
Submitted:
15 March 2023
Posted:
15 March 2023
You are already at the latest version
Abstract
Background: Sleeping Sickness, Human African Trypanosomiasis (HAT) is a vector- borne disease caused by Trypanosoma brucei (T.b). Sleeping sickness in Ethiopia was reported in 1967 for the first time. Recently in Southern parts of Ethiopia, in August 2022, five (5) cases of sleeping sickness (T. b. rhodesiense) were confirmed. Following this outbreak, the current investigation was aimed to identify the entomological and epidemiological drivers for the reemergence of HAT outbreak and recommend appropriate interventions.
Methods: A cross sectional study design with descriptive data analysis was used. Tsetse fly collection and blood samples from cattle animals were taken. NGU and bio-conical traps were used to determine the distribution (density and abundance) of the vector. About 10μl of blood was collected from the marginal ear vein of 301 cattle animals using the heparinized microhematocrit capillary. The parasite detection was carried out through vector dissection under binocular stereo-microscope (magnification of 60X) and microscopic examination from serum of Animals using the Buffy coat method.
Results: A total of 329 tsetse flies were captured and identified to Glosina (G.) pallidipes 259 (60.4%) and Glossina fuscipes 70 (16.3%). 188 (51.1%) of tsetse flies were collected from Dembagofa with 94 apparent density. Among all captured Tsetse flies, 39 (11.8%) of Tsetse were fed with high female apparent density in each ecological variation: wood land (51), Bush land (20) and grassland (11). Overall, the apparent density of tsetse fly was high in Wood land (93): G. pallidipes (76.5) and Bush land (36.5). Among all examined cattles for the presence of parasite, 9 cattles were detected positive with an overall prevalence of 3%. T. congolense 6 (2%) and T. vivax 1 (0.3%) with 2 (0.7%) suspected brucei. The parasite prevalence Trypanosoma was 4 (4.6%) in poor body condition (Bcs) cattle. The animals in the age range 5 - 9 years were infected high with 7 (5.3%) prevalence.
Conclusion: The current study revealed that there are high-risk factors that predispose the community to Human African Trypanosomiasis (HAT) due to the presence of two different species of Tsetse flies and many animal reservoirs. The transmissions of Human African trypanosomiasis (HAT) are related to environmental, Vector, and human factors. Further geographically expanded investigation should be conducted throughout the country.
Keywords:
Trypanosomiasis
; Tsetse fly
; Trypanosoma brucei rhodesiense
; Human
; Animal
; Ethiopia
Background
Sleeping sickness, so called Human African Trypanosomiasis (HAT) is a parasitic vector-borne disease caused by the genus Trypanosoma (T.) and species brucei [1]. The disease is transmitted to humans by the bite of an infected both female and male tsetse fly (Glossina species) [2], hematophagous and, capable of transmitting infection [3]. T. b. rhodesiense is one of the HAT subspecies found in eastern and southern Africa, causing an acute and rapidly progressive disease [4]. Rhodesiense is another subspecies of HAT that has epidemic potential in humans, as it has been responsible for large outbreaks in the past decade [5]. The Trypanosoma genus are also pathogenic to animals (both in wild and domestic) and cause African trypanosomiasis (AAT), called Nagana [6]. The spread out of parasites from wildlife to domestic livestock and humans, and from domestic animal species to wildlife, compromises health [7]. Most trypanosome infections in wildlife do not cause apparent harm to their hosts, but some wildlife species are extremely at risk to trypanosome infections [8]. T. b. rhodesiense, can be found in both domestic and wild animals, contract T. b. gambiense and most likely serve as reservoirs [9]. However, it is still unclear how exactly the animal reservoir contributes to the epidemiology of the disease of the gambiense type. T. brucei, T. congolense, T. simiae, and T. uniforme are transmitted within the tsetse belts of Africa. Within and outside tsetse fly-infested zones, T. vivax and T. evansi can be transmitted mechanically[10]. Sleeping sickness threatens millions of people in 36 sub-Saharan Africa living in remote rural areas with limited access to adequate health services [11]. This complicates the surveillance and, therefore, the diagnosis and treatment of cases is neglected. Additionally, factors like population displacement, war, and poverty are facilitating the transmission. HAT, without distinction between T. b. gambiense and T. b. rhodesiense, was considered in the first Global Burden of Disease assessments and estimated to result in 1.78 million DALYs lost across Africa and the good reason for numerous of the parameters pertaining to HAT and used in the global assessments (e.g., disability weighting, and estimates of incidence) is not transparent and is a major problem. [12].
Currently, in Africa, animal Trypanosoma species such as T. congolense (found in tsetse-infested areas), T. viva x (found in both tsetse-infested and tsetse-free areas except in the highlands >2500 m above sea level), and T. brucei are commonly known. T. evansi (surra) and T. equiperdum can occur in tsetse-free areas [13]. The transmission of Trypanosoma vivax is cyclical (transmitted by the tsetse fly) and mechanical (by tabanidae and stomoxys). T. evansi in camels and T. equiperdum in horses are transmitted by the tsetse fly, Stomoxys and Tabanus, but mechanical transmission to domestic animals [14].
Cattle movement is a threat for the disease transmission, spread, and occurrences of outbreaks [14]. Wildlife hosts: bushbucks (Tragelaphusscriptus), impalas (Aepycerosmelampus), lions (Pantheraleo), zebras (Equus quagga boehma), warthogs (Phacocoerusafricanus), and duikers (Sylvicapragrimmia) carry the human-infective zoonotic trypanosome strain T. b. rhodesiense [13]. People can be bitten by an infective tsetse fly while farming, fishing, hunting, collecting water or wood, or engaging in any other activity. More focus is needed on Tsetse fly or vector control (bush clearing, game animal elimination, and ground and aerial spray) [3]. Insecticide Treated Cattle (1% Deltamethrin pour on Animal), deltamethrin 1% pour-on on domestic animals to prevent the animals from insect bites and to act like mobile targets [15]. Blue -Black-Blue target impregnated with insecticides and deployed in tsetse-infested areas by using deltamethrin 20% impregnated targets and ground spray have been used [16]. A ground spray in areas where targets are difficult to deploy [17] and SIT is a very specific method to the target species, non-pollutant, more effective, and has no adverse effect on other living organisms [18]. Barrier Establishment avoid the dangers of re-invasion, effort should be made to close the possible reinvasion site with artificial barriers (using traps and stationary targets) [19].
AAT is also a major constraint to the development of the livestock sector, and induces a decrease of livestock productivity. Tsetse flies are the main vectors of both HAT and AAT, a major factor in trypanosomiasis epidemiology by their central role in transmission of trypanosoma to vertebrate hosts [20]. In Ethiopia, animal trypanosomiasis is a significant obstruction to livestock and farm production [21] with approximately 75% of the area favorable for the proliferation of tsetse flies in the southern nation's nationalities and people's region states (SNNPR). And also Trypanosomiasis is prevalent in two main regions of Ethiopia: the northwest and the southwest [8]. Most of the areas in Gamo and Gofa zones in the region are suitable (with their savannah-covered national parks, river basins, ambient temperature, and bushy land nature) for tsetse multiplication [22]. In 1967, a confirmed HAT case was reported for the first in Ethiopia. The distribution of endemic human trypanosomiasis and its tsetse vectors appears to be limited to southern and southwestern Ethiopia (Oromia Region (Illubabor and Wollega), Gambella, and SNNPR). No major outbreak of the disease was recorded until the 1969-1970 epidemics and then after, only sporadic cases have been reported [23].
Currently, in August 2022, the SNNP Regional Health Bureau has reported 5 cases of HAT from Kucha Alpha woreda, Gamo zone, to the ministry of health Ethiopia. Dried blood spot samples were collected and transported to the Institute of Tropical Medicine, Antwerp, Belgium, through DHL and confirmed to be T. b. rhodesiense. HAT reoccurrence is linked to absence of surveillance system, the presence of favorable ecology for vector distribution and disease transmission, the presence of agricultural activities in proximity to vector habitat and the presence of reservoir wild animals in proximity with the human residence, the absence of road accessibility for controlling activities and lack of collaboration (one health). The epidemiology of HAT in Ethiopia remains unclear and will be essential to clarify in order to develop more effective and accessible diagnostic, treatment and control programs. Analyzing HAT data by place of infection on a longitudinal basis would provide valuable data on local risk and transmission characteristics and indicate whether control programs were effective. Without an accurate picture of the true epidemiology of HAT in Ethiopia, associations with environmental, vector, animal, and human factors cannot be analyzed and used to inform policy [24].
Identification of trypanosomes in tsetse flies and reservoir hosts could be a good indicator for HAT and AAT within an area [25]. Detection of pathogenic trypanosomes in animals and identification of tsetse flies and their distribution status within different environmental variables will highlight the areas at risk [20, 26]. Such type of information will help in the use of adapted control methods in this increasingly anthropized habitat within vector human activity overlap. Thus, the current investigation was initiated to identify the root cause of HAT reemergence in Ethiopia after 55 years.
Material and Methods
Study periods and Setting
The investigation was conducted in Kucha alpha (Gamo zone) and Dembagofa (Gofa zone) in SNNPR of Ethiopia from August – November 2022 following HAT outbreak (case) report. SNNPR is the most diverse region in the country in terms of language, culture, and ethnic background [27]. The Kucha district is located 172 kilometers from Arba Minch town of GamoGofa Zone and located 440 kms away from Addis Ababa, Ethiopia [28]. The capital town is Selamber[29]. The DembaGofa district is located 510 km south of Addis Ababa that divided into 38 rural kebeles. The capital town of the woreda is Sawla. The study area has three agro-climatic zones: highland altitude/Dega/ (7.8%), mid-altitude/WoinaDega/(15.8%), and lowland altitude/Kola/(76.4%) [30] (Figure 1).
A cross sectional study design was employed taking into consideration: all the historical and current information regarding the HAT case report, tsetse distribution history, environmental variables, and animal case surveillance variables. The entomological investigation was conducted in purposively and case-based selected sites.
Tsetse Fly sampling
Tsetse flies were caught using bi-conical, small sticky targets, and NGU traps in and around the HAT case report areas. A total of 30 traps (15 NGU and 15 bi-conical) were installed at roughly 4/km2 for two days in each grid depending on vegetation type, drainage system, accessibility, and human activities (settlements or cultivated land). All traps were baited with cow urine dispensed at 100 mg/hr from plastic bottles and acetone, dispensed at 500 mg/h from glass bottles and greased to prevent ants from damaging the catch [14]. Traps were checked after 12 hours, and the flies were removed from the trap. Tsetse flies were identified using the tsetse control personnel training manual key for Glossina species identification, through the analysis of the fly’s morphological traits [31]. Tabanids and Stomoxis were identified at the genus level using the morphological keys [32]. Tsetse flies were then preserved in the vials using ethanol for further molecular detection. All the bottles containing trapped flies were labeled with location, trap number, date, and time. The GPS coordinates of each trap were recorded using GARMIN (72H). Collected anesthetized flies or freshly dead tsetse flies were counted per species and sex and then dissected in the refrigerator with the objective of searching for the possible presence of trypanosomes. Dissection was carried out under a binocular stereo-microscope (magnification of 60X) using saline solution. All dissected organs of the tsetse were collected individually in Eppendorf® microtubes containing 50μL of sterile distilled water and kept on the field at about 8°C and then at the laboratory, at −20° C until DNA extraction. Additionally, un-dissected Tsetse flies were preserved for further parasite analysis with PCR.
Animal Sample Collection and parasite detection
A total of 301 cattle animals were randomly selected in local breeders based on the differences in body conditions and colors were sampled to collect blood samples. Cattle are the most common domestic animals in the study areas and are mainly bred in the periphery of villages or along rivers crossing the villages and forest, where tsetse flies are often abundant.
About 10μl of blood was collected from the marginal ear vein using the heparinized microhematocrit capillary [33]. All the blood samples were examined using the Buffy coat method (BCM). After screwing the rotary cover and closing the centrifuge lid, the specimens were allowed to centrifuge at 12,000 rpm for 5 min [34]. After the centrifugation, tubes were placed in the hematocrit reader and the readings were expressed as a percentage of packed red blood cells to the total volume of whole blood. Animals with PCV < 24% were considered to be anemic and animals with >/=25 were considered non-anemic. Finally, the slide was examined under ×40 objectives and ×10 eyepieces for the movement of parasites [35].
Method of statistical analysis
All the data was entered and cleaned in Microsoft Excel 2019 and exported to SPSS version 23. A descriptive statistical analysis was applied to summarize the data by sex and abdominal condition, animal, and laboratory results. The site, trap type, tsetse species, vegetation type, and collection were the independent variables. The ecological variation distribution of the vectors and its species were analyzed with descriptive statistics. Tsetse catches were used as a dependent variable. The density of tsetse fly population was calculated by dividing the number of flies trapped by the number of traps deployed and number of days of deployment expressed as Fly/Trap/Day [34]. Cross tabulations, Chi-square test, and Logistic regression were applied. The total prevalence rate or infection rate (IR) was calculated by dividing the number of positives by the total number of animals examined and multiplied by 100. The association between the trypanosome infection and associated risk factors were assessed by logistic regression. Chi-square test was employed to determine prevalence with sex, age and body condition scores, whereas two sample T-tests were used to compare the Mean PCV values between trypanosoma positive and negative animals [36]. In all analyses, the confidence interval level was 95% and P value P<0.05 was considered significant.
Ethics statement
Blood sampling on cattle was done following the framework of epidemiological surveillance activities of world organization for Animal Health. No ethical statement is required by local authorities. Blood sampling was carried out by veterinarian on domestic animals, with the authorization of the owner, as it is performed during prophylaxis or diagnostic campaigns in the response of Human African Trypanosomiasis outbreak. No samples other than those for outbreak response screening and diagnostic procedures were collected. The objectives of the study were explained for breeders to give their consent for animal sampling. For animal care, venous sampling was performed by a veterinarian of the Ethiopian Public Health and Animal Health Institutes. A deworming treatment was provided free to all animals sampled and those positive with BCT were treated for trypanosomiasis.
Results
HAT case reports
A total of 5 confirmed cases and 4 deaths were reported with CFR of 80%. Among confirmed number (5) cases by age category: 5 – 14 years (2) and 15 -44 (2) were male and one (1) case was female (between 0 and 4 years) (Table 1). Most of the confirmed cases were above 5 years in age (Table 2).
The Attack Rate (AR) HAT in Kucha Alpha and Damba Gofa districts were 38 and 37 per 100,000 populations respectively (Table.2).
Fly composition and density
A total of 429 flies belonging to three genera: tsetse flies 329 (76.7%), Stomoxis 76 (17%) and Tabanids 27 (6.3%) were captured. Two species of Tsetse flies: G. pallidipes 259 (60.4%) and G. fuscipes 70 (16.3%) were identified from the outbreak reported areas with 129.5 and 35 AD respectively within a day (Table 3).
In areas of tsetse fly collection, 188 (51.1%) of tsetse flies were collected from Demba Gofa with 94 AD. Of these 153 (41.6%; AD = 76.5) of Tsetse flies were identified to be G. pallidipes. 28.8% (n = 106; AD = 53) of the same Tsetse fly species G. pallidipes were captured from Kucha Alpha district. G. fuscipes was also collected from both study sites (Figure 2).
Abdominal status of tsetse flies
In a total of collected Tsetse species, 89.4% (294) of tsetse fly were collected with Bi-conical traps whereas small 10.6% (35) tsetses were collected by NGU trap techniques. High population density (AD = 116; N = 232, 70.5 %) of G. pallidipes were observed with Bi-conical traps. Large numbers of unfed tsetse fly 283 (86.0%) were collected with 141.5 AD (Table 4).
Ecological distribution of Tsetse fly
The AD of female tsetse fly species was found to be high in each ecological variation: wood land (51), Bush land (20) and grassland (11). High male tsetse fly (G. pallidipes) AD was observed in Wood land the same as the female density. Generally, the AD (93) of tsetse fly was high in Wood land mostly by G. pallidipes(76.5) which was relatively followed by Bush land (36.5) by the same species, G. pallidipes (Figure 3).
Animal investigation result
301 cattles were examined to determine the presence of trypanosomiasis and the parasites were detected in 9 cattle with an overall prevalence of 3%. Two species of trypanosomes were identified with T. congolense 6 (2%) and T. vivax 1 (0.3%) with 2 (0.7%) suspected brucei. The risk prevalence of trypanosomosis was in poor body conditions (Bcs) cattle 4 (4.6%) specially at the age range of 5 - 9 yrs 7 (5.3%) (Table 5).
High prevalence of trypanosomiasis was detected in Demba Gofa (6.7%) in Black color cattle in comparison to Kucha Alpha (3.3%) from white color cattle (Table 6).
Discussion
Cases of HAT were confirmed by parasitological testing after active medical work-up and five cases were confirmed in these areas after 55 years of HAT occurrence in 1967 in Ethiopia. Among all five cases four deaths were recorded. The attack rate (AR) and case fatality rate (CFR) of the human African trypanosomiasis were between 37 and 38 and 80% respectively in the study population, with the majority of patients being male (15 – 44 years old). In Africa, approximately, 70 million people (1.55 million km2) are estimated to be at risk of HAT and this corresponds to 10% of the total population and 7.4% of the total area of the endemic countries [37]. It was reported that HAT infected children rate is generally less than that observed in adults because of their lower exposure to flies during daily activities except those children who often accompany their parents during any activities [38]. As in outbreak areas of the Southwestern Ethiopia, sleeping sickness in Kucha Alpha and Demba Gofa is a rural disease mainly affecting the working population and productivity (farmer). Almost all patients in our study areas were from the native population that is self-employed (from farming households) and lives in villages with no immigrant population. The study reports from the Republic of Congo confirmed that uneducated participants as well as those aged more than 15 years were significantly at high risk of HAT infection compared to their corresponding person [38]. The majority of the positive cases were from households working in farming (animal and agricultural farming) [39]. All age groups and both sexes are at risk, the prevalence is higher in adults and the sex distribution varies in relation to gender-specific at-risk activities (e.g., mainly male hunting and fishing or principally female water fetching and small crop growing) [40].
This finding could provide a useful diagnostic sign for screening populations living in these Mazie forest regions and as a source for parasitological diagnosis and establishing better control programs. So the main objective of the current study was to screen the population at risk and risk factors and provide information on human African trypanosomiasis transmission in outbreak areas Kucha Alpha (Gamo Zone) and Demba Gofa (Gofa Zone) of Ethiopia. The findings of this study emphasize the call for not only to go on surveillance in these regions but also to expand surveillance throughout the country as a means of avoiding reemergence and spread of the disease.
In the current study areas, high-risk factors that predispose the community to Human African Trypanosomiasis (HAT) were identified in the HAT outbreak investigation areas. Cropping is considered to play a major role in maintaining HAT reemergence. This risk zone is in association with the Maze National Park and the Omo and Gibe River. There was a high movement of cattle and people to the Omo River for castling and fetching water, where the favorable environment for the breeding of tsetse flies vectors with high density. At the areas limit of trypanosomiasis distribution, nature reserves shape the patterns of HAT risk [37], most notably in around near the developmental corridor, cattle farming and agricultural intensification within the natural parks and Forest of Omo River and gibe catchment in agreement with the Geo-referenced layers of sleeping sickness occurrence and human population for the period 2000–2009 [37].
In this study, entomological investigation was conducted to improve our knowledge on the distribution of trypanosomes responsible for HAT and AAT. The tsetse flies were caught and dissected along the outbreak areas revealed that two species of Glossina (G. pallidipes and G. fuscipes) were found to be the predominant species. The presence of these two different species of Tsetse flies around where HAT cases were reported, areas implied that the disease is infecting the communities from animal reservoirs. It is likely that the identified Tsetse fly species are feeding on both small and large wild and domestic animals to facilitate the transmission of the HAT [41]. The findings in the present study corroborate entomological and epizootic information from the national atlas of tsetse flies and animal trypanosomosis in Ethiopia. In particular, our study confirms that in the study zone two species of tsetse flies occur (i.e. G. pallidipes and G. fuscipes), and that T. vivax and T. congolense are the dominant species of trypanosomes circulating in cattle [42]. The overall apparent density of tsetse fly was found to be very much high (AD = 164.5) per trap per day which is in contrast to the study reported that less AD of tsetse fly was recorded in Benishangul Gumuz in Ethiopia [43]. As in the AD of species comparison of previous study, G. pallidipes abundance in the current study area is higher than the previous study in Jima by Lemuet al. [34]. This higher abundance of tsetse species might be linked to suitable vegetation and available host density [44] and differences in agro ecology [34] and bioclimatic conditions during the trapping periods. The pathogenic trypanosomes identification in tsetse flies has helped to highlight human and animal trypanosomiasis associated with different biotopes (ecology) potential risks [26]. In the current study findings, the densities of the tsetse flies were also high in wood land, relatively less in bush land and grassland. This may be due to a combination of factors including favorable climatic conditions for the survival and dispersal ability of this species and abundant vegetation attracting hosts [20]. The current finding is in agreement with the study report from BenishangulGumuz region of Ethiopia by Mekonen et al. [43]who stated that the vegetation is vital for providing suitable conditions; the savanna, forest and riverine tsetse flies concentrate in the wooden savanna, in the bush vegetation and near the edge of the river, where the vegetation is dense.
High numbers of flies were observed unfed in the current study areas. This may impact the parasite density within the tsetse fly population and trypanosomiasis transmission. This may be an important factor in natural situations where densities of hosts commonly fed on by tsetse, can vary between periods [45]. Host population movement can determine the feeding status of the vectors. Moreover, as one moves from protected areas into farmland, host densities may drop below those required for sustaining tsetse populations but increasing host densities in others areas [46]. Similarly, wild and domestic host’s densities are essential factors for trypanosome transmission, affecting vector density, and mortality rates. The existence of hungrier flies in regions where wild hosts are scarce can have implications for trypanosome transmission. First hungrier flies may be more likely to bite less preferential hosts, including humans, thereby ever-increasing the trypanosomiasis transmission risk [47].
None of dissected tsetse fly species were positive for HAT parasite. The absence of infected salivary glands in the present study confirms that the natural infection rate of tsetse by trypanosomes of brucei complex remains very low in agreement with study report by Vincent et al. [20] in Cote D'ivoire. The absence of infected tsetse flies in HAT case reported areas could not be guaranteed for the absences of future infection in the area, similarly, zero prevalence most probably would not mean any parasite infected tsetse flies were present in the area [45,48].
The overall prevalence of Trypanosomiasis in animals was 3% (T. congolense 6 (2%) and T. vivax 1 (0.3%)) with 2 (0.7%) suspected brucei in the investigated areas. The presence of T. congolense and T. vivax is an indicator of AAT transmission magnitude near the Maze National Park in agreement to other studies [49]. The prevalence of bovine trypanosomosis in the present study areas was significantly lower than the previous study reports [43]. This suggests that livestock expansion will be seriously hampered by the Nagana if control measures against tsetse flies and trypanosomes are not taken prior to cattle introduction along with this river and park [43]. This study result showed that risk prevalence of trypanosomosis was high in poor body conditions (Bcs) cattle compared the other body conditions )medium and good) which indicates that infection rates in poor body condition animals were significantly higher than that of medium and good body condition animals. This is due to poor body condition, animals are at risk of the infectious disease, reduced animal presentation created by lack of essential nutrients and poor management [34].
There are several weaknesses in the current study design, which need to be considered when interpreting the findings of the current study report. Moreover, the current study has not covered large geographical areas within short data collection periods to diversify the species and get positive tsetse flies. As this study was employed to use only three collection techniques, it would not have been appropriate to collect a high number of blood-fed mosquitoes for blood meal origin analysis and seasonal variation data was not included.
Conclusion
The current study revealed that there are high-risk factors that predispose the community to Human African Trypanosomiasis (HAT) with the presence of highly abundant Tsetse flies species and animal reservoirs. The transmissions of Human African trypanosomiasis (HAT) are related to environmental, Vector, and human factors. Proper identification of vectors is important in the surveillance of arthropod-borne diseases. Further geographically expanded investigation on the transmission patterns from these fly vectors to livestock and humans will be important to understanding the epidemiology and management of AAT throughout the country. All concerned bodies should work on the minimization of its impact. .Communities from most affected areas in the design and implementation of locally appropriate interventions for HAT will be crucial to the success of future interventions, and to reducing the local burden of this important neglected disease. Designing appropriate risk reduction strategies in partnership with community stakeholders would facilitate efficient and effective program design.
Author Contributions
AW: Conceptualization, Methodology, Resource, Investigation, Visualization, Validation, Data curation, Formal analysis, wrote the main manuscript (Writing-original draft, and Writing-review and editing). EA: AG, HT, BM, MH, HD, AE, AK, GG, TW, NM, TH, KK, NA, JH, WM, DT, AA, BB, FL, TM, FT, KA, TM, YM, MW and FR: Conceptualization, Methodology, Visualization, Validation, Formal analysis and Writing-review and editing, Methodology, Visualization, Validation, Formal analysis, Writing-original draft and Writing-review and editing. GT and AA: Supervision, Visualization, Validation, Formal analysis, Writing-review, and editing. All authors have read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research is part of routine government work as an outbreak investigation has received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the required analyzed and used in this research paper are within the manuscript and the supporting information not present within the manuscript will be available at corresponding authors and provided upon reasonable request.
Acknowledgements
We are very grateful to South Nation, Nationalities of Peoples of regional bureau and Health department office leaders, the village head, Health extension workers, and volunteers for their cooperation during data collection. We are so proud to express our deep gratitude to the data collectors for their strong commitment to collecting required data.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Mitashi P, Hasker E, Lejon V, Kande V, Muyembe JJ, Lutumba P, et al. Human African Trypanosomiasis Diagnosis in First-Line Health Services of Endemic Countries, a Systematic Review. PLoS Negl Trop Dis. 2012;6(11).
- Simo G, Mbida JAM, Eyenga VEO, Asonganyi T, Njiokou F, Grébaut P. Challenges towards the elimination of Human African Trypanosomiasis in the sleeping sickness focus of Campo in southern Cameroon. Parasites and Vectors. 2014;7(1):1–7.
- Malvy D, Chappuis F. Sleeping sickness. Clin Microbiol Infect. 2011;17(7):986–95. [CrossRef]
- Adungo F, Mokaya T, Makwaga O, Mwau M. Tsetse distribution, trypanosome infection rates, and small-holder livestock producers’ capacity enhancement for sustainable tsetse and trypanosomiasis control in Busia, Kenya. Trop Med Health. 2020;48(1).
- Franco JR, Cecchi G, Priotto G, Paone M, Diarra A, Grout L, et al. Monitoring the elimination of human African trypanosomiasis: Update to 2016. PLoS Negl Trop Dis. 2018;12(12):1–16.
- Odera JO. Population genetic studies of the tsetse fly, Glossina pallidipes (Diptera: Glossinidae). ProQuest Diss Theses. 2004;152. http://ezproxy.nottingham.ac.uk/login?url=https://search.proquest.com/docview/305169097?accountid=8018%5Cnhttp://sfx.nottingham.ac.uk/sfx_local/?url_ver=Z39.88-2004&rft_val_fmt=info:ofi/fmt:kev:mtx:dissertation&genre=dissertations+%26+theses&sid=ProQ:ProQ.
- Seck MT, Sall B, Ndiaye EY, Guerrini L, Vreysen MJB, Bouyer, J Seck, MT Sall, B Ndiaye, EY Guerrini, L Vreysen M. Stratified entomological sampling in preparation for an area-wide integrated pest management program : the example of Glossinapalpalisgambiensis ( Diptera : Glossinidae ) in the Niayes of. J Med Entomol. 2010;47(4):543–52.
- Geremew B, Zelalem A, Aster T, Ayantu H. Survey of Apparent Density of Tsetse and other Biting flies in Gimbi district, West Wollega, Western Ethiopia. SOJ Vet Sci. 2018.
- Longbottom J, Wamboga C, Bessell PR, Torr SJ, Stanton MC. Optimising passive surveillance of a neglected tropical disease in the era of elimination: A modelling study. PLoS Negl Trop Dis. 2021;15(3).
- World Health Organization. Control and surveillance of human African trypanosomiasis. World Health Organ Tech Rep Ser. 2013;(984):1–237.
- Serap A. Control of tsetse flies and trypanosomes using molecular genetics. Vet Parasitol. 2003;115(2003):125–45.
- Nagagi YP, Silayo RS, Kweka EJ. Advancements in bait technology to control Glossina swynnertoni Austen, the species of limited distribution in Kenya and Tanzania border : A review Advancements in bait technology to control Glossina swynnertoni Austen, the species of limited distributi. J Vector Borne Dis. 2017;54:16–24.
- Shereni W, Anderson NE, Nyakupinda L, Cecchi G. Spatial distribution and trypanosome infection of tsetse flies in the sleeping sickness focus of Zimbabwe in Hurungwe District. Parasit Vectors. 2016;1–9. [CrossRef]
- Solano, P. Need of entomological criteria to assess zero transmission of gambiense hat. PLoS Negl Trop Dis. 2021;15(3):1–8. [CrossRef]
- Pan African TsetseEradication Campaign Trypanosomosis (PATTEC). Pan African Tsetse and Trypanosomosis Eradication Campaign.
- Mihok S, Carlson DA, Krafsur ES, Foil LD, Walker C, Macquart H. Performance of the Nzi and other traps for biting flies in North America. Bull Entomol Res. 2006;96(2006):387–97.
- LSTM LS of TM. Use of Tiny Targets to control tsetse flies in Gambian HAT foci : standard operating procedures. 2016;(October):1–32.
- Vreysen MJB. Prospects for area-wide integrated control of tsetse flies ( Diptera : Glossinidae ) and trypanosomosis in sub-Saharan Africa. Rev Soc Entomol Argent 65. 2006;65(1–2):1–21.
- Vale GA, Hargrove JW, Lehane MJ, Solano P, Torr SJ. Optimal Strategies for Controlling Riverine Tsetse Flies Using Targets : A Modelling Study. PLoS Negl Trop Dis. 2015;9(3):1–20. [CrossRef]
- Vincent D, Dramane K, Jean-baptiste R, Guiguigbaza-kossigan D, Bamoro C, Ernest S, et al. Detection and identification of pathogenic trypanosome species in tsetse flies along the Comoé River in Côte d ’ Ivoire. Parasite. 2015;22(18).
- Asmamaw Aki, Getachew D. Trypanosomosis in Cattle Population of Pawe District of Benishangul Gumuz Regional State, Western Ethiopia: Anemia, Vector Density and Associated Risks. Researcher. 2016;8(4):79–85.
- Zekarias T, Kapitano B, Mekonnen S, Zeleke G. The Dynamics of TseTse Fly in and Around Intensive Suppression Area of Southern Tsetse Eradication Project Site, Ethiopia. Ethiop J Agric science. 2014;24(2):59–67.
- Mesfin SM. Full Length Research Paper A cross-sectional study on the apparent density of tsetse flies and prevalence of bovine Trypanosomosis. Glob J Med Surg. 2016;4(10):151–5.
- Reid H, Kibona S, Rodney A, McPherson B, Sindato C, Malele I, et al. Assessment of the burden of human African trypanosomiasis by rapid participatory appraisal in three high-risk villages in Urambo District, northwest Tanzania. Afr Health Sci. 2012;12(2):104–13.
- Boundenga L, Mombo IM, Augustin MO, Barthélémy N, Nzassi PM, Moukodoum ND, et al. Molecular Identification of Trypanosome Diversity in Domestic Animals Reveals the Presence of Trypanosoma brucei gambiense in Historical Foci of Human African Trypanosomiasis in Gabon. Pathogens. 2022;11(9).
- Bemba I, Lenga A, Awono-Ambene HP, Antonio-Nkondjio C. Tsetse Flies Infected with Trypanosomes in Three Active Human African Trypanosomiasis Foci of the Republic of Congo. Pathogens. 2022;11(11):1275.
- A G, B Z, H K, A J. Characterization of Gofa Cattle Population, Production System, Production and Reproduction Performance in Southern Ethiopia. J Fish Livest Prod. 2017;05(03).
- Mutiso SM, Qureshi Z, Kinuthia J. Birth Preparedness Among Antenatal Clients in Rurar Health Centers in Kucha Woreda, Gamo-Gofa Zone, Southern Ethiopia. East Afr Med J. 2008;85(6):275–83.
- Senbeta Bedane A, Tanto TK, Asena TF. Malaria Distribution in Kucha District of Gamo Gofa Zone, Ethiopia: A Time Series Approach. Am J Theor Appl Stat. 2016;5(2):70.
- Zekarias B, Mesfin F, Mengiste B, Tesfaye A, Getacher L. Prevalence of goiter and associated factors among women of reproductive age group in Demba gofa woreda, Gamo Gofa zone, southwest Ethiopia: A community-based cross-sectional study. J Nutr Metab. 2020;2020.
- Rosemary B, Jingwen W, Yineng W, L WB, C WW, A MG, et al. Tsetse fly ( Glossina pallidipes ) midgut responses to Trypanosoma brucei challenge. Parasite and vectors. 2017;10(615):1–12.
- Ngomtcho SCH, Weber JS, Ngo Bum E, Gbem TT, Kelm S, Achukwi MD. Molecular screening of tsetse flies and cattle reveal different Trypanosoma species including T. grayi and T. theileri in northern Cameroon. Parasites and Vectors. 2017;10(1):1–16.
- Adugna T, Lamessa A, Hailu S, Habtamu T, Kebede B. A Cross-Sectional Study on the Prevalence of Bovine Trypanosomosis in Ankesha District of Awi Zone, Northwest Ethiopia. Austin J Vet Sci Anim Husb. 2017;4(2):1–4.
- Lemu M, Bekuma F, Abera D, Meharenet B. Prevalence of Bovine Trypanosomosis and Apparent Density of Tsetse Fly in Botor Tolay District, Jimma. Biomed J Sci Tech Res. 2019;13(3):9976–83.
- Nakamura Y, Hayashida K, Delesalle V, Qiu Y, Omori R, Simuunza M, et al. Genetic Diversity of African Trypanosomes in Tsetse Flies and Cattle From the Kafue Ecosystem. Front Vet Sci. 2021;8(January).
- Efrem D, Hagos A, Getachew T, Tesfu K, Nigatu K, Workineh S, et al. A Cross-sectional Study of Bovine Trypanosomosis in Sayo District, Oromia Regional State, Western Ethiopia. Int J Nutr Food Sci. 2018;7(2):56–64.
- Franco R, Paone M, Diarra A, Ruiz-postigo A, Fe EM. Estimating and Mapping the Population at Risk of Sleeping Sickness. 2012;6(10).
- Elenga VA, Lissom A, Ornelle D, Elion A, Vouvoungui JC, Djontu JC, et al. Risk factors and prevalence of human African trypanosomiasis in individuals living in remote areas of the republic of Congo. BMC Public Health. 2022;1–9. [CrossRef]
- Uba BV, Aliyu A, Abubakar A, Uba SA, Gidado S, Edukugho A, et al. Knowledge and prevalence of human african trypanosomiasis among residents of kachia grazing reserve, Kachia local government area, Kaduna state, Nigeria, 2012. Pan Afr Med J. 2016;23:1–8.
- Büscher P, Cecchi G, Jamonneau V, Priotto G. Human African trypanosomiasis. Lancet. 2017;390(10110):2397–409.
- Crump RE, Huang CI, Knock ES, Spencer SEF, Brown PE, Miaka EM, et al. Quantifying epidemiological drivers of gambiense human African Trypanosomiasis across the Democratic Republic of Congo. PLoS Comput Biol. 2021;17(1):1–23. [CrossRef]
- Gebre T, Kapitano B, Beyene D, Alemu D, Beshir A, Worku Z, et al. The national atlas of tsetse flies and African animal trypanosomosis in Ethiopia. Parasites and Vectors. 2022;15(1):1–15. [CrossRef]
- Mekonnen G, Mekonnen AN. Vector identification and prevalence of bovine trypanosomosis in Oda Buldigilu district of Benishangul Gumuze regional state , Western Ethiopia Vector identification and prevalence of bovine trypanosomosis in Oda Buldigilu district of Benishangul Gumuze r. J Entomol Zool Stud. 2017;5(5):1178-1183 E-ISSN:.
- Shumago N, Tekalign W. Distribution of Tsetse Fly in Selected Sites of Upper Omo Belt, Southern Ethiopia. Adv Life Sci Technol. 2016;44(1976):30–7.
- Opiro R, Opoke R, Angwech H, Nakafu E, Oloya FA, Openy G, et al. Apparent density, trypanosome infection rates and host preference of tsetse flies in the sleeping sickness endemic focus of northwestern Uganda. BMC Vet Res. 2021;17(1):1–12. [CrossRef]
- Aksoy E, Telleria EL, Echodu R, Wu Y, Okedi LM, Weiss BL, et al. Analysis of multiple tsetse fly populations in Uganda reveals limited diversity and species-specific gut microbiota. Appl Environ Microbiol. 2014;80(14):4301–12.
- Lord JS, Mthombothi Z, Lagat VK, Atuhaire F, Hargrove JW. Host-seeking efficiency can explain population dynamics of the tsetse fly Glossina morsitans morsitans in response to host density decline. PLoS Negl Trop Dis. 2017;11(7).
- Mekonnen G, Mekonnen AN, Golessa M, Asrese NM. Vector identification and prevalence of bovine trypanosomosis in Oda Buldigilu district of Benishangul Gumuze regional state, Western Ethiopia Vector identification and prevalence of bovine trypanosomosis in Oda Buldigilu district of Benishangul Gumuze r. J Entomol Zool Stud. 2017;5(5):1178–83.
- Regasa T, Bedada M, Workine M. Study on Spatial Distribution of Tsetse Fly and Prevalence of Bovine Trypanosomosis and Other Risk Factors : Case Study in Bedele Woreda, Ilu Aba Bora Zone, South Western Ethiopia. Acta Parasitol Glob 6. 2018;6(3):174–81.
Figure 1.
Map of outbreak areas in Ethiopia, SNNP and HAT investigation sites (Kucha Alpha and Demba Gofa), 2022.
Figure 1.
Map of outbreak areas in Ethiopia, SNNP and HAT investigation sites (Kucha Alpha and Demba Gofa), 2022.
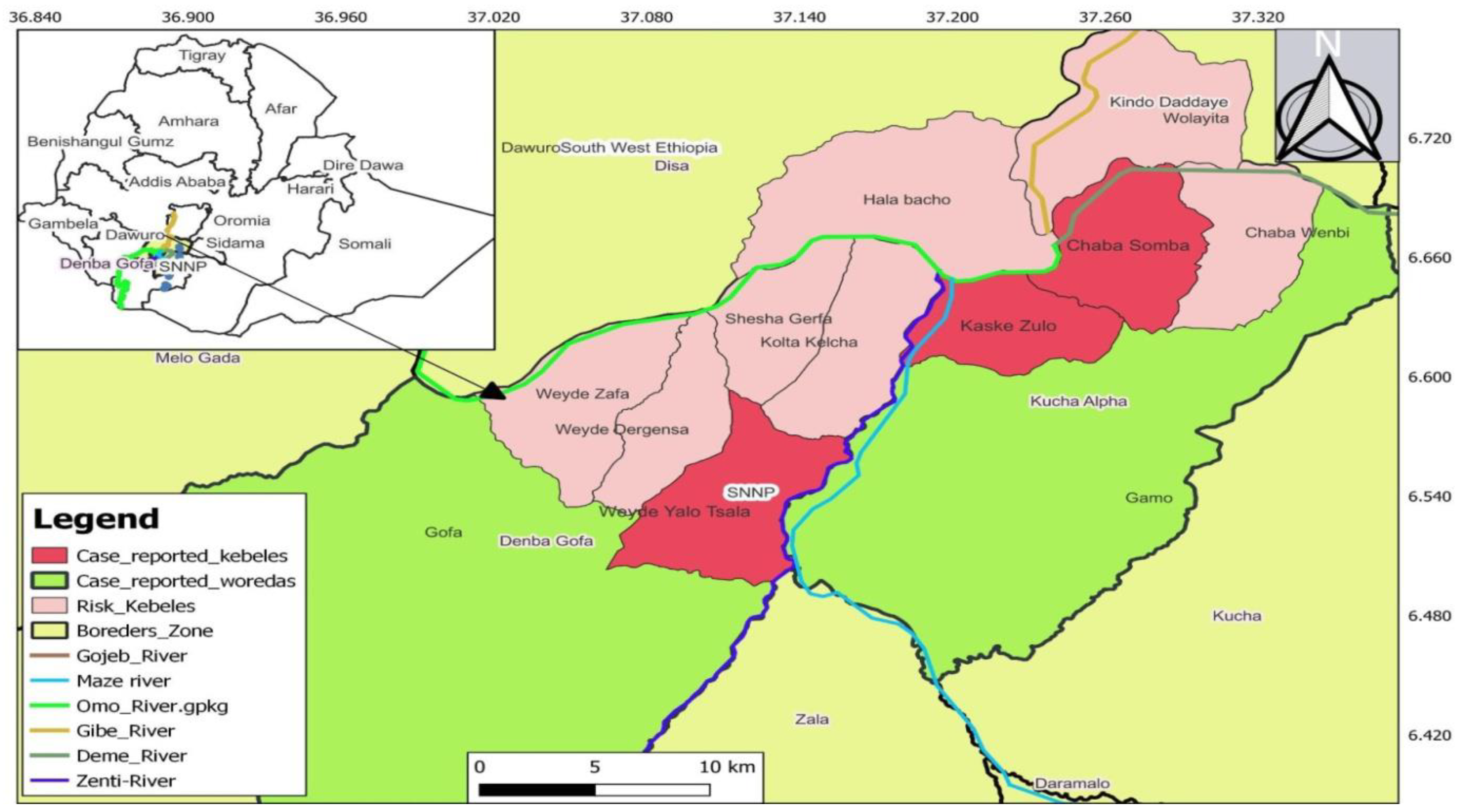
Figure 2.
Distribution of Tsetse fly species in fly sampling sites (Demba Gofa and Kucha Alpha), SNNPR, Ethiopia.
Figure 2.
Distribution of Tsetse fly species in fly sampling sites (Demba Gofa and Kucha Alpha), SNNPR, Ethiopia.
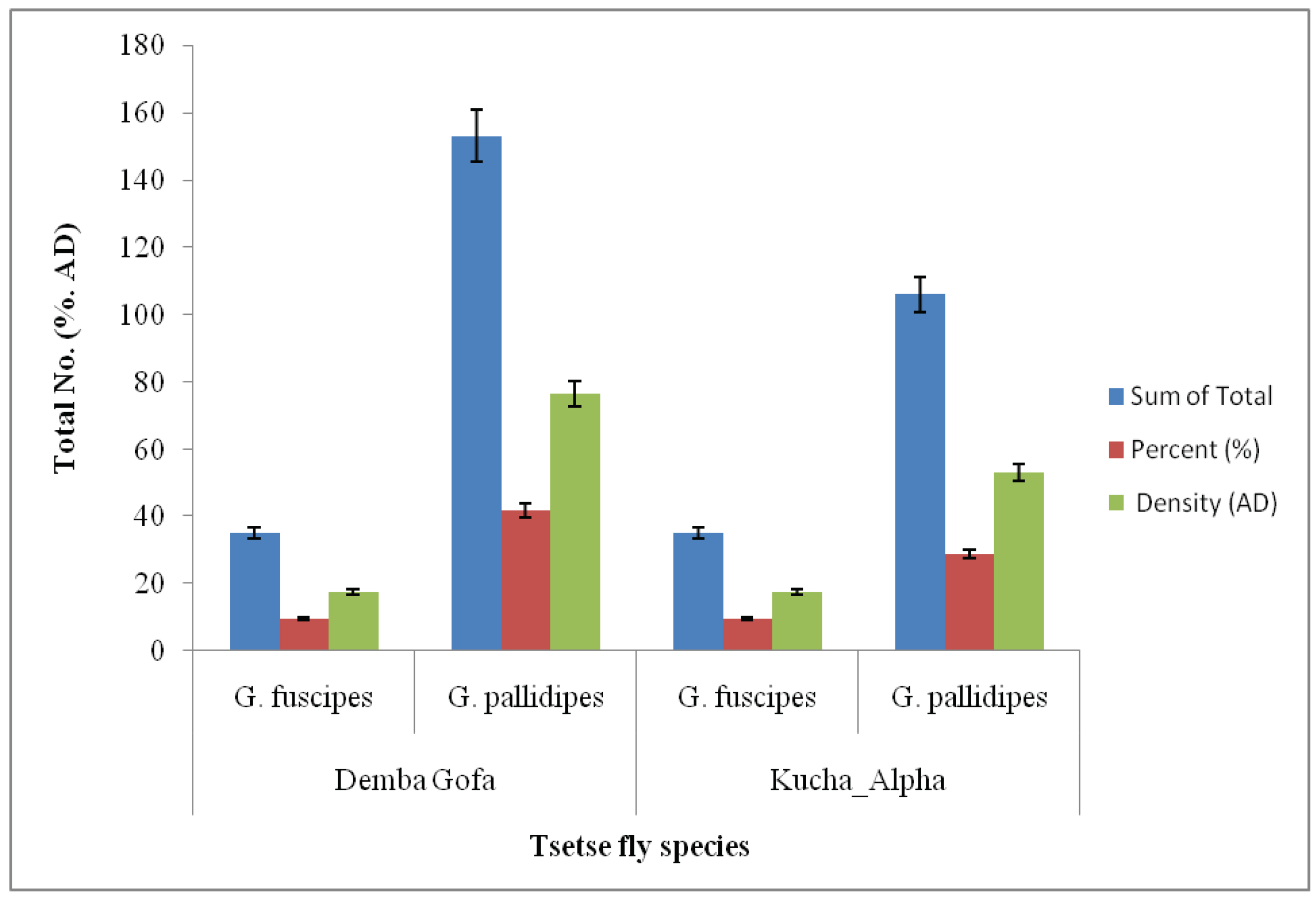
Figure 3.
The distribution of Tsetse fly species by sex and ecological variation, SNNPR, Ethiopia.
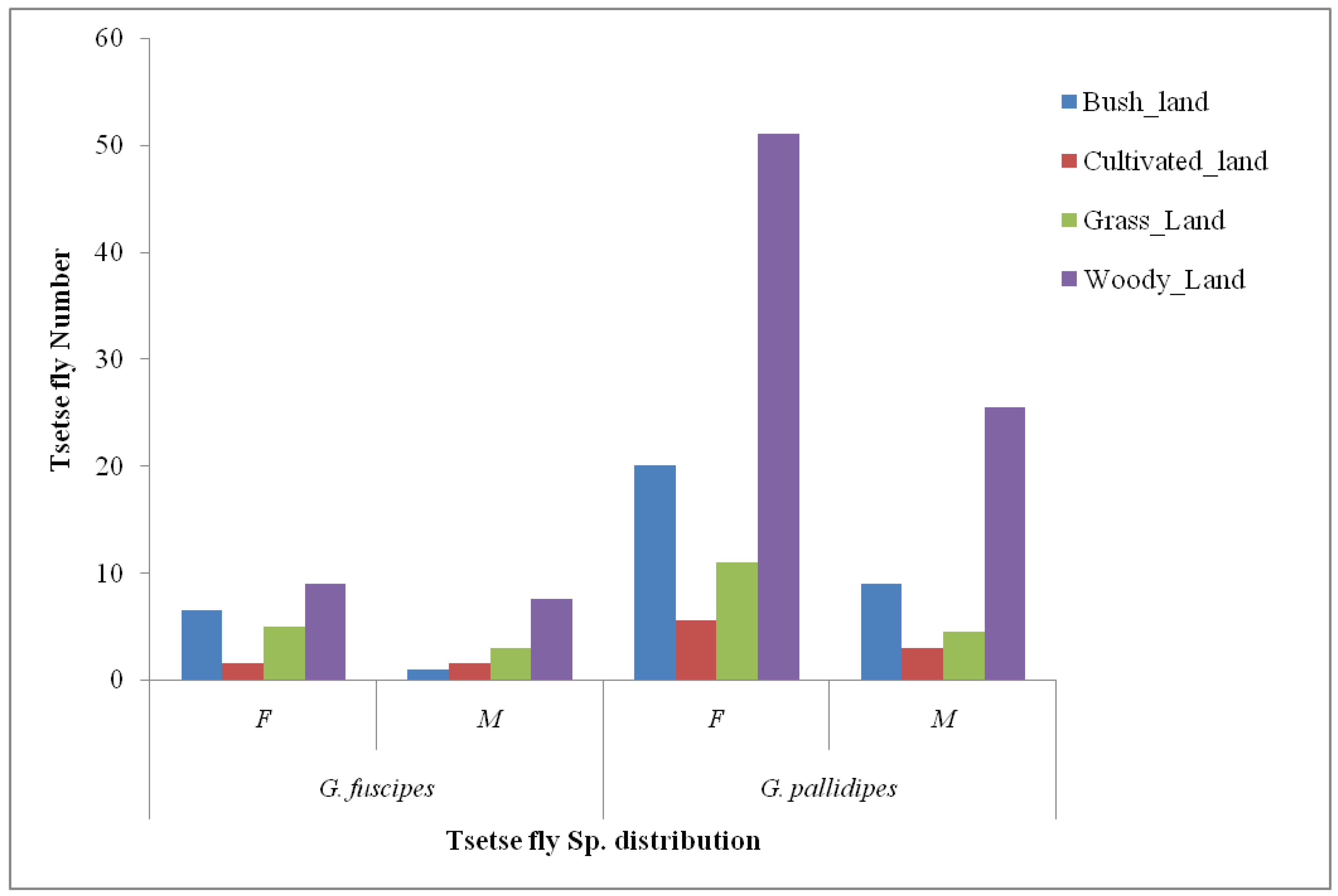

Table 1.
Age distributions of Confirmed cases in Kucha alpha Demba Gofa districts, SNNPR, November 2022.
Table 1.
Age distributions of Confirmed cases in Kucha alpha Demba Gofa districts, SNNPR, November 2022.
| Sex | Age-group | Number of confirmed Cases |
|---|---|---|
| Female | 0 - 4 years | 1 |
| Male | 5 - 14 years | 2 |
| Male | 15 - 44 years | 2 |
| Total | 5 |
Table 2.
HAT cases by geographical area with AR in Kucha alpha Demba Gofa Districts, SNNPR, November 2022.
Table 2.
HAT cases by geographical area with AR in Kucha alpha Demba Gofa Districts, SNNPR, November 2022.
| Zone | Districts | AR/100,000 pop |
|---|---|---|
| Gamo | Kucha Alpha | 38 |
| Gofa | DembaGofa | 37 |
Table 3.
Fly species collected and their apparent densities, Gamo and Gofa Zone, SNNPR, Ethiopia.
| Fly species | Number (%) | Apparent Density (AD) |
| G. Fuscipes | 70 (16.3) | 35 |
| G. pallidipes | 259 (60.4) | 129.5 |
| Total | 329 (76.7) | 164.5 |
| Stomoxys | 73 (17) | 34.5 |
| Tabanids | 27 (6.3) | 13.5 |
| Total | 100 (23.3) | 50 |
| Grand Total | 429 (100) | 214.5 |
Table 4.
Tsetse fly species abdominal status with the two tsetse fly trapping methods.
| Tsetse species | Fed (%, AD) | Unfed (%, AD) | Total (%, AD) | |
| Bi-conical | 39 (11.8, 19.5) | 255 (77.5, 127.5) | 294 (89.4, 147) | |
| G. Fuscipes | 7 (2.1, 3.5)) | 55 (16.7, 27.5) | 62 (18.8, 31) | |
| G. pallidipes | 32 (9.7, 16) | 200 (60.8, 100) | 232 (70.5, 116) | |
| NGU | 7 (2.1, 3.5) | 28 (8.5, 14) | 35 (10.6, 17.5) | |
| G. Fuscipes | 2 (0.6, 1) | 6 (1.8, 3) | 8 (2.4, 4) | |
| G. pallidipes | 5 (1.5, 2.5) | 22 (6.7, 11) | 27 (8.2, 13.5) | |
| Grand Total | 46 (14, 23) | 283 (86.0, 141.5) | 329 (100, 164.5) | |
Table 5.
Results showing association of potential factors for the prevalence of Trypanosomosis.
|
Factors |
Level |
Total tested |
-ve |
Positive (%) |
Total +ve |
||
| Brucei_suspected | T_congo | T_vivax | |||||
| Sex | Female | 139 | 134 | 1(0.7) | 4 (2.9) | 0 (0) | 5 (3.6) |
| Male | 162 | 158 | 1(0.6) | 2 (1.2) | 1(0.6) | 4 (2.5) | |
| Bcs | Good | 55 | 54 | 0 (0) | 1(1.8) | 0 (0) | 1(1.8) |
| Medium | 159 | 155 | 1 (0.6) | 2 (1.3) | 1(0.6) | 4 (2.5) | |
| Poor | 87 | 83 | 1((1.1) | 3 (3.4) | 0 (0) | 4 (4.6) | |
| Age | 1 - 4 yrs | 160 | 158 | 1 ((0.6) | 0 (0) | 1(0.6) | 2 (1.3) |
| 5 - 9 yrs | 132 | 125 | 1(0.8) | 6 (4.5) | 0 (0) | 7 (5.3) | |
| 10 - 15 yrs | 9 | 9 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | |
| Grand Total | 301 | 292 | 2 (0.7) | 6 (2) | 1(0.3) | 9 (3) | |
Table 6.
prevalence of Animal trypanosomiasis species in Damba Gofa and Kucha Alpha districts, SNNP, Ethiopia.
Table 6.
prevalence of Animal trypanosomiasis species in Damba Gofa and Kucha Alpha districts, SNNP, Ethiopia.
|
Animal color |
Total tested |
-ve |
Positive (+ve) |
Total +ve |
IR (%) |
|||
| Brucei | T_congo | T_vivax | ||||||
| Dambagofa | 96 | 94 | 0 | 2 | 0 | 2 | 2.1 | |
| Black | 15 | 14 | 0 | 1 | 0 | 1 | 6.7 | |
| Red | 44 | 43 | 0 | 1 | 0 | 1 | 2.3 | |
| White | 37 | 37 | 0 | 0 | 0 | 0 | 0.0 | |
| Kucha Alpha | 205 | 198 | 2 | 4 | 1 | 7 | 3.4 | |
| Black | 31 | 29 | 1 | 1 | 0 | 0 | 0.0 | |
| Red | 114 | 111 | 1 | 2 | 0 | 0 | 0.0 | |
| White | 60 | 58 | 0 | 1 | 1 | 2 | 3.3 | |
| Grand Total | 301 | 292 | 2 | 6 | 1 | 9 | 3.0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



