
Submitted:
07 February 2023
Posted:
08 February 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
In the recent days, lipid nanoparticles have been successfully emerged as one of the most advanced technology for highly efficient in vivo delivery of exogenous mRNA, especially for delivery of COVID-19 vaccines. For the vaccines to be successful or protective, they require highly efficient mRNA delivery systems. However, developing effective, translatable vaccines with better safety against some of the SARS‐CoV‐2 variants is still a challenge. Lipid nanoparticles (LNPs) are composed of four different types of lipids including ionizable lipids, helper or neutral lipids, cholesterol and polyethylene glycol (PEG) attached lipids. In this review, we present recent advancements and insights in designing the advanced LNPs and their composition and properties, with a subsequent discussion on the development of COVID-19 vaccines. In particular, as the ionizable lipids are most important drivers for complexing the mRNA and in vivo delivery, the role of ionizable lipids in mRNA vaccines discussed in detail. Furthermore, the use of LNPs as effective delivery vehicles for vaccination, genome editing, and protein replacement therapy were discussed. Finally, expert opinion of LNPs for mRNA vaccines were discussed which might address the future challenges in the development of mRNA vaccines employing highly efficient LNPs using novel set of ionizable lipids.
Keywords:
vaccine
; mRNA
; COVID 19
; lipid nanoparticles
1. Introduction
Advances in messenger RNA (mRNA)-based delivery technologies have demonstrated therapeutic potential in a wide range of biomedical applications, including protein replacement therapies, vaccines, cellular reprogramming, and cancer immunotherapies among others.[1,2,3] To achieve ideal therapeutic benefits, mRNA molecules must be safeguarded from degradation andreach specific target cells to produce proteins of interest. However, targetability, stability, and endosomal escape remain key challenges for mRNA delivery systems, emphasizing the need for safe and effective mRNA delivery.[4] Numerous biocompatible, biodegradable lipids, polymers, and protein-based derivatives have been used in the development of mRNA-based therapeutics.[5]
In particular, lipid nanoparticles (LNPs) have emerged as a thoroughly investigated category of vehicles in the biopharmaceutical industries as potential delivery systems for various nucleic acid therapeutics including oligonucleotides. Compared to viral vectors, LNPs are advantageous for gene therapy due to their low or no immunogenicity, high payload, ease of synthesis, and scale-up. Besides, LNPs are tailormade using lipid to mRNA ratios, can be surface functionalized with ligand of interest for various cell targets and disease-related applications. LNPs, which are gaining attention as an important component of COVID-19 mRNA vaccines, are essential for preserving and delivering the payload (mRNA) to specific targeted cells.[4] Notably, LNPs have successfully reached the clinics for the delivery of various biologics including mRNA; in particular, mRNA-LNP vaccines against COVID-19 are currently in clinical use, representing a ground-breaking path for mRNA-based therapies.[6,7]
Recent years have seen a phenomenal expansion of RNA therapies, which has greatly profited from decades of research on LNPs. The delivery of RNAs to target cells by clinically translatable LNPs opens up new avenues for treating a variety of life-threatening diseases, such as Onpattro® (an RNA interference drug that was the first FDA-approved LNP-nucleic acid therapeutic for the treatment of polyneuropathy),[8] and COVID-19 vaccine emergency use authorization (Pfizer-BioNTech and Moderna mRNA-LNPs).[9] Before Onpattro® was approved in 2018, scientists investigated more than a decade testing different ionizable lipids and LNP-based formulations. The success of MC3 (ionizable lipid used for Onpattro® product by Alnylam® Pharmaceuticals) has reignited the interest in RNA delivery and greatly accelerated the clinical development of other LNP-based RNA therapeutics, especially mRNA vaccines [including two recent breakthrough LNP-based mRNA vaccines mRNA-1273 (Moderna) and BNT162b2 (BioNTech) for COVID 19].[10,11] However, still it is a big challenge to develop more efficient vaccines against some of newly emerging of the SARS-CoV-2 variants.[7]
In this review, we present an overview of the current knowledge about LNP design composition and characteristics, focusing on their pharmacological use. We then discuss the role of ionizable lipids in mRNA delivery. Furthermore, we present a landscape of classical methods to the importance of latest microfluidics technology for LNPs preparation. Finally, we present insights of mRNA release from LNP, its pharmacokinetics on immunogenicity and provided future perspectives for LNPs and mRNA therapeutics.
2. Role and Importance of LNPs Structural Composition in Designing mRNA Therapeutics
LNPs are generally spherical systems with at least one lipid bilayer and an aqueous interior compartment, which provide several benefits, including formulation simplicity, self-assembly, biocompatibility, high bioavailability, and the potential to transport large payloads.[12] Due to these factors, LNPs are the most recommended formulation class of FDA-approved nanomedicines.[13] LNPs have been thoroughly investigated and successfully used in clinics for the delivery of various macromolecules and small-molecule drugs, including nucleic acids.
Unlike classical bilayered liposomes, LNPs have a complex internal architecture with enhanced physical stability due to their structural properties. LNPs' physicochemical properties such as nanoparticle size, charge, and ligand surface functionalization, which can be altered during the synthesis, which could allow for enhanced in vitro, in vivo stability and impart specialized nucleic acid delivery functions.[14,15] Typically, upon administration, LNPs are internalized by host cells, the encapsulated mRNA cargo will be delivered inside the cytosol and mRNA sequences are further translated into targeted protein by the ribosome of host cells (discussed in detail in next sections).[16] One of the key obstacles of mRNA vaccines is that naked mRNA is quickly degraded upon administration by enzymes ribonucleases (RNase), reducing intracellular stability and further minimizing translation efficiency. Despite mRNA's immense therapeutic applicability for pharmaceutical uses, intracellular targetability remains difficult due to its high molecular weight, polyanionic nature, and intrinsic chemical instability. Henceforth, LNPs and other lipid-based nanomedicine systems play a key role in effectively protecting and transporting exogenous mRNA to targeted cells.
2.1. Material Aspects and Structural design of LNPs for mRNA Therapeutics
The following four lipid components (please see Figure 1 and Table 1 for more details) are majorly used in the synthesis of LNPs - i) ionizable lipids mainly needed to complex with mRNA; (ii) helper lipid to improve the properties of LNPs (stability) and delivery efficiency; (iii) cholesterol to impart structural stability of the vehicle; and (iv) surface PEGylation with poly(ethylene glycol) (PEG) or PEG-derivatives to minimize the host-immune recognition and enhance the systemic circulation.[17,18,19] Among all these lipids, ionizable lipids are the most essential components of the majority of LNPs acting as a main driver to deliver mRNA through high mRNAs encapsulation, enhance stability and pH-sensitive in vivo delivery.[19] The role of each component of LNPs in mRNA delivery is discussed below in detail.
i).Ionizable cationic lipids: Ionizable lipids for example OF-Deg-Lin and FTT5 (Table 1) are protonated at low pH which makes them positively charged inside the cell and uncharged/neutral at physiological pH in the bloodstream.[19,20] These positively charged lipids stabilize the nucleic acid and prevent degradation by nucleases.[6] In addition, ionizable lipids facilitate in the cellular uptake of the LNPs and aid in endosomal escape by acquiring the positive charge in the endosomal pH (discussed in detail in next sections).[19,21]
ii). Neutral/helper phospholipids: In addition to cationic ionizable lipids, LNPs also comprise helper lipids, such as phospholipids such as phosphatidylcholine (e.g. 1,2- distearoyl- sn-glycero-
3- phosphocholine-DSPC and 1,2-Dioleoyl-sn-glycero-3-phosphoethanolamine-DOPE, respectively). These phospholipids improve the properties of LNPs such as particle stability, membrane integrity, and delivery efficacy.[4] These lipids (e.g., DOPE, DSPC) stabilizes the LNPs by forming different geometries using acyl chains, and enhances the transfection efficiency by promoting membrane fusion.[6] They destabilize the endosomal membrane and promote the endosomal escape of LNPs, the ratios of various types of lipid composition(s) influence the effectiveness of the nanoparticles. LNPs with higher amounts of helper lipids are used for mRNA delivery; mRNA delivery is enhanced in DOPE than in DSPC as it can undergo a conformational change from a stable lamellar phase to an unstable hexagonal phase, resulting in membrane fusion, whereas the phosphocholine-containing lipids inhibit membrane fusion-mediated endosomal escape.[4,22]
iii). Cholesterol: Emerging evidences have shown incorporation of cholesterol (which is hydrophobic and rigid) help in filling the gaps between lipids within the vesicle membrane, and has the potential to modulate membrane integrity and stiffness, which in turn can increase particle stability.[4,6,23] Besides, incorporating the cholesterol showed improved delivery efficacy and increased the circulation half-life of nanoparticles and enhances the transfection efficiency of LNPs by promoting membrane fusion and endosomal release.[4,13,24] For example, Patel et al. demonstrated that incorporation of cholesterol analogues with C-24 alkyl phytosterols into LNPs (eLNPs) could enhance gene transfection due to higher cellular uptake and retention followed by steady release of encapsulated mRNA.[25] An increase in cholesterol percentage lowers the transitional temperature of the membranes of LNPs.[25] Moreover, the biocompatibility of the LNPs is enhanced as cholesterol is one of the major components of biological membranes.[26]
iv). Lipid anchored polyethylene glycol (PEG) constructs: PEG-lipids constructs (PEG molecule attached to alkyl chains) acts as an anchor inside the LNP layer, incorporation of PEG lipids in LNPs inhibit opsonization by serum proteins as well as reticuloendothelial clearance, both of which are particularly important for increasing the biodistribution.[27,28] In addition, incorporating PEG-lipid into LNPs can affect the size, surface charge of the nanoparticles and improve surface properties by restricting access through steric hindrance to the surface.[6,29] PEG-lipids can further prevents the blood plasma protein surface adsorption, thereby increasing the circulation lifetime in the bloodstream and minimize aggregation behavior.[4] Also, PEG-lipid composite coating on LNPs facilitate the receptor mediated cellular uptake by exchanging with the serum proteins including ApoE.[23,25] Furthermore, PEG-lipids can also prevent endosomal escape by blocking the interaction between liposome and endosomal membrane sterically and electrostatically.[21,23,30,31].
2.2. Role of Ionizable Lipids in mRNA Delivery
Among four types of lipids, ionizable lipids are the most important class of lipids which complex with the negatively charged mRNA and responsible for endosomal escape of mRNA subsequently releasing in the cytosol. Thanks to the unique properties of ionizable lipids and their pH-dependent surface charge. Notably, these lipids are positively charged in the low pH and are neutral in the physiological pH environment.[18,37] Hence, at the physiological pH the neutral lipids have least interaction with the anionic cell membrane which enhances the biocompatibility of the LNPs. At the same time, inside the endosome (acidic pH), the cationic ionizable lipids interact with the anionic endosomal phospholipids to form cone shaped ion pairs known as inverted hexagonal HII phase. These structures distort the bilayer of LNPs and facilitate the disruption of membrane and release of payload into the cytosol.[4] Literature evidences showed that, ionizable lipids are broadly classified into five types; i) unsaturated, ii) multi-tail, iii) polymeric, iv) biodegradable, and v) branched tail ionizable lipids as shown in Figure 2.[37]
Unsaturated ionizable lipids constructs have increased cis double bonds in tail which makes the bilayer structure unstable and helps to easily form inverted hexagonal structures.[38] Hence, it enhances the destruction of membranes and thereby facilitates the release of cargo. Some of the examples of such lipids are OF-02 and A6.[36,37] Multi-tail ionizable lipids, as the name suggests, have more than two tails (e.g., C12-200). These multi-tail ends help to form non-bilayer, cone shaped structures that amplifies the endosomal destruction and promotes highly efficient cargo release compared to the two tailed ionizable lipids.[37,39] On the other hand, polymeric ionizable lipids are synthesized by replacing the free amine groups in cationic polymers with alkyl tails.[37] These lipids stimulate the formation of cone shaped structures through hydrophobic interaction. G0-C14 is a polymeric ionizable lipid used for lipid nanoparticle formation that has application in RNA therapeutics against cancer.[37,40] However, the main disadvantage of polymeric ionizable lipids is that increasing the complexity of the lipid nanoparticle formation due to unwanted substitution compounds even after purification. Moreover, the poly cationic core and non-biodegradable polymeric structure increases the cytotoxicity.[4] One of the approaches to develop biodegradable ionizable lipids is the addition of biocleavable ester groups within in the hydrophilic alkyl chains of the lipid structure.[41,42,43] These ester groups need to be easily hydrolyzed in the cytosol but should be stable at physiological pH. For example, 304O13 (Figure 2) is a biodegradable lipid that showed similar potency to its non-biodegradable analogue C12-200 and has lower toxicity even at higher doses.[37,42] A problem faced by these lipids is the lower potency of gene (protein) expression when compared with their non-biodegradable analogue as a result of rapid hydrolysis. However, by using the lipids containing secondary esters (i.e. composed of ethanolamine headgroup, and a primary ester at C8 position along with the secondary esters in the second lipid tail) resulted to give ideal balance of in vivo protein expression and clearance of the lipids.[37,44] Increased branching of the tails can also enhance the lipid performance, e.g., acrylate based ionizable lipids showed increased efficacy than the methacrylate tailed ones.[45,46] These lipids showed higher cationic charges at endosomal acidic pH that enhanced the endosomal escape, and also due to the wider lipid tails, the cone structure formation was easier compared to other lipids. Further, branched tail ionizable lipids are the ionizable lipids used to deliver the mRNA in the case of COVID-19 vaccines.[18] FTT5 is a one of highly studied ionizable lipid that showed higher mRNA delivery for protein supplementation application.[37,47] However, how these branched tails affect quality of LNPs has been studied poorly due to lack of commercial availability of these branched tail components.[18] These examples demonstrate the importance of ionizable lipids in LNPs based therapeutics delivery.
More recently, Chen K et al. 2022 reported a novel set of unique ionizable lipids termed 4N4T to formulate new series of LNPs known as 4N4T-LNPs by squeezing the lipid organic phase with aqueous mRNA solution into microfluidic chip.[48] Compared to the approved SM-102-LNPs, 4N4T-LNPs have greater mRNA translation efficiency against SARS-CoV-2 and its variations, including Delta and Omicron. Overall, the study revealed that 4N4T-based lipid delivery technology might contribute to bring advanced, efficacious mRNA therapeutics for infectious diseases.[48]
Interestingly, Hashiba et al. 2022 reported the branching of ionizable lipids impact on enhancement of stability, fusogenicity, and functional mRNA delivery. Branched lipids (CL4F 8-6) with a high level of symmetry contributed ideal features for efficient intracellular distribution and stable formulations, providing new insights into rational lipid design and successful gene therapy applications.[18] Riley S. R et al. 2021 developed a library of ionizable LNPs for in utero mRNA delivery to mouse fetuses, demonstrated prenatal LNP delivery platform for erythropoietin (EPO) mRNA to hepatocytes in the fetal circulation for protein replacement therapy.[49]
3. Moving beyond Classical Lipid Carriers to Specialized LNPs Design
The classical liposomes, an early version of LNPs are versatile nanocarriers that deliver both hydrophobic and hydrophilic therapeutic agents. In recent years, a lot of attention has been paid to developing subsequent generations of LNPs, such as solid LNPs (SLNPs), nanostructured lipid carriers (NLCs), and cationic lipid-nucleic acid complexes with more structural complexity and greater physical stability of the delivery carrier.[6,50] Notably, LNPs take advantage of spontaneous formation or self-assembly due to the interactions between the charged groups in the lipidic mixture and charged nucleic acids. Intermolecular interactions aid in the formation of spherical nanostructures from lipid components. In general, various methods have been used to formulate LNPs, including nanoprecipitation, thin-film hydration, extrusion, homogenization, microfluidic mixing, and others. Among the existing methods, the latest approach is microfluidic mixing, since it produces uniform, size-controlled, stable, and reproducible LNPs. In addition, recent advancements have shown microfluidic process is one of the most reliable method compared to conventional methods.[14]
The development of more adaptable, highly effective, and biocompatible systems is made possible by ongoing efforts to synthesize and evaluate a variety of LNPs by chemically modifying their molecular architectures. The rapid development of mRNA vaccines would not have been conceivable without advances in screening the latest lipid constructs and LNP technologies to deliver nucleic acids. In particular, cationic and ionizable lipids are preferred because of their inherent tendency to self-assemble into LNPs with nucleic acids via intermolecular interactions, which will help in the efficient delivery of the payload.[51]
3.1. Microfluidics over Other Conventional Methods
The traditional lipid thin film hydration method was used for the preparation of liposomes and LNP, however this approach has several disadvantages, including heterogeneous particle size distribution due to the formation of nanoparticles of varied particle sizes. Henceforth, a subsequent size tuning approach is necessary to formulate uniform, homogenous LNPs. Besides, the application of the post processes, including extrusion, sonication, and others may result in morphological changes to the final nanoparticle product.[52] To address these limitations, recently researchers developed microfluidics tool, which has simplified production processes.[53]
Microfluidics tool offer many advantages with tunable optimization parameters (flow rate, controlled mixing, temperature, time), which help in manufacturing precise LNPs to enhance the transfection efficiency.[54,55] Importantly, the flow of ingredients and the speed of mixing during the rapid production process control LNP production processes. Scale up is also feasible with minimum time and cost, which adds a major advantage in the formulation development especially in emerging situations like Covid-19 pandemic for easy clinical translation.[56] Numerous articles published over the last five years demonstrated the importance of microfluidics approach for the synthesis of LNPs with significant benefits for the precise and rapid manufacturing. In addition, the recently developed mRNA based LNPs for COVID-19 vaccine by Pfizer/BioNTech and Moderna is produced in large scale using this microfluidic technique, which highlights the importance of the technique in the field of vaccine production for usage in emergency situations.[57]
Although, predictable and rational design of LNPs for nucleic acid delivery to extrahepatic tissues remains challenging (LNPs resemble as very-low-density lipoprotein, adsorb apolipoprotein E in blood plasma, resulting in preferential accumulation in the liver and uptake into hepatocytes), which must be solved in order to realize the full potential of mRNA LNP technology for wide therapeutic developments.[58] However, such mechanisms are effective in treating in liver diseases, including cancer. A recent interesting new technology termed as selective organ targeting (SORT) was developed, whereby nanoparticles are methodically tailored to deliver therapeutic compounds to both hepatic and extrahepatic regions.[59] Wang X et al. 2022 reported the SORT LNPs include tunable SORT molecules that target the payload to the liver, lungs, and spleen of mice following intravenous administration. The engineered SORT LNPs able to target specific cells and organs in the body via passive, active, and endogenous targeting methods, each of which necessitates a different set of design requirements.[58] The authors published procedures for the preparation of 4A3-SC8- and MC3-based liver, lung, and spleen SORT LNPs using three techniques (pipette mixing, vortex mixing, and microfluidic mixing) to provide readers with methods for small-, medium-, and large-scale manufacturing (see Figure 3).
4. Applications of LNPs in mRNA Delivery
Before discussing LNPs applications, a brief discussion on mRNA and its therapeutic potential is placed here. Since its first isolation in 1961, mRNA (that encodes the protein of interest) research has taken several paths, which made us understand its diversified functions and modification-mediated potential for therapeutic applications.[1], [60] As a result of the COVID-19 pandemic, nucleic acid therapeutics (NATs), particularly mRNA vaccines, potentials have been enabled for emerging infectious diseases. The translation of host genetic information (DNA) into proteins by ribosomes in the cytoplasm is mediated by mRNA (Figure 4).
Use of mRNA as Prophylactics and Therapeutics
Despite use of DNA vaccines, RNA-based vaccines have become popular in the 21st century. Interestingly, mRNA vaccine technology purely depends on the genetic code of the virus, since the SARS-CoV-2 genetic sequence was published,[61] virally generated self-amplifying RNA and non-replicating mRNA are being used abundantly for vaccine development. In fact, during the COVID-19 pandemic, mRNA vaccines have become the only source of vaccines for preventing the rapid spread of the disease. While several pharmaceutical companies have been explored the different vaccine technologies against COVID-19, two vaccines have been approved (Initially for emergency use and further full approval) Pfizer/BioNTech and Moderna’s Spikevax by FDA (Table 2).[62] Both use the complete sequence of spike and receptor-binding domains of SARS-CoV-2 as target antigens. Although, mRNA-based preclinical studies have been reported, since 1990. Its poor-stability as naked mRNA and tremendous capacity to induce innate immune response (particularly double stranded RNA) has halted the progress of RNA-based therapeutics. The emergence of advanced nanotechnologies in the protection and targeted delivery of water-soluble molecules have shaded a light on the development of the LNP-mRNA vaccines. Nucleic acid delivery to the cytosol without hampering the cell wall integrity and degradation by nucleases is a rate liming step for targeted delivery. Despite mechanical and viral particle mediated targeted delivery, chemical based targeted delivery via LNPs have been the ease of process and less toxic, less immunogenicity. During the start of the decade several clinical trials have been initiated on mRNA therapeutics, which covers a range of different types of diseases, such as cancer, infectious diseases, protein replacement therapy, and others. Correspondingly, promising results have been shown in terms of diseases alleviation, reduction of pathogenicity, and others. Interestingly, several diseases or disorders are targeted by using mRNA therapeutics. For the proof-of-concept studies, in vitro transcription produced mRNA is more suitable (Figure 5). To characterize the efficiency of mRNA in vitro several transfection reagents are available for facilitate the mRNA translation and targeted protein production, although not efficient as targeted delivery systems. The Table 2 shows the most successful mRNA vaccines. As explained earlier, a suitable (perfect mix) vehicle is needed for the mRNA vaccine delivery. LNPs have become the one of the most characterised mRNA delivery vehicles.
LNP Role in mRNA Delivery
Using lipid-encapsulated forms of sequence-optimized mRNA vaccines shown strong immunity (both in preclinical and clinical) against infectious disease targets like the flu virus, Zika virus, rabies virus, and others, especially in the last few years.[63] Of note, the LNPs act as potential vaccine adjuvants,[62] which helps in shaping the antigen-specific immunity.[64] Despite several components of the LNP, ionizable lipids have a prominent role, including the formation of LNPs and targeted delivery of mRNA into the cytosol. The ionizable lipid imparts the neutral charge to the LNPs at physiological pH and ionizes at lower pH in the endosome/lysosome followed by the delivery of intact mRNA to the cytoplasm, where it can then be translated into the encoded protein. Despite mRNA, LNPs also induce the innate immune signalling pathways that aided in the adaptive immune responses. Like other potential adjuvants, LNPs (based on the composition) also induces activation of several signalling pathways, including, nuclear factor kappa B, interferons, inflammasome activation, and other.[49]
In the future, it will be important to compare and understand the immune pathways that are activated by different mRNA vaccine platforms, to improve current approaches based on these mechanisms, and to start new clinical trials against more disease targets.
Salient Features of LNP-Based Therapeutics[2], [4], [62]
- Naked mRNA is not suitable for the therapeutic purpose, as it rapidly degraded by extracellular RNases. Several nanotechnology platforms have been set and optimized for mRNA targeted delivery.
- As mRNA is thermolabile, LNPs would enhance its stability and half-life at room-temperature, and also useful for avoiding vaccine cold-chain.
- Modifications of the type of delivery system (carrier molecules) rule out the organ specific mRNA delivery (for lung, spleen, liver, etc.) and its in vivo half-life.
- Composition of LNP could decide the type of immune response induces.
- LNPs acts as adjuvants system for mRNA vaccines.
- The stable LNPs make the less dose of mRNA to work.
- Composition of LNPs also decides the number of booster doses required, if it admixed with suitable adjuvant.
- Moreover, for chronic treatments, multiple administration via different routes of administration is possible.
Mechanism of mRNA Vaccines
Even though needle free route of administration is long awaited[65]-[67], mRNA encapsulated by LNPs are administered mostly by intramuscular route, and several study results support this route of administration[68]-[70] ;. Upon injection, local innate immune cells are recruited to the site of injection followed by the uptake of LNPs by innate immune cells[71] . Once LNPs reaches the cytosol, where lower pH enables the LNPs to release mRNA followed by the translation of targeted antigens. The antigens are presented to the T helper cells (CD4+ T cells) and release the several cytokines and chemokines, which further helps in the shaping the antigen-specific immune responses, including B cell differentiation and plasma cell development.[72] The two successful vaccines (Pfizer/BioNTech and Moderna’s Spikevax) have confirmed this mechanism in both preclinical and clinical studies.[73]
Salient Features of mRNA Based Therapeutics.[2], [4], [62]
Figure 6.
mRNA vaccine development. The figure illustrates the sequential steps, from design to preclinical studies, involved in the mRNA vaccine development.
Figure 6.
mRNA vaccine development. The figure illustrates the sequential steps, from design to preclinical studies, involved in the mRNA vaccine development.
Expert Opinion
Since its discovery, NATs (siRNA) started seeing light in twentieth century by the approval of patisiran (2019) and mipomersen (2013). Even though, NATs have been used in 19th century, the COVID-19 pandemic more witnessed its potential for public health in protecting from diseases. However, the delivery or targeted delivery of nucleic acid therapeutics is a major concern for maintaining it in stable condition until it reachs targeted site to prevent from off target side effects.
Since 1995, the approval of first liposomal drug delivery system, i.e., Doxil, several nanomedicine platforms for drug delivery have been emerged, such as micelles, lipid nanocapsules, solid LNPs lipid nanoparticles, and many other polymeric nanoparticles. Interestingly, few polymers (e.g., Poly (lactic-co-glycolic acid) (PLGA), Polyethyleneimine (PEI), chitosan, and others) that have been used to deliver the NATs, particularly DNA vaccines, have been shown significant results in both preclinical and clinical studies.[76]
Nevertheless, LNPs succeeded to protect NATs degradation and site-specific delivery. Therefore, NATs and LNPs have been the way out in targeting many diseases, which does not have any acceptable, approved, and available medical alternatives.
Despite efficacy of mRNA vaccines, the mutation rate of SARS-CoV-2 is alarming the mRNA technology to focus more on the multi-antigenic vaccines, which could prevent the virus escape, induced by the low mutations, from the host immune response. Similarly, the use of potential vaccine adjuvants also should be considered, which not only enhances the antigen-specific immune response, but it also helps for the dose, cost, scalability, and mass production of mRNA vaccines.[64] On the other hand, unlike DNA vaccines, mRNA vaccines always require ultra-cool temperature or cold-chain cycle (-20°C– - 80°C) to deliver the low-income countries. Despite mRNA vaccine efficacy, the adverse effects should not be overlooked. Even though, the less incidence of Myocarditis and pericarditis have been reported, the incidence rate in young adults with age range 18-30 years are considerable.[74] Furthermore, either with booster doses or after bivalent immunization, there was a higher rate of inability to work and use of PRN (pro re nata) medications in healthcare workers are reported.[75] Therefore, focused research for findings the reasons for these adverse reactions should be more encouraged.
Unanswered questions
- Is the ionizable lipids deciding the fate of immune response against mRNA encoded protein. Changing the ionizable lipid composition will vary the type of immune response, B or T cell response. Perhaps some mRNA vaccines show T cell-mediated protection without having notable neutralizing antibody inducing capacity.
- As documentation of evidence showing the mRNA vaccine induced local and system adverse events,[77] are these events are related to the vaccine or presence of minute amounts of dsRNA.
- Several new strategies have been developed in codon optimization by keeping the mRNA template intact, are these lead to transient immunogenicity (if not immunosilent) rather than durability as they lack naïve uridine or modified uridines.
- Do we have enough data to support the best route of mRNA vaccine administration for durable immune responses with less boosters?
References
- Sahin, U.; Karikó, K.; Türeci, Ö. , mRNA-based therapeutics — developing a new class of drugs. Nature Reviews Drug Discovery 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M. J.; Porter, F. W.; Weissman, D. , mRNA vaccines — a new era in vaccinology. Nature Reviews Drug Discovery 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, P. S.; Rudra, A.; Miao, L.; Anderson, D. G. , Delivering the Messenger: Advances in Technologies for Therapeutic mRNA Delivery. Molecular therapy : the journal of the American Society of Gene Therapy 2019, 27, 710–728. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. , Lipid nanoparticles for mRNA delivery. Nature Reviews Materials 2021, 6, 1078–1094. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, K. J.; Webber, M. J.; Anderson, D. G. , Materials for non-viral intracellular delivery of messenger RNA therapeutics. J Control Release 2016, 240, 227–234. [Google Scholar] [CrossRef]
- Tenchov, R.; Bird, R.; Curtze, A. E.; Zhou, Q. , Lipid Nanoparticles─From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano 2021, 15, 16982–17015. [Google Scholar] [CrossRef]
- Chen, K.; Fan, N.; Huang, H.; Jiang, X.; Qin, S.; Xiao, W.; Zheng, Q.; Zhang, Y.; Duan, X.; Qin, Z.; Liu, Y.; Zeng, J.; Wei, Y.; Song, X. , mRNA Vaccines Against SARS-CoV-2 Variants Delivered by Lipid Nanoparticles Based on Novel Ionizable Lipids. Adv Funct Mater 2022, 32, 2204692. [Google Scholar] [CrossRef] [PubMed]
- Akinc, A.; Maier, M. A.; Manoharan, M.; Fitzgerald, K.; Jayaraman, M.; Barros, S.; Ansell, S.; Du, X.; Hope, M. J.; Madden, T. D.; Mui, B. L.; Semple, S. C.; Tam, Y. K.; Ciufolini, M.; Witzigmann, D.; Kulkarni, J. A.; van der Meel, R.; Cullis, P. R. , The Onpattro story and the clinical translation of nanomedicines containing nucleic acid-based drugs. Nature Nanotechnology 2019, 14, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Swingle, K. L.; Hamilton, A. G.; Mitchell, M. J. , Lipid Nanoparticle-Mediated Delivery of mRNA Therapeutics and Vaccines. Trends Mol Med 2021, 27, 616–617. [Google Scholar] [CrossRef]
- Baden, L. R.; El Sahly, H. M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S. A.; Rouphael, N.; Creech, C. B.; McGettigan, J.; Khetan, S.; Segall, N.; Solis, J.; Brosz, A.; Fierro, C.; Schwartz, H.; Neuzil, K.; Corey, L.; Gilbert, P.; Janes, H.; Follmann, D.; Marovich, M.; Mascola, J.; Polakowski, L.; Ledgerwood, J.; Graham, B. S.; Bennett, H.; Pajon, R.; Knightly, C.; Leav, B.; Deng, W.; Zhou, H.; Han, S.; Ivarsson, M.; Miller, J.; Zaks, T. , Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. The New England journal of medicine 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Polack, F. P.; Thomas, S. J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J. L.; Pérez Marc, G.; Moreira, E. D.; Zerbini, C.; Bailey, R.; Swanson, K. A.; Roychoudhury, S.; Koury, K.; Li, P.; Kalina, W. V.; Cooper, D.; Frenck, R. W., Jr.; Hammitt, L. L.; Türeci, Ö.; Nell, H.; Schaefer, A.; Ünal, S.; Tresnan, D. B.; Mather, S.; Dormitzer, P. R.; Şahin, U.; Jansen, K. U.; Gruber, W. C. , Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. The New England journal of medicine 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Sercombe, L.; Veerati, T.; Moheimani, F.; Wu, S. Y.; Sood, A. K.; Hua, S. , Advances and Challenges of Liposome Assisted Drug Delivery. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M. J.; Billingsley, M. M.; Haley, R. M.; Wechsler, M. E.; Peppas, N. A.; Langer, R. , Engineering precision nanoparticles for drug delivery. Nature Reviews Drug Discovery 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Eygeris, Y.; Gupta, M.; Kim, J.; Sahay, G. , Chemistry of Lipid Nanoparticles for RNA Delivery. Accounts of Chemical Research 2022, 55, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Hu, C. M.; Fang, R. H.; Zhang, L. , Liposome-like Nanostructures for Drug Delivery. Journal of materials chemistry. B 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J. A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D. J. A. , mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. International Journal of Pharmaceutics 2021, 601, 120586. [Google Scholar] [CrossRef] [PubMed]
- Swingle, K. L.; Hamilton, A. G.; Mitchell, M. J. , Lipid Nanoparticle-Mediated Delivery of mRNA Therapeutics and Vaccines. Trends in Molecular Medicine 2021, 27, 616–617. [Google Scholar] [CrossRef] [PubMed]
- Hashiba, K.; Sato, Y.; Taguchi, M.; Sakamoto, S.; Otsu, A.; Maeda, Y.; Shishido, T.; Murakawa, M.; Okazaki, A.; Harashima, H. , Branching Ionizable Lipids Can Enhance the Stability, Fusogenicity, and Functional Delivery of mRNA. Small Science 2023, 3, 2200071. [Google Scholar] [CrossRef]
- Tilstra, G.; Couture-Senécal, J.; Lau, Y. M. A.; Manning, A. M.; Wong, D. S. M.; Janaeska, W. W.; Wuraola, T. A.; Pang, J.; Khan, O. F. , Iterative Design of Ionizable Lipids for Intramuscular mRNA Delivery. Journal of the American Chemical Society 2023. [Google Scholar] [CrossRef]
- Hajj, K. A.; Whitehead, K. A. , Tools for translation: non-viral materials for therapeutic mRNA delivery. Nature Reviews Materials 2017, 2, 17056. [Google Scholar] [CrossRef]
- Kanasty, R.; Dorkin, J. R.; Vegas, A.; Anderson, D. , Delivery materials for siRNA therapeutics. Nature Materials 2013, 12, 967–977. [Google Scholar] [CrossRef]
- Harper, P. E.; Mannock, D. A.; Lewis, R. N. A. H.; McElhaney, R. N.; Gruner, S. M. , X-Ray Diffraction Structures of Some Phosphatidylethanolamine Lamellar and Inverted Hexagonal Phases*. Biophysical Journal 2001, 81, 2693–2706. [Google Scholar] [CrossRef]
- Cheng, X.; Lee, R. J. , The role of helper lipids in lipid nanoparticles (LNPs) designed for oligonucleotide delivery. Advanced Drug Delivery Reviews 2016, 99, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, D.; Marchini, C.; Cardarelli, F.; Amenitsch, H.; Garulli, C.; Bifone, A.; Caracciolo, G. , Transfection efficiency boost of cholesterol-containing lipoplexes. Biochimica et Biophysica Acta (BBA) - Biomembranes 2012, 1818, 2335–2343. [Google Scholar] [CrossRef]
- Patel, S.; Ashwanikumar, N.; Robinson, E.; Xia, Y.; Mihai, C.; Griffith, J. P.; Hou, S.; Esposito, A. A.; Ketova, T.; Welsher, K.; Joyal, J. L.; Almarsson, Ö.; Sahay, G. , Naturally-occurring cholesterol analogues in lipid nanoparticles induce polymorphic shape and enhance intracellular delivery of mRNA. Nature Communications 2020, 11, 983. [Google Scholar] [CrossRef] [PubMed]
- Evers, M. J. W.; Kulkarni, J. A.; Van Der Meel, R.; Cullis, P. R.; Vader, P.; Schiffelers, R. M. , State-of-the-Art Design and Rapid-Mixing Production Techniques of Lipid Nanoparticles for Nucleic Acid Delivery. Small Methods 2018, 2, 1700375. [Google Scholar] [CrossRef]
- Jokerst, J. V.; Lobovkina, T.; Zare, R. N.; Gambhir, S. S. , Nanoparticle PEGylation for imaging and therapy. Nanomedicine 2011, 6, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Knop, K.; Hoogenboom, R.; Fischer, D.; Schubert, U. S. , Poly(ethylene glycol) in Drug Delivery: Pros and Cons as Well as Potential Alternatives. Angewandte Chemie International Edition 2010, 49, 6288–6308. [Google Scholar] [CrossRef]
- Kim, J.; Eygeris, Y.; Gupta, M.; Sahay, G. , Self-assembled mRNA vaccines. Advanced Drug Delivery Reviews 2021, 170, 83–112. [Google Scholar] [CrossRef]
- Akinc, A.; Goldberg, M.; Qin, J.; Dorkin, J. R.; Gamba-Vitalo, C.; Maier, M.; Jayaprakash, K. N.; Jayaraman, M.; Rajeev, K. G.; Manoharan, M.; Koteliansky, V.; Röhl, I.; Leshchiner, E. S.; Langer, R.; Anderson, D. G. , Development of Lipidoid–siRNA Formulations for Systemic Delivery to the Liver. Molecular Therapy 2009, 17, 872–879. [Google Scholar]
- Zhu, X.; Tao, W.; Liu, D.; Wu, J.; Guo, Z.; Ji, X.; Bharwani, Z.; Zhao, L.; Zhao, X.; Farokhzad, O. C.; Shi, J. , Surface De-PEGylation Controls Nanoparticle-Mediated siRNA Delivery In Vitro and In Vivo. Theranostics 2017, 7, 1990–2002. [Google Scholar] [CrossRef] [PubMed]
- Fenton, O. S.; Kauffman, K. J.; Kaczmarek, J. C.; McClellan, R. L.; Jhunjhunwala, S.; Tibbitt, M. W.; Zeng, M. D.; Appel, E. A.; Dorkin, J. R.; Mir, F. F.; Yang, J. H.; Oberli, M. A.; Heartlein, M. W.; DeRosa, F.; Langer, R.; Anderson, D. G. , Synthesis and Biological Evaluation of Ionizable Lipid Materials for the In Vivo Delivery of Messenger RNA to B Lymphocytes. Advanced materials (Deerfield Beach, Fla.) 2017, 29. [Google Scholar] [CrossRef] [PubMed]
- Fenton, O. S.; Kauffman, K. J.; McClellan, R. L.; Kaczmarek, J. C.; Zeng, M. D.; Andresen, J. L.; Rhym, L. H.; Heartlein, M. W.; DeRosa, F.; Anderson, D. G. , Customizable Lipid Nanoparticle Materials for the Delivery of siRNAs and mRNAs. Angewandte Chemie (International ed. in English) 2018, 57, 13582–13586. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, W.; Nguyen, G. N.; Zhang, C.; Zeng, C.; Yan, J.; Du, S.; Hou, X.; Li, W.; Jiang, J.; Deng, B.; McComb, D. W.; Dorkin, R.; Shah, A.; Barrera, L.; Gregoire, F.; Singh, M.; Chen, D.; Sabatino, D. E.; Dong, Y. , Functionalized lipid-like nanoparticles for in vivo mRNA delivery and base editing. Science advances 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Lin, J.; Huang, Y.; Li, L.; Delcassian, D.; Ge, Y.; Shi, Y.; Anderson, D. G. , Synergistic lipid compositions for albumin receptor mediated delivery of mRNA to the liver. Nature Communications 2020, 11, 2424. [Google Scholar] [CrossRef]
- Fenton, O. S.; Kauffman, K. J.; McClellan, R. L.; Appel, E. A.; Dorkin, J. R.; Tibbitt, M. W.; Heartlein, M. W.; DeRosa, F.; Langer, R.; Anderson, D. G. , Bioinspired Alkenyl Amino Alcohol Ionizable Lipid Materials for Highly Potent In Vivo mRNA Delivery. Advanced materials (Deerfield Beach, Fla.) 2016, 28, 2939–2943. [Google Scholar] [CrossRef]
- Han, X.; Zhang, H.; Butowska, K.; Swingle, K. L.; Alameh, M.-G.; Weissman, D.; Mitchell, M. J. , An ionizable lipid toolbox for RNA delivery. Nature Communications 2021, 12, 7233. [Google Scholar] [CrossRef]
- Heyes, J.; Palmer, L.; Bremner, K.; MacLachlan, I. , Cationic lipid saturation influences intracellular delivery of encapsulated nucleic acids. Journal of Controlled Release 2005, 107, 276–287. [Google Scholar] [CrossRef]
- Kauffman, K. J.; Dorkin, J. R.; Yang, J. H.; Heartlein, M. W.; DeRosa, F.; Mir, F. F.; Fenton, O. S.; Anderson, D. G. , Optimization of Lipid Nanoparticle Formulations for mRNA Delivery in Vivo with Fractional Factorial and Definitive Screening Designs. Nano Letters 2015, 15, 7300–7306. [Google Scholar] [CrossRef]
- Zhu, X.; Xu, Y.; Solis, L. M.; Tao, W.; Wang, L.; Behrens, C.; Xu, X.; Zhao, L.; Liu, D.; Wu, J.; Zhang, N. ; Wistuba, II; Farokhzad, O. C.; Zetter, B. R.; Shi, J., Long-circulating siRNA nanoparticles for validating Prohibitin1-targeted non-small cell lung cancer treatment. Proceedings of the National Academy of Sciences of the United States of America 2015, 112, 7779–7784. [Google Scholar] [CrossRef]
- Maier, M. A.; Jayaraman, M.; Matsuda, S.; Liu, J.; Barros, S.; Querbes, W.; Tam, Y. K.; Ansell, S. M.; Kumar, V.; Qin, J.; Zhang, X.; Wang, Q.; Panesar, S.; Hutabarat, R.; Carioto, M.; Hettinger, J.; Kandasamy, P.; Butler, D.; Rajeev, K. G.; Pang, B.; Charisse, K.; Fitzgerald, K.; Mui, B. L.; Du, X.; Cullis, P.; Madden, T. D.; Hope, M. J.; Manoharan, M.; Akinc, A. , Biodegradable Lipids Enabling Rapidly Eliminated Lipid Nanoparticles for Systemic Delivery of RNAi Therapeutics. Molecular Therapy 2013, 21, 1570–1578. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, K. A.; Dorkin, J. R.; Vegas, A. J.; Chang, P. H.; Veiseh, O.; Matthews, J.; Fenton, O. S.; Zhang, Y.; Olejnik, K. T.; Yesilyurt, V.; Chen, D.; Barros, S.; Klebanov, B.; Novobrantseva, T.; Langer, R.; Anderson, D. G. , Degradable lipid nanoparticles with predictable in vivo siRNA delivery activity. Nature Communications 2014, 5, 4277. [Google Scholar] [CrossRef]
- Paunovska, K.; Loughrey, D.; Dahlman, J. E. , Drug delivery systems for RNA therapeutics. Nature Reviews Genetics 2022, 23, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Sabnis, S.; Kumarasinghe, E. S.; Salerno, T.; Mihai, C.; Ketova, T.; Senn, J. J.; Lynn, A.; Bulychev, A.; McFadyen, I.; Chan, J.; Almarsson, Ö.; Stanton, M. G.; Benenato, K. E. , A Novel Amino Lipid Series for mRNA Delivery: Improved Endosomal Escape and Sustained Pharmacology and Safety in Non-human Primates. Molecular Therapy 2018, 26, 1509–1519. [Google Scholar] [CrossRef]
- Knapp, C. M.; Guo, P.; Whitehead, K. A. , Lipidoid Tail Structure Strongly Influences siRNA Delivery Activity. Cellular and Molecular Bioengineering 2016, 9, 305–314. [Google Scholar] [CrossRef]
- Hajj, K. A.; Ball, R. L.; Deluty, S. B.; Singh, S. R.; Strelkova, D.; Knapp, C. M.; Whitehead, K. A. , Branched-Tail Lipid Nanoparticles Potently Deliver mRNA In Vivo due to Enhanced Ionization at Endosomal pH. Small 2019, 15, 1805097. [Google Scholar] [CrossRef] [PubMed]
- Sabnis, S.; Kumarasinghe, E. S.; Salerno, T.; Mihai, C.; Ketova, T.; Senn, J. J.; Lynn, A.; Bulychev, A.; McFadyen, I.; Chan, J.; Almarsson, Ö.; Stanton, M. G.; Benenato, K. E. , A Novel Amino Lipid Series for mRNA Delivery: Improved Endosomal Escape and Sustained Pharmacology and Safety in Non-human Primates. Molecular therapy : the journal of the American Society of Gene Therapy 2018, 26, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Fan, N.; Huang, H.; Jiang, X.; Qin, S.; Xiao, W.; Zheng, Q.; Zhang, Y.; Duan, X.; Qin, Z.; Liu, Y.; Zeng, J.; Wei, Y.; Song, X. , mRNA Vaccines Against SARS-CoV-2 Variants Delivered by Lipid Nanoparticles Based on Novel Ionizable Lipids. Advanced Functional Materials 2022, 32, 2204692. [Google Scholar] [CrossRef]
- Riley, R. S.; Kashyap, M. V.; Billingsley, M. M.; White, B.; Alameh, M.-G.; Bose, S. K.; Zoltick, P. W.; Li, H.; Zhang, R.; Cheng, A. Y.; Weissman, D.; Peranteau, W. H.; Mitchell, M. J. , Ionizable lipid nanoparticles for in utero mRNA delivery. Science Advances 7 eaba1028. [CrossRef]
- Samaridou, E.; Heyes, J.; Lutwyche, P. , Lipid nanoparticles for nucleic acid delivery: Current perspectives. Advanced Drug Delivery Reviews 2020, 154-155, 37–63. [Google Scholar] [CrossRef]
- Suzuki, Y.; Miyazaki, T.; Muto, H.; Kubara, K.; Mukai, Y.; Watari, R.; Sato, S.; Kondo, K.; Tsukumo, S. I.; Yasutomo, K.; Ito, M.; Tsukahara, K. , Design and lyophilization of lipid nanoparticles for mRNA vaccine and its robust immune response in mice and nonhuman primates. Molecular therapy. Nucleic acids 2022, 30, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Dhawan, V.; Holm, R.; Nagarsenker, M. S.; Perrie, Y. , Liposomes: Advancements and innovation in the manufacturing process. Advanced Drug Delivery Reviews 2020, 154-155, 102–122. [Google Scholar] [CrossRef] [PubMed]
- Matsuura-Sawada, Y.; Maeki, M.; Nishioka, T.; Niwa, A.; Yamauchi, J.; Mizoguchi, M.; Wada, K.; Tokeshi, M. , Microfluidic Device-Enabled Mass Production of Lipid-Based Nanoparticles for Applications in Nanomedicine and Cosmetics. ACS Applied Nano Materials 2022, 5, 7867–7876. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, H.; Cito, S.; Fan, J.; Mäkilä, E.; Salonen, J.; Hirvonen, J.; Sikanen, T. M.; Weitz, D. A.; Santos, H. A. , Core/Shell Nanocomposites Produced by Superfast Sequential Microfluidic Nanoprecipitation. Nano Letters 2017, 17, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Utharala, R.; Tseng, Q.; Furlong, E. E. M.; Merten, C. A. , A Versatile, Low-Cost, Multiway Microfluidic Sorter for Droplets, Cells, and Embryos. Analytical Chemistry 2018, 90, 5982–5988. [Google Scholar] [CrossRef] [PubMed]
- Maeki, M.; Uno, S.; Niwa, A.; Okada, Y.; Tokeshi, M. , Microfluidic technologies and devices for lipid nanoparticle-based RNA delivery. Journal of Controlled Release 2022, 344, 80–96. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, B. G.; Ceccato, B. T.; Michelon, M.; Han, S. W.; de la Torre, L. G. , Advanced Microfluidic Technologies for Lipid Nano-Microsystems from Synthesis to Biological Application. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef]
- Wang, X.; Liu, S.; Sun, Y.; Yu, X.; Lee, S. M.; Cheng, Q.; Wei, T.; Gong, J.; Robinson, J.; Zhang, D.; Lian, X.; Basak, P.; Siegwart, D. J. , Preparation of selective organ-targeting (SORT) lipid nanoparticles (LNPs) using multiple technical methods for tissue-specific mRNA delivery. Nature Protocols 2023, 18, 265–291. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Wei, T.; Farbiak, L.; Johnson, L. T.; Dilliard, S. A.; Siegwart, D. J. , Selective organ targeting (SORT) nanoparticles for tissue-specific mRNA delivery and CRISPR–Cas gene editing. Nature Nanotechnology 2020, 15, 313–320. [Google Scholar] [CrossRef]
- Gros, F.; Hiatt, H.; Gilbert, W.; Kurland, C. G.; Risebrough, R. W.; Watson, J. D. , Unstable Ribonucleic Acid Revealed by Pulse Labelling of Escherichia Coli. Nature 1961, 190, 581–585. [Google Scholar] [CrossRef]
- Oude Munnink, B. B.; Nieuwenhuijse, D. F.; Stein, M.; O’Toole, Á.; Haverkate, M.; Mollers, M.; Kamga, S. K.; Schapendonk, C.; Pronk, M.; Lexmond, P.; van der Linden, A.; Bestebroer, T.; Chestakova, I.; Overmars, R. J.; van Nieuwkoop, S.; Molenkamp, R.; van der Eijk, A. A.; GeurtsvanKessel, C.; Vennema, H.; Meijer, A.; Rambaut, A.; van Dissel, J.; Sikkema, R. S.; Timen, A.; Koopmans, M.; Oudehuis, G. J. A. P. M.; Schinkel, J.; Kluytmans, J.; Kluytmans-van den Bergh, M.; van den Bijllaardt, W.; Berntvelsen, R. G.; van Rijen, M. M. L.; Schneeberger, P.; Pas, S.; Diederen, B. M.; Bergmans, A. M. C.; van der Eijk, P. A. V.; Verweij, J. J.; Buiting, A. G. N.; Streefkerk, R.; Aldenkamp, A. P.; de Man, P.; Koelemal, J. G. M.; Ong, D.; Paltansing, S.; Veassen, N.; Sleven, J.; Bakker, L.; Brockhoff, H.; Rietveld, A.; Slijkerman Megelink, F.; Cohen Stuart, J.; de Vries, A.; van der Reijden, W.; Ros, A.; Lodder, E.; Verspui-van der Eijk, E.; Huijskens, I.; Kraan, E. M.; van der Linden, M. P. M.; Debast, S. B.; Naiemi, N. A.; Kroes, A. C. M.; Damen, M.; Dinant, S.; Lekkerkerk, S.; Pontesilli, O.; Smit, P.; van Tienen, C.; Godschalk, P. C. R.; van Pelt, J.; Ott, A.; van der Weijden, C.; Wertheim, H.; Rahamat-Langendoen, J.; Reimerink, J.; Bodewes, R.; Duizer, E.; van der Veer, B.; Reusken, C.; Lutgens, S.; Schneeberger, P.; Hermans, M.; Wever, P.; Leenders, A.; ter Waarbeek, H.; Hoebe, C.; The Dutch-Covid-19 response, t. , Rapid SARS-CoV-2 whole-genome sequencing and analysis for informed public health decision-making in the Netherlands. Nature Medicine 2020, 26, 1405–1410. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, R.; Hogan, M. J.; Loré, K.; Pardi, N. , Innate immune mechanisms of mRNA vaccines. Immunity 2022, 55, 1993–2005. [Google Scholar] [CrossRef]
- Arevalo, C. P.; Bolton, M. J.; Le Sage, V.; Ye, N.; Furey, C.; Muramatsu, H.; Alameh, M.-G.; Pardi, N.; Drapeau, E. M.; Parkhouse, K.; Garretson, T.; Morris, J. S.; Moncla, L. H.; Tam, Y. K.; Fan, S. H. Y.; Lakdawala, S. S.; Weissman, D.; Hensley, S. E. , A multivalent nucleoside-modified mRNA vaccine against all known influenza virus subtypes. Science 2022, 378, 899–904. [Google Scholar] [CrossRef]
- Bonam, S. R.; Partidos, C. D.; Halmuthur, S. K. M.; Muller, S. , An Overview of Novel Adjuvants Designed for Improving Vaccine Efficacy. Trends in Pharmacological Sciences 2017, 38, 771–793. [Google Scholar] [CrossRef] [PubMed]
- Vaca, G. B.; Meyer, M.; Cadete, A.; Hsiao, C. J.; Golding, A.; Jeon, A.; Jacquinet, E.; Azcue, E.; Guan, C. M.; Sanchez-Felix, X.; Pietzsch, C. A.; Mire, C. E.; Hyde, M. A.; Comeaux, M. E.; Williams, J. M.; Sung, J. C.; Carfi, A.; Edwards, D. K.; Bukreyev, A.; Bahl, K. , Intranasal mRNA-LNP vaccination protects hamsters from SARS-CoV-2 infection. bioRxiv 2023. 2023.01.11.523616. [Google Scholar]
- Jansen, E. M.; Frijlink, H. W.; Hinrichs, W. L. J.; Ruigrok, M. J. R. , Are inhaled mRNA vaccines safe and effective? A review of preclinical studies. Expert Opinion on Drug Delivery 2022, 19, 1471–1485. [Google Scholar] [CrossRef]
- Phua, K. K. L.; Leong, K. W.; Nair, S. K. , Transfection efficiency and transgene expression kinetics of mRNA delivered in naked and nanoparticle format. Journal of Controlled Release 2013, 166, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Hassett, K. J.; Benenato, K. E.; Jacquinet, E.; Lee, A.; Woods, A.; Yuzhakov, O.; Himansu, S.; Deterling, J.; Geilich, B. M.; Ketova, T.; Mihai, C.; Lynn, A.; McFadyen, I.; Moore, M. J.; Senn, J. J.; Stanton, M. G.; Almarsson, Ö.; Ciaramella, G.; Brito, L. A. , Optimization of Lipid Nanoparticles for Intramuscular Administration of mRNA Vaccines. Molecular Therapy - Nucleic Acids 2019, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ols, S.; Yang, L.; Thompson, E. A.; Pushparaj, P.; Tran, K.; Liang, F.; Lin, A.; Eriksson, B.; Karlsson Hedestam, G. B.; Wyatt, R. T.; Loré, K. , Route of Vaccine Administration Alters Antigen Trafficking but Not Innate or Adaptive Immunity. Cell Reports 2020, 30, 3964–3971.e7. [Google Scholar] [CrossRef]
- Davies, N.; Hovdal, D.; Edmunds, N.; Nordberg, P.; Dahlén, A.; Dabkowska, A.; Arteta, M. Y.; Radulescu, A.; Kjellman, T.; Höijer, A.; Seeliger, F.; Holmedal, E.; Andihn, E.; Bergenhem, N.; Sandinge, A.-S.; Johansson, C.; Hultin, L.; Johansson, M.; Lindqvist, J.; Björsson, L.; Jing, Y.; Bartesaghi, S.; Lindfors, L.; Andersson, S. , Functionalized lipid nanoparticles for subcutaneous administration of mRNA to achieve systemic exposures of a therapeutic protein. Molecular Therapy - Nucleic Acids 2021, 24, 369–384. [Google Scholar] [CrossRef]
- Alameh, M.-G.; Tombácz, I.; Bettini, E.; Lederer, K.; Ndeupen, S.; Sittplangkoon, C.; Wilmore, J. R.; Gaudette, B. T.; Soliman, O. Y.; Pine, M.; Hicks, P.; Manzoni, T. B.; Knox, J. J.; Johnson, J. L.; Laczkó, D.; Muramatsu, H.; Davis, B.; Meng, W.; Rosenfeld, A. M.; Strohmeier, S.; Lin, P. J. C.; Mui, B. L.; Tam, Y. K.; Karikó, K.; Jacquet, A.; Krammer, F.; Bates, P.; Cancro, M. P.; Weissman, D.; Luning Prak, E. T.; Allman, D.; Igyártó, B. Z.; Locci, M.; Pardi, N. , Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity 2021, 54, 2877–2892.e7. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, J.; Xu, Q. , Current Developments and Challenges of mRNA Vaccines. Annual Review of Biomedical Engineering 2022, 24, 85–109. [Google Scholar] [CrossRef] [PubMed]
- Barbier, A. J.; Jiang, A. Y.; Zhang, P.; Wooster, R.; Anderson, D. G. , The clinical progress of mRNA vaccines and immunotherapies. Nature Biotechnology 2022, 40, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Naveed, Z.; Li, J.; Wilton, J.; Spencer, M.; Naus, M.; Velásquez García, H. A.; Kwong, J. C.; Rose, C.; Otterstatter, M.; Janjua, N. Z. , Comparative Risk of Myocarditis/Pericarditis Following Second Doses of BNT162b2 and mRNA-1273 Coronavirus Vaccines. Journal of the American College of Cardiology 2022, 80, 1900–1908. [Google Scholar] [CrossRef] [PubMed]
- Wagenhäuser, I.; Reusch, J.; Gabel, A.; Krone, L. B.; Kurzai, O.; Petri, N.; Krone, M. , Bivalent BNT162b2mRNA original/Omicron BA.4-5 booster vaccination: adverse reactions and inability to work compared to the monovalent COVID-19 booster. medRxiv 2022. 2022.11.07.22281982. [Google Scholar]
- Bolhassani, A.; Javanzad, S.; Saleh, T.; Hashemi, M.; Aghasadeghi, M. R.; Sadat, S. M. , Polymeric nanoparticles: potent vectors for vaccine delivery targeting cancer and infectious diseases. Human vaccines & immunotherapeutics 2014, 10, 321–332. [Google Scholar]
- Rosenblum, H. G.; Gee, J.; Liu, R.; Marquez, P. L.; Zhang, B.; Strid, P.; Abara, W. E.; McNeil, M. M.; Myers, T. R.; Hause, A. M.; Su, J. R.; Markowitz, L. E.; Shimabukuro, T. T.; Shay, D. K. , Safety of mRNA vaccines administered during the initial 6 months of the US COVID-19 vaccination programme: an observational study of reports to the Vaccine Adverse Event Reporting System and v-safe. The Lancet Infectious Diseases 2022, 22, 802–812. [Google Scholar] [CrossRef]
- Ju, Y.; Lee, W. S.; Pilkington, E. H.; Kelly, H. G.; Li, S.; Selva, K. J.; Wragg, K. M.; Subbarao, K.; Nguyen, T. H. O.; Rowntree, L. C.; Allen, L. F.; Bond, K.; Williamson, D. A.; Truong, N. P.; Plebanski, M.; Kedzierska, K.; Mahanty, S.; Chung, A. W.; Caruso, F.; Wheatley, A. K.; Juno, J. A.; Kent, S. J. , Anti-PEG Antibodies Boosted in Humans by SARS-CoV-2 Lipid Nanoparticle mRNA Vaccine. ACS Nano 2022, 16, 11769–11780. [Google Scholar] [CrossRef]
- Szebeni, J.; Storm, G.; Ljubimova, J. Y.; Castells, M.; Phillips, E. J.; Turjeman, K.; Barenholz, Y.; Crommelin, D. J. A.; Dobrovolskaia, M. A. , Applying lessons learned from nanomedicines to understand rare hypersensitivity reactions to mRNA-based SARS-CoV-2 vaccines. Nature Nanotechnology 2022, 17, 337–346. [Google Scholar] [CrossRef]
Figure 1.
Composition of LNPs and importance of each component.
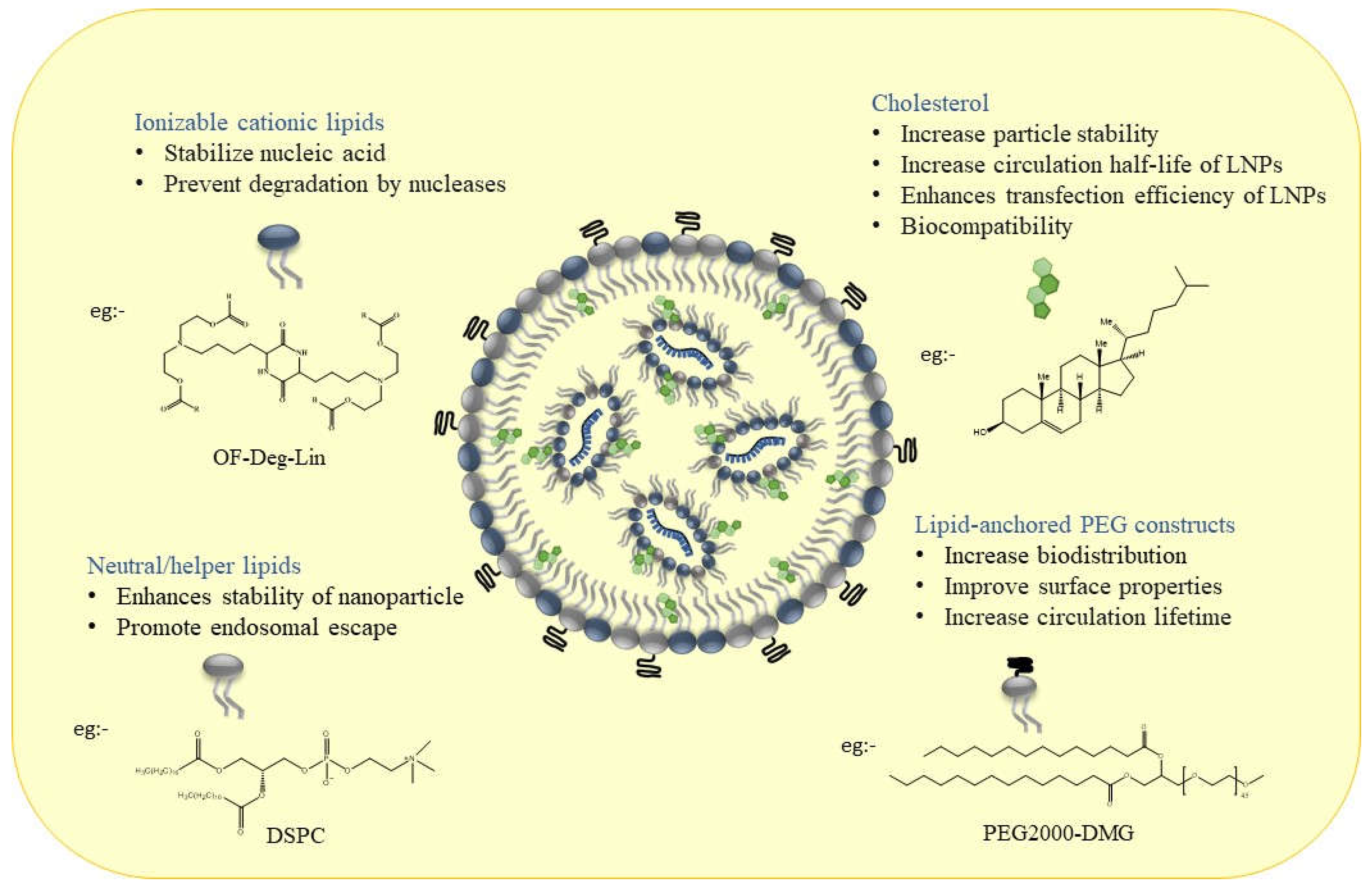
Figure 2.
Shows the different types of ionizable lipids used in the composition of LNPs.
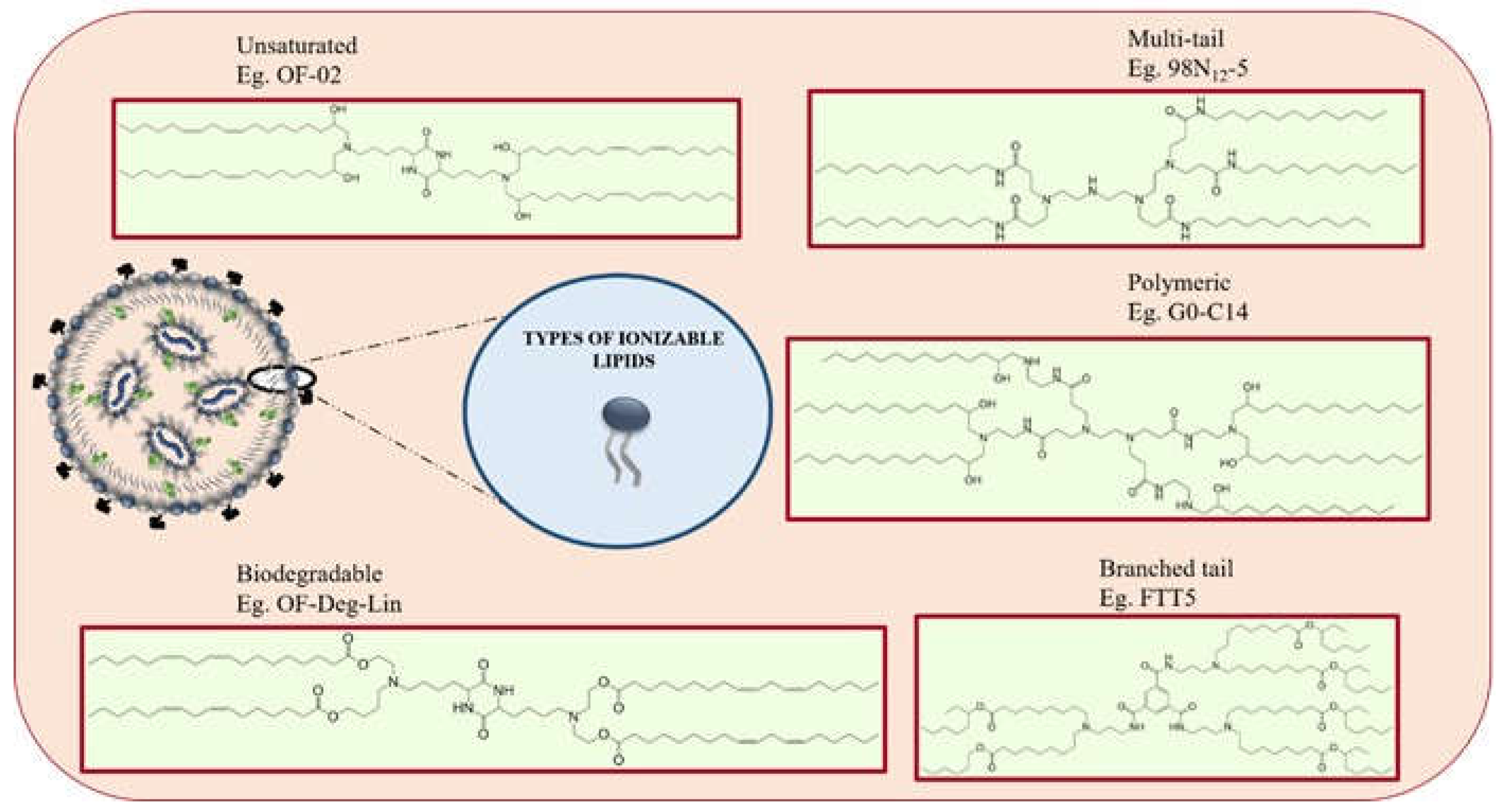
Figure 3.
a). The addition of a SORT molecule to typical four-component LNPs alters the in vivo delivery profile of the resultant five-component SORT LNPs, allowing for tissue-specific distribution of mRNA to the liver, lungs, and spleen of mice following IV injection. b). Chemical structures of the lipids used in this procedure. c-e). Three ways of mixing methods for SORT LNP formulation preparation: pipette mixing, vortex mixing, and microfluidic mixing. (Adapted from Ref with Permission.[59]).
Figure 3.
a). The addition of a SORT molecule to typical four-component LNPs alters the in vivo delivery profile of the resultant five-component SORT LNPs, allowing for tissue-specific distribution of mRNA to the liver, lungs, and spleen of mice following IV injection. b). Chemical structures of the lipids used in this procedure. c-e). Three ways of mixing methods for SORT LNP formulation preparation: pipette mixing, vortex mixing, and microfluidic mixing. (Adapted from Ref with Permission.[59]).
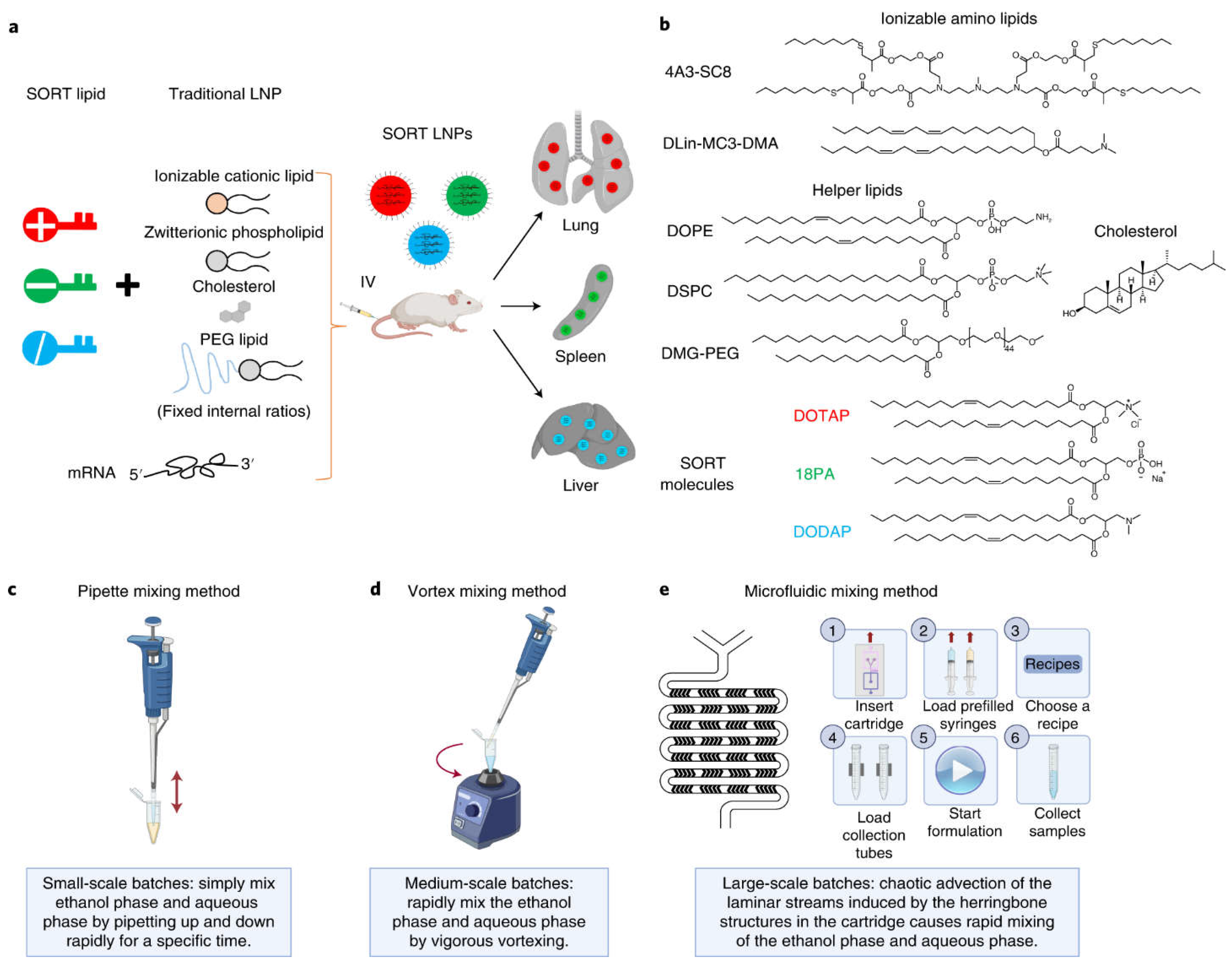
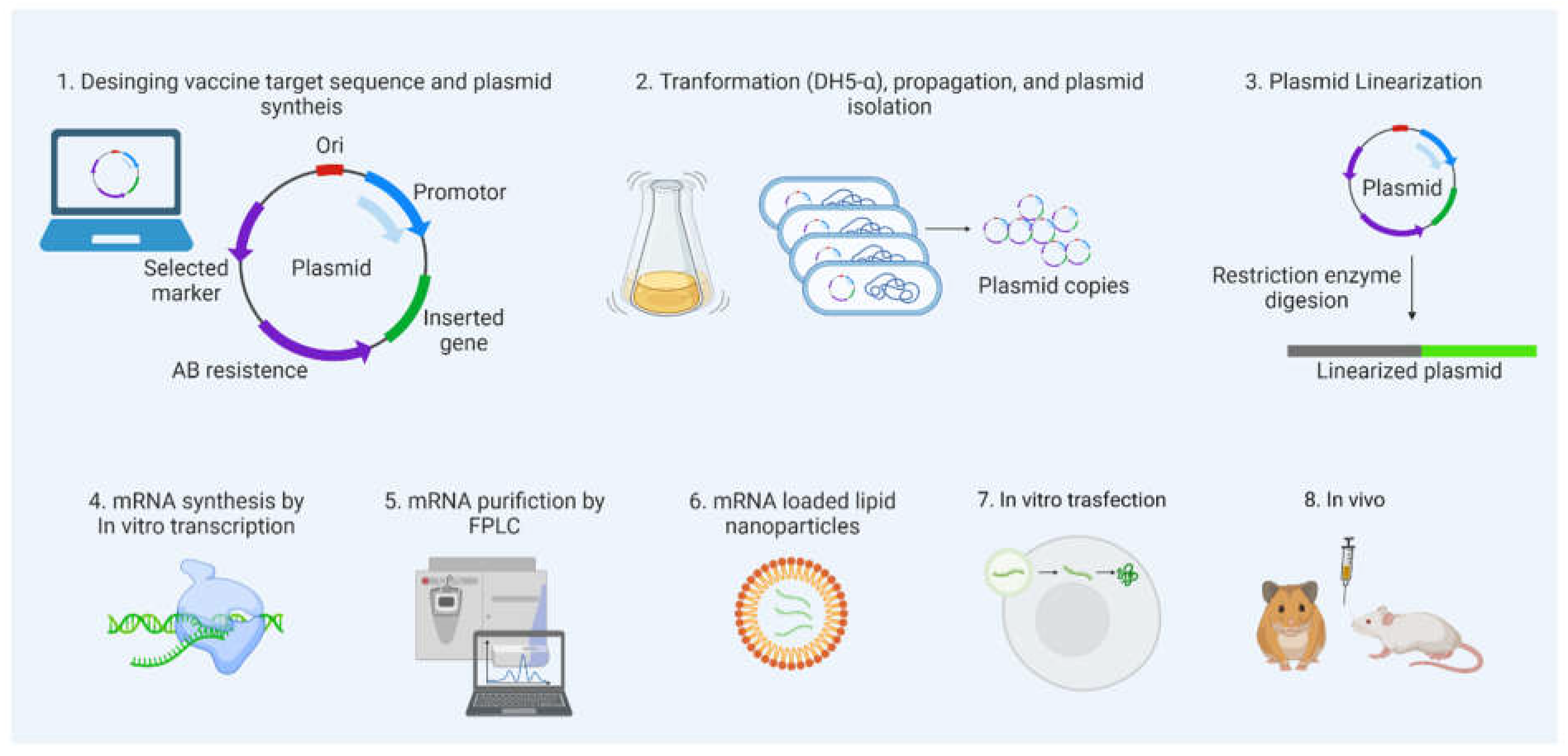
Figure 4.
(A). Structure and in vitro synthesis of mRNA. A. Basic structure of mRNA for therapeutics development. Cap, 5’ UTR, ORF, 3’UTR, PolyA are the five structural components of mRNA. (B). In Vitro Synthesis of Modified mRNA. Mostly, the vector DNA is used to construct the mRNA sequence. RNA polymerase uses the linearized DNA template containing either T7 or T3 or another promoter sequence for in vitro transcription of mRNA. In some instances, caping and poly A addition can be done after the IVT process. However, some advanced kits are engineered to do the capping and poly A during the IVT process, as a single step. More details are reviewed elsewhere.[1].
Figure 4.
(A). Structure and in vitro synthesis of mRNA. A. Basic structure of mRNA for therapeutics development. Cap, 5’ UTR, ORF, 3’UTR, PolyA are the five structural components of mRNA. (B). In Vitro Synthesis of Modified mRNA. Mostly, the vector DNA is used to construct the mRNA sequence. RNA polymerase uses the linearized DNA template containing either T7 or T3 or another promoter sequence for in vitro transcription of mRNA. In some instances, caping and poly A addition can be done after the IVT process. However, some advanced kits are engineered to do the capping and poly A during the IVT process, as a single step. More details are reviewed elsewhere.[1].
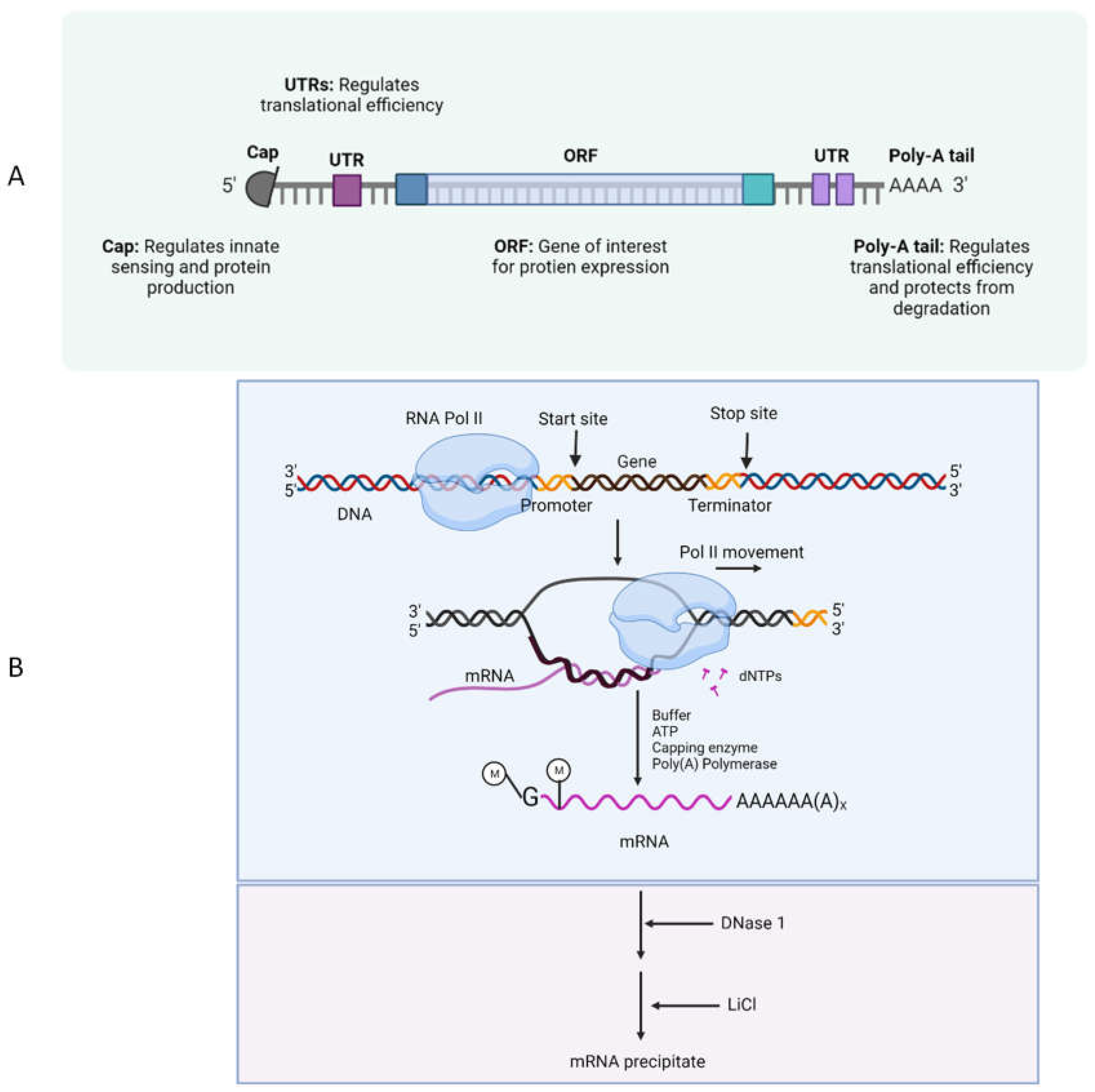
Figure 5.
Immune response by mRNA vaccines. LNPs are prepared by encapsulating mRNA, which encodes the viral protein of interest. Upon injection of vaccines, muscular cells taken up the LNPs following the release of mRNA into the cytosol and translation of target protein with the help of host machinery. In parallel, the danger associated signals produced by the LNPs recruit the innate immune cells, including neutrophils, monocytes, macrophages, dendritic cells, and others. The antigen-presenting cells process and present the antigen to the T cells, which further polarizes into effector T cells, and helps in B cell mediated responses. The cytotoxic T cells produced upon activation kills the infected cells and antibody (produced by B cells or plasma cells) neutralizes the virus.
Figure 5.
Immune response by mRNA vaccines. LNPs are prepared by encapsulating mRNA, which encodes the viral protein of interest. Upon injection of vaccines, muscular cells taken up the LNPs following the release of mRNA into the cytosol and translation of target protein with the help of host machinery. In parallel, the danger associated signals produced by the LNPs recruit the innate immune cells, including neutrophils, monocytes, macrophages, dendritic cells, and others. The antigen-presenting cells process and present the antigen to the T cells, which further polarizes into effector T cells, and helps in B cell mediated responses. The cytotoxic T cells produced upon activation kills the infected cells and antibody (produced by B cells or plasma cells) neutralizes the virus.
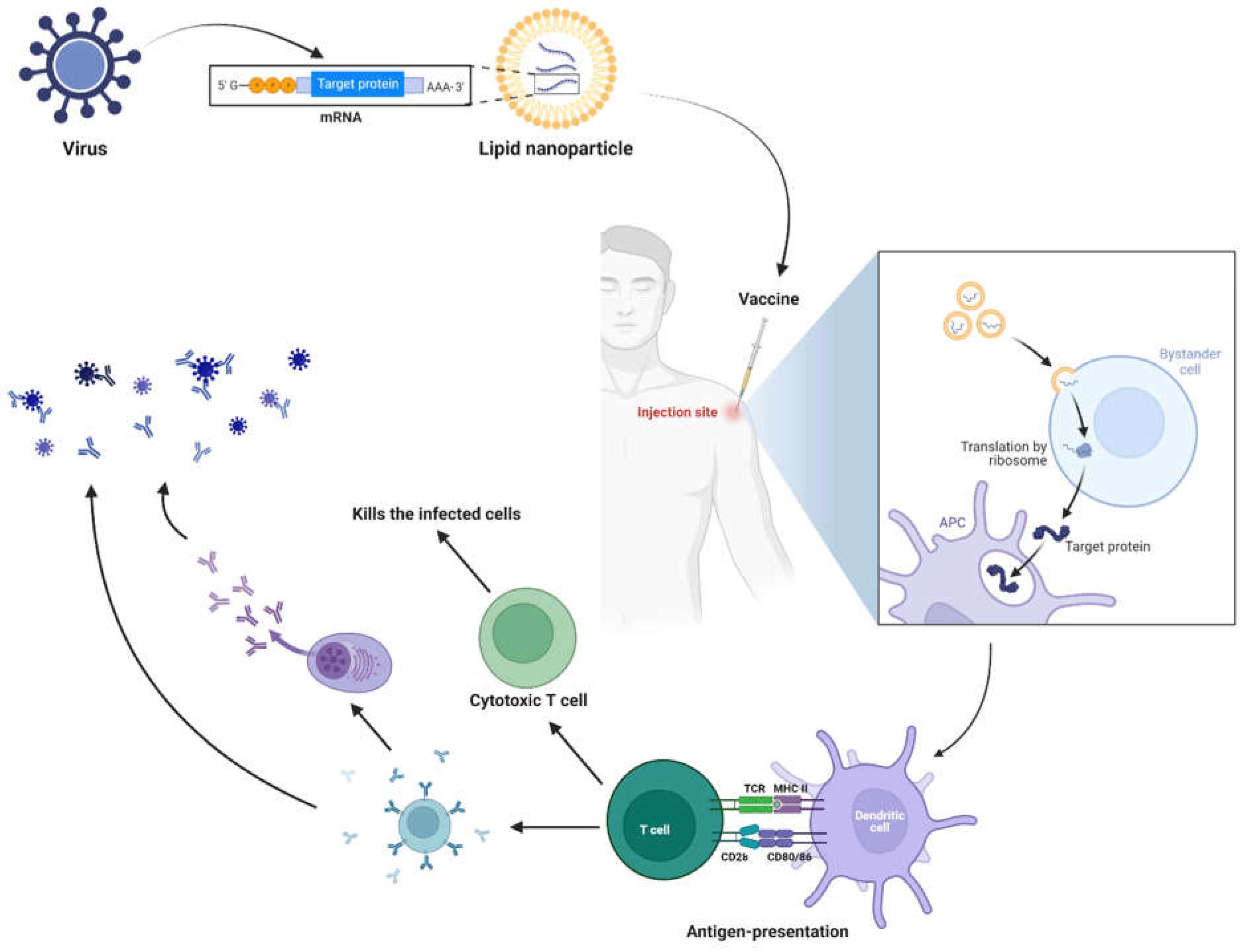

Table 1.
Shows the chemical structures of different class of lipids used in LNPs for mRNA delivery.
| Lipid | Function | Reference |
|---|---|---|
| Cationic ionizable lipids | ||
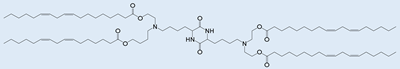 OF-Deg-Lin OF-Deg-Lin |
Selective delivery of mRNA into B lymphocytes | [32] |
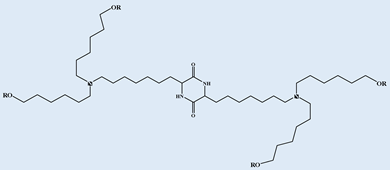 OF-C4-Deg-Lin OF-C4-Deg-Lin |
Selective delivery of siRNAs and mRNAs | [33] |
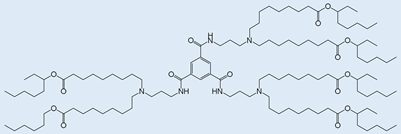 FTT5 FTT5 |
In vivo delivery of mRNA encoding human factor VIII and base editing components | [34] |
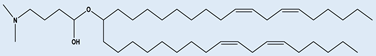 Dlin-MC3-DMA Dlin-MC3-DMA |
Used in albumin receptor mediated delivery of mRNA to the liver | [35] |
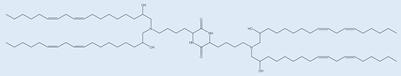 OF-02 OF-02 |
Enhanced hepatic mRNA delivery | [36] |
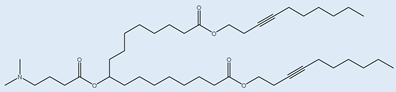 A6 A6 |
Albumin receptor mediated mRNA delivery | [35] |
| Neutral/helper lipids | ||
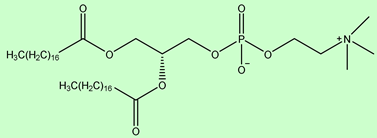 DSPC DSPC |
Used in mRNA vaccines and vaccine candidates, including COVID19. | [6] |
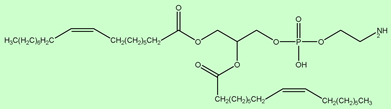 DOPE DOPE |
Delivery of variety of nucleic acids. | [26] |
| PEG lipids | ||
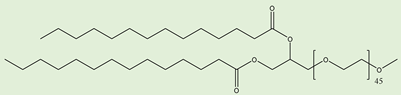 PEG2000-DMG PEG2000-DMG |
Used in mRNA vaccines and vaccine candidates, including COVID19. | [6] |
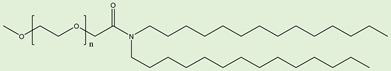 ALC-0159 ALC-0159 |
Delivery of mRNA vaccines. | [6] |
| Name | mRNA specific to | LNP Composition | Adverse effects |
|---|---|---|---|
| BNT162b2 | Spike glycoprotein of SARS-CoV-2 | NT162b2 LNPs composition: lipids ((4- hydroxybutyl)azanediyl) bis(hexane-6,1- diyl)bis(2- hexyldecanoate), 2 [(polyethylene glycol)- 2000]-N,Nditetradecylacetamide, 1,2-distearoyl-snglycero- 3-phosphocholine, and cholesterol), potassium chloride, monobasic potassium phosphate, sodium chloride, dibasic sodium phosphate dihydrate, and sucrose |
Myocarditis Pericarditis |
| mRNA- 1273 |
Spike glycoprotein of SARS-CoV-2 | LNP: Proprietary Ionic lipid SM-102, polyethylene glycol (PEG) 2000, dimyristoylglycerol (DMG), cholesterol, 1,2- distearoyl-sn-glycero-3- phosphocholine [DSPC]), tromethamine hydrochloride, acetic acid, sodium acetate, and sucrose |
Myocarditis Pericarditis |
| LUNARCOv- 19 (ARCT- 021) |
Self-replicating mRNA specific to Spike glycoprotein of SARS-CoV-2 |
Arcturus Therapeutics proprietary ionizable lipid, DSPC, cholesterol, and PEG2000-DMG dissolved in ethanol |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



