
Submitted:
14 February 2023
Posted:
16 February 2023
You are already at the latest version
Abstract
To date, 41 species of Gyrodactylus have been described from Africa. However, none of these have been reported in Morocco. After identifying and examining 738 cyprinid host specimens, 26 specimens belonging to Gyrodactylus were found to parasitize the gills of nine species of Luciobarbus, Carasobarbus, and Pterocapoeta. The current study describes in detail 12 specimens of Gyrodactylus isolated from the gills of Luciobarbus pallaryi (Pellegrin, 1919) and Luciobarbus ksibi (Boulenger, 1905). Based on morphoanatomical observations, the characterization of the specimens collected suggests a species of Gyrodactylus new to science, described here as Gyrodactylus nyingiae n. sp. The new species is different from previously described gyrodactylids infecting African cyprinid hosts because it has a longer hamulus total length, a longer hamulus root, a downward projecting toe, a trapezium shaped ventral bar membrane with slightly striated median portion and small rounded anterolateral processes. This study brings the total number of Gyrodactylus spp. found in African cyprinids to four.
Keywords:
Cyprinidae
; Ectoparasite
; Gyrodactylidea
; Luciobarbus
; Monopisthocotylea
; North Africa
; Parasite
; Platyhelminthes
1. Introduction
Morocco is considered to be one of the top producers of fisheries resources occupying the 13th place after Chile [1]. In 2018, the national fisheries production totaled a volume of 1,371,683 tons for a turnover of 11,579,544 thousand dirhams [1]. In freshwater, culture-based fisheries which are projected to generate 13,000 tons of fish annually in Morocco, is the main source of fish protein. This production is based on the routine stocking of cultured organisms, mainly cyprinids, into lakes and reservoirs [2]. Biogeographers in the Maghreb region have often focused on ichthyofaunal studies because of its geographical position between the African and Eurasian plates. The primary freshwater fishes are a suitable subject for historical biogeography due to their limited dispersal that is strictly restricted to fluvial basins showing less capacity for trans-watershed dispersal [3]. However, the freshwater fish fauna of North Africa shows low diversity which could probably reflect a long period of isolation during the Cenozoic Era [3].
The high level of endemism of cyprinid fish in Morocco (20 endemic species) noted by [4] is linked to the geological and climatic history of the Mediterranean biome, which have led to the endemic status of many species (animal or plant) present in these zones [5,6]. Studies on freshwater fish parasites have increased globally due to the growing interest in developing fisheries and aquaculture as cheap sources of protein to sustain the rapidly growing human population, especially in some African communities [7]. For management of this resource, a thorough knowledge of the taxonomy, distribution, biology and ecology of parasites is of paramount importance [7].
The cyprinid host/parasite system constitutes a good model for studying evolutionary phenomena and determining speciation mechanisms. Previous studies [8] on representatives of the monogenean parasite Dactylogyrus Diesing, 1850 recorded 17 Dactylogyrus spp. on 17 cyprinid species from four genera (Luciobarbus Heckel, 1843; Carasobarbus Karaman, 1971; Labeobarbus Rüppel, 1835 and Pterocapoeta Günther, 1902) in Morocco. These studies revealed that these fishes arrived in Morocco through three different routes; from Europe, Asia and Sub-Saharan Africa [4,9]. Despite there being various studies on Gyrodactylus von Nordmann, 1832 in Africa, also on cyprinids, these often do not include species-level identifications (e.g. [10]) and only a few of its representatives have been identified to species level in the whole continent. Currently, there are over 400 valid species of Gyrodactylus described [11]. In African freshwater fishes, only 41 species of Gyrodactylus have been described [12]. The known species represent only a fraction of the expected Gyrodactylus spp. in Africa [13]. Only three Gyrodactylus species have been described from small African cyprinids, always with a host belonging to Enteromius Cope, 1867, namely G. ivindoensis Price & Gery, 1968 from Enteromius cf. holotaenia (Boulenger, 1904) in Gabon, G. kyogae Paperna, 1973 from Enteromius neumayeri (Fischer, 1884) and Enteromius perince (Rüppell, 1835) in Uganda and G. paludinosus Truter, Smit, Malherbe & Přikrylová, 2021 [12] from Enteromius paludinosus (Peters, 1852) in South Africa. In Morocco, no research has documented species of Gyrodactylus to date. Monogeneans belonging to Gyrodactylus are major pathogens in fishes as well as a major challenge in both fisheries and aquaculture. Gyrodactylids have an economic significance that outweighs that of any other monogenean family. In Norway, for example, the introduction of Gyrodactylus salaris Malmberg, 1957 into the salmon industry resulted in uncontrollable epidemics and mortalities, leading to massive economic losses [14].
Despite the numerous economic benefits a country may achieve from introduction of living organisms, they can also be detrimental to the native species [15]. In Lake Naivasha, Kenya, for example, common carp Cyprinus carpio Linnaeus, 1758 was thought to have reached the lake in 1999 during the heavy rains from juveniles that escaped in the Malewa River [16]. Parasitological studies on parasites of C. carpio in Lake Naivasha discovered it to be dominated by representatives of Dactylogyrus [17] with high prevalence of 99.3% [17]. Moroccan irrigation channels and reservoirs have also been stocked with non-native freshwater fish species such as the silver carp Hypophthalmichthys molitrix (Valenciennes, 1844), common carp Cyprinus carpio and grass carp Ctenopharyngodon idella (Valenciennes, 1844) which could pose a threat to the native fishes by providing a perfect opportunity for parasite transmission [18]. For this reason, it is important to have a baseline for the Moroccan native monogenean fauna of cyprinids. Therefore, and in view of the importance of the cyprinid-monogenean system in investigating the aquatic biodiversity and biogeography of North Africa, the present study aims to identify Gyrodactylus species infecting cyprinids across Morocco and contribute to the parasite species inventory of these fishes. The current study focuses on parasites belonging to Gyrodactylus isolated from the gills of Luciobarbus pallaryi (Pellegrin, 1919) and Luciobarbus ksibi (Boulenger, 1905).
2. Materials and Methods
2.1. Sample Collection
During September 2014 and June 2021, a total of 28 localities covering nine different watersheds in Morocco were sampled on five different occasions for cyprinid specimens as shown in Figure 1. The fish specimens were collected after getting the required permit from the Ministry of Water, Forestry and Desertification Control (sampling permit no: 62 HCEFLCD/DLCDPN/CPC/PPC). These fish samples were collected using a backpack electrofisher (Samus-725G) or gill nets where the physico-chemical water parameters could not allow sampling using the electrofisher. Fish hosts were identified morphologically following [19], euthanized by severing their spinal cords and dissected immediately. The gills were fixed according to [20] and some fish specimens were frozen in a portable freezer and analyzed in the laboratory. The nomenclature and the classification of fishes are those provided in [21]. The map showing sampling localities (Figure 1) was created using QGIS v3.22.8 (QGIS Development Team 2022. QGIS Geographic Information System. Open Source Geospatial Foundation Project. http://qgis.osgeo.org, accessed on 20 January 2023).
2.2. Parasitological Examination
The fish samples were transported to the laboratory for parasitological examination. Monogeneans were isolated under a dissecting microscope (Wild Heerbrugg) from the gills (gill arches from the right side of the excised fish). With the aid of a fine needle, the parasites were picked out one by one and subsequently mounted on a glass slide then covered with a coverslip to ‘flatten’ the specimen. The slides were mounted according to [22]. For worms fixed in ethanol, Hoyer’s chloral hydrate was used [23] while ammonium picrate glycerine was used for frozen parasites [24]. The glass slide is left to dry for 24 hours in a horizontal position before sealing the coverslip with Glyceel [25]. Type material was deposited in the collections of the Research Group Zoology: Biodiversity & Toxicology at Hasselt University (HU) (Diepenbeek, Belgium) (HU 838-841) and the Institut Scientifique of the Mohammed V University in Rabat (Rabat, Morocco) (ZA PPM 0101).
2.3. Identification of Representatives of Gyrodactylus
Gyrodactylus was distinguished from the other monogeneans as its members have a cylindrical body bearing two small cephalic lobes on the exterior part of the body, lack eyes and possess an opisthaptor armed with a single pair of hamuli linked by dorsal and ventral bars with 16 articulated marginal hooks (14 in members of Dactylogyrus, the other monogenean genus most common on Moroccan cyprinids) [26].
2.4. Infection Parameters
2.5. Morphological Characterization of Members of Gyrodactylus
Light microscopy using both phase and differential interference contrast approaches were used to study the shape and dimensions of sclerotized structures, which were viewed under a x100 oil immersion objective on a Leica DM2500 optical microscope using Las X software v3.6.0.20104 fitted with a Leica DMC4500 camera. The whole mount, attachment organ, and male copulatory organ (MCO) (when present) on each specimen were photographed. The haptoral morphometrics (26 point-to-point measurements) followed the measurements proposed by [28]; these were taken using ImageJ v1.53k software (available at http://imagej.nih.gov/ij). These measurements are given in micrometers (µm) as the mean, followed by the range in parentheses and number of structures (n) measured for each metric. The micrographs taken were used to draw taxonomically important structures using Inkscape v1.2.
2.6. Statistical Analyses
For statistical analysis, a Principal Component Analysis (PCA) was carried out in R Studio v4.1.0. The analyses included 19 measurements of the haptoral hard parts of hamuli and marginal hooks only. The MCO, ventral bar and dorsal bar measurements were excluded from the analysis due to the large number of missing data.
3. Results
3.1. Specimens Examined and Individuals of Gyrodactylus Isolated
A total of 738 fish specimens belonging to three genera (Luciobarbus Heckel, 1843, Carasobarbus Karaman, 1871 and Pterocapoeta Günther, 1902) were collected. Thirteen cyprinid fish species were identified and their gills examined for infection with species of Gyrodactylus (Table 1). Nine out of the 13 species were found to be infected with representatives of Gyrodactylus (n=26).
3.2. Infection Parameters
The infection parameters of examined hosts are as shown in Table 2.

Table 2.
Prevalence, mean intensity and mean abundance of Gyrodactylus infecting cyprinid hosts, based on examination of the right-side gill arches only.
Table 2.
Prevalence, mean intensity and mean abundance of Gyrodactylus infecting cyprinid hosts, based on examination of the right-side gill arches only.
| Locality | Species | H | N | n | P=(N/H)*100 | M. I=n/N | M.A=n/H |
|---|---|---|---|---|---|---|---|
| Oued Guir |
Luciobarbus pallaryi |
157 | 1 | 14 | 0.64 | 14 | 0.09 |
| Oued Bouregreg |
Luciobarbus rabatensis |
24 | 1 | 3 | 4.17 | 3 | 0.13 |
| Oued Sebou |
Luciobarbus maghrebensis |
55 | 1 | 1 | 1.82 | 1 | 0.02 |
| Oued Zendoula |
Luciobarbus rifensis |
19 | 1 | 3 | 5.26 | 3 | 0.16 |
| Oued Moulouya |
Luciobarbus zayanensis |
25 | 1 | 1 | 4.00 | 1 | 0.04 |
| Oued Moulouya |
Carasobarbus moulouyensis |
44 | 1 | 1 | 2.27 | 1 | 0.02 |
| Oued Ksob | Luciobarbus ksibi | 40 | 1 | 1 | 2.50 | 1 | 0.03 |
| Oued Souss |
Luciobarbus massaensis |
21 | 1 | 1 | 4.76 | 1 | 0.05 |
| Oued OumEr′Rabia |
Pterocapoeta maroccana |
3 | 1 | 1 | 33.33 | 1 | 0.33 |
H, number of examined hosts; N, number of infected hosts; n, number of individuals of Gyrodactylus in infected host; P, prevalence; M.I, mean infection intensity; M.A, mean abundance.
3.3. Characterization of a New Species of Gyrodactylus
All the isolated flatworms belonging to Gyrodactylus showed the diagnostic features of this genus: gyrodactylid monogeneans with opisthaptor having one pair of haptoral anchors surrounded up by 16 marginal hooks. The measurements are given in Table 3.
Class: Monogenea Van Beneden, 1858
Subclass: Polyonchoinea Bychowsky, 1937
Order: Gyrodactylidea Bychowsky, 1937
Family: Gyrodactylidae Van Beneden & Hesse, 1863
Subfamily: Gyrodactylinae Van Beneden & Hesse, 1863
Genus: Gyrodactylus von Nordmann, 1832
Species: Gyrodactylus nyingiae n. sp.
Type material: holotype (HU_838_IV.1.18) and five paratypes (HU_839_IV.1.19, HU_840_IV.1.20, HU_841_IV.1.21, ZA PPM 0101)
- Type host: Luciobarbus pallaryi (Pellegrin, 1919) (Teleostei: Cyprinidae)
- Other host: Luciobarbus ksibi (Boulenger, 1905) (Teleostei: Cyprinidae)
- Type locality: Oued Guir (31°52′12′′N, 003°0′00′′W) (on type host)
- Other locality: Oued Ksob (31°27′50.7′′N, 009°45′25.3′′W) (on L. ksibi)
- Site of infection: Gill filaments
- ZooBank registration: The Life Science Identifier (LSID) of the article is xxx. The LSID for Gyrodactylus nyingiae Shigoley, Rahmouni, Louizi, Pariselle & Vanhove n. sp. is xxx.
- Studied material: 12 mounted individuals were measured; 11 of these were isolated from L. pallaryi and one from L. ksibi
Etymology: The species epithet honors Dr. Dorothy Wanja Nyingi, ichthyologist at the National Museums of Kenya and author of the first Guide to Common Freshwater Fishes of Kenya
Description: Elongated body. Male copulatory organ (MCO) observed in five specimens, spherical (Figure 2a, i; 3 ii), positioned posterior to the pharynx and armed with one principal spine and single row of 5-6 smaller spines (Figure 2a, i; 3 ii). Hamuli slightly slender with a pointed tip with a superficial root (Figure 2a, ii; 3 i). Anterior end where dorsal bar attaches on hamulus prominent, creating a notch between the root and dorsal bar attachment point. Dorsal bar simple and flexible. Ventral bar with small rounded anterolateral processes; trapezoid-like membrane with a slightly striated median portion (Figure 2a, iii). Marginal hook shaft approximately perpendicular to the base of the marginal hook sickle (Figure 2a, iv; 3 iii). Sickle point is slightly curved and perpendicular to the base with its tip in line with the distal end of the toe. Overlapping measurements (Table 3) and the similarity in the shape of the marginal hook sickle (Figure 4) suggest the worms infecting the two host species are conspecific.
Figure 2.
Gyrodactylus nyingiae n. sp. isolated from Luciobarbus pallaryi. (a) Micrograph of hamuli, male copulatory organs (MCO), ventral bar and marginal hooks. (b) Drawings of sclerotized structures of the haptor with (A); hamuli, (B); ventral bar, (C); marginal hook, and of the (D); male copulatory organ. Scale bars in µm.
Figure 2.
Gyrodactylus nyingiae n. sp. isolated from Luciobarbus pallaryi. (a) Micrograph of hamuli, male copulatory organs (MCO), ventral bar and marginal hooks. (b) Drawings of sclerotized structures of the haptor with (A); hamuli, (B); ventral bar, (C); marginal hook, and of the (D); male copulatory organ. Scale bars in µm.
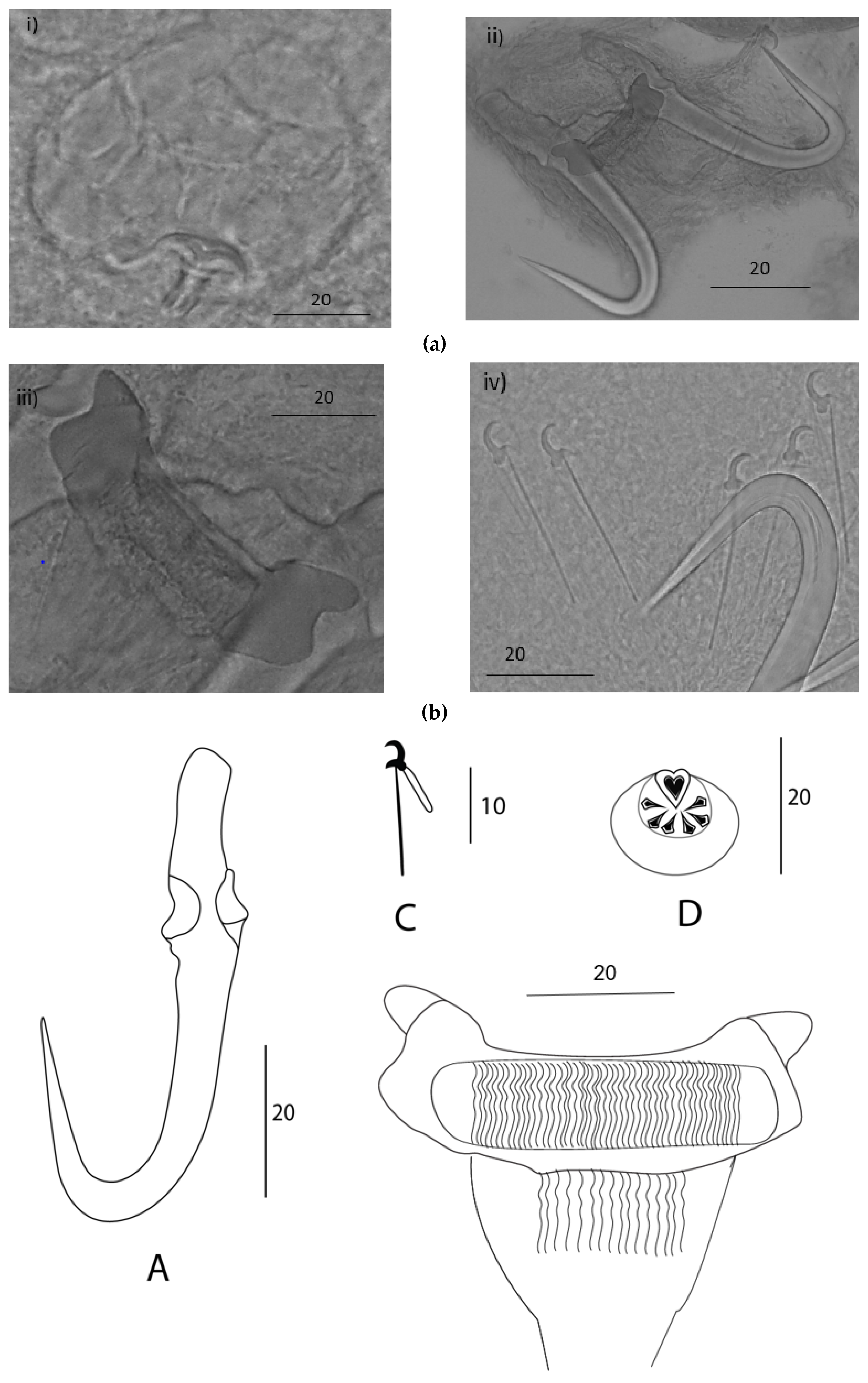
Figure 3.
Micrograph and drawings of sclerotized structures of Gyrodactylus nyingiae n. sp. isolated from Luciobarbus ksibi.
Figure 3.
Micrograph and drawings of sclerotized structures of Gyrodactylus nyingiae n. sp. isolated from Luciobarbus ksibi.
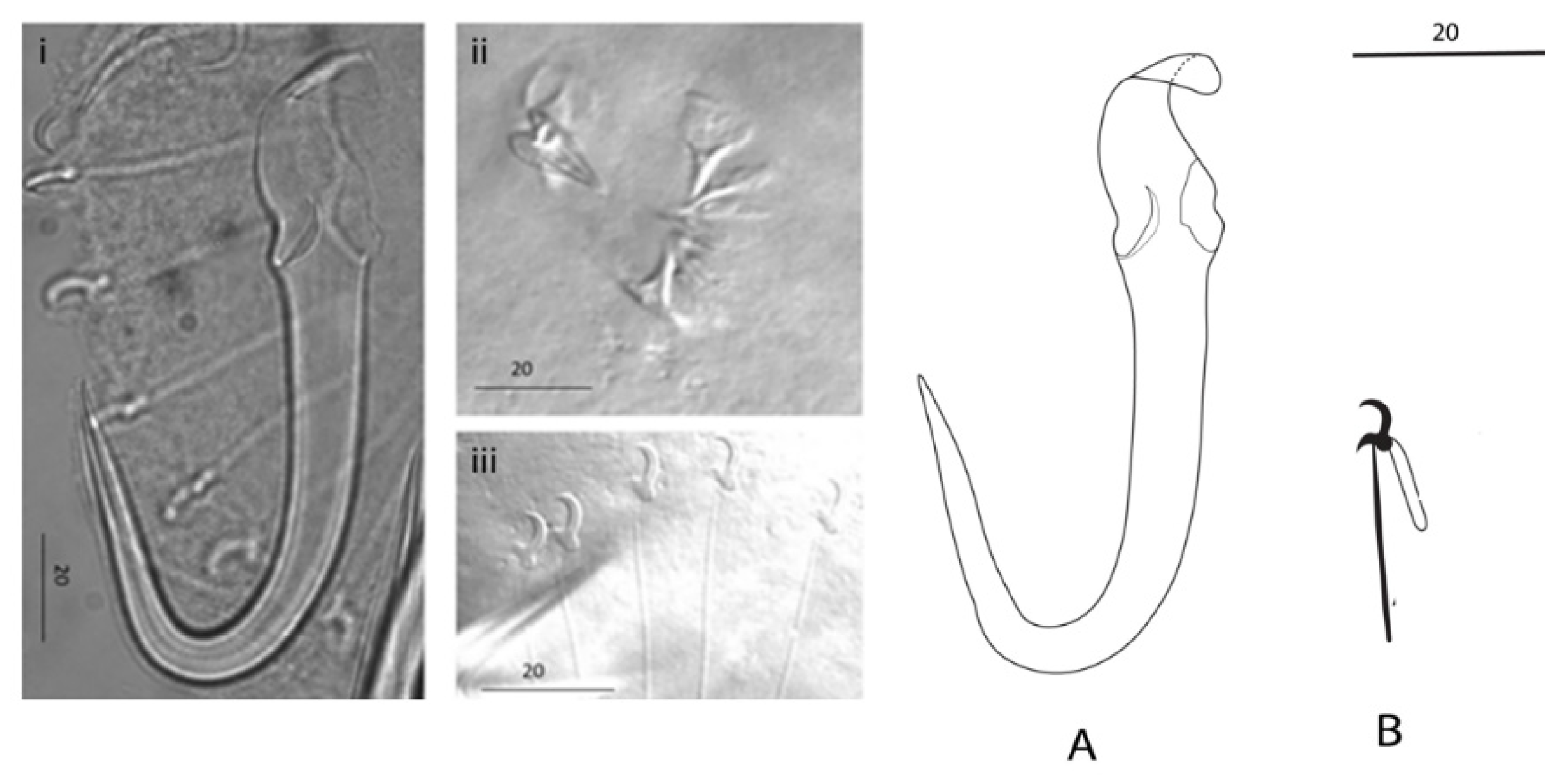
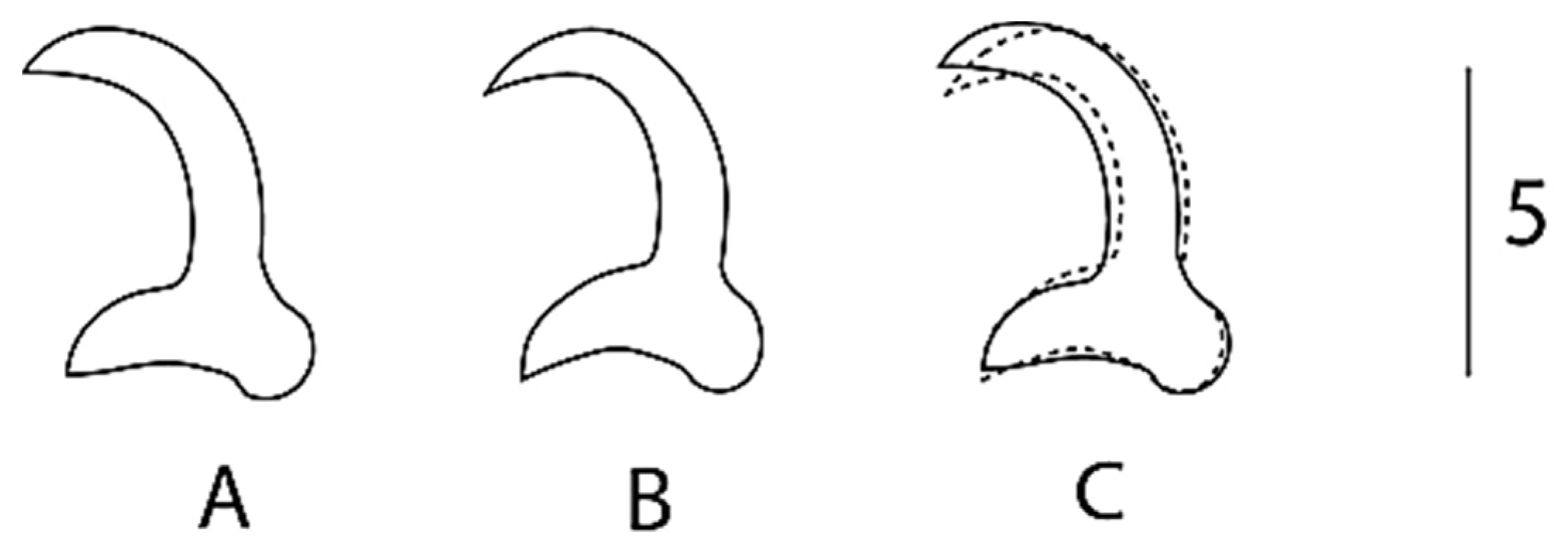
Figure 4.
Overlay of marginal hook sickles (4C) of G. nyingiae n. sp. from L. ksibi (4A) and L. pallaryi (4B) (dotted outline). Scale bar represents 5µm.
Figure 4.
Overlay of marginal hook sickles (4C) of G. nyingiae n. sp. from L. ksibi (4A) and L. pallaryi (4B) (dotted outline). Scale bar represents 5µm.
Remarks:
The comparison with other gyrodactylid species is based on the phenotypic similarities with known parasite species and their occurrence from related hosts. From the three species of Gyrodactylus recorded from cyprinids in Africa, the here newly described species of Gyrodactylus can be differentiated by the longer hamuli: G. nyingiae n. sp. 76.5 (65.9-88.2) compared to a hamulus total length in G. ivindoensis of 55 (52-58), 32.1 (23-33) in G. kyogae and 43.3 (35.1-51.5) in G. paludinosus. Like G. paludinosus, G. kyogae has an upward projecting toe as opposed to that of G. nyingiae n. sp. whose toe points downwards. Also, the MCO of G. nyingiae n. sp. has one principal spine and five to six smaller spines arranged in a single row (Figure 2 and Figure 3) as opposed to G. kyogae which has an unarmed MCO [29]. Gyrodactylus kyogae, in contrast to the other three species, lacks a ventral bar membrane. Gyrodactylus ivindoensis has shorter marginal hooks and a marginal hook total length of 22 (21-24) when compared to G. nyingiae n. sp. 34.8 (31.7-42.1). When comparing the relative length of the root to hamulus total length respectively, G. nyingiae n. sp. (26.7 vs 76.4), G. ivindoensis (19.4 vs 55) and G. paludinosus (15.4 vs 43.3) have similar ratios of the root length and total hamulus length (ca. 1:2.8). Gyrodactylus kyogae (9.2 vs 33.1) on the other hand has a different ratio of the relative root length to hamulus total length (1:3.5).
Due to the important biogeographical connections between the Middle East and Maghreb region during the Cenozoic period in the dispersal of freshwater fish fauna, it is interesting to compare the Gyrodactylus fauna of the Iranian region with the North African ones [3,30]. The freshwater species of Gyrodactylus mentioned by [31] and [32] were either known from Europe or Central Asia, or undescribed. It is therefore instructive to compare G. nyingiae n. sp. with widespread Palearctic species of Gyrodactylus infecting cyprinids, several of which are reminiscent of G. nyingiae n. sp. in marginal hook morphology. This includes Gyrodactylus mutabilitas Bychowsky, 1957 and Gyrodactylus scardiniensis Glaeser, 1974, which can both be distinguished from G. nyingiae n. sp. by virtue of their shorter hamulus root (max. 20 in G. mutabilitas and max. 23 in G. scardiniensis versus min. 24 in G. nyingiae n. sp.), and Gyrodactylus schulmani Ling, 1962 which has a hamulus of a total length of max. 44, shorter than the minimally 66 of G. nyingiae n. sp. A species described from a fish species endemic to Iran is Gyrodactylus jalalii Vanhove, Boeger, Muterezi Bukinga, Volckaert, Huyse & Pariselle, 2012, parasite of the cichlid host Iranocichla hormuzensis Coad, 1982. It can easily be distinguished from G. nyingiae n. sp. by its more pronounced ventral bar auricles and the sub rectangular ventral bar membrane as opposed to G. nyingiae n. sp. having small rounded anterolateral processes and a trapezium shaped ventral bar membrane. Following [33] Gyrodactylus molnari Ergens, 1978 infecting Cyprinus carpio Linnaeus, 1758 in Iraq is smaller in hamuli length (55-65) versus G. nyingiae n. sp. (65.9-88.2). Also, G. molnari has a longer dorsal bar (15-18), compared to G. nyingiae n. sp. (9.9-13.4), and an entirely different shape of marginal hook sickle. Therefore, G. nyingiae n. sp. can be distinguished from the aforementioned Gyrodactylus species by virtue of its longer hamulus total length, longer hamulus root, small rounded anterolateral process and trapezium shaped ventral bar membrane.
3.4. Multivariate Statistics
The morphological variation of the 12 specimens of Gyrodactylus was visualized based on a PCA performed on 19 standardized haptoral morphometric characters. The first two principal component axes contributed 25.9% and 19.6% of the variation respectively (Figure 5).
The biplot shows a single cloud which includes all the 12 specimens belonging to Gyrodactylus. From the PCA biplot we can confirm that we are dealing with a single species described herein as Gyrodactylus nyingiae n. sp.
4. Discussion
Gyrodactylus nyingiae n. sp. is the first described species of Gyrodactylus in Morocco. It is also the first gyrodactylid to be described from Luciobarbus in Africa, as previous studies on gyrodactylids infecting cyprinids in Africa have focused on the small barbs belonging to Enteromius, with only three Gyrodactylus species described so far [12].
In addition to the low prevalence and possible seasonality of members of Gyrodactylus, less research, a lack of reports on infections, and a lack of understanding of relationships between these monogeneans and cyprinid hosts [12], the current study’s low number of gyrodactylids isolated from cyprinid hosts could be due to the fact that only the gills were examined for parasites. However, on native cichlid fishes in Morocco, [34] also found a species depauperate fauna and low abundances of gill-infecting dactylogyrids: hence, we cannot exclude that environmental conditions in Morocco’s freshwater ecosystems limit the species richness and abundance of certain monogenean taxa. On the other hand, Dactylogyrus reaches higher species richness and higher infection intensities on Moroccan cyprinid-monogenean systems [4,35]
5. Conclusions
Based on the morphoanatomical observation of opisthaptoral parts of 12 individuals of Gyrodactylus in the current study, we described a new species infecting two cyprinid hosts for the first time in Morocco. The new species is different from previously described gyrodactylids infecting cyprinid hosts because it has a longer hamulus total length, a longer hamulus root, a downward projecting toe, trapezium shaped ventral bar membrane with slightly striated median portion and small rounded anterolateral processes.
Author Contributions
Conceptualization, M.S. and A.P.; methodology, I.R.; investigation, M.S., H.L. and I.R.; resources, I.R.; data curation, M.S. and M.P.M.V; writing—original draft preparation, M.S.; writing—review and editing, M.S, A.P and M.P.M.V; supervision, A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Special Research Fund of Hasselt University, grant number BOF22DOCLI04 to MIS; BOF20TT06 and BOF21INCENT09 to MPMV and by research grant 1513419N of the Research Foundation – Flanders (FWO-Vlaanderen).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Type material was deposited in the collection of the research group Zoology: Biodiversity and Toxicology of Hasselt University (Diepenbeek, Belgium) (HU 838-841) and the Institut Scientifique of the Mohammed V University in Rabat (Rabat, Morocco) (ZA PPM 0101).
Acknowledgments
The authors would like to sincerely thank Prof. Abdelaziz Benhoussa for allowing us to work in the Biodiversity, Ecology and Genome Laboratory (Mohammed V University, Rabat). Prof. Bouabid Badaoui and Hocein Bazairi are thanked for their statistical advice. Daniel Mungai Ndegwa, Kelly Thys, Tiziana Gobbin and Hervé Mandtoumbi for their laboratory assistance and help with statistical analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dahani, S.; Bouchriti, N.; Benabbes, I.; Boudakkou, A.; Chiaar, A. Occurrence Des Parasites Dans Les Poissons Collectés Au Niveau Du Littoral Marocain. Eur. Sci. J. ESJ. 2019, 15, 497–506. [Google Scholar] [CrossRef]
- Mohammed, B.; Salwa, O.; Yahyaoui, A. Perceived Barriers to Consumption of Freshwater Fish in Morocco. Br. Food J. 2015, 117, 274–285. [Google Scholar] [CrossRef]
- Doadrio, I. Freshwater Fish Fauna of North Africa and Its Biogeography. 1994, 275, 21-34.
- Rahmouni, I.; Řehulková, E.; Pariselle, A.; Berrada Rkhami, O.; Šimková, A. Four New Species of Dactylogyrus Diesing, 1850 (Monogenea: Dactylogyridae) Parasitising the Gills of Northern Moroccan Luciobarbus Heckel (Cyprinidae): Morphological and Molecular Characterisation. Syst. Parasitol. 2017, 94, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Cowling, R.M.; Rundel, P.W.; Lamont, B.B.; Arroyo, M.K.; Arianoutsou, M. Plant Diversity in Mediterranean-Climate Regions. Trends Ecol. Evol. 1996, 11, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Sauquet, H.; Weston, P.H.; Anderson, C.L.; Barker, N.P.; Cantrill, D.J.; Mast, A.R.; Savolainen, V. Contrasted Patterns of Hyperdiversification in Mediterranean Hotspots. Proc. Natl. Acad. Sci. USA 2009, 106, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Scholz, T.; Smit, N.; Jayasundera, Z.; Gelnar, M. A Guide to the Parasites of African Freshwater Fishes; 2018; Vol. 18; ISBN 9789073242388.
- Řehulková, E.; Rahmouni, I.; Pariselle, A.; Šimková, A. Integrating Morphological and Molecular Approaches for Characterizing Four Species of Dactylogyrus (Monogenea: Dactylogyridae) from Moroccan Cyprinids, with Comments on Their Host Specificity and Phylogenetic Relationships. PeerJ. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Šimková, A.; Benovics, M.; Rahmouni, I.; Vukić, J. Host-Specific Dactylogyrus Parasites Revealing New Insights on the Historical Biogeography of Northwest African and Iberian Cyprinid Fish. Parasites and Vectors. 2017, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Allalgua, A.; Guerfi, S.; Kaouachi, N.; Boualleg, C.; Boucenna, I.; Barour, C. L’infestation de Cyprinus carpio (Cyprinides) Peuplant Le Barrage Foum El-Khanga (Souk Ahras, Algerie) Par Les Monogenes Parasites. Bull. la Soc. Zool. Fr. 2015, 140, 217–232. [Google Scholar]
- Bakke; Harris, P.D.; Cable, J. Host Specificity Dynamics: Observations on Gyrodactylid Monogeneans. Int. J. Parasitol. 2002; 32, 281–308. [CrossRef]
- Truter, M.; Smit, N.J.; Malherbe, W.; Přikrylová, I. Description of Gyrodactylus paludinosus sp. nov. (Monogenea: Gyrodactylidae) from the Straightfin Barb, Enteromius paludinosus (Peters, 1852), in South Africa. Acta Parasitol. 2022, 67, 446–453. [Google Scholar] [CrossRef]
- Dos Santos, Q.M.; Maina, J.N.; Avenant-Oldewage, A. Gyrodactylus magadiensis n. sp. (Monogenea, Gyrodactylidae), Parasitising the Gills of Alcolapia grahami (Perciformes, Cichlidae), a Fish Inhabiting the Extreme Environment of Lake Magadi, Kenya. Parasite. 2019, 26, 76. [Google Scholar] [CrossRef] [PubMed]
- Bakke, T.A.; Harris, P.D.; Hansen, H.; Cable, J.; Hansen, L.P. Susceptibility of Baltic and East Atlantic Salmon Salmo salar Stocks to Gyrodactylus salaris (Monogenea). Dis. Aquat. Organ. 2004, 58, 171–177. [Google Scholar] [CrossRef]
- Kerr, S.J.; Grant, R.E. Ecological Impacts of Fish Introductions: Evaluating the Risk; 2000; ISBN 0777893169.
- Hickley, P.; Muchiri, S.M. Management and Ecological Note Discovery of Carp, Cyprinus carpio, in the Already Stressed Fishery of Lake Naivasha, Kenya. Mar. Freshw. Res. 2004, 139–142. [Google Scholar]
- Otachi, E.O.; Magana, A.E.M.; Jirsa, F.; Fellner-Frank, C. Parasites of Commercially Important Fish from Lake Naivasha, Rift Valley, Kenya. Parasitol. Res. 2014, 113, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, F.; Robert Britton, J.; Mavuti, K.M.; Pacini, N.; Grey, J.; Tricarico, E.; Harper, D.M. A Review of Allodiversity in Lake Naivasha, Kenya: Developing Conservation Actions to Protect East African Lakes from the Negative Impacts of Alien Species. Biol. Conserv. 2011, 144, 2585–2596. [Google Scholar] [CrossRef]
- Pellegrin, J. Poissons Du Gribingui Recueillis Par M. Baudon, Description de Sept Especes Nouvelles. Bull. la Société Zool. Fr. 1919, 44, 201–214. [Google Scholar]
- Justine, J. L.; Briand, M.J.; Bray, R.A. A Quick and Simple Method, Usable in the Field, for Collecting Parasites in Suitable Condition for Both Morphological and Molecular Studies. Parasitol. Res. 2012, 111, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Fricke, R.; Eschmeyer, W.; Van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References Available online: https://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp.
- Pariselle, A.; Euzet, L. Three New Species of Cichlidogyrus Paperna, 1960 (Monogenea, Ancyrocephalidae) Parasitic on Tylochromis jentinki (Steindachner, 1895) (Pisces, Cichlidae) in West Africa. Syst. Parasitol. 1994, 29, 229–234. [Google Scholar] [CrossRef]
- Humason, G.L. Animal Tissue Techniques; W.H. Freeman and Company: United States of America, 1979. [Google Scholar]
- Malmberg, G. The Excretory Systems and the Marginal Hooks as a Basis for the Systematics of Gyrodactylus (Trematoda, Monogenea). Arkiv for Zoologi, 1970. [Google Scholar]
- Bates, J.W. The Slide-Sealing Compound "Glyceel". 1997, 29, 565–566.
- Pugachev, O.N.; Gerasev, P.; Gussev, A.; Ergens, R.; Khotenowsky, I. Guide to Monogenoidea of Freshwater Fish of Palaeartic and Amur Regions, Ledizoni‐Ledipublishing, 2009.
- Bush, A.O.; Lattertyf, K.D.; Lotz, J.M.; Shostakll, A.W.; Brandon, Z.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et Al. Revisited*. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Shinn, A.P.; Hansen, H.; Olstad, K.; Bachmann, L.; Bakke, T.A. The Use of Morphometric Characters to Discriminate Specimens of Laboratory-Reared and Wild Populations of Gyrodactylus salaris and G. thymalli (Monogenea). Folia Parasitol. (Praha). 2004, 51, 239–252. [Google Scholar] [CrossRef]
- Paperna, I. New Species of Monogenea (Vermes) from African Freshwater Fish. A Preliminary Report. Rev. Zool. Bot. Africaines. 1973, 87, 505–518. [Google Scholar]
- Benovics, M.; Nejat, F.; Abdoli, A.; Šimková, A. Molecular and Morphological Phylogeny of Host-Specific Dactylogyrus Parasites (Monogenea) Sheds New Light on the Puzzling Middle Eastern Origin of European and African Lineages. Parasites and Vectors. 2021, 14, 1–15. [Google Scholar] [CrossRef]
- Jalali, B.; Shamsi, S.; Barzegar, M. Occurrence of Gyrodactylus spp (Monogenea: Gyrodactylidae) from Iranian Freshwater Fishes. Iran. J. Fish. Sci. 2005, 4, 19–30. [Google Scholar]
- Barzegar, M.; Ebrahimzadeh Mousavi, H.; Rahmati-holasoo, H.; Taheri Mirghaed, A.; Bozorgnia, A. Gyrodactylus (Monogenea, Gyrodactylidae) Parasite Fauna of Fishes in Some Rivers of the Southern Caspian Sea Basin in Mazandaran Province. Iran. J. Vet. Med. 2017, 12, 35–44. [Google Scholar] [CrossRef]
- Younis, S.A.; Shamall, M.A.A. Monogenean Infections on Fishes from Darbandikhan Lake in Kurdistan Region, Iraq. Basrah J. Agric. Sci. 2013, 26, 117–131. [Google Scholar] [CrossRef]
- Louizi, H.; Vanhove, M.P.M.; Rahmouni, I.; Berrada Rkhami, O.; Benhoussa, A.; Van Steenberge, M.; Pariselle, A. Species Depauperate Communities and Low Abundances of Monogenean Gill Parasites at the Edge of the Natural Distribution Range of Their Cichlid Hosts in Northern Africa. Hydrobiologia. 2022, Advances in Cichlid Research V, 1. 1–11. [Google Scholar] [CrossRef]
- El Gharbi, S.; Birgi, E.; Lambert, A. Monogènes Dactylogyridae Parasites de Cyprinidae du Genre Barbus d’Afrique Du Nord. Syst. Parasitol. 1994, 27, 45–70. [Google Scholar] [CrossRef]
- Přikrylová, I.; Blažek, R.; Vanhove, M.P.M. An Overview of the Gyrodactylus (Monogenea: Gyrodactylidae) Species Parasitizing African Catfishes, and Their Morphological and Molecular Diversity. Parasitol. Res. 2012, 110, 1185–1200. [Google Scholar] [CrossRef]
- Vanhove, M.P.M.; Snoeks, J.; Volckaert, F.A.M.; Huyse, T. First Description of Monogenean Parasites in Lake Tanganyika: The Cichlid Simochromis diagramma (Teleostei, Cichlidae) Harbours a High Diversity of Gyrodactylus Species (Platyhelminthes, Monogenea). Parasitology. 2011, 138, 364–380. [Google Scholar] [CrossRef]
Figure 1.
An overview of the sampling locations of cyprinid hosts examined for the presence of members of Gyrodactylus. The type host and additional host localities for the newly described Gyrodactylus species in the present study are depicted by blue and green circles respectively.
Figure 1.
An overview of the sampling locations of cyprinid hosts examined for the presence of members of Gyrodactylus. The type host and additional host localities for the newly described Gyrodactylus species in the present study are depicted by blue and green circles respectively.
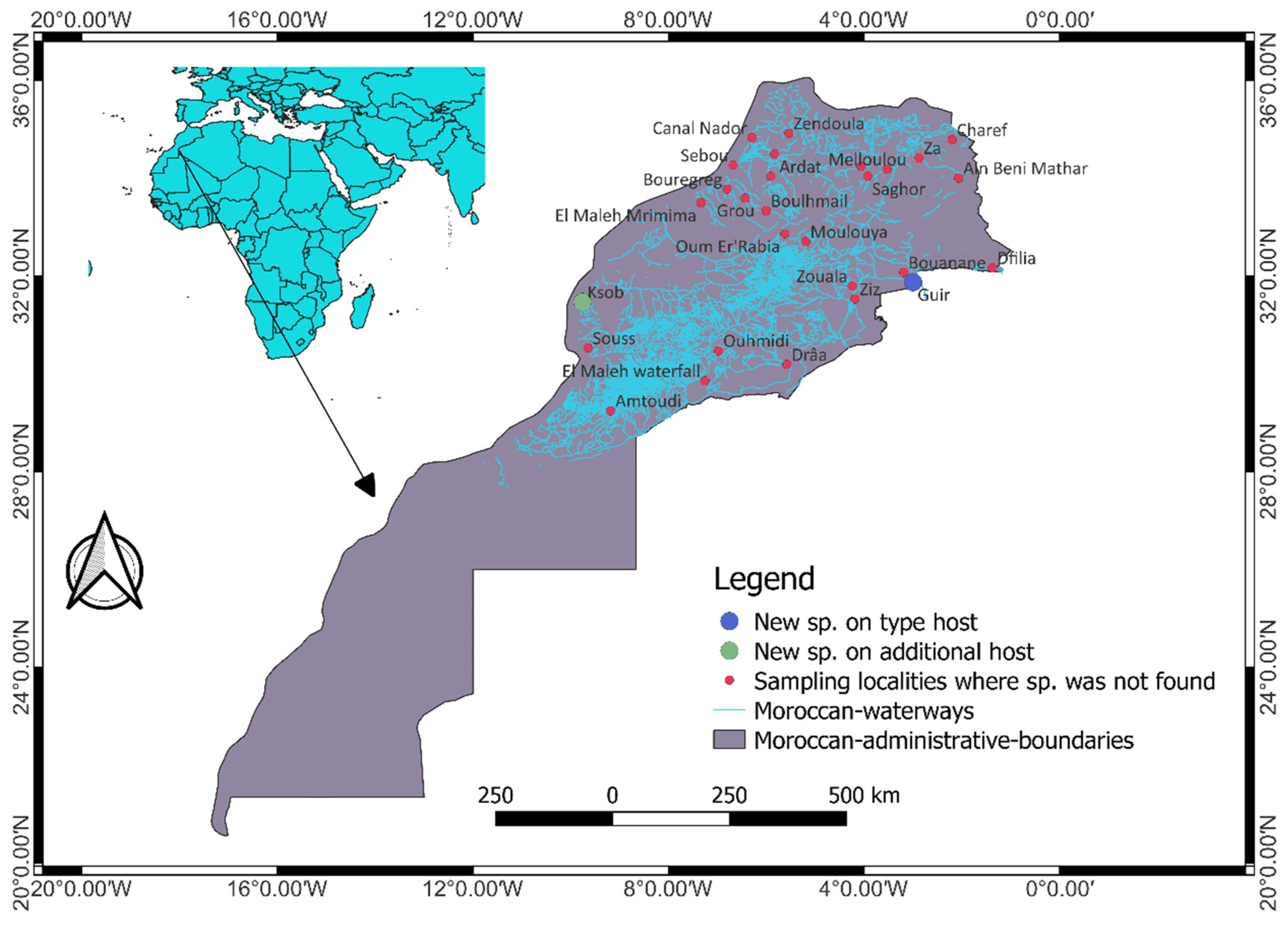
Figure 5.
Biplot of PCA (first two axes) of all the 12 specimens of Gyrodactylus sp.
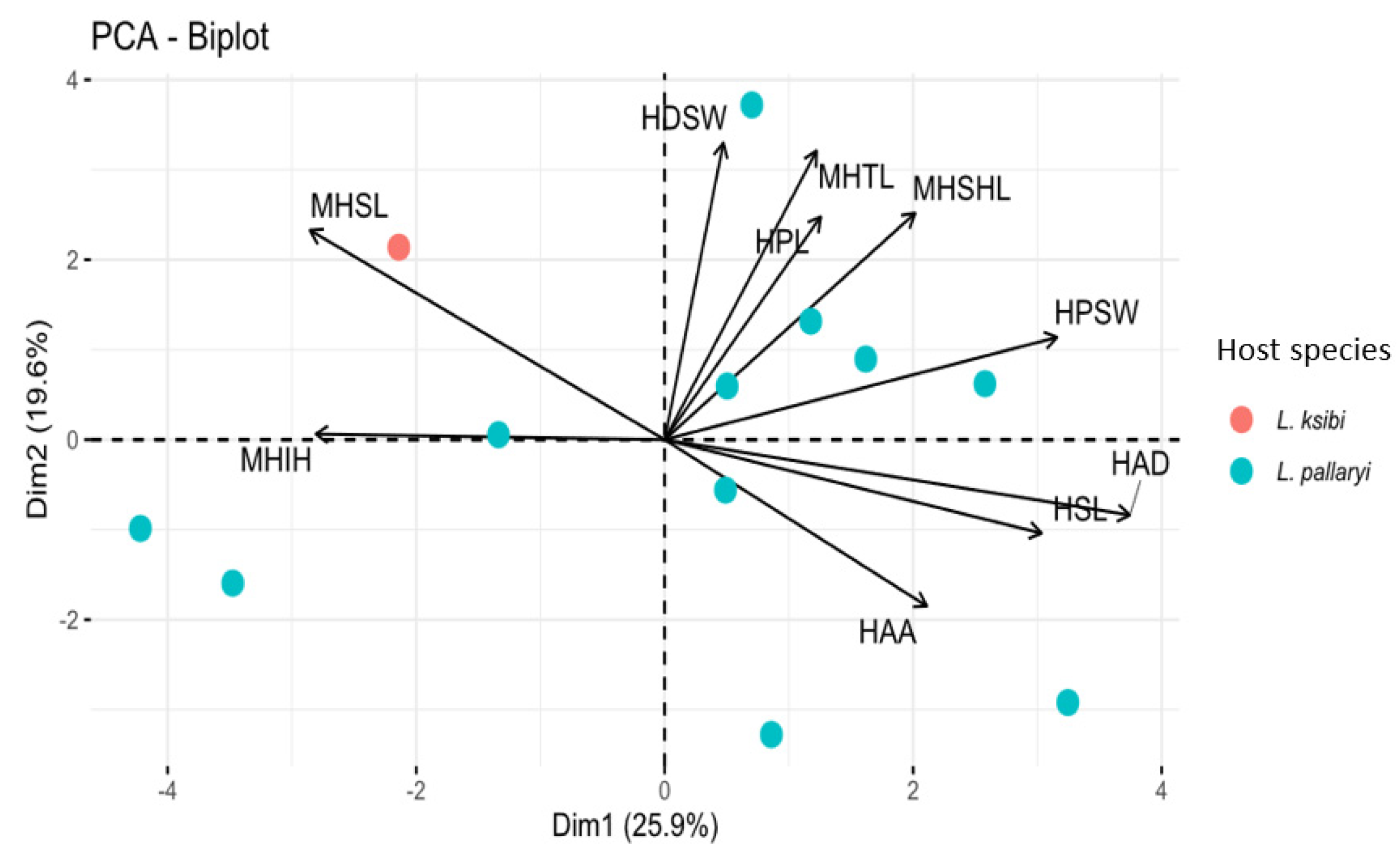
Table 1.
Cyprinid specimens collected, their localities and number of individuals belonging to Gyrodactylus infecting the hosts.
Table 1.
Cyprinid specimens collected, their localities and number of individuals belonging to Gyrodactylus infecting the hosts.
| Host | Locality | Watershed | Coordinates | Nb. of hosts sampled | Nb. of specimens of Gyrodactylus isolated from the hosts | |
|---|---|---|---|---|---|---|
| Latitude | Longitude | |||||
| Luciobarbus pallaryi (Pellegrin, 1919) | Oued Guir | Ziz | 31°52′12′′N | 003°0′00′′W | 157 | 14 |
| Oued Bouanane | Ziz | 32°04′04′′N | 003°11′23.9′′W | |||
| Oued Dfilia | Ziz | 32°9′48.892′′N | 001°22′37.4′′W | |||
| Luciobarbus rabatensis Doadrio, Perea & Yahyaoui, 2015 | Oued Grou | Bouregreg | 33°35′28.0′′N | 006°25′49.6′′W | 24 | 3 |
| Oued Bouregreg | Bouregreg | 33°46′18.0′′N | 006°48′16.6′′W | |||
| Oued Boulhmail | Bouregreg | 33°19′49.6′′N | 006°00′15.1′′W | |||
| Luciobarbus maghrebensis Doadrio, Perea & Yahyaoui, 2015 | Oued Lahdar | Sebou | 34°14′32.7′′N | 004°03′53.9′′W | 55 | 1 |
| Oued Saghor | Sebou | 34°02′4.0′′ N | 003°55′45.5′W | |||
| Oued Ardat | Sebou | 34°29′26.8′′N | 005°49′49.2′′W | |||
| Oued Beht | Sebou | 34°01′55.5′′N | 005°54′43.2′′W | |||
| Oued Sebou | Sebou | 34°15′48.0′′N | 006°40′42.0′′W | |||
| Canal Nador | Sebou | 34°49′19.7′′N | 006°17′36.7′′W | |||
| Luciobarbus rifensis Doadrio, Casal-López & Perea 2015 | Oued Zendoula | Loukkos | 34°54′57.6′′N | 005°32′17.2′′W | 19 | 3 |
| Luciobarbus guercifensis Doadrio, Perea & Yahyaoui, 2016 | Oued Melloulou | Moulouya | 34°10′51.7′′N | 003°31′59.6′′W | 4 | 0 |
| Oued Za | Moulouya | 34°24′38.9′′N | 002°52′28.1′′W | |||
| Luciobarbus yahyaouii Doadrio, Casal-López & Perea 2016 | Oued Za | Moulouya | 34°24′38.9′′N | 002°52′28.1′′W | 62 | 0 |
| Oued Charef | Moulouya | 34°46′44.0′′N | 002°11′56.0′′W | |||
| Oued Melloulou | Moulouya | 34°10′51.7′′N | 003°31′59.6′′W | |||
| Ain Beni Mathar | Moulouya | 34°00′00.3′′N | 002°03′58.6′′W | |||
| Luciobarbus zayanensis Doadrio, Casal-López & Yahyaoui, 2016 | Oued Oum Er′Rabia | Oum Er′Rabia | 32°51′32.8′′N | 005°37′18.9′′W | 25 | 1 |
| Oued Moulouya | Moulouya | 32°41′55.4′′N | 005°11′51.2′′W | |||
| Luciobarbus lepineyi (Pellegrin, 1939) | Oued Ziz | Ziz | 31°31′34.7′′N | 004°11′10.0′′W | 127 | 0 |
| Oued Zouala | Ziz | 31°47′31.9′′N | 004°14′43.5′′W | |||
| Oued Dfilia | Ziz | 32°9′48.892′′N | 001°22′37.4′′W | |||
| Oued Drâa | Draa | 30°11′12.24′′N | 005°34′47.34′′W | |||
| Oued Ouhmidi | Draa | 30°28′5.64′′N | 006°58′36.12′′W | |||
| Oued El Maleh Mrimima | Draa | 33°29’34.8’’N | 007°19′58.1′′W | |||
| Oued El Maleh Waterfall | Draa | 29°51′108′′N | 007°15′23′′W | |||
| Oued Amtoudi | Draa | 29°14′32.42′′ N | 009°11′8.71′′W | |||
| Carasobarbus moulouyensis (Pellegrin, 1924) | Oued Moulouya | Moulouya | 32°41′55.4′′N | 005°11′51.2′′W | 44 | 1 |
| Luciobarbus ksibi (Boulenger, 1905) | Oued Oum Er′Rabia | Oum Er′Rabia | 32°51′32.8′′N | 005°37′18.9′′W | 40 | 1 |
| Oued Ksob | Ksob | 31°27′50.7′′N | 009°45′25.3′′W | |||
| Luciobarbus massaensis (Pellegrin, 1922) | Oued Souss | Souss-Massa | 30°31′33.6′′N | 009°38′53.6′′W | 21 | 1 |
| Carasobarbus fritschii (Günther, 1874) | Oued Grou | Bouregreg | 33°35′28.0′′N | 006°25′49.6′′W | 157 | 0 |
| Oued Boulhmail | Bouregreg | 33°19′49.6′′N | 006°00′15.1′′W | |||
| Oued Lahdar | Sebou | 34°14′32.7′′N | 004°03′53.9′′W | |||
| Oued Oum Er′Rabia | Oum Er′Rabia | 32°41′03.8′′N | 005°13′00.3′′W | |||
| Oued Za | Moulouya | 34°24′38.9′′N | 002°52′28.1′′W | |||
| Oued Charef | Moulouya | 34°46′44.0′′N | 002°11′56.0′′W | |||
| Oued Ksob | Ksob | 31°27′50.7′′N | 009°45′25.3′′W | |||
| Oued Ardat | Sebou | 34°29′26.8′′N | 005°49′49.2′′ W | |||
| Oued Beht | Sebou | 34°01′55.5′′N | 005°54′43.2′′W | |||
| Oued Sebou | Sebou | 34°15′48.0′′N | 006°40′42.0′′ W | |||
| Pterocapoeta maroccana Günther, 1902 | Oued Oum Er′Rabia | Oum Er′Rabia | 32°51′32.8′′N | 005°37′18.9′′W | 3 | 1 |
| Total | 738 | 26 | ||||
Table 3.
Morphometric measurements of sclerotized parts of Gyrodactylus nyingiae n. sp. The number of structures measured is given in superscript.
Table 3.
Morphometric measurements of sclerotized parts of Gyrodactylus nyingiae n. sp. The number of structures measured is given in superscript.
| Host | Luciobarbus pallaryi (n=1) | Luciobarbus ksibi (n=1) | Both host species combined |
|---|---|---|---|
| Total body length | 386.8 (278.3-456)5 | 443.7 | 396.3 (278.3-456)6 |
| Total body width | 133 (115.8-145.9)6 | 158 | 136.6 (115.8-158.4)7 |
| Hamulus total length | 76.5 (65.9-88.2)10 | 75.3 | 76.4 (65.9-88.2)11 |
| Hamulus sickle length | 47.6 (42.5-54.8)8 | 45.1 | 47.4 (42.5-54.8)9 |
| Hamulus aperture distance | 27.5 (21.1-30.2)9 | 22.6 | 26.7 (21.1-30.2)10 |
| Hamulus point length | 36.9 (31.7-41.3)9 | 36.3 | 36.2 (31.7-41.3)10 |
| Hamulus inner curve length | 1.7 (1.4-2.7)6 | 4 | 2.1 (1.4-4)7 |
| Hamulus distal shaft width | 5.5 (4.6-6.7)10 | 6 | 5.7 (4.6-7.3)11 |
| Hamulus root length | 26.7 (24.2-28.3)7 | - | 26.7 (24.2-28.3)7 |
| Hamulus aperture angle (in degrees) | 36.9 (31.5-45.4)7 | 32.4 | 36.4 (31.5-45.4)8 |
| Hamulus point curve angle (in degrees) | 4.4 (3.4-5.4)4 | - | 4.4 (3.4-5.4)4 |
| Hamulus inner angle (in degrees) | 40.4 (36-45.4)7 | 37.2 | 40 (36-45.4)8 |
| Hamulus proximal shaft width | 10.4 (8.2-12.1)10 | 10.3 | 10.2 (8.2-12.1)11 |
| Marginal hook total length | 34.4 (31.7-42.1)8 | 35.2 | 34.8 (31.7-42.1)9 |
| Marginal hook shaft length | 28.6 (26.1-33.4)9 | 29.2 | 28.7 (26.1-33.4)10 |
| Marginal hook sickle length | 6.2 (5.5-6.5)9 | 6.6 | 6.3 (5.5-6.6)10 |
| Marginal hook sickle proximal width | 4.6 (3.9-5)9 | 5.5 | 4.7 (3.9-5.5)10 |
| Marginal hook sickle distal width | 4.5 (3.9-5.1)9 | 5 | 4.5 (3.9-5.1)10 |
| Marginal hook sickle toe length | 1.9 (1.8-2.1)9 | 2.1 | 2 (1.8-2.1)10 |
| Marginal hook aperture distance | 5.5 (5-5.9)8 | 5.3 | 5.4 (5-5.9)9 |
| Marginal hook in-step height | 0.6 (0.5-0.9)8 | 0.7 | 0.6 (0.5-0.9)9 |
| Ventral bar total length | 19.6 (18.6-20.5)2 | - | 19.6 (18.6-20.5)2 |
| Ventral bar total width | 25.1 (24.8-25.4)2 | - | 25.1 (24.8-25.4)2 |
| Ventral bar median length | 6.1 (5.5-6.8)3 | - | 6.1 (5.5-6.8)3 |
| Ventral bar membrane lengthVentral bar process length | 13.6 (12.7-14.5)33.7 (3.6-3.8)2 | - - | 13.6 (12.7-14.5)33.7 (3.6-3.8)2 |
| Male copulatory organ diameter | 18.4 (16.5-19.5)4 | 21.2 | 18.9 (16.5-21.2)5 |
| Principal spine length | 6.5 (6.3-6.6)3 | 6.5 | 6.5 (6.3-6.6)4 |
| Small spine length | 3.3 (3.1-3.5)3 | 5.4 | 4.4 (3.1-5.4)4 |
| Dorsal bar length | 11.9 (9.9-13.4)3 | - | 11.9 (9.9-13.4)3 |
| Dorsal bar width | 1.6 (1.2-1.9)3 | - | 1.6 (1.2-1.9)3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



