
Submitted:
10 February 2023
Posted:
13 February 2023
You are already at the latest version
Abstract
Methane (CH4) has attracted attention as not only synthetic natural gas, but also one of the hy-drogen carriers in terms of energy density. On the other hand, there exist bacterial ecosystems in nature that can decompose organic compounds to produce CH4 and CO2. In this study, Clostridi-um cellulovorans was first cultivated with pig manure (PM) as an unused biomass. Regarding the measurement of organic acids by high-performance liquid chromatography (HPLC), acetate and butyrate were increased in the C. cellulovorans medium containing 0.5% PM, while formate and lactate were decreased in it. Next, in comparison with carbon sources such as glucose, cellobiose, and acetate, cocultivation of C. cellulovorans and Methanosarcina mazei or microbial flora of me-thane production (MFMP) was performed in the C. cellulovorans medium. These results revealed that 0.5% acetate as the sole carbon source produced CH4 only by cocultivating C. cellulovorans and MFMP. Furthermore, MFMP was only cultivated with 1% acetate or 1% methane as a carbon source after precultivated with 0.5% glucose medium for 12 h. As a result, methane productivity of MFMP with 1% methanol medium was approximately eight times higher than that with 1% acetate medium. Finally, next-generation sequencing (NGS) analysis of MFMP after cultivation with 1% acetate or 1% methane was carried out. Interestingly, Methanofollis (0.211%) belonging to H2/CO2 -using methanogens (CO2 reduction pathway) was dominant in the 1% acetate medium for 72 h cultivation, whereas Methanosarcina siciliae (1.178%), M. barkeri (0.571%), and Methano-follis (0.490%) were major species in 1% methanol medium for 72 h cultivation. Since Methano-sarcina spp. are belonging to acetoclasts (acetoclastic pathway), methanol could promote to grow Methanosarcina spp. rather than acetate. Therefore, it seemed Methanosarcina spp. may play a key methanogenesis in MFMP. Thus, these results will provide important information for low cost biomethane production.
Keywords:
methanogenesis
; pig manure
; carbon sources
; C. cellulovorans
; methanogens
1. Introduction
Anaerobic digestion (AD) consists of a series of biochemical processes (i.e., hydrolysis, fermentation (acidogenesis), acetogenesis and methanogenesis) performed by various interacting microorganisms, including bacteria (i.e., acidogens, acetogens) and archaea (methanogens). It is also obvious that the cumulative CH4 production from the three different substrates varied significantly and was not in agreement with the expected according to the theoretical value calculated (Table 1) (formate 82.35 N mLCH4/gVS, acetate 273.17 N mLCH4/gVS, H2/CO2 414.81 N mLCH4/gVS) [1]. Since methanogenesis is the final step in anaerobic carbon transformation and is of critical concern in thawing permafrost peatland systems where CH4 release is increasing rapidly, prediction of the magnitude of carbon loss as CO2 or CH4 is hampered by our limited knowledge of microbial metabolism of organic matter in these environments [2]. Genome-centric metagenomic analysis of microbial communities provides the necessary information to examine how specific lineages transform organic matter during permafrost thaw [3]. The biomethanation process in nature relies on the microbial interactions between three main metabolic groups of anaerobes such as fermentative, acetogenic, and methanogenic microorganisms [4,5,6]. Whereas the first two groups decompose complex organic matters to acetate, H2 and CO2, which are the key precursors for methanogenesis, methanogens further convert these metabolites to CH4 by two major routes, i.e., acetoclastic pathway and CO2 reduction pathway [7]. On the other hand, although the growth behavior of a donor bacterium, Sulfurospirillum multivorans in the modified Methanococcus voltae (acceptor) medium with pyruvate alone as substrate was similar to that in the medium originally used for cultivation of S. multivorans, the morphology of S. multivorans cells was unaltered in the M. voltae medium and independent from the type of cultivation—fermentatively or respiratory [8]. In this case, the new medium with lactate as the sole growth substrate instead of formate and acetate could not promote growth for pure S. multivorans cultures. In the corresponding coculture, 15mM lactate was consumed in approximately 2 weeks while methane was produced, indicating lactate fermentation by S. multivorans and H2 transfer to M. voltae as syntrophic partner. Therefore, the coculture system seems to include system unique advantages, composition, products, and interaction mechanisms.
Elaboration of the underlying mechanism in microbial communities such as the exchange of intermediate metabolites, cell-to-cell electrical connections, communications, etc. would guide the design of artificial microbial consortia and further improve the robustness and stability of the cocultivation systems [9,10,11,12]. Therefore, these artificial microbial consortia interact mutually through the interaction of synergism, commensalism, competition, mutualism, and so on [12]. Diverse microbial communities within the same or different species have been set up to realize more complicated tasks [8,13,14]. In particular, the greatest advantage of coculture systems is that the combination of the metabolic capacity of two or more microorganisms allows for the utilization of more complex substrates and the production of specific products [14]. In addition to treatment of waste-water, biodegradation of textile azo dye and dispose of contaminated soil, recently, cocultivation systems were also applied to produce biofuels, bulk chemicals, and natural products [15,16,17,18,19,20,21,22,23,24,25,26].
Cellulose is most abundant on the Earth and not easily degraded and utilized. In addition to cellulosic sources, various other carbohydrates, carbon monoxide and syngas can also be processed using these systems [27]. The cellulolytic system of Clostridium cellulovorans mainly consists of a cellulosome which synergistically collaborates with non-complexed enzymes [28,29]. By the cocultivation of C. cellulovorans and C. beijerinckii, IBE fermentation was performed using mandarin orange wastes [30]. Moreover, methane was produced from sugar beet pulp [31] and mandarin orange peel [32] under cocultivation with C. cellulovorans and methanogens. Furthermore, two coculture models combining C. cellulovorans with Methanosarcina barkeri Fusaro or M. mazei Gö1 were established for the direct conversion of cellulose to CH4 [33]. Coculturing C. cellulovorans with M. barkeri or M. mazei not only enabled direct conversion of cellulose to CH4, but also stabilized pH for C. cellulovorans, resulting in a metabolic shift and enhanced cellulose degradation. The other approach was by implementing nanotechnology in combination with C. cellulovorans through consolidated bioprocessing (CBP) method to produce hydrogen from raw corn cob [34].
In this study, we observed the cocultivation of C. cellulovorans and M. mazei or microbial flora of methane production (MFMP) for the different carbon sources between sugars such as glucose and cellobiose that are the products from cellulose degraded by C. cellulovorans and acetate metabolized from glucose through TCA cycle. Furthermore, pig manure (PM) was used for the C. cellulovorans cultivation and was analyzed with organic acids. In addition, we investigated the cultivation manner of MFMP in comparison with acetate and methanol as the sole carbon source. Finally, 16S rRNA analysis in MFMP was performed by next generation sequencing (NGS) after cultivations with acetate or methanol as a carbon source.
2. Materials and Methods
2.1. Microorganism and culture condition
Clostridium cellulovorans 743B (ATCC35296) was grown anaerobically as described previously [28], with pig manure (PM) (Mie University, Tsu, Japan) as a carbon source. M. mazei (DSM# 3647) was purchased from the German Collections of Microorganisms and Cell Cultures (DSMZ, Germany) and was cultivated with the JCM230 medium [35]. 0.5% (w/v) Glucose, 0.5% acetic acid (FUJIFILM Wako Chemicals, Japan), and 0.5% (w/v) cellobiose (Sigma, MO, USA) were used as the sole carbon source in 10 ml or 50 ml of C. cellulovorans media and was anaerobically cultivated. The microbial flora of methane production (MFMP) was obtained from methane fermentation digested liquid on January, 2017 at Gifu in Japan [32]. C. cellulovorans (C.c) was precultured with 0.5% cellobiose for 12 h at 37 °C and M. mazei (M.m) and MFMP were done with 0.5% glucose for 12 h at 37 °C, respectively. Co-cultivation was performed as approximately 1000 RLU of C.c cells and approximately 20000 RLU of MFMP cells (C.c : MFMP=1:20) and approximately 1000 RLU of C.c cells and approximately 3000 RLU of M.m cells (C.c:M.m=1:3), respectively.
2.2. 16S rRNA sequencing
Samples were crashed by Shake Master Neo (bms, Tokyo, Japan) and DNA was extracted by Fast DNA spin kit (MP Bio, CA, USA). iSeq 100 (Illumina, CA, USA) was used for sequencing under the condition of 2 × 150 bp. The 16S Metagenomics App performs taxonomic classification of 16S rRNA targeted amplicon reads using a version of the GreenGenes taxonomic database curated by Illumina. The primer sequences used in the protocol are: PCR1_Forward (50 bp): 5′–TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG–3′ and PCR1_Reverse (55 bp): 5′–GTCTCGTGG GCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC–3′, respectively The 16S rRNA sequences of MFMP previously reported [31] has been deposited in the DDBJ database (accession no. DRR160954).
2.3. Gas and organic acid concentrations
The total gas amount and the concentration of organic acids were measured as previously described [31]. The produced gas after the cultivation was recovered by downward displacement of water by a syringe (Terumo, Tokyo, Japan) and measured by gas chromatography (Shimadzu, Kyoto, Japan). The concentration of organic acids was measured by high-performance liquid chromatography (Shimadzu, Kyoto, Japan) with UV detector.
3. Results
3.1. Cultivation of C. cellulovorans with pig manure
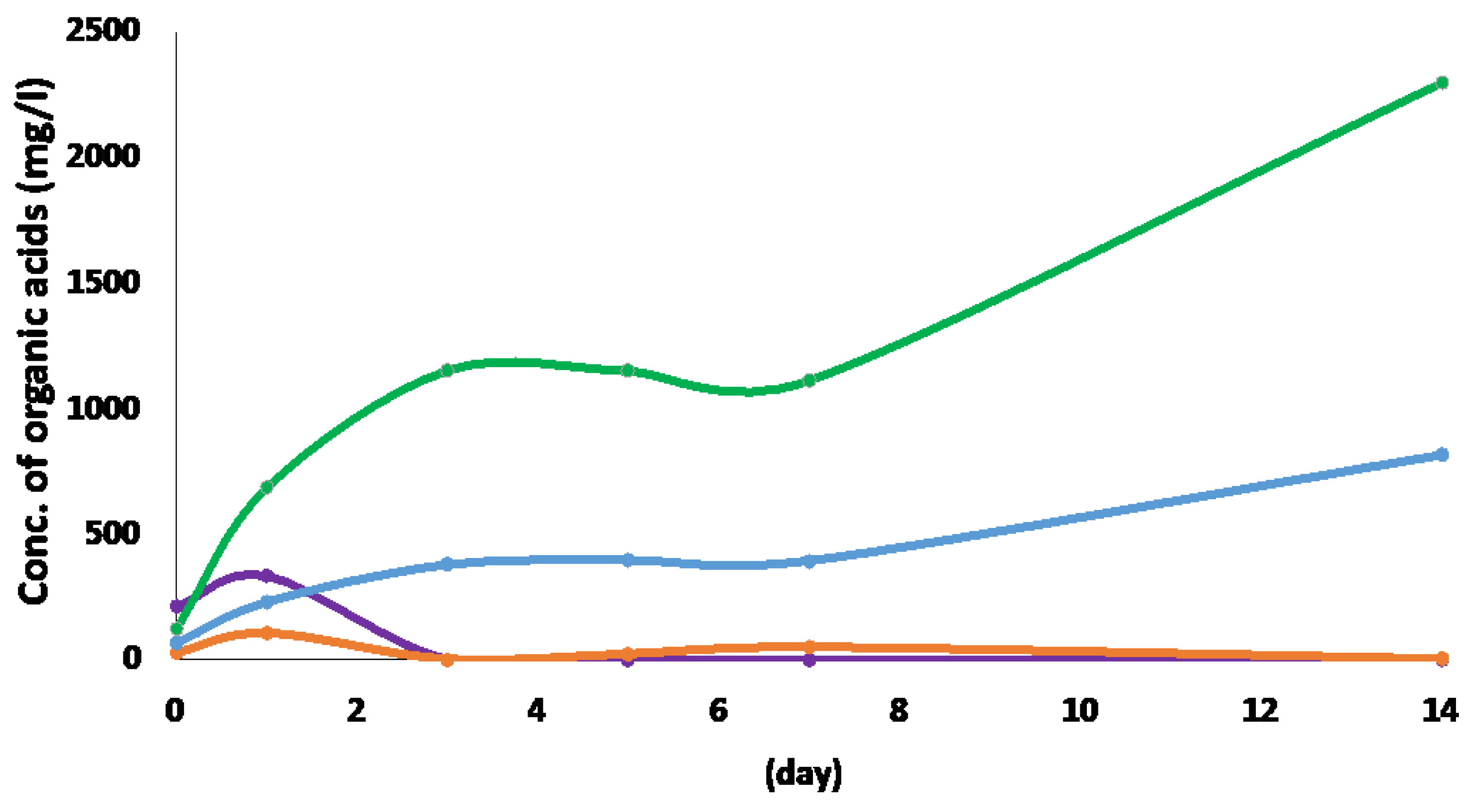
In order to promote the utilization of pig manure (PM) as an unused biomass, the cultivation of C. cellulovorans was carried out. PM was pretreated with 0.45 mm filter to remove the inhibitor for bacterial cell growth and 0.5% (w/v) pretreated PM was used as the sole carbon source in the C. cellulovorans medium. C. cellulovorans was inoculated into the PM medium and then organic acids were measured by HPLC. The result suggested C. cellulovorans was able to grow in the 0.5% PM medium and acetate and butyrate were increased, while formate and lactate were decreased after once increased at 1 day cultivation (Figure 1). Total concentrations of acetate and butyrate at 14 days was approximately 2300 mg/L and 820 mg/L, respectively, resulting that PM would be an excellent biomass for methanogenesis.
3.2. Co-cultivation of C. cellulovorans with methanogens or M. mazei
CH4 production by coculturing C. cellulovorans–methanogens (MFMP) was examined with 0.5% (w/v) glucose, 0.5% (w/v) cellobiose, and 0.5% (v/v) acetate, respectively, while cocultivation of C. cellulovrorans–M. mazei was done with 0.5% cellobiose as the sole substrate. As shown in Figure 2A, the cell growth in each coculture was observed and different patterns. On the other hand, the cocultivation of C. cellulovorans-MFMP showed CH4 production only with 0.5% acetate, whereas the cocultivation of C. cellulovorans-M. mazei with the 0.5% cellobiose medium led to no methanogenesis during the cultivation period, resulting that M. mazei could never use cellobiose for its growth (Figure 2B). These results suggested methanogenesis promotes not sugars such as glucose or cellobiose but acetate as the carbon source.
Figure 1.
Measurement of organic acids from 0.5% pig manure (PM) cultivated by C. cellulovorans. Lines: orange, formate; purple, lactate, green, acetate; blue, butyrate.
Figure 1.
Measurement of organic acids from 0.5% pig manure (PM) cultivated by C. cellulovorans. Lines: orange, formate; purple, lactate, green, acetate; blue, butyrate.
Bars: blue, 0.5% cellobiose cultivated with C. cellulovorans and MFMP; orenge, 0.5% acetate cultivated with C. cellulovorans and MFMP; gray, 0.5% glucose cultivated with C. cellulovorans and MFMP; yellow, 0.5% cellobiose cultivated with C. cellulovorans and M. mazei.
3.3. Effect of carbon sources with methanogens
In order to produce CH4 efficiently, MFMP was examined with the culture media of 1.0% (v/v) acetate and 1.0% (v/v) methanol, respectively (Figure 3). The cell growth in the medium of 1.0% acetic acid was a peak at 1 day, while that in the medium of 1.0% methanol was a peak at 16 days (Figure 3A). On the other hand, CH4 production on the methanol medium was increased from 8 days, and then the maximum production of methane was a peak at 16 days (Figure 3B). In case of the acetic acid medium, CH4 production was lower than that of the methane medium, resulting in the difference of metabolic pathway of methanogenesis in MFMP. These results indicated methanogenesis easily occurs for not acetate but methanol and the production of methane by 1.0% methanol was 8 times higher than that by 1.0% acetate.
3.4. Identification of methanogens for different carbon sources
MFMP was precultivated with 0.5% glucose medium for 12 h at 37°C and then 1,000 RLU of MFMP cells was inoculated into the C. cellulovorans medium containing 1% acetate or 1% methanol at 37 °C for 72 h. After DNA extraction from the growth cells of each medium, 16S rRNA analyses were carried out by next generation sequencer. As shown in Table 2, Methanofollis was a majority of archaea and was 0.211% in 1% acetate medium for 72 h cultivation. On the other hand, Methanofollis in 1% methanol medium was found, i.e., 0.007% for 24 h cultivation and 0.490% for 72 h cultivation, respectively. On the other hand, Methanosarcina barkeri was a typical methanogen and was 0.011% for 24 h cultivation and 0.015% for 72 h cultivation, respectively, in 1% acetate medium. Interestingly, for 72 h cultivation 0.004% of M. mazei was found in 1% methanol medium, while 0.571% of M. barkeri was detected in the same medium. These results indicated the growth of methanogens was dependent on the carbon sources and their growth trend of individual methanogens seemed remarkably different under the sole carbon sources.
4. Discussion
In Japan, around 25.31 million tons of food waste was generated in 2018 from food manufacturing, retail, and consumer households [36]. Appropriate food waste management practices should be implemented to minimize the environmental impacts and maximize social and economic benefits. Since recycling food waste as compost and animal feed is preferred in Japan, composting of food waste still presents high-quality demand by farmers, relatively low price, and a shortage of cropland for application [37,38,39]. Therefore, since the most successful application so far at the commercial scale has been anaerobic digestion (AD), which has been widely adopted for waste treatment, pig manure (PM) is a plentiful source of organic compounds that can be used as feedstock in AD. Namely, recycling food waste into fermented liquid feed (FLF) for pigs that contains several nutrients required for bacterial growth was considered a possible alternative for many years. Also, PM has a high buffering capacity, which possibly protects AD against failures due to the accumulation of volatile fatty acids (VFAs) [40,41,42]. It was reported that the effect of varying PM with food waste mixing ratio was evaluated on methane yield, suggesting that the feedstock composition of 60:40 (volatile solid basis) enhanced methane yield significantly [43]. On the other hand, the other group reported that using vegetable processing wastes as co-substrate with a feedstock ratio of 50:50 (dry weight basis) could improve methane yield up to 3-fold [44]. Thus, since several potential co-substrates have been examined to assess the effect of varying feedstock composition on increasing methane yield and improving the AD process performance, the VFAs of the C. cellulovorans medium containing PM were measured in this study. As a result, acetic acid (approx. 2300 mg/mL) and butyric acid (approx. 820 mg/mL) were accumulated for 14 days, respectively (Figure 1). As another possibility to efficient methane production, since the high ammonia concentration might inhibit bacterial activity in AD [45,46,47,48,49], PM was pretreated with 0.45 mm filtration before inoculation of C. cellulovorans in this study. Therefore, by adjusting the carbon-to-nitrogen (C/N) ratio, co-digestion of PM with organic waste containing high carbon dilute seemed to improve the inhibitory effect of ammonia and to enhance the macro and micronutrient balance in the feedstocks [50,51]. Besides, cow manure (CM) is rich in nutrients and can provide strong buffer capacity, and thus, CM seems more robust than other manures in AD [52]. Therefore, the alleviation of ammonia inhibition when CM is used in AD seems not that urgent and should not be the priority of co-digestion. Additionally, CM is categorized as lignocellulosic waste due to its high amount of lignocellulose (50% in dry matter), which is relatively low in other types of manure [53]. Hence, to make full use of CM to produce more methane via co-digestion, attention should be paid to how to improve the degradation of recalcitrant lignocellulose in CM. In addition, the current study determined biogas production in single-stage and two-stage AD using sheep manure (SP) as substrate and yak rumen fluid as the inoculum. Yak rumen fluid is rich in hydrolytic bacteria [54] and, consequently, its inclusion should improve the degradation of lignocellosic biomass, leading to high biogas production.
Pathways related to methanogenesis and relevant energy conservation systems were reconstructed in all archaeal the metagenome-assembled genomes (MAGs) [55]. The holistic microbial community activity could be evaluated by the average RPKM of genes in each KEGG module [56]. Thus, in order to maintain the methanogenic activity of the microbial community, a syntrophic behavior is needed to synthesize numerous metabolites. An overall shift of the microbial activity was observed in the majority of the KEGG modules after H2 addition. Moreover, H2 also enhanced the activity of the glyoxylate cycle and the biosynthesis of lipids and specific amino acids. Besides H2, also formate, similarly formed during fermentative metabolism, is an important electron carrier in e.g., syntrophic fatty acid-degrading methanogenic consortia [57]. In fact, formate was low concentration and immediately consumed in the PM medium (Figure 1). Therefore, other anaerobes may use both H2 and formate as an electron donor for sulfate respiration or methanogenesis.
Clostridium coculture systems are typically used to produce biofuels such as H2 and CH4, solvents, and organic acids [58]. Because cellulosic materials are commonly found in nature [18], the specific metabolic capacities of cellulolytic strains and producers in coculture systems have attracted significant attention and offered many long-term prospects for development. Furthermore, since the combination of genome-centric metagenomics and metatranscriptomics successfully revealed individual functional roles of microbial members in methanogenic microcosms, these results assigned a multi-trophic role to Methanosarcina ssp., suggesting its ability to perform simultaneous methanogenesis from acetate, CO2 and methanol/methylamine [55]. MFMP used in this study originally consisted of C. butyricum (0.005%) identified as the same genus of C. celulovorans and M. mazei (1.34%) found among methanogens [32]. Furthermore, other methanogens such as Methanosaetaceae, Methanosaeta, and Methanospirillaceae were also identified in MFMP. The genus Methanosaeta, which utilizes only acetate, was a large portion of ratio next to Methanosarcina. On the other hand, 1% acetate or 1% methanol was used as the sole carbon source for MFMP cultivation in this study. As a result, Methanofollis (0.211%) was dominant in the 1% acetate medium for 72 h cultivation, whereas Methanosarcina siciliae (1.178%), M. barkeri (0.571%), and Methanofollis (0.490%) were major species in the 1% methanol medium for 72 h cultivation (Table 2). It is thought that all methanogens are physiologically specialized and able to scavenge the electrons from H2, formate, acetate, and methanol, having CH4 as the final product [49]. The Clostridium coculture system can also produce CH4 in addition to producing H2 and solvents, in particular the coculture of cellulolytic Clostridia and methanogens including M. barkeri Fusaro, M. mazei, and Methanothermobacter thermautotrophicus, the methanogens utilized H2 and CO2, acetate, and even formate that was generated by the cellulolytic Clostridia from cellulose to produce CH4 [33,59]. In this study, CH4 production by cellobiose was not found in the cocultivation of C. cellulovorans-M. mazei (C.c : M.m = 1:3), while only acetate led to methanogenesis in the cocultivation of C. cellulovorans-MFMP (Figure 2). In addition, since M. barkeri was more dominant than M. mazei in MFMP cultivation according to the 16S rRNA analysis (Table 2), it seemed that Methanosarcina spp. may play a key methanogenesis in MFMP. So far, it has been reported that CH4 production was investigated with sugar beet pulp [16] and mandarin orange peel [17] in the cocultivation of C. cellulovorans-MFMP (C.c : MFMP= 1:20). Therefore, carbon sources such as acetic acid and methanol were compared by the production of CH4 in this study. As expected, CH4 production from methanol was approximately eight times higher than that from acetic acid, with related to the cell growth of MFMP (Figure 3). Thus, methanogens seemed to be altered in their flora dependent on the sole carbon source.
5. Conclusions
In this study, C. cellulovorans was cultivated with PM and cocultivation of C. cellulovoroans-M. mazei or C. cellulovorans-MFMP was performed with different carbon sources. Since the cultivation of C. cellulovorans with PM had much acetic acid, it was thought to be one of excellent biomass for methane production. On the other hand, methanol was a best carbon source for CH4 production with MFMP. Regarding next generation sequence analysis of MFMP, Methanofollis (0.211%) was dominant in the 1% acetic acid medium for 72 h cultivation, whereas Methanosarcina siciliae (1.178%), M. barkeri (0.571%), and Methanofollis (0.490%) were major species in 1% methanol medium for 72 h cultivation. Therefore, it seemed Methanosarcina spp. may play a key methanogenesis in MFMP.
Author Contributions
Performed the experiments: H.S. and H.T.; experimental design: Y.T.; writing—original draft preparation, H.S. and F.O.; writing—review and editing, Y.T.; supervision, Y.T.; funding acquisition, Y.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was a part supported by Chubu Electric Power Co., Inc., Mitsui Chemicals, Inc., and Yanmar Holdings CO., LTD.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Mie University.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Profs. Dr. Hiroki Matsui and Dr. Tomomi Ban at Mie University for providing PM. The authors also thank Ms. Mayu Hayashi and Ms. Midori Kosaka at Tamaru’s laboratory for technical support of experimental studies.
Conflicts of Interest
The authors have no conflicts of interest relevant to the content of this article.
References
- Pan, X.; Angelidaki, I.; Alvarado-Morales, M.; Liu, H.; Liu, Y.; Huang, X.; Zh, G. Methane production from formate, acetate and H2/CO2; focusing on kinetics and microbial characterization. Bioresour. Technol. 2016, 218, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Christensen, T.R.; Johansson, T.; Åkerman, H.J.; Mastepanov, M.; Malmer, N.; Friborg, T.; Crill, P.; Svenssonet, B.H. Thawing sub-arctic permafrost: Effect sonvegetation and methane emissions. Geophys. Res. Lett. 2004, 31, L04501. [Google Scholar] [CrossRef]
- Woodcroft, B.J.; Singleton, C.M.; Boyd, J.A.; Evans, P.N.; Emerson, J.B.; Zayed, A.F.Z.; Hoelzle, R.D.; Lamberton, T.O.; Mccalley, C.K.; Hodgkins, S.B.; et al. Genome-centric view of carbon processing in thawing permafrost. Nature 2018, 560, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Hattori, S. Syntrophic acetate-oxidizing microbes in methanogenic environments. Microbes Environ. 2008, 23, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.K.; Kaster, A.K.; Seedorf, H.; Buckel, W.; Hedderich, R. Methanogenic archaea: Ecologically relevant differences in energy conservation. Nat. Rev. Microbiol. 2008, 6, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.L.; Patel, B.K.C.; Ollivier, B. Taxonomic, phylogenetic, and ecological diversity of methanogenic Archaea. Anaerobe 2000, 6, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Deppenmeier, U.; Müller, V.; Gottschalk, G. Pathways of energy conservation in methanogenic archaea. Arch. Microbiol. 1996, 165, 149–163. [Google Scholar] [CrossRef]
- Kruse, S.; Goris, T.; Westermann, M.; Adrian, L.; Diekert, G. Hydrogen production by Sulfurospirillum species enables syntrophic interactions of Epsilonproteobacteria. Nat. Commun. 2018, 9, 4872. [Google Scholar] [CrossRef]
- Fischbach, M.A.; Segre, J.A. Signaling in host-associated microbial com- munities. Cell 2016, 164, 1288–1300. [Google Scholar] [CrossRef]
- Kenny, D.J.; Balskus, E.P. Engineering chemical interactions in microbial communities. Chem Soc Rev. 2018, 47, 1705–1729. [Google Scholar] [CrossRef]
- Brenner, K.; You, L.; Arnold, F.H. Engineering microbial consortia: A new frontier in synthetic biology. Trends Biotechnol. 2008, 26, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.Z.; Song, H.; Wang, E.X.; Liu, Y.; Yuan, Y.J. Design and construction of synthetic microbial consortia in China. Synth Syst Biotechnol. 2016, 1, 230–235. [Google Scholar] [CrossRef]
- De Bok, F.A.; Plugge, C.M.; Stams, A.J. Interspecies electron transfer in methanogenic propionate degrading consortia. Water Res. 2004, 38, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zou, W.; Zhang, K.; Ye, G.; Yang, J. Advances and applications of Clostridium co-culture systems in biotechnology. Front. Microbiol. 2020, 11, 560223. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Hu, W.; Li, Y. Biodegradation mechanisms and kinetics of azo dye 4BS by a microbial consortium. Chemosphere 2004, 57, 293–301. [Google Scholar] [CrossRef]
- Khouni, I.; Marrot, B.; Amar, R.B. Treatment of reconstituted textile waste-water containing a reactive dye in an aerobic sequencing batch reactor using a novel bacterial consortium. Sep Purif Technol. 2012, 87, 110–119. [Google Scholar] [CrossRef]
- Safonova, E.; Kvitko, K.V.; Iankevitch, M.I.; Surgko, L.F.; Afti, I.A.; Reisser, W. Biotreatment of industrial wastewater by selected algal-bacterial consortia. Eng Life Sci. 2004, 4, 347–353. [Google Scholar] [CrossRef]
- Xu, X.H.; Liu, X.M.; Zhang, L.; Mu, Y.; Zhu, X.Y.; Fang, J.Y.; Li, S.P.; Jiang, J.D. Bioaugmentation of chlorothalonil-contaminated soil with hydrolytically or reductively dehalogenating strain and its effect on soil microbial community. J Hazard Mater. 2018, 351, 240–249. [Google Scholar] [CrossRef]
- Sabra, W.; Dietz, D.; Tjahjasari, D.; Zeng, A.P. Biosystems analysis and engineering of microbial consortia for industrial biotechnology. Eng Life Sci. 2010, 10, 407–421. [Google Scholar] [CrossRef]
- Bertrand, S.; Bohni, N.; Schnee, S.; Schumpp, O.; Gindro, K.; Wolfender, J.L. Metabolite induction via microorganism co-culture: A potential way to enhance chemical diversity for drug discovery. Biotechnol Adv. 2014, 32, 1180–1204. [Google Scholar] [CrossRef]
- Eiteman, M.A.; Lee, S.A.; Altman, R.; Altman, E. A substrate-selective co-fermentation strategy with Escherichia coli produces lactate by simultaneously consuming xylose and glucose. Biotechnol Bioeng. 2009, 102, 822–827. [Google Scholar] [CrossRef]
- Wang, E.X.; Ding, M.Z.; Ma, Q.; Dong, X.T.; Yuan, Y.J. Reorganization of a synthetic microbial consortium for one-step vitamin C fermentation. Microb Cell Fact. 2016, 15, 21. [Google Scholar] [CrossRef]
- Schroeckh, V.; Scherlach, K.; Nützmann, H.W.; Shelest, E.; Schmidtheck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc Natl Acad Sci USA. 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.X.; He, J.Z. Characterization of a thermostable xylanase from a newly isolated Kluyvera species and its application for biobutanol production. Bioresour Technol. 2013, 135, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.L.; Goyal, G.; Chen, W. Surface display of a functional minicelluloome by intracellular complementation using a synthetic yeast consortium and its application to cellulose hydrolysis and ethanol production. Appl Environ Microbiol. 2010, 76, 7514–7520. [Google Scholar] [CrossRef]
- Lovley, D.R. Happy together: Microbial communities that hook up to swap electrons. ISME J. 2016, 11, 327–336. [Google Scholar] [CrossRef]
- Charubin, K.; Bennett, R.K.; Fast, A.G.; Papoutsakis, E.T. Engineering Clostridium organisms as microbial cell-factories: Challenges & opportunities. Metab. Eng. 2018, 50, 173–191. [Google Scholar] [PubMed]
- Tamaru, Y.; Miyake, H.; Kuroda, K.; Nakanishi, A.; Kawade, Y.; Yamamoto, K.; Uemura, M.; Fujita, Y.; Doi, R.H.; Ueda, M. Genome sequence of the cellulosome-producing mesophilic organism Clostridium cellulovorans 743B. J. Bacteriol. 2010, 192, 901–902. [Google Scholar] [CrossRef] [PubMed]
- Tamaru, Y.; Miyake, H.; Kuroda, K.; Nakanishi, A.; Matsushima, C.; Doi, R.H.; Ueda, M. Comparison of the mesophilic cellulosome- producing Clostridium cellulovorans genome with other cellulosome-related clostridial genomes. Micro. Biotechnol. 2011, 4, 64–73. [Google Scholar] [CrossRef]
- Tomita, H.; Okazaki, F.; Tamaru, Y. Direct IBE fermentation from mandarin orange wastes by combination of Clostridium cellulovorans and Clostridium beijerinckii. AMB Express 2019, 9, 1. [Google Scholar] [CrossRef]
- Tomita, H.; Okazaki, F.; Tamaru, Y. Biomethane production from sugar beet pulp under cocultivation with Clostridium cellulovorans and methanogens. AMB Express 2019, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Tomita, H.; Tamaru, Y. The second-generation biomethane from mandarin orange peel under cocultivation with methanogens and the armed Clostridium cellulovorans. Fermentation 2019, 5, 95. [Google Scholar] [CrossRef]
- Lu, H.; Ng, S.-K.; Jia, Y.; Cai, M.; Lee, P.K.H. Physiological and molecular characterizations of the interactions in two cellulose-to-methane cocultures. Biotechnol. Biofuels 2017, 10, 37. [Google Scholar] [CrossRef]
- Singhvi, M.; Kim, B.S. Green hydrogen production through consolidated bioprocessing of lignocellulosic biomass using nanobiotechnology approach. Biores. Technol. 2022, 365, 128108. [Google Scholar] [CrossRef]
- Goevert, D.; Conrad, R. Effect of substrate concentration on carbon isotope fractionation during acetoclastic methanogenesis by Methanosarcina barkeri and M. acetivorans and in rice field soil. Appl. Environ. Microbiol. 2009, 75, 2605–2612. [Google Scholar] [CrossRef]
- Ministry of the Environment. MOE Discloses the Estimated Amount of Japan’s Food Loss and Waste Generated in FY 2018. 2021. Available online: https://www.env.go.jp/en/headline/2515.html (accessed on 10 November 2022).
- Watanabe, E.; Seike, N.; Motoki, Y.; Inao, K.; Otani, T. Potential application of immunoassays for simple, rapid and quantitative detections of phytoavailable neonicotinoid insecticides in cropland soils. Ecotoxicol. Environ. Saf. 2016, 132, 288–294. [Google Scholar] [CrossRef]
- Cheung, H.N.B.; Huang, G.H.; Yu, H. Microbial-growth inhibition during composting of food waste: Effect of organic acids. Bioresour. Technol. 2010, 101, 5925–5934. [Google Scholar] [CrossRef] [PubMed]
- Nakasaki, K.; Hirai, H. Temperature control strategy to enhance the activity of yeast inoculated into compost raw material for accelerated composting. Waste Manag. 2017, 65, 29–36. [Google Scholar] [CrossRef]
- Wang, H.; Lim, T.T.; Duong, C.; Zhang, W.; Xu, C.; Yan, L.; Mei, Z.; Wang, W. Long-Term Mesophilic Anaerobic Co-Digestion of Swine Manure with Corn Stover and Microbial Community Analysis. Microorganisms 2020, 8, 188. [Google Scholar] [CrossRef] [PubMed]
- Córdoba, V.; Fernández, M.; Santalla, E. The effect of different inoculums on anaerobic digestion of swine wastewater. J. Environ. Chem. Eng. 2016, 4, 115–122. [Google Scholar] [CrossRef]
- Cuetos, M.J.; Fernández, C.; Gómez, X.; Moran, A. Anaerobic co-digestion of swine manure with energy crop residues. Biotechnol. Bioprocess Eng. 2011, 16, 1044–1052. [Google Scholar] [CrossRef]
- Dennehy, C.; Lawlor, P.G.; McCabe, M.S.; Cormican, P.; Sheahan, J.; Jiang, Y.; Zhan, X.; Gardiner, G.E. Anaerobic co-digestion of pig manure and food waste: Effects on digestate dewaterability, and microbial community dynamics. Waste Manag. 2018, 71, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Molinuevo-Salces, B.; González-Fernández, C.; Gómez, X.; García-González, M.C.; Morán, A. Vegetable processing wastes addition to improve swine manure anaerobic digestion: Evaluation in terms of methane yield and SEM characterization. Appl. Energy 2012, 91, 36–42. [Google Scholar] [CrossRef]
- Mata-Alvarez, J.; Dosta, J.; Macé, S.; Astals, S. Codigestion of solid wastes: A review of its uses and perspectives including modeling. Crit. Rev. Biotechnol. 2011, 31, 99–111. [Google Scholar] [CrossRef]
- Yenigün, O.; Demirel, B. Ammonia inhibition in anaerobic digestion: A review. Process Biochem. 2013, 48, 901–911. [Google Scholar] [CrossRef]
- Astals, S.; Nolla-Ardèvol, V.; Mata-Alvarez, J. Anaerobic co-digestion of pig manure and crude glycerol at mesophilic conditions: Biogas and digestate. Bioresour. Technol. 2012, 110, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, G.; Feng, Y.; Ren, G.; Han, X. Optimizing feeding composition and carbon–nitrogen ratios for improved methane yield during anaerobic co-digestion of dairy, chicken manure and wheat straw. Bioresour. Technol. 2012, 120, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Moestedt, J.; Müller, B.; Westerholm, M.; Schnürer, A. Ammonia threshold for inhibition of anaerobic digestion of thin stillage and the importance of organic loading rate. Microb. Biotechnol. 2016, 9, 180–194. [Google Scholar] [CrossRef]
- Hartmann, H.; Ahring, B.K. Strategies for the anaerobic digestion of the organic fraction of municipal solid waste: An overview. Water Sci. Technol. 2006, 53, 7–22. [Google Scholar] [CrossRef]
- Zamanzadeh, M.; Hagen, L.H.; Svensson, K.; Linjordet, R.; Horn, S.J. Biogas production from food waste via co-digestion and digestion- effects on performance and microbial ecology. Sci. Rep. 2017, 7, 17664. [Google Scholar] [CrossRef]
- Font-Palma, C. Methods for the treatment of cattle manure. A review. C-J. Carbon Res. 2019, 5, 27. [Google Scholar] [CrossRef]
- Kafle, G.K.; Chen, L. Comparison on batch anaerobic digestion of five different live-stock manures and prediction of biochemical methane potential (BMP) using different statistical models. Waste Manag. 2016, 48, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Rabee, A.E.; Sayed Alahl, A.A.; Lamara, M.; Ishaq, S.L. Fibrolytic rumen bacteria of camel and sheep and their applications in the bioconversion of barley straw to soluble sugars for biofuel production. PLoS ONE 2022, 17, e0262304. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Campanaro, S.; Treu, L.; Seshadri, R.; Ivanova, N.; Kougias, P.G.; Kyrpides, N.; Angelidaki, I. Metabolic dependencies govern microbial syntrophies during methanogenesis in an anaerobic digestion ecosystem. Microbiome 2020, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621. [Google Scholar] [CrossRef]
- De Bok, F.A.; Plugge, C.M.; Stams, A.J. Interspecies electron transfer in methanogenic propionate degrading consortia. Water Res. 2004, 38, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wu, R.; Zhou, J.; He, A.; Xu, J.; Xin, F.; Zhang, W.; Ma, J.; Jiang, M.; Dong, W. Recent advances of biofuels and biochemicals production from sustainable resources using co-cultivation systems. Biotechnol. Biofuels 2019, 12, 155. [Google Scholar] [CrossRef]
- Sasaki, D.; Morita, M.; Sasaki, K.; Watanabe, A.; Ohmura, N. Acceleration of cellulose degradation and shift of product via methanogenic co-culture of a cellulolytic bacterium with a hydrogenotrophic methanogen. J. Biosci. Bioeng. 2012, 114, 435–439. [Google Scholar] [CrossRef]
Figure 2.
Measurement of ATP amount (RLU) and methane production (B) with cocultivation of C. cellulovorans and MFMP or M. mazei.
Figure 2.
Measurement of ATP amount (RLU) and methane production (B) with cocultivation of C. cellulovorans and MFMP or M. mazei.
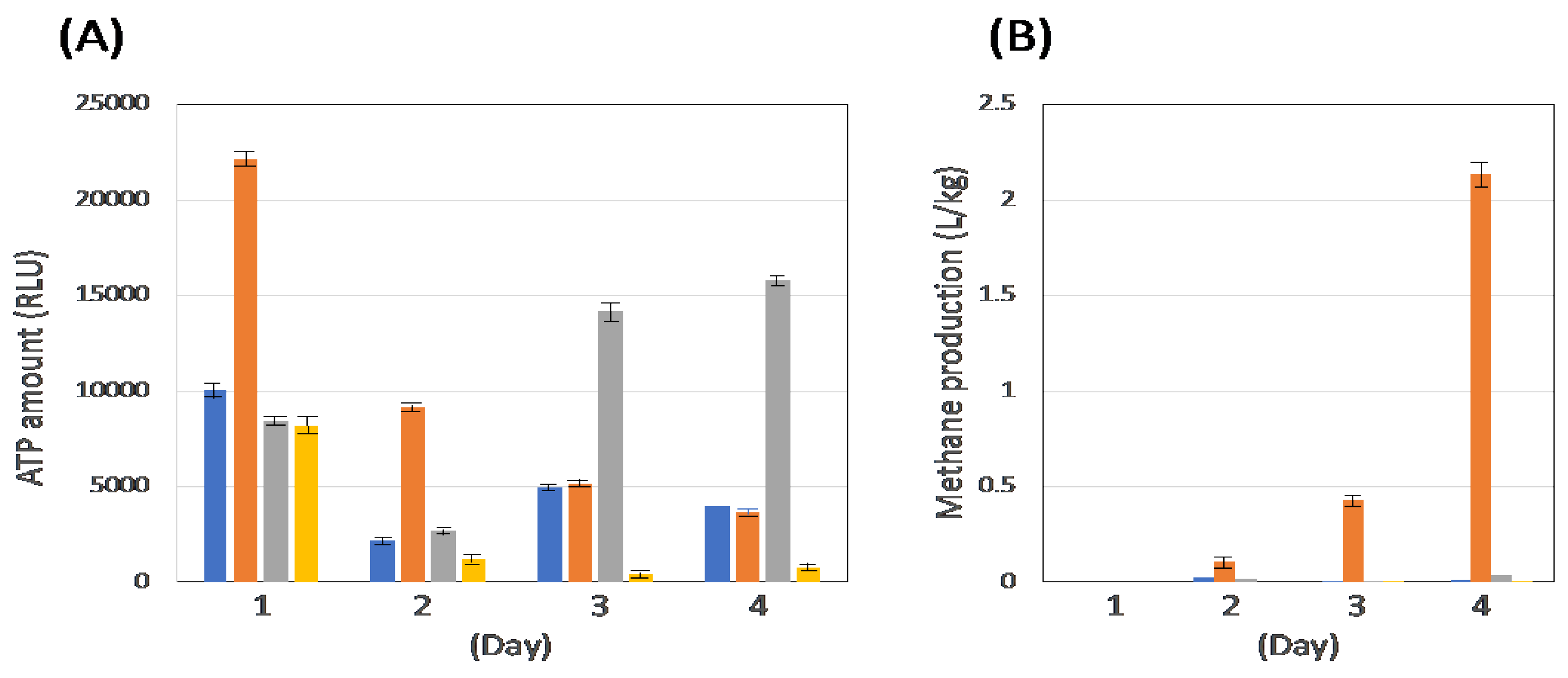
Figure 3.
Measurement of ATP amount (RLU) (A) and methane production (B) in MFMP cultivation. (A) black line, 1% methanol; wavey line, 1% acetic acid. (B) black bar, 1% methanol; gray bar, 1% acetic acid.
Figure 3.
Measurement of ATP amount (RLU) (A) and methane production (B) in MFMP cultivation. (A) black line, 1% methanol; wavey line, 1% acetic acid. (B) black bar, 1% methanol; gray bar, 1% acetic acid.
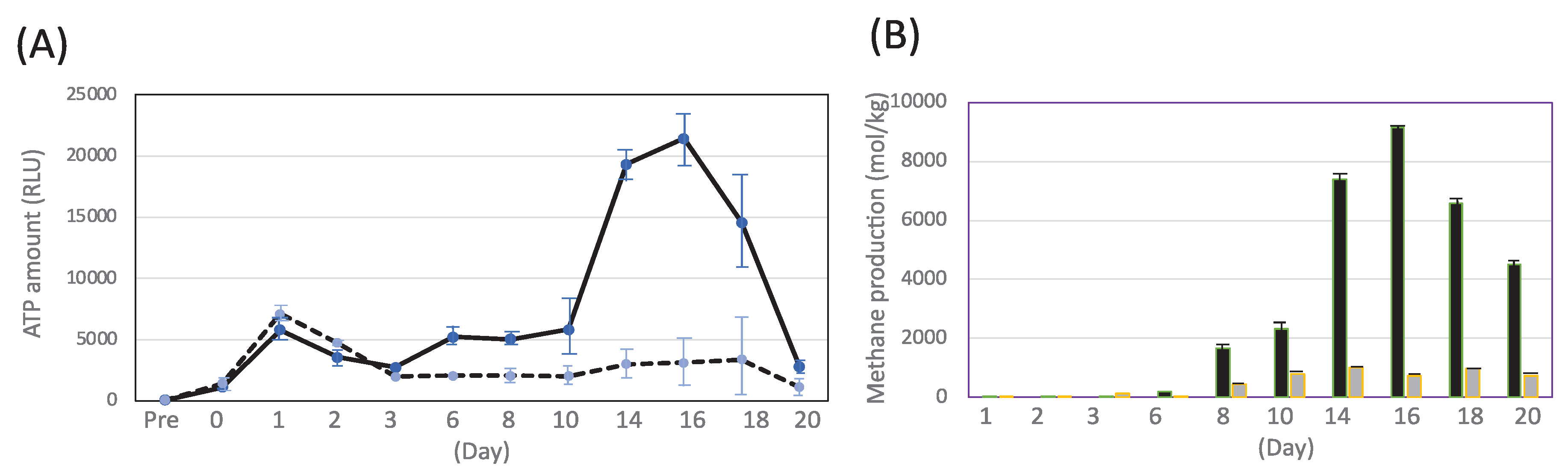

Table 1.
Methanogenic reactions from typical substrates
| Reactions | DG0′ (kJ/mol CH4) |
Microorganisms |
|
I. Hydrogen 4H2 +CO2→CH4 +2H2O |
-135 | Most methanogens |
|
II. Formate 4HCOOH→ CH4+3CO2+2H2O |
-130 | Many hydrogenotrophic methanogens |
|
III. Acetate CH3COOH→CH4+CO2 |
-33 | Methanosarcina and Methanosaeta |
Table 2.
16S rRNA analysis of archaea in MFMP after cultivated with the different carbon sources.
| 1% Acetic acid | 1% Methanol | ||||||
| 24 h | 72 h | 24 h | 72 h | ||||
| Archaea | Ratio (%) | Archaea | Ratio (%) | Archaea | Ratio (%) | Archaea | Ratio (%) |
| Methanosarcina barkeri | 0.011 | Methanofollis | 0.211 | Methanofollis | 0.007 | Methanosarcina siciliae | 1.178 |
| Methanofollis | 0.008 | Methanofollis ethanolicus | 0.076 | Methanosarcina barkeri | 0.571 | ||
| Methanofollis ethanolicus | 0.006 | Methanosarcina siciliae | 0.055 | Methanofollis | 0.490 | ||
| Methanosarcina | 0.004 | Methanosarcina barkeri | 0.015 | Methanosarcina | 0.244 | ||
| Methanosarcina siciliae | 0.002 | Methanosarcina | 0.006 | Methanofollis ethanolicus | 0.131 | ||
| Methanosarcina vacuolate | 0.001 | Methanosarcina vacuolate | 0.027 | ||||
| Methanosarcina mazei | 0.004 | ||||||
| Methanofollis liminatans | 0.001 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



