
Submitted:
06 February 2023
Posted:
08 February 2023
You are already at the latest version
Abstract
Mass cells (MCs) are tissue cells that are derived from bone marrow stem cells that contribute to allergic reactions, inflammatory diseases, innate and adaptive immunity, autoimmunity, and mental disorders. MCs located near the meninges communicate with microglia through the production of mediators such as histamine and tryptase, but also through the secretion of IL-1, IL-6 and TNF, which can create pathological effects in the brain. Preformed chemical mediators of inflammation and tumor necrosis factor (TNF) are rapidly released from the granules of MCs, the only immune cells capable of storing the cytokine TNF, although it can also be produced later through mRNA. The role of MCs in nervous system diseases has been extensively studied and reported in scientific literature and is of great clinical interest. However, many of the published articles concern studies on animals (mainly rats or mice) and not on humans. MCs are known to interact with neuropeptides that mediate endothelial cell activation, resulting in central nervous system (CNS) inflammatory disorders. In the brain, MCs interact with neurons causing neuronal excitation with the production of neuropeptides and the release of inflammatory mediators such as cytokines and chemokines. This article explores the current understanding of MC activation by neuropeptides and the role of pro-inflammatory cytokines, suggesting a therapeutic effect of the anti-inflammatory cytokines IL-37 and IL-38.
Keywords:
Neuropeptide
; Inflammation
; Immunity
; Mast cell: Pro-inflammatory cytokines
; Anti-inflammatory cytokines
1. Introduction
Mast cells (MCs) derive from bone marrow progenitors and after maturation, they migrate into the tissues where they carry out various biological responses, including innate and acquired immunity (1). In addition, MCs are immune cells involved in a number of disorders including inflammatory, autoimmune, and allergic diseases (2). The maturation of MCs has been reported to occur in the presence of stem cell factor (SCF), IL-3, IL-4, and IL-9, in vitro and in vivo (3). MCs are ubiquitous in the human body but are predominantly localized in perivascular tissue, and in the central nervous system (CNS) are found in corticotropin-releasing hormone (CRH)-positive nerve endings (4). The meninges also possess MCs that can be activated by insults such as stress and toxins, via vascular permeability, an effect that does not occur in MC-deficient rodents (5).
The generation, survival and development of MCs depends on the c-kit receptor which binds SCF by exerting the biological response (6). c-kit/CD117 is a proto-oncogene transmembrane tyrosine kinase receptor that has been immunolocalized in various cells including MCs. CD117 is a 145-Kd glycoprotein that is the product of the kit-gene, and SCF is the c-kit ligand, named MC growth factor, which promotes autophosphorylation of the c-kit receptor, which mediates signal transduction, critical for the survival of MCs (7). Moreover, CD117 can be instrumental in the diagnosis of some tumors, such as gastrointestinal ones where MCs are abundant (8). Experiments in mice have shown that c-kit is encoded at the mouse locus where it affects immature germ cells (9).
However, for ethical and practical reasons, there is not much research on human MCs in vivo and the most significant results on this issue have been obtained from research on rodents. In fact, the biological effects of MCs are studied in MC deficient mice, such as KitW-f/KitW-f, KitW/KitW-v or KitW-sh/KitW-sh, which possess a c-kit receptor dysfunction (10).
MCs express various surface receptors that allow them to respond to stimuli such as IL-3, SCF, neuropeptides, and others (11). Bacterial products such as lipopolysaccharide (LPS) (a potent macrophage stimulator) also activate MCs, demonstrating that these cells have anti-bacterial functions (12). The classical high specific activation of MCs occurs through the antigenic reaction with the IgE antibody bound on the cellular high-affinity IgE Fc receptor (FcRI) (13). This interaction occurs at high affinity (1 × 1010 M−1), provoking the aggregation of receptors and eliciting an intracellular biological response (14). FcεRI is composed of 4 subunits called alpha, beta and two gammas, responsible for initiating the biological cascade that leads to the generation of proteins that mediate the inflammatory and allergic responses (15). The beta subunit leads to the amplification of the antigen reaction with the IgE antibody, while the alpha subunit binds IgE and activates the MC (16). The biochemical cascade of reactions, leading to the transcription and activation of inflammatory cytokines, thus begins with the activation of the FcRI and the phosphorylation of tyrosine kinases (Src, Syk and Tec family) (17). Next, the 76 kDa SH2-containing leukocyte protein (SLP-76), the non-T cell activation linker (NTAL), and the phospholipase Cγ (PLCγ) (which regulates calcium in the cell) contribute to recruit protein kinase C (PKC) (18). The phospholipid PI-4,5-P2 membrane is subsequently hydrolyzed and soluble inositol 1,4,5-triphosphate (IP3) and therefore, diacylglycerol (DAG) is formed (19). Other signaling reactions include mitogen-activated protein kinase (MAPK), extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38 that lead to the transcription and production of inflammatory cytokines (20, 21) (Figure 1).
Furthermore, the activation of MC-phospholipases leads to the generation of arachidonic acid inflammatory products, such as prostaglandin D2 (PGD2), leukotrienes LTC4, LCD4, and LTE4. PGD2 is an unstable prostaglandin detected as 11β-PGF2α, a more stable compound, that is related to systemic inflammation at non-physiological concentration in peripheral blood (22, 23). Therefore, in the brain, PGD2 mediates inflammation and pain; while LTC4, LCD4, and LTE4 are slow reacting substances of anaphylaxis (SRS-A) involved in asthma and other inflammatory diseases (24, 25). For example, leukotriene E4 is produced by MCs and increases in mastocytosis, fueling inflammation. In fact, the inhibitor of this leukotriene is used in asthma and for shortness of breath (26).
After the MCs have been activated, they degranulate and release the inflammatory mediators stored in their granules in seconds (rapid release) (27). However, most of the production of the potent inflammatory cytokine tumor necrosis factor (TNF) results from the induction of the corresponding mRNA upon MC activation (late secretion) (28). Cytokines such as IL-5, IL-6, IL-31 and IL-33, and the chemokines CCL2, CCL5 and CXCL8, are synthesized by MCs through mRNA after activation (29) (Table 1).
Injury in the brain can stimulate pro-IL-1 which is cleaved by caspase-1 to form mature IL-1, which binds its receptor IL-1R in the cell membrane and activates gene expression pathways (Figure 2).
The secretion of IL-1 and TNF in the brain induces fever which is produced in the hypothalamus (30). The mechanism of the production of fever is not yet fully understood, however, the antigen which can be a neuropeptide and other pyrogens (IL-1, TNF, IL-6, etc.) activates MCs with the production of IL-1 (31, 32). This cytokine acts on the anterior hypothalamus to generate prostaglandin E2 (PGE2), which stimulates the vasomotor center with the heat and fever production (33). MCs secrete neuropeptides CRH and Substance P (SP), which activate microglia in the brain to generate IL-1 and the chemokines CXCL8 (34). In fact, MCs cross-talk with microglia by releasing histamine and tryptases which induce the secretion of pro-inflammatory cytokines in MCs (35).
IL-6 (also called "myokine") is involved in fever and is mostly secreted by T lymphocytes, macrophages, and MCs. It stimulates the immune response by binding to its receptor gp130, increasing intracellular calcium and causing a transduction cascade that leads to the activation of the signal transcription factors signal transducer and activator of transcription 3 (STAT3) and MAPK (36). IL-6 can be secreted by macrophages and MCs in response to specific microbial molecules called pathogen associated molecular patterns (PAMPs), which bind a group of important receptors of the innate immune system, the pattern recognition receptors (PRRs), to which the Toll-like receptors (TLRs) belong (37). Furthermore, IL-6 initiates PGE2 synthesis in the hypothalamus, thereby causing an increase in body temperature and mediating systemic inflammation (38). The relationship between MC-mediated allergic diseases and some neurological pathologies is demonstrated by the higher frequency of allergies and elevated IgE in children with CNS disorders (39).
2. Neuropeptides
2.1. Substance P (SP)
SP, discovered in 1931 by Von Euler, was characterized in 1970 by Leeman and Chang. It is a highly conserved neuropeptide, isolated from the rat brain, that is mainly secreted by neurons and is involved in nociception, hypotension, muscle contraction, and inflammation (40). The pro-inflammatory effect of SP, acting through its specific neurokinin-1 (NK-1), was confirmed in a large number of studies (41-43).
SP is a member of the tachykinin peptide hormone family, located on human chromosome 7 and encoded by the TAC1 gene (44, 45). SP activates MCs through specific receptors without degranulation and causes the infiltration of granulocytes through the synthesis of some cytokines such as TNF and IL-8 (46). In addition, the neuropeptides nerve growth factor (NGF) and NT can activate MCs and participate in inflammatory processes (47). In fact, activation of MCs, in some neuronal diseases, leads to the activation of NK1 receptors following an increase of vascular permeability (48) (Table 2).
SP stimulation can cause focal inflammation in the hypothalamus and amygdala, with pathological symptoms in several neurological diseases (49). Neuropeptides such as SP, CRH, and NT may have a synergistic inflammatory action through the activation of cytokines (50). Therefore, SP is an important neuropeptide and neuromodulatory compound involved in neurogenic inflammation (51). It is an activator of several immune cells, including macrophages and MCs, causing pro-inflammatory compounds including interleukins, chemokines and growth factors (31, 52). In the brain, SP is involved in pain through the stimulation of the trigeminal nerve. In in vitro studies, SP can activate MCs to secrete pro-inflammatory mediators such as cytokines, chemokines, arachidonic acid products and proteases. All these compounds participate to the local and systemic inflammatory response (53, 54).
In addition, SP induces vascular endothelial growth factor (VEGF) and IL-33 in a dose-dependent manner in MCs and acts synergistically with them in inflammatory responses (41, 55). The cytokine IL-33, also called "alarmin”, is generated and secreted by MCs and increases the ability of the SP to stimulate MCs to release VEGF, TNF, and IL-1 (56). This effect of SP on VEGF and TNF production demonstrates a crosstalk between neuropeptides and pro-inflammatory cytokines (55, 57). Moreover, human MCs stimulated with SP and anti-IgE produce IL-33 which increases the release of IL-31, a cytokine involved in atopic dermatitis (AD) and transmitting itch diseases involving the CNS (58). This demonstrates that SP is a potent pro-inflammatory neuropeptide and can activate and contribute to the secretion of certain pro-inflammatory cytokines (59). In fact, serum elevations of SP and its analogue hemokine-1, along with serum levels of the cytokines TNF and IL-6, have been found in inflammatory diseases including fibromyalgia (60). Additionally, in mouse models it has been observed that SP stimulates angiogenesis through the proliferation of endothelial cells, causing neurogenesis and peripheral inflammation (61).
SP performs its biological action by binding to tachykinin NK1 receptors located on the membrane of vascular endothelial cells, inducing inflammation, vascular permeability and edema (62). SP mediates pain through the trigeminal nerve and activates MCs to produce inflammatory mediators including leukotrienes and prostaglandins (63).
In some neurological and psychiatric disorders such as depression, bipolar mood disorder and anxiety, where MCs are in contact with SP positive nerves, an increase of SP, TNF and VEGF are observed (64). However, some neuropeptides such as epinephrine, a neurotransmitter beta-receptor agonist, are capable of inhibiting TNF, histamine, and PGD2 released by activated MCs, an effect that impedes some diseases including asthma (65).
2.2. Corticotropin-Releasing Hormone (CRH)
As early as 1948, Harris reported that the hypothalamus is an important connecting organ between the nervous and endocrine systems (66). Information is generated directly from the nuclei of the hypothalamus to the Locus Coureuleus, the main center for the release of norepinephrine, and from here an efferent pathway starts that goes directly to the adrenal medulla causing the release of catecholamines, including adrenaline, norepinephrine, and dopamine (67, 68). Catecholamines can interact with pro-inflammatory cytokines secreted after cognitive (stress) or non-cognitive (microorganisms) stimuli (69). Circulating cytokines reach the CNS and bind to their receptors, exerting an inflammatory effect (70). CRH acts through two receptors, corticotropin-releasing hormone receptor (CRHR)-1 (71) and CRHR-2 (72) which is subdivided into CRHR-2α and CRHR-2β (73).
In 1981, Vale et al. isolated and characterized CRH from the sheep hypothalamus. It is a very similar peptide to the human one and is widely expressed in the brain (74). The hypothalamus secretes CRH, allowing the pituitary gland to release adreno-corticotrophic hormone (ACTH) which, at the level of the cortex of the adrenal glands, will allow the release of cortisol, an anti-inflammatory marker (75, 76). Cortisol and catecholamines produced by the adrenal gland act at an anti-inflammatory level by inhibiting pro-inflammatory cytokines such as IL-1, TNF and IL-6 (77). The parvocellular neurons of the paraventricular nucleus are the major source of CRH in rats and humans (78). This hormone is secreted and then transported to the anterior part of the pituitary gland (79).
In humans, CRH can also be found outside the CNS, such as in the adrenal medulla, stomach, placenta, pancreas, duodenum, and in some tumors (80). The administration of CRH in vivo causes an elevation of ACTH and cortisol in plasma, with side effects depending on the administered dose (81). Thus, CRH is secreted by the hypothalamus after antigen stimulation and activates the hypothalamic-pituitary-adrenal (HPA) axis, but it can also be released from nerves outside the CNS and produced by immune cells including MCs (82).
Immune cells such as MCs and other cells are interconnected to the CNS through the production of cytokines influencing the physiological behavior of the body (83). CRH secretion increases during stress and may lead to hypercortisolism. Moreover, CRH can promote MC maturation and induces neurogenic inflammation (84). In fact, the histamine produced by MCs increases CRH both at the protein level and by inducing mRNA (29, 85). MCs can produce and be activated by neuropeptides which include SP, CRH, NGF and neurotensin (NT), an effect that can be altered by cytokines (86) (Table 3).
Some CNS diseases can lead to an increase in the expression of CRH-1 receptor in MCs (87), with an elevation of CRH levels and the generation of neurological disorders. In some neurological disorders, there may be an increase in MCs expressing the CRH-1 receptor with induction of inflammation (88).
The secretion of IL-1 and IL-6 by MCs also causes stimulation of the CRH that can further stimulate the release of VEGF (90) and PGE2α (91).
2.3. Neurotensin (NT)
In 1973, Susan Leeman (today a leading researcher in our group) isolated NT for the first time. NT is a 13 amino acid peptide which plays the role of neuromodulator and neurotransmitter in the CNS (92). There are 4 cell receptors that bind NT: NTSR 1, NTSR2 and the type 1 receptors Sortilin 1 (Sort 1) and SorLA (93). The NTSR1, with high neuron affinity, and the NTSR2, with low activity, were discovered first and therefore they are the most studied (94). NTSR1 is expressed more in neurons, while NTSR2 is poorly expressed, but this data still needs to be confirmed (95). NT regulates the digestive tract and cardiovascular system and is a mediator of neurological responses such as pain, psychosis, temperature regulation, sensitivity to ethanol, analgesia, etc. (96). Some studies show that NT can act as an antipsychotic-induced dopamine against schizophrenia without modifying NTS1 receptors (97). It has also been observed that low levels of NT in the thalamus lead to a stimulation of greater alcohol consumption (98).
NT is mostly found in the brain and gut where it becomes involved with inflammatory processes (99). It induces hormone secretion from the anterior pituitary gland and when it is administered in mice, causes side effects such as antinociception and hypothermia (100). NT secreted under stress acts synergistically with CRH to stimulate MCs, resulting in increased vascular permeability, generation of VEGF, and disruption of the blood-brain barrier (BBB) (101). This tridecapeptide which impacts the brain and other organs is fundamental in some inflammatory processes (102).
In addition, NT induces the expression of CRHR-1 and CRH protein which stimulates MCs in allergic diseases (103). In an interesting article, we reported that NT stimulates the gene expression and release of IL-1β and CXCL8 from cultured human microglia, underlining that the NT is a proinflammatory peptide (104) and therefore, confirming that NT plays an important role in inflammation. Immune cells such as lymphocyte macrophages and MCs are activated by NT with secretion of inflammatory cytokines and B cell immunoglobulin production, an effect that emphasizes the crosstalk of NT with the immune system (105). At the cellular level, NT increases calcium levels and the production of nitric acid, a pentavalent nitrogen oxyacid, with strong oxidizing power. NT is therefore a brain neurotransmitter that interacts with MCs, both in innate and acquired immunity, and has a synergistic action with CRH on the stimulation of MCs, activating a neuro-immune mechanism (106). These effects, which play a stress-mediating role, were observed in vivo in rodents (107, 108). In fact, NT and CRH released in the brain mediate CNS-related inflammatory disorders, such as stress, psoriasis, and AD, where MCs play a crucial role (109).
MC activation by neurotransmitters can lead to the release of histamine and increases vascular permeability with the generation of headache (110). There are several neuropeptides that can activate cerebral MCs that can produce pathological phenomena. For example, SP is capable of inducing itch due to the interaction of SP with neurokinin-1 receptor (NK1R) on neurons. NT binding its receptor NTR1 of the MC, stimulates the production of cytokines and chemokines through the activation of RAS, a signaling protein associated with the cell membrane, with phosphorylation of the Raf gene family which activates MEK 1/2 and ERK ½, which promote the transcription factor AP-1 in the nucleus and consequently, the production of cytokines/chemokines (111, 112) (Figure 3). Stress-induced NT can mediate neurogenic inflammation in SP-enhanced mice, causing activation and degranulation of MCs via the NK-1 receptor which induces, in turn, the upregulation of SCF or IL-4, important for proliferation of the MCs (113).
In conclusion, NT in the brain causes the activation and proliferation of microglia with brain inflammation, disruption of the blood-brain and the intestinal barrier, along with MC activation and the generation of pro-inflammatory cytokines with neuronal damage (114).
2.4. Anti-Inflammatory IL-37
In the late 1970s, IL-1α and IL-1β were shown to be pro-inflammatory cytokines that we now know can drive inflammation of Th1 and Th17 cells (115). Inflammatory neuropeptides stimulate IL-1 and other cytokines in the brain. For example, NT stimulates gene expression and secretion of IL-1β and the chemokine CXCL8 in cultured microglia (116). The IL-1 family includes IL-1α, IL-1β, and other inflammatory cytokines such as IL-18, IL-33, IL-36a, IL-36b, and IL-36 (117). Later, other cytokines were discovered, but with anti-inflammatory activity such as IL-1Ra, IL-37, and later, IL-38 and the anti-receptor IL-36Ra. IL-37 is a member of the IL-1 family and is a human anti-inflammatory cytokine, which suppresses innate immunity, modulates acquired immunity, and functions in mice (118). Five isoform splice variants (“a to e”) of IL-37 have been reported, of which “b” is the most studied. IL-37 is expressed in the nucleus as IL-1 and IL-33, but in contrast to these last two cytokines, IL-37 has anti-inflammatory power, dumping the innate immunity (119). Activation of the TLR in macrophages leads to the production and secretion of the IL-37 precursor (pro-IL37), which is cleaved by caspase-1 to form mature IL-37. Some IL-37 is transferred into the nucleus, while another part, together with pro-IL-37 is transferred out of the cell, and all are biologically active (120). In addition, it appears that extracellular proteases may influence pro-IL-37 to become more active. Once generated, IL-37 binds to the IL-18 receptor alpha chain (IL-18R), exercising the down regulation of inflammation (121). Human monocytes treated with various stimuli such as IL-1 or bacterial LPS are known to produce abundant pro-inflammatory cytokines, an effect enhanced in genetically IL-37 deficient monocytes (122). This data suggests that IL-37 controls the generation of pro-inflammatory cytokines of the IL-1 family. Indeed, in experimental models such as rheumatoid arthritis, where mice are treated with human IL-37, inflammation is inhibited (123). The IL-37 gene has only been identified in human cells, but not in mouse cells. However, accurate and sophisticated studies have allowed the generation of transgenic mice expressing human IL-37 (IL-37tg), allowing to better understand the functions and role of this cytokine in the inflammatory process (124). In these studies, IL-37 confirmed its inflammation-suppressing power even in clinical experimental models (125). In fact, in mice with inflammatory diseases, such as asthma or ulcerative colitis, treatment with IL-37 improved their pathological condition (126). IL-37 not only suppresses inflammation and innate immunity, but also plays a regulatory role in acquired immunity by acting on the inhibition of antigen-stimulated T lymphocytes (127). Thus, IL-37 is produced to protect the body's tissues against pro-inflammatory stimuli (128). A number of human cells, including immune cells (such as monocytes, macrophages, B cells, dendritic cells, etc.) express IL-37. Upon stimulation, immune cells treated with IL-37 in vitro produce less IL-1β, IL-6, and TNF (129). In addition, the migration of inflammatory cells is also inhibited by IL-37, an effect that confirms its anti-inflammatory power. However, the inhibitory mechanisms of IL-37 still need to be clarified, although some authors have hypothesized that IL-37 inhibits mammalian target of rapamycin (mTOR) (130).
IL-37 is generated as a protective, anti-inflammatory effect in the CNS in patients with ischemic stroke and other inflammatory diseases (131). Therefore, the inhibition of IL-1 family proinflammatory cytokines requires the IL-1 family decoy receptor (IL-1R8) (120).
In our recent studies we reported that IL-37 plays a fundamental role in autism spectrum disorder where it enhances and inhibits NT secretion and gene expression of IL-1 and CXCL8 in microglia (104). However, the mechanism of action of IL-37 is still unclear even though it is thought that the anti-inflammatory molecule SMAD family member 3 (SMAD3) of the cytokine TGF is involved in the nucleus (127, 132). In light of these studies, we can certainly say that IL-37 opens up therapeutic hope for acute and chronic inflammatory diseases, including autoimmune disorders (133).
2.5. IL-38 Dumping IL-1 Induced Inflammation
IL-1 is the endogenous pyrogen mediator of fever, with activating effects on CNS neurons and therefore is crucial in neurophysiological functions (134). Increased IL-1 in the brain may result in a number of biological effects including the elevation of inflammatory mediators and the inhibition of gamma-aminobutyric acid (GABA) receptor responses and calcium fluxes (135).
The IL-1 family member genes include pro-inflammatory and anti-inflammatory molecules (Figure 4). The last two cytokines discovered, IL-37 and IL-38, are anti-inflammatory and are located on chromosome 2. IL-38 is thought to be an ancestral gene intended to counteract the pro-inflammatory effects of IL-1 (136). In humans, IL-38 is found in several organs such as the tonsils, skin, fetal liver, heart, and placenta, but in future studies it will certainly be found in many other organs (137). The anti-inflammatory effect of IL-38 is homologous with two anti-receptors, IL-36Ra (about 40%) and IL-1Ra (about 40%), sharing the anti-inflammatory effect (138). to be active, IL-38 must be processed, but at the moment, the protease/s responsible for the generation of its mature form are unknown. IL-38 has similar anti-inflammatory potency to IL-37 and IL-36Ra, and works best at low concentrations, while at higher concentrations it may have opposite effects (139). In vitro studies demonstrated that IL-38 is capable of inhibiting TNF and IL-1 in LPS stimulated THP-1 cells. Moreover, it has been shown that IL-38 used in the truncated form is capable of inhibiting TNF and IL-1 in LPS stimulated THP-1 cells, whereas when IL-38 used at full length can have a stimulatory effect, especially on IL-6. In certain fungal infections such as Candida albicans, IL-38 suppresses TH17 lymphocytes and inhibits IL-17A, a pro-inflammatory cytokine that is produced in certain immune reactions (140). The same effect occurred after treatment with IL-36Ra and IL-1Ra (141). IL-17 comprises six subgroups ranging from IL-17A to IL-17F and among these the most studied cytokine is IL-17A (142). Immune cells and glial cells of the CNS express IL-17A receptors which, once activated, can mediate inflammatory brain disorders (143). IL-17 generated by γδ T lymphocytes, is a cytokine with pro-inflammatory potential that can act synergistically on astrocytes with other cytokines such as TNF, causing the secretion of CXCL1 which recruits neutrophils to the inflammatory site (144). In addition, in some cases, IL-17 expression may be due to bacterial infections or parasitic infestations which can activate TLR2. In these pathological conditions, treatment with IL-38 could have a therapeutic role (145).
Therefore, it is deduced that the activation of IL-17 in the CNS is very important in inflammatory pathologies (146). However, in many inflammatory brain disorders where IL-17 is involved, the mechanism of action or whether IL-17 directly causes inflammation is not clear yet, and therefore future studies are needed to clarify this dilemma (147).
In light of the results that have been published so far, IL-38 has been shown to play a regulatory role in rheumatic diseases such as rheumatoid arthritis and psoriasis, which are IL-1-mediated diseases (148).
Since, neuropeptides such as CRH, SP, and others, can activate microglia leading to secretion of the proinflammatory cytokines IL-1β, IL-6 and TNF (60), it is pertinent therefore, to think that inhibiting IL-1 by IL-38 can have a therapeutic effect in inflammatory disorders mediated by IL-1 pro-inflammatory members (138).
Author Contributions
Experiments D.L.; discussion, suggestions and critical contributions for the issues treated in the manuscript F.M., C.D., G.R., A.C., C.E.G., I.F., S.K.K., M.T., F.C.; writing, review and editing of the manuscript, P.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Conflicts of Interest
The authors declare no conflict of interest.
References
- Collington, S.J.; Westwick, J.; Williams, T.J.; Weller, C.L. The Function of CCR3 on Mouse Bone Marrow-Derived Mast Cells in Vitro. Immunology 2009, 129, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Nagata, K.; Nishiyama, C. IL-10 in Mast Cell-Mediated Immune Responses: Anti-Inflammatory and Proinflammatory Roles. International Journal of Molecular Sciences 2021, 22, 4972. [Google Scholar] [CrossRef] [PubMed]
- Eklund, K.K.; Ghildyal, N.; Austen, K.F.; Stevens, R.L. Induction by IL-9 and Suppression by IL-3 and IL-4 of the Levels of Chromosome 14-Derived Transcripts That Encode Late-Expressed Mouse Mast Cell Proteases. Journal of Immunology (Baltimore, Md.: 1950) 1993, 151, 4266–4273. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C. The Impact of Psychological Stress on Mast Cells. Annals of Allergy, Asthma & Immunology: Official Publication of the American College of Allergy, Asthma, & Immunology 2020, 125, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Polyzoidis, S.; Koletsa, T.; Panagiotidou, S.; Ashkan, K.; Theoharides, T.C. Mast Cells in Meningiomas and Brain Inflammation. Journal of Neuroinflammation 2015, 12. [Google Scholar] [CrossRef] [PubMed]
- Ammendola, M.; Sacco, R.; Sammarco, G.; Luposella, M.; Patruno, R.; Gadaleta, C.D.; De Sarro, G.; Ranieri, G. Mast Cell-Targeted Strategies in Cancer Therapy. Transfusion Medicine and Hemotherapy 2016, 43, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Caraffa, A.; Kritas, S.K.; Ronconi, G.; Lessiani, G.; Toniato, E.; Theoharides, T.C. Mast Cell, Pro-Inflammatory and Anti-Inflammatory: Jekyll and Hyde, the Story Continues. Journal of Biological Regulators and Homeostatic Agents 2017, 31, 263–267. [Google Scholar] [PubMed]
- de Silva, M.C.; Reid, R. Gastrointestinal Stromal Tumors (GIST): C-Kit Mutations, CD117 Expression, Differential Diagnosis and Targeted Cancer Therapy with Imatinib. Pathology Oncology Research 2003, 9, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.; Valent, P.; Galli, S.J. KIT as a Master Regulator of the Mast Cell Lineage. Journal of Allergy and Clinical Immunology 2022, 149, 1845–1854. [Google Scholar] [CrossRef]
- Wang, X.; Liu, W.; O’Donnell, M.; Lutgendorf, S.; Bradley, C.; Schrepf, A.; Liu, L.; Kreder, K.; Luo, Y. Evidence for the Role of Mast Cells in Cystitis-Associated Lower Urinary Tract Dysfunction: A Multidisciplinary Approach to the Study of Chronic Pelvic Pain Research Network Animal Model Study. PLoS ONE 2016, 11, e0168772. [Google Scholar] [CrossRef]
- Mencarelli, A.; Gunawan, M.; Yong, K.S.M.; Bist, P.; Tan, W.W.S.; Tan, S.Y.; Liu, M.; Huang, E.K.; Fan, Y.; Chan, J.K.Y.; et al. A Humanized Mouse Model to Study Mast Cells Mediated Cutaneous Adverse Drug Reactions. Journal of Leukocyte Biology 2020, 107, 797–807. [Google Scholar] [CrossRef] [PubMed]
- De Zuani, M.; Dal Secco, C.; Tonon, S.; Arzese, A.; Pucillo, C.E.M.; Frossi, B. LPS Guides Distinct Patterns of Training and Tolerance in Mast Cells. Frontiers in Immunology 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Moñino-Romero, S.; Erkert, L.; Schmidthaler, K.; Diesner, S.C.; Sallis, B.F.; Pennington, L.; Jardetzky, T.; Oettgen, H.C.; Bohle, B.; Fiebiger, E.; et al. The Soluble Isoform of Human FcɛRI Is an Endogenous Inhibitor of IgE-Mediated Mast Cell Responses. Allergy 2018, 74, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Sabban, S.; Ye, H.; Helm, B. Development of an in Vitro Model System for Studying the Interaction of Equus Caballus IgE with Its High-Affinity Receptor FcɛRI. Veterinary Immunology and Immunopathology 2013, 153, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Asai, K.; Fujimoto, K.; Harazaki, M.; Kusunoki, T.; Korematsu, S.; Ide, C.; Ra, C.; Hosoi, S. Distinct Aggregation of β- and γ-Chains of the High-Affinity IgE Receptor on Cross-Linking. Journal of Histochemistry & Cytochemistry 2000, 48, 1705–1715. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Mortaz, E.; Amani, S.; Tiotiu, A.; Folkerts, G.; Adcock, I.M. The Role of Mast Cells in IgE-Independent Lung Diseases. Clinical Reviews in Allergy & Immunology 2020, 58, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Yip, K.H.; Kolesnikoff, N.; Hauschild, N.; Biggs, L.; Lopez, A.F.; Galli, S.J.; Kumar, S.; Grimbaldeston, M.A. The Nedd4-2/Ndfip1 Axis Is a Negative Regulator of IgE-Mediated Mast Cell Activation. Nature Communications 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Mitroulis, I.; Alexaki, V.I.; Kourtzelis, I.; Ziogas, A.; Hajishengallis, G.; Chavakis, T. Leukocyte Integrins: Role in Leukocyte Recruitment and as Therapeutic Targets in Inflammatory Disease. Pharmacology & therapeutics 2015, 147, 123–135. [Google Scholar] [CrossRef]
- Zhou, Y.; Wing, Michele R.; Sondek, J.; Harden, T. Kendall Molecular Cloning and Characterization of PLC‐η2. Biochemical Journal, 2005; 391, 667–676. [CrossRef]
- Johnson, G.L. Mitogen-Activated Protein Kinase Pathways Mediated by ERK, JNK, and P38 Protein Kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef]
- Roux, P.P.; Blenis, J. ERK and P38 MAPK-Activated Protein Kinases: A Family of Protein Kinases with Diverse Biological Functions. Microbiology and Molecular Biology Reviews 2004, 68, 320–344. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Fergusson, J.; Salimi, M.; Panse, I.; Ussher, J.E.; Hegazy, A.N.; Vinall, S.L.; Jackson, D.G.; Hunter, M.G.; Pettipher, R.; et al. Prostaglandin D2 and Leukotriene E4 Synergize to Stimulate Diverse TH2 Functions and TH2 Cell/Neutrophil Crosstalk. Journal of Allergy and Clinical Immunology 2015, 135, 1358–1366e11. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and Inflammation. Arteriosclerosis, Thrombosis, and Vascular Biology 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Arima, M.; Fukuda, T. Prostaglandin D2and TH2 Inflammation in the Pathogenesis of Bronchial Asthma. The Korean Journal of Internal Medicine 2011, 26, 8. [Google Scholar] [CrossRef] [PubMed]
- Peinhaupt, M.; Sturm, E.M.; Heinemann, A. Prostaglandins and Their Receptors in Eosinophil Function and as Therapeutic Targets. Frontiers in Medicine 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Lazarinis, N.; Bood, J.; Gomez, C.; Kolmert, J.; Lantz, A.-S.; Gyllfors, P.; Davis, A.; Wheelock, C.E.; Dahlén, S.-E.; Dahlén, B. Leukotriene E4 Induces Airflow Obstruction and Mast Cell Activation through the Cysteinyl Leukotriene Type 1 Receptor. Journal of Allergy and Clinical Immunology 2018, 142, 1080–1089. [Google Scholar] [CrossRef]
- Iskarpatyoti, J.A.; Shi, J.; Abraham, M.A.; Rathore, A.P.S.; Miao, Y.; Abraham, S.N. Mast Cell Regranulation Requires a Metabolic Switch Involving MTORC1 and a Glucose-6-Phosphate Transporter. Cell Reports 2022, 40, 111346. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-M.; Kim, Y.-M.; Lee, Y.M.; Kim, H.-S.; Kim, J.D.; Choi, Y.; Kim, K.-W.; Lee, S.-Y.; Kwon, Y.-G. TNF-Related Activation-Induced Cytokine (TRANCE) Induces Angiogenesis through the Activation of Src and Phospholipase c (PLC) in Human Endothelial Cells. Journal of Biological Chemistry 2002, 277, 6799–6805. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast Cells as Sources of Cytokines, Chemokines, and Growth Factors. Immunological Reviews 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Cartmell, T.; Luheshi, G.N.; Rothwell, N.J. Brain Sites of Action of Endogenous Interleukin-1 in the Febrile Response to Localized Inflammation in the Rat. The Journal of Physiology 1999, 518, 585–594. [Google Scholar] [CrossRef]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide Substance P and the Immune Response. Cellular and molecular life sciences : CMLS 2016, 73, 4249–4264. [Google Scholar] [CrossRef] [PubMed]
- Oleszycka, E.; Kwiecien, K.; Kwiecinska, P.; Morytko, A.; Pocalun, N.; Camacho, M.; Brzoza, P.; Zabel, B.A.; Cichy, J. Soluble Mediators in the Function of the Epidermal-Immune-Neuro Unit in the Skin. Frontiers in Immunology 2022, 13. [Google Scholar] [CrossRef]
- Tan, C.L.; Knight, Z.A. Regulation of Body Temperature by the Nervous System. Neuron 2018, 98, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Tsilioni, I.; Russell, I.J.; Stewart, J.M.; Gleason, R.M.; Theoharides, T.C. Neuropeptides CRH, SP, HK-1, and Inflammatory Cytokines IL-6 and TNF Are Increased in Serum of Patients with Fibromyalgia Syndrome, Implicating Mast Cells. Journal of Pharmacology and Experimental Therapeutics 2016, 356, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Peng, Y.; Zhong, C.; Cai, Y.; Zhou, S.; Chen, H.; Zhuang, J.; Zeng, H.; Xu, C.; Xu, H.; et al. Mast Cells Mediate Inflammatory Injury and Aggravate Neurological Impairment in Experimental Subarachnoid Hemorrhage through Microglial PAR-2 Pathway. Frontiers in Cellular Neuroscience 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Scheller, J.; Rose-John, S.; Born, J.; Marshall, L. Enhancing Influence of Intranasal Interleukin-6 on Slowwave Activity and Memory Consolidation during Sleep. The FASEB Journal 2009, 23, 3629–3636. [Google Scholar] [CrossRef] [PubMed]
- Vijay, K. Toll-like Receptors in Immunity and Inflammatory Diseases: Past, Present, and Future. International Immunopharmacology 2018, 59, 391–412. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, A.; Engblom, D. Neural Mechanisms of Inflammation-Induced Fever. The Neuroscientist 2018, 24, 381–399. [Google Scholar] [CrossRef] [PubMed]
- Gurney, J.G.; McPheeters, M.L.; Davis, M.M. Parental Report of Health Conditions and Health Care Use among Children with and without Autism. Archives of Pediatrics & Adolescent Medicine 2006, 160, 825. [Google Scholar] [CrossRef]
- Chang, M.M.; Leeman, S.E. Isolation of a Sialogogic Peptide from Bovine Hypothalamic Tissue and Its Characterization as Substance P. The Journal of Biological Chemistry 1970, 245, 4784–4790. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Zhang, B.; Kempuraj, D.; Tagen, M.; Vasiadi, M.; Angelidou, A.; Alysandratos, K.-D.; Kalogeromitros, D.; Asadi, S.; Stavrianeas, N.; et al. IL-33 Augments Substance P–Induced VEGF Secretion from Human Mast Cells and Is Increased in Psoriatic Skin. Proceedings of the National Academy of Sciences 2010, 107, 4448–4453. [Google Scholar] [CrossRef] [PubMed]
- Taracanova, A.; Tsilioni, I.; Conti, P.; Norwitz, E.R.; Leeman, S.E.; Theoharides, T.C. Substance P and IL-33 Administered Together Stimulate a Marked Secretion of IL-1β from Human Mast Cells, Inhibited by Methoxyluteolin. Proceedings of the National Academy of Sciences of the United States of America 2018, 115, E9381–E9390. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.S.; von Mentzer, B.; Geppetti, P.; Pothoulakis, C.; Bunnett, N.W. Tachykinins and Their Receptors: Contributions to Physiological Control and the Mechanisms of Disease. Physiological Reviews 2014, 94, 265–301. [Google Scholar] [CrossRef] [PubMed]
- Campo, A.; Lafont, A.-G.; Lefranc, B.; Leprince, J.; Tostivint, H.; Kamech, N.; Dufour, S.; Rousseau, K. Tachykinin-3 Genes and Peptides Characterized in a Basal Teleost, the European Eel: Evolutionary Perspective and Pituitary Role. Frontiers in Endocrinology 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.M.; Seda, M.; Candenas, L. Neurokinin B. xPharm: The Comprehensive Pharmacology Reference 2007, 1–4. [CrossRef]
- Conti, P.; Pregliasco, F.E.; Bellomo, R.G.; Gallenga, C.E.; Caraffa, A.; Kritas, S.K.; Lauritano, D.; Ronconi, G. Mast Cell Cytokines IL-1, IL-33, and IL-36 Mediate Skin Inflammation in Psoriasis: A Novel Therapeutic Approach with the Anti-Inflammatory Cytokines IL-37, IL-38, and IL-1Ra. International Journal of Molecular Sciences 2021, 22, 8076. [Google Scholar] [CrossRef] [PubMed]
- Minnone, G.; De Benedetti, F.; Bracci-Laudiero, L. NGF and Its Receptors in the Regulation of Inflammatory Response. International Journal of Molecular Sciences 2017, 18, 1028. [Google Scholar] [CrossRef] [PubMed]
- van der Kleij, H.P.M.; Ma, D.; Redegeld, F.A.M.; Kraneveld, A.D.; Nijkamp, F.P.; Bienenstock, J. Functional Expression of Neurokinin 1 Receptors on Mast Cells Induced by IL-4 and Stem Cell Factor. The Journal of Immunology 2003, 171, 2074–2079. [Google Scholar] [CrossRef]
- Cai, D.; Liu, T. Hypothalamic Inflammation: A Double-Edged Sword to Nutritional Diseases. Annals of the New York Academy of Sciences 2011, 1243, E1–E39. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Alysandratos, K.-D.; Angelidou, A.; Delivanis, D.-A.; Sismanopoulos, N.; Zhang, B.; Asadi, S.; Vasiadi, M.; Weng, Z.; Miniati, A.; et al. Mast Cells and Inflammation. Biochimica et Biophysica Acta 2012, 1822, 21–33. [Google Scholar] [CrossRef]
- Zieglgänsberger, W. Substance P and Pain Chronicity. Cell and Tissue Research 2019, 375, 227–241. [Google Scholar] [CrossRef]
- Montana, G.; Lampiasi, N. Substance P Induces HO-1 Expression in RAW 264. 7 Cells Promoting Switch towards M2-like Macrophages. PLOS ONE 2016, 11, e0167420. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Shi, X.; Li, X.; Zou, J.; Zhou, C.; Liu, W.; Shao, H.; Chen, H.; Shi, L. Neurotransmitter and Neuropeptide Regulation of Mast Cell Function: A Systematic Review. Journal of Neuroinflammation 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Traina, G. The Role of Mast Cells in the Gut and Brain. Journal of Integrative Neuroscience 2021, 20, 185. [Google Scholar] [CrossRef] [PubMed]
- Franke, K.; Wang, Z.; Zuberbier, T.; Babina, M. Cytokines Stimulated by IL-33 in Human Skin Mast Cells: Involvement of NF-ΚB and P38 at Distinct Levels and Potent Co-Operation with FcεRI and MRGPRX2. International Journal of Molecular Sciences 2021, 22, 3580. [Google Scholar] [CrossRef] [PubMed]
- Eissmann, M.F.; Buchert, M.; Ernst, M. IL33 and Mast Cells—the Key Regulators of Immune Responses in Gastrointestinal Cancers? Frontiers in Immunology 2020, 11. [Google Scholar] [CrossRef]
- Taracanova, A.; Alevizos, M.; Karagkouni, A.; Weng, Z.; Norwitz, E.; Conti, P.; Leeman, S.E.; Theoharides, T.C. SP and IL-33 Together Markedly Enhance TNF Synthesis and Secretion from Human Mast Cells Mediated by the Interaction of Their Receptors. Proceedings of the National Academy of Sciences of the United States of America 2017, 114, E4002–E4009. [Google Scholar] [CrossRef] [PubMed]
- Murdaca, G.; Greco, M.; Tonacci, A.; Negrini, S.; Borro, M.; Puppo, F.; Gangemi, S. IL-33/IL-31 Axis in Immune-Mediated and Allergic Diseases. International Journal of Molecular Sciences 2019, 20, 5856. [Google Scholar] [CrossRef]
- Carniglia, L.; Ramírez, D.; Durand, D.; Saba, J.; Turati, J.; Caruso, C.; Scimonelli, T.N.; Lasaga, M. Neuropeptides and Microglial Activation in Inflammation, Pain, and Neurodegenerative Diseases. Mediators of Inflammation 2017, 2017, 1–23. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Tsilioni, I.; Bawazeer, M. Mast Cells, Neuroinflammation and Pain in Fibromyalgia Syndrome. Frontiers in Cellular Neuroscience 2019, 13. [Google Scholar] [CrossRef]
- Holzer, P. Local Effector Functions of Capsaicin-Sensitive Sensory Nerve Endings: Involvement of Tachykinins, Calcitonin Gene-Related Peptide and Other Neuropeptides. Neuroscience 1988, 24, 739–768. [Google Scholar] [CrossRef]
- Eglezos, A.; Giuliani, S.; Viti, G.; Maggi, C.A. Direct Evidence That Capsaicin-Induced Plasma Protein Extravasation Is Mediated through Tachykinin NK1 Receptors. European Journal of Pharmacology 1991, 209, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, M.; Neri, G.; Maccauro, G.; Tripodi, D.; Varvara, G.; Saggini, A.; Potalivo, G.; Castellani, M.L.; Fulcheri, M.; Rosati, M.; et al. Impact and Neuropeptide Substance Pan Inflammatory Compound on Arachidonic Acid Compound Generation. International Journal of Immunopathology and Pharmacology 2012, 25, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Maletic, V.; Raison, C. Integrated Neurobiology of Bipolar Disorder. Frontiers in Psychiatry 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Okayama, Y.; Church, M.K. Comparison of the Modulatory Effect of Ketotifen, Sodium Cromoglycate, Procaterol and Salbutamol in Human Skin, Lung and Tonsil Mast Cells. International Archives of Allergy and Immunology 1992, 97, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Harris, G.W. NEURAL CONTROL of the PITUITARY GLAND. Physiological Reviews 1948, 28, 139–179. [Google Scholar] [CrossRef] [PubMed]
- Pacak, K.; Palkovits, M.; Kopin, I.J.; Goldstein, D.S. Stress-Induced Norepinephrine Release in the Hypothalamic Paraventricular Nucleus and Pituitary-Adrenocortical and Sympathoadrenal Activity: In Vivo Microdialysis Studies. Frontiers in Neuroendocrinology 1995, 16, 89–150. [Google Scholar] [CrossRef] [PubMed]
- Wrońska, D.; Kania, B.F.; Błachuta, M. Direct Effect of Hypothalamic Neuropeptides on the Release of Catecholamines by Adrenal Medulla in Sheep – Study Ex Vivo. Polish Journal of Veterinary Sciences 2017, 20, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.A.; Carson, M.J.; Nair, M.G. Non-Traditional Cytokines: How Catecholamines and Adipokines Influence Macrophages in Immunity, Metabolism and the Central Nervous System. Cytokine 2015, 72, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. International Journal of Molecular Sciences 2019, 20, 6008. [Google Scholar] [CrossRef]
- Chen, R.; Lewis, K.A.; Perrin, M.H.; Vale, W.W. Expression Cloning of a Human Corticotropin-Releasing-Factor Receptor. Proceedings of the National Academy of Sciences 1993, 90, 8967–8971. [Google Scholar] [CrossRef]
- Lovenberg, T.W.; Liaw, C.W.; Grigoriadis, D.E.; Clevenger, W.; Chalmers, D.T.; De Souza, E.B.; Oltersdorf, T. Cloning and Characterization of a Functionally Distinct Corticotropin-Releasing Factor Receptor Subtype from Rat Brain. Proceedings of the National Academy of Sciences 1995, 92, 836–840. [Google Scholar] [CrossRef] [PubMed]
- Lovenberg, T.W.; Chalmers, D.T.; Liu, C.; De Souza, E.B. CRF2 Alpha and CRF2 Beta Receptor MRNAs Are Differentially Distributed between the Rat Central Nervous System and Peripheral Tissues. Endocrinology 1995, 136, 4139–4142. [Google Scholar] [CrossRef] [PubMed]
- Vale, W.; Spiess, J.; Rivier, C.; Rivier, J. Characterization of a 41-Residue Ovine Hypothalamic Peptide That Stimulates Secretion of Corticotropin and Beta-Endorphin. Science (New York, N.Y.) 1981, 213, 1394–1397. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.J.; Sharma, S. Physiology, Adrenocorticotropic Hormone (ACTH) Available online: https://www.ncbi.nlm.nih.gov/books/NBK500031/.
- Miller, T.; Gibbison, B.; Russell, G.M. Hypothalamic–Pituitary–Adrenal Function during Health, Major Surgery, and Critical Illness. BJA Education 2017, 17, 16–21. [Google Scholar] [CrossRef]
- Silverman, M.N.; Pearce, B.D.; Biron, C.A.; Miller, A.H. Immune Modulation of the Hypothalamic-Pituitary-Adrenal (HPA) Axis during Viral Infection. Viral Immunology 2005, 18, 41–78. [Google Scholar] [CrossRef] [PubMed]
- Wamsteeker Cusulin, J.I.; Füzesi, T.; Watts, A.G.; Bains, J.S. Characterization of Corticotropin-Releasing Hormone Neurons in the Paraventricular Nucleus of the Hypothalamus of Crh-IRES-Cre Mutant Mice. PLoS ONE 2013, 8, e64943. [Google Scholar] [CrossRef]
- Ferguson, A.V.; Latchford, K.J.; Samson, W.K. The Paraventricular Nucleus of the Hypothalamus – a Potential Target for Integrative Treatment of Autonomic Dysfunction. Expert Opinion on Therapeutic Targets 2008, 12, 717–727. [Google Scholar] [CrossRef]
- Emeric-Sauval, E. Corticotropin-Releasing Factor (CRF) — a Review. Psychoneuroendocrinology 1986, 11, 277–294. [Google Scholar] [CrossRef] [PubMed]
- OELKERS, W.; BOELKE, T.; BÄHR, V.; EXNER, P.; FAUST, B.; HARENDT, H. Dose-Response Relationships between Plasma Adrenocorticotropin (ACTH), Cortisol, Aldosterone, and 18-Hydroxycorticosterone after Injection of ACTH-(1–39) or Human Corticotropin-Releasing Hormone in Man*. The Journal of Clinical Endocrinology & Metabolism 1988, 66, 181–186. [Google Scholar] [CrossRef]
- McCann, S.M.; Antunes-Rodrigues, J.; Franci, C.R.; Anselmo-Franci, J.A.; Karanth, S.; Rettori, V. Role of the Hypothalamic Pituitary Adrenal Axis in the Control of the Response to Stress and Infection. Brazilian Journal of Medical and Biological Research 2000, 33, 1121–1131. [Google Scholar] [CrossRef]
- Wieseler-Frank, J.; Jekich, B.M.; Mahoney, J.H.; Bland, S.T.; Maier, S.F.; Watkins, L.R. A Novel Immune-To-CNS Communication Pathway: Cells of the Meninges Surrounding the Spinal Cord CSF Space Produce Proinflammatory Cytokines in Response to an Inflammatory Stimulus. Brain, Behavior, and Immunity 2007, 21, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, H.; Shichi, H.; Yamamoto, M.; Takahashi, Y. The Mechanisms Underlying Autonomous Adrenocorticotropic Hormone Secretion in Cushing’s Disease. International Journal of Molecular Sciences 2020, 21, 9132. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Lu, M.; Yuan, H.; Chen, T.; Han, X. Mast Cell-Mediated Neuroinflammation May Have a Role in Attention Deficit Hyperactivity Disorder (Review). Experimental and Therapeutic Medicine 2020, 20, 714–726. [Google Scholar] [CrossRef] [PubMed]
- DESIMONE, R. Nerve Growth Factor Released into the Bloodstream Following Intraspecific Fighting Induces Mast Cell Degranulation in Adult Male Mice. Brain, Behavior, and Immunity 1990, 4, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kempuraj, D.; Marchand, J.; Tzianoumis, L.; Vasiadi, M.; Katsarou-Katsari, A.; Makris, M.; Kalogeromitros, D. Urticaria Pigmentosa Associated with Acute Stress and Lesional Skin Mast-Cell Expression of CRF-R1. Clinical and Experimental Dermatology 2009, 34, e163–e166. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Dong, H.-Q.; Liu, Y.-H.; Ji, M.-H.; Zhang, X.; Dai, H.-Y.; Sun, Z.-C.; Liu, L.; Zhou, J.; Sha, H.-H.; et al. Laparotomy-Induced Peripheral Inflammation Activates NR2B Receptors on the Brain Mast Cells and Results in Neuroinflammation in a Vagus Nerve-Dependent Manner. Frontiers in Cellular Neuroscience 2022, 16. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, I.; Inoue, Y.; Shimada, T.; Aikawa, T. Brain Mast Cells Act as an Immune Gate to the Hypothalamic-Pituitary-Adrenal Axis in Dogs. Journal of Experimental Medicine 2001, 194, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Papadopoulou, N.; Kempuraj, D.; Boucher, W.S.; Sugimoto, K.; Cetrulo, C.L.; Theoharides, T.C. Human Mast Cells Express Corticotropin-Releasing Hormone (CRH) Receptors and CRH Leads to Selective Secretion of Vascular Endothelial Growth Factor. The Journal of Immunology 2005, 174, 7665–7675. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Majid, R.M.; Marshall, J.S. Prostaglandin E2 Induces Degranulation-Independent Production of Vascular Endothelial Growth Factor by Human Mast Cells. The Journal of Immunology 2004, 172, 1227–1236. [Google Scholar] [CrossRef]
- Leeman, S.E.; Mroz, E.A. Substance P. Life Sciences 1974, 15, 2033–2044. [Google Scholar] [CrossRef]
- Christou, N.; Blondy, S.; David, V.; Verdier, M.; Lalloué, F.; Jauberteau, M.-O.; Mathonnet, M.; Perraud, A. Neurotensin Pathway in Digestive Cancers and Clinical Applications: An Overview. Cell Death & Disease 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Martinez-Fong, D.; Trédaniel, J.; Forgez, P. Neurotensin and Its High Affinity Receptor 1 as a Potential Pharmacological Target in Cancer Therapy. Frontiers in Endocrinology 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, L.E.; Leinninger, G.M. Role of Central Neurotensin in Regulating Feeding: Implications for the Development and Treatment of Body Weight Disorders. Biochimica et biophysica acta 2018, 1864, 900–916. [Google Scholar] [CrossRef] [PubMed]
- St-Gelais, F.; Jomphe, C.; Trudeau, L.-É. The Role of Neurotensin in Central Nervous System Pathophysiology: What Is the Evidence? Journal of Psychiatry and Neuroscience 2006, 31, 229–245. [Google Scholar] [PubMed]
- Servonnet, A.; Minogianis, E.-A.; Bouchard, C.; Bédard, A.-M.; Lévesque, D.; Rompré, P.-P.; Samaha, A.-N. Neurotensin in the Nucleus Accumbens Reverses Dopamine Supersensitivity Evoked by Antipsychotic Treatment. Neuropharmacology 2017, 123, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Tuladhar, A.M.; de Leeuw, F.-E. Thalamus: A Key Player in Alcohol Use Disorder and Korsakoff’s Syndrome. Brain 2019, 142, 1170–1172. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Annals of gastroenterology 2015, 28, 203–209. [Google Scholar] [PubMed]
- Boules, M.; Li, Z.; Smith, K.; Fredrickson, P.; Richelson, E. Diverse Roles of Neurotensin Agonists in the Central Nervous System. Frontiers in Endocrinology 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kempuraj, D.; Tagen, M.; Conti, P.; Kalogeromitros, D. Differential Release of Mast Cell Mediators and the Pathogenesis of Inflammation. Immunological Reviews 2007, 217, 65–78. [Google Scholar] [CrossRef]
- Mustain, W.C.; Rychahou, P.G.; Evers, B.M. The Role of Neurotensin in Physiologic and Pathologic Processes. Current Opinion in Endocrinology, Diabetes & Obesity 2011, 18, 75–82. [Google Scholar] [CrossRef]
- Zhang, B.; Asadi, S.; Weng, Z.; Sismanopoulos, N.; Theoharides, T.C. Stimulated Human Mast Cells Secrete Mitochondrial Components That Have Autocrine and Paracrine Inflammatory Actions. PLoS ONE 2012, 7, e49767. [Google Scholar] [CrossRef] [PubMed]
- Tsilioni, I.; Patel, A.B.; Pantazopoulos, H.; Berretta, S.; Conti, P.; Leeman, S.E.; Theoharides, T.C. IL-37 Is Increased in Brains of Children with Autism Spectrum Disorder and Inhibits Human Microglia Stimulated by Neurotensin. Proceedings of the National Academy of Sciences 2019, 116, 21659–21665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M.; An, J. Cytokines, Inflammation, and Pain. International Anesthesiology Clinics 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Andrea Predonzani, B.C.; Andrielly HR Agnellini, B.M. Spotlights on Immunological Effects of Reactive Nitrogen Species: When Inflammation Says Nitric Oxide. World Journal of Experimental Medicine 2015, 5, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Wink, D.A.; Hines, H.B.; Cheng, R.Y.S.; Switzer, C.H.; Flores-Santana, W.; Vitek, M.P.; Ridnour, L.A.; Colton, C.A. Nitric Oxide and Redox Mechanisms in the Immune Response. Journal of Leukocyte Biology 2011, 89, 873–891. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Stewart, J.M.; Taracanova, A.; Conti, P.; Zouboulis, C.C. Neuroendocrinology of the Skin. Reviews in Endocrine & Metabolic Disorders 2016, 17, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Marek-Jozefowicz, L.; Czajkowski, R.; Borkowska, A.; Nedoszytko, B.; Żmijewski, M.A.; Cubała, W.J.; Slominski, A.T. The Brain–Skin Axis in Psoriasis—Psychological, Psychiatric, Hormonal, and Dermatological Aspects. International Journal of Molecular Sciences 2022, 23, 669. [Google Scholar] [CrossRef] [PubMed]
- Redegeld, F.A.; Yu, Y.; Kumari, S.; Charles, N.; Blank, U. Non-IgE Mediated Mast Cell Activation. Immunological Reviews 2018, 282, 87–113. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Lal, G. Neurokinin Receptors and Their Implications in Various Autoimmune Diseases. Current Research in Immunology 2021, 2, 66–78. [Google Scholar] [CrossRef]
- Fulenwider, H.D.; Smith, B.M.; Nichenko, A.S.; Carpenter, J.M.; Nennig, S.E.; Cheng, K.; Rice, K.C.; Schank, J.R. Cellular and Behavioral Effects of Lipopolysaccharide Treatment Are Dependent upon Neurokinin-1 Receptor Activation. Journal of Neuroinflammation 2018, 15. [Google Scholar] [CrossRef]
- Aich, A.; Afrin, L.B.; Gupta, K. Mast Cell-Mediated Mechanisms of Nociception. International Journal of Molecular Sciences 2015, 16, 29069–29092. [Google Scholar] [CrossRef] [PubMed]
- da Fonseca, A.C.C.; Matias, D.; Garcia, C.; Amaral, R.; Geraldo, L.H.; Freitas, C.; Lima, F.R.S. The Impact of Microglial Activation on Blood-Brain Barrier in Brain Diseases. Frontiers in Cellular Neuroscience 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Damsker, J.M.; Hansen, A.M.; Caspi, R.R. Th1 and Th17 Cells. Annals of the New York Academy of Sciences 2010, 1183, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.B.; Tsilioni, I.; Leeman, S.E.; Theoharides, T.C. Neurotensin Stimulates Sortilin and MTOR in Human Microglia Inhibitable by Methoxyluteolin, a Potential Therapeutic Target for Autism. Proceedings of the National Academy of Sciences 2016, 113, E7049–E7058. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Mu, R.; Wei, X. The Roles of IL-1 Family Cytokines in the Pathogenesis of Systemic Sclerosis. Frontiers in Immunology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Boutet, M.-A.; Nerviani, A.; Pitzalis, C. IL-36, IL-37, and IL-38 Cytokines in Skin and Joint Inflammation: A Comprehensive Review of Their Therapeutic Potential. International Journal of Molecular Sciences 2019, 20, 1257. [Google Scholar] [CrossRef]
- Zhao, M.; Li, Y.; Guo, C.; Wang, L.; Chu, H.; Zhu, F.; Li, Y.; Wang, X.; Wang, Q.; Zhao, W.; et al. IL-37 Isoform D Downregulates Pro-Inflammatory Cytokines Expression in a Smad3-Dependent Manner. Cell Death & Disease 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Amo-Aparicio, J.; Neff, C.P.; Tengesdal, I.W.; Azam, T.; Palmer, B.E.; López-Vales, R.; Bufler, P.; Dinarello, C.A. Role for Nuclear Interleukin-37 in the Suppression of Innate Immunity. Proceedings of the National Academy of Sciences 2019, 116, 4456–4461. [Google Scholar] [CrossRef] [PubMed]
- Nold-Petry, C.A.; Lo, C.Y.; Rudloff, I.; Elgass, K.D.; Li, S.; Gantier, M.P.; Lotz-Havla, A.S.; Gersting, S.W.; Cho, S.X.; Lao, J.C.; et al. IL-37 Requires the Receptors IL-18Rα and IL-1R8 (SIGIRR) to Carry out Its Multifaceted Anti-Inflammatory Program upon Innate Signal Transduction. Nature Immunology 2015, 16, 354–365. [Google Scholar] [CrossRef]
- Parihar, A.; Eubank, T.D.; Doseff, A.I. Monocytes and Macrophages Regulate Immunity through Dynamic Networks of Survival and Cell Death. Journal of Innate Immunity 2010, 2, 204–215. [Google Scholar] [CrossRef]
- Su, Z.; Tao, X. Current Understanding of IL-37 in Human Health and Disease. Frontiers in Immunology 2021, 12. [Google Scholar] [CrossRef]
- Shuai, X.; Wei-min, L.; Tong, Y.; Dong, N.; Sheng, Z.; Yao, Y. Expression of IL-37 Contributes to the Immunosuppressive Property of Human CD4+CD25+ Regulatory T Cells. Scientific Reports 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Lonnemann, N.; Hosseini, S.; Ohm, M.; Geffers, R.; Hiller, K.; Dinarello, C.A.; Korte, M. IL-37 Expression Reduces Acute and Chronic Neuroinflammation and Rescues Cognitive Impairment in an Alzheimer’s Disease Mouse Model. eLife 2022, 11. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, S.; Li, L.; Huang, Z.; Ma, K. Anti-Inflammatory Effect of IL-37-Producing T-Cell Population in DSS-Induced Chronic Inflammatory Bowel Disease in Mice. International Journal of Molecular Sciences 2018, 19, 3884. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Nold-Petry, C.; Nold, M.; Fujita, M.; Li, S.; Kim, S.; Bufler, P. Suppression of Innate Inflammation and Immunity by Interleukin-37. European Journal of Immunology 2016, 46, 1067–1081. [Google Scholar] [CrossRef]
- Cavalli, G.; Tengesdal, I.W.; Gresnigt, M.; Nemkov, T.; Arts, R.J.W.; Domínguez-Andrés, J.; Molteni, R.; Stefanoni, D.; Cantoni, E.; Cassina, L.; et al. The Anti-Inflammatory Cytokine Interleukin-37 Is an Inhibitor of Trained Immunity. Cell Reports 2021, 35, 108955. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Neff, C.P.; Barber, K.; Hong, J.; Luo, Y.; Azam, T.; Palmer, B.E.; Fujita, M.; Garlanda, C.; Mantovani, A.; et al. Extracellular Forms of IL-37 Inhibit Innate Inflammation in Vitro and in Vivo but Require the IL-1 Family Decoy Receptor IL-1R8. Proceedings of the National Academy of Sciences 2015, 112, 2497–2502. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-T.; Zhu, D.; Mou, T.; Guo, Z.; Pu, J.-L.; Chen, Q.-S.; Wei, X.-F.; Wu, Z.-J. IL-37 Induces Autophagy in Hepatocellular Carcinoma Cells by Inhibiting the PI3K/AKT/MTOR Pathway. Molecular Immunology 2017, 87, 132–140. [Google Scholar] [CrossRef]
- Zhang, S.R.; Nold, M.F.; Tang, S.-C.; Bui, C.B.; Nold, C.A.; Arumugam, T.V.; Drummond, G.R.; Sobey, C.G.; Kim, H.A. IL-37 Increases in Patients after Ischemic Stroke and Protects from Inflammatory Brain Injury, Motor Impairment and Lung Infection in Mice. Scientific Reports 2019, 9. [Google Scholar] [CrossRef]
- McCurdy, S.; Liu, C.A.; Yap, J.; Boisvert, W.A. Potential Role of IL-37 in Atherosclerosis. Cytokine 2019, 122, 154169. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Li, M.; Tan, S.; Wu, D. IL-37 as a Potential Biotherapeutics of Inflammatory Diseases. Current Drug Targets 2020, 21. [Google Scholar] [CrossRef]
- Liu, X.; Quan, N. Microglia and CNS Interleukin-1: Beyond Immunological Concepts. Frontiers in Neurology 2018, 9. [Google Scholar] [CrossRef]
- Pringle, A.K.; Gardner, C.R.; Walker, R.J. Reduction of Cerebellar GABAA Responses by Interleukin-1 (IL-1) through an Indomethacin Insensitive Mechanism. Neuropharmacology 1996, 35, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Goldstein, J.D.; Mermoud, L.; Diaz-Barreiro, A.; Palmer, G. IL-1 Family Antagonists in Mouse and Human Skin Inflammation. Frontiers in Immunology 2021, 12. [Google Scholar] [CrossRef]
- Esmaeilzadeh, A.; Bahmaie, N.; Nouri, E.; Hajkazemi, M.J.; Zareh Rafie, M. Immunobiological Properties and Clinical Applications of Interleukin-38 for Immune-Mediated Disorders: A Systematic Review Study. International Journal of Molecular Sciences 2021, 22, 12552. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Peng, X.; Li, Y.; Li, M. Role of IL-38 and Its Related Cytokines in Inflammation. Mediators of Inflammation 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Huang, Z.; Li, H.; Liu, X.; Zheng, S.; Su, W. IL-38: A New Player in Inflammatory Autoimmune Disorders. Biomolecules 2019, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-D.; Huang, A.-F. Role of Interleukin-38 in Chronic Inflammatory Diseases: A Comprehensive Review. Frontiers in Immunology 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- van de Veerdonk, F.L.; Stoeckman, A.K.; Wu, G.; Boeckermann, A.N.; Azam, T.; Netea, M.G.; Joosten, L.A.B.; van der Meer, J.W.M.; Hao, R.; Kalabokis, V.; et al. IL-38 Binds to the IL-36 Receptor and Has Biological Effects on Immune Cells Similar to IL-36 Receptor Antagonist. Proceedings of the National Academy of Sciences 2012, 109, 3001–3005. [Google Scholar] [CrossRef]
- Brevi, A.; Cogrossi, L.L.; Grazia, G.; Masciovecchio, D.; Impellizzieri, D.; Lacanfora, L.; Grioni, M.; Bellone, M. Much More than IL-17A: Cytokines of the IL-17 Family between Microbiota and Cancer. Frontiers in Immunology 2020, 11. [Google Scholar] [CrossRef]
- Waisman, A.; Hauptmann, J.; Regen, T. The Role of IL-17 in CNS Diseases. Acta Neuropathologica 2015, 129, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, M.; Weymar, A.; Bernreuther, C.; Velden, J.; Arunachalam, P.; Steinbach, K.; Orthey, E.; Arumugam, T.V.; Leypoldt, F.; Simova, O.; et al. Neutralization of the IL-17 Axis Diminishes Neutrophil Invasion and Protects from Ischemic Stroke. Blood 2012, 120, 3793–3802. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.R.; Aldrich, A.L.; Mariani, M.M.; Vidlak, D.; Esen, N.; Kielian, T. Toll-like Receptor 2 (TLR2) Deficiency Leads to Increased Th17 Infiltrates in Experimental Brain Abscesses. Journal of immunology (Baltimore, Md. : 1950) 2009, 182, 7119–7130. [Google Scholar] [CrossRef] [PubMed]
- Milovanovic, J.; Arsenijevic, A.; Stojanovic, B.; Kanjevac, T.; Arsenijevic, D.; Radosavljevic, G.; Milovanovic, M.; Arsenijevic, N. Interleukin-17 in Chronic Inflammatory Neurological Diseases. Frontiers in Immunology 2020, 11. [Google Scholar] [CrossRef]
- Siffrin, V.; Radbruch, H.; Glumm, R.; Niesner, R.; Paterka, M.; Herz, J.; Leuenberger, T.; Lehmann, S.M.; Luenstedt, S.; Rinnenthal, J.L.; et al. In Vivo Imaging of Partially Reversible Th17 Cell-Induced Neuronal Dysfunction in the Course of Encephalomyelitis. Immunity 2010, 33, 424–436. [Google Scholar] [CrossRef]
- Migliorini, P.; Italiani, P.; Pratesi, F.; Puxeddu, I.; Boraschi, D. The IL-1 Family Cytokines and Receptors in Autoimmune Diseases. Autoimmunity Reviews 2020, 19, 102617. [Google Scholar] [CrossRef]
Figure 1.
Here we depict a mast cell (MC) activated by IgE binding the receptor FcRI, leading a biochemical cascade which results in the transcription and the generation of inflammatory cytokines.
Figure 1.
Here we depict a mast cell (MC) activated by IgE binding the receptor FcRI, leading a biochemical cascade which results in the transcription and the generation of inflammatory cytokines.
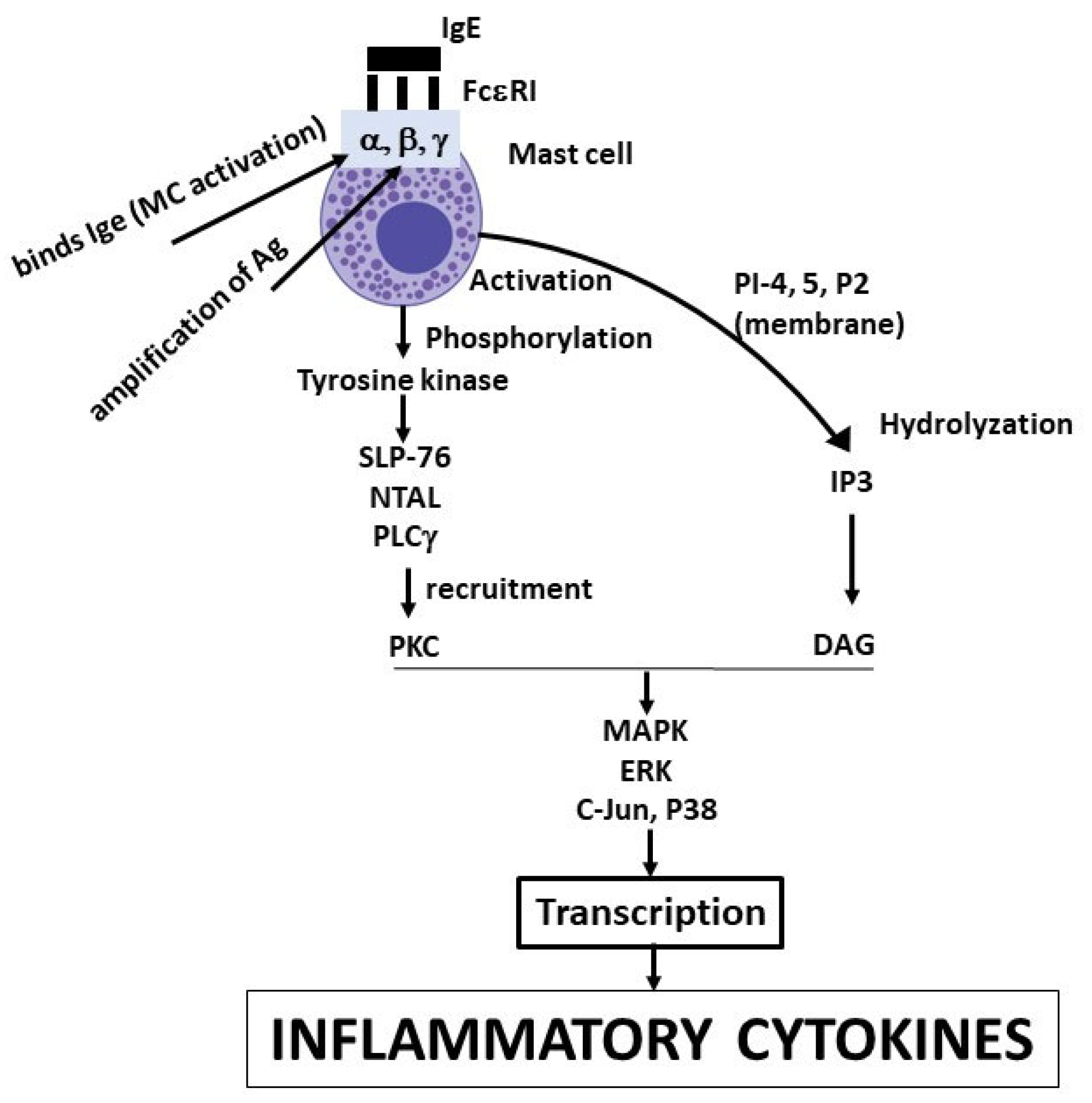
Figure 2.
Brain injury causes the activation of pro-IL-1 which is cleaved by caspase-1, leading to mature IL-1 which binds its receptor on the cell membrane, resulting in NF-B activation and gene expression.
Figure 2.
Brain injury causes the activation of pro-IL-1 which is cleaved by caspase-1, leading to mature IL-1 which binds its receptor on the cell membrane, resulting in NF-B activation and gene expression.
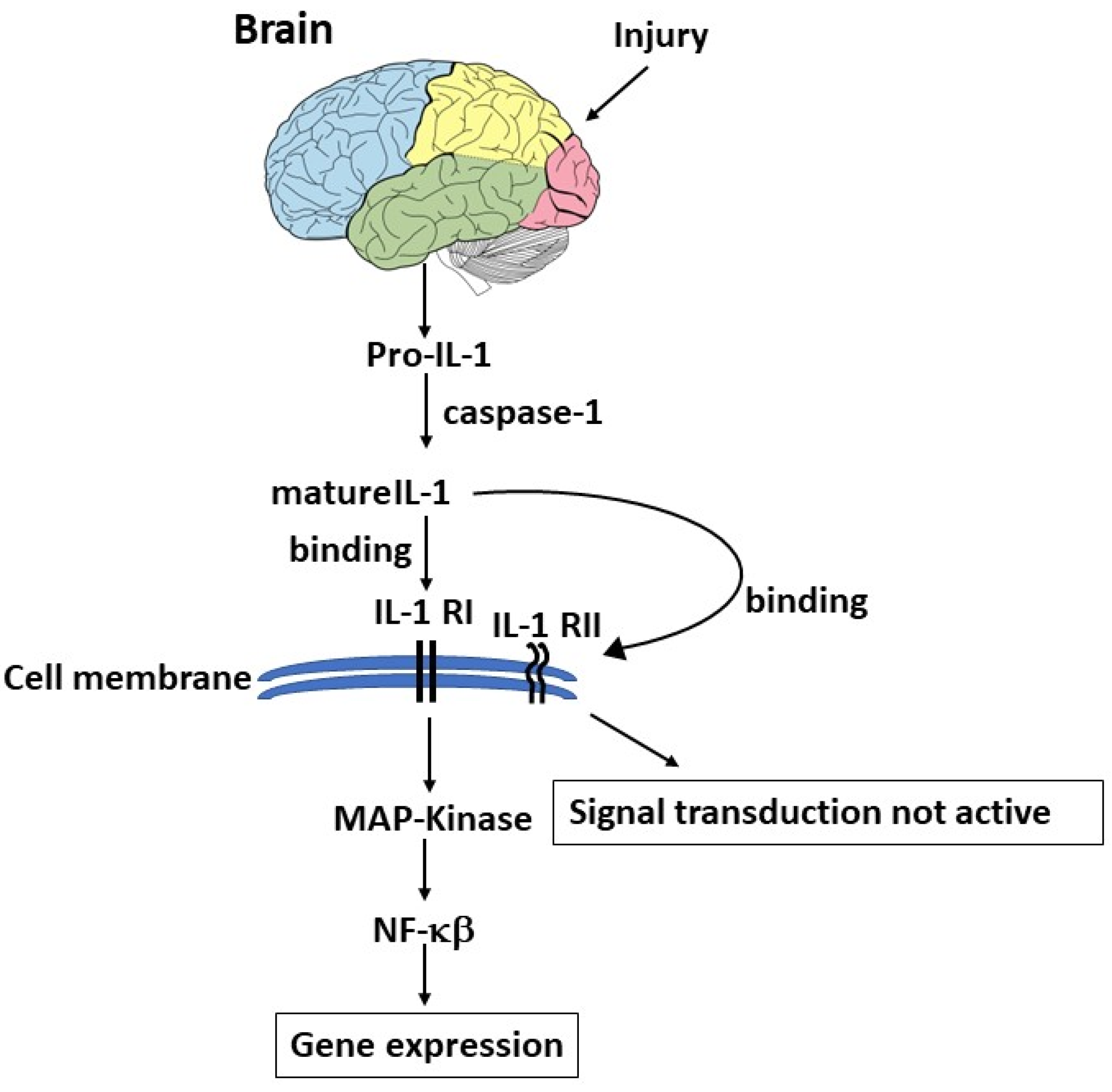
Figure 3.
The mast cell (MC) can be activated by neurotensin (NT) through its receptor NTR1, leading to a biochemical cascade in the cytoplasm and nucleus that results in transcription proteins.
Figure 3.
The mast cell (MC) can be activated by neurotensin (NT) through its receptor NTR1, leading to a biochemical cascade in the cytoplasm and nucleus that results in transcription proteins.
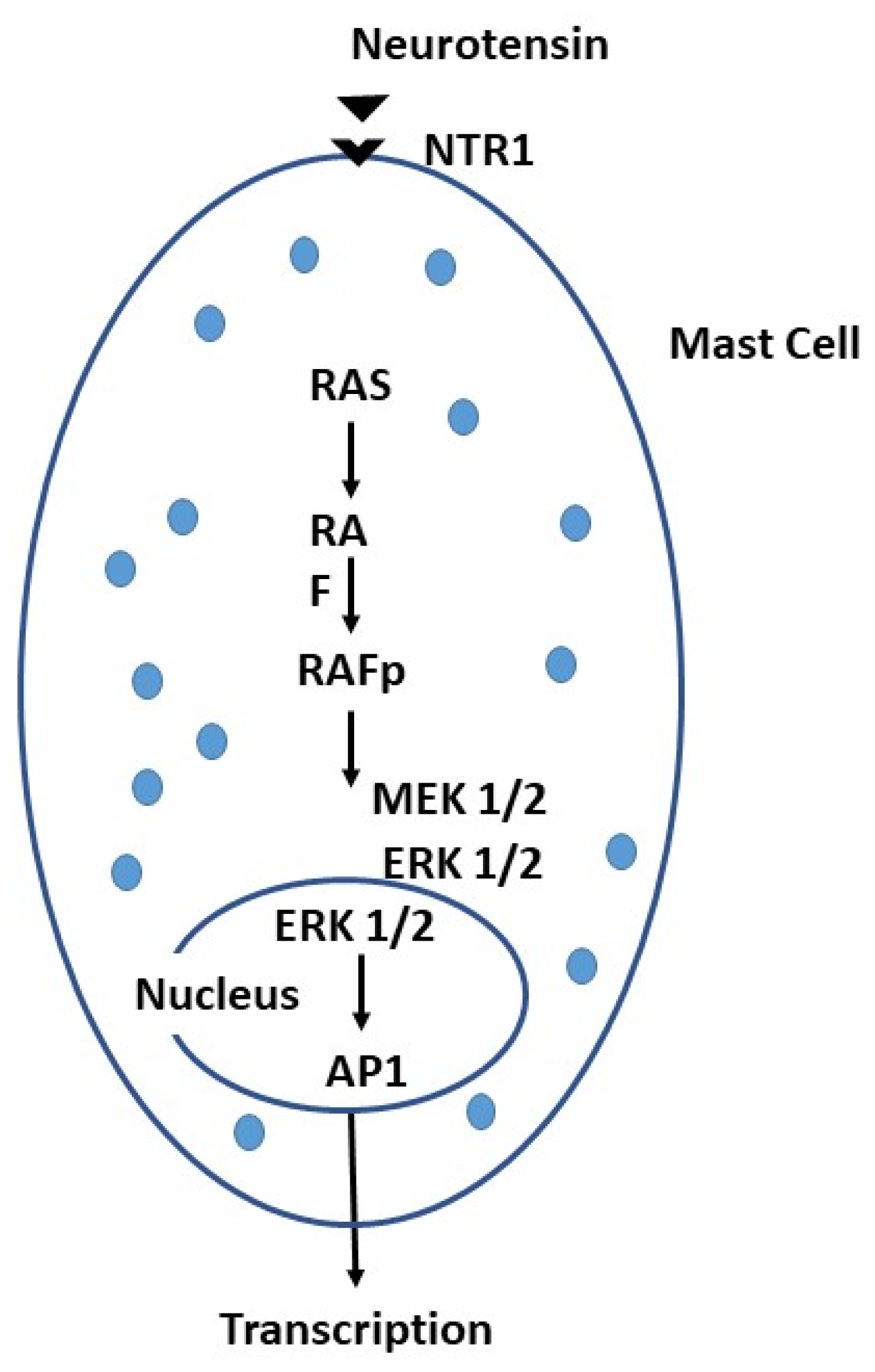
Figure 4.
Brain microglia can be activated by neuropeptides, which can also activate mast cells (MCs), leading to pro-inflammatory cytokines, brain disorders, and neuron disorders, an effect that can be inhibited by IL-37 or IL-38.
Figure 4.
Brain microglia can be activated by neuropeptides, which can also activate mast cells (MCs), leading to pro-inflammatory cytokines, brain disorders, and neuron disorders, an effect that can be inhibited by IL-37 or IL-38.
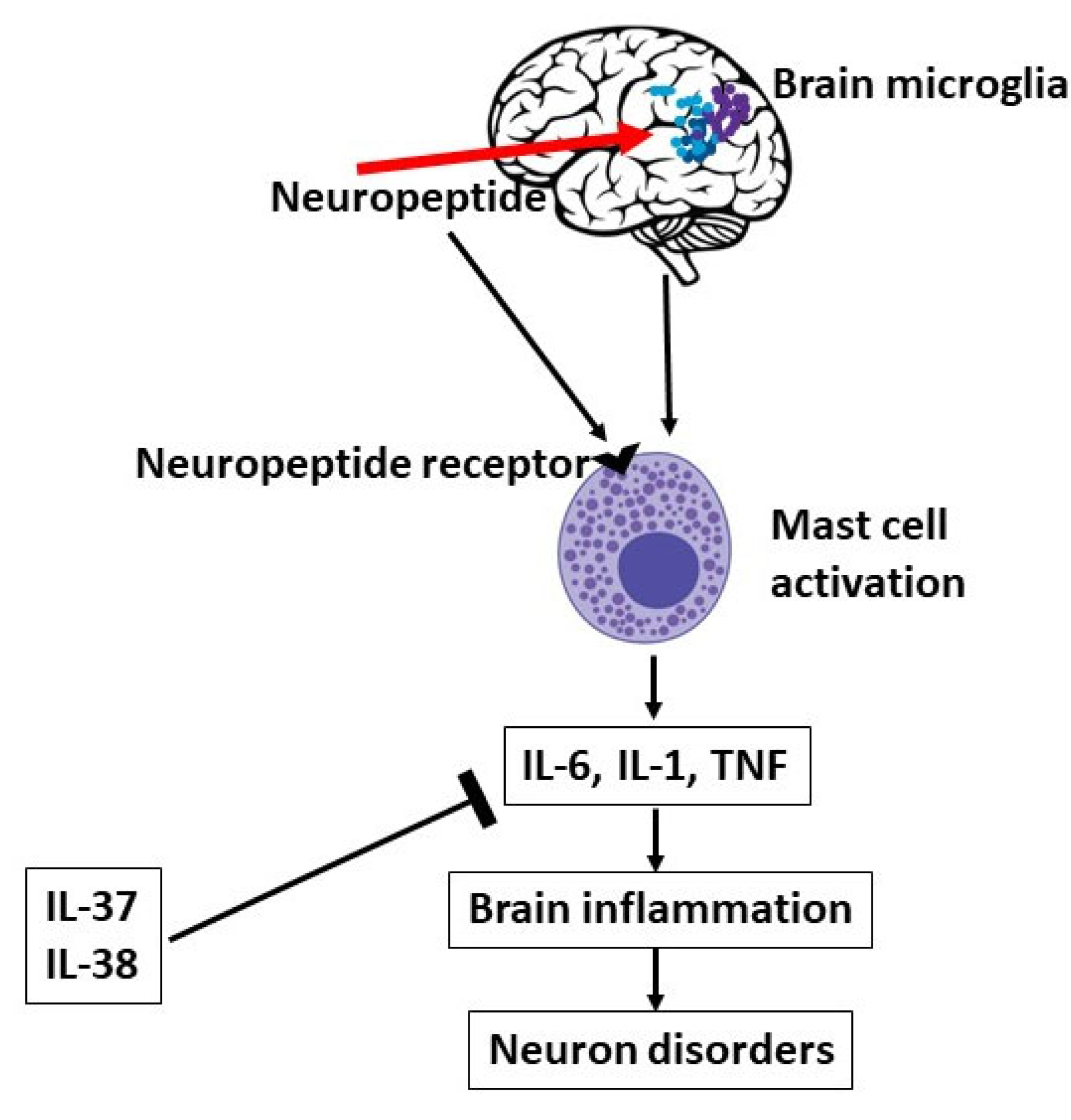

Table 1.
Compounds Released by Mast Cells after Activation.
| Compound de novo synthesis: IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-10, IL-13, TNF, NO, VEGF.Arachidonic acid products: prostaglandin PGD2, leukotriene LTB4, LTC4, Chemokines: IL-8 (CXCL8), MCP-1 (CCL2), MCP-3 (CCL7), MCP-4, RANTES (CCL5), Eotaxin (CCL11) |
| Prestored mediators: chymase, tryptase, CRH, GM-CSF, SCF, NGF, TGF-β, chondroitin, heparin, histamine, serotonin, β-endorphin, SP, VIP, NT |
Table 2.
Mast Cell Triggers without Degranulation.
| IL-6, TNF or VEGF, NT (neurotensin), CRH (corticotropin releasing hormone), LPS (lipopolysaccharide), VIP (vasoactive intestinal peptide), PACAP (pituitary adenylate cyclase activating polypeptide), PCBs (polychlorinated biphenols), PTH (parathyroid hormone), SP (substance P) |
| Heavy metals: Aluminum, Cadmium, Mercury |
Table 3.
Some peptides released by MCs.
| Peptide | Function |
| Neurotensin | Digestive tract and cardiovascular regulation |
| Substance P | Inflammation, pain |
| Kinins (bradykinin) | Inflammation, pain, vasodilation |
| Corticotropin-releasing hormone | Inflammation: vasodilation |
| VEGF | Neovascularization, vasodilation |
| Angiogenin | Neovascularization |
| Endorphins | Analgesia |
| Endothelin | Sepsis |
| Renin | Angiotensin synthesis |
| Urocortin | Inflammation, vasodilation |
| Vasoactive intestinal peptide | Vasodilation, mast cell activation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



