Melanoconion of Culex (Diptera: Culicidae). Contrib. Am. Entomol. Inst. USA 1992. [Google Scholar]
- Sirivanakarn, S. A Review of the Systematics and a Proposed Scheme of Internal Classification of the New World Subgenus Melanoconion of Culex (Diptera, Culicidae). Mosq Syst 1982, 70. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Ratnasingham, S.; Hebert, P.D.N. Bold: The Barcode of Life Data System (Http://Www.Barcodinglife.Org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Ostell, J.; Pruitt, K.D.; Sayers, E.W. GenBank. Nucleic Acids Res. 2018, 46, D41–D47. [Google Scholar] [CrossRef] [PubMed]
- BLAST.
- Briolant, S.; Costa, M.M.; Nguyen, C.; Dusfour, I.; Pommier de Santi, V.; Girod, R.; Almeras, L. Identification of French Guiana Anopheline Mosquitoes by MALDI-TOF MS Profiling Using Protein Signatures from Two Body Parts. PLoS ONE 2020, 15, e0234098. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rúa, A.; Pagès, N.; Fontaine, A.; Nuccio, C.; Hery, L.; Goindin, D.; Gustave, J.; Almeras, L. Improvement of Mosquito Identification by MALDI-TOF MS Biotyping Using Protein Signatures from Two Body Parts. Parasit. Vectors 2018, 11, 574. [Google Scholar] [CrossRef] [PubMed]
- Nebbak, A.; Willcox, A.C.; Bitam, I.; Raoult, D.; Parola, P.; Almeras, L. Standardization of Sample Homogenization for Mosquito Identification Using an Innovative Proteomic Tool Based on Protein Profiling. PROTEOMICS 2016, 16, 3148–3160. [Google Scholar] [CrossRef] [PubMed]
- Lafri, I.; Almeras, L.; Bitam, I.; Caputo, A.; Yssouf, A.; Forestier, C.-L.; Izri, A.; Raoult, D.; Parola, P. Identification of Algerian Field-Caught Phlebotomine Sand Fly Vectors by MALDI-TOF MS. PLoS Negl. Trop. Dis. 2016, 10, e0004351. [Google Scholar] [CrossRef]
- Diarra, A.Z.; Almeras, L.; Laroche, M.; Berenger, J.-M.; Koné, A.K.; Bocoum, Z.; Dabo, A.; Doumbo, O.; Raoult, D.; Parola, P. Molecular and MALDI-TOF Identification of Ticks and Tick-Associated Bacteria in Mali. PLoS Negl. Trop. Dis. 2017, 11, e0005762. [Google Scholar] [CrossRef] [PubMed]
- Seng, P.; Drancourt, M.; Gouriet, F.; La Scola, B.; Fournier, P.-E.; Rolain, J.M.; Raoult, D. Ongoing Revolution in Bacteriology: Routine Identification of Bacteria by Matrix-Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2009, 49, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Karger, A. Current Developments to Use Linear MALDI-TOF Spectra for the Identification and Typing of Bacteria and the Characterization of Other Cells/Organisms Related to Infectious Diseases. PROTEOMICS – Clin. Appl. 2016, 10, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Sevestre, J.; Diarra, A.Z.; Laroche, M.; Almeras, L.; Parola, P. Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry: An Emerging Tool for Studying the Vectors of Human Infectious Diseases. Future Microbiol. 2021, 16, 323–340. [Google Scholar] [CrossRef] [PubMed]
- Yssouf, A.; Socolovschi, C.; Flaudrops, C.; Ndiath, M.O.; Sougoufara, S.; Dehecq, J.-S.; Lacour, G.; Berenger, J.-M.; Sokhna, C.S.; Raoult, D.; et al. Matrix-Assisted Laser Desorption Ionization - Time of Flight Mass Spectrometry: An Emerging Tool for the Rapid Identification of Mosquito Vectors. PLoS ONE 2013, 8, e72380. [Google Scholar] [CrossRef] [PubMed]
- Cupp, E.W.; Hassan, H.K.; Yue, X.; Oldland, W.K.; Lilley, B.M.; Unnasch, T.R. West Nile Virus Infection in Mosquitoes in the Mid-South USA, 2002–J. Med. Entomol. 2007, 44, 117–125. [Google Scholar] [CrossRef]
- Swei, A.; Couper, L.I.; Coffey, L.L.; Kapan, D.; Bennett, S. Patterns, Drivers, and Challenges of Vector-Borne Disease Emergence. Vector Borne Zoonotic Dis. 2020, 20, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; O’Guinn, M.L.; Jones, J.W.; Sardelis, M.R.; Dohm, D.J.; Watts, D.M.; Fernandez, R.; Travassos da Rosa, A.; Guzman, H.; Tesh, R.; et al. Isolation of Viruses from Mosquitoes (Diptera: Culicidae) Collected in the Amazon Basin Region of Peru. J. Med. Entomol. 2005, 42, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Torres-Gutierrez, C.; de Oliveira, T.M.P.; Emerson, K.J.; Sterlino Bergo, E.; Mureb Sallum, M.A. Molecular Phylogeny of Culex Subgenus Melanoconion (Diptera: Culicidae) Based on Nuclear and Mitochondrial Protein-Coding Genes. R. Soc. Open Sci. 2018, 5, 171900. [Google Scholar] [CrossRef] [PubMed]
- Wilai, P.; Namgay, R.; Made Ali, R.S.; Saingamsook, J.; Saeung, A.; Junkum, A.; Walton, C.; Harbach, R.E.; Somboon, P. A Multiplex PCR Based on Mitochondrial COI Sequences for Identification of Members of the Anopheles Barbirostris Complex (Diptera: Culicidae) in Thailand and Other Countries in the Region. Insects 2020, 11, 409. [Google Scholar] [CrossRef]
- Kweka, E.J.; Kamau, L.; Munga, S.; Lee, M.-C.; Githeko, A.K.; Yan, G. A First Report of Anopheles Funestus Sibling Species in Western Kenya Highlands. Acta Trop. 2013, 128, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Vezenegho, S.B.; Bass, C.; Puinean, M.; Williamson, M.S.; Field, L.M.; Coetzee, M.; Koekemoer, L.L. Development of Multiplex Real-Time PCR Assays for Identification of Members of the Anopheles Funestus Species Group. Malar. J. 2009, 8, 282. [Google Scholar] [CrossRef] [PubMed]
- Jourdain, F.; Picard, M.; Sulesco, T.; Haddad, N.; Harrat, Z.; Sawalha, S.S.; Günay, F.; Kanani, K.; Shaibi, T.; Akhramenko, D.; et al. Identification of Mosquitoes (Diptera: Culicidae): An External Quality Assessment of Medical Entomology Laboratories in the MediLabSecure Network. Parasit. Vectors 2018, 11, 553. [Google Scholar] [CrossRef]
- Jinbo, U.; Kato, T.; Ito, M. Current Progress in DNA Barcoding and Future Implications for Entomology. Entomol. Sci. 2011, 14, 107–124. [Google Scholar] [CrossRef]
- Paskewitz, S.M.; Wesson, D.M.; Collins, F.H. The Internal Transcribed Spacers of Ribosomal DNA in Five Members of the Anopheles Gambiae Species Complex. Insect Mol. Biol. 1993, 2, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Huynh, L.N.; Diarra, A.Z.; Nguyen, H.S.; Tran, L.B.; Do, V.N.; Ly, T.D.A.; Ho, V.H.; Nguyen, X.Q.; Parola, P. MALDI-TOF Mass Spectrometry Identification of Mosquitoes Collected in Vietnam. Parasit. Vectors 2022, 15, 39. [Google Scholar] [CrossRef] [PubMed]
- Bamou, R.; Costa, M.M.; Diarra, A.Z.; Martins, A.J.; Parola, P.; Almeras, L. Enhanced Procedures for Mosquito Identification by MALDI-TOF MS. Parasit. Vectors 2022, 15, 240. [Google Scholar] [CrossRef] [PubMed]
- Nabet, C.; Kone, A.K.; Dia, A.K.; Sylla, M.; Gautier, M.; Yattara, M.; Thera, M.A.; Faye, O.; Braack, L.; Manguin, S.; et al. New Assessment of Anopheles Vector Species Identification Using MALDI-TOF MS. Malar. J. 2021, 20, 33. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.V.; Bogale, H.N.; Bhalerao, D.; Keita, K.; Camara, D.; Barry, Y.; Keita, M.; Coulibaly, D.; Kone, A.K.; Doumbo, O.K.; et al. High-Throughput Detection of Eukaryotic Parasites and Arboviruses in Mosquitoes. Biol. Open 2021, 10, bio058855. [Google Scholar] [CrossRef] [PubMed]
- Kent, R.J. Molecular Methods for Arthropod Bloodmeal Identification and Applications to Ecological and Vector-Borne Disease Studies. Mol. Ecol. Resour. 2009, 9, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Niare, S.; Berenger, J.-M.; Dieme, C.; Doumbo, O.; Raoult, D.; Parola, P.; Almeras, L. Identification of Blood Meal Sources in the Main African Malaria Mosquito Vector by MALDI-TOF MS. Malar. J. 2016, 15, 87. [Google Scholar] [CrossRef] [PubMed]
- Tahir, D.; Almeras, L.; Varloud, M.; Raoult, D.; Davoust, B.; Parola, P. Assessment of MALDI-TOF Mass Spectrometry for Filariae Detection in Aedes Aegypti Mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0006093. [Google Scholar] [CrossRef]
- Rakotonirina, A.; Pol, M.; Kainiu, M.; Barsac, E.; Tutagata, J.; Kilama, S.; O’Connor, O.; Tarantola, A.; Colot, J.; Dupont-Rouzeyrol, M.; et al. MALDI-TOF MS: Optimization for Future Uses in Entomological Surveillance and Identification of Mosquitoes from New Caledonia. Parasit. Vectors 2020, 13, 359. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, A.L.; Batovska, J.; Webb, C.E.; Lynch, S.E.; Blacket, M.J.; Šlapeta, J.; Parola, P.; Laroche, M. Accurate identification of Australian mosquitoes using protein profiling. Parasitology 2019, 146, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Tandina, F.; Laroche, M.; Davoust, B.; K Doumbo, O.; Parola, P. Blood Meal Identification in the Cryptic Species Anopheles Gambiae and Anopheles Coluzzii Using MALDI-TOF MS. Parasite 2022, 25, 40. [Google Scholar] [CrossRef] [PubMed]
- Dieme, C.; Yssouf, A.; Vega-Rúa, A.; Berenger, J.-M.; Failloux, A.-B.; Raoult, D.; Parola, P.; Almeras, L. Accurate Identification of Culicidae at Aquatic Developmental Stages by MALDI-TOF MS Profiling. Parasit. Vectors 2014, 7, 544. [Google Scholar] [CrossRef] [PubMed]
- Nebbak, A.; Koumare, S.; Willcox, A.C.; Berenger, J.-M.; Raoult, D.; Almeras, L.; Parola, P. Field application of MALDI-TOF MS on mosquito larvae identification. Parasitology 2018, 145, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Nebbak, A.; Willcox, A.C.; Koumare, S.; Berenger, J.-M.; Raoult, D.; Parola, P.; Fontaine, A.; Briolant, S.; Almeras, L. Longitudinal Monitoring of Environmental Factors at Culicidae Larval Habitats in Urban Areas and Their Association with Various Mosquito Species Using an Innovative Strategy. Pest Manag. Sci. 2019, 75, 923–934. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparison of MALDI-TOF MS spectra from legs of Culex mosquitoes. (a) Representative legs MS spectra of Culex species. The name of each species was indicated on the right corner of each spectrum. (b) Composite Correlation Index (CCI) matrix value representing the levels of legs MS spectra reproducibility among Culex specimens of the same species and between species. The 24 specimens from the 12 Culex species, which were selected as reference MS spectra introduced in the MS DB, are shown. The levels of MS spectra reproducibility are indicated in red and blue, revealing relatedness and incongruence between spectra, respectively. The values correspond to the mean coefficient correlation and respective standard deviations obtained for paired condition comparisons. CCI was calculated with MALDI-Biotyper v.3.0 software. (c) MSP dendrogram of MALDI-TOF MS spectra from legs of Culex mosquitoes, from the 24 Culex specimens selected as reference MS spectra. The Culex and Melanoconion subgenus were indicated at the right part. The distance units correspond to the relative similarity of MS spectra. The dendrogram was created by Biotyper v.3.0 software. The same color code was used for specimens of the same Culex species between panels (a) and (c). a.u., arbitrary units; Cx., Culex; MSP, Main Spectrum Profile; m/z, mass-to-charge ratio; SD, standard deviation.
Figure 1.
Comparison of MALDI-TOF MS spectra from legs of Culex mosquitoes. (a) Representative legs MS spectra of Culex species. The name of each species was indicated on the right corner of each spectrum. (b) Composite Correlation Index (CCI) matrix value representing the levels of legs MS spectra reproducibility among Culex specimens of the same species and between species. The 24 specimens from the 12 Culex species, which were selected as reference MS spectra introduced in the MS DB, are shown. The levels of MS spectra reproducibility are indicated in red and blue, revealing relatedness and incongruence between spectra, respectively. The values correspond to the mean coefficient correlation and respective standard deviations obtained for paired condition comparisons. CCI was calculated with MALDI-Biotyper v.3.0 software. (c) MSP dendrogram of MALDI-TOF MS spectra from legs of Culex mosquitoes, from the 24 Culex specimens selected as reference MS spectra. The Culex and Melanoconion subgenus were indicated at the right part. The distance units correspond to the relative similarity of MS spectra. The dendrogram was created by Biotyper v.3.0 software. The same color code was used for specimens of the same Culex species between panels (a) and (c). a.u., arbitrary units; Cx., Culex; MSP, Main Spectrum Profile; m/z, mass-to-charge ratio; SD, standard deviation.
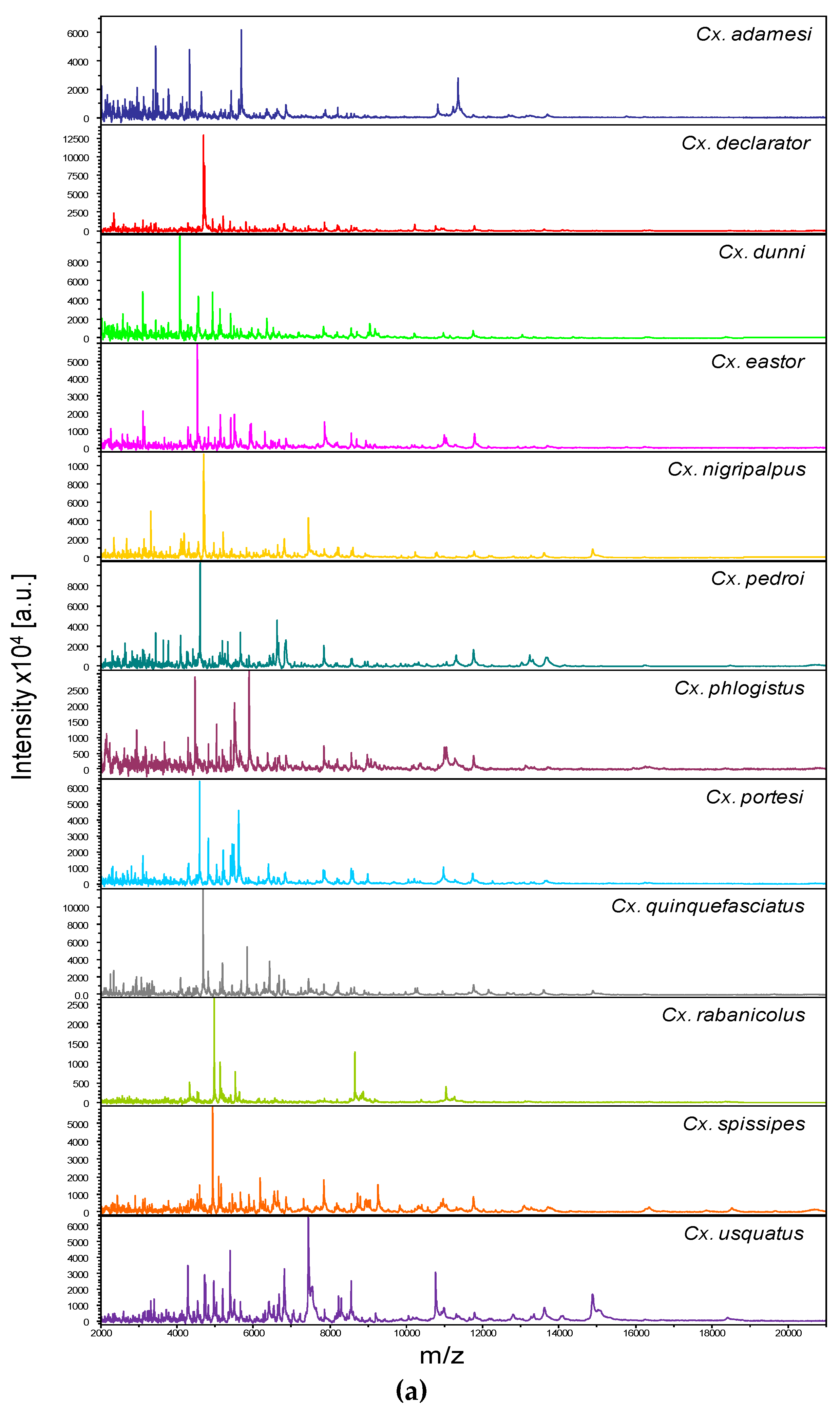
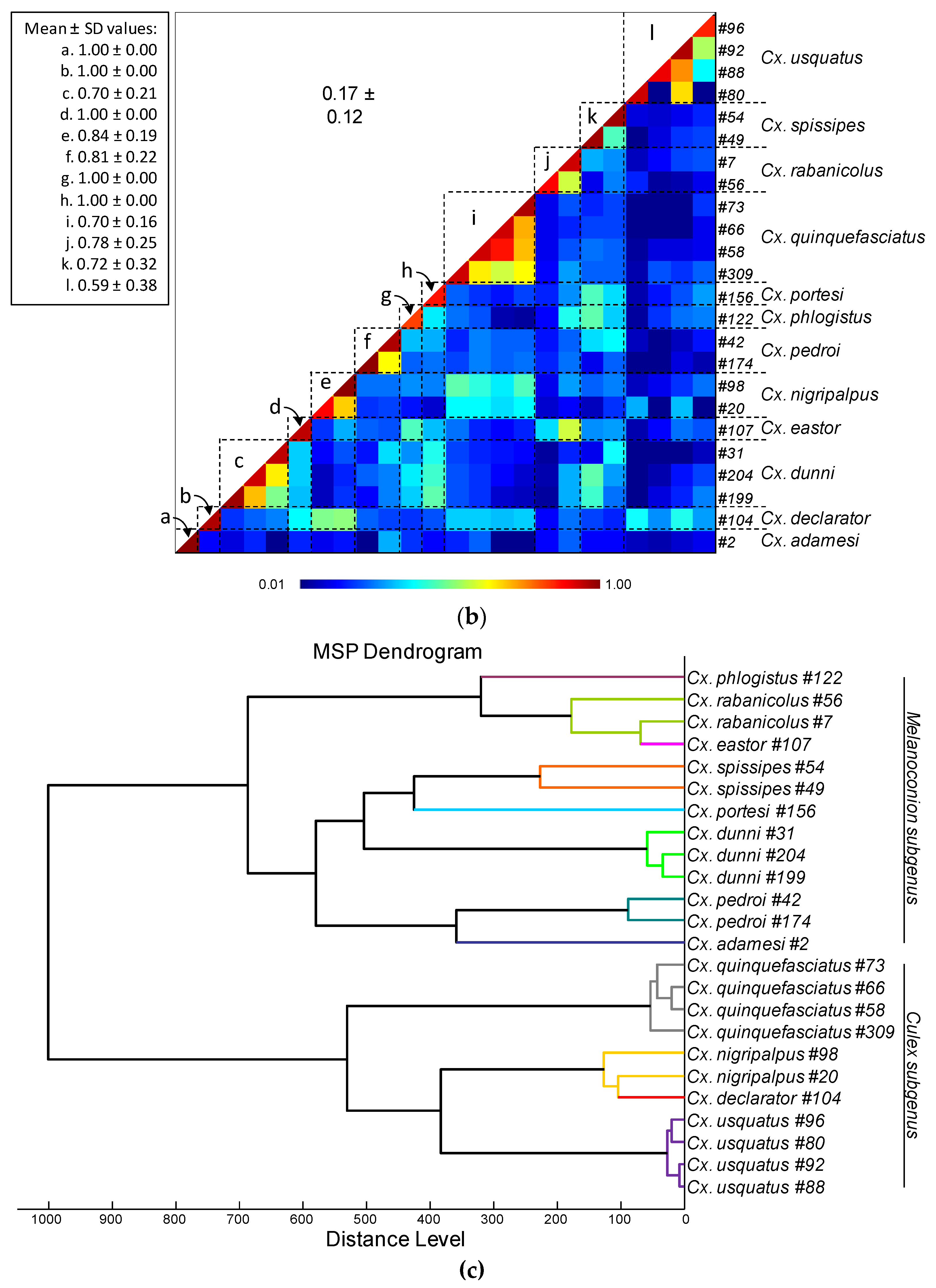
Figure 2.
Comparison of MALDI-TOF MS spectra from thoraxes of Culex mosquitoes. (a) Representative thoraxes MS spectra of Culex species. The name of each species was indicated on the right corner of each spectrum. (b) Composite Correlation Index (CCI) matrix value representing the levels of thoraxes MS spectra reproducibility among Culex specimens of the same species and between species. The 34 specimens from the 13 Culex species, which were selected as reference MS spectra introduced in the MS DB, are shown. The levels of MS spectra reproducibility are indicated in red and blue, revealing relatedness and incongruence between spectra, respectively. The values correspond to the mean coefficient correlation and respective standard deviations obtained for paired condition comparisons. CCI was calculated with MALDI-Biotyper v.3.0 software. (c) MSP dendrogram of MALDI-TOF MS spectra from thoraxes of Culex mosquitoes, from the 34 Culex specimens selected as reference MS spectra. The Culex and Melanoconion subgenus were indicated at the right part. The distance units correspond to the relative similarity of MS spectra. The dendrogram was created by MALDI-Biotyper v.3.0 software. The same color code was used for specimens of the same Culex species between panels (a) and (c). a.u., arbitrary units; Cx., Culex; MSP, Main Spectrum Profile; m/z, mass-to-charge ratio; SD, standard deviation.
Figure 2.
Comparison of MALDI-TOF MS spectra from thoraxes of Culex mosquitoes. (a) Representative thoraxes MS spectra of Culex species. The name of each species was indicated on the right corner of each spectrum. (b) Composite Correlation Index (CCI) matrix value representing the levels of thoraxes MS spectra reproducibility among Culex specimens of the same species and between species. The 34 specimens from the 13 Culex species, which were selected as reference MS spectra introduced in the MS DB, are shown. The levels of MS spectra reproducibility are indicated in red and blue, revealing relatedness and incongruence between spectra, respectively. The values correspond to the mean coefficient correlation and respective standard deviations obtained for paired condition comparisons. CCI was calculated with MALDI-Biotyper v.3.0 software. (c) MSP dendrogram of MALDI-TOF MS spectra from thoraxes of Culex mosquitoes, from the 34 Culex specimens selected as reference MS spectra. The Culex and Melanoconion subgenus were indicated at the right part. The distance units correspond to the relative similarity of MS spectra. The dendrogram was created by MALDI-Biotyper v.3.0 software. The same color code was used for specimens of the same Culex species between panels (a) and (c). a.u., arbitrary units; Cx., Culex; MSP, Main Spectrum Profile; m/z, mass-to-charge ratio; SD, standard deviation.
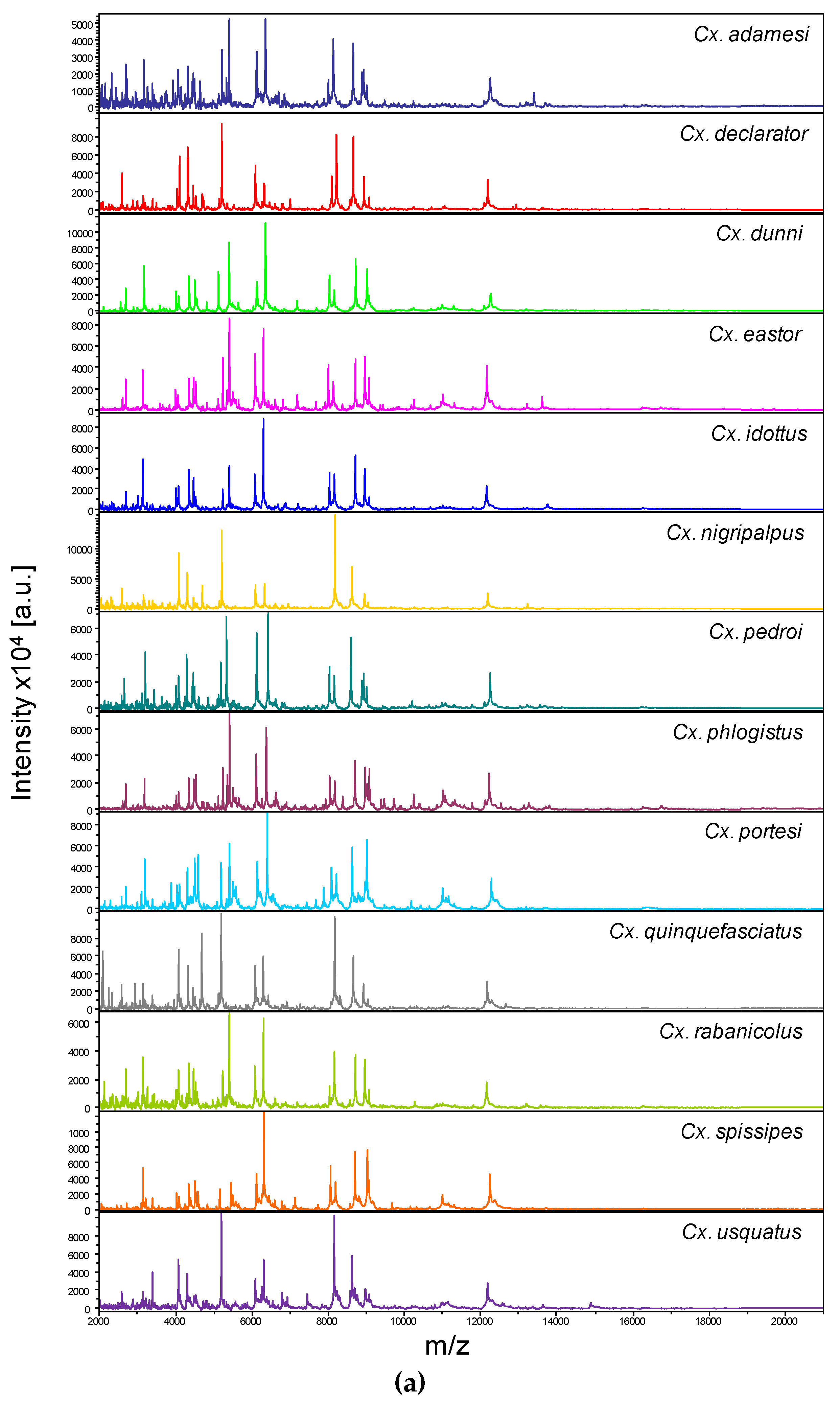
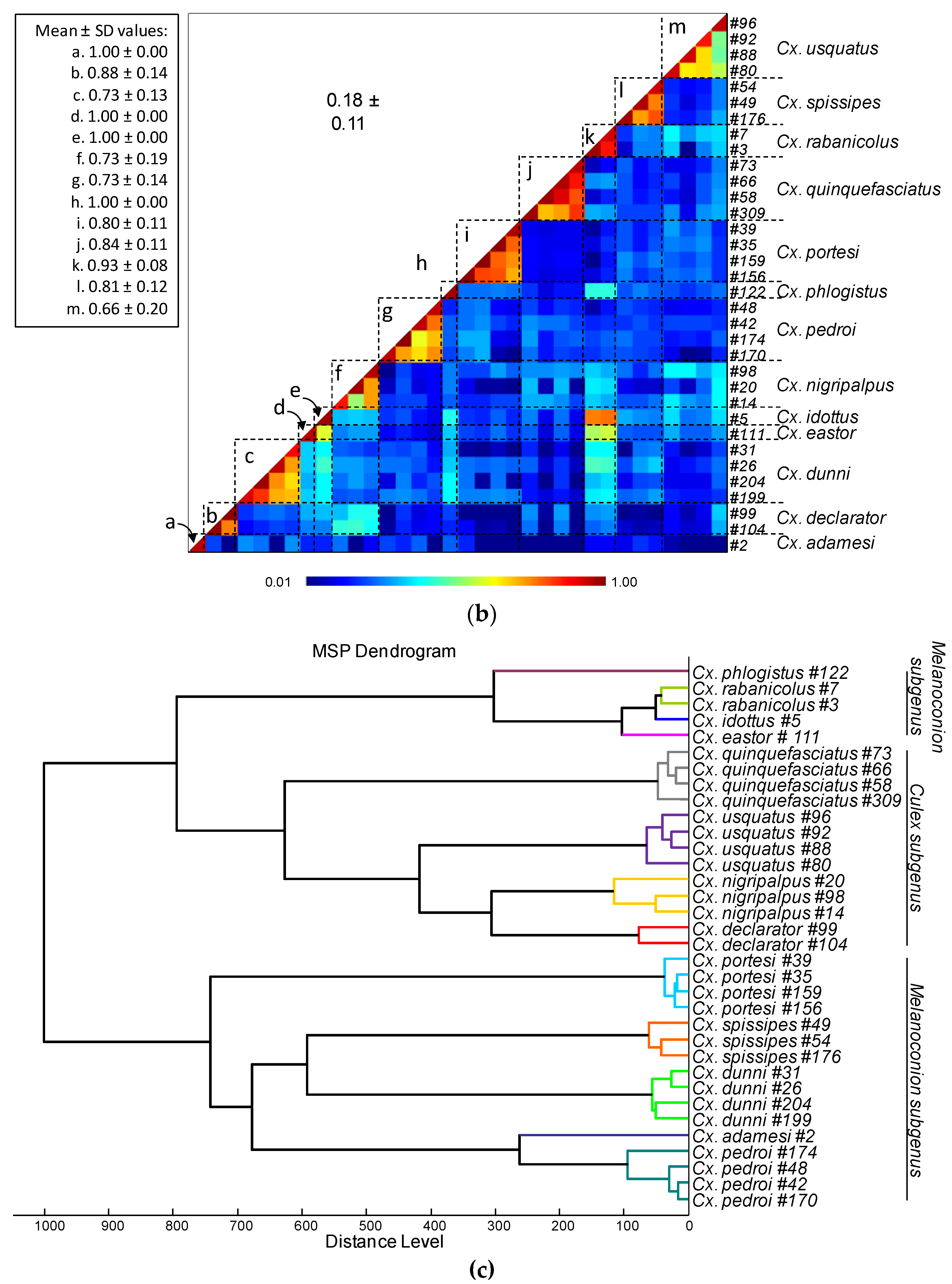
Figure 3.
LSVs from query against homemade MS reference database. MS spectra of legs (a) and thoraxes (b) from Culex mosquitoes are presented. Dashed line represents the threshold value (LSV ≥ 1.8), for relevant identification. a.u., arbitrary units; LSV, log score value.
Figure 3.
LSVs from query against homemade MS reference database. MS spectra of legs (a) and thoraxes (b) from Culex mosquitoes are presented. Dashed line represents the threshold value (LSV ≥ 1.8), for relevant identification. a.u., arbitrary units; LSV, log score value.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



