Submitted:
13 February 2023
Posted:
14 February 2023
You are already at the latest version
Abstract
Berberine (BBR) is an isoquinoline that inhibits the proliferation of transformed cells in vitro, but due to its poor solubility and bioavailability, it has only moderate therapeutic potential in vivo. Increasing evidence indicates that BBR specifically targets several metabolic, signaling and gene transcription events in transformed cells, altering their progression through the cell cycle and decreasing their metabolic rate and proliferation. In order to further develop BBR as a therapeutic, its mode of cellular internalization and localization within the cell needs to be further examined. BBR’s molecular targets and interactions with kinases, transcription factors and some enzymes are discussed in an attempt to better understand BBR’s role in these important pathways and how they may lead to changes in metabolism. Lastly, this review examines some of the benefits and challenges of using BBR as an inhibitor in cancer cell proliferation in breast cancer and glioblastoma, as two examples. BBR is a potent drug with multiple targets and it is this multiplicity of function that makes BBR such a promising drug for targeting cell metabolism and proliferation.
Keywords:
nanoparticles
; berberine
; metabolism
; mitochondria
; apoptosis
1. Introduction
Berberine (BBR; Natural Yellow 18, 5,6-dihydro-9,10-dimethoxybenzo(g)-1,3-benzodioxolo(5,6-a)quinolizinium; molecular formula: C20H19NO5; molecular weight: 353.36) is a naturally occurring small 3.7 kDa nonbasic quaternary ammonium salt derived from the protoberberine group of benzylisoquinoline alkaloids (Figure 1). It is found in many medicinal plants and vegetable species, including but not limited to, Coptis Chinensis (Chinese goldthread), Hydrastis canadensis (goldenseal), Berberis vulgaris (barberry), Berberis aristata (tree turmeric), meadow rue (Thalictrum), celandine (Chelidonium), Mahonia aquifolium (Oregon grape), Xanthorhiza simplicissima (yellowroot), Phellodendron amurense (Amur cork tree), Argemone mexicana (prickly poppy), and Eschscholzia californica (Californian poppy) [1,2,3,4]. About 450-500 species belonging to the Berberidaceae family in the Berberis genus which represent the main natural source of BBR [5]. BBR is present in barks, leaves, twigs, rhizomes, roots and stems of medicinal plants with a wide distribution in barks and roots. It naturally exists in several different chemical forms such as hydrochloride, sulfate, citrate, phosphate, or bromide salt [6]. BBR products are available from numerous commercial suppliers worldwide and are sold in either pure forms or extracts. Geographically, China and India are the main manufacturers and commercial suppliers of BBR while several other countries also supply BBR products.
BBR has been traditionally used for thousands of years in Indian, Chinese and Middle-Eastern medicines against a variety of infections caused by microbes, (including bacteria, fungi, viruses and protozoa) and helminthes [1,7]. There have been no obvious side-effects reported from these home remedies or clinical applications, although dosage and duration of treatment is not always clear and there have been few double-blind clinical trials of BBR on controlled patient disease cohorts [8,9,10,11]. However, several studies on functions and health benefits of BBR have been reported in the past two decades, and they have shown some beneficial properties including anti-inflammatory, immunoregulatory, anticancer, hypoglycemic, lipid lowering and hepatoprotective in some in vitro and in vivo models [12,13,14,15,16,17,18,19,20,21]. One of the cellular effects of BBR is its ability to block the metabolic activity, proliferation and viability of transformed cells. For this reason, BBR has been proposed as an effective chemotherapy, particularly for cancers in the gastrointestinal tract. However, the efficacy of BBR is largely limited by its poor solubility and thus low bioavailability, and new technologies and drug delivery systems to improve its solubility and bioavailability are currently underway. This review will examine some of the challenges of using BBR to target cells and tissue and then provide a brief overview of BBR’s effect on cell metabolism and proliferation. These data will be discussed in the context BBR modulation of cell signaling pathways in an effort to better understand the molecular targets and pathways that are modulated in transformed cells.
2. Aqueous solubility of berberine and oral bioavailability
2.1. Aqueous solubility of berberine
It is well-known that berberine is a poorly soluble compound; however, the actual solubility data is limited. A study that examined the solubility of BBR in solutions at different conditions including pH, temperature and surfactants [22], found that the aqueous solubility of BBR chloride at 25°C and 37°C was 1.96 mg/ml (5.3 mM) and 3.16 mg/ml (8.5 mM), respectively. There were significant interactions between pH and types of buffer in determining the solubility of BBR chloride. For example, in a range of pH 1.2 - 9.0, the maximum solubility of BBR chloride was seen in phosphate buffer at pH 7.0 (4.05 mM and 9.69 mM at 25°C and 37°C, respectively), while at pH 1.2 in HCI solution, 3.0 and 5.0 in phthalate buffer, and pH 9.0 in borate buffer it was nearly 20-fold lower than that at pH 7.0 in phosphate buffer. Nevertheless, in the same buffer, such as phosphate buffer, the solubility was pH independent in the tested pH range of 2.0-9.0. Moreover, it was noted that the aqueous solubility of BBR chloride increases with temperature. A similar effect of temperature on BBR’s solubility was seen in different solvents [23].
2.2. Oral bioavailability of berberine
Numerous studies have shown that the intestinal absorption rate of BBR is very low. The absolute bioavailability of BBR is reported to be as low as 0.68% in rats [24]. An even lower bioavailability rate of 0.37% was seen in a more recent study in rats that were given intragastrically a single dose of 48.2 mg [25]. In rats, the plasma Cmax (peak plasma concentration) of BBR was 11 ng/ml after a single oral administration of 50 mg/kg body weight [26]. In humans, the plasma Cmax of BBR was 0.4 ng/ml after a single oral dose of 400 mg [27]. A human pharmacokinetic study showed that when the African and Chinese volunteers were given a single oral dose of 600 mg BBR hydrochloride (net dose of 542.8 mg), the Cmax were 0.16 ng/mL and 0.06 ng/mL, respectively [28]. Interestingly, African study participants had a higher Cmax than Chinese participants. Further examinations indicated that the difference between the Africans and Chinese was possibly attributed to variations in gut microbiota and its corresponding metabolic capacity [28].
Poor water solubility is a commonly accepted cause of low oral bioavailability of BBR. In addition, the low oral bioavailability of BBR could be attributed to additional factors [29] such as: 1) self-aggregation, which decreases the solubility of BBR in the fluids of gastrointestinal tract; 2) poor permeability across the intestinal mucous membrane, which has been reported to be related to the structure of berberine; 3) the fact that BBR is a P-glycoprotein (P-gp) substrate, limiting its transport through the gut mucus membrane by promoting efflux of BBR in the enterocytes; and 4) the intestinal first-pass elimination and hepatic degradation [30,31].
The quaternary ammonium group of BBR has low permeability through the cell membrane, which hinders the transmembrane transport and intestinal absorption of BBR [32]. It has been demonstrated in mice and humans that liver metabolizes BBR quickly by hepatic P450 enzymes such as cytochrome P450 2D6 (CYP2D) [33]. These enzymes are also active in the degradation of BBR in the intestinal tissues [34]. Following the initial report that BBR had three metabolites, namely demethyleneberberine-2-O-sulfate (HM1 and RM3), jatrorrhizine-3-O-sulfate (HM5) and thalifendine (RM5), six additional metabolites have been identified, which are jatrorrhizine-3-O-β-D-glucuronide (HM2), thalifendine-10-O-β-D-glucuronide (HM3), berberrubine-9-O-β-D- glucuronide (HM4 and RM2), 3,10-demethylpalmatine-10-O-sulfate (HM6 and RM4), columbamin-2-O-β-D-glucuronide (HM7), and demethyleneberberine-2,3-di-O- β-D-glucuronide (RM1) in human and rat urine samples [35]. It is emerging that BBR undergoes a similar post absorption biotransformation and metabolism in the intestine and liver in rats and humans, contributing in part to its low bioavailability after oral supplementation [34,36,37]. Amongst the multi-factors, poor absorption in the small intestine is a key factor that presently limits the efficacy of BBR and therefore, a number of new formulations or delivery systems have been developed to improve the intestinal absorption and oral bioavailability of BBR, including emulsions, nanoparticles, and enhancers, with substantial progresses being achieved in nearly the past two decades [29,38,39].
3. Internalization of berberine into cells and molecular targets
3.1. Internalization of berberine into cells
Despite its many biological effects, the underlying molecular mechanism of BBR internalization into cells is poorly understood. It is presumed that BBR binds to the surface of the cell membrane and is internalized in some manner into the cytoplasmic space, either within vesicles or through diffusion. Although very few studies are available that examine the internalization of BBR into cells, there is some evidence from our laboratory that this is an energy-dependent process. We have used both fluorescence microscopy (Figure 2A) and flow cytometry (Figure 2B,C) to show that BBR is internalized into bone marrow derived mouse mast cells (BMMC). Fluorescent images show cytoplasmic labelling suggesting that BBR is internalized into BMMC (Figure 2A). Flow cytometric analysis of BBR treated BMMC shows that there is an increase in BBR fluorescence as indicated by a shift of histogram towards the right (Figure 2B). Moreover, with increasing concentration of BBR there is an increase in the % of BMMC that are positive for BBR fluorescence (Figure 2C).
We have demonstrated that BBR internalizes into resting and IL-3-activated BMMC by utilizing energy-dependent active transport mechanism that is dependent upon glucose metabolism and temperature [40]. Moreover, internalization of BBR into metabolically active BMMC is time-dependent. Our time course (0.01 hr- 24 hr) experiment show that BBR internalization increased over time, requiring at least 4 hr. Since BBR internalization is not immediate and requires several hours of incubation, it may be that cells require protein synthesis or structural reorganization in order to internalize BBR. Methanol fixation almost completed blocked BBR internalization. Since methanol extracts cholesterol from the plasma membrane and effectively “fixes” cells, this further supports the hypothesis that BBR does not enter cells through passive diffusion across the cell membrane, but involves energy-dependent active transport mechanisms. Surprisingly, incorporation of BBR into Lipofectamine 2000 also significantly inhibited BBR internalization. This was unexpected since Lipofectamine is widely used as a DNA and RNA transfection reagent and its efficacy comes from its ability to bypass the microtubule-dependent endocytic pathway and move through the cell cytoplasm via Brownian diffusion [41]. This observation suggests that BBR requires specific interactions with the plasma membrane, and that Lipofectamine 2000 disrupts these interactions, therefore blocking BBR internalization. Does this suggest that BBR might be binding a surface receptor that facilitates its internalization? Possibly, but if a receptor, or group of receptors are involved in BBR internalization, it is likely that these receptors are not specific for only BBR but facilitate the internalization of similar compounds with similar chemical structures. There are several examples of such receptors like the ATP-binding cassette (ABC) transporters and solute carriers which facilitate the internalization of other small molecules [42,43]. Nevertheless, our data indicates that reagents such as Lipofectamine 2000 might not be ideal reagents for facilitating delivery of drug cargos into cells. Therefore, designing alternative approaches that augment these BBR interactions with the plasma membrane might provide a better drug delivery strategy for BBR.
Our data also shows that BBR’s internalization is dependent upon actin polymerization and lysosomal pH. We further show that BBR is internalized via activation of phosphoionositide 3-kinase (PI3-K), a feature of the autophagy dependent endocytic pathway. In contrast, when BMMC were pre-treated with IL-3, BBR endocytosis was enhanced suggesting that its internalization is dependent upon actively dividing cells. Using inhibitors of the clathrin-mediated endocytic pathway, we show some data to suggest that BBR’s internalization may be partly dependent upon clathrin [44]. Taken together, our data suggests that BBR internalization may also be linked to the progression of cells into the proliferative stages of the cell cycle which is enhanced upon IL-3 treatment. Figure 3 shows possible endocytic routes of BBR into BMMC.
BBR endocytosis might be partially dependent upon clathrin (1). BBR endocytosis is drastically reduced by the phosphoinositide 3-kinase (PI3K) inhibitor 3-methyladenine, that plays a prominent role in fusion of macropinosomes with the plasma membrane. Hence, it is possible that majority of BBR might utilize a clathrin independent (2) pinocytic pathway to internalize into BMMC (3). Methanol fixation that removes cholesterol, the integral structural component of caveole, substantially reduces BBR internalization into BMMC. Therefore, involvement of caveolin (4) in BBR’s endocytosis might be a possibility and requires further investigation. IL-3 treatment significantly enhanced BBR endocytosis in BMMC (5). We speculate that IL-3 might also: (5a) activate a specific pinocytic pathway and that in turn enhances BBR endocytosis or (5b) promotes the IL-3 receptor’s clathrin-mediated endocytosis that could perhaps be utilized as a trojan horse by BBR to internalize into BMMC or (5c) engage partially quiescent BMMC into the proliferative stages of the cell cycle. Regardless of the mode of internalization, BBR endocytosis is dependent on actin cytoskeleton (shown in red), that also plays a role in vesicular transport inside the cell. Once internalized into early endosomes (EE) by utilizing a potential internalization pathway, the EE ripens into late endosomes (LE, 6) that ultimately matures to endolysosomes (EL, 7) via further lowering of the pH. The EL and the autophagosomes (AP) fuse to form the autolysosomal compartment (AL), where PI3K plays an important role (8). BBR could also be internalized via interacting with a cell surface receptor shown in brown. Therefore, internalization of BBR might be inhibited when it is incorporated into lipoplexes (9).
Based on our observation, we speculate that BBR can be internalized by BMMC either via clathrin-mediated or clathrin-independent pathways. While clathrin-mediated pathway depends on the formation of clathrin coated endocytic vesicles around the cargo, clathrin-independent pathways depend upon PI3-K (pinocytosis) or cholesterol (caveolin mediated endocytosis). Once within the cell, BBR has a variety of molecular targets.
3.2. Molecular targets of berberine
Using the PANTHER (http://www.pantherdb.org/) classification system, in vitro BBR targets were analyzed and it was found that BBR acts on a diverse range of molecular targets. The most common function of these molecular targets included receptor binding (including epidermal growth factor receptor, EGFR; vascular endothelial growth factor A; interleukin-1;; interleukin-6; growth/differentiation factor 15, NAG-1 and glucagon-like peptide, GLP1), kinase activity (such as cyclin-dependent kinases CDK1 and CDK6; receptor tyrosine kinase, ERBB2; mitogen activated protein kinases, ERK1, ERK2, MAPK8), protein binding, transcription activity, DNA binding and kinase regulator activity [45]. Based on the algorithm, these targets were related to intracellular signal transduction, cell surface receptor linked signaling, cell motion, cell cycle control, immunity and protein metabolism.
4. Effects of berberine on metabolic pathways, apoptosis and necrosis of transformed cells
4.1. Effects of berberine on metabolic pathways of transformed cells
The proliferation of transformed cancerous cells is mainly driven by energy generation in the mitochondria. When the metabolic pathways in the mitochondria are disrupted, the cell cycle is arrested, apoptotic pathways are initiated and cell growth is stalled. Chemotherapeutic research, particularly work addressing multidrug resistance (MDR), has been focused on targeting the mitochondria and inhibiting the metabolic activity of mitochondrial enzymes [46]. This approach has been successful at reducing the growth of MDR cells. BBR can selectively accumulate in the mitochondria due to its amphiphilic structure and positive charge [47,48,49]. It arrests the cell cycle and causes apoptosis of malignant cell lines, possibly by disruption of the mitochondrial membrane potential [50,51,52,53]. Electron microscopy analysis reveals that BBR selectively accumulates in the mitochondria of transformed cells due to increase of mitochondrial membrane potential and respiration [53,54]. The accumulation of BBR reduces the formation of mitochondrial structures, increasing complex I-dependent oxidative stress and mitochondrial permeability without a direct effect on ATPase activity [54].
Although oral administration of BBR has few side effects, there are some limitations in its clinical applications due to low bioavailability as a result of short half-life, poor absorption in the gastrointestinal tract, and high susceptibility to metabolic degradation in the liver and other tissues [55]. These characteristics dramatically reduce the delivery efficiency of BBR to various cellular microenvironments [55]. It is, therefore, desirable to design a BBR complex that is capable of selectively targeting the mitochondria of transformed cells and there are several groups actively working in this area [56,57,58].
BBR has anti-proliferative activities and can be used to inhibit the growth of transformed cells in situations such as neoplasia [59,60,61,62]. As discussed earlier in this review, one of the limitations in the use of BBR in clinical settings involves its poor aqueous solubility and thus, poor absorption by the gastrointestinal tract [55]. The low bioavailability of BBR reduces its ability to reach and penetrate into target tissues [63,64]. Previous studies have shown that BBR-nanoparticles are stable and can be used as prophylaxis for osteoarthritis [65] among other pathologies.
4.2. Effect of berberine on apoptosis and necrosis of transformed cells
BBR has been studied in various types of cancerous cells for its effects and underling mechanisms cell functions, including apoptosis and necrosis. For instance, breast cancer cells, which is the second leading cause of cancer-related death in women and characterized by a malignant proliferation of mammary tissues [66]. Currently, there are many methods used comprehensively and extensively in breast cancer therapy; however, the majority of the patients develop drug resistance and experience severe side effects due to a single target [67]. Therefore, there is a need to discover and develop alternatives that work through multi-targets of cancer cells and used as a single or co-therapeutic agent with other drugs to treat cancers. In this regard, BBR has arisen from the traditional Chinese medicine [68]. BBR is a multi-target drug and has been proven effective in treating many cancers and inflammatory diseases. It binds to diverse molecular targets due to the heterogeneity among different cancers or different subtypes of same cancer. It has been shown in an in vitro study that BBR inhibits the proliferation of MCF-7 breast cancer cells by upregulating p53 and a mitochondria-dependent apoptotic pathway [67]. It also binds directly to several proteins and DNA sequences to exert its anticancer activities, such as salt-inducible kinase 3 (SIK3) [69], a single transmembrane cell surface protein ephrin-B2 [70], histone lysine demethylase 1 (LSD1) [71], DNA, TATA box and ply(A) tails [72,73,74], microRNA-214-3p and secretin [75,76,77]. BBR suppresses proliferation and migration of breast cancer cells through ZO-1 mediated Wnt/β-catenin signaling pathway, induces cell cycle arrest through upregulating p21 and p27, sensitizes chemical agents and overcomes drug resistance via activating AMPLK signaling, and inhibits TPA-induced PKC-α signaling in breast cancer cells [68]. In MCF-7/DOXFluc cells, BBR reverses multi-drug resistance by inhibiting the efflux function of ATP-binding cassette transporters and downregulating their expression [78].
BBR appears to target different pathways in different types of cancers. Glioblastoma is the most common primary tumor in the brain or spinal cord that originates from astrocytes, oligodendrocytes, and mixed cell types [79]. It is one of the most aggressive and deadliest brain cancers, killing millions of people worldwide [80]. Current therapeutics, including surgical removal of tumor, radiotherapy, and chemotherapies, or their combinations, are unable to stop the progression of this disease. The poor prognosis of gliomas is likely attributed to high invasiveness and recurrence and in fact, the invasion of glioma cells into the normal brain tissue and extracranial metastasis are the major clinical challenges [81]. Accordingly, exploring novel alternative therapeutics that can control the aggressive malignant behavior of glioma cells and improve prognosis becomes critical.
As mentioned above, BBR inhibits breast and other types of cancer through multiple pathways by targeting multiple molecular targets. In glioblastoma, BBR induces cell death as determined in U-87 MG and U-251 MG cell lines derived from human malignant glioblastoma [82]. Recent work in our lab shows that BBR internalizes into U-87 MG cells (Figure 4). Fluorescent images show intracellular labelling in the cytoplasm. Also, internalization of BBR appears to be variable, with some cells showing strong fluorescence, while others being weakly fluorescent.
Fluorescence microscopic images acquired at 20X showing internalization of BBR by U-87 MG cells treated with 1 µg/mL BBR for 24 hr.
BBR decreased ATP levels and mitochondrial membrane potential in both glioblastoma cell lines, and inhibited PI3K/Akt/mTOR pathway [82]. BBR inhibits glioma cells by inducing endoplasmic reticulum stress, disrupting mitochondrial membrane potential, downregulating ERK1/2 activity, and thus triggering apoptosis [83,84]. BBR and its derivatives are able to cross blood-brain barrier and target mitochondria as determined in a zebrafish model [85]. TGF-β/SMAD signaling pathway is hyperactivated in gliomas, and promotes tumor cell proliferation, migration, and invasion, leading to poor prognosis [86]. A functional link between BBR and TGF-β/SMAD signaling pathway has been observed in various diseases. For example, BBR can attenuate airway inflammation by down-regulating the TGF-β1/SMAD signaling pathway, which plays a key role in tumor cell proliferation and apoptosis [87]. BBR inhibited the proliferation of human glioma U-87 MG cells and induced apoptosis in the U-87 MG and LN229 cells by downregulating Bcl-2 and upregulating Bax and caspase-3. BBR exerted its effects on the proliferation, migration, invasion, and apoptosis of glioma cells by inhibiting the TGF-β1/SMAD2/3 signaling pathway [83]. Similar results have been reported in another study, showing that BBR reduce cell viability of U-87 MG cells in a concentration- and time-dependent manner, suppressed proliferation, and increased oxidative stress and early apoptosis [88]. BBR induces apoptosis in human glioblastoma T98G cells via endoplasmic reticulum stress through the elevation of reactive oxygen species and mitochondria -dependent pathway [89]. It has also been shown that BBR promoted the phosphorylation of wild-type p53 (wtp53), increased the expression of p21 protein, reduced cyclin D1 content, and caused G1 phase arrest in U-87 MG cells. BBR also reduced mutant p53 (mutp53) content and caused G2 phase arrest in U250 cells with concurrent decrease in p21, cyclin D1, and cyclin B1 content. The inhibitory effect of BBR on glioma growth was confirmed in a in vivo mouse tumor model [90]. P53 is a well-established tumor suppressor protein, which inhibits tumor cell proliferation and invasion by participating in DNA repair and cell cycle regulation, differentiation, metabolism, autophagy and apoptosis [91,92]. The results of this study indicate that BBR can inhibit the proliferation of glioma cells by interfering with wtp53 and mutp53, and can be a potential drug to treat wild type and mutant p53 related glioma. BBR induces autophagy and impairs the glycolytic capacity in glioblastoma by targeting the AMPK/mTOR/ULK1-pahtway [93]. All these changes reduce the invasiveness of glioma cells and induce cell death.
Treatment of glioblastoma using therapeutic agents such as temozolomide has been hampered by the development of drug resistance. It has been shown that BBR enhances autophagy and apoptosis in temozolomide-resistance cells by increasing sensitivity to temozolomide through ERK1/2 signaling pathways [94]. A similar phenomenon has been reported in the treatment of other cancers with BBR. For instance, BBR has exhibited the ability to overcome multidrug resistance and shown potential in tumor chemotherapy. Co-administration of BBR and cisplatin results in potentiation and sensitization of the breast cancer cells to cisplatin treatment, and BBR increased the extent of DNA damage and apoptotic and genotoxic effects induced by cisplatin [95]. It reported that BBR interferes with DNA replication by binding directly to DNA [96], explaining the ability of BBR to decrease cellular DNA repair and sensitize breast cancer cells to genotoxic cisplatin [97].
In addition to breast cancer and glioblastoma, BBR has shown promising effects on other types of cancers, such as lung, gastric, liver, ovarian, colorectal, cervical, prostate, pancreatic bladder, endometrial, and esophageal cancers. The common mechanisms of action of BBR on cancer involve primarily inhibiting inflammation, suppressing cell proliferation and migration/metastasis, arresting cell cycle, and inducing apoptosis [98].
Recent studies have indicated that BBR induces apoptotic cell death via the activation of capase-9/cytochrome c, the cell cleavage of poly ADP-ribose polymerase (PARP), caspase-7 and caspase-8 proteins, and the upregulation of pro-apoptotic B-cell pymphoma-2 (BCL-2) proteins via p53 [99,100,101]. Cleavage of chromosomal DNA into oligonucleosomal size fragments is an hall mark of cell apoptosis [102]. The induction of DNA fragmentation by BBR in cancer cells promotes p53 expression, leading to the activation of apoptosis [103]. BBR exerts pro-apoptotic effects on multiple cancers, potentially through AMPK, MAPK, and PKB pathways [104]. It is reported that in both MCF-7 and MDA-MB-231 cancer cells, BBR increases the production of reactive oxygen species (ROS), which activated the pro-apoptotic JNK signaling. Phosphorylated JNK triggers mitochondria membrane potential depolarization and downregulates the expression of anti-apoptotic protein Bcl-2, concomitant with upregulation in the expression of pro-apoptotic protein Bax. Downregulation of anti-apoptotic Bcl-2 family protein in parallel with loss of mitochondrial membrane potential results in the increased release of cytochrome c and apoptosis-inducing factor from mitochondria, triggering eventually the caspase-dependent and -independent apoptotic processes [105].
5. Conclusions
BBR is an effective inhibitor of proliferation and metabolism of transformed cells and the use of this drug for targeting of neoplasia shows some promise. Our understanding of BBR’s molecular targets is still developing, but it is known that BBR modifies metabolism by targeting specific electron transport pathways in the mitochondria. In addition, BBR binds to and modifies signaling pathways through transcription factors such as NF-B, leading to alterations in gene expression. Often, these changes can alter the cell cycle or reduce the production of pro-inflammatory mediators and proliferation promoting growth factors. Although BBR crosses the cell membrane and targets specific intracellular pathways, its mode of internalization is still not fully understood and any therapies that rely on BBR will need to examine BBR’s internalization and transport within cells. Once the mechanism of BBR’s internalization is known and the specific receptors as well as internalization pathways are delineated, better drug delivery vehicles can be designed that perhaps target only specific cell types. The other major challenge with the use of BBR as a therapeutic is its poor solubility and bioavailability. This is an area of active research that is not the focus of this review per se, but will need to be considered by the BBR research field moving forward. BBR has significant potential as an important, cost-effective and relatively safe therapeutic for a variety of disease where there is a need for targeting cell metabolism and proliferation.
Author Contributions
Conceptualization, S.B.A., M.K. and Y.W.; writing—original draft preparation, S.B.A., M.K. and Y.W.; writing—review and editing, S.B.A., M.K. and Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Research Council of Canada.
Institutional Review Board Statement
All animal studies were conducted in accordance with the Canadian Council on Animal Care Guidelines and Policies (https://ccac.ca/en/about-the-ccac/) with approval from the Health Science Animal Care and Use Committee for the University of Alberta.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Feng Wang and Ashley Wagner for technical support. We also thank Dr. Frank Wuest, Department of Oncology, University of Alberta for providing the U-87 MG cells.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tang, J., et al., Berberine and Coptidis rhizoma as novel antineoplastic agents: a review of traditional use and biomedical investigations. J Ethnopharmacol, 2009, 126, 5–17. [CrossRef]
- Ye, X., et al., Hepatoprotective effects of Coptidis rhizoma aqueous extract on carbon tetrachloride-induced acute liver hepatotoxicity in rats. J Ethnopharmacol, 2009, 124, 130–136. [CrossRef]
- Q, Z., et al., - [Simultaneous determination of jatrorrhizine, palmatine, berberine, and obacunone in Phellodendri Amurensis Cortex by RP-HPLC]. Zhongguo Zhong Yao Za Zhi, 2010, 35, 2061–2064.
- Germán-Acacio, J.M., D.E. Meza-Sánchez, and D. Morales-Morales, Chapter 3 - Therapeutically relevant natural products as AMPK activators in the treatment of diabetes, in Studies in Natural Products Chemistry, R. Atta ur, Editor. 2020, Elsevier. 57-90.
- Neag, M.A., et al., Berberine: Botanical Occurrence, Traditional Uses, Extraction Methods, and Relevance in Cardiovascular, Metabolic, Hepatic, and Renal Disorders. Front Pharmacol, 2018, 9, 557. [CrossRef] [PubMed]
- Berberine, "Bureau CMMICoNMM" in Handbook of Effective Compositions in Plants. Vol. 1. 1991: Beijing: People’s Medical Publishing House. pp. 12-18.
- Gu, Y., et al., Effect of traditional Chinese medicine berberine on type 2 diabetes based on comprehensive metabonomics. Talanta, 2010, 81, 766–772. [CrossRef]
- Zeng, X.H., X.J. Zeng, and Y.Y. Li, Efficacy and safety of berberine for congestive heart failure secondary to ischemic or idiopathic dilated cardiomyopathy. Am J Cardiol, 2003, 92, 173–176. [CrossRef]
- Jagetia, G.C. and M.S. Baliga, Effect of Alstonia scholaris in enhancing the anticancer activity of berberine in the Ehrlich ascites carcinoma-bearing mice. J Med Food, 2004, 7, 235–244. [CrossRef] [PubMed]
- Zhang, Q., et al., Preventive effect of Coptis chinensis and berberine on intestinal injury in rats challenged with lipopolysaccharides. Food Chem Toxicol, 2011, 49, 61–69. [CrossRef] [PubMed]
- Singh, S.P. and K.V. Sashidhara, Lipid lowering agents of natural origin: An account of some promising chemotypes. Eur J Med Chem, 2017, 140, 331–348. [CrossRef] [PubMed]
- Pirillo, A. and A.L. Catapano, Berberine, a plant alkaloid with lipid- and glucose-lowering properties: From in vitro evidence to clinical studies. Atherosclerosis, 2015, 243, 449–461. [CrossRef]
- Zhang, H., et al., Berberine lowers blood glucose in type 2 diabetes mellitus patients through increasing insulin receptor expression. Metabolism, 2010, 59, 285–292. [CrossRef]
- Li, X., et al., Berberine hydrochloride IL-8 dependently inhibits invasion and IL-8-independently promotes cell apoptosis in MDA-MB-231 cells. Oncol Rep, 2014, 32, 2777–2788. [CrossRef]
- Li, Z., et al., Antioxidant and Anti-Inflammatory Activities of Berberine in the Treatment of Diabetes Mellitus. Evidence-Based Complementary and Alternative Medicine, 2014, 2014, 289264.
- Zhang, Y., et al., Berberine hydrochloride prevents postsurgery intestinal adhesion and inflammation in rats. J Pharmacol Exp Ther, 2014, 349, 417–426. [CrossRef]
- Wang, Y., et al., Berberine inhibits free fatty acid and LPS-induced inflammation via modulating ER stress response in macrophages and hepatocytes. PLoS One, 2020, 15, e0232630.
- Habtemariam, S., Berberine and inflammatory bowel disease: A concise review. Pharmacol Res, 2016, 113 Pt A, 592–599. [CrossRef]
- Wang, Q., et al., Effect of berberine on proinflammatory cytokine production by ARPE-19 cells following stimulation with tumor necrosis factor-α. Invest Ophthalmol Vis Sci, 2012, 53, 2395–2402. [CrossRef] [PubMed]
- Ehteshamfar, S.-M., et al., Anti-inflammatory and immune-modulatory impacts of berberine on activation of autoreactive T cells in autoimmune inflammation. Journal of Cellular and Molecular Medicine, 2020, 24, 13573–13588. [CrossRef] [PubMed]
- Lu, Z., et al., Anti-inflammatory activity of berberine in non-alcoholic fatty liver disease via the Angptl2 pathway. BMC Immunology, 2020, 21, 28. [CrossRef]
- Battu, S.K., et al., Physicochemical Characterization of Berberine Chloride: A Perspective in the Development of a Solution Dosage Form for Oral Delivery. AAPS PharmSciTech, 2010, 11, 1466–1475. [CrossRef]
- Lu, Y.-C., et al., Solubility of Berberine Chloride in Various Solvents. Journal of Chemical & Engineering Data, 2006, 51, 642–644.
- Chen, W., et al., Bioavailability Study of Berberine and the Enhancing Effects of TPGS on Intestinal Absorption in Rats. AAPS PharmSciTech, 2011, 12, 705–711. [CrossRef]
- Feng, X., et al., Pharmacokinetics and Excretion of Berberine and Its Nine Metabolites in Rats. Frontiers in Pharmacology, 2021, 11. [CrossRef]
- Xue, M., et al., Characterization, pharmacokinetics, and hypoglycemic effect of berberine loaded solid lipid nanoparticles. Int J Nanomedicine, 2013, 8, 4677–4687. [CrossRef]
- Hua, W., et al., Determination of berberine in human plasma by liquid chromatography-electrospray ionization-mass spectrometry. J Pharm Biomed Anal, 2007, 44, 931–937. [CrossRef] [PubMed]
- Alolga, R., et al., Significant pharmacokinetic differences of berberine are attributable to variations in gut microbiota between Africans and Chinese. Scientific Reports, 2016, 6, 27671. [CrossRef]
- Wang, Y. and J.A. Zidichouski, Update on the Benefits and Mechanisms of Action of the Bioactive Vegetal Alkaloid Berberine on Lipid Metabolism and Homeostasis. Cholesterol, 2018, 2018, 7173920. [CrossRef] [PubMed]
- Liu, Y.-T., et al., Extensive Intestinal First-Pass Elimination and Predominant Hepatic Distribution of Berberine Explain Its Low Plasma Levels in Rats. Drug Metabolism and Disposition, 2010, 38, 1779–1784. [CrossRef]
- Liu, C.S., et al., Research progress on berberine with a special focus on its oral bioavailability. Fitoterapia, 2016, 109, 274–282. [CrossRef]
- Battu, S.K., et al., Physicochemical Characterization of Berberine Chloride: A Perspective in the Development of a Solution Dosage Form for Oral Delivery. AAPS PharmSciTech, 2010, 11, 1466–1475. [CrossRef] [PubMed]
- Guo, Y., et al., CYP2D plays a major role in berberine metabolism in liver of mice and humans. Xenobiotica, 2011, 41, 996–1005. [CrossRef]
- Li, Y., et al., Bioactivities of berberine metabolites after transformation through CYP450 isoenzymes. Journal of Translational Medicine, 2011, 9, 62. [CrossRef]
- Qiu, F., et al., Isolation and identification of urinary metabolites of berberine in rats and humans. Drug Metab Dispos, 2008, 36, 2159–2165. [CrossRef]
- Chen, C.M. and H.C. Chang, Determination of berberine in plasma, urine and bile by high-performance liquid chromatography. J Chromatogr B Biomed Appl, 1995, 665, 117–123. [CrossRef] [PubMed]
- Zhang, Y., et al., Treatment of type 2 diabetes and dyslipidemia with the natural plant alkaloid berberine. J Clin Endocrinol Metab, 2008, 93, 2559–2565. [CrossRef]
- Xu, H.-Y., et al., Nanoemulsion improves hypoglycemic efficacy of berberine by overcoming its gastrointestinal challenge. Colloids and Surfaces B: Biointerfaces, 2019, 181, 927–934. [CrossRef]
- Gui, S., et al., Preparation and evaluation of a microemulsion for oral delivery of berberine. Die Pharmazie, 2008, 63, 516–519. [CrossRef]
- Alam, S.B. and M. Kulka, Internalization of benzylisoquinoline alkaloids by resting and activated bone marrow-derived mast cells utilizes energy-dependent mechanisms. Inflamm Res, 2022, 71, 343–356. [CrossRef]
- Cardarelli, F., et al., The intracellular trafficking mechanism of Lipofectamine-based transfection reagents and its implication for gene delivery. Sci Rep, 2016, 6, 25879. [CrossRef] [PubMed]
- Chen, C., et al., Organic anion-transporting polypeptides contribute to the hepatic uptake of berberine. Xenobiotica, 2015, 45, 1138–1146. [CrossRef] [PubMed]
- Sakai, K., et al., Characterization of berberine transport into Coptis japonica cells and the involvement of ABC protein. J Exp Bot, 2002, 53, 1879–1886. [CrossRef]
- Alam, S.B. and M. Kulka, Is Clathrin Involved in Internalization of The Small Molecule Berberine by Primary Mouse Mast Cells?. Archives of Microbiology and Immunology, 2022, 6, 141–148. [CrossRef]
- Chen, X.W., et al., Interaction of herbal compounds with biological targets: a case study with berberine. Scientific World Journal, 2012, 2012, 708292. [CrossRef]
- Jeena, M.T., et al., Recent Progress in Mitochondria-Targeted Drug and Drug-Free Agents for Cancer Therapy. Cancers (Basel), 2019, 12. [CrossRef]
- Chen, H., et al., Mitochondria-targeting fluorescent molecules for high efficiency cancer growth inhibition and imaging. Chem Sci, 2019, 10, 7946–7951. [CrossRef]
- Jean, S.R., et al., Peptide-Mediated Delivery of Chemical Probes and Therapeutics to Mitochondria. Acc Chem Res, 2016, 49, 1893–1902. [CrossRef] [PubMed]
- Ruan, L., et al., Thermoresponsive drug delivery to mitochondria in vivo. Chem Commun (Camb), 2019, 55, 14645–14648. [PubMed]
- Pereira, G.C., et al., Mitochondrially targeted effects of berberine [Natural Yellow 18, 5,6-dihydro-9,10-dimethoxybenzo(g)-1,3-benzodioxolo(5,6-a) quinolizinium] on K1735-M2 mouse melanoma cells: comparison with direct effects on isolated mitochondrial fractions. J Pharmacol Exp Ther, 2007, 323, 636–649. [CrossRef]
- Zhao, H.L., et al., Sustained antidiabetic effects of a berberine-containing Chinese herbal medicine through regulation of hepatic gene expression. Diabetes, 2012, 61, 933–943. [CrossRef]
- Lin, C.C., et al., Berberine induces apoptosis in human HSC-3 oral cancer cells via simultaneous activation of the death receptor-mediated and mitochondrial pathway. Anticancer Res, 2007, 27, 3371–3378.
- Peng, L, et al., Synergistic tumor-killing effect of radiation and berberine combined treatment in lung cancer: the contribution of autophagic cell death. Int J Radiat Oncol Biol Phys, 2008, 70, 529–542. [CrossRef]
- Turner, N., et al., Berberine and its more biologically available derivative, dihydroberberine, inhibit mitochondrial respiratory complex I: a mechanism for the action of berberine to activate AMP-activated protein kinase and improve insulin action. Diabetes, 2008, 57, 1414–1418. [CrossRef] [PubMed]
- Mirhadi, E., M. Rezaee, and B. Malaekeh-Nikouei, Nano strategies for berberine delivery, a natural alkaloid of Berberis. Biomed Pharmacother, 2018, 104, 465–473. [PubMed]
- Ravera, S., et al., Berberine affects mitochondrial activity and cell growth of leukemic cells from chronic lymphocytic leukemia patients. Sci Rep, 2020, 10, 16519. [CrossRef]
- Yan, X.J., et al., Mitochondria play an important role in the cell proliferation suppressing activity of berberine. Sci Rep, 2017, 7, 41712. [CrossRef] [PubMed]
- Shi, X., et al., Anticancer Effects of Honokiol via Mitochondrial Dysfunction Are Strongly Enhanced by the Mitochondria-Targeting Carrier Berberine. J Med Chem, 2020, 63, 11786–11800. [CrossRef] [PubMed]
- Khemani, R.G., et al., Comparison of SpO2 to PaO2 based markers of lung disease severity for children with acute lung injury. Crit Care Med, 2012, 40, 1309–1316. [CrossRef] [PubMed]
- Park, J.J., et al., Berberine inhibits human colon cancer cell migration via AMP-activated protein kinase-mediated downregulation of integrin beta1 signaling. Biochem Biophys Res Commun, 2012, 426, 461–467. [CrossRef]
- Ho, Y.T., et al., Berberine suppresses in vitro migration and invasion of human SCC-4 tongue squamous cancer cells through the inhibitions of FAK, IKK, NF-kappaB, u-PA and MMP-2 and -9. Cancer Lett, 2009, 279, 155–162. [CrossRef]
- Zhao, Y., et al., Berberine inhibits the chemotherapy-induced repopulation by suppressing the arachidonic acid metabolic pathway and phosphorylation of FAK in ovarian cancer. Cell Prolif, 2017, 50. [CrossRef]
- Esposito, C.L., Kirilov, and V.G. Roullin, Organogels, promising drug delivery systems: an update of state-of-the-art and recent applications. J Control Release, 2018, 271, 1–20. [CrossRef]
- Mayr, J., C. Saldias, and D. Diaz Diaz, Release of small bioactive molecules from physical gels. Chem Soc Rev, 2018, 47, 1484–1515. [CrossRef] [PubMed]
- Zhou, Y., et al., In vivo anti-apoptosis activity of novel berberine-loaded chitosan nanoparticles effectively ameliorates osteoarthritis. Int Immunopharmacol, 2015, 28, 34–43. [CrossRef] [PubMed]
- Siegel, R.L., et al., Cancer statistics, 2022. CA: A Cancer Journal for Clinicians, 2022, 72, 7–33. [CrossRef]
- Patil, J.B., J. Kim, and G. K. Jayaprakasha, Berberine induces apoptosis in breast cancer cells (MCF-7) through mitochondrial-dependent pathway. European Journal of Pharmacology, 2010, 645, 70–78. [CrossRef]
- Zhong, X.-D., et al., Berberine as a potential agent for breast cancer therapy. Frontiers in Oncology, 2022, 12. [CrossRef]
- Ponnusamy, L., G. Kothandan, and R. Manoharan, Berberine and Emodin abrogates breast cancer growth and facilitates apoptosis through inactivation of SIK3-induced mTOR and Akt signaling pathway. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease, 2020, 1866, 165897. [CrossRef] [PubMed]
- Ma, W., et al., Berberine inhibits the proliferation and migration of breast cancer ZR-75-30 cells by targeting Ephrin-B2. Phytomedicine, 2017, 25, 45–51. [CrossRef]
- Fu, D.-J., J. Li, and B. Yu, Annual review of LSD1/KDM1A inhibitors in 2020. European Journal of Medicinal Chemistry, 2021, 214, 113254. [CrossRef]
- Bhadra, K. and G.S. Kumar, Therapeutic potential of nucleic acid-binding isoquinoline alkaloids: Binding aspects and implications for drug design. Medicinal Research Reviews, 2011, 31, 821–862. [CrossRef]
- Parra-Marín, O., et al., The highly diverse TATA box-binding proteins among protists: A review. Molecular and Biochemical Parasitology, 2020, 239, 111312. [CrossRef]
- Yuan, Z.-Y., et al., TATA boxes in gene transcription and poly (A) tails in mRNA stability: New perspective on the effects of berberine. Scientific Reports, 2015, 5, 18326. [CrossRef] [PubMed]
- Vander Heiden, M.G., L.C. Cantley, and C.B. Thompson, Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science, 2009, 324, 1029–1033. [CrossRef]
- Kang, S., et al., SCTR regulates cell cycle-related genes toward anti-proliferation in normal breast cells while having pro-proliferation activity in breast cancer cells. Int J Oncol, 2015, 47, 1923–1931. [CrossRef] [PubMed]
- Han, L.C., et al., Effect miR-214-3p on proliferation and apoptosis of breast cancer cells by targeting survivin protein. European review for medical and pharmacological sciences, 2019, 23, 7469–7474. [CrossRef] [PubMed]
- Qian, K., et al., Berberine Reverses Breast Cancer Multidrug Resistance Based on Fluorescence Pharmacokinetics In Vitro and In Vivo. ACS Omega, 2021, 6, 10645–10654. [CrossRef]
- Roy, L.-O., M.-B. Poirier, and D. Fortin, Chloroquine inhibits the malignant phenotype of glioblastoma partially by suppressing TGF-beta. Investigational New Drugs, 2015, 33, 1020–1031. [CrossRef]
- SORDILLO, L.A., P. SORDILLO, and L. HELSON, Curcumin for the Treatment of Glioblastoma. Anticancer Research, 2015, 35, 6373–6378.
- Lin, T.-H., et al., Berberine enhances inhibition of glioma tumor cell migration and invasiveness mediated by arsenic trioxide. BMC Cancer, 2008, 8, 58. [CrossRef]
- Maiti, P., A. Plemmons, and G. Dunbar, Combination treatment of berberine and solid lipid curcumin particles increased cell death and inhibited PI3K/Akt/mTOR pathway of human cultured glioblastoma cells more effectively than did individual treatments. PLOS ONE, 2019, 14, e0225660. [CrossRef]
- Jin, Y., et al., Berberine Suppressed the Progression of Human Glioma Cells by Inhibiting the TGF-β1/SMAD2/3 Signaling Pathway. Integrative Cancer Therapies, 2022, 21, 15347354221130303. [CrossRef]
- Asemi, Z., et al., Therapeutic Potential of Berberine in the Treatment of Glioma: Insights into Its Regulatory Mechanisms. Cellular and Molecular Neurobiology, 2021, 41, 1195–1201. [CrossRef] [PubMed]
- Wang, L., et al., Treatment of Parkinson’s disease in Zebrafish model with a berberine derivative capable of crossing blood brain barrier, targeting mitochondria, and convenient for bioimaging experiments. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2021, 249, 109151. [CrossRef]
- Jiang, L., et al., Overexpression of SMC4 activates TGFβ/Smad signaling and promotes aggressive phenotype in glioma cells. Oncogenesis, 2017, 6, e301–e301. [CrossRef] [PubMed]
- Wang, W., et al., Berberine Attenuates Cigarette Smoke Extract-induced Airway Inflammation in Mice: Involvement of TGF-β1/Smads Signaling Pathway. Current Medical Science, 2019, 39, 748–753. [CrossRef] [PubMed]
- Palma, T.V., et al., Berberine induces apoptosis in glioblastoma multiforme U87MG cells via oxidative stress and independent of AMPK activity. Molecular Biology Reports, 2020, 47, 4393–4400. [CrossRef]
- Eom, K.S., et al., Berberine-Induced Apoptosis in Human Glioblastoma T98G Cells Is Mediated by Endoplasmic Reticulum Stress Accompanying Reactive Oxygen Species and Mitochondrial Dysfunction. Biological and Pharmaceutical Bulletin, 2010, 33, 1644–1649. [CrossRef]
- Z, L., et al., - Berberine Inhibits Cell Proliferation by Interfering with Wild-Type and Mutant P53 in Human Glioma Cells. Onco Targets Ther, 2020, 13, 12151–12162. [CrossRef]
- Vousden, K.H. and C. Prives, Blinded by the Light: The Growing Complexity of p53. Cell, 2009, 137, 413–431. [CrossRef] [PubMed]
- Li, Y., et al., Functional Diversity of p53 in Human and Wild Animals. Frontiers in Endocrinology, 2019, 10. [CrossRef]
- Wang, J., et al., Berberine induces autophagy in glioblastoma by targeting the AMPK/mTOR/ULK1-pathway. Oncotarget, 2016, 7, 66944–66958. [CrossRef]
- Qu, H., et al., Berberine reduces temozolomide resistance by inducing autophagy via the ERK1/2 signaling pathway in glioblastoma. Cancer Cell International, 2020, 20, 592. [CrossRef] [PubMed]
- Zhao, Y., et al., Berberine in combination with cisplatin suppresses breast cancer cell growth through induction of DNA breaks and caspase-3-dependent apoptosis. Oncol Rep, 2016, 36, 567–572. [CrossRef] [PubMed]
- Liu, Q., et al., Berberine Radiosensitizes Human Esophageal Cancer Cells by Downregulating Homologous Recombination Repair Protein RAD51. PLOS ONE, 2011, 6, e23427. [CrossRef]
- Zhu, Q., et al., Post-translational modifications of proliferating cell nuclear antigen: A key signal integrator for DNA damage response (Review). Oncol Lett, 2014, 7, 1363–1369. [CrossRef] [PubMed]
- RG, X., et al., - Anticancer Effects and Mechanisms of Berberine from Medicinal Herbs: An Update Review. LID - 10.3390/molecules27144523 [doi] LID - 4523. Molecules, 2022, 27. [CrossRef]
- Zhao, Y., et al., Berberine activates caspase-9/cytochrome c-mediated apoptosis to suppress triple-negative breast cancer cells in vitro and in vivo. Biomedicine & Pharmacotherapy, 2017, 95, 18–24. [CrossRef]
- Carneiro, B.A. and W.S. El-Deiry, Targeting apoptosis in cancer therapy. Nature Reviews Clinical Oncology, 2020, 17, 395–417. [CrossRef] [PubMed]
- El Khalki, L., et al., Berberine Impairs the Survival of Triple Negative Breast Cancer Cells: Cellular and Molecular Analyses. Molecules, 2020, 25, 506. [CrossRef]
- Zhang, J.H. and M. Xu, DNA fragmentation in apoptosis. Cell Research, 2000, 10, 205–211. [CrossRef]
- Hafner, A., et al., The multiple mechanisms that regulate p53 activity and cell fate. Nature Reviews Molecular Cell Biology, 2019, 20, 199–210. [CrossRef] [PubMed]
- Zhu, Y., et al., Apoptosis Induction, a Sharp Edge of Berberine to Exert Anti-Cancer Effects, Focus on Breast, Lung, and Liver Cancer. Frontiers in Pharmacology, 2022, 13. [CrossRef]
- Xie, J., et al., Berberine-induced apoptosis in human breast cancer cells is mediated by reactive oxygen species generation and mitochondrial-related apoptotic pathway. Tumor Biology, 2015, 36, 1279–1288. [CrossRef] [PubMed]
Figure 1.
Structure of berberine.
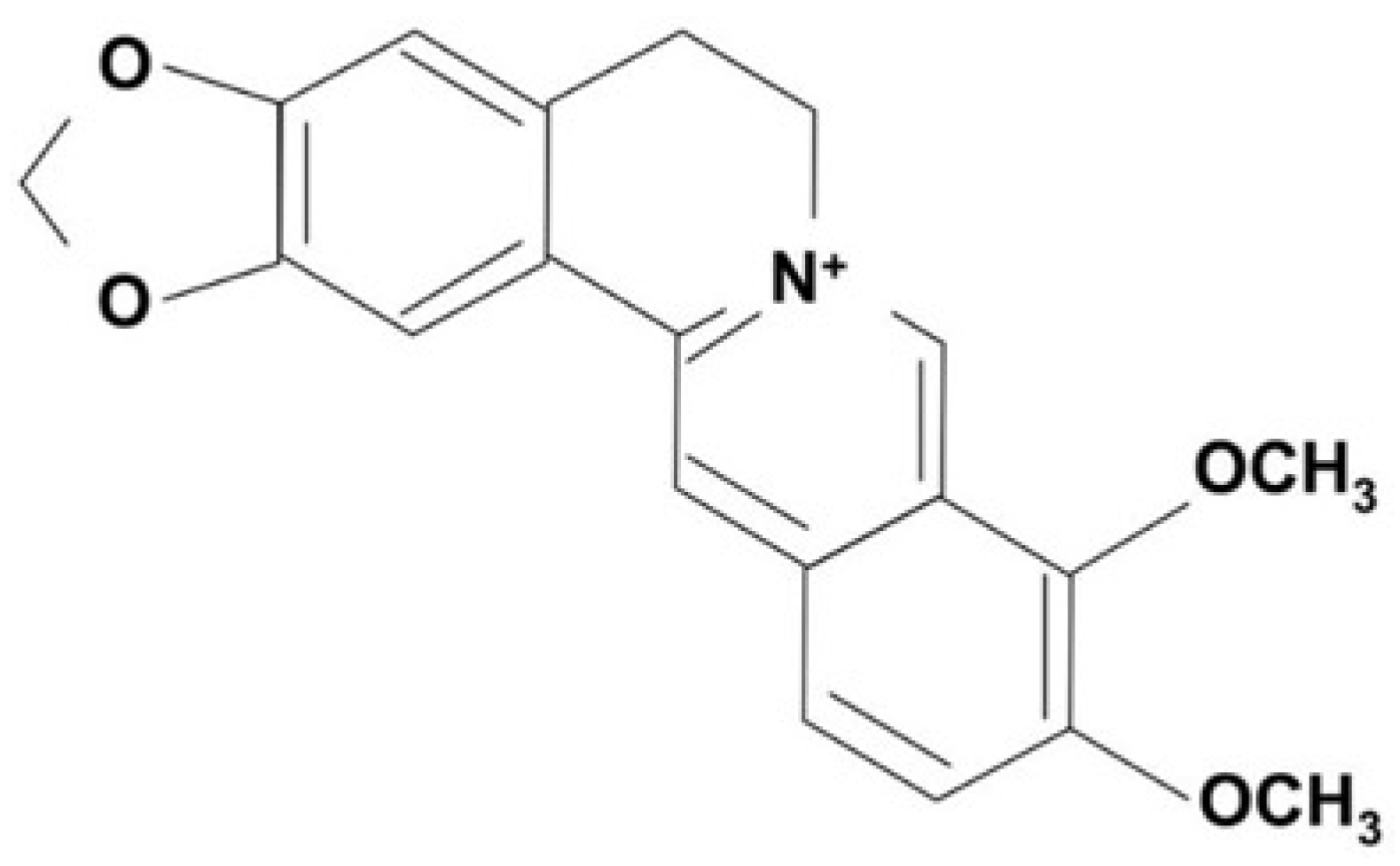
Figure 2.
BBR is internalized into BMMC. (A) Fluorescence microscopic images acquired at 20X showing internalization of BBR by BMMC treated with 10 µg/mL BBR. (B) Flow cytometric analysis of BMMC treated with BBR. The red histogram represents Ut (untreated) cells and the blue histogram represents cells treated with 10 µg/ml BBR. Gate “G1” was used to determine % positive cells compared to Ut cells. (C) Percent of G1 cells that are fluorescent after treatment with different concentrations of BBR (0-100 µg/ml) (n=4).
Figure 2.
BBR is internalized into BMMC. (A) Fluorescence microscopic images acquired at 20X showing internalization of BBR by BMMC treated with 10 µg/mL BBR. (B) Flow cytometric analysis of BMMC treated with BBR. The red histogram represents Ut (untreated) cells and the blue histogram represents cells treated with 10 µg/ml BBR. Gate “G1” was used to determine % positive cells compared to Ut cells. (C) Percent of G1 cells that are fluorescent after treatment with different concentrations of BBR (0-100 µg/ml) (n=4).
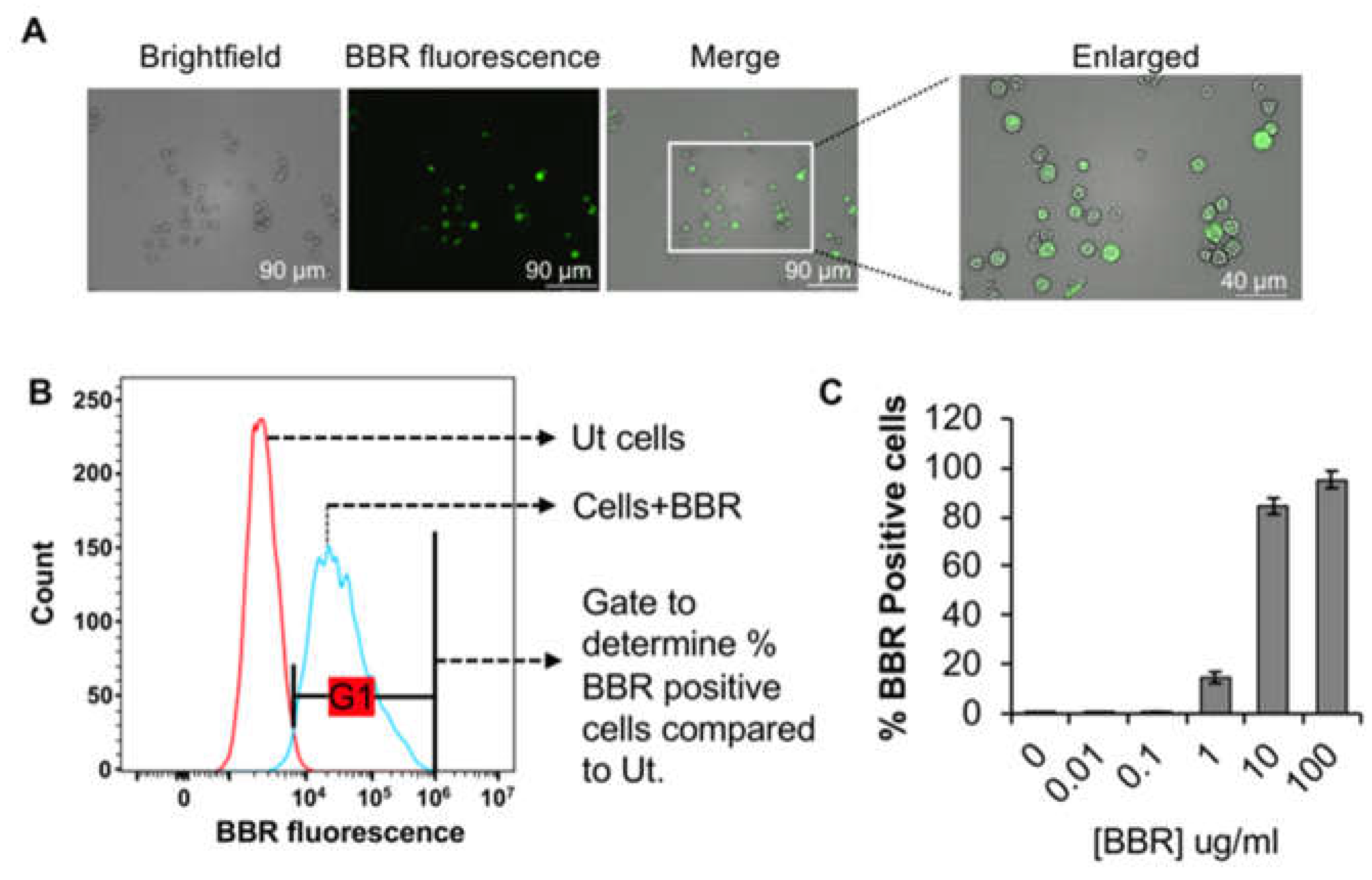
Figure 3.
Possible endocytic routes of BBR in BMMC: BBR endocytosis might be partially dependent upon clathrin (1). BBR endocytosis is drastically reduced by the phosphoinositide 3-kinase (PI3K) inhibitor 3-methyladenine, that plays a prominent role in fusion of macropinosomes with the plasma membrane. Hence, it is possible that majority of BBR might utilize a clathrin independent (2) pinocytic pathway to internalize into BMMC (3). Methanol fixation that removes cholesterol, the integral structural component of caveole, substantially reduces BBR internalization into BMMC. Therefore, involvement of caveolin (4) in BBR’s endocytosis might be a possibility and requires further investigation. IL-3 treatment significantly enhanced BBR endocytosis in BMMC (5). We speculate that IL-3 might also: (5a) activate a specific pinocytic pathway and that in turn enhances BBR endocytosis or (5b) promotes the IL-3 receptor’s clathrin-mediated endocytosis that could perhaps be utilized as a trojan horse by BBR to internalize into BMMC or (5c) engage partially quiescent BMMC into the proliferative stages of the cell cycle. Regardless of the mode of internalization, BBR endocytosis is dependent on actin cytoskeleton (shown in red), that also plays a role in vesicular transport inside the cell. Once internalized into early endosomes (EE) by utilizing a potential internalization pathway, the EE ripens into late endosomes (LE, 6) that ultimately matures to endolysosomes (EL, 7) via further lowering of the pH. The EL and the autophagosomes (AP) fuse to form the autolysosomal compartment (AL), where PI3K plays an important role (8). BBR could also be internalized via interacting with a cell surface receptor shown in brown. Therefore, internalization of BBR might be inhibited when it is incorporated into lipoplexes (9).
Figure 3.
Possible endocytic routes of BBR in BMMC: BBR endocytosis might be partially dependent upon clathrin (1). BBR endocytosis is drastically reduced by the phosphoinositide 3-kinase (PI3K) inhibitor 3-methyladenine, that plays a prominent role in fusion of macropinosomes with the plasma membrane. Hence, it is possible that majority of BBR might utilize a clathrin independent (2) pinocytic pathway to internalize into BMMC (3). Methanol fixation that removes cholesterol, the integral structural component of caveole, substantially reduces BBR internalization into BMMC. Therefore, involvement of caveolin (4) in BBR’s endocytosis might be a possibility and requires further investigation. IL-3 treatment significantly enhanced BBR endocytosis in BMMC (5). We speculate that IL-3 might also: (5a) activate a specific pinocytic pathway and that in turn enhances BBR endocytosis or (5b) promotes the IL-3 receptor’s clathrin-mediated endocytosis that could perhaps be utilized as a trojan horse by BBR to internalize into BMMC or (5c) engage partially quiescent BMMC into the proliferative stages of the cell cycle. Regardless of the mode of internalization, BBR endocytosis is dependent on actin cytoskeleton (shown in red), that also plays a role in vesicular transport inside the cell. Once internalized into early endosomes (EE) by utilizing a potential internalization pathway, the EE ripens into late endosomes (LE, 6) that ultimately matures to endolysosomes (EL, 7) via further lowering of the pH. The EL and the autophagosomes (AP) fuse to form the autolysosomal compartment (AL), where PI3K plays an important role (8). BBR could also be internalized via interacting with a cell surface receptor shown in brown. Therefore, internalization of BBR might be inhibited when it is incorporated into lipoplexes (9).
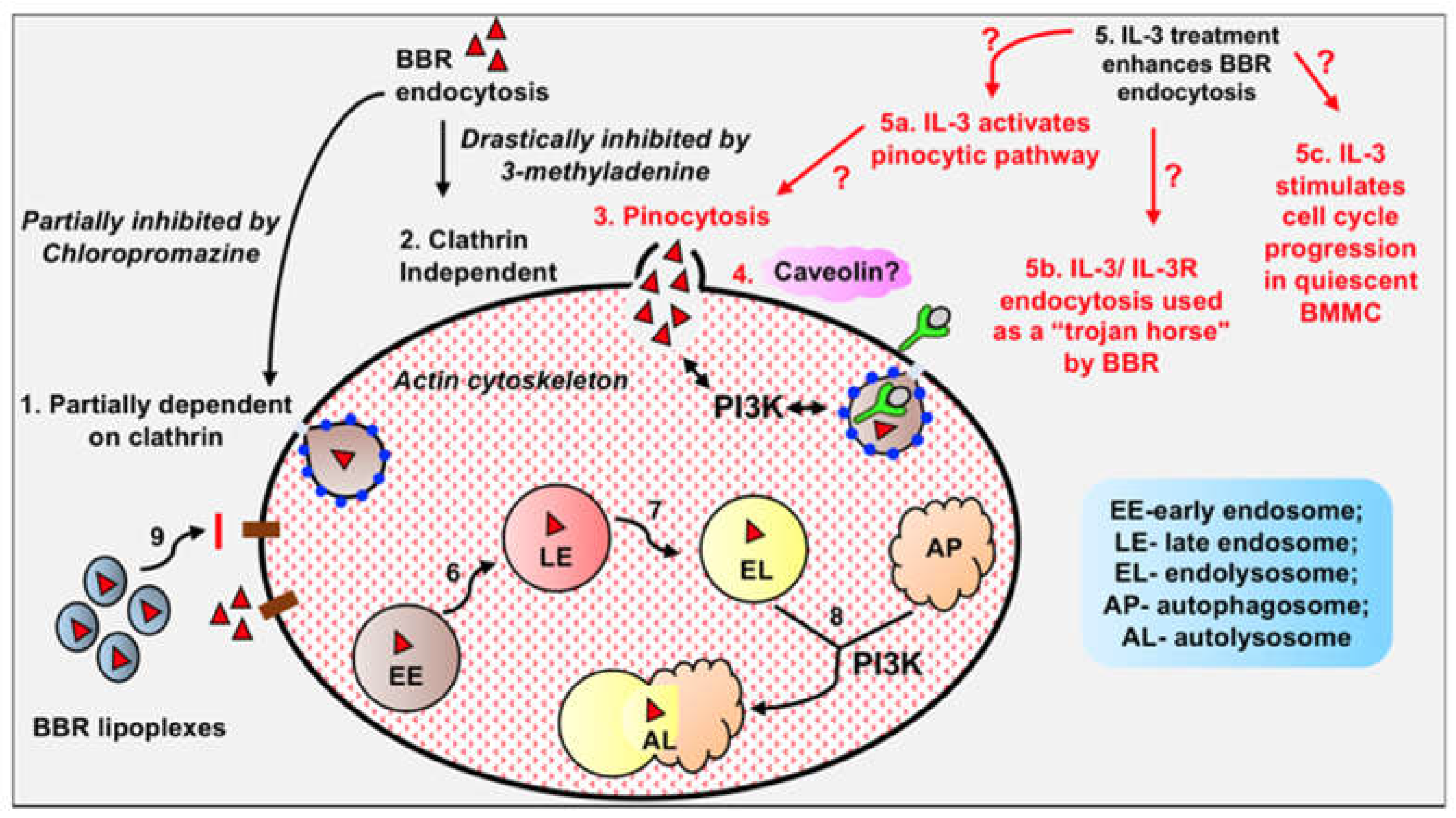
Figure 4.
BBR internalizes into U-87 MG cells. Fluorescence microscopic images acquired at 20X showing internalization of BBR by U-87 MG cells treated with 1 μg/mL BBR for 24 hr.
Figure 4.
BBR internalizes into U-87 MG cells. Fluorescence microscopic images acquired at 20X showing internalization of BBR by U-87 MG cells treated with 1 μg/mL BBR for 24 hr.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



