
Submitted:
27 February 2023
Posted:
28 February 2023
You are already at the latest version
Abstract
The objective of this study was to evaluate the effect of the addition of a phytogenic compounds blend (PHA) containing hydrolyzable tannins, carvacrol and cinnamaldehyde oil in the mineral salt or energy supplement on the rumen microbiota and nitrogen metabolism of grazing Nellore cattle. Eight castrated Nellore steers were distributed in a double Latin square 4 × 4 design, with a 2 × 2 factorial arrangement (two type supplements and the addition or not of PHA) as follows: energy supplement without PHA addition (EW); energy supplement with PHA addition (EPHA); mineral supplement without the addition of PHA (MW); and mineral supplement with PHA addition (MPHA). Steers that received supplements with PHA have a lower ruminal proportion of Valerate (with PHA 1.06%; without PHA 1.15%), a lower ruminal abundance of Verrucomicrobia and a tendency for lower DM digestibility (with PHA 62.8%; without PHA 64.8%). Energy supplements allowed higher ammonia concentrations (+2.28 mg of NH3-N/dL), increased the propionate proportion (+0.29% of total VFA), and showed a tendency to have a higher abundance of Bacteroidetes and a lower abundance of Firmicutes in the rumen. The PHA addition in the supplement did not improve the use of nitrogen in grazing cattle, but negatively affected the diet digestibility.
Keywords:
digestibility
; essential oils
; hydrolyzable tannins
; rumen fermentation
; ruminal bacteria.
1. Introduction
The ruminal microbiota activity is important for the supply of nutrients to the host, involving a relevant association between diet utilization efficiency and rumen microbial diversity[1]. Antimicrobials feed additives can potentially improve feed efficiency through rumen fermentation modulation, but the possible generation of bacterial resistance and residues in animal products has banned their use in ruminant nutrition [2,3]. Thus, phytogenic compounds such as tannins and essential oils emerge as safer alternatives to modulate the ruminal environment and improve the efficiency of diet utilization in cattle.
Hydrolyzable tannins are water-soluble polyphenolic compounds that do not bind proteins in the rumen but may result in monomeric subunits of low molecular weight with antimicrobial potential by reacting with secreted extracellular enzymes and microbial cell walls [4]. Tannin extracts may influence the rumen microbiota, by reducing the diversity and increasing richness of the ruminal bacteria population [5], and consequently, alter the rumen fermentation of diet [6,7]. Thus, low to moderate inclusion levels (≤ 20 g/kg DM) of hydrolyzable tannins have been used in cattle diets [8,9,10], to avoid toxicity due to its degradation by some rumen microorganisms [11] and obtain beneficial effects, as a higher concentration of ruminal volatile fatty acids [12], lower acetate: propionate ratio [13], and the reduction of nitrogen losses by urine [14].
Compounds present in essential oils such as cinnamaldehyde, derived from cinnamon, have shown high antioxidant power and antimicrobial activity [15], due to the presence of chemical compounds of the phenylpropanoids class which can damage the enzymes necessary for energy production in bacteria or even denature proteins [16]. Cinnamaldehyde may improve energy efficiency and utilization of nitrogen in the rumen [17], reducing Prevotella.spp abundance [18]. Carvacrol is a phenolic compound from oregano, with nonspecific antimicrobial activity, due to the hydroxyl group present in its molecule, which functions as an ion transporter, and its high hydrophobicity [18]. Carvacrol has the potential to influence gram-positive and gram-negative bacteria, and modulate ruminal fermentation, reducing both ruminal protein degradability and the acetate: butyrate ratio [18,19]. The association of essential oils has shown promising effects on ruminal fermentation modulation depending on the dosage and basal diet [20]. A blend of essential oils, bioflavonoids, and tannins improved diet digestibility and feed conversion rate in dairy cattle [21]. But the association of hydrolyzable tannins and essential oils during the supplementation of grazing beef cattle resulted in a negative effect on forage intake and digestibility [22].
Energy supplementation in tropical pasture-based systems well managed can promote greater uptake of nitrogen compounds in the rumen and improve ruminal fermentation and digestibility in grazing cattle [23]. To date, the response of the rumen microbiota and nitrogen metabolism in steers grazing tropical forages when supplemented with energy and phytogenic compounds has not been reported. Thus, was hypothesized that the use of a blend of hydrolyzable tannins and essential oils in the energy supplement of grazing steers could improve nitrogen metabolism through modulation of the ruminal microbiome and reduction of nitrogen excretion via urine. The objective of this study was to evaluate the effect of the inclusion of a phytogenic compounds blend (PHA) containing hydrolyzable tannins, carvacrol, and cinnamaldehyde oil supplied at a dose of 1.5 g/kg of ingested dry matter (DM) in the mineral salt or energy supplement on the rumen microbiota and nitrogen metabolism of grazing Nellore cattle during the rainy season.
2. Materials and Methods
2.1. Grazing area and animals
The experiment was carried out at the Faculty of Agricultural and Veterinary Sciences (FCAV) of the São Paulo State University “Júlio de Mesquita Filho” (UNESP), Jaboticabal campus, São Paulo (Brazil), during the rainy season from January to April of 2018 (21◦15′22′ ′ S latitude and 48◦18′58′ ′W longitude). The climate of this region is subtropical type AW according to the Köppen classification. Animal care and handling followed the guidelines of the Brazilian College of Animal Experimentation (COBEA) and was approved by the Ethics Committee on the Use of Animals (CEUA) of FCAV/UNESP (Protocol No. 12703/15).
Eight castrated Nellore steers, cannulated in the rumen (silicone-type cannula), with an average body weight (BW) of 456.6 kg ± 32.8 kg, were randomly distributed in a double Latin square 4 × 4 design, with a 2 × 2 factorial arrangement of treatments (energy or mineral supplement and with or without phytogenic compounds), and four experimental periods. Steers were housed in eight paddocks of 0.7 and 1.3 ha (one steer in each paddock) formed by Urochloa brizantha cv. Marandu, delimited by electric fencing and served by a trough with access from both sides (30 cm linear per animal) and a drinking fountain. The meteorological data and maintenance of fertilization in the experimental area during the experimental period were described by Teobaldo et al. [22]. Pastures were managed under continuous stocking with variable stocking rates and a canopy target of approximately 25 cm high, using the “put and take” technique [24], and animals’ regulators to adjust and maintain the height target, in addition to the steers testers. The forage height was measured weekly from 80 random points per paddock to the stocking rate adjustment. Every 28 days, three points per paddock were sampled (using a frame of 0.25 m2) representing the average forage height to evaluate the forage mass and morphological components, thus, the average total herbage mass was 6.52 t of DM per ha with an average herbage allowance of 3.86 kg of DM per kg of BW.
The steers were distributed into the paddocks two weeks before the beginning of the experiment to adapt to the conditions. Then, four experimental periods of 21 days were conducted, considering 14 days of treatment adaptation and seven days of sampling phase, to test four supplements as follows: energy supplement without phytogenic compounds addition (EW); energy supplement with phytogenic compounds addition (EPHA) (Table 1); mineral supplement without the addition of phytogenic compounds (MW); and mineral supplement with the addition of phytogenic compounds (MPHA). Both energy supplements were offered daily at 09:00 a.m. at 0.3% BW, while mineral supplements were provided ad libitum. The phytogenic compounds blend (PHA) contained 10% of carvacrol and cinnamaldehyde oil, and 90% hydrolyzable tannins extracted from berries and grapes, supplied at a dose of 1.5 g/kg of ingested DM, as recommended by commercial suppliers. The energy supplements used were commercial, and suppliers were not authorized to disclose the percentage composition of the ingredients. These supplements did not contain urea, containing corn gluten meal and minerals (Ca 20.0 g/kg; Na 20.0 g/kg; S 10.0 g/kg; Cu 133.0 mg/kg; Mn 49 mg/kg; Zn 340.0 mg/kg; Co 1.4 mg/kg; I 7.0 mg/kg; Se 2.25 mg/kg). Mineral salt supplements warranty levels were Ca 123.0 g/kg; P 90.0 g/kg; Cu 1040.0 mg/kg; Mn 500 mg/kg; Zn 2000.0 mg/kg; Co 15.0 mg/kg; I 67.0 mg/kg; Se 14.0 mg/kg.
2.2. Intake and digestibility
The intake and digestibility of the forage were evaluated on days 16–28 of each experimental period using Chromium oxide (Cr2O3) as an external marker to estimate the excretion of fecal matter and the iNDF as an internal marker. One cellulose capsule containing 10 g of chromium oxide was placed in the rumen of each animal through the cannula, daily at 9:00 for 10 days (7 of adaptation and 3 days of fecal sampling). Fecal samples were collected directly from the rectum, once daily alternating at the following times: 5:00 p.m., 11:00 a.m., and 6:00 a.m. The forage bromatological composition was evaluated in forage samples collected by the manual grazing simulation method [25] each 28 d. The average of each supplement intake was calculated from the average amount of supplement provided (% BW) per paddock.
Forage, energy supplements, and feces samples were dried under forced air with forced at 55° C for 72 h, and ground in a Willey mill with a 1- or 2-mm mesh sieve for bromatological and indigestible neutral detergent fiber analysis (iNDF), respectively. Then, the DM, organic matter (OM), mineral matter (MM), and ether extract (EE) were determined according to AOAC [26]. The crude protein (CP) was determined using the Dumas method in a LECO FP-528 N analyzer (Leco Corporation, St. Joseph, MI, USA). Neutral (NDF) and acid (ADF) detergent fiber were determined following the recommendations of Van Soest et al. [27] and Goering and Van Soest [28], respectively, using Ankom® 2000 equipment (Ankom Technologies, Fairport, NY, USA). α-amylase was added during the NDF procedures of the energy supplements samples. In situ incubation procedure for 288 h [29] was used to obtain iNDF content. The gross energy (GE) content was obtained by the combustion of samples in an adiabatic calorimetric pump (PARR Instrument Company 6300, Moline, IL, USA). Chromium oxide in feces was measured after acid digestion, using an atomic absorption spectrophotometer [30].
2.3. Nitrogen metabolism
Urine spot samples were collected once daily at the same moment as feces collection, during steer spontaneous urination. Approximately 50 mL of urine were filtered through three layers of cheesecloth and two aliquots were immediately stored at - 20 ◦C, for the later analysis of creatinine, uric acid, and urea by a colorimetric-enzymatic method using commercial kits (Labtest®, Lagoa Santa, MG, Brazil), total N concentration through Dumas method in a LECO FP-528 N analyzer (Leco Corporation, St. Joseph, MI, USA), and allantoin by the colorimetric method described by Chen and Gomes [31].
Daily urinary volume was estimated according to Costa e Silva et al. [32] based on the relationship between daily urinary creatinine excretion and BW. Then, the daily urinary excretion of N compounds (urea and total nitrogen) was calculated as the product of the total N compounds concentration in the samples and the estimated daily urinary volume. The daily N intake was obtained as a summatory of N intake from forage and N intake from the supplement. The daily fecal excretion of N was calculated as the de product of the total N concentration in feces and the estimated excretion of fecal matter.
The total purine derivates were calculated as the summatory of allantoin and uric acid and expressed in mmol/d [30]. Purine derivates absorbed (Y, mmol/d) were calculated using the equation proposed by Verbic et al. [33]: Y= (X – (0.30×BW0.75))/0.80, where 0.80 correspond to the recovery of absorbed purines as purine derivatives in the urine (mmol/mmol), and 0.30*BW0.75 is the endogenous excretion of purine derivatives (mmol) in the urine per unit of metabolic body size [33]. The ruminal microbial nitrogen synthesis (Nmic, g N/day) was estimated using the equation proposal by Barbosa et al. [34]: Y=70×X / (0.83×0.137×1000), where 70 is the N content of purines (mg/mmol); 0.137 corresponds to the purine N: total N ratio in bacteria, and 0.83 is the true digestibility of microbial purines. The digestible organic matter apparently fermented in the rumen (OMFR) was calculated as the product of the digestible OM intake and the factor 0.65 [35] and, later, used to calculate the efficiency of microbial protein synthesis, expressed in g of Nmic/kg of OMFR.
Blood samples were collected before supplementation (0 hours) and 4 hours after supplementation on day 21 of each experimental period. Blood was sampled directly from the caudal vein using vacuum tubes and clotting accelerator gel and stored at -20ºC to determine the serum urea level later through commercial kits (Labtest®, Lagoa Santa, MG, Brazil). The conversion of urea values into urea N was obtained as the product of values obtained by commercial kits and the factor 0.466.
2.4. Rumen fermentation parameters and microbiota population
Rumen content was collected through the ruminal cannula on day 20 of each experimental period at 0, 6, 9, and 12 h after supplementation, and immediately filtered through double layers of cheesecloth, then, approximately 100 mL of ruminal fluid was recovered. The pH was measured by a digital pH meter (TEC 7, Tecnal, Piracicaba, SP, Brazil), and two aliquots of approximately 50 mL were stored at - 20 ◦C. Later, one aliquot was used for ruminal ammonia (NH3-N) concentration analysis in a Kjeldahl system [26], and the second aliquot was used for volatile fatty acid (VFA) quantification following the recommendations of Famme and Knudsen [36] by gas chromatography (2014AF, Shimatzu Corporation, Kyoto, Japan).
Approximately 100 g of rumen content (solid + liquid) was collected from several regions of the rumen before supplementation (0h) on day 20. These samples were immediately stored in coolers with ice and transported to the laboratory. Then, a bacterial pellet was obtained as described by Granja-Salcedo et al. [37]. The DNA extraction was carried out using the Quick-DNA™ Fecal/Soil Microbe Miniprep kit (Zymo Research Corporation, CA, USA) and the bead beating method for cell lysis (FastPrep-24, MP, Biomedicals, Illkirch, France). The DNA concentrations were checked by spectrophotometer (NanoDropR ND-1000 Spectrophotometer, Thermo Fisher Scientific, Waltham, MA, United States) and fluorometer (QubitR3.0, kit Qubit RdsDNA Broad Range Assay Kit, Life Technologies, Carlsbad, CA, USA). The DNA purity was checked through absorbance ratios (260/230 and 260/280 nm), and DNA integrity was assessed by 0.8% (w/v) agarose gel electrophoresis stained with SYBR Safe DNA Gel Stains (Thermo Fisher Scientific, Waltham, MA, EUA).
Libraries in duplicate were prepared by PCR amplification of the V4 -V5 regions of the 16S ribosomal RNA gene 16S rRNA, using barcoded 16S Illumina primers [38]. PCR product length and amplicon size were checked by 1% (w/v) agarose gel electrophoresis using a 1 kb plus DNA ladder (Invitrogen, Carlsbad, CA, USA). Then, PCR fragments were purified using a Zymoclean TM Gel DNA Recovery kit (Zymo Research Corporation, CA, United States). Sequencing was performed using the MiSeq Reagent v2 (2 × 250 bp; Illumina®, USA) kit in an Illumina MiSeq® Machine.
Bioinformatics analyses were performed with QIIME 2 [39]. Raw sequence data were demultiplexed and quality filtered using the q2-demux plugin followed by denoising with DADA2 [40]. The q2-diversity plugin was used to estimate the diversity metrics (after the samples were rarefied (subsampled without replacement) to 4994 sequences per sample): alpha-diversity metrics (observed OTUs, evenness, and Faith’s Phylogenetic Diversity [41], beta diversity metrics (unweighted UniFrac [42] and Bray-Curtis dissimilarity). Taxonomy was assigned to ASVs using the q2-feature-classifier classify-consensus-vsearch taxonomy classifier against the Silva 128 database with 97% OTUs reference sequences [43]. Predictive functional profiling of microbial communities was assessed by PICRUSt [44]. OTU representative sequences were captured from the Greengenes database using q2 feature-classifier classify-sklearn [45]. Predicted functional genes were categorized into Clusters of Kyoto Encyclopedia of Genes and Genome (KEGG) Orthology (KO).
2.5. Statistical analyses
Intake, digestibility, and nitrogen metabolism data were analyzed using the PROC MIXED do SAS v.9.2 (SAS Inst. Inc., Cary, NC), as a double 4 × 4 Latin square design in a factorial arrangement (A × B), where the fixed effect of factor A corresponded to the type of supplement as mineral or energy supplement, and the factor B to the phytogenic compounds blend (PHA) addition (yes or no). The ANOVA also included the factors (A×B) interactions, treatments error, the random effects of Latin square, period, animal, the period × animal interaction, and residues corresponding to the model. Tukey’s post hoc test was applied when ANOVA indicated a significant difference.
Rumen fermentation parameters data were analyzed as repeated-measures ANOVA, as a double 4 × 4 Latin square design in a split-plot factorial arrangement (A × B). The model included the fixed effects of factor A and factor B that were considered as independent variables; the sampling times considered as dependent variable (covariate), factors interactions (A × B), factors and time interactions; and the random effects of Latin square, period, animal, the period × animal interaction, and residues corresponding to the model, and the Tukey’s post hoc test was applied when ANOVA indicated a significant difference.
Microbiota population data were compared between factor A and factor B groups by the Friedman rank test. The interactions of the factors were evaluated by Kruskal–Wallis and Dunn’s post hoc tests using R Software version 4.1.1 (R Core Team, 2015). A principal component analysis (PCA) was performed to extract the important microbial OTUs associated with predicted functional genes, ruminal fermentation parameters, intake, digestibility, and N metabolism through the Factor Miner package in R and visualized on a biplot PCA.
For all tests, a probability of P ≤ 0.05 was considered significant, and tendencies were considered when 0.05 < P ≤ 0.10.
3. Results
3.1. Intake and digestibility
No effects of the type of supplement × addition of PHA interactions were observed on intake and apparent total digestibility (Table 2). The total DM intake expressed as % of BW or as kg/d, and the intake of OM, CP, neutral detergent fiber corrected for ash and protein (apNDF), gross energy (GE), digestible energy, and metabolizable energy increased in steers supplemented with energy supplements when compared to steers supplemented with mineral supplements (P <0.05). In addition, the intake of forage expressed as % of BW (P = 0.059) or kg/d (P = 0.070) showed a tendency to increase in steers supplemented with energy supplements than those supplemented with mineral supplements.
The apparent total digestibility of DM, OM, CP, apNDF, and GE increased in steers supplemented with energy supplements compared to mineral supplements (P ≤ 0.01). In addition, a tendency for lower total digestibility of DM was observed with the addition of PHA (62.8%) in the supplements, when compared to steers feed supplements without the addition of PHA (64.8%) (P=0.073).

Table 2.
Effect of phytogenic compounds in mineral or energy supplementation on intake and apparent total digestibility of Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season.
Table 2.
Effect of phytogenic compounds in mineral or energy supplementation on intake and apparent total digestibility of Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season.
| Supplements1 | SEM | P-value2 | ||||||
|---|---|---|---|---|---|---|---|---|
| EW | EPHA | MW | MPHA | ST | PHA | ST × PHA | ||
| Intake (% BW) | ||||||||
| Total DM | 2.52 | 2.42 | 1.87 | 1.97 | 0.08 | 0.001 | 0.977 | 0.623 |
| Forage DM | 2.25 | 2.15 | 1.87 | 1.97 | 0.07 | 0.059 | 0.983 | 0.474 |
| apNDF | 1.30 | 1.26 | 1.16 | 1.10 | 0.10 | 0.121 | 0.595 | 0.949 |
| Intake (kg/d) | ||||||||
| Total DM | 11.60 | 10.92 | 8.75 | 9.15 | 0.38 | 0.001 | 0.810 | 0.386 |
| Forage DM | 10.36 | 9.68 | 8.65 | 9.04 | 0.32 | 0.070 | 0.816 | 0.392 |
| Supplement DM | 1.24 | 1.23 | 0.10 | 0.11 | 0.02 | <0.001 | 0.747 | 0.702 |
| OM | 10.27 | 9.63 | 7.88 | 8.24 | 0.33 | 0.003 | 0.803 | 0.417 |
| CP | 1.79 | 1.66 | 1.39 | 1.39 | 0.06 | 0.006 | 0.551 | 0.559 |
| apNDF | 6.02 | 5.66 | 4.76 | 5.04 | 0.19 | 0.017 | 0.911 | 0.381 |
| GE, MJ/d | 201.5 | 189.0 | 156.4 | 162.3 | 6.35 | 0.004 | 0.761 | 0.411 |
| Digestible energy, MJ/d | 138.6 | 125.6 | 102.8 | 98.51 | 5.45 | 0.002 | 0.340 | 0.627 |
| Metabolizable energy, MJ/d | 117.6 | 107.3 | 85.44 | 79.43 | 8.76 | 0.001 | 0.321 | 0.791 |
| g CP/kg DOM | 247.5 | 249.5 | 259.4 | 262.8 | 13.87 | 0.111 | 0.721 | 0.927 |
| Digestibility (%) | ||||||||
| DM | 67.0 | 65.1 | 62.5 | 60.6 | 0.70 | <0.001 | 0.073 | 0.177 |
| OM | 71.0 | 69.5 | 64.0 | 64.1 | 0.90 | <0.001 | 0.555 | 0.364 |
| CP | 70.0 | 67.3 | 58.9 | 59.7 | 1.31 | <0.001 | 0.515 | 0.235 |
| apNDF | 72.5 | 71.9 | 66.2 | 67.2 | 0.91 | <0.001 | 0.877 | 0.560 |
| GE | 68.5 | 66.3 | 60.0 | 60.6 | 1.02 | <0.001 | 0.536 | 0.219 |
1 EW= energy supplement containing corn gluten meal and minerals without phytogenic compounds addition; EPHA= energy supplement containing corn gluten meal and minerals with phytogenic compounds addition (containing 10% of carvacrol and cinnamaldehyde oil, and 90% hydrolyzable tannins extracted from berries and grapes) at a dose of 1.5 g/kg of ingested DM. MW= mineral supplement warranty levels: Ca 123.0 g/kg; P 90.0 g/kg; Cu 1040.0 mg/kg; Mn 500 mg/kg; Zn 2000.0 mg/kg; Co 15.0 mg/kg; I 67.0 mg/kg; Se 14.0 mg/kg. MPHA= Guarantee levels similar to DM include phytogenic compounds at a dose of 1.5 g/kg of ingested DM. 2 ST = effect of the type of supplement as mineral or energy supplement. PHA = effect of the addition of phytogenic compounds blend. SEM = Standard error of the mean. DM = dry matter, OM = organic matter, CP = crude protein, apNDF = neutral detergent fiber corrected for ash and protein, GE = gross energy, DOM = digestible organic matter.
3.2. Nitrogen metabolism
There was a type of supplement × addition of PHA interaction effect on urinary N excreted (Table 3; P = 0.033). Thus, steers supplemented with EW presented higher values of urinary N excreted (g of N per day) when compared to steers supplemented with MW and MPHA supplements.
Steers supplemented with energy supplements showed higher N intake, higher N retained expressed as g/d and as % of N intake, and higher plasma urea N than steers supplemented with mineral supplements (P<0.05). The plasma urea N concentration increased from 11.1 mg/dL at 0h to 16.43 mg/dL at 4h after supplementation (P<0.001). The fecal N excreted tended to increase in steers supplemented with energy supplements (P= 0.081). The efficiency of microbial protein synthesis expressed as g Nmic/kg OMFR and as g Pmic/kg DOM tended to increase in steers supplemented with mineral supplements than in steers supplemented with energy supplements (P=0.055).
Table 3.
Effect of phytogenic compounds in mineral or energy supplementation on nitrogen balance and efficiency of ruminal microbial nitrogen synthesis in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season.
Table 3.
Effect of phytogenic compounds in mineral or energy supplementation on nitrogen balance and efficiency of ruminal microbial nitrogen synthesis in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season.
| Supplements1 | SEM | P-value2 | ||||||
|---|---|---|---|---|---|---|---|---|
| EW | EPHA | MW | MPHA | ST | PHA | ST × PHA | ||
| N balance | ||||||||
| N intake, g/d | 286.0 | 266.1 | 222.6 | 222.4 | 9.69 | 0.006 | 0.551 | 0.559 |
| Fecal N excreted, g/d | 89.77 | 91.59 | 79.37 | 84.30 | 2.94 | 0.081 | 0.407 | 0.906 |
| Urinary N excreted, g/d | 116.3a | 96.6ab | 89.7b | 96.1b | 3.93 | 0.006 | 0.271 | 0.033 |
| N retained, g/d | 94.51 | 99.73 | 54.46 | 45.82 | 7.59 | <0.001 | 0.611 | 0.992 |
| N retained, % of N intake | 31.10 | 33.99 | 21.97 | 18.96 | 2.04 | 0.050 | 0.987 | 0.409 |
| Ruminal microbial N synthesis | ||||||||
| Nmic, g N/d | 60.15 | 54.43 | 55.92 | 56.63 | 6.48 | 0.848 | 0.636 | 0.544 |
| Pmic, g protein/d | 375.95 | 340.18 | 349.51 | 353.95 | 40.47 | 0.848 | 0.636 | 0.544 |
| ENmic, g Nmic/kg OMFR | 13.10 | 13.36 | 17.39 | 17.00 | 2.29 | 0.055 | 0.974 | 0.868 |
| EPmic, g Pmic/kg DOM | 53.21 | 54.28 | 70.64 | 69.06 | 9.29 | 0.055 | 0.974 | 0.868 |
| Plasma urea N3, mg/dL | 15.95 | 16.01 | 14.52 | 14.61 | 0.22 | <0.001 | 0.786 | 0.939 |
1 EW= energy supplement containing corn gluten meal and minerals without phytogenic compounds addition; EPHA= energy supplement containing corn gluten meal and minerals with phytogenic compounds addition (containing 10% of carvacrol and cinnamaldehyde oil, and 90% hydrolyzable tannins extracted from berries and grapes) at a dose of 1.5 g/kg of ingested DM. MW= mineral supplement warranty levels: Ca 123.0 g/kg; P 90.0 g/kg; Cu 1040.0 mg/kg; Mn 500 mg/kg; Zn 2000.0 mg/kg; Co 15.0 mg/kg; I 67.0 mg/kg; Se 14.0 mg/kg. MPHA= Guarantee levels similar to DM include phytogenic compounds at a dose of 1.5 g/kg of ingested DM. 2 ST = effects of the type of supplement as mineral or energy supplement. 3 Time affected the plasma urea concentration (0h = 11.1 vs. 4h = 16.43 mg/dL; P < 0.001). SEM = Standard error of the mean. PHA = effect of the addition of phytogenic compounds blend. Nmic = ruminal microbial nitrogen synthesis, Pmic = ruminal microbial protein synthesis, OMFR = digestible organic matter apparently fermented in the rumen, DOM = digestible organic matter.
3.3. Rumen fermentation parameters
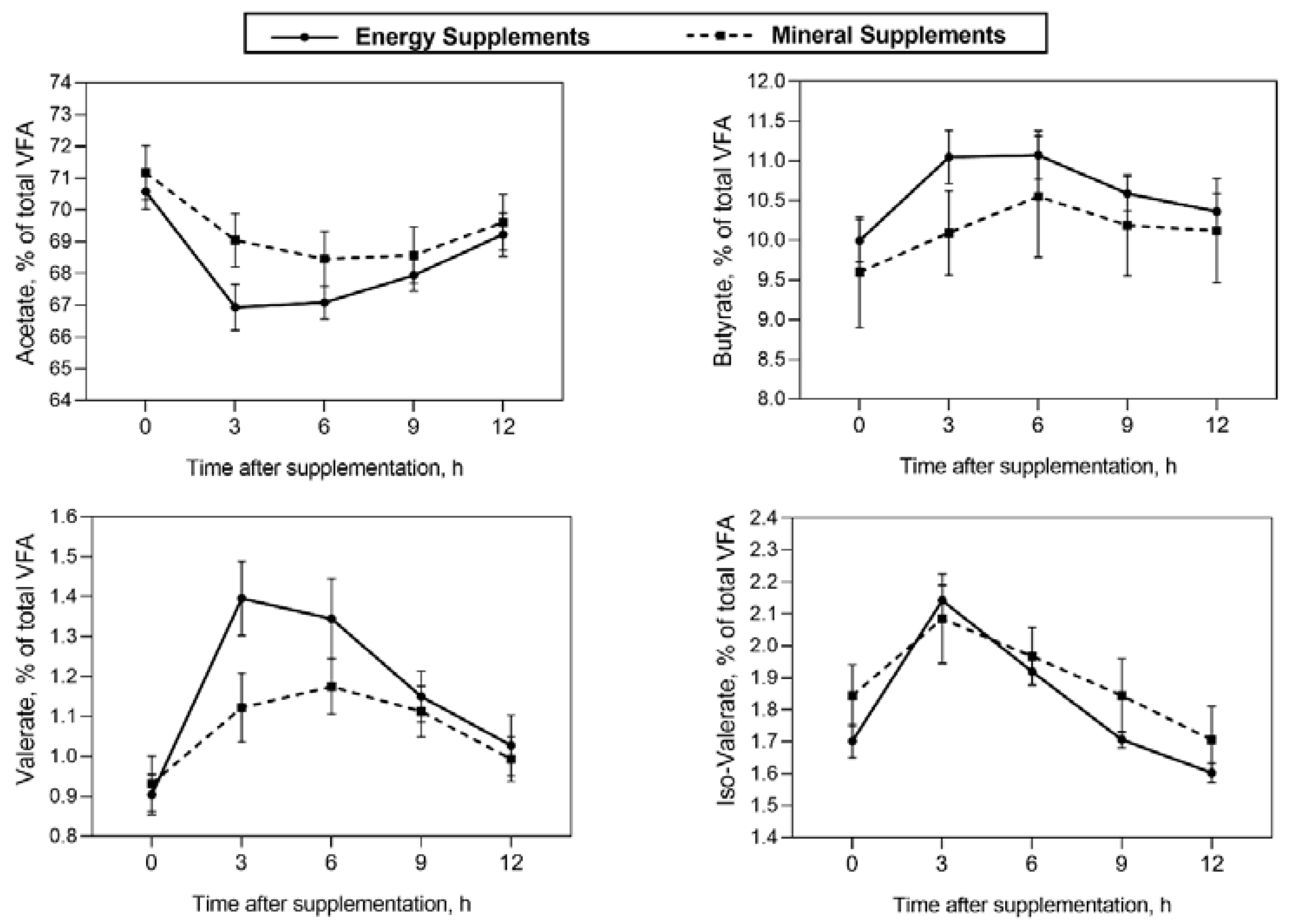
No effects or tendencies of type of supplement × the addition of PHA, and type of supplement × the addition of PHA × time interactions were observed on ruminal fermentation parameters (Table 4; P>0.10). Energy supplements increased ruminal ammonia concentrations (16.51 vs 14.23 mg of NH3-N /dL; P = 0.003) and increased the ruminal propionate proportion (16.89 vs 16.60 %; P = 0.010). Steers that received supplements with PHA showed a lower ruminal proportion of Valerate (with PHA 1.06 % vs without PHA 1.15 %; P=0.015) and tended to have a higher ruminal proportion of propionate (with PHA 16.80 % vs without PHA 16.69 %; P=0.074) than steers fed supplements without PHA. There was an interaction effect of type of supplement × time on the ruminal proportion of acetate, butyrate, valerate, and iso-Valerate, and on the acetate: propionate ratio (Table 4; P<0.05). Thus, at 3h after supplementation steers fed energy supplements showed lower both acetate proportion and lower acetate: propionate ratio, and higher valerate, iso-Valerate, and butyrate proportions in the rumen than steers supplemented with mineral supplements (Figure 1).
Table 4.
Effect of phytogenic compounds in mineral or energy supplementation on ruminal fermentation parameters in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season.
Table 4.
Effect of phytogenic compounds in mineral or energy supplementation on ruminal fermentation parameters in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season.
| Supplements1 | SEM | P-value2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| EW | EPHA | MW | MPHA | ST | PHA | Time | ST × PHA | ST × Time | ||
| pH | 6.25 | 6.14 | 6.35 | 6.34 | 0.02 | 0.115 | 0.691 | <0.001 | 0.679 | 0.726 |
| NH3-N, mg/dL | 17.56 | 15.46 | 14.24 | 14.23 | 0.46 | 0.003 | 0.228 | <0.001 | 0.233 | 0.149 |
| Total VFA, mmol/L | 117.0 | 112.9 | 115.8 | 118.7 | 1.16 | 0.723 | 0.740 | <0.001 | 0.394 | 0.671 |
| Individual VFA, % of total VFA | ||||||||||
| Acetate | 68.30 | 66.74 | 69.70 | 68.99 | 0.15 | 0.003 | 0.335 | <0.001 | 0.190 | <0.001 |
| Propionate | 17.05 | 16.74 | 16.34 | 16.87 | 0.08 | 0.010 | 0.074 | <0.001 | 0.235 | 0.106 |
| Butyrate | 10.66 | 10.31 | 10.06 | 10.16 | 0.06 | 0.198 | 0.792 | <0.001 | 0.360 | <0.001 |
| iso-Butyrate | 0.95 | 0.93 | 0.93 | 1.13 | 0.01 | 0.406 | 0.635 | <0.001 | 0.978 | 0.458 |
| Valerate | 1.22 | 1.08 | 1.08 | 1.05 | 0.02 | 0.001 | 0.015 | <0.001 | 0.149 | <0.001 |
| iso-Valerate | 1.83 | 1.76 | 1.89 | 1.89 | 0.02 | 0.058 | 0.672 | <0.001 | 0.749 | 0.008 |
| A:P ratio | 4.02 | 3.91 | 4.29 | 4.10 | 0.03 | 0.006 | 0.085 | <0.001 | 0.141 | 0.009 |
1 EW= energy supplement containing corn gluten meal and minerals without phytogenic compounds addition; EPHA= energy supplement containing corn gluten meal and minerals with phytogenic compounds addition (containing 10% of carvacrol and cinnamaldehyde oil, and 90% hydrolyzable tannins extracted from berries and grapes) at a dose of 1.5 g/kg of ingested DM. MW= mineral supplement warranty levels: Ca 123.0 g/kg; P 90.0 g/kg; Cu 1040.0 mg/kg; Mn 500 mg/kg; Zn 2000.0 mg/kg; Co 15.0 mg/kg; I 67.0 mg/kg; Se 14.0 mg/kg. MPHA= Guarantee levels similar to DM include phytogenic compounds at a dose of 1.5 g/kg of ingested DM. 2 ST = effects of the type of supplement as mineral or energy supplement. PHA = effect of the addition of phytogenic compounds blend, no effects, or tendencies of PHA × time and ST × PHA × Time interactions were observed (P>0.10). SEM = Standard error of the mean. Nmic = ruminal microbial nitrogen synthesis, Pmic = ruminal microbial protein synthesis, OMFR = digestible organic matter apparently fermented in the rumen, DOM = digestible organic matter.
Figure 1.
Interaction effect of type of supplement × time after supplementation on the ruminal proportion of acetate (P<0.001), butyrate (P<0.001), valerate (P<0.001), and iso-valerate (P=0.008) in Nellore cattle grazing Urochloa brizantha cv. Marandu supplemented with energy supplements or mineral supplements.
Figure 1.
Interaction effect of type of supplement × time after supplementation on the ruminal proportion of acetate (P<0.001), butyrate (P<0.001), valerate (P<0.001), and iso-valerate (P=0.008) in Nellore cattle grazing Urochloa brizantha cv. Marandu supplemented with energy supplements or mineral supplements.
3.4. Ruminal microbiota population
The richness index (ACE and Chao 1) and diversity estimators (Fisher, Simpson, and Shannon) of the ruminal microbial population were not affected by supplements (P>0.10; Table S1). The beta diversity Bray-Curtis distance showed that rumen microbial communities were different as a function of supplement type (P=0.015; Figure 2).
Eleven phyla were identified (Table 5), and 12.66 ± 2.05% of the OTUs identified in each sample could not be classified at the phylum level. In the Archaea domain, only the phylum Euryarchaeota was identified and was not influenced by the supplements tested. The ruminal abundance of Proteobacteria and Spirochaetae phyla were higher in steers supplemented with energy supplements than in steers supplemented with mineral supplements (P<0.05). Steers supplemented with energy supplements showed a tendency to have a higher ruminal abundance of Bacteroidetes (P=0.089) and showed a lower ruminal abundance of Firmicutes (P=0.069) than steers supplemented with mineral supplements. Steers supplemented with PHA presented a lower ruminal abundance of Verrucomicrobia phylum than steers fed supplements without PHA (P = 0.022).
At the family level, 16 bacterial families were identified, the most abundant were Lachnospiraceae (17.73%), Ruminococcaceae (14.90%), and Prevotellaceae (13.46%). The interaction between the type of supplement provided and the use of phytogenic compounds influenced the relative ruminal abundance of Bacteroidales S24-7 group, and Veillonellaceae families (Figure 3A). Thus, the relative abundance of the Veillonellaceae family was lower in the rumen of steers supplemented with EPHA when compared with other supplements (P = 0.007), and the Bacteroidales S24-7 group (P = 0.084) tended to be lower when compared with steers supplemented with MW. In addition, steers supplemented with PHA showed a higher ruminal abundance of families Bacteroidales BS11 gut group (P = 0.024).
The ruminal abundance of three Archaea OTU was influenced at the genera level by the interaction type of supplement × the addition of PHA (Figure 3B). Thus, the relative abundance of the Methanosphaera was lower in the rumen of steers supplemented with MW when compared to other supplements (P=0.041). Thermoplasmatales Incertae Sedis abundance was lower in the rumen of steers supplemented with MW when compared to EW and EPHA supplements (P=0.011). While uncultured archaeon from Thermoplasmatales was not identified in the rumen of steers supplemented with MPHA, its abundance was lower in the rumen of steers supplemented with MW when compared to EW and EPHA supplements (P=0.018).
Figure 3.
Relative abundance of operational taxonomic units (OTU) classified at family (A) and genera level of ruminal Archaea (B) and bacteria (C) domain in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season influencing by the supplementation with energy supplement without phytogenic compounds addition (EW), energy supplement with phytogenic compounds addition (EPHA), mineral supplement without phytogenic compounds addition (MW), or mineral supplement with phytogenic compounds addition (MPHA). Just significative (P < 0.05) or tendencies (P < 0.10) values are shown.
Figure 3.
Relative abundance of operational taxonomic units (OTU) classified at family (A) and genera level of ruminal Archaea (B) and bacteria (C) domain in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season influencing by the supplementation with energy supplement without phytogenic compounds addition (EW), energy supplement with phytogenic compounds addition (EPHA), mineral supplement without phytogenic compounds addition (MW), or mineral supplement with phytogenic compounds addition (MPHA). Just significative (P < 0.05) or tendencies (P < 0.10) values are shown.
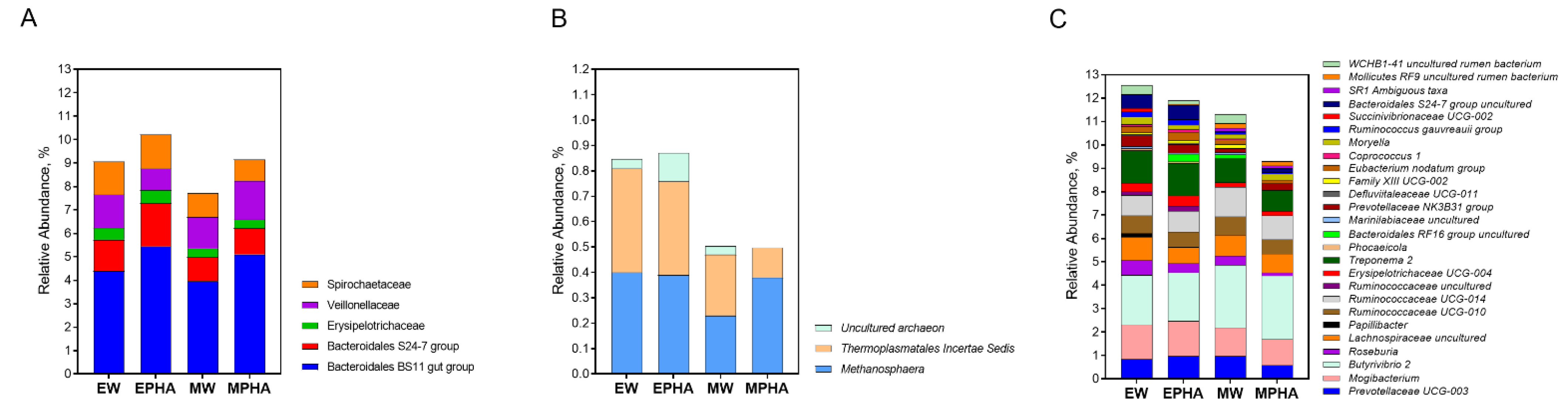
Several bacterial OTUS at the genera level were influenced by the interaction between the type of supplement × the addition of PHA (Figure 3C). Both Roseburia (P = 0.033) and Family XIII UCG-002 (P = 0.034) abundance was lower in the rumen of steers supplemented with MPHA when compared with those supplemented with EW and MW, respectively. The relative abundance of the genus Moryella was higher in steers supplemented with EW than in other groups (P = 0.044). The ruminal abundance of Succinivibrionaceae UCG-002 was lower in steers supplemented with MW when compared to both energy supplements EW and EPHA (P = 0.045). A higher ruminal abundance of SR1 Ambiguous taxa was observed in the rumen of steers supplemented with both mineral supplements (MW and MPHA) when compared to steers supplemented with EPHA (P = 0.017). The relative abundance of OTU class WCHB1-41 uncultured rumen bacterium was higher in the rumen of steers supplemented with EW when compared to both supplements with PHA (P = 0.041). The relative abundance of Prevotellaceae UCG-003 tended to be lower in the rumen of animals that received MPHA when compared to MW and EPHA (P = 0.071). Bacteroidales RF16 group uncultured (P = 0.060) and Ruminococcaceae uncultured (P = 0.071) tended to be lower in the rumen of animals supplemented with EW when compared to those that received EPHA and MW. A tendency of a lower ruminal abundance of the genus Prevotellaceae NK3B31 group (P = 0.076) and Treponema 2 (P = 0.058) was also observed in MW when compared to steers supplemented with EW.
In addition, energy supplements allowed a lower ruminal abundance of genera Butyrivibrio 2 (P = 0.033), Mollicutes RF9 uncultured rumen bacterium (P=0.038), and Ruminococcaceae UCG-014 (P = 0.040), and a higher ruminal abundance of genera Papillibacter (P = 0.031), Erysipelotrichaceae UCG-004 (P = 0.047), and Phocaeicola (P = 0.030). There was also a trend towards an increase in the abundance of Bacteroidales S24-7 group (P = 0.056), Mogibacterium (P = 0.063), Defluviitaleaceae UCG-011 (P = 0.056), Eubacterium nodatum group (P = 0.082), Coprococcus 1 (P = 0.056), Ruminococcus gauvreauii group (P = 0.090) in the rumen of steers supplemented with energy supplements when compared to those supplemented with mineral supplements.
Steers supplemented with PHA showed a lower ruminal abundance of Ruminiclostridium 6 when compared to those supplemented without PHA (P = 0.011). The ruminal abundance of Ruminococcaceae UCG-010 (P = 0.089), Lachnospiraceae uncultured (P = 0.056), and Marinilabiaceae uncultured (P = 0.075) tended to be lower in the rumen of steers supplemented with PHA when compared to steers fed supplements without PHA.
Through PCA was possible to extract the most important variables that explained 68.1% of the total variance and revealed a separation among animals supplemented with MW and other supplements evaluated (Figure 4). Steers supplemented with MW were distributed mainly in the negative region of dimensions 1 and showed a positive association with the KEGG functional pathways predicted metabolism of pyruvate (r=0.71), propionate (r=0.78), butyrate (r=0.86), glycerophospholipids (r=0.74), membrane transporters (r=0.98), the ruminal acetate concentration (r=0.61), and members of the phylum Firmicutes (r=0.87) and family Christensenellaceae (r=0.68). In contrast, steers supplemented with MPHA were distributed in positive regions of both dimensions 1 and 2, and were positively associated with 17 variables as the metabolism of purine (r=0.92), energy (r =0.88), nitrogen (r =0.59), cyano amino acids (r=0.84), alanine, aspartate, and glutamate (r=0.78), and glycine, serine, and threonine (r=0.89), the biosynthesis of lipopolysaccharide (r=0.84), vitamin B5 and CoA (r= 0.75), folic acid (r=0.87), phenylalanine, tyrosine, and tryptophan (r=0.67), the degradation of glycosaminoglycan (r=0.77) and other glycans (r=0.65), the DNA replication (r=0.64), the ruminal butyrate concentration (r=0.64), peptidases (r=0.72), the ruminal abundance of the phylum Bacteroidetes (r=0.82), and Prevotellaceae family (r=0.66).
In addition, steers supplemented with EW were negatively associated with members of the Lachnospiraceae family (r=0.65), Lachnospiraceae 10 (r=0.73), and Butyrivibrio 2 (r=0.60). Also were positively associated with the predicted KEGG functional pathways of metabolism pyrimidine (r=0.93), vitamin B2 (r=0.85), CH4 (r=0.66), cofactors and vitamins (r=0. 75), D-glutamine and D-glutamate (r=0.63), cysteine and methionine (r=0.70), degradation (r=0.60) and biosynthesis (r=0.56) of valine, leucine and isoleucine, carbon fixation pathways in prokaryotes (r=0.94), amino acid-related enzymes (r=0.95), the tricarboxylic acid cycle (r=0.93) and the ruminal abundance of Archaea as the phylum Euryarchaeota (r= 0.71) and Methanobrevibacter (r=0.66).
4. Discussion
It is well known that supplementation can modulate the forage intake in grazing cattle by associative effects as substitutive, additive, or combined, due to metabolic and digestive interactions [46]. In the present study, the energy supplementation trend allows an additive effect on forage intake and consequently, increased the total DM intake and higher intake of OM, CP, apNDF, GE, digestible energy, and metabolizable energy. A similar additive effect was reported in young beef bulls grazing U. brizantha cv Xaraés with energy-protein supplementation at a similar rate than used in this study [47]. Due to the greater input of nitrogen from the energy supplements, there was also a greater excretion of urinary N and a trend toward greater excretion of N in the feces. However, these steers also had a higher proportion of N retained/N ingested. The efficiency of nitrogen use in ruminants shows high variation, being able to change from 15% to 40% [48]. Hoffmann et al. [49] reported an efficiency of nitrogen use of 42.94% in young Nellore bulls grazing tropical pastures intensively managed and supplemented with 0.3% BW, possibly due to greater synchrony between the degradation of carbohydrates and nitrogenous compounds in the rumen.
Among the various factors that can influence the production of volatile fatty acid in the rumen are the ruminal microbiota population. The positive association observed in the rumen of steers supplemented with MW between the ruminal acetate concentration and the Firmicutes phylum is related to the higher proportion of acetate verified in the rumen of these animals and the tendency to a higher ruminal abundance of Firmicutes when compared to energy supplements. The association of the Firmicutes and Bacteroidetes phyla abundance and the A:P ratio in the rumen were previously observed by Min et al. [50]. Then, steers that were fed energy supplements had a higher proportion of propionate in the rumen, and a lower A:P ratio 3h after supplementation. Among the energy supplement ingredients, corn gluten meal has an energy value equivalent to 92-95% of corn with high levels of isoleucine (0.73%), leucine (2.15%), and valine (1.19%) [51,52], which could be used as a substrate to propionate production in the rumen. Thus, an improvement in ruminal fermentation allows a greater animal performance [23]. In addition, ruminal valerate is a glycogenic fatty acid [53], and the highest concentration of valerate (1.40%) and iso-valerate (2.14%) 3h after supplementation in the rumen of steers that received energy supplements can be justified by the deamination of these amino acids as substrate for the formation of branched-chain fatty acid [54]. The PCA corroborates these results, as it identified a positive association between the energy supplement and the degradation and biosynthesis of valine, leucine, and isoleucine and the metabolism of other amino acids such as D-glutamine and D-glutamate, cysteine, and methionine. Similarly, Camargo et al. [52] reported that grazing beef cattle supplemented with corn gluten meal had an increase in the ruminal molar proportion of valerate and iso-valerate compared with cattle with ad libitum mineral supplementation.
The inclusion of PHA in the supplement allows a lower ruminal proportion of Valerate and tends to increase the ruminal proportion of propionate. Similarly, Yang et al. [13] observed that the supply of tannic acid at doses of 6.5, 13.0, and 26.0 g/kg DM resulted in an increase in the proportion of propionate and a reduction in the proportion of valerate, and the results were also monitored by reduction of enteric methane emissions in cattle. In vitro approach also supports the reduction of valerate proportion with increasing doses of tannin extracts while increasing the Propionate proportion during rumen fermentation [55]. Steers supplemented with EPHA showed a positive association with the Bacteroidetes phylum and the Prevotellaceae family in the PCA, this may be related to the lower A:P ratio observed in the rumen of these steers. Bacteroidetes were also associated by Min et al. [50] with enhanced propionate production and the reduction in the A:P ratio in goats. According to Calsamiglia et al. [18], compounds such as thymol and carvacrol do not selectively inhibit Gram-positive or Gram-negative bacteria. Some specific compounds of essential oils, such as carvacrol, can begin loss of cellular content and lysis of Gram-negative bacteria cells due to the presence of the carbonyl group [56]. Bacteroidetes phylum members are generally Gram-negative, therefore, no effect of the carvacrol supply in the PHA was verified on these bacterial groups. Either were not detected effects on ruminal butyrate production, contrary to those observed by Garcia et al. [19] in an in vitro study evaluating the addition of 250 mg/L of carvacrol. However, the concentration of carvacrol used in the present study may not be sufficient to cause this effect.
The different supplements evaluated provided the potential to modify ruminal NH3-N concentration, since hydrolyzable tannins can modulate protein-degrader microorganisms, decreasing the degradation of soluble protein in the rumen, and potentially improving nitrogen use [57]. However, in this study, the PHA inclusion did not affect the ruminal concentration of NH3-N. But a higher NH3-N concentration was observed in the rumen of steers supplemented with energy supplements when compared to mineral supplements. This is associated with the energy supplements composition, once ingredients such as corn gluten meal allowed higher CP intake and consequently, higher protein degradation in the rumen. In this sense, the relationship between the CP content of the diet and the amount of digestible organic matter (DOM) could influence the efficiency of the transfer of ingested CP to the intestine. According to Poppi and McLennan [58], a higher efficient transfer occurs when these values are lower than 160 g CP/kg DOM, and values higher than 210 g CP/kg DOM may result in losses and/or incomplete transfer. In the present study, U. brizantha cv. Marandu intensively managed, allowing CP values above 15% and values higher than 210 g CP/kg MOD (around 254.8 g CP/ kg MOD). In addition, serum urea N levels may be indicative of both the protein status in the animal and the maximum microbial efficiency. According to Vendramini et al. [59], serum urea N levels between 15 and 19 mg/dL are considered threshold levels to prevent protein loss. Thus, the highest ruminal NH3-N concentration corroborates the premise of a higher concentration of N-urea in the serum [60], withal, the levels were close to the threshold even steers that received energy supplements (around 15.98 mg/dL).
The ability to form complexes with proteins, which could lead to a lower ruminal NH3-N concentration and a higher flux of rumen undegraded protein to the small intestine, is generally greater for animals supplemented with hydrolyzable tannins with higher molecular weight [61]. However, both Nmic and efficiency of microbial protein synthesis were not influenced by the PHA inclusion in the supplements, and the average value of microbial efficiency observed (15.21 g Nmic/kg OMFR) was below 30 g Nmic/kg OMFR, the value recommended by the ARC [35]. The efficiency of microbial protein synthesis details the energy toward nitrogen assimilation by rumen microorganisms, lower values are representative of cattle-fed tropical conditions due to the nutritional unbalance of pastures [23].
The inclusion of PHA in the supplements reduced the ruminal relative abundance of the phylum Verrucomicrobia. This phylum is more abundant in ruminants fed forage diets and its low tolerance to tannins extracts has been reported [62,63]. The PHA did not influence the abundance of other phyla, contrary to the observed in grazing goats fed extracts of chestnut and quebracho, which reduced the abundance of Firmicutes and increased the abundance of Bacteroidetes [64], and in grazing cattle supplemented with condensed tannin from quebracho and hydrolyzable tannins from Castanea.spp, which reduced the relative ruminal abundances of Firmicutes, Fusobacteria, and Fibrobacteres, and increased the relative abundances of Armatimonadetes and Actinobacteria [5]. At the family level, the PHA promoted the ruminal abundance of Bacteroidales BS11 gut group, a common and abundant group of bacteria of the rumen, specialized to ferment different hemicellulosic monomers producing acetate and butyrate [65].
In addition, the PHA allowed a reduction of genera Ruminiclostridium 6, Ruminococcaceae UCG-010, and Lachnospiraceae uncultured belonging to the Firmicutes phylum, Clostridiales order, and typical of cellulolytic bacteria [66]. This deleterious effect of PHA also was observed in the rumen of steers supplemented with EPHA, which had a lower relative abundance of both genera Roseburia, Family XIII UCG-002, and the Veillonellaceae family, and may be associated with the lower total digestibility of DM observed with the addition of PHA in this study and the lower fiber digestibility observed by Teobaldo et al. [22] in performance trial. Witzig et al. [8] also observed a reduction in the relative abundance of Ruminococcus albus by chestnut and valonea tannins. The inclusion of hydrolyzable tannins from chestnut tannin extracts also reduced the abundance of Ruminococcus flavefaciens [6].
The lower ruminal abundance of Archaea genus Methanosphaera in steers supplemented with MW may be associated with the fact that Methanosphaera uses methyl groups derived from methoxyl substitutes of plant material as the main substrate, thus, the production of CH4 derived from the methyl group is limited by the availability of methyl donors, and the relative abundance of Methanosphaera was negatively related to CH4 production in sheep [67]. In contrast, steers supplemented with EW were positively associated with the ruminal abundance of Archaea as phylum Euryarchaeota and the genus Methanobrevibacter, and CH4 metabolism. This can be justified by the sulfur content of the corn gluten meal, due to the industrial process for obtaining that uses sulphited water in one of its stages, because many Archaea use sulfur compounds as electron donors or acceptors for energy production [68]. This also is in line with the increase in the enteric CH4 daily production reported in grazing beef cattle when supplemented with energy supplements [22]. In contrast, Methanobrevibacter abundance was reduced when 1.5 g of hydrolyzable tannins were evaluated in vitro [7], indicating that tannin extract also can affect the archaea population in the rumen. In this study, the negative effect of PHA on the archaea population was observed just on an uncultured archaeon OTU belonging to Thermoplasmatales order.
5. Conclusions
The inclusion of a blend of phytogenic compounds containing hydrolyzable tannins, carvacrol, and cinnamaldehyde oil in the supplement of grazing Nellore steers did not influence the nitrogen balance and efficiency of ruminal microbial nitrogen synthesis, even tending to increase the ruminal production of propionate. The inclusion of a blend of phytogenic compounds negatively affected the total dry matter digestibility and the abundance of several ruminal bacterial groups belonging to the Firmicutes and Verrucomicrobia phyla.
Energy supplementation is an effective strategy to improve the intake of nutrients and nitrogen efficiency in grazing Nellore steers during the rainy season by the modulation of ruminal fermentation parameters and bacteria diversity, leading to an increase in the nitrogen retained and propionate, valerate, and iso-valerate production in the rumen.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Median ± interquartile range of alpha diversity as richness index (ACE and Chao 1) and diversity estimators (Fisher, Simpson, and Shannon) of ruminal microbial population in Nellore cattle grazing Urochloa brizantha cv. Marandu and supplemented or not with phytogenic compounds during the rainy season.
Author Contributions
Conceptualization, R.W.T., A.d.S.C., and R.A.R.; methodology Y.T.G.-S and R.A.R.; formal analysis R.W.T., M.T.L.C., and Y.T.G.-S; investigation R.W.T., T.R.B., E.P.R., and A.d.S.C.; writing—original draft preparation R.W.T. and Y.T.G.-S; writing—review and editing R.W.T., A.d.S.C., Y.T.G.-S., and R.A.R.; visualization Y.T.G.-S; supervision R.A.R.; project administration A.d.S.C. and R.A.R.; funding acquisition R.A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fundação de Amparo à Pesquisa do Estado de São Paulo (Grants numbers: 16/16631-5 and 17/11274-5). The authors received scholarships of Coordenação de Aperfeiçoamento do Pessoal de Nível Superior and Conselho Nacional de desenvolvimento e Tecnologico.
Data Availability Statement
Data will be made available upon request directly to the authors.
Acknowledgments
This work was conducted as a part of the first author’s thesis. We are grateful for the help of UnespFor (study group) for their assistance in conducting the experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gomes, K.L.C.A.; Granja-Salcedo, Y.T.; Messana, J.D.; Carneiro de Souza, V.; Generoso Ganga, M.J.; Detogni Colovate, P.H.; Kishi, L.T.; Berchielli, T.T. Rumen bacterial diversity in relation to nitrogen retention in beef cattle. Anaerobe 2021, 67, 102316. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, L.O.; Callaway, T.R.; Muir, J.P.; Anderson, R.C. Potential environmental benefits of feed additives and other strategies for ruminant production. Rev. Bras. Zootec. 2011, 40, 291–309. [Google Scholar]
- Clark, S.; Daly, R.; Jordan, E.; Lee, J.; Mathew, A.; Ebner, P. The future of biosecurity and antimicrobial use in livestock production in the United States and the role of extension. J. Anim. Sci. 2012, 90, 2852–2861. [Google Scholar] [CrossRef] [PubMed]
- Aboagye, I.A.; Oba, M.; Castillo, A.R.; Koenig, K.M.; Iwaasa, A.D.; Beauchemin, K.A. Effects of hydrolyzable tannin with or without condensed tannin on methane emissions, nitrogen use, and performance of beef cattle fed a high-forage diet. J. Anim. Sci. 2018, 96, 5276–5286. [Google Scholar] [CrossRef]
- Cidrini, I.A.; Granja-Salcedo, Y.T.; Prados, L.F.; Kishi, L.T.; Siqueira, G.R.; Resende, F.D. Effect of tannin extract associated with two levels of non-protein nitrogen in the supplement on performance, ruminal parameters, and microbial diversity of grazing Nellore cattle during the growing phase at dry season. Anim. Feed Sci. Technol. 2022, 286, 115269. [Google Scholar] [CrossRef]
- Jayanegara, A.; Goel, G.; Makkar, H.P.S.; Becker, K. Divergence between purified hydrolysable and condensed tannin effects on methane emission, rumen fermentation and microbial population in vitro. Anim. Feed Sci. Technol. 2015, 209, 60–68. [Google Scholar] [CrossRef]
- Witzig, M.; Zeder, M.; Rodehutscord, M. Effect of the ionophore monensin and tannin extracts supplemented to grass silage on populations of ruminal cellulolytics and methanogens in vitro. Anaerobe 2018, 50, 44–54. [Google Scholar] [CrossRef]
- Krueger, W.K.; Gutierrez-Banuelos, H.; Carstens, G.E.; Min, B.R.; Pinchak, W.E.; Gomez, R.R.; Anderson, R.C.; Krueger, N.A.; Forbes, T.D.A. Effects of dietary tannin source on performance, feed efficiency, ruminal fermentation, and carcass and non-carcass traits in steers fed a high-grain diet. Anim. Feed Sci. Technol. 2010, 159, 1–9. [Google Scholar] [CrossRef]
- Toral, P.G.; Hervas, G.; Bichi, E.; Belenguer, A.; Frutos, P. Tannins as feed additives to modulate ruminal biohydrogenation: Effects on animal performance, milk fatty acid composition and ruminal fermentation in dairy ewes fed a diet containing sunflower oil. Anim. Feed Sci. Technol. 2011, 164, 199–206. [Google Scholar] [CrossRef]
- Stewart, E.K.; Beauchemin, K.A.; Dai, X.; MacAdam, J.W.; Christensen, R.G.; Villalba, J.J. Effect of tannin-containing hays on enteric methane emissions and nitrogen partitioning in beef cattle. J. Anim. Sci. 2019, 97, 3286–3299. [Google Scholar] [CrossRef]
- Nelson, K.E.; Pell, A.N.; Schofield, P.; Zinder, S. Isolation and Characterization of an Anaerobic Ruminal Bacterium Capable of Degrading Hydrolyzable Tannins. Appl. Environ. Microbiol. 1995, 61, 3293–3298. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Alves, S.P.; Cappucci, A.; Cook, S.R.; Duarte, A.; Caldeira, R.M.; McAllister, T.A.; Bessa, R.J.B. Effects of condensed and hydrolysable tannins on rumen metabolism with emphasis on the biohydrogenation of unsaturated fatty acids. J. Agric. Food Chem. 2018, 66, 3367–3377. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wei, C.; Zhao, G.Y.; Xu, Z.W.; Lin, S.X. Effects of dietary supplementing tannic acid in the ration of beef cattle on rumen fermentation, methane emission, microbial flora and nutrient digestibility. J. Anim. Physiol. Anim. Nutr. 2016, 101, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Aboagye, I.A.; Oba, M.; Koenig, K.M.; Zhao, G.Y.; Beauchemin, K.A. Use of gallic acid and hydrolyzable tannins to reduce methane emission and nitrogen excretion in beef cattle fed a diet containing alfalfa silage. J. Anim. Sci. 2019, 97, 2230–2244. [Google Scholar] [CrossRef]
- Patra, A.K.; Yu, Z. Effects of essential oils on methane production and fermentation by and abundance and diversity of, rumen microbial populations. Appl. Environ. Microbiol. 2012, 78, 4271–4280. [Google Scholar] [CrossRef]
- Geraci, J.I.; Garciarena, A.D.; Gagliostro, G.A.; Beauchemin, K.A.; Colombatto, D. Plant extracts containing cinnamaldehyde, eugenol and capsicum oleoresin added to feedlot cattle diets Ruminal environment, short term intake pattern and animal performance. Anim. Feed Sci. Technol. 2012, 176, 123–130. [Google Scholar] [CrossRef]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Kamel, C. Plant extracts effect in vitro rumen microbial fermentation. J. Dairy Sci. 2006, 89, 761–771. [Google Scholar] [CrossRef]
- Calsamiglia, S.; Busquet, M.; Cardozo, P.W.; Castillejos, L.; Ferret, A. Essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 2007, 90, 2580–2595. [Google Scholar] [CrossRef]
- García, V.; Catalá-Gregori, P.; Madrid, J.; Hernández, F.; Megías, M.D.; Andrade-Montemayor, H.M. Potential of carvacrol to modify in vitro rumen fermentation as compared with monensin. Animal 2007, 1, 675–680. [Google Scholar] [CrossRef]
- Durmic, Z.; Moate, P.J.; Eckard, R.; Revell, D.K.; Williams, R.; Vercoe, P.E. In vitro screening of selected feed additives, plant essential oils and plant extracts for rumen methane mitigation. J. Sci. Food Agric. 2014, 94, 1191–1196. [Google Scholar] [CrossRef]
- Rossi, C.A.S.; Grossi, S.; Dell’Anno, M.; Compiani, R.; Rossi, L. Effect of a Blend of Essential Oils, Bioflavonoids and Tannins on In Vitro Methane Production and In Vivo Production Efficiency in Dairy Cows. Animals 2022, 12, 728. [Google Scholar] [CrossRef]
- Teobaldo, R.W.; Cardoso, A.d.S.; Brito, T.R.; Leite, R.G.; Romanzini, E.P.; Granja-Salcedo, Y.T.; Reis, R.A. Response of Phytogenic Additives on Enteric Methane Emissions and Animal Performance of Nellore Bulls Raised in Grassland. Sustainability 2022, 14, 9395. [Google Scholar] [CrossRef]
- Detmann, E.; Valente, É.E.L.; Batista, E.D.; Huhtanen, P. An evaluation of the performance and efficiency of nitrogen utilization in cattle fed tropical grass pastures with supplementation. Livest. Sci. 2014, 162, 141–153. [Google Scholar] [CrossRef]
- Mott, G.O.; Lucas, H.L. The design conduct and interpretation of grazing trials on cultivated and improved pastures. In Proceedings of the 6th International Grassland Congress, State College, PA, USA, 17–23 August 1952. [Google Scholar]
- Halls, L.K. The approximation of cattle diet through herbage sampling. Rang. Ecol. Manag. 1954, 7, 269–270. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists; Association of Official Agricultural Chemists: Rockville, MD, USA, 1990; pp. 1–771. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, and no starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Goering, H.K.; Van Soest, P.J. Forage Fibre Analyses (Apparatus. Reagents. Procedures. and Some Applications); Agriculture Handbook No 379; Agricultural Research Service: Washington, DC, USA, 1970.
- Valente, T.N.P.; Detmann, E.; Queiroz, A.C.D.; Valadares Filho, S.D.C.; Gomes, D.I.; Figueiras, J.F. Evaluation of ruminal degradation profiles of forages using bags made from different textiles. Rev. Bras. Zootec. 2011, 40, 2565–2573. [Google Scholar] [CrossRef]
- Souza, N.K.P.; Detmann, E.; Pina, D.S.; Valadares Filho, S.C.; Sampaio, C.B.; Queiroz, A.C.; Veloso, C.M. Evaluation of chromium concentration in cattle feces using different acid digestion and spectrophotometric quantification techniques. Arq. Bras. Med. Vet. Zoote. 2013, 65, 1472–1482. [Google Scholar] [CrossRef]
- Chen, X.B.; Gomes, M.J. Estimation of Microbial Protein Supply to Sheep and Cattle Based on Urinary Excretion of Purine Derivatives—An Overview of the Technical Details; Occasional Publication; Rowett Research Institute: Bucksburn Aberdeen, Scotland, 1995. [Google Scholar]
- Costa e Silva, L.F.; Valadares Filho, S.C.; Chizzotti, M.L.; Rotta, P.P.; Prados, L.F.; Valadares, R.F.D.; Zanetti, D.; Braga, J.M.S. Creatinine excretion and relationship with body weight of Nellore cattle. Rev. Bras. Zootec. 2012, 41, 807–810. [Google Scholar] [CrossRef]
- Verbic, J.; Chen, X.; MacLeod, N.; Ørskov, E. Excretion of purine derivatives by ruminants. Effect of microbial nucleic acid infusion on purine derivative excretion by steers. J. Agric. Sci. 1990, 114, 243–248. [Google Scholar] [CrossRef]
- Barbosa, A.M.; Valadares, R.F.D.; Valadares Filho, S.C.; Pina, D.S.; Detmann, E.; Leão, M.I. Endogenous fraction and urinary recovery of purine derivatives obtained by different methods in Nellore cattle. J. Anim. Sci. 2011, 89, 510–519. [Google Scholar] [CrossRef]
- ARC. The Nutrient Requirements of Ruminant Livestock; Technical review by an Agricultural Research Council Working Party; Commonwealth Agricultural Bureau: Farnham Royal, UK, 1984. [Google Scholar]
- Famme, P.; Knudsen, J. Total heat balance study of anaerobiosis in Tubiflex (Muller). J. Comp. Physiol. 1984, 154, 587–591. [Google Scholar] [CrossRef]
- Granja-Salcedo, Y.T.; Ramirez-Uscategui, R.A.; Machado, E.G.; Messana, J.D.; Kishi, L.T.; Dias, A.V.L.; Berchielli, T.T. Studies on bacterial community composition are affected by the time and storage method of the rumen content. PLoS ONE 2017, 12, e0176701. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.; Walters, W.A.; Lyons, D.B.; Luzupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2010, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible interactive scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.; McMurdie, P.; Rosen, M.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Opens external link in new window. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 8, 1–10. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Moore, J.E. Forage crops. In Crop Quality, Storage, and Utilization; Hoveland, C.S., Ed.; Crop Science Society of America: Madison, WI, USA, 1980; pp. 61–91. [Google Scholar]
- Simioni, T.A.; Messana, J.D.; Silva, L.G.; Granja-Salcedo, Y.T.; Torrecilhas, J.A.; San Vito, E.; Lage, J.F.; Reis, R.A.; Berchielli, T.T. Effects of mineral or protein-energy supplementation and genetic group on metabolism parameters of young beef bulls grazing tropical grass during the rainy season. Livest. Sci. 2022, 255, 104805. [Google Scholar] [CrossRef]
- Calsamiglia, S.; Ferret, A.; Reynolds, C.K.; Kristensen, N.B.; Van Vuuren, A.M. Strategies for optimizing nitrogen use by ruminants. Animal 2010, 4, 1184–1196. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.; Cardoso, A.S.; Fonseca, N.V.B.; Romanzini, E.P.; Siniscalchi, D.; Berndt, A.; Ruggieri, A.C.; Reis, R.A. Effects of supplementation with corn distillers’ dried grains on animal performance, nitrogen balance, and enteric CH4 emissions of young Nellore bulls fed a high-tropical forage diet. Animal 2021, 15, 100155. [Google Scholar] [CrossRef]
- Min, B.R.; Gurung, N.; Shange, R.; Solaiman, S. Potential role of rumen microbiota in altering average daily gain and feed efficiency in meat goats fed simple and mixed pastures using bacterial tag-encoded FLX amplicon pyrosequencing. J. Anim. Sci. 2019, 97, 3523–3534. [Google Scholar] [CrossRef]
- Firkins, J.L.; Berger, L.L.; Fahey, G.C. Evaluation of wet and dry distillers grains and dry corn gluten feeds for ruminants. J. Anim. Sci. 1985, 60, 847–860. [Google Scholar] [CrossRef]
- Camargo, K.D.V.; Messana, J.D.; Silva, L.G.; Granja-Salcedo, Y.T.; Dias, A.V.L.; Alves, K.L.G.C.; Gonçalves, P.H.; Souza, W.A.; Reis, R.A.; Berchielli, T.T. Intake, metabolism parameters, and performance of growing beef cattle on pasture supplemented with different rumen undegradable protein with different amino acid profile. Anim. Feed Sci. Technol. 2022, 286, 115258. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Merchen, N.R.; Titgemeyer, E.C. Manipulation of amino acid supply to the growing ruminant. J. Anim. Sci. 1992, 70, 3238–3247. [Google Scholar] [CrossRef] [PubMed]
- Menci, R.; Coppa, M.; Torrent, A.; Natalello, A.; Valenti, B.; Luciano, G.; Priolo, A.; Niderkorn, V. Effects of two tannin extracts at different doses in interaction with a green or dry forage substrate on in vitro rumen fermentation and biohydrogenation. Anim. Feed Sci. Technol. 2021, 278, 114977. [Google Scholar] [CrossRef]
- Cobellis, G.; Trabalza-Marinucci, M.; Marcotullio, M.C.; Yu, Z. Evaluation of different essential oils in modulating methane and ammonia production, rumen fermentation, and rumen bacteria in vitro. Anim. Feed Sci. Technol. 2016, 215, 25–36. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition—Review. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Poppi, D.P.; McLennan, S.R. Protein and energy utilization by ruminants at pasture. J. Anim. Sci. 1995, 73, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Vendramini, J.M.B.; Sanchez, J.M.D.; Cooke, R.F.; Aguiar, A.D.; Moriel, P.; da Silva, W.L.; Cunha, O.F.R.; Ferreira, P.D.S.; Pereira, A.C. Stocking rate and monensin supplemental level effects on growth performance of beef cattle consuming warm-season grasses. J. Anim. Sci. 2015, 93, 3682–3689. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.M.; Milligan, L.P. The degradation and utilization of endogenous urea the gastrointestinal tract of ruminants: A review. Can. J. Anim. Sci. 1980, 60, 205–221. [Google Scholar] [CrossRef]
- Kawamoto, H.; Nakatsubo, F.; Murakami, K. Quantitative determination of tannin and protein in the precipitates by high-performance liquid chromatography. Phytochemistry 1995, 40, 1503–1505. [Google Scholar] [CrossRef]
- Rabee, A.E.; Kewan, K. Z.; Lamara, M. Identification of Micro-Organisms that Tolerant to Anti-Nutritional Factors in the Rumen of Camel. J. Anim. Poult. Prod. 2022, 13, 7–13. [Google Scholar] [CrossRef]
- Auffret, M.D.; Dewhurst, R.J.; Duthie, C.A.; Rooke, J.A.; Wallace, R.J.; Freeman, T.C.; Stewart, R.; Watson, M.; Roehe, R. The rumen microbiome as a reservoir of antimicrobial resistance and pathogenicity genes is directly affected by diet in beef cattle. Microbiome 2017, 5, 159. [Google Scholar] [CrossRef]
- Min, B.R.; Wright, C.; Ho, P.; Eun, J.S.; Gurung, N.; Shange, R. The effect of phytochemical tannins-containing diet on rumen fermentation characteristics and microbial diversity dynamics in goats using 16S rDNA amplicon pyrosequencing. Agric. Food Anal. Bacteriol. 2014, 4, 195–211. [Google Scholar]
- Solden, L.M.; Hoyt, D.W.; Collins, W.B.; Plank, J.E.; Daly, R.A.; Hildebrand, E.; Beavers, T.J.; Wolfe, R.; Nicora, C.D.; Purvine, S.O.; et al. New roles in hemicellulosic sugar fermentation for the uncultivated Bacteroidetes family BS11. ISME J. 2017, 11, 691–703. [Google Scholar] [CrossRef]
- Wu, K.; Cheng, L. Ruminiclostridium. In Bergey’s Manual of Systematics of Archaea and Bacteria; Trujillo, M.E., Dedysh, S., DeVos, P., Hedlund, B., Kämpfer, P., Rainey, F.A., Whitman, W.B., Eds.; 2022. [Google Scholar]
- Kittelmann, S.; Pinares-Patino, C.S.; Seedorf, H.; Kirk, M.R.; Ganesh, S.; McEwan, J.C.; Janssen, P. Two different bacterial community types are linked with the low-methane emission trait in sheep. PLoS ONE. 2014, 9, e103171. [Google Scholar] [CrossRef]
- Kletzin, A.; Urich, T.; Müller, F.; Bandeiras, T.M.; Gomes, C.M. Dissimilatory oxidation and reduction of elemental sulfur in thermophilic archaea. J. Bioenerg. Biomembr. 2004, 36, 77–91. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Beta diversity Bray-Curtis index computed for nonparametric statistical tests between rumen microbial communities of Nellore cattle grazing Urochloa brizantha cv. Marandu and supplemented with energy supplements (SE) or mineral supplements (SM). There were no observed effects of the addition of a phytogenic compounds blend containing 10% of carvacrol and cinnamaldehyde oil, and 90% hydrolyzable tannins extracted from berries and grapes at a dose of 1.5 g/kg of ingested dry matter.
Figure 2.
Beta diversity Bray-Curtis index computed for nonparametric statistical tests between rumen microbial communities of Nellore cattle grazing Urochloa brizantha cv. Marandu and supplemented with energy supplements (SE) or mineral supplements (SM). There were no observed effects of the addition of a phytogenic compounds blend containing 10% of carvacrol and cinnamaldehyde oil, and 90% hydrolyzable tannins extracted from berries and grapes at a dose of 1.5 g/kg of ingested dry matter.
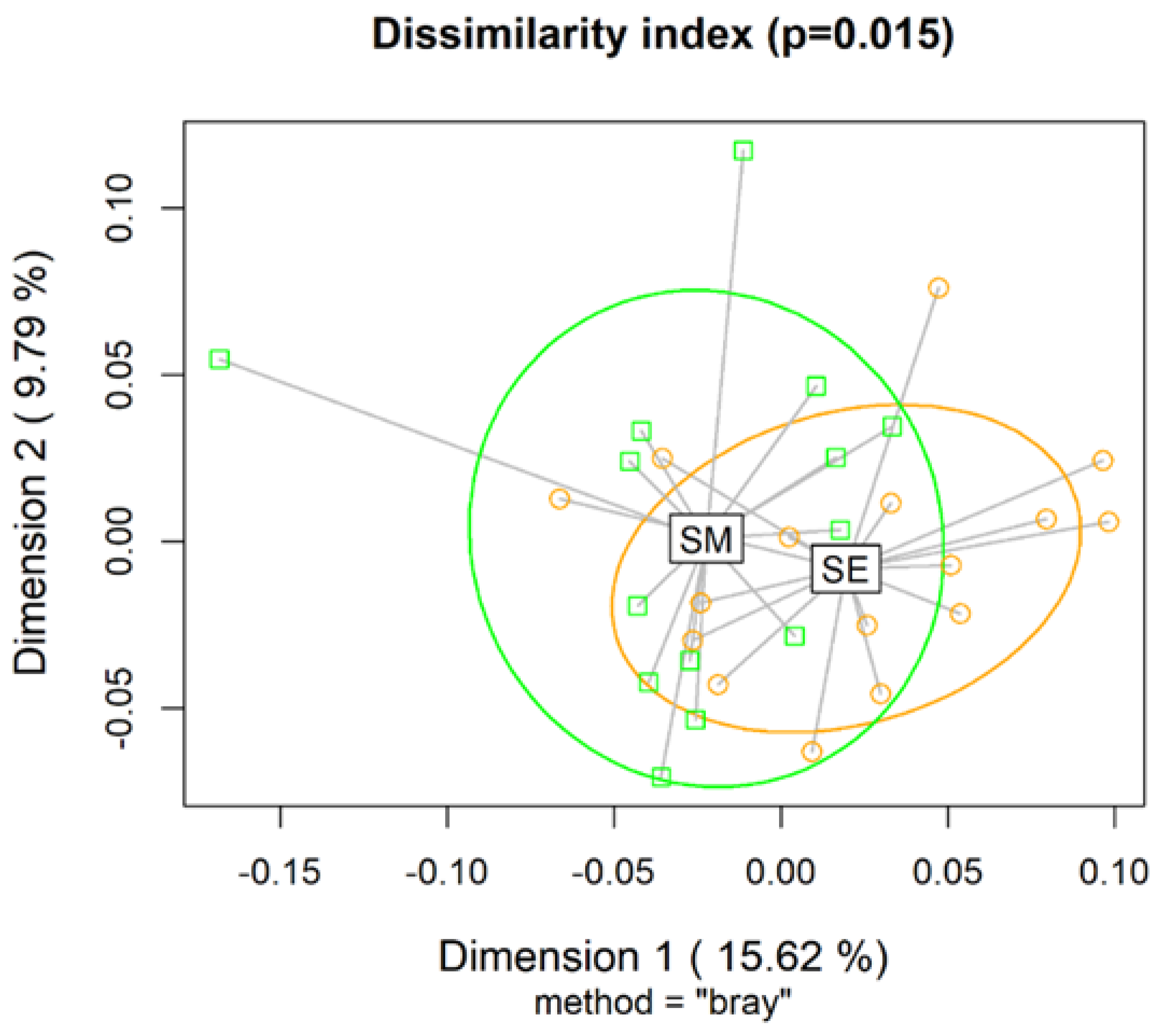
Figure 4.
Microbial operational taxonomic units associated with predicted functional genes, ruminal fermentation parameters, intake, digestibility, and N metabolism in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season influencing by the supplementation with energy supplement without phytogenic compounds addition (EW), energy supplement with phytogenic compounds addition (EPHA), mineral supplement without phytogenic compounds addition (MW), or mineral supplement with phytogenic compounds addition (MPHA). The ellipses represent the Euclidian distance among groups through principal component analysis. Blue variables represent higher contribution. CFP = Carbon fixation pathways in prokaryotes, M = metabolism, DEG = degradation, BIO = biosynthesis, AC = acid, AA = amino acids, TCA = tricarboxylic acid, PPP = pentose phosphate pathway.
Figure 4.
Microbial operational taxonomic units associated with predicted functional genes, ruminal fermentation parameters, intake, digestibility, and N metabolism in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season influencing by the supplementation with energy supplement without phytogenic compounds addition (EW), energy supplement with phytogenic compounds addition (EPHA), mineral supplement without phytogenic compounds addition (MW), or mineral supplement with phytogenic compounds addition (MPHA). The ellipses represent the Euclidian distance among groups through principal component analysis. Blue variables represent higher contribution. CFP = Carbon fixation pathways in prokaryotes, M = metabolism, DEG = degradation, BIO = biosynthesis, AC = acid, AA = amino acids, TCA = tricarboxylic acid, PPP = pentose phosphate pathway.
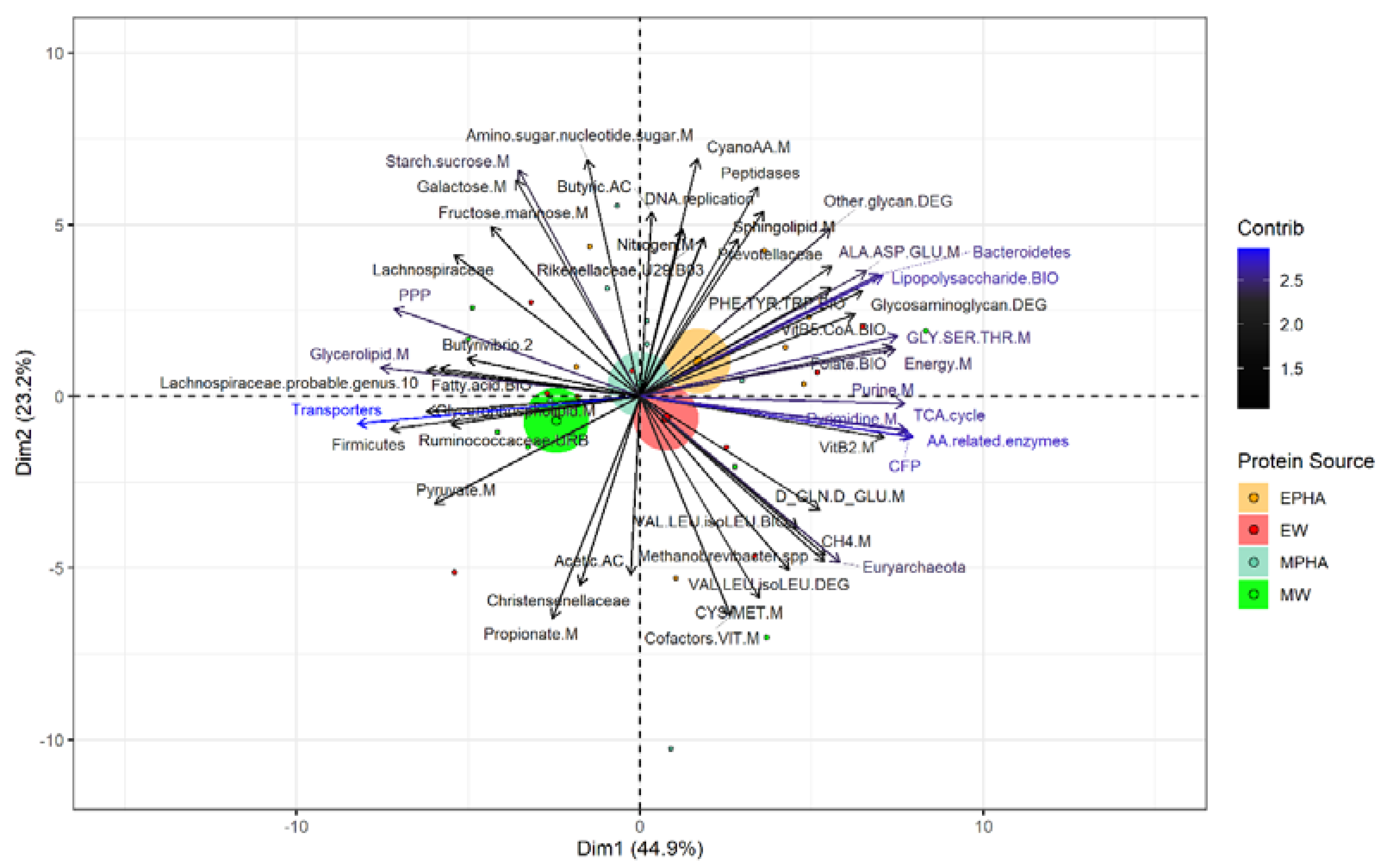
Table 1.
Chemical composition of forage (Urochloa brizantha cv. Marandu) and energy supplements.
| Chemical composition, % DM | Supplements1 | Forage2 | |||
|---|---|---|---|---|---|
| EW | EPHA | MW | MPHA | ||
| Mineral matter, % | 34.02 | 33.12 | - | - | 8.91±0.06 |
| Organic matter, % | 65.98 | 66.88 | - | - | 91.08±0.07 |
| apNDF3, % | 30.32 | 30.31 | - | - | 54.85±0.38 |
| iNDF4, % | 15.75 | 15.61 | - | - | 17.75±0.26 |
| Ether extract, % | 1.81 | 2.11 | - | - | 2.47±0.05 |
| Gross energy, MJ/kg DM | 12.58 | 13.33 | - | - | 17.95±0.04 |
| Crude protein, % | 14.87 | 15.02 | - | - | 15.28±0.22 |
| Fraction5, as % of crude protein | |||||
| A | 29.01 | 36.98 | - | - | 26.64 |
| B1 | 6.33 | 6.48 | - | - | 7.40 |
| B2 | 50.92 | 45.01 | - | - | 45.70 |
| B3 | 10.89 | 8.76 | - | - | 14.20 |
| C | 2.85 | 2.77 | - | - | 6.06 |
1 EW= energy supplement containing corn gluten meal and minerals without phytogenic compounds addition; EPHA= energy supplement containing corn gluten meal and minerals with phytogenic compounds addition (containing 10% of carvacrol and cinnamaldehyde oil, and 90% hydrolyzable tannins extracted from berries and grapes) at a dose of 1.5 g/kg of ingested DM. MW= mineral supplement warranty levels: Ca 123.0 g/kg; P 90.0 g/kg; Cu 1040.0 mg/kg; Mn 500 mg/kg; Zn 2000.0 mg/kg; Co 15.0 mg/kg; I 67.0 mg/kg; Se 14.0 mg/kg. MPHA= Guarantee levels similar to DM include phytogenic compounds at a dose of 1.5 g/kg of ingested DM. 2 Chemical composition of samples obtained by simulated grazing technique. 3apNDF: neutral detergent fiber corrected for ash and protein.4 iNDF: indigestible neutral detergent fiber. 5A = non-protein nitrogen, B1 = rapidly degradable protein in the rumen, B2 = moderately degradable protein in the rumen, B3 = slowly degradable protein in the rumen, and C = non-degradable protein in the rumen and unavailable to the animal.
Table 5.
Effect of phytogenic compounds in mineral or energy supplementation on the abundance of operational taxonomic units (OTU) classified at phylum level of ruminal bacteria and Archaea domain (expressed as median ± interquartile range) of the in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season.
Table 5.
Effect of phytogenic compounds in mineral or energy supplementation on the abundance of operational taxonomic units (OTU) classified at phylum level of ruminal bacteria and Archaea domain (expressed as median ± interquartile range) of the in Nellore cattle grazing Urochloa brizantha cv. Marandu during the rainy season.
| Supplements1 | P-value2 | ||||||
|---|---|---|---|---|---|---|---|
| Domain; Phylum (%) | EW | EPHA | MW | MPHA | ST | PHA | ST × PHA |
| Archaea; Euryarchaeota | 4.84 ± 1.55 | 4.37 ± 1.32 | 3.07 ± 3.20 | 3.80 ± 1.52 | 0.343 | 0.782 | 0.626 |
| Bacteria; Firmicutes | 48.88 ± 5.80 | 46.24 ± 5.12 | 51.22 ± 2.91 | 47.99 ± 4.84 | 0.069 | 0.418 | 0.254 |
| Bacteria; Bacteroidetes | 27.71 ± 3.53 | 30.93 ± 4.62 | 25.87 ± 1.58 | 27.88 ± 2.90 | 0.089 | 0.244 | 0.248 |
| Bacteria; Proteobacteria | 1.42 ± 0.63 | 1.40 ± 0.39 | 1.02 ± 0.57 | 1.05 ± 0.64 | 0.046 | 0.828 | 0.259 |
| Bacteria; Spirochaetae | 1.42 ± 0.50 | 1.45 ± 0.51 | 1.02 ± 0.44 | 0.91 ± 0.40 | 0.022 | 0.953 | 0.152 |
| Bacteria; Chloroflexi | 1.09 ± 0.40 | 0.99 ± 0.39 | 0.83 ± 0.90 | 0.98 ± 0.63 | 0.812 | 0.953 | 0.953 |
| Bacteria; Actinobacteria | 0.98 ± 0.43 | 0.95 ± 0.33 | 0.91 ± 0.52 | 0.88 ± 1.15 | 0.782 | 0.984 | 0.978 |
| Bacteria; Tenericutes | 0.74 ± 0.24 | 0.65 ± 0.11 | 0.94 ± 0.44 | 1.03 ± 0.38 | 0.114 | 0.664 | 0.360 |
| Bacteria; SR1 (Absconditabacteria) | 0.45 ± 0.36 | 0.65 ± 0.45 | 0.77 ± 0.21 | 0.52 ± 0.45 | 0.332 | 0.722 | 0.584 |
| Bacteria; Verrucomicrobia | 0.44 ± 0.30 | 0.32 ± 0.22 | 0.51 ± 0.23 | 0.15 ± 0.43 | 0.635 | 0.022 | 0.133 |
| Bacteria; Fibrobacteres | 0.20 ± 0.14 | 0.29 ± 0.08 | 0.21 ± 0.21 | 0.15 ± 0.18 | 0.275 | 0.551 | 0.404 |
1 EW= energy supplement containing corn gluten meal and minerals without phytogenic compounds addition; EPHA= energy supplement containing corn gluten meal and minerals with phytogenic compounds addition (containing 10% of carvacrol and cinnamaldehyde oil, and 90% hydrolyzable tannins extracted from berries and grapes) at a dose of 1.5 g/kg of ingested DM. MW= mineral supplement warranty levels: Ca 123.0 g/kg; P 90.0 g/kg; Cu 1040.0 mg/kg; Mn 500 mg/kg; Zn 2000.0 mg/kg; Co 15.0 mg/kg; I 67.0 mg/kg; Se 14.0 mg/kg. MPHA= Guarantee levels similar to DM include phytogenic compounds at a dose of 1.5 g/kg of ingested DM. 2 ST = effect of the type of supplement as mineral or energy supplement by a Friedman test. PHA = effect of the addition of phytogenic compounds by a Friedman test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



